
Cytoplasmic Immunoreactivity of Thyroid Transcription ... · yielded a 300-base-pair (bp) product...
7
382 Am J Clin Pathol 2007;128:382-388 382 DOI: 10.1309/CADCVWHR2QF6JMVN © American Society for Clinical Pathology Anatomic Pathology / CYTOPLASMIC STAINING OF TTF-1 IN LIVER Cytoplasmic Immunoreactivity of Thyroid Transcription Factor-1 (Clone 8G7G3/1) in Hepatocytes True Positivity or Cross-Reaction? Keni Gu, MD, PhD, 1,4 Veena Shah, MD, 1 Chan Ma, MD, 1 Lixin Zhang, PhD, 3 and Maozhou Yang, PhD 2 Key Words: Thyroid transcription factor-1; TTF-1; Gene structure; Gene expression; Hepatocytes; Cytoplasmic immunoreactivity DOI: 10.1309/CADCVWHR2QF6JMVN Abstract The nuclear immunoreactivity for thyroid transcription factor-1 (TTF-1) is a useful marker for identification of carcinomas of thyroid and lung origin. Our aim was to determine whether cytoplasmic staining in the liver is a result of cross-reaction of anti–TTF-1 antibody (clone 8G7G3/1, DAKO, Carpinteria, CA) or true positivity resulting from aberrant expression of TTF-1 or products of the alternatively sliced TTF-1 gene. Fresh tissue samples from liver, thyroid, and lung were obtained for H&E-stained sections, TTF-1 immunostaining, and RNA and protein analyses. Western blot revealed an abundant band corresponding to an approximately 160-kd protein from liver but not either thyroid or lung tissue samples. By reverse transcriptase–polymerase chain reaction, messenger RNA of TTF-1 was not detectable in liver tissue. Our study demonstrates that TTF-1 immunoreactivity (clone 8G7G3/1) in the hepatocyte cytoplasm is due to an approximately 160-kd protein; this unique protein is not an alternative splicing product of TTF-1 and neither is it expressed in thyroid and lung tissues. Thyroid transcription factor-1 (TTF-1), a homeodomain containing DNA-binding protein, was first described in rat thyroid follicular cells as a regulator of thyroid-specific genes. 1-3 Subsequently, it was identified in lung (type II cells and Clara cells) and ventral forebrain. 4,5 TTF-1 is a member of the NKx2 family of homeodomain transcription factors and is also called NKx2.1 and TITF1. Human TTF-1 (locus, NC_000014) is a single polypep- tide 371 amino acids long ❚Figure 1❚; the gene has been mapped to chromosome 14q13. This gene contains 2 exons that produce an approximately 2.4-kb messenger RNA (mRNA) transcript coding for an approximately 41-kd pro- tein. The mouse TTF-1 gene contains 3 exons producing 2 iso- forms; the predominant transcript includes exon 2 and exon 3 coding for a 371–amino acid protein, whereas a minor tran- script includes all 3 exons that encode a 401–amino acid iso- form of TTF-1. 6,7 TTF-1 is a useful immunohistochemical marker for carci- nomas of thyroid and lung origin. 8-11 Normal hepatocytes and hepatocellular carcinoma often display cytoplasmic TTF-1 immunoreactivity (mainly from clone 8G7G3/1, DAKO, Carpinteria, CA). This immunoreactivity has been used in the differential diagnosis of primary and metastatic carcinomas of the liver. 12,13 TTF-1 cytoplasmic staining (clone 8G7G3/1), together with cytokeratin 19, might also serve as a useful marker for the differential diagnosis of hepatocellular carcino- ma from cholangiocarcinoma. 14 Our aim was to determine whether this cytoplasmic staining is a result of a cross-reaction with anti–TTF-1 anti- body (clone 8G7G3/1) or true positivity resulting from aber- rant expression of TTF-1 or products of alternative splicing of the TTF-1 gene.
