
D inosaur Paleobiology
30
Stephen L. Brusatte Dinosaur Paleobiology Series Editor: Professor Michael J. Benton Topics in Paleobiology
Transcript of D inosaur Paleobiology

Stephen L. Brusatte
Dinosaur Paleobiology
Series Editor: Professor Michael J. Benton
Topics in PaleobiologyThe study of dinosaurs has been experiencing a remarkable renaissance over the past few decades.
Scientifi c understanding of dinosaur anatomy, biology, and evolution has advanced to such a degree
that paleontologists often know more about 100-million-year-old dinosaurs than many species of
living organisms. This book provides a contemporary review of dinosaur science intended for students,
researchers, and dinosaur enthusiasts. It reviews the latest knowledge on dinosaur anatomy and phylogeny,
how dinosaurs functioned as living animals, and the grand narrative of dinosaur evolution across the
Mesozoic. A particular focus is on the fossil evidence and explicit methods that allow paleontologists to
study dinosaurs in rigorous detail. Scientifi c knowledge of dinosaur biology and evolution is shifting fast,
and this book aims to summarize current understanding of dinosaur science in a technical, but accessible,
style, supplemented with vivid photographs and illustrations.
The Topics in Paleobiology Series is published in collaboration with the Palaeontological Association,
and is edited by Professor Mike Benton, University of Bristol.
Stephen Brusatte is a vertebrate paleontologist and PhD student at Columbia University and the American
Museum of Natural History. His research focuses on the anatomy, systematics, and evolution of fossil
vertebrates, especially theropod dinosaurs. He is particularly interested in the origin of major groups such
as dinosaurs, birds, and mammals. Steve is the author of over 40 research papers and three books, and his
work has been profi led in The New York Times, on BBC Television and NPR, and in many other
press outlets.
Photography: Mick Ellison
Life reconstructions: Jason Brougham
Skeletal reconstructions: Scott Hartmann
A companion website with additional resources is available at:www.wiley.com/go/brusatte/dinosaurpaleobiology
Cover image: The small feathered dromaeosaurid Sinornithosaurus (“Dave” specimen) from the Early Cretaceous of China.Photograph by Mick Ellison.
Cover design by Design Deluxe
Dinosau
r Paleobiology
Bru
satte


Dinosaur Paleobiology

Books in the Topics in Paleobiology series will feature key fossil groups, key events, andanalytical methods, with emphasis on paleobiology, large-scale macroevolutionary studies,and the latest phylogenetic debates.
The books will provide a summary of the current state of knowledge and a trusted routeinto the primary literature, and will act as pointers for future directions for research. As well asvolumes on individual groups, the Series will also deal with topics that have a cross-cuttingrelevance, such as the evolution of significant ecosystems, particular key times and events in thehistory of life, climate change, and the application of new techniques such as molecularpaleontology.
The books are written by leading international experts and will be pitched at a levelsuitable for advanced undergraduates, postgraduates, and researchers in both thepaleontological and biological sciences.
The Series Editor is Mike Benton, Professor of Vertebrate Palaeontology in the Schoolof Earth Sciences, University of Bristol.
The Series is a joint venture with the Palaeontological Association.
COMPANION WEBSITE
This book has a companion website:
www.wiley.com/go/br usatte/dinosaurpaleobiology
with Figures and Tables from the book for downloading

DinosaurPaleobiologyStephen L. Brusatte

This edition first published 2012 � 2012 by John Wiley & Sons, Ltd.
Wiley-Blackwell is an imprint of John Wiley & Sons, formed by the merger of Wiley’s global Scientific,Technical and Medical business with Blackwell Publishing.
Registered office: John Wiley & Sons, Ltd, The Atrium, Southern Gate, Chichester, West Sussex,PO19 8SQ, UK
Editorial offices: 9600 Garsington Road, Oxford, OX4 2DQ, UKThe Atrium, Southern Gate, Chichester, West Sussex, PO19 8SQ, UK111 River Street, Hoboken, NJ 07030-5774, USA
For details of our global editorial offices, for customer services and for information about how toapply for permission to reuse the copyright material in this book please see our websiteat www.wiley.com/wiley-blackwell.
The right of the author to be identified as the author of this work has been asserted in accordance with theUK Copyright, Designs and Patents Act 1988.
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, ortransmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise,except as permitted by the UK Copyright, Designs and Patents Act 1988, without the prior permission ofthe publisher.
Designations used by companies to distinguish their products are often claimed as trademarks.All brand names and product names used in this book are trade names, service marks, trademarks orregistered trademarks of their respective owners. The publisher is not associated with any product orvendor mentioned in this book. This publication is designed to provide accurate and authoritativeinformation in regard to the subjectmatter covered. It is sold on the understanding that the publisher is notengaged in rendering professional services. If professional advice or other expert assistance is required,the services of a competent professional should be sought.
Library of Congress Cataloging-in-Publication Data
Brusatte, Stephen.Dinosaur paleobiology / Stephen L. Brusatte.
p. cm. – (Topics in paeobiology)Includes bibliographical references and index.ISBN 978-0-470-65657-0 (hardcover : alk. paper) 1. Dinosaurs. 2. Paleobiology. I. Title.QE861.4.B786 2012567.9–dc23 2011050466
A catalogue record for this book is available from the British Library.
Wiley also publishes its books in a variety of electronic formats. Some content that appears in printmaynotbe available in electronic books.
Set in 9/11.5pt Trump Mediaeval-Roman by Thomson Digital, Noida, India
1 2012

DedicationTo my wife, Anne


Contents
Foreword ix
Preface xi
Acknowledgments xiii
Chapter 1 An Introduction to Dinosaurs 1
Chapter 2 Hard Tissues 29
Chapter 3 Soft Tissues 65
Chapter 4 Phylogeny 88
Chapter 5 Form 117
Chapter 6 Locomotion and Posture 135
Chapter 7 Feeding and Diet 159
Chapter 8 Reproduction, Growth, and Physiology 191
Chapter 9 Paleoecology and Dwelling 227
Chapter 10 Macroevolution and Extinction 242
References 262
Index 309
Plate section between pp. 130 and 131
vii


Foreword
Paleobiology is a vibrant discipline that addressescurrent concerns about biodiversity and aboutglobal change. Further, paleobiology opens unim-agined universes of past life, allowing us to exploretimes when the world was entirely different andwhen some organisms could do things that are notachieved by anything now living.
Much current work on biodiversity addressesquestions of origins, distributions, and future con-servation. Phylogenetic trees based on extant or-ganisms can give hints about the origins of cladesand help answer questions about why one clademight be more species-rich (“successful”) than an-other. The addition of fossils to such phylogeniescan enrich them immeasurably, thereby giving afuller impression of early clade histories, and soexpanding our understanding of the deep origins ofbiodiversity.
In the field of global change, paleobiologists haveaccess to the fossil record and this gives accurateinformation on the coming and going of majorgroups of organisms through time. Such detailedpaleobiological histories can be matched to evi-dence of changes in the physical environment, suchas varying temperatures, sea levels, episodes ofmid-ocean ridge activity,mountain building, volcanism,continental positions, and impacts of extraterres-trial bodies. Studies of the influence of such eventsand processes on the evolution of life address corequestions about the nature of evolutionary pro-cesses on the large scale.
As examples of unimagined universes, one needonly think of the life of the Burgess Shale or thetimes of the dinosaurs. The extraordinary arthro-
pods and other animals of the Cambrian sites ofexceptional preservation sometimes seem morebizarre than the wildest imaginings of a sciencefiction author. During the Mesozoic, the sauropoddinosaurs solved basic physiological problems thatallowed them to reach bodymasses ten times thoseof the largest elephants today. Further, the giantpterosaur Quetzalcoatlus was larger than any fly-ing bird, and so challenges fundamental assump-tions in biomechanics.
Books in the Topics in Paleobiology series willfeature key fossil groups, key events, and analyticalmethods, with emphasis on paleobiology, large-scale macroevolutionary studies, and the latestphylogenetic debates.
The bookswill provide a summary of the currentstate of knowledge, a trusted route into the primaryliterature, and will act as pointers for future direc-tions for research. As well as volumes on individualgroups, the Series will also deal with topics thathave a cross-cutting relevance, such as the evolu-tion of significant ecosystems, particular key timesand events in the history of life, climate change, andthe application of new techniques such as molecu-lar paleontology.
The books are written by leading internationalexperts and will be pitched at a level suitable foradvanced undergraduates, postgraduates, and re-searchers in both the paleontological and biologicalsciences.
Michael Benton,Bristol,
November 2011
ix


