
Copyright by Isagani Damasco Padolina 2002 · The Dissertation Committee for Isagani Damasco...
138
Copyright by Isagani Damasco Padolina 2002
Transcript of Copyright by Isagani Damasco Padolina 2002 · The Dissertation Committee for Isagani Damasco...

Copyright
by
Isagani Damasco Padolina
2002

The Dissertation Committee for Isagani Damasco Padolina
Certifies that this is the approved version of the following dissertation:
Chitin-Induced Biosynthesis of Phytoalexin 4´- Deoxyaurone
In Cell Suspension Cultures of “Old Man” Cactus,
Cephalocereus senilis
Committee:
Tom J. Mabry: Supervisor
Stanley J. Roux, Jr.
Alan M. Lloyd
Billie L. Turner
Ben A. Shoulders

Chitin-Induced Biosynthesis of Phytoalexin 4´- Deoxyaurone
In Cell Suspension Cultures of “Old Man” Cactus,
Cephalocereus senilis
by
Isagani Damasco Padolina, B.S.
Dissertation
Presented to the Faculty of the Graduate School of
The University of Texas at Austin
in Partial Fulfillment
of the Requirements
for the Degree of
Doctor of Philosophy
The University of Texas at Austin
May, 2002

Dedication
Para kay Nanay at Tatay, sa aking mga kapatid- kay Ibarra at Linglingay, at para
sa minamahal kong Anneke

Acknowledgements
I thank my supervising professor, Dr. Tom J. Mabry, for his patience, guidance
and support. I cannot think of anything better than the training he has given me as
his teaching assistant and graduate student. His no-nonsense approach to the
politics and nuances of research in graduate school will always be remembered. It
is both an honor and privilege to work with the “Father of Phytochemistry”.
Without Dr. Mabry, there would not be a dissertation, only a good idea.
I am grateful to Dr. Richard Dixon and Dr. Suzanne Rasmussen of the Samuel
Roberts Noble Foundation and Dr. Paul Paré of Texas Tech for their ideas on the
hypotheses that I am testing and for helping me design my pulse-chase
experiments. My special thanks go to Dr. Delia Brownson for helping me get
started in Dr. Mabry’s Phytochemistry lab. Her initial guidance saved me a lot of
time in finding where things are and how things work.
I would like to thank the other members of my dissertation committee: Drs.
Stanley Roux Jr., Alan Lloyd, Ben Shoulders and Billie Turner for reviewing this
dissertation as well as for their guidance during my graduate program.
My dissertation project was supported financially through Dr. Mabry’s F-0130
grant from the Robert A. Welch Foundation. I am grateful for the travel grants
v

and research fellowships awarded to me by the Molecular Cell and
Developmental Biology Department and the Plant Biology graduate program.
I am privileged to spend my many summers in the Mabry lab working with some
of the best phytochemists in world. I thank Drs. Nabil El-Sayed, Ahmed Ahmed,
Eman Haggag and Mahmound Nasser for helping me learn the biochemical
techniques of extraction, isolation and characterization of natural products.
Much of my time in graduate school is spent in the lab and I would like to thank
my fellow graduate students in the Mabry group- Schalk van Rooyen, Malgorzata
Wojcinska, and Jeff Williams, for the sharing of ideas and for the company of
helping each other.
Thanks to the laboratories and graduate students of Drs. Bob Jansen, Stan Roux,
Su Dharmawardhane, Malcolm Brown, James Mauseth and Jerry Brand for letting
me use their equipment and for lending me some of the chemicals that I needed. I
thank Dr. Greg Clark for the exchange of great ideas in doing good science.
The support staff of the Plant Biology program has been a great help in weaving
through the intricate logistics and bureaucracy of graduate school. I would like to
thank the staff of the former Botany department- Cathy Prescott, John Lucio,
Theresa Barnes, and Bill Baker, and the new Molecular Cell and Developmental
Biology staff, Tamra Rogers, Maureen Meko, Al Alvarado, Melody Villareal,
vi

Francisco Valledares and Adam Polanco. My special thanks go to Scott Schulz
for his assistance in doing my illustrations and figures.
My main source of support throughout my graduate career is my teaching
assistantship. My special thanks to Susan Cook of the School of Biological
Sciences for all her help in my TA assignments every semester.
My roommates through all these years in the Eiler house, Brian vanden Heuvel,
Jan Barber, Amy Bystrom and Mandi Vaughn have made living in Austin more
fun. I thank them for the dinners, the hikes, the rows and for their fantastic
company. It makes me sad to see them go.
The Beckman-Coulter HPLC team was instrumental (no pun intended) in helping
me set up my HPLC system for my experiments. Thanks to Leiko Dahlgren for
helping me with intricacies of the 171A radiodetector, to Marcia Witt for
troubleshooting software and to Don Tjernlund for all the plumbing lessons.
I wish to thank the numerous outstanding undergraduate students who did
research in the Mabry labs and have helped me so well in dealing with the
technical difficulties involved in my experiments. My special thanks go to the
students who showed up and went the extra mile- Sebastian Paige, Vaishali
Trivedi, Mitch Luna, Tanya Ong, Deepa Vyas, Kaye Charpentier; and to the
students of homestretch semester, Spring 2002- Bradley Eng, Manny Zapata,
vii

Amy Chen, Yenabi Keflemariam, Darrell Brown, Otis Tucker, Hannaise Cruz and
Sarah Rodgers.
My cello classes at the UT School of Music not only helped me in relieving some
of the stresses in grad school but has also made my education more well-rounded.
I wish to thank Professor Young for all her patience in her master classes when I
play out of tune and my many, many thanks goes to her TA, Tomasz Ryzezcki,
the guru of cello pedagogy, for his steady guidance and unrelenting patience for a
beginner like me.
My fellow graduate students, not just in the botany program but also in other
fields such as chemistry and zoology have been valuable for their company and
exchange of ideas, trials and tribulations and other goofy stuff- Rafe Brown,
Vaughan Symonds and Jen Tate, James and Grace Javier, Joey and Ruela
Pabalan, Shawn and Dylan Stricklin and Wowie Ebron. I am especially grateful to
the Tropa and Ari Bautista, CB and Tasha Mamaril, and Chris and Kristy Mateo
for their prayers and for their friendship.
I wish to thank my all of my friends from the bottom of my heart, my new friends
here in the US and old friends in the Philippines, you know who you are and you
viii