Transcript of Cytoplasmic Immunoreactivity of Thyroid Transcription ... · yielded a 300-base-pair (bp) product...

382 Am J Clin Pathol 2007;128:382-388382 DOI: 10.1309/CADCVWHR2QF6JMVN
© American Society for Clinical Pathology
Anatomic Pathology / CYTOPLASMIC STAINING OF TTF-1 IN LIVER
Cytoplasmic Immunoreactivity of Thyroid TranscriptionFactor-1 (Clone 8G7G3/1) in Hepatocytes
True Positivity or Cross-Reaction?
Keni Gu, MD, PhD,1,4 Veena Shah, MD,1 Chan Ma, MD,1 Lixin Zhang, PhD,3
and Maozhou Yang, PhD2
Key Words: Thyroid transcription factor-1; TTF-1; Gene structure; Gene expression; Hepatocytes; Cytoplasmic immunoreactivity
DOI: 10.1309/CADCVWHR2QF6JMVN
A b s t r a c t
The nuclear immunoreactivity for thyroidtranscription factor-1 (TTF-1) is a useful marker foridentification of carcinomas of thyroid and lung origin.Our aim was to determine whether cytoplasmic stainingin the liver is a result of cross-reaction of anti–TTF-1antibody (clone 8G7G3/1, DAKO, Carpinteria, CA) ortrue positivity resulting from aberrant expression ofTTF-1 or products of the alternatively sliced TTF-1gene. Fresh tissue samples from liver, thyroid, and lung were obtained for H&E-stained sections, TTF-1immunostaining, and RNA and protein analyses.Western blot revealed an abundant band correspondingto an approximately 160-kd protein from liver but noteither thyroid or lung tissue samples. By reversetranscriptase–polymerase chain reaction, messengerRNA of TTF-1 was not detectable in liver tissue. Ourstudy demonstrates that TTF-1 immunoreactivity(clone 8G7G3/1) in the hepatocyte cytoplasm is due toan approximately 160-kd protein; this unique protein isnot an alternative splicing product of TTF-1 and neitheris it expressed in thyroid and lung tissues.
Thyroid transcription factor-1 (TTF-1), a homeodomaincontaining DNA-binding protein, was first described in ratthyroid follicular cells as a regulator of thyroid-specificgenes.1-3 Subsequently, it was identified in lung (type II cellsand Clara cells) and ventral forebrain.4,5 TTF-1 is a member ofthe NKx2 family of homeodomain transcription factors and isalso called NKx2.1 and TITF1.
Human TTF-1 (locus, NC_000014) is a single polypep-tide 371 amino acids long ❚Figure 1❚; the gene has beenmapped to chromosome 14q13. This gene contains 2 exonsthat produce an approximately 2.4-kb messenger RNA(mRNA) transcript coding for an approximately 41-kd pro-tein. The mouse TTF-1 gene contains 3 exons producing 2 iso-forms; the predominant transcript includes exon 2 and exon 3coding for a 371–amino acid protein, whereas a minor tran-script includes all 3 exons that encode a 401–amino acid iso-form of TTF-1.6,7
TTF-1 is a useful immunohistochemical marker for carci-nomas of thyroid and lung origin.8-11 Normal hepatocytes andhepatocellular carcinoma often display cytoplasmic TTF-1immunoreactivity (mainly from clone 8G7G3/1, DAKO,Carpinteria, CA). This immunoreactivity has been used in thedifferential diagnosis of primary and metastatic carcinomas ofthe liver.12,13 TTF-1 cytoplasmic staining (clone 8G7G3/1),together with cytokeratin 19, might also serve as a usefulmarker for the differential diagnosis of hepatocellular carcino-ma from cholangiocarcinoma.14
Our aim was to determine whether this cytoplasmicstaining is a result of a cross-reaction with anti–TTF-1 anti-body (clone 8G7G3/1) or true positivity resulting from aber-rant expression of TTF-1 or products of alternative splicing ofthe TTF-1 gene.