Preface
Dinosaurs are everywhere these days. They are themost popular exhibits in many museums, the starsof movies and the focus of television documen-taries, the pitchmen in advertising campaigns andthe subject of gushing articles in magazines andnewspapers. Looking at how dinosaurs are por-trayed in the popular press, it is easy to lump themtogether with leprechauns, unicorns, and dragons –creatures of myth and iconic lore that only exist inthe imaginations of children and thewhimsy of popculture. But dinosaurs are not creatures of fantasy –they were real animals, of many fantastic shapesand sizes, that dominated terrestrial ecosystems foran astounding span of over 160 million years. Theywere living, breathing, feeding, moving, reprodu-cing, evolving organisms that originated in theaftermath of the worst mass extinction in earthhistory, rose to dominance as a supercontinent wassplitting and climates were fluctuating, evolvedinto some of the largest andmost fearsome animalsthe planet has ever seen, and then suddenly wentextinct right at the same time that a giant comet orasteroid slammed into the earth and supervolca-noeswere belching rivers of lava. And perhapsmostastonishing of all, these ancient creatures, so oftensymbols of lethargy and failure, were the ancestorsof one of the most successful groups of livinganimals: the birds.
The scientific study of dinosaurs has beenexperiencing a remarkable renaissance over thepast couple of decades. Scientific understanding ofdinosaur anatomy, biology, and evolution has ad-vanced to such a degree that paleontologists oftenknow more about 100 million-year-old dinosaursthan many species of living organisms. Research is
proceeding at a frenetic pace, as illustrated by asimple statistic: during 2010, the year I proposedand beganwriting this book, some 63 new species ofdinosaurswere discovered. That’s a stupendous rateof over one new species per week, which has largelybeen fueled by a great increase in fieldwork explora-tion (especially in China and South America) andan ever-expanding roster of graduate students andother young researchers choosing to studydinosaurs. And not only is our stockpile of dinosaurfossils growing at an exponential pace, but so is thedevelopment of new research techniques. It used tobe that paleontologists could pontificate on thebiology, evolution, andextinctionof dinosaursbasedonly on the flimsiest scraps of evidence, interpretedwith a healthy dose of imagination and a snickeringdismissal of the explicit, quantitative, repeatablemethodologies thathave longbeen thenorminmostother sciences. Those days are long gone. Today,dinosaur paleontology is a dynamic science thatdemands evidence-based rigor and is firmly integr-ated with many other scientific disciplines. Indeed,researchers often draw from a diverse repertoire ofanatomy, geology, chemistry, physics,mathematics,and statistics when studying dinosaurs. It is notuncommon to see advanced calculus used to esti-matedinosaurbodymasses, computerized engineer-ing analyses marshaled to test whether certaindinosaurs were capable of feeding or moving a cer-tain way, or statistics utilized to explicitly assesswhether some dinosaurs were evolving faster orslower than others.
The breadth of current dinosaur research is vast.Some scientists spend their careers discovering anddescribing new species, others may focus solely on
xi

anatomy or genealogy, and others concentrateon studying dinosaur locomotion or feeding. Ingeneral, though, all contemporary work on dino-saurs provides evidence for addressing two mainquestions. First, how did dinosaurs function asliving animals? Second, what is the grand narrativeof dinosaur evolution across the Mesozoic? Theonly way to attack these questions in a defensiblecompetent manner is to interpret the primaryevidence – the actual dinosaur fossils that providea bedrock for the entire enterprise of dinosaurresearch – using explicit quantitative methodolo-gies. The emerging answers to these questions, andthe evidence and methods that are revealing them,are the focus of this book. Like any science, dino-saur paleontology is constantly changing as newfossils are found and new research techniques aredeveloped and refined. Our knowledge of dinosaurbiology and evolution is shifting fast, and this bookis an attempt to capture what is currently knownabout this remarkable group of ancient creaturesthat dominated our planet for so long.
From a more personal standpoint, this book isalso a young, perhaps brazen, researcher’s exami-nation of his field of study. I am in the somewhatunusual position of writing this book as a PhDstudent – a scientist without an advanced degree,with less than a decade of research experience,whohas not had the time and wisdom to (at least yet)make a substantial mark on the field. But althoughI may not be the most traditional author of atechnical dinosaur book, and although perhaps Ishould be focusing more on my thesis than onwriting books, I feel that I am able to present a
perspective that has yet to be tapped by the over-saturated dinosaur book market. I have beenbrought up and trained within the dynamism ofcontemporary dinosaur research, and have beenexperiencing the explosive growth of this field as adizzying cocktail of new discoveries and techni-ques have enabled modern scientists to under-stand dinosaurs in unprecedented detail. Inmany ways this book is a personal journey. I donot pretend that this book is an exhaustive ency-clopedia of everything that is currently knownabout dinosaurs, or a technical critique of theminutiae of every method and each piece of evi-dence. Instead, what I present ismy understandingof dinosaur biology and evolution – the under-standing of a student actively learning aboutdinosaurs and in the midst of planning his ownresearch program and career. I present what I findinteresting and empowering, what I think is mostimportant and exciting about contemporaryresearch, and where I think the field is heading.
So, then, what do scientists actually know aboutdinosaurs? As it turns out, the truth about dinosaurbiology and evolution is surely more fascinatingthan even themost sensational dinosaur documen-tary or movie, andmore than fascinating enough tofuel the passion of this member of theMTV genera-tion. Indeed, without even a hint of hyperbole, thestory of dinosaur evolution is one of the greateststories ever told.
Steve BrusatteNew York, USA
September 1, 2011
xii P R E F A C E