know me well, I’m hurrying up to finish right now so I don’t have time to put all
your names. You’ve seen me complain and ponder my way through the course of
my dissertation project and you’ve always managed to put a smile on my face
even when the rain pours.
Filipinos have big extended families they are always there for you no matter what.
I am grateful for the support that they have given- Auntie Nesy and Uncle Lito,
Auntie Bibing and Uncle Tante, the Casabar/Damasco clan in Fresno, Papa Bob
and Mama Wency, Ninang Nancy and Tita Lina, Auntie Gertrude, and my cousin
Marlo Casabar and his family.
My siblings whose significant experiences I have missed by being in a faraway
country. I wish to thank Ibarra and Linglingay for being there and for their quiet
support.
Without them I would not be here, literally, in every sense of the word. My Nanay
and Tatay, aside from being my parents, met and graduated at the University of
Texas. Without UT in the equation, definitely no Gani and most definitely no
Ph.D. No amount of words would be able to express my thanks to them for their
patience and support. I am proud of them, proud to be their son and I give them
my love.
ix

I would like to express my deepest appreciation to Anneke. She is the love of my
life and my closest friend. Her many sacrifices, understanding, patience, help and
support has made this scholarly journey more wonderful than I have imagined.
She has diluted the chemistry part of me with botany (not just some but lots of it),
she has balanced my diet with lots of vegetables, she has opened my mind to a lot
more than just science and she has filled my life with happiness.
Last, but far from least, I thank the Lord, Almighty and All Knowing. I am all but
human and I am very humbled by my accomplishments in the duration of this
work.
x

Chitin-Induced Biosynthesis of Phytoalexin 4´- Deoxyaurone In Cell Suspension Cultures of “Old Man” Cactus,
Cephalocereus senilis
Publication No._____________
Isagani Damasco Padolina, Ph.D.
The University of Texas at Austin, 2002
Supervisor: Tom J. Mabry
Cell suspension cultures of Cephalocereus senilis develop red
pigmentation when infected by fungus. Previous work in our laboratory has
established that chitin, a fungal cell wall component, can elicit the same response.
These elicited cell cultures also produce several unusual flavonoids lacking the
normal 4´-hydroxyl substitution in the B-ring, including the red pigment
cephalocerone, a 4´-deoxyaurone. Previous enzymological studies showed that the
elicited cell suspension cultures have the necessary enzymatic activities required
for the conversion of L-phenylalanine to the 4´-deoxyflavonoids. Explanations for
the 4´-deoxyflavonoid biosynthesis include (1) a bypass of cinnamic acid 4-
hydroxylase therefore leading to an accumulation of 4´-deoxy precursors
available for the 4´-deoxyflavonoid production and (2) a dehydroxylation step
within the flavonoid biosynthetic pathway.
xi

The current biochemical investigations into the biosynthesis of the
phytoalexin cephalocerone using HPLC metabolic profiling of isotope dilution
experiments established the existence of a novel 4´-dehydroxylation step instead
of a 4´-hydroxyl “bypass”. Our results strongly suggest that the metabolic flux
involved in the selective production of 4´-deoxyflavonoids may be controlled by a
specific enzyme activity that dehydroxylates the 4´-position of the
tetrahydroxychalcone B-ring.
xii

Table of Contents
List of Figures ...................................................................................................... xvi
Abbreviations ....................................................................................................... xix
Chapter 1. Introduction ........................................................................................... 1 A. General Comments on Secondary Plant Chemistry .................................. 1 B. The Phenylpropanoid Pathway.................................................................. 2 C. Previous Work on the Cephalocereus senilis Elicitor System .................. 4 D. Two Theories for the 4´-deoxyflavonoid Phenomenon ............................ 6
1. Enzyme Complexes and the “Bypass” Theory ................................. 6 2. A New Enzyme Activity for the 4´-Dehydroxylation Pathway ...... 10
E. Significance of the Current Study............................................................ 12 F. Outline of the Present Dissertation .......................................................... 13 Literature Cited ............................................................................................ 15
Chapter 2. Results: Novel Dehydroxylation Pathway in Elicited Suspension Cultures of Cephalocereus senilis................................................................ 35 A. Comments on the Test System: Chitin-Elicited C. senilis Cell
Suspension Cultures ............................................................................ 35 B. Results and Discussion ............................................................................ 36
1. Analysis of the Phytoalexin Red Pigment....................................... 37 2. Metabolic Fate of trans-Cinnamic Acid in “Old Man” Cactus
Cell Suspension Cultures ........................................................... 37 3. Metabolic Fate of p-Coumaric Acid in “Old Man” Cactus Cell
Suspension Cultures ................................................................... 41 4. The Proposed 4´-Deoxyflavonoid Bypass Theory .......................... 43 5. Evidence for the 4´-Dehydroxylation Step Leading to the
Biosynthesis of Cephalocerone .................................................. 46 6. Comments on the 4´-Deoxyflavonoid Phenomenon in
Cephalocereus senilis................................................................. 49
xiii

C. Experimental............................................................................................ 50 1. Plant Material .................................................................................. 50 2. Chemicals ........................................................................................ 51 3. Precusor Dilution Experiments ....................................................... 51 4. Separation of Phenolics by Reverse Phase HPLC .......................... 52 5. Liquid Chromatography Electrospray Ionisation Mass
Spectrometry (LC/ESI-MS) analysis ......................................... 53 6. Other Experimental Activities......................................................... 54
Literature Cited ............................................................................................ 55
Chapter 3. Conclusions and Summary .................................................................. 74 A. Studies Completed................................................................................... 74
1. Metabolic Profiling ......................................................................... 74 2. Evidence for the Novel Chitin- Elicited 4´-Dehydroxylation Step . 75
B. Future Research ....................................................................................... 77 1. Isolation of 4´-Deoxyflavonoid Standards ...................................... 77 2. Metabolic Profiling with Other Labeled Precursors ....................... 77 3. Dual-Labeling Experiments ............................................................ 78 5. 4´-Deoxy-Biosynthesis Enzymes .................................................... 79 6. 4´-Deoxyflavonoid Survey............................................................. 80 7. Molecular and Genetic Studies ....................................................... 80
Literature Cited ............................................................................................ 81
Appendix A. Phytoalexin biosynthesis ................................................................. 86 A. Elicitor Signal.......................................................................................... 86 B. Elicitor Release........................................................................................ 88 C. Signal Transduction................................................................................. 89 Literature Cited ............................................................................................ 91
xiv