Am J Clin Pathol 2007;128:382-388 383383 DOI: 10.1309/CADCVWHR2QF6JMVN 383
© American Society for Clinical Pathology
Anatomic Pathology / ORIGINAL ARTICLE
Materials and Methods
We obtained 6 cases with fresh tissue samples, 3 explant-ed livers (2 for cirrhosis and 1 for acute rejection), 1 liver froma lobectomy for metastatic colon adenocarcinoma, 1 thyroid(goiter), and 1 lung from a lobectomy for adenocarcinoma.Studies included immunohistochemical staining, RNA extrac-tion and reverse transcriptase–polymerase chain reaction (RT-PCR), and protein extraction and Western blot.
Immunohistochemical Stains
Immunohistochemical stains were performed from for-malin-fixed, paraffin-embedded tissue sections using a com-mercially available primary antibody (monoclonal mouseanti–TTF-1, clone 8G7G3/1) and a detection kit (DAKO) perthe manufacturer’s protocol. Briefly, 4-µm-thick sections weredeparaffinized, quenched with 3% hydrogen peroxide for 6minutes, and heated for 40 minutes at 95°C to 99°C in an anti-gen retrieval solution in a steamer. The slides were stained for60 minutes with primary antibodies (1:200 dilution) followedby a detection process (horseradish peroxidase–liquid DAB+System, DAKO).
RNA Extraction and RT-PCR
Total RNA was isolated from freshly obtained surgicalspecimens using TRIzol reagent (Invitrogen LifeTechnologies, Grand Island, NY). Briefly, 100 mg of tissuewas pulverized under liquid nitrogen; 1 mL of TRIzol reagentwas added, and the lysate was further homogenized for about5 minutes. Phase separation and RNA purification were thenperformed. Next, 2 µg of total RNA was primed with randomhexamers and reverse transcribed into complementary DNA(cDNA) per standard protocol. Only 1 µL of the diluted cDNAmixture was used for the subsequent PCR analysis.15
The TTF-1 mRNA level was measured by RT-PCR using3 pairs of primers (Figure 1). Also, a pair of primers for glyc-eraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA, a
housekeeping gene, was used as an internal control for verify-ing the quality and quantity of mRNA. The PCR primers forthe TTF-1 and GAPDH genes were designed using Oligo v4.1software (Molecular Biology Insights, Cascade, CO).
All PCR primer sequences used in the study for TTF-1cDNA and GADPH cDNA were as follows: P1 sense primer,5'-ATG TCG ATG AGT CCA AAG CAC-3'; P2 antisenseprimer, 5'-CGC TGT TCC TCA TGG TGT CC-3'; P3 senseprimer, 5'- CGC TTC CCC GCC ATC TCC-3'; P4 antisenseprimer, 5'-GGC CTG GCG CTT CAT TTT GTA G-3'; P5sense primer, 5'-CGA GCG GCA TGA ACA TGA G-3'; P6antisense primer, 5'-CTT GGC CTG GCG CTT CAT T-3';GAPDH sense primer, 5'-TGA AGG TCG GAG TCA ACGGAT TTG GT-3'; and GAPDH antisense primer, 5'-CAT GTGGGC CAT GAG GTC CAC CAC-3'.
PCR was initiated by adding 1 µL of the diluted cDNAinto a 19-µL master PCR reaction mixture per standard proto-col by using an annealing temperature of 63°C for all primers.To keep the PCR in exponential phase, RT-PCR for GAPDHmRNA was conducted for only 25 cycles. RT-PCR for TTF-1mRNA was performed for 28 to 35 cycles.