Acknowledgments
Writing a book is a lot of fun, but also a lot of work.Although my name may appear on the cover, thisbook could never have been written without thehelp of so many friends and colleagues across theglobe. One of my favorite aspects of paleontologi-cal research is the friendships and collaborationsthat are fostered, and I’m pleased to be able (even ifin a small way) to showcase the work of many ofmy colleagues and include their contributions inthe form of photos and illustrations. This bookrelects my personal journey studying paleontology,and building a career in research, over the pastdecade. I have tried, wherever possible, to includephotographs and figures that I have compiled dur-ing my research work, fieldwork, and museumvisits, or those provided by trusted colleagues andfriends. These colleagues are too numerous tothank here, but individuals who helped withimages are credited in the figure captions. To allof them, let me apologize one final time for all mynagging questions.
I have been very fortunate in my young careerto have been mentored by three very excellentadvisors: my undergraduate advisor Paul Serenoat the University of Chicago, my Master’s advisorMike Benton at the University of Bristol, and myPhD advisor Mark Norell at Columbia Universityand the American Museum of Natural History. Irecognize how lucky I have been to study underthree of the most prominent luminaries in thefield, and thank them for all of heir guidance,advice, and support over the years. Specific to thisbook, I would like to offer my sincere thanks toMike Benton, the editor of the Topics in Paleobi-ology series, who invited me to write a book on
dinosaurs for Wiley-Blackwell. I am humbled thathe would place such trust in me at such a youngstage in my career, and I hope that I have seizedthis remarkable opportunity and written a bookthat justifies his confidence in me. And to MarkNorell, my current advisor, please know that I willalways appreciate the freedom that you have pro-vided me as a student to pursue whatever interestsme. Not every PhD advisor would allow his stu-dent to put a doctoral thesis on the backburner towrite a book.
Although many colleagues helped with imagesand advice, a few people deserve special mentionhere. I am very pleased to feature the skeletalreconstructions of Scott Hartman, the photographsofMick Ellison, and the artistic life reconstructionsof Jason Brougham. Scott, Mick, and Jason arethree of the best artists in the business, and areconsistently producing beautiful and scientificallyaccurate work that, at least in my opinion, sets abenchmark for the field. Without their contribu-tions this book would be little more than a jumbleof 130,000 words; if this book does succeed inbringing dinosaurs to life, it is largely due to theirreconstructions, photos, and illustrations. A lot ofthe work they have provided here has not beenreproduced before, and all three worked tirelesslyto help make this book something more than just arun-of-the-mill dinosaur tome. Several trusted col-leagues also read large portions of this book, includ-ing Roger Benson, Mike Benton, Richard Butler,Matt Carrano, Greg Erickson, Paul Gignac, JohnHutchinson, and Pat O’Connor, as well as theformal reviewers (Paul Barrett and Larry Witmer).Their advice was instrumental, and I thank them
xiii

for their encouragement, suggestions, and frankcriticism that helped tighten my writing and im-prove the text. All mistakes, however, are of coursemine.
My continuing development as a scientist hasbeen facilitated by the friendship, collaboration,advice, and assistance of many trusted colleagues.I would like to especially acknowledge my twoclosest colleagues, Roger Benson and Richard But-ler, whom I consider something of scientific bloodbrothers. I’ve been fortunate to work on manyprojects with Richard and Roger, and have sharedmany long car journeys, evenings over beers, andhours in museum collections learning from them.They are two of the most dynamic, thoughtfulyoung researchers in the field, and I have no doubtthat they will emerge as among the most respectedvoices in dinosaur research as their careers unfold.I’ve also enjoyed fruitful collaborations and friend-ships with many other close colleagues, includingThomas Carr, Zolt�an Csiki, Phil Currie, GraemeLloyd, Octavio Mateus, Josh Mathews, GrzegorzNied�zwiedzki, Marcello Ruta, Steve Wang, ScottWilliams, and Tom Williamson. My fellow gradu-ate students and advisors have provided constantinspiration, including Carol Abraczinskas, MarcoAndrade, Amy Balanoff, Robin Beck, Mark Bell,Gabe Bever, Jianye Chen, Jonah Choiniere, JohnFlynn, Andres Giallombardo, Christian Kam-merer, Mike LaBarbera, Shaena Montanari,Sterling Nesbitt, Paul Olsen, Rui Pei, AlbertPrieto-M�arquez, Manabu Sakamoto, MichelleSpaulding, Mark Webster, Hongyu Yi, and MarkYoung. Three colleagues that I have only workedwith briefly, but have long admired for their
adherence to quantitative rigor and ability to askand answer interesting questions, are Matt Carra-no, Greg Erickson, and John Hutchinson. Manyother colleagues also have helped me along, in-cluding Robert Bronowicz, Dan Chure, Julia Des-ojo, Phil Donoghue, Gareth Dyke, Jerzy Dzik,Martin Ezcurra, Dave Gower, Mike Henderson,Dave Hone, Steve Hutt, Randy Irmis, Max Langer,Pete Makovicky, Darren Naish, Chris Organ, Em-ily Rayfield, Nate Smith, Tomasz Sulej, CorwinSullivan, Alan Turner, M�atyas Vremir, Anne Weil,Jessica Whiteside, Zhao Xijin, and Xu Xing.
And finally, but most importantly, I must thankmy family and close personal friends. My parents(Jim and Roxanne Brusatte) have long fostered mypassion for paleontology and writing, even going asfar as letting me plan whole days of family vaca-tions dedicated to museum hopping (which mybrothers, Mike and Chris, must have really en-joyed). My parents and in-laws (Peter and MaryCurthoys) have generously provided space in theirhomes for me to write this book. A handful of goodpersonal friends have helped fuel my passion forwriting and have molded me (gradually, and surelywith much pain) into a competent author: FredBervoets, Lonny Cain, Lynne Clos, Allen Debus,Mike Fredericks, Richard Green, Joe Jakupcak,Mike Murphy, and Dave Wischnowsky. I sincerelythank all of the help thatmy editor, Ian Francis, hasprovided with this book. And last, but certainly notleast, I dedicate this book to my patient and beau-tiful wife Anne, who someday (I hope) will under-stand that it isn’t too strange for a grown man tospend his days thinking about 65 million-year-old,6-tonne, killer-toothed megapredators.
xiv A C K N OW L E D G M E N T S

1 An Introductionto Dinosaurs
It is necessary to begin with a straightforward, if not pedantic, question: what is a
dinosaur? In popular parlance a dinosaur is often anything that is old, big, or
frightening. Any kindergartner could identity Tyrannosaurus or Triceratops as
dinosaurs, and they would be correct, but newspapers will often sloppily use the
term ‘‘dinosaur’’ to refer to flying reptiles (pterosaurs), marine reptiles
(plesiosaurs, ichthyosaurs, etc.), or even large mammals (such as the woolly
mammoth). ‘‘Dinosaur’’ has become a cultural and political idiom as well: out-of-
touch politicians or washed-up celebrities are often mockingly ridiculed as
‘‘dinosaurs,’’ a synonym for lethargy, obsolescence, and inevitable extinction.
Although the term ‘‘dinosaur’’ is firmly established in the popular lexicon, it is
also a scientific term that refers to a specific group of organisms that shared
particular anatomical features and lived during a certain period of time. While the
popular definition of ‘‘dinosaur’’ is amorphous, the scientific definition is precise.
Wewill get to that definition in amoment, but first it is necessary to reviewexactly
where dinosaurs fit in the tree of life – when they evolved, what they evolved
from, and who their closest relatives are – so it is easier to comprehend the
explicit distinction between dinosaur and non-dinosaur. Some of the following
discussion may seem elementary to more advanced readers, and I intentionally
use a more conversational tone in this introduction to appeal to non-specialists
and younger students. It is important, however, to set the stage for this book by
first painting in broad strokes, before progressing to a more nuanced discussion
of dinosaur anatomy, ecology, behavior, and function.
Dinosaur Paleobiology, First Edition. Stephen L. Brusatte.
� 2012 John Wiley & Sons, Ltd. Published 2012 by John Wiley & Sons, Ltd.
1