Appendix B ........................................................................................................... 95
Appendix C ......................................................................................................... 107
Bibliography........................................................................................................ 109
Vita .................................................................................................................... 118
xv

List of Figures
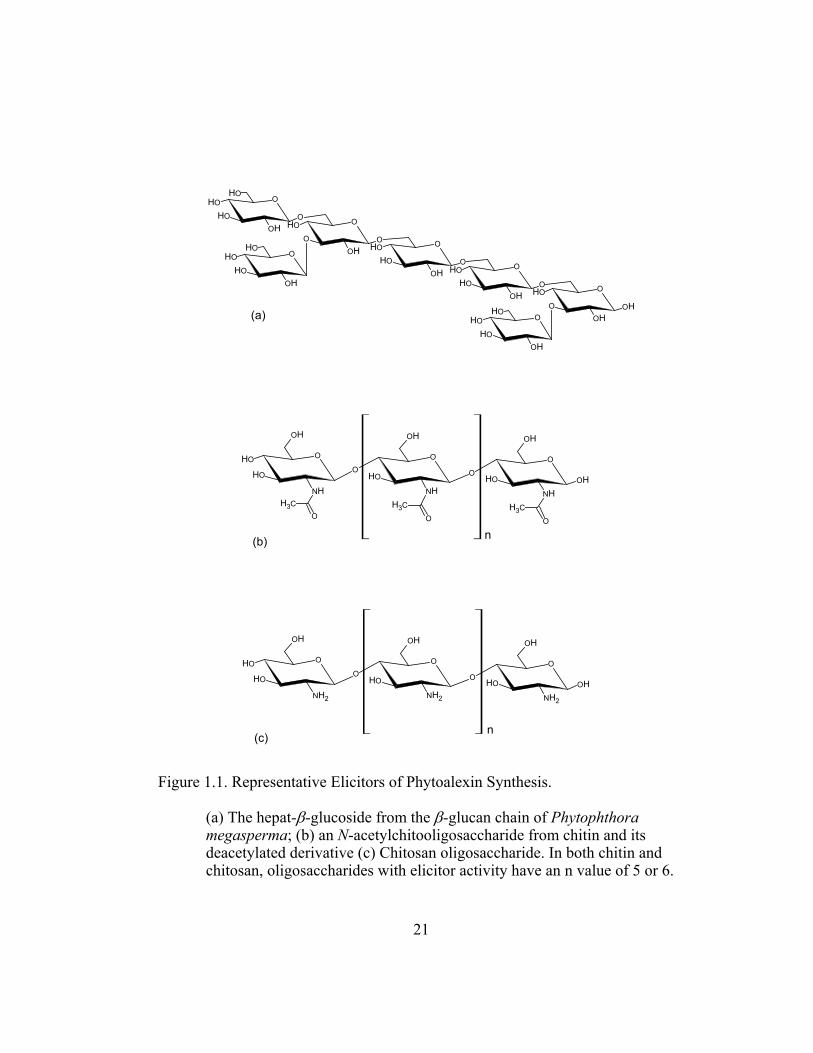
Figure 1.1. Representative Elicitors of Phytoalexin Synthesis. ............................ 21
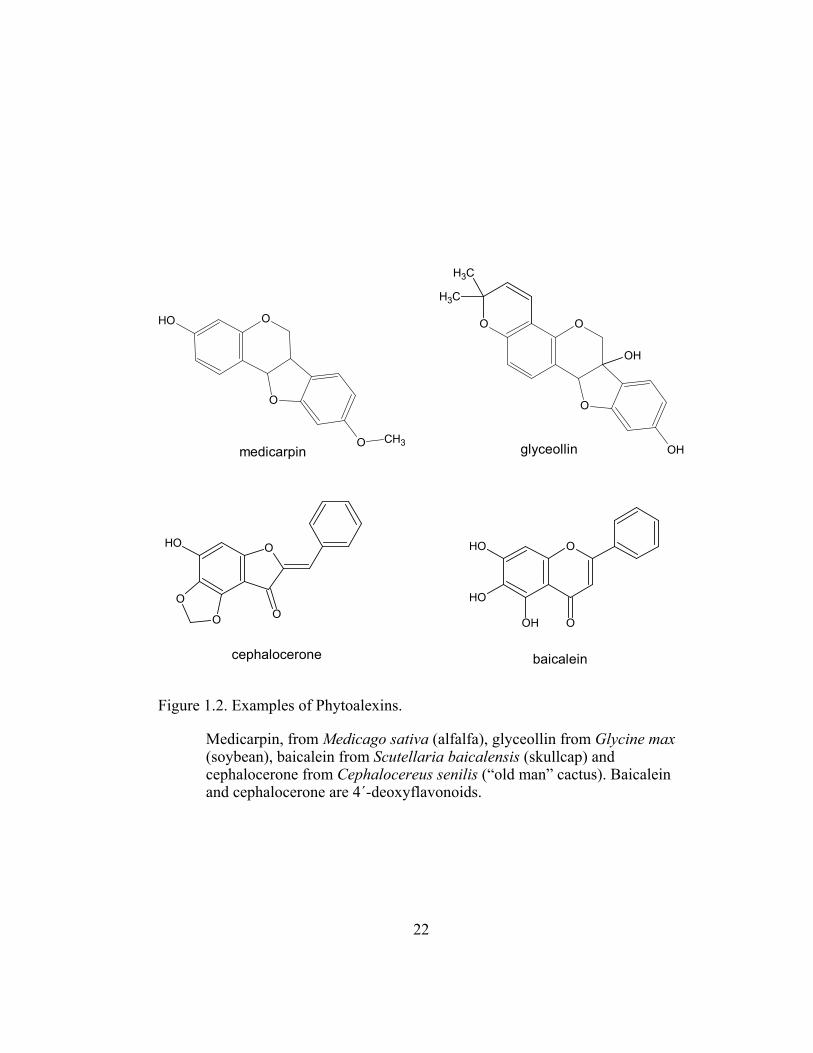
Figure 1.2. Examples of Phytoalexins................................................................... 22
Figure 1.3. “Old Man” Cactus (Cephalocereus senilis)........................................ 23
Figure 1.4. “Old Man” Cactus Callus and Cell Suspension Cultures. .................. 24
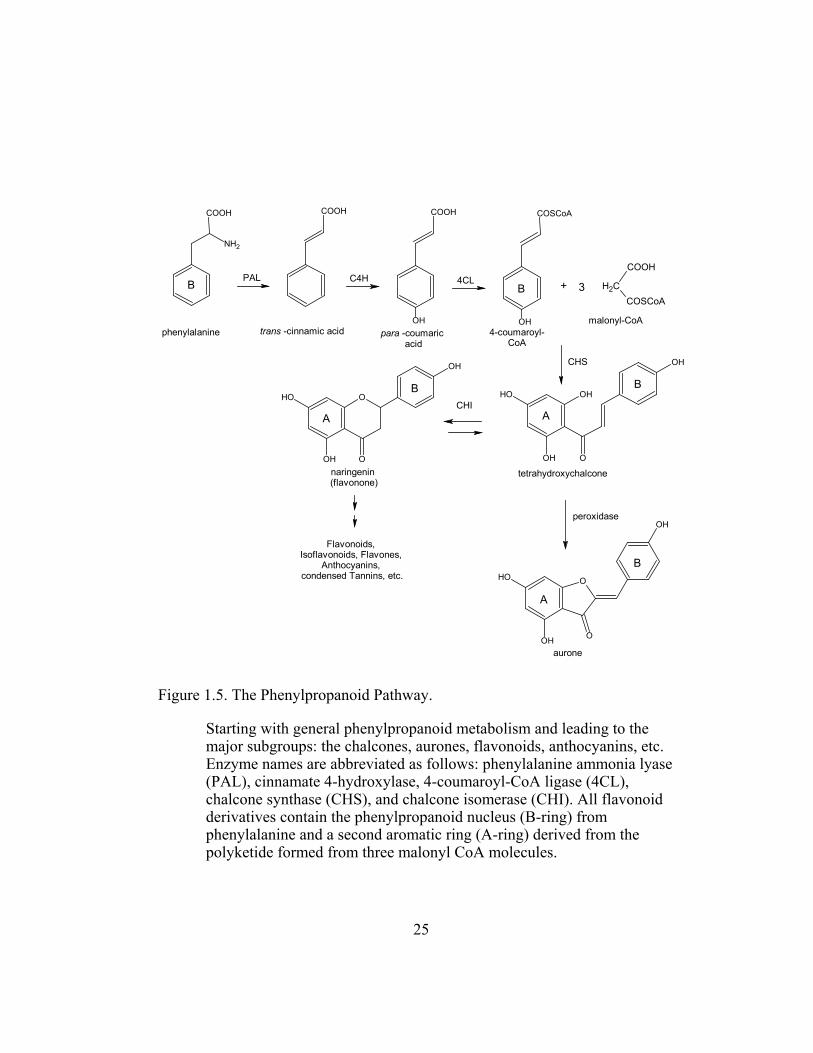
Figure 1.5. The Phenylpropanoid Pathway. .......................................................... 25
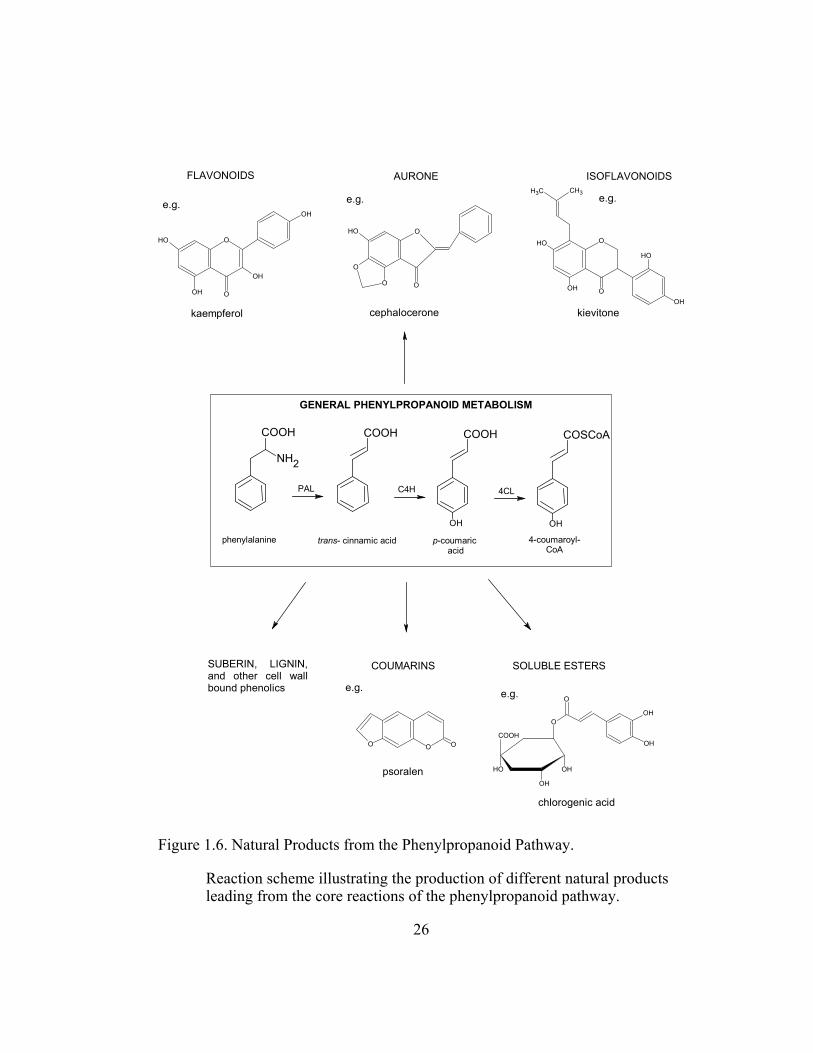
Figure 1.6. Natural Products from the Phenylpropanoid Pathway........................ 26
Figure 1.7. Mold-Infected Callus Cultures of “Old Man” Cactus. ....................... 27
Figure 1.8. Structures of Major 4´-Deoxyflavonoids from Elicited “Old Man”
Cactus Cell Suspension Cultures...................................................... 28
Figure 1.9. HPLC Profile of Radioactive Compounds Extracted from Elicited
“Old Man” Cactus Cultures.............................................................. 29
Figure 1.10. Metabolic “Bypass” of Cinammate 4-Hydroxylase.......................... 30
Figure 1.11. Numbering Convention Scheme for Phenylpropanoid
Derivatives. ...................................................................................... 31
Figure 1.12. Suggested Bypass Pathway in Elicited “Old Man” Cactus Cell
Suspension Cultures vs. Usual Phenylpropanoid Pathway. ............. 32
Figure 1.13. Proposed Dehydroxylation Pathway of 4´-Deoxyflavonoids in
Chitin-Elicited Cell Suspension Cultures of C. senilis. ................... 33
Figure 1.14. Biosynthetic Reactions Leading to Formation of Calycosin and
other 5-Deoxy(iso)flavonoids. ......................................................... 34
xvi