Protein Extraction and Western Blot
Surgical specimens were pulverized and homogenizedin cold radioimmunoprecipitation assay buffer per standardprotocol. The tissue lysates were centrifuged at 16,000g for30 minutes at 4°C. The total protein concentration wasdetermined, and 20 µg of total protein was analyzed onto an8% polyacrylamide gel. Precision Plus Protein Standardswere from Bio-Rad, Hercules, CA. The proteins from thegels were electrotransferred onto a polyvinylidene fluoridemembrane (Bio-Rad) using a semidry electrotransfer sys-tem (Bio-Rad). The membrane was then incubated with pri-mary antibody against β-actin (Sigma A5316, dilution1:5,000; Sigma, St Louis, MO) or primary antibody againstTTF-1 (clone 8G7G3/1, dilution 1:500) for 3 hours at roomtemperature. Afterward, the membranes were incubated with
5� 3�Gene structure
Functional domains
P1 P2
1 31 123 160 220 253 270 295 371
D1 D2 D3 D4
P3 P4
P5 P6
❚Figure 1❚ Gene structure and functional domain of human thyroid transcription factor-1 and positions of polymerase chainreaction (PCR) primers. Gene structure, open boxes, exon, untranslated region; solid boxes, exon, coding region; horizontal linebetween the 2 exons, intron of the gene; horizontal arrows, PCR primer positions. Functional domain, D1, N-terminal activationdomain; D2, homeodomain; D3, NK-specific domain; D4, C-terminal activation domain. Vertical arrows with numbers markamino acid positions.
384 Am J Clin Pathol 2007;128:382-388384 DOI: 10.1309/CADCVWHR2QF6JMVN
© American Society for Clinical Pathology
Gu et al / CYTOPLASMIC STAINING OF TTF-1 IN LIVER
A B
C D
E F

Am J Clin Pathol 2007;128:382-388 385385 DOI: 10.1309/CADCVWHR2QF6JMVN 385
© American Society for Clinical Pathology
Anatomic Pathology / ORIGINAL ARTICLE
peroxidase-conjugated goat antimouse IgG and were devel-oped with PIERCE SuperSignal West Pico Substrate(PIERCE, Rockford, IL).
Results
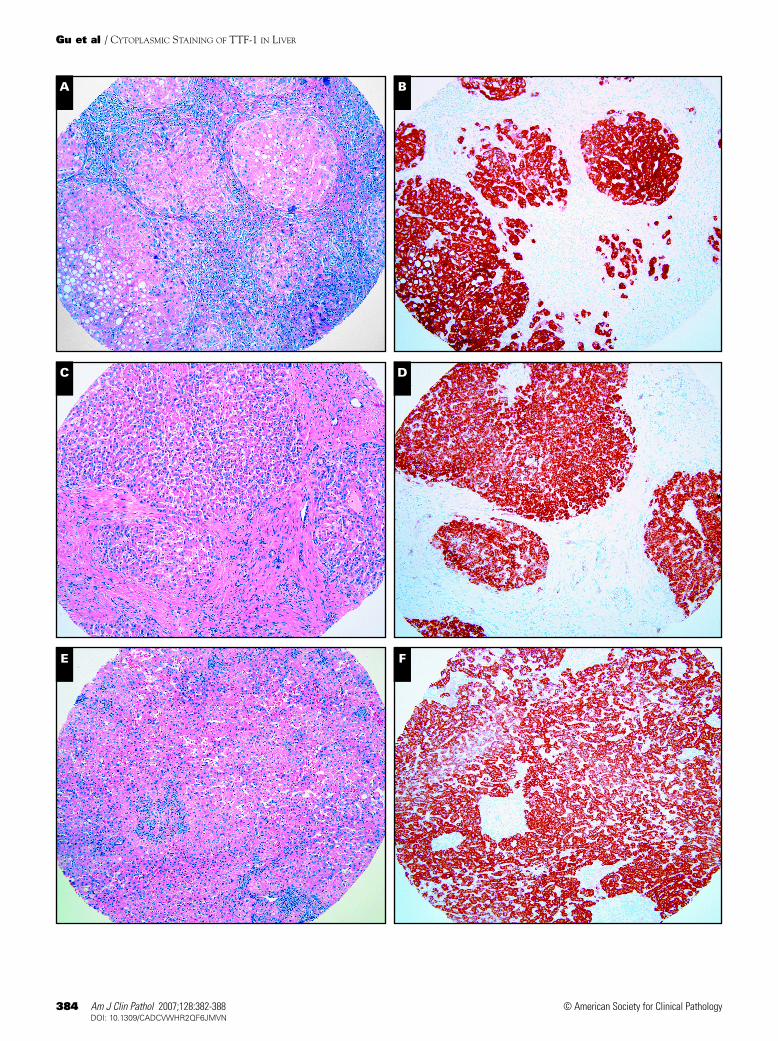
Strong and diffuse cytoplasmic immunoreactivity forTTF-1 (clone 8G7G3/1) was detected in hepatocytes of cir-rhotic liver ❚Image 1B❚ and ❚Image 1D❚ and noncirrhotic liverresected for metastatic adenocarcinoma of colon ❚Image 1F❚
(nontumor area). Weak cytoplasmic immunoreactivity forTTF-1 was detected in a liver explanted owing to severe rejec-tion ❚Image 1H❚. High magnification (×600) ❚Image 1I❚
revealed diffuse and granular cytoplasmic immunoreactivityfor TTF-1 (clone 8G7G3/1) in hepatocytes. Thyroid tissueshowed strong nuclear staining as a staining control sample