Dinosaurs: A BriefBackground
Dinosaurs are one of the best-known, most inten-sively studied, and most successful groups of tetra-pods: animalswith a backbone that have limbswithdigits (fingers and toes) (Fig. 1.1). Within the tetra-pod group, dinosaurs are members of a speciosesubgroup of reptiles called the Archosauria, whichliterally means ‘‘ruling reptiles’’ in Greek (Cope,1869; Romer, 1956; Carroll, 1988; Benton, 2005)(Figs 1.1–1.6). This is a fitting moniker, as archo-saurs have been a major component of terrestrialecosystems since the early Mesozoic, and for largeswaths of time have been ecologically dominantand incredibly diverse (Benton, 1983; Fraser, 2006).Living archosaur subgroups include two majorclades, birds and crocodylomorphs, which areamong the most familiar and successful groups ofextant vertebrates (note that a ‘‘clade’’ refers to agroup of animals that includes an ancestor and all ofits descendants; Fig. 1.5) (Gauthier, 1986; Sereno,1991a; Nesbitt, 2011). However, the great majorityof archosaur diversity is extinct, and the two main
living groups merely represent two highly aberrantbody types (fliers and semiaquatic sprawlers) thatwere able to endure several mass extinction eventsthat pruned most other lineages on the archosaurfamily tree. Dinosaurs, without a doubt, are themost familiar of these extinct archosaurs.
The archosaur clade is an ancient group thatoriginated approximately 250 million years ago(Nesbitt, 2003, 2011; Brusatte et al., 2010a,2011a; Nesbitt et al., 2011). Some of the closestarchosaur relatives are known from the Late Perm-ian (e.g. Dilkes, 1998; Nesbitt et al., 2009a), andarchosaurs themselves arose within the first fewmillion years after the devastating Permo-Triassicmass extinction, the largest instance of mass deathin earth history, estimated to have eradicated75–95% of all species (Raup, 1979; Stanley andYang, 1994; Benton, 2003; Erwin, 2006; Claphamet al., 2009). The Permo-Triassic extinction intervalwas a time of death and destruction on a massivescale, but its aftermath was a time of equally large-scale rebirth: ecosystems were reshuffled, organ-isms thatwere once overshadowed had the freedomto flower, and entirely new groups originated and
Lizar
ds an
d Snak
es
Amniota
Diapsida
Archosauria
Lissa
mphib
ians
Mam
mal
s
Non-Avian Dinosaurs Birds
Tetrapoda
Dinosauria
Cro
codyl
omorp
hs
Figure 1.1 A simplified genealogical tree (cladogram) of tetrapods (limbed vertebrates) showing the position of dinosaurs
and their closest relatives. Artwork by Simon Powell, University of Bristol.
2 A N I N T R O D U C T I O N T O D I N O S A U R S

diversified in the barren, post-extinction landscape(Benton et al., 2004; Sahney and Benton, 2008).Among these entirely new groups were ‘‘modern’’lineages such as turtles, mammals, lepidosaurs (li-zards and their relatives), lissamphibians (frogs and
salamanders), and archosaurs. It is no wonder thatthe Triassic Period is often called the ‘‘birth ofmodern ecosystems,’’ as so many of today’s mostdistinctive and successful clades originated duringthis time.
Figure 1.2 A montage of the skulls of various archosaurs, including the rauisuchian crurotarsan Batrachotomus (A), the
aetosaurian crurotarsan Aetosaurus (B), the phytosaurian crurotarsan Nicrosaurus (C), the poposauroid crurotarsan
Lotosaurus (D), the ornithosuchid crurotarsan Riojasuchus (E), and the sauropodomorph dinosaur Plateosaurus (F).
D I N O S A U R S : A B R I E F B A C K G R O U N D 3

Figure 1.3 A montage of life reconstructions of various crurotarsan (crocodile-line) archosaurs. Illustrations courtesy of
Dr Jeff Martz, National Park Service.
Tria
ssic
Jura
E
M
L
E
Ind
Ole
Anis
Lad
Crn
Nor
Rha
Het
201.5
228
235
242
247.2
251.3
252.3
~208
Permo-Triassic Extinction
Highest Rates of Morphological
Character Evolution
Major Increase in Crurotarsan Disparity
(Morphological Variability)
Major Increase in Dinosaur
Abundance and DiversitySin
Plb
Toa
197
190
183
Major Increase in Dinosaur Disparity
Substantial Increase in Archosaur
Faunal Abundance and Diversity
End Triassic Extinction
Crurotarsan Disparity Crashes
Major Increase in Archosaur Diversity
(Crurotarsi and Avemetatarsalia)
First Well-Preserved
Archosaur Body Fossils
First Archosaur Fossils:
Fragmentary Bones and Footprints
Figure 1.4A general timeline of important events during the first 70 million years of archosaur evolution during the Triassic
and early Jurassic. Image based on illustration in Brusatte et al. (2011a).
4 A N I N T R O D U C T I O N T O D I N O S A U R S

The archosaur clade diversified rapidly after itsorigination, as most of the major archosaur sub-clades and body planswere established by the end ofthe Early Triassic, a mere 5 million years after themass extinction (Brusatte et al., 2011b) (Fig. 1.4).The oldest unequivocal archosaur body fossilwith awell-constrained age and phylogenetic position isXilousuchus, from the late Olenekian/earlyAnisian (c.247–248 million years ago) of China
(Nesbitt et al., 2011). This species is a derivedmember of the ‘‘crocodile line’’ of archosaur phy-logeny, which is properly referred to as Crurotarsi(also sometimes called Pseudosuchia). Crurotarsiincludes crocodylomorphs and their closest extinctrelatives, whereas the other half of the archosaurclade, the ‘‘bird-line’’ groupAvemetatarsalia (some-times also called Ornithodira), includes birds,dinosaurs, and pterosaurs (the familiar flying
Lizar
ds
Aet
osaurs
“Rau
isuch
ians”
Cro
codyl
omorp
hs
Phyt
osaurs
Amniota
Diapsida
Archosauria
Lissa
mphib
ians
Mam
mal
s
Non-Avian Dinosaurs Birds
Tetrapoda
Pte
rosa
urs
Crurotarsi Avemetatarsalia
Dinosauria
Figure 1.5 A simplified genealogical tree (cladogram) of archosaurs, showing the position of dinosaurs and their closest
relatives. Artwork by Simon Powell, University of Bristol.
Figure 1.6 Life reconstructions of the basal non-dinosaurian dinosauromorphs Marasuchus and Silesaurus, two of the
closest relatives to dinosaurs. Illustrations courtesy of Dr Jeff Martz, National Park Service.
D I N O S A U R S : A B R I E F B A C K G R O U N D 5