Figure 2.1. HPLC Profile of Radioactive Compounds.......................................... 59
Figure 2.2. HPLC Online Absorption Spectra of Cephalocerone. ........................ 60
Figure 2.3. HPLC Metabolic Profiles of “Old Man” Cactus Cell Suspension
Cultures. ........................................................................................... 61
Figure 2.4. HPLC Traces Showing Incorporation of Tritium from 3H-L-Phe
into Phenylpropanoid Compounds in Elicited “Old Man” Cactus
Cell Suspension Cultures. ................................................................ 62
Figure 2.5. Suggested Bypass Pathway in Elicited “Old Man” Cactus Cell
Suspension Cultures vs. Usual Phenylpropanoid pathway. ............. 63
Figure 2.6. Induction of 4CL Activity in “Old Man” Cactus Cell Cultures. ........ 64
Figure 2.7. In Vivo Labeling Experiments Demonstrate 3H-Isotope Dilution in
Cephalocerone Upon Addition of trans-Cinnamic Acid in
Elicited “Old Man” Cactus Cell Suspension Cultures. .................... 65
Figure 2.8. In Vivo Labeling Experiments Demonstrate 3H-Isotope Dilution on
Cephalocerone Upon Addition of Unlabeled trans-Cinnamic and
para-Coumaric Acids in Elicited “Old Man” Cactus Suspension
Cultures. ........................................................................................... 66
Figure 2.9. Amount of Cephalocerone in In Vivo Labeling Experiments............. 67
Figure 2.10. Primary and Secondary Pathways..................................................... 68
Figure 2.11. Proposed Dehydroxylation Pathway of 4´-Deoxyflavonoids in
Chitin-Elicited Cell Suspension Cultures of C. senilis. ................... 69
Figure 2.12. Compounds Produced from Dehydroxylation Biosynthesis............. 70
Figure 2.13. Tissue Culture Propagation of “Old Man” Cactus............................ 71
xvii

Figure 2.14. HPLC with Radioisotope Detector. .................................................. 72
Figure 2.15. Liquid Chromatography/Electrospray Ionisation- Mass
Spectrometry. ................................................................................... 73
Figure 3.1. The Proposed 4′-Dehydroxylation Pathway vs. the Unlikely C4H
Bypass Pathway................................................................................ 83
Figure 3.2. Synthesis Pathway of Lignin Monomers from Phenylpropanoid
Precursors. ........................................................................................ 84
Figure 3.3. Partial Structure of Spruce Lignin. ..................................................... 85
xviii

Abbreviations
4CL 4-coumarate CoA ligase 3H Tritium
3H-L-Phe Tritium-labeled L-phenylalanine 14C Carbon- 14
C4H or CA4H Cinnamic acid 4-hydroxylase
CHI Chalcone isomerase
CHS Chalcone synthase
Ci/mCi Curies/milliCuries
CPM Counts per minute
DCHI Deoxychalcone isomerase
DNA Deoxyribonucleic acid
DPM Disintegrations per minute
ER Endoplasmic reticulum
g fr wt gram fresh weight
Glc Glucose
HPLC High performance (or pressure) liquid chromatography
kat Katal, enyme activity effecting conversion of 1 mole
substrate per second
LC/ESI-MS Liquid chromatography/ electrospray ionization mass
spectrometry
mAu milliabsorbance units
xix

mRNA messenger ribonucleic acid
MS Murashige and Skoog
nm nanometers
NADPH Nicotinamide adenine dinucleotide phosphate (reduced
form)
PAL Phenylalanine ammonia lyase
pCA p-coumaric acid
PKR Polyketide reductase
rpm revolutions per minute
tCA trans-cinnamic acid
UV Ultraviolet
xx

Chapter 1
Introduction
A. GENERAL COMMENTS ON SECONDARY PLANT CHEMISTRY
Secondary plant metabolites play a major part in plants’ essential
biochemical processes with numerous roles in plant growth and development and
plant-pathogen interactions. These natural products, most of which are derived
from the phenylpropanoid, alkaloid, and isoprenoid pathways, are widespread
throughout the plant kingdom and account for much of its biochemical diversity
(Dixon, 1999). Much of the variation in plant secondary metabolism is under
genetic control (Dixon, 1998). The secondary metabolic pathways give a chemical
diversity in plants not produced by the more universal, uniformly organized and
controlled primary metabolism (Hartmann, 1996,1999). One of the more
interesting groups of natural products in this vast collection of secondary plant
compounds are the flavonoid derivatives that perform a variety of important
functions such as plant pigmentation, signal molecules in insect-plant interactions
and as plant defense compounds or phytoalexins. Phytoalexin biosynthesis in
plant-pathogen interactions is initiated by elicitor compounds produced by the
invading microorganism (see Appendix A for phytoalexin biosynthesis). The most
common of the biotic elicitors are the components of the pathogen that are
released exogenously; these may represent parts of the cell wall or metabolites
that secrete out of the cell wall (Hammerschmidt, 1999,1999; Smith, 1996).
1