(data not shown). Western blot analysis using the same anti-body (clone 8G7G3/1) revealed a very prominent band corre-sponding to an approximately 160-kd protein from liver tissuebut not from thyroid ❚Image 2A❚ or nontumor lung tissue (datanot shown) samples. Thyroid tissue contained an approxi-mately 41-kd protein in Western blot (Image 2A). The primaryantibody, anti–β-actin, was also used as an internal control inWestern blot analysis to verify that the same amount of totalprotein was loaded in the gel (data not shown).
The mRNA of TTF-1 was not detected in liver tissuesamples by RT-PCR in up to 35 cycles using all 3 pairs ofprimers, whereas the positive control thyroid tissue sampleyielded a 300-base-pair (bp) product using primers P3 andP4, a 269-bp product using primers P5 and P6, and a 322-bpproduct using primers P1 and P2 ❚Image 2B❚. The expectedband (980-bp) of internal control GAPDH mRNA can be eas-ily amplified by RT-PCR from both thyroid and liver tissues
G H
I ❚Image 1❚ Thyroid transcription factor-1 (TTF-1) cytoplasmicimmunoreactivity (clone 8G7G3/1) in hepatocytes fromcirrhotic liver (A and C, H&E, ×20; B and D, TTF-1immunostain, ×20), noncirrhotic liver parenchyma (E, H&E,×20; F, TTF-1 immunostain, ×20); and liver explanted owing toacute rejection (G, H&E, ×20; H, TTF-1 immunostain, ×20). I,Diffuse and granular cytoplasmic immunoreactivity of TTF-1 inhepatocytes at high magnification (×600).

386 Am J Clin Pathol 2007;128:382-388386 DOI: 10.1309/CADCVWHR2QF6JMVN
© American Society for Clinical Pathology
Gu et al / CYTOPLASMIC STAINING OF TTF-1 IN LIVER
with similar intensity for only 25 cycles (Image 2B, lanes 2and 3, respectively).
Discussion
The incidence and significance of cytoplasmic TTF-1immunoreactivity in a variety of tumors have been investigat-ed. One study concluded that occasional cytoplasmic stainingfor TTF-1 (clone 8G7G3/1)in tumors was a nonspecific find-ing and should be disregarded for diagnostic purposes.16
However, other recent studies have indicated the usefulness ofcytoplasmic immunoreactivity for TTF-1 (clone 8G7G3/1) inthe differential diagnosis of primary and metastatic carcino-mas of the liver.12-14
The cytoplasmic staining of hepatocytes for TTF-1depends on the type of antibody used, and it varies from 0%(Novocastra, Newcastle upon Tyne, England) to up to 70%positivity (DAKO) (clone 8G7G3/1) in hepatocytes.17 Theantibody we used was from DAKO (clone 8G7G3/1), and ourfindings were consistent with the findings in the literature. Ourcases demonstrated strong and diffuse cytoplasmic staining incirrhotic and noncirrhotic liver parenchyma. The case withsevere acute cellular rejection revealed only weak staining.