reptiles) (Gauthier, 1986; Sereno, 1991a; Benton,1999, 2004; Irmis et al., 2007a; Brusatte et al.,2010a, 2011b; Nesbitt, 2011) (Fig. 1.5). BecauseXilousuchus is a member of the crocodile lineage,then the bird line (but not true birds themselves)must have also been present by approximately248 million years ago, because these two lineagesare each other’s closest relative, and the presence ofone implies the contemporary existence of the other(see Norell, 1992, 1993 for details of such ‘‘ghostlineages,’’ which will be discussed later in the text).
Although the bird lineage, ofwhich dinosaurs area part,must have been present by the Early Triassic,the first body fossils of truly dinosaur-like animalsare not known until the late Anisian (c.243–244million years ago) (Nesbitt et al., 2010). Thesefossils do not belong to true dinosaurs, as willbecome clear below, but are among the handful ofclosest relatives to dinosaurs, and likely resembledand behaved like their more famous cousins(Fig. 1.6). More properly, they are members of the‘‘dinosaur stem clade,’’ technically known as Dino-sauromorpha (Sereno, 1991a; Benton, 1999, 2004;
Ezcurra, 2006; Brusatte et al., 2010a;Nesbitt, 2011).Among the best known species are Lagerpeton(Sereno and Arcucci, 1993), Marasuchus (Serenoand Arcucci, 1994), Dromomeron (Irmis et al.,2007a), Silesaurus (Dzik, 2003), and Asilisaurus(Nesbitt et al., 2010). Middle to Late Triassic dino-sauromorphs were small animals, no bigger than asmall dog, and were incredibly rare in their ecosys-tems. The tiny fragile footprints of some of theseclose dinosaur cousins are known from severalfossils sites in the western United States (Peabody,1948) and Europe (Haubold, 1999; Ptaszynski, 2000;Klein andHaubold, 2007; Brusatte et al., 2011a), andthese are remarkably scarce compared with thefootprints of other characteristic Triassic animals,especially crocodile-line archosaurs (Fig. 1.7). Itseems therefore that these dinosaur stem taxa weresmall, rare, only represented by a few species, andovershadowed by other contemporary reptiles.From such a humble beginning came the dinosaurs.
True dinosaurs likely originated some time inthe Middle Triassic, although it is difficult to pin-point the exact time. The first dinosaur body fossils
Figure 1.7 A montage of photographs and illustrations of the footprints (A–D) and handprint (E) of a small-bodied
quadrupedal dinosauromorph from the Early Triassic of Poland. These fossils are currently the oldest known fossil
evidence of the dinosauromorph lineage. Scale bars equal 1 cm. Images by Grzegorz Nied�zwiedzki and modified from
Brusatte et al. (2011a).
6 A N I N T R O D U C T I O N T O D I N O S A U R S

are known from rocks that were deposited in Ar-gentina at approximately the Carnian–Norianboundary (c.228 million years ago) (Rogers et al.,1993; Shipman, 2004; Brusatte et al., 2010b; Ezcur-ra, 2010a; Langer et al., 2010; Martinez et al., 2011)(Figs 1.8 and 1.9). However, it is almost certain thatdinosaurs arose several million years earlier. First,the closest relatives of dinosaurs were clearly pres-ent by at least 243 million years ago, as outlinedabove, and it is reasonable to hypothesize thatdinosaurs originated around this time (Nesbitt etal., 2010). Second, there are a number of provocativefootprints, which closely match the feet of primi-tive dinosaurs, that have recently been describedfrom the Ladinian (c.242–235 million years ago) ofEurope and South America (Gand and Demathieu,2005; Melchor and de Valais, 2006). Regardless ofthe exact timing of dinosaur origins, which willsurely become clearer as new fossils are found, it isundeniable that dinosaurs began to diversify quick-ly once they originated. By the time the first dino-saur body fossils appear in the fossil record,representatives of the three major subgroups ofdinosaurs – the carnivorous theropods, long-necked
sauropodomorphs, and herbivorous and oftenarmored or crested ornithischians – are alreadypresent (Sereno and Novas, 1992; Sereno et al.,1993; Langer et al., 1999, 2010; Butler et al., 2007;Martinez and Alcober, 2009; Brusatte et al., 2010b;Ezcurra and Brusatte, 2011; Martinez et al., 2011).
Therefore, by the Late Triassic, the Age of Di-nosaurs was in full swing, and over the course ofthe next 50 million years dinosaurs would contin-ue to diversify into new species and body types,before ultimately becoming the dominant mid-to-large size vertebrates in terrestrial ecosystemsglobally in the Early Jurassic, about 176 millionyears ago (Benton, 1983; Brusatte et al., 2008a,2008b, 2010b) (Fig. 1.4). From this point on,throughout the remainder of the Jurassic and theCretaceous, from approximately 175 to 65 millionyears ago, dinosaurs truly were ‘‘ruling reptiles’’ inevery sense of the phrase. They lived in all cornersof the globe, including the Arctic highlands, andreached some of the most stupendous sizes everseen in land-living animals. Some species devel-oped absurdly long necks, others extravaganthorns and armor that would make a medieval
Figure 1.8 Skeletal reconstructions of four early dinosaurs from the Late Triassic to Early Jurassic: the theropod
Herrerasaurus, the theropod Dilophosaurus, the sauropodomorph Saturnalia, and the ornithischian Heterodontosaurus.
Illustrations by Frank Ippolito (American Museum of Natural History) and modified from Brusatte et al. (2010b).
D I N O S A U R S : A B R I E F B A C K G R O U N D 7

knight blush, and yet others grotesque skulls,longer than an averageman is tall and packed withdagger-like teeth, perfect for delivering bone-crunching bites. This fantastic array of dinosaurs –predators and herbivores, dwarves and 50-m longbehemoths and all sizes in between – continued toevolve in concert with the slow drift of the con-tinents and the roller-coaster wiggles of climatechange, until an unexpected visitor from outerspace smashed into the planet 65 million yearsago, snuffing out the Age of Dinosaurs and permit-ting the survival of only one marginal, aberrantdinosaur subgroup: the birds.
The Scientific Definition ofDinosaurs
The above review liberally used terms like ‘‘truedinosaur’’ and ‘‘close dinosaur cousin.’’ Vagueterminology like this can often be maddening,and can sadly obstruct communication betweenscientists. Thankfully, however, there is an ex-plicit definition of what constitutes a dinosaur(the ‘‘true dinosaurs’’). Dinosaurs are defined by
scientists as ‘‘members of the least inclusive cladecontaining Triceratops horridus and Passer do-mesticus (the living house sparrow)’’ (Padian andMay, 1993; Sereno, 1998; Sereno et al., 2005).At first this definition may seem confusing, andperhaps even counterintuitive, but in fact it isquite straightforward.
Most modern biologists define groups of organ-isms, such as dinosaurs or mammals or birds, basedon ancestry, not on the possession of certain char-acteristics (e.g. de Queiroz and Gauthier, 1990,1992; Sereno, 2005). An animal is a dinosaur if itfalls in a certain place on the family tree of life, inthis case that group of organisms that includesTriceratops, the living sparrow (Passer), and alldescendants of their common ancestor. This hypo-thetical common ancestor can be visually traced ona family tree (properly called a cladogram, or aphylogeny) of reptiles: simply findTriceratops, thenPasser, and then trace the branches leading to bothspecies down to their common meeting point(Fig. 1.10). Any species that can also be traced downto this common ancestor – in other words, anyspecies that descended from this ancestor – is bydefinition a dinosaur.
Figure 1.9 Life reconstructions of early dinosaurs from the Late Triassic. Illustrations courtesy of Dr Jeff Martz, National
Park Service.
8 A N I N T R O D U C T I O N T O D I N O S A U R S