Compounds belonging to the exogenous biotic elicitor class include
glycoproteins, lipids, and lipopolysaccharides, and the most studied group being
the poly- and oligosaccharides (Figure 1.1).
Different model systems have been developed to study the enzymology
and biochemistry of phytoalexin biosynthesis. For example, alfalfa (Medicago
sativa L.) cell suspension cultures, once treated with a fungal elicitor,
immediately produces high concentrations of the phytoalexin medicarpin (Figure
1.2), which is accompanied by an increase in extractable activities of all
phenylpropanoid enzymes needed for its synthesis (Dalkin et al., 1990; Ni et al.,
1996). In soybean (Glycine max L.) cells, this rapid phytoalexin response system
is characterized by the early and strong induction of PAL and CHS mRNAs upon
elicitation and the production of glyceollin (Habereder et al., 1989) (Figure 1.2).
The Mabry group, in collaboration with Dr. Richard Dixon’s lab at the Samuel
Roberts Noble Foundation, established a novel system for investigating
phenylpropanoid regulation and phytoalexin elicitation using cell cultures of the
cactus Cephalocereus senilis (Haworth) Pfeiffer (Figure 1.3 and 1.4).
B. THE PHENYLPROPANOID PATHWAY
The first enzyme of the phenylpropanoid pathway is phenylalanine
ammonia lyase (PAL), which catalyzes the deamination of phenylalanine to trans-
cinnamic acid, precursor of many phenolics and isoflavonoids (Figure 1.5). The
second step involves the hydroxylation of trans-cinnamic acid at C-4 to yield
trans-p-coumaric acid. This reaction is catalyzed by a cytochrome P450- mixed
2

function oxygenase, cinnamate 4-hydroxylase (C4H), with the aid of O2 and
NADPH as is common for such enzymes. It is trans-p-coumaric acid, with its 4-
hydroxyl group, that is the immediate precursor of lignin in all plants. The
enzyme 4-coumaroyl CoA ligase (4CL) uses trans-p-coumaric acid to produce 4-
coumaroyl CoA. This leads to the central reaction in flavonoid biosynthesis, the
formation of the C15 skeleton resulting from the combination of 4-coumaroyl CoA
(C9) with three molecules of malonyl CoA (3 x C2) catalyzed by chalcone
synthase (CHS). Thus all flavonoid derivatives contain the phenylpropanoid
nucleus (B-ring) from phenylalanine and a second aromatic ring (A-ring) derived
from the three malonyl CoA molecules (Figure 1.5). Further modification to other
flavonoid and isoflavonoid derivatives are then initiated by chalcone isomerase
followed by isoflavone synthase (Goodwin and Mercer, 1983). A vast array of
phenylalanine-derived intermediates such as flavonoids, isoflavonoids,
coumarins, soluble esters, suberin, lignin and other cell-wall bound phenolics are
produced via branching from the core reactions of general phenylpropanoid
metabolism (Hahlbrock and Scheel, 1989) (Figure 1.6).
The phenylpropanoid pathway is complex and dynamic, controlled by a
myriad of tightly coordinated signals at the transcriptional level (Dixon, 1998).
Control mechanisms such as exposure to light, wounding, plant growth regulators,
pathogens, and other biotic and abiotic factors are responsible for the induction of
phenylpropanoid enzyme activity in most plants (Bucciarelli et al., 1998; Dixon,
1999,2001).
3

C. PREVIOUS WORK ON THE CEPHALOCEREUS SENILIS ELICITOR SYSTEM
Cephalocereus senilis, commonly known as “old man” cactus (Family
Cactaceae, Tribe Pachycereae) is endemic to southern Mexico. Callus and
suspension cultures of Cephalocereus senilis were developed from inner stem
tissues of young plants (Bonness and Mabry, 1992). Cell suspension cultures of
C. senilis turn red (Figure 1.4 and 1.7) and accumulate a set of 4´-
deoxyflavonoids (Figure 1.8) with phytoalexin activity upon fungal elicitation or
treatment with chitin (Figure 1.1). A fungal species of Cladosporium and
autoclaved cultures of other fungi as well as yeast extracts also show elicitor
activity. Closely related oligomers of chitin such as chitosan turn the cultures red
while their respective monomers do not (Liu, 1994). The elicited compounds so
far identified were shown to be derived from L-[2,6-3H] phenylalanine as
determined by HPLC with a radioisotope detector (Paré et al., 1992) (Figure 1.9).
Elicitation was also correlated with increased activity of consecutive enzymes in
the phenylpropanoid pathway including PAL, C4H, 4CL, CHI, and CHS (Liu et
al., 1995; Paré et al., 1992).
Most of the phenolics isolated and characterized from “old man” cactus
elicited cell cultures lack the usual hydroxyl group at the 4´-position of the B-ring
(Liu et al., 1993) (Figure 1.6). In contrast, only flavonoids with the expected
hydroxylation pattern were isolated from healthy “old man” cactus plant material
(Liu et al., 1994). Cephalocerone, a 4´-deoxyaurone, was isolated from the
elicited cultures and was suggested to be responsible for the red pigmentation
(Paré et al., 1991) (Figure 1.2). Some of the isolated 4´-deoxyflavonoids,
4

including the red pigment aurone showed antibacterial activities in quantitative
microbial assays just as expected for induced phytoalexin compounds (Paré et al.,
1991). Preliminary qualitative studies have shown the same red pigmentation
response in whole C. senilis plants. The same response is also observed upon
initiation of callus cultures and wounding.
It has been shown that the elicited “old man” cactus cell suspension
cultures contain the necessary enzymatic activities required for the conversion of
L-phenylalanine to the B-ring deoxyflavonoids (Liu et al., 1995; Paré et al.,
1992). Further studies have determined that the enzymes do not show preferential
activity on the usual substrates or on the deoxy precursors (Liu et al., 1995). The
4´-deoxy phenomena may represent a form of metabolic compartmentalization
involving the close physical association of consecutive pathway enzymes
(Hrazdina and Jensen, 1992). The unique synthesis of B-ring deoxyflavonoids
(Figure 1.6) from elicited cultures of C. senilis with normal induction levels of
C4H activity suggests a form of metabolite channeling from PAL to 4CL that
involves bypassing microsomal C4H (Liu et al., 1995) (Figure 1.10). Another
alternative mechanism could involve the usual biosynthesis of 4´-
hydroxychalcones followed by the induction of a 4´-dehydroxylation activity
allowing the selective formation of 4´-deoxyflavonoids.
5