The weak staining may result from severe injury of the hepa-tocytes by rejection. In contrast, there was completely nega-tive staining for TTF-1 in liver mesenchymal cells and inflam-matory cells (Image 1).
There are several possible explanations for the cytoplas-mic TTF-1 staining (clone 8G7G3/1) in hepatocytes. First, thestaining may be due to high level of endogenous biotin pres-ent in hepatocytes. The detection kit used for immunohisto-chemical staining used a refined avidin-biotin technique inwhich a biotinylated secondary antibody reacts with severalperoxidase-conjugated streptavidin molecules, raising thepossibility that the cytoplasmic TTF-1 staining could resultfrom a high level of endogenous biotin in hepatocytes. Thispossibility, however, is excluded because our Western blotanalysis that does not use the avidin-biotin technique unequiv-ocally demonstrated intensive staining corresponding to anapproximately 160-kd protein (Image 2A).
Second, cytoplasmic TTF-1 staining (clone 8G7G3/1)could come from aberrant expression of TTF-1. As a tran-scription factor, TTF-1 shows only nuclear staining in thyroid,lung, and ventral forebrain according to a considerable num-ber of previously published studies.2-5 After its phosphoryla-tion, the TTF-1 protein moves into cell nuclei and binds to atissue-specific promoter of downstream target genes such as
160 kd
(41 kd)
A B
❚Image 2❚ Western blot and reverse transcriptase–polymerase chain reaction: A, Western blot analysis using anti–thyroidtranscription factor-1 (TTF-1) antibody (clone 8G7G3/1). Lane 1, thyroid tissue; lane 2, liver tissue. Left, Precision Plus ProteinStandards (kd; Bio-Rad, Hercules, CA). B, Reverse transcriptase–polymerase chain reaction results. Lanes 2, 4, 6, and 9, thyroidtissue; lanes 3, 5, 7, and 10, liver tissue; lanes 2 and 3 using glyceraldehyde-3-phosphate dehydrogenase primers (980 base pairs[bp]); lanes 4 and 5, TTF-1 P3 and P4 primers (300 bp); lanes 6 and 7, TTF-1 P5 and P6 primers (269 bp); and lanes 9 and 10, TTF-1P1 and P2 primers (322 bp); lanes 1, 8, and 11, 100-bp DNA ladder.

Am J Clin Pathol 2007;128:382-388 387387 DOI: 10.1309/CADCVWHR2QF6JMVN 387
© American Society for Clinical Pathology
Anatomic Pathology / ORIGINAL ARTICLE
surfactant proteins and thyroglobulin, activating their expres-sions.18 Therefore, any nonnuclear staining of TTF-1 or stain-ing outside the aforementioned specific tissues should be con-sidered an aberrant expression of TTF-1 if the mRNA of TTF-1 is indeed transcribed. However, we were unable to detectTTF-1 mRNA in the liver tissue samples by RT-PCR using apair of primers (P3 and P4) that span an intron of the TTF-1gene (Image 2B, lanes 4 and 5). By using the aforementionedpair of primers, we were able to detect true TTF-1 mRNA,rather than DNA contamination, because the latter wouldshow a much larger PCR band if possible (TTF-1 intron, 964bp; locus, NC_000014).