The definition of Dinosauria given above iscalled a phylogenetic definition, and it is a generaldefinition that can be applied to any cladogram.Clearly, however, this definition needs a phylogenyfor context, and it is unintelligible without a clado-gram to refer to. The first scientists to study dino-saurs did not define them this way, which isunsurprising given that these pioneering paleontol-ogists were working in a pre-Darwinian world inwhich evolution (and hence common ancestry)was regarded as heresy. The man who namedDinosauria, Richard Owen (1842), followed thecustom of the time and defined dinosaurs as those
animals possessing a certain set of anatomical fea-tures, which included various traits relating to bodysize, posture, and locomotion (see below). Owensaw these features as essential characteristics – anunchangeable blueprint that set dinosaurs apartfrom other reptiles – but today we simply recognizethem as products of common ancestry, as traits thatall dinosaurs inherited from that distant ancestorthat unites Triceratops and Passer. These are so-called synapomorphies: shared derived characters –evolutionary novelties – that unite a group on thetree of life.
This clarifies an important point: animals such asdinosaurs arenot strictly defined by their anatomicalfeatures, but everygroupon the treeof lifepossesses acharacteristic set of traits inherited from their com-mon ancestor and thus absent in other organisms.These features are said to diagnose dinosaurs, ratherthandefinethem.Ananalogycanbeseeninmedicine:cancer is defined as a disease in which cells growuncontrollably (a process), but is diagnosed by symp-toms such as headaches, swelling, or abnormalbreathing.Doctorsneverrigidlydefineadiseasebasedon symptoms, but a certain disease usually has acharacteristic set of symptoms, and by noticing andstudying these symptoms a doctor can pinpoint thediseasethatiscausingthem.Dinosaurs, therefore,aredefined based on ancestry, but share a common set offeatures, and by identifying and studying these fea-tures scientists canbe sure that a certain specimenororganism is truly a dinosaur.
Characteristic Featuresof Dinosaurs
With the above semantics out of the way, we cannow focus on those features that distinguish dino-
Phylogenetic definitions may seem confusing, but they can be understood with analogies to our own family histories.
Some of my ancestors, for instance, immigrated to the United States from northern Italy. As the story goes, my great
grandfather, upon hearing distressing rumors of anti-Italian sentiment in his soon-to-be new homeland, decided to
change his surname from the obviously Italian ‘‘Brusatti’’ to the somewhat more ambiguous ‘‘Brusatte’’ when
registering as a new citizen. This name change can be thought of as the origin of a new group of organisms, in this
case the Brusatte family, and anybody who has descended from my great grandfather is by definition a Brusatte. It
doesn’t matter what we look like – whether we are tall, short, fat, thin, or bald – or when or where we live. We are
simply Brusattes by definition.
Tricera
tops
Passe
r
Tyran
nosauru
s
Stegosa
urus
Velocir
aptor
Plateo
sauru
s
Brachiosa
urus
Silesa
urus
Marasu
chus
Cro
codyl
omorp
hs
Lizar
ds an
d
Snak
es
DINOSAURIA
Figure 1.10 A schematic illustration showing how a group
(such as Dinosauria) is defined in a phylogenetic sense.
Dinosauria is formally defined as ‘‘members of the least
inclusive clade containing Triceratops horridus and Pass-
er domesticus.’’ This definition requires a genealogical
tree, or phylogeny, to make sense. In this case, locate
Triceratops and Passer on the tree and then trace both
branches back to their common ancestral meeting point
(denoted by a circle). All species that also descended from
this common ancestor are dinosaurs by definition (those
species shown in black), whereas other species that fall
outside this group are not dinosaurs by definition (those
species shown in gray).
C H A R A C T E R I S T I C F E A T U R E S O F D I N O S A U R S 9

saurs. After all, these anatomical features, and theirbiological and ecological significance, are muchmore interesting than the subtleties of cladograms,and the mundane quibbles about whether a certainspecies did or did not descend from a certain com-mon ancestor. This criticism is not to trivializephylogenetic definitions – their strength is in theirexplicitness and stability – but unfortunately te-dious debates have raged over whether a certainspecies is a proper dinosaur or falls just outsideof thegroupdefinedbyTriceratopsandPasser.Theseacademic quarrels can be maddening, because thefocus is on a technicality of nomenclature ratherthan much more illuminating discussions of bio-logy, function, andevolution.And inone sense thesedebates miss the point, because even if an animal isnot quite a dinosaur by definition, it may still havemany features common to other dinosaurs, andmayhave resembled and behaved like true dinosaurs.
A prime example concerns a recently describedgroup of peculiarMiddle to Late Triassic archosaurscalledthesilesaurids.Thereisnoquestionthattheseanimalswereverysimilar todinosaurs, as theyshareseveral derived features with species that are un-equivocally part of the Triceratops–Passer group.But there is debate over whether they are true dino-saurs: whether they descended from the commonancestor of Passer and Triceratops, or whether theyare the closest relatives of true dinosaurs (i.e., areimmediately outside the Triceratops–Passer group)(Dzik, 2003; Ferigolo and Langer, 2007; Irmis et al.,2007a; Brusatte et al., 2010a; Nesbitt et al., 2010;Nesbitt, 2011). This debate is indeed important forthat narrow group of specialists which focuses onreptile phylogeny, and does have important ramifi-cations for understanding patterns of character evo-lution,but isof littleconcerneven formostdinosaurpaleontologists. Therefore, in this section, I take amore catholic view of dinosaurs and focus not onlyon those features that precisely diagnose Dinosaur-ia, but also features that are seen in a handful of theclosest dinosaur relatives, which are not dinosaursby definition but likely were very similar to dino-saurs in a biological sense. Throughout the remain-der of this book the focus will be on true dinosaurs,but close dinosaur cousins (‘‘stem dinosaurs’’) willsometimes be discussed for context or to flesh outexploration of biology, function, or large-scale evo-lutionary patterns.
When outlining features common to all dino-saurs, it is wise to begin with some historicalbackground. Dinosauria was first established as adistinctive group by Owen (1842), who recognizedthat three extinct genera of large reptiles –Megalosaurus, Iguanodon, and Hylaeosaurus –shared several unusual features that were unknownin other reptiles, both living and extinct. Theseincluded features of the hips, limbs, and bodyposture, which generally indicated that dinosaurshad a more upright stance than other reptiles (seereview in Cadbury, 2002). Discoveries of new fossilscontinued at a frenzied pace during the remainder ofthe 19th century, and by the dawn of the 20thcentury paleontologists had recognized that notonly did all known dinosaurs share many features– including several additional hallmarks revealed bythe new finds – but that they could be divided intotwo major subgroups: the ‘‘lizard-hipped’’ saur-ischians, which include theropods and sauropodo-morphs, and the ‘‘bird-hipped’’ ornithischians(Seeley, 1887). These groups are recognized to thisday as the twomajor subdivisions of dinosaurs. Overthe next several decades, however, scientists gradu-allychanged their conceptionofdinosaurs. Formuchof the 20th century, paleontologists considered saur-ischians and ornithischians to be separate lineages,which independently diverged long ago from sepa-rate ‘‘thecodont’’ (primitive archosaur) ancestors.Therefore, all the features common to saurischiansand ornithischians were not seen as the product ofcommon ancestry – characteristics that united alldinosaurs relative to other animals – but rather asinsignificant nuances of the anatomy that evolved inparallel in both groups. The very idea of a single,distinctive dinosaur group had fallen out of favor.
This view began to change in the mid 1970s andwithina fewyearswaswidelydismissedasoutdatedand incorrect. A new generation of paleontologists,motivated by new discoveries and conceptual ad-vances, resurrectedOwen’s (1842) original notionofa single, unique group of Mesozoic reptiles – Dino-sauria – that could be distinguished from all otherorganisms based on their possession of shared de-rived characters. This revolution in thinking wasdrivenbytwomajor factors.First, if saurischiansandornithischiansweredescended fromseparate ances-tors, then the most primitive members of bothgroups should look very different from each other.
10 A N I N T R O D U C T I O N T O D I N O S A U R S