D. TWO THEORIES FOR THE 4´-DEOXYFLAVONOID PHENOMENON
1. Enzyme Complexes and the “Bypass” Theory
The validity of the “bypass” theory for the production of 4´-
deoxyflavonoids in chitin-elicited “old man” cactus cell cultures depends on the
formation of enzyme complexes in the phenylpropanoid pathway and the unusual
activity of these enzymes to 4´-deoxy substrates. A single form of CoA ligase
(4CL) was detected in the elicited cactus cell cultures. This enzyme showed
activity to a variety of substrates including p-coumaric, caffeic, ferulic, 4-
methoxycinnamic acids and, unexpectedly, it also showed activity to trans-
cinnamic acid (Liu et al., 1995). Chalcone synthase activity towards cinnamoyl
CoA has been detected in Scots pine (Pinus sylvestris L.) (Fliegmann et al., 1992)
and a similar CHS activity towards cinnamoyl CoA was observed in our cactus
cell cultures (Liu et al., 1995). Two isoforms of chalcone isomerase (CHI I and II)
were identified and both isoforms were able to activate 2´,4,4´,6´-
tetrahydroxychalcone (4´-hydroxy form) and 2´,4´,6´-trihydroxychalcone (4´-
deoxy form) as substrates (Liu et al., 1995) (chalcone conventional numbering is
used, i.e. the number primes actually refer to the chalcone A ring; Figure 1.11),
providing evidence that all the enzymes downstream of 4CL are capable of
activating any 4´-deoxy substrates produced if C4H is bypassed (Figure 1.12).
One explanation for the “bypass” pathway would be the lack of induction
of C4H in the elicited cell cultures. However, this is not the case, in fact, C4H is
strongly induced in the elicited cell cultures (Liu et al., 1995). Another possible
mechanism for the “bypass” of C4H is the metabolic channeling of substrates
6

from PAL to 4CL, probably facilitated by a close association of PAL and 4CL.
PAL and 4CL are cytosolic enzymes while C4H is a membrane-bound
cytochrome P450 enzyme (Czichi and Kindl, 1975,1977; O'Neal and Keller,
1970). A mechanism resulting in a substrate channel bypass of membrane- bound
C4H may be induced in chtin-elicited cultures. While substrate channeling of
trans-cinnamic acid usually occurs between PAL and C4H, it has been shown that
this channeling is disrupted in elicited tobacco cell cultures (Rasmussen and
Dixon, 1997,1999). The post-transcriptional organization of the phenylpropanoid
enzymes, depending on different environmental cues, may result in the formation
of different metabolic grids that would produce the necessary metabolic products
(Dixon et al., 2001).
The phenylpropanoid enzymes may be arranged in a linear fashion,
localized in the endoplasmic reticulum (ER) and anchored to the membrane via
cytochrome P450 enzymes such as C4H and flavanone 3´ hydroxylase (F3H)
(Hrazdina and Wagner, 1985; Shirley, 1999). This model is a result of labeling
studies performed on microsomes of potato (Czichi and Kindl, 1975,1975,1977)
and cucumber cotyledons (Hrazdina and Wagner, 1985) showing evidence that
PAL and C4H activities are co-localized in the ER membrane. These experiments
also show that exogenous cinnamic acid is less preferred as substrate for C4H
compared to the cinnamic acid already formed through PAL, suggesting a form of
metabolic channeling in the conversion of L-Phe to p-coumaric acid (Czichi and
Kindl, 1975; Hrazdina and Jensen, 1992; Hrazdina and Wagner, 1985). Also the
addition of more PAL in the microsomes only converts L-Phe to cinnamic acid
7

and does not contribute to any increase in p-coumaric acid in the overall reaction.
These biochemical studies laid a strong foundation for the metabolic channeling
model and were further strengthened by evidence from modern transgenic studies.
In Arabidopsis, almost all the enzymes of the general phenylpropanoid and central
flavonoid pathways are encoded by single copy genes except for PAL (encoded
by three genes) and flavonol synthase (encoded by five genes) (Shirley, 1999).
These suggest that the single copy gene enzymes are already arranged in specific
branched complexes that produce specific flavonoid derivatives and different
isoforms of PAL, for example, turn these arrangements on and off, depending on
the signals encountered by the plant (Shirley, 1999). Different isoforms of PAL
were isolated from tobacco microsomes and exhibit different levels of association
with the membrane bound C4H (Rasmussen and Dixon, 1999). It is suggested that
PAL isoforms can be deactivated through some form of phosphorylation carried
out by a phenylpropanoid protein kinase (Allwood et al., 1999).
In chitin-elicited “old man” cactus cell cultures, it could be that a PAL
isoform that associates less with C4H and more with 4CL is activated thereby
disrupting the channeling of trans-cinnamic acid for hydroxylation by C4H
(Figure 1.10 and 1.12). This creates a cytosolic pool of trans-cinnamic acid that
may be accessible to 4CL. Cinnamoyl-CoA (4´-deoxy) instead of coumaroyl-CoA
(4´-hydroxy) is then formed from the activation of trans-cinnamic acid by 4CL.
Cinnamoyl-CoA is then processed by CHS and CHI to initiate the formation of
4´-deoxyflavonoids.
8