Finally, cytoplasmic TTF-1 staining could result from across-reaction of anti–TTF-1 antibody (clone 8G7G3/1). OurWestern blot showed an approximately 160-kd protein fromliver tissue samples using anti–TTF-1 antibody (clone8G7G3/1). This protein is apparently much larger than theTTF-1 protein (41 kd) in terms of molecular weight.According to currently released information from NationalCenter for Biotechnology Information, the human TTF-1 genecontains only 2 exons, although a third exon, upstream of thecurrent TTF-1 gene, was reported in 1998,19 which would bevery similar to the gene structure of mouse TTF-1. The largerisoform of mouse TTF-1, which contains all 3 exons, is only30 amino acids longer than the predominant form (approxi-mately 41 kd), producing an approximately 46-kd protein atmost.6 Therefore, hepatocyte cytoplasmic TTF-1 stainingapparently results from a cross-reaction of anti–TTF-1 anti-body (clone 8G7G3/1) rather than gene expression of TTF-1.
To reach the preceding conclusion, however, we shouldexclude the possibility that either of the 2 currently identifiedhuman TTF-1 exons may alternately splice with other not yetidentified downstream or upstream exons, which produce ahigh-molecular-weight protein shown on Western blot.Therefore, we designed 2 additional pairs of PCR primers,with P1 and P2 deriving from the coding region of exon 1 andP5 and P6 from the coding region of exon 2 (Figure 1) to testthe possibility. We were still not able to detect any mRNAfrom liver tissue samples (Image 2B, lanes 6 and 7 and 9 and10), whereas the quality of RNA preparation was verified byan internal control, GAPDH.
Recently, a study showed the binding of TTF-1 (clone8G7G3/1) and hepatocytes paraffin (Hep Par) 1 to mitochon-drial proteins in hepatocytes.20 In that study, Western blotanalysis with cellular proteins extracted from normal humanliver demonstrated that Hep Par 1 and TTF-1 bound to a pro-tein band of approximately 150 kd. The authors speculatedthat different epitopes to Hep Par 1 and TTF-1 share the sameprotein molecules in hepatocyte mitochondria, and we com-pletely agree. In our Western blot analysis using the same anti-body, we showed binding to an approximately 160-kd protein;the discrepancy of estimated molecular weight between our
study and the aforementioned study20 is probably due to a lackof accurate protein markers around that region or to variationfrom different techniques.
TTF-1 immunoreactivity in hepatocyte cytoplasm is dueto an approximately 160-kd protein that reacts with anti–TTF-1 antibody (clone 8G7G3/1). This protein is not from TTF-1aberrant expression or an alternative slicing product of TTF-1in hepatocytes. It was not expressed in thyroid or lung tissuesamples in our study. The identity of this high-molecular-weight protein needs to be further studied.
From the 1Department of Pathology and 2Bone and Joint Center,Henry Ford Hospital, Detroit, MI; 3Department of Epidemiology,School of Public Health, University of Michigan, Ann Arbor; and4Department of Pathology, Medical College of Georgia, Augusta.
Address reprint requests to Dr Gu: Dept of Pathology, MedicalCollege of Georgia, 1120 15th St, BA-2576, Augusta, GA 30912.
References1. Guazzi S, Price M, De Felice M, et al. Thyroid nuclear factor 1
(TTF-1) contains a homeodomain and displays a novel DNAbinding specificity. EMBO J. 1990;9:3631-3639.
2. Musti AM, Ursini VM, Avvedimento EV, et al. A cell typespecific factor recognizes the rat thyroglobulin promoter.Nucleic Acids Res. 1987;15:8149-8166.
3. Sinclair AJ, Lonigro R, Civitareale D, et al. Thyroid specificgene expression. Adv Exp Med Biol. 1989;261:373-389.
4. Kimura S, Hara Y, Pineau T, et al. The T/ebp null mouse:thyroid-specific enhancer-binding protein is essential for theorganogenesis of the thyroid, lung, ventral forebrain, andpituitary. Genes Dev. 1996;10:60-69.