However, as new fossil finds of early saurischiansandornithischianswerediscovered inTriassic rocksacross theworld, thispredictionwasutterly rejected(Welles, 1954; Crompton and Charig, 1962; Reig,1963; Colbert, 1970). Instead, primitive theropods,sauropodomorphs, andornithischianswere remark-ably similar to each other, exactly as would be pre-dicted if they diverged from a single commonancestor. Second, the advent of an explicit, numeri-cal methodology for inferring genealogical relation-ships – cladistics – swept through the field of biologyin the 1970s and 1980s (Hennig, 1965, 1966). Cladis-tic principles hold that a lengthy roster of sharedanatomical features between two groups is muchmore likely to indicate close relationship than par-allelevolution,anditwouldtakequiteabitofspecialpleadingtoretainsaurischiansandornithischiansasseparate entities that evolved somanyeerily similarfeatures independent of each other.
Itwasmore plausible, therefore, that themyriadsimilarities between saurischians and or-nithischians meant that these two groups des-cended from a common ancestor, and could beunited as a single, larger group: Dinosauria. Thisview was persuasively articulated in a seminal1974 paper by Robert Bakker and Peter Galton. Indoing so, Bakker and Galton (1974: 168–169)highlighteda surprisingly long listof characteristicdinosaur features,manyofwhichhadbeenrevealedby new discoveries during that long dark periodwhen saurischians and ornithischians were as-sumed to be nothing but distant, convergent rela-tives. These features included an upright and fullyerectposture,anenlargeddeltopectoralcrestonthehumerus (which anchors large shoulder and chestmuscles), a perforated hip socket for articulationwith theheadof the femur, awell-developed fourthtrochanter and lesser trochanter on the femur(which anchor hindlimb muscles), and an anklejoint in which the proximal tarsals (astragalus andcalcaneum) were ‘‘fixed immovably on the ends ofthe tibia and fibula [resulting in a] simple unidirec-tional hinge between the astragalus–calcaneumand distal tarsals.’’ As is evident, many of thesefeatures have to do with the posture, strength, andrange of motion of the forelimbs and hindlimbs:compared with their closest relatives, dinosaurshad amore upright stance and stronger,moremus-cular legs, which moved in a more restricted fore–
aft direction, ideal for fast running and keen bal-ance. Importantly, Bakker andGalton (1974) acute-ly recognizedthatmanyof thesehallmarkdinosaurfeatures are also present in living birds, and thussupport a close relationshipbetweendinosaurs andbirds. This was not a new idea, but one that wasrapidly gaining traction in the field at the time. Ithad been proposed as early as the 1860s (Huxley,1868, 1870a, 1870b), but had largely been ignoreduntil the pioneering studies of John Ostrom in the1960s and 1970s (Ostrom, 1969, 1973).
It is a great testament to the work of Bakker andGalton (1974) that many of the features they de-scribed as dinosaur trademarks are still consideredvalid today. This is no small feat, as the exactcharacteristics that diagnose a clade on the tree oflife, such as Dinosauria, are constantly changing asnew fossils are discovered and ideas are reinter-preted. At one point in time a certain character,such as a large deltopectoral crest, may only beknown in one group, such as dinosaurs. It is easyto envision, however, how a single new fossil dis-covery, such as a new close dinosaur cousin with alarge crest, could reveal that this feature is morewidely distributed. This has, in fact, happened toseveral of Bakker and Galton’s diagnostic charac-ters but, importantly, most of the features theydescribed are still only known in dinosaurs and ahandful of their closest cousins, and their generalargument that dinosaurs are distinguished fromother reptiles by their posture and hindlimb anato-my still stands. But perhaps most important of all,Bakker and Galton’s (1974) paper was a catalyst forfuture studies, and authors continue to activelydebate exactly what characters unite dinosaurs.
Over the past four decades, beginning with Bak-ker and Galton’s (1974) paper, approximately 50characters have been identified as potential dino-saur synapomorphies. Many of these have emergedfrom detailed, higher-level cladistic analyses ofarchosaur phylogeny (Benton, 1984, 1999, 2004;Gauthier, 1986; Benton and Clark, 1988; Novas,1989, 1992, 1996; Sereno, 1991a, 1999; Sereno andNovas, 1994; Fraser et al., 2002; Ezcurra, 2006,2010a; Langer and Benton, 2006; Irmis et al.,2007a; Nesbitt et al., 2009b, 2010; Brusatte et al.,2010a; Martinez et al., 2011; Nesbitt, 2011).Of course, different phylogenies may imply differ-ent patterns of character evolution, and the exact
C H A R A C T E R I S T I C F E A T U R E S O F D I N O S A U R S 11

characters that diagnose Dinosauria often differdepending on the phylogeny being considered. Toavoid the risk of gettingmired in a tedious catalogueof different phylogenies, the discussion here usesthe recent phylogeny of Brusatte et al. (2010a) andthe review of dinosaur origins by Brusatte et al.(2010b) as guides. This phylogenetic context isgraphically shown in Fig. 1.11.
Taking at first a reductionist view, seven fea-tures are currently recognized as unequivocalsynapomorphies of Dinosauria. In other words,these characters are only known in true dinosaurs,and are absent even in the closest dinosaur cou-sins. These bona fide dinosaur hallmarks areknown from across the skeleton, and include thefollowing.
1 Temporal musculature that extends anteriorlyonto the skull roof. The mandibular adductors(temporal muscles) are among the fundamentalmuscles ofmastication in vertebrates: when theycontract they elevate the lower jaw, allowing themouth to close. Dinosaurs have an unusuallylarge and extensive set of mandibular adductormuscles, which expand anteriorly onto the top ofthe skull (see Holliday, 2009 for review). Al-though muscle tissue is rarely preserved in dino-saur fossils, the location and size of themandibular adductors can be deduced based onthe position and size of a smooth fossa on theskull roof, to which these muscles attached. Inmost reptiles, including most archosaurs andeven close dinosaur kin such as Silesaurus, the
fossa is restricted to the parietal bone, and is onlyexpressed as a narrow depression in front of thesupratemporal fenestra (one of the main diapsidskull openings, which will be described in moredetail below) (Dzik, 2003). In dinosaurs, however,the fossa extends further anteriorly onto thefrontal bone, and is a much deeper and morediscrete depression (Fig. 1.12A,B). This indicatesthat the mandibular adductor muscles were larg-er and more powerful in dinosaurs than in closerelatives, and probably implies that dinosaurshad a stronger bite than most other archosaurs.
2 Posterior process of the jugal bifurcates to artic-ulate with the quadratojugal. The jugal boneforms the lateral ‘‘cheek’’ region of the skullunderneath the eye and articulates posteriorlywith the quadratojugal bone. Together these twobones define the ventral margin of the lateraltemporal fenestra, the second of the two maindiapsid skull openings. In all archosaurs otherthan dinosaurs, including Silesaurus, the poste-rior process of the jugal tapers and meets thequadratojugal at a simple overlapping joint (Dzikand Sulej, 2007). In dinosaurs, by contrast, theposterior process bifurcates into two prongs,which clasp the anterior process of the quadra-tojugal (Fig. 1.12C,D). The biological significanceof these two conditions is uncertain, but it islikely that dinosaurs had a stronger jugal–qua-dratojugal articulation, and thismay be function-ally associated with their larger mandibularadductor musculature and inferred strongerbite force.
Ave
met
atar
salia
Dinosauria
Dinosauromorpha
Arc
hosa
uria
Basal Archosauromorphs
CRUROTARSI
ScleromochlusPTEROSAURIA
LagerpetonDromomeron
Marasuchus
PseudolagosuchusLewisuchus
Sacisaurus
EucoelophysisSilesaurusSAURISCHIA
ORNITHISCHIA
Figure 1.11 The genealogical relationships of ‘‘bird-line’’ archosaurs (Avemetatarsalia) based on the phylogenetic
analysis of Brusatte et al. (2010a).
12 A N I N T R O D U C T I O N T O D I N O S A U R S

Figure 1.12 Distinctive features of dinosaurs. (A, B) Portions of the skulls of two theropod dinosaurs in dorsal view
(Dubreuillosaurus and Guanlong) showing the anterior extension of the fossa for the temporal jaw muscles onto the
frontal. (C, D) The bifurcated posterior process of the jugal, for articulationwith the quadratojugal (jugal and quadratojugal
of the theropod Allosaurus shown in articulation in C, only the jugal of the tyrannosaurid theropod Alioramus shown in D).
(E, F) The epipophysis, a bump-like projection of bone on the dorsal surface of the postzygapophysis of the cervical
vertebrae of the large theropod Aerosteon (E) and the tyrannosaurid Alioramus (F). Photographs (D) and (F) by Mick
Ellison; image (E) courtesy of Dr Roger Benson.
C H A R A C T E R I S T I C F E A T U R E S O F D I N O S A U R S 13
3 Epipophyses on the cervical vertebrae. Epipo-physes are projections of bone, which range fromsmall mounds to more elaborate flanges, thatprotrude from the dorsal surfaces of the postzy-gapophyses of the cervical vertebrae (those partsof the vertebra that articulate with the followingvertebra) (Fig. 1.12E,F). These are present in alldinosaurs, but not close relatives such as Mara-suchus (Sereno and Arcucci, 1994) or Silesaurus(Dzik, 2003; Piechowski andDzik, 2010). Variousmuscles of the neckwould have attached to thesestructures, as well as some muscles that mayhave extended onto the back and thorax (Tsuihiji,2005; Snively and Russell, 2007a, 2007b). Theprimary function of these muscles is to extend,rotate, and reinforce the neck and back.Althoughthese muscles would have been present in otherarchosaurs, the epipophyses in dinosaurs wouldhave increased their available attachment area,perhaps indicating that these muscles werestronger or capable of a greater range of motion(see Snively and Russell 2007a, 2007b for func-tional considerations).
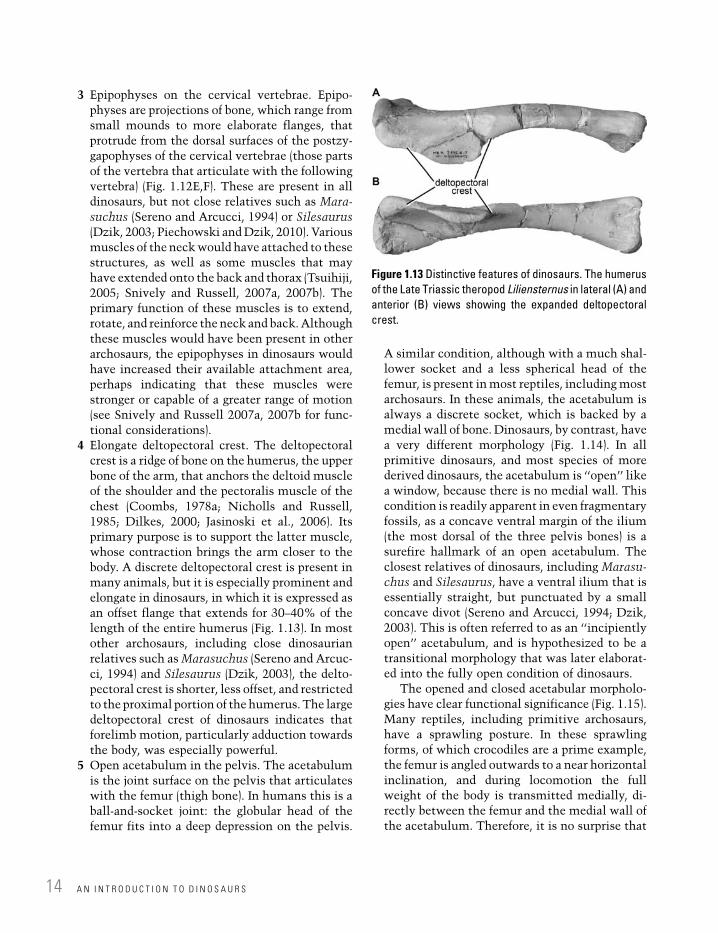
4 Elongate deltopectoral crest. The deltopectoralcrest is a ridge of bone on the humerus, the upperbone of the arm, that anchors the deltoid muscleof the shoulder and the pectoralis muscle of thechest (Coombs, 1978a; Nicholls and Russell,1985; Dilkes, 2000; Jasinoski et al., 2006). Itsprimary purpose is to support the latter muscle,whose contraction brings the arm closer to thebody. A discrete deltopectoral crest is present inmany animals, but it is especially prominent andelongate in dinosaurs, in which it is expressed asan offset flange that extends for 30–40% of thelength of the entire humerus (Fig. 1.13). In mostother archosaurs, including close dinosaurianrelatives such asMarasuchus (Sereno and Arcuc-ci, 1994) and Silesaurus (Dzik, 2003), the delto-pectoral crest is shorter, less offset, and restrictedto the proximal portion of the humerus. The largedeltopectoral crest of dinosaurs indicates thatforelimb motion, particularly adduction towardsthe body, was especially powerful.
5 Open acetabulum in the pelvis. The acetabulumis the joint surface on the pelvis that articulateswith the femur (thigh bone). In humans this is aball-and-socket joint: the globular head of thefemur fits into a deep depression on the pelvis.
A similar condition, although with a much shal-lower socket and a less spherical head of thefemur, is present inmost reptiles, includingmostarchosaurs. In these animals, the acetabulum isalways a discrete socket, which is backed by amedial wall of bone. Dinosaurs, by contrast, havea very different morphology (Fig. 1.14). In allprimitive dinosaurs, and most species of morederived dinosaurs, the acetabulum is ‘‘open’’ likea window, because there is no medial wall. Thiscondition is readily apparent in even fragmentaryfossils, as a concave ventral margin of the ilium(the most dorsal of the three pelvis bones) is asurefire hallmark of an open acetabulum. Theclosest relatives of dinosaurs, including Marasu-chus and Silesaurus, have a ventral ilium that isessentially straight, but punctuated by a smallconcave divot (Sereno and Arcucci, 1994; Dzik,2003). This is often referred to as an ‘‘incipientlyopen’’ acetabulum, and is hypothesized to be atransitional morphology that was later elaborat-ed into the fully open condition of dinosaurs.
The opened and closed acetabular morpholo-gies have clear functional significance (Fig. 1.15).Many reptiles, including primitive archosaurs,have a sprawling posture. In these sprawlingforms, of which crocodiles are a prime example,the femur is angled outwards to a near horizontalinclination, and during locomotion the fullweight of the body is transmitted medially, di-rectly between the femur and the medial wall ofthe acetabulum. Therefore, it is no surprise that
Figure 1.13 Distinctive features of dinosaurs. The humerus
of the Late Triassic theropod Liliensternus in lateral (A) and
anterior (B) views showing the expanded deltopectoral
crest.
14 A N I N T R O D U C T I O N T O D I N O S A U R S











![Video Transcript - Paleobiology - Unearthing Fossil Whales · Video Transcript - Paleobiology - Unearthing Fossil Whales Maggy Benson: [00:00:30] A special group of scientists called](https://static.fdocuments.us/doc/165x107/5fcc5e3cc244bb291a3b60e4/video-transcript-paleobiology-unearthing-fossil-whales-video-transcript-paleobiology.jpg)