5. Lazzaro D, Price M, de Felice M, et al. The transcription factorTTF-1 is expressed at the onset of thyroid and lungmorphogenesis and in restricted regions of the foetal brain.Development. 1991;113:1093-1104.
6. Li C, Cai J, Pan Q, et al. Two functionally distinct forms ofNKX2.1 protein are expressed in the pulmonary epithelium.Biochem Biophys Res Commun. 2000;270:462-468.
7. Kolla V, Gonzales LW, Gonzales J, et al. Thyroid transcriptionfactor in differentiating type II cells: regulation, isoforms, andtarget genes. Am J Respir Cell Mol Biol. 2007;36:213-225.
8. Zhang P, Zuo H, Nakamura Y, et al. Immunohistochemicalanalysis of thyroid-specific transcription factors in thyroidtumors. Pathol Int. 2006;56:240-245.
9. Chang YL, Lee YC, Liao WY, et al. The utility and limitation ofthyroid transcription factor-1 protein in primary and metastaticpulmonary neoplasms. Lung Cancer. 2004;44:149-157.
10. Moldvay J, Jackel M, Bogos K, et al. The role of TTF-1 indifferentiating primary and metastatic lung adenocarcinomas.Pathol Oncol Res. 2004;10:85-88.
11. Nakamura N, Miyagi E, Murata S, et al. Expression of thyroidtranscription factor-1 in normal and neoplastic lung tissues.Mod Pathol. 2002;15:1058-1067.
12. Wieczorek TJ, Pinkus JL, Glickman JN, et al. Comparison of thyroid transcription factor-1 and hepatocyte antigenimmunohistochemical analysis in the differential diagnosis ofhepatocellular carcinoma, metastatic adenocarcinoma, renalcell carcinoma, and adrenal cortical carcinoma. Am J ClinPathol. 2002;118:911-921.

388 Am J Clin Pathol 2007;128:382-388388 DOI: 10.1309/CADCVWHR2QF6JMVN
© American Society for Clinical Pathology
Gu et al / CYTOPLASMIC STAINING OF TTF-1 IN LIVER
13. Gokden M, Shinde A. Recent immunohistochemical markersin the differential diagnosis of primary and metastaticcarcinomas of the liver. Diagn Cytopathol. 2005;33:166-172.
14. Lei JY, Bourne PA, diSant’Agnese PA, et al. Cytoplasmicstaining of TTF-1 in the differential diagnosis ofhepatocellular carcinoma vs cholangiocarcinoma andmetastatic carcinoma of the liver. Am J Clin Pathol.2006;125:519-525.
15. Yang M, Hayashi K, Hayashi M, et al. Cloning and develop-mental expression of a membrane-type matrix metalloproteinasefrom chicken. J Biol Chem. 1996;271:25548-25554.
16. Bejarano PA, Mousavi F. Incidence and significance ofcytoplasmic thyroid transcription factor-1 immunoreactivity.Arch Pathol Lab Med. 2003;127:193-195.
17. Pan CC, Chen PC, Tsay SH, et al. Cytoplasmic immuno-reactivity for thyroid transcription factor-1 in hepatocellularcarcinoma: a comparative immunohistochemical analysis of fourcommercial antibodies using a tissue array technique. Am JClin Pathol. 2004;121:343-349.
18. DeFelice M, Silberschmidt D, DiLauro R, et al. TTF-1phosphorylation is required for peripheral lung morphogenesis,perinatal survival, and tissue-specific gene expression. J BiolChem. 2003;278:35574-35583.
19. Hamdan H, Liu H, Li C, et al. Structure of the human Nkx2.1gene. Biochim Biophys Acta. 1998;1396:336-348.
20. Pang Y, von Turkovich M, Wu H, et al. The binding of thyroidtranscription factor-1 and hepatocyte paraffin 1 tomitochondrial proteins in hepatocytes: a molecular andimmunoelectron microscopic study. Am J Clin Pathol.2006;125:722-726.


