Other mechanisms of differential control maybe arranged in each plant by
flavonoid enzymes that are multi-copy encoded for several isoforms. In alfalfa,
there are seven or more genes encoding for chalcone synthase (CHS) with the
lack of gene duplications for the enzymes thereafter, suggesting that different
isoforms of CHS could aid in controlling the flux of phenylpropanoid
intermediates into several flavonoid pathways (Dixon, 1998). The flavonoid
production process has been characterized to be more dynamic than linear, with
different branches switching to specific end products as needed by the plant
(Hartmann, 1996).
Metabolic channeling in the phenylpropanoid pathway was hypothesized
by Stafford (1974) and Hrazdina (1985) as a form of metabolic regulation
involving close association of enzymes to form enzyme complexes. Although
most studies are geared to characterize the regulation of transcriptional activation,
a growing body of evidence confirms the Hrazdina hypothesis and suggests that
the efficiency and plasticity of phenylpropanoid metabolism is spatially
coordinated by the presence of inducible membrane-bound enzyme complexes
(Allwood et al., 1999; Burbulis and Winkel-Shirley, 1999; Rasmussen and Dixon,
1997,1999; Shirley, 1996; Shirley, 1999; Spivey and Ovadi, 1999; Stafford,
1974). The use of transgenic plants in studying flavonoid biosynthesis has
contributed new methods in determining how the metabolites in the
phenylpropanoid pathway are being partitioned (Essenberg, 2001). Flavonoid
pathway research over the past 30 years, strongly supported by classical
9

biochemical techniques, has laid a solid foundation for the macromolecular
organization model (Hrazdina and Wagner, 1985; Shirley, 1999).
2. A New Enzyme Activity for the 4´-Dehydroxylation Pathway
The accumulation of 4´-deoxyflavonoids may not be due to metabolic
channeling as discussed above but by the elicitation of a specific enzyme activity
that catalyzes a 4´-dehydroxylation step (Figure 1.13). A novel dehydroxylase-
like enzyme, downstream of 4CL, could be induced by elicitation to remove the
4´-hydroxyl group, thereby allowing the selective formation of B-ring
deoxyflavonoids. A dehydroxylation step within the flavonoid pathway would
still be consistent with the observed heightened activity of the phenylpropanoid
enzymes in elicited “old man” cactus cultures (Liu et al., 1995; Paré et al., 1992).
This means that the phenylpropanoid biosynthesis proceeds as expected, up to the
production of the tetrahydroxychalcones. The tetrahydroxychalcones may then be
converted by a “dehydroxylase” to a trihydroxychalcone lacking a hydroxyl group
at the 4´-position of the B-ring. Although there is almost no known
dehydroxylation activity for the 4´-position, removal of the hydroxyl groups at
other positions in the A and B rings of flavonoids has been observed in various
plant systems (Goodwin and Mercer, 1983).
The basic hydroxylation pattern observed in chalcones is C-2´, 4´, 6´ for
the A-ring and C- 4 for the B ring (chalcone numbering; Figure 1.11) and the
same pattern is found in most flavonoids. Hydroxylation at the C-4-position of the
B-ring is almost universally indicative of the phenylpropanoid residue being
10

derived from p-coumaric acid. The hydroxylation pattern most frequently
encountered in ring A (C- 2´, 4´, 6´, chalcone numbering) is what is expected via
the polyketide formation from three molecules of malonyl-CoA (Goodwin and
Mercer, 1983). However, compounds with a missing hydroxyl substitution at C-5
position in the A-ring (flavone numbering; see Figure 1.11) are known to exist,
mostly in the isoflavones of Leguminosea; an example is calycosin from Baptisia
calycosa Canby (Kimura et al., 2001; Markham and Mabry, 1968; Markham et
al., 1968,1970; Markham et al., 1968) (Figure 1.14).
Previous genetic studies suggest that the dehydroxylation at the C-5
position (C-6´, chalcone numbering) probably occurs at the aromatic ring
formation at the polyketide stage or at the chalcone stage at the latest because the
presence of the step controlling the removal of the hydroxyl group results in all
flavonoids being equally affected (Goodwin and Mercer, 1983; Kimura et al.,
2001) (Figure 1.14). Flavonoid exudates in the roots are also known to be
dehydroxylated by Rhizobacteria at the 3 and 3´-position (Pillai and Swarup,
2002).
The occurrence of 4´-deoxyflavonoids in plants exists but is uncommon as
all flavonoids are known to be solely derived from p-coumaric acid in the
phenylpropanoid pathway (Goodwin and Mercer, 1983). Baicalein, a major 4´-
deoxyflavonoid from roots of a skullcap (Scutellaria baicalensis Georgi; Figure
1.2), is also found in other plants and is also one of the 4´-deoxyflavonoid
compounds isolated from elicited “old man” cactus cell cultures (Liu et al., 1993;
Rossi et al., 2001). Baicalein has been shown to exhibit biochemical activity
11

against certain types of cancer cells (Gao et al., 2001; Rossi et al., 2001), however
the details of baicalein biosynthesis in plants are not well established (Kuzovkina
et al., 2001).
E. SIGNIFICANCE OF THE CURRENT STUDY
The elicited cell suspension culture system of C. senilis is a well suited
model for studying regulation of metabolic flux within the phenylpropanoid and
flavonoid pathways, most especially in the biosynthesis of 4´-deoxyflavonoids.
The techniques developed and information gathered from this study are not only
relevant for the “old man” cactus metabolic system but for other metabolic
systems as well. The methods developed for metabolic profile screening and
isotope feeding experiments under various conditions can be applied in studying
similar systems such as the cinnamic acid derivation of salicylic acid in tobacco
(Nicotiana tabacum L.) (Leon et al., 1993; Raskin, 1992) and the biosynthesis of
baicalein (a 4´-deoxyflavonoid well studied for its biochemical activity against
certain types of cancer) in Scutellaria baicalensis (Chen et al., 2000; Kuzovkina
et al., 2001). The enzymatic events that control the biosynthesis of salicylic acid
and baicalein are yet to be determined.
The “old man” cactus 4´-deoxyflavonoid phenomenon represents a useful
model on how plants control their metabolism in general. This study may provide
evidence about how plants organize their metabolic grid in response to internal
and external signals. On a larger scale, the information derived from the
enzymology and organization of biosynthetic pathways and the molecular
12

“switches” that plants use to make their metabolic “choices” may lead to a better
understanding of how pathways can be engineered, especially in crop plants, to
increase flux into specific compounds for improved nutrition and plant defense.
Most of the research on metabolic compartmentation in flavonoid
biosynthesis has focused on crop plants such as alfalfa and tobacco (Dixon et al.,
1995; Rasmussen and Dixon, 1997,1999). Aside from their immediate economic
importance, it is also easy to perform transgenic studies on these crop plants
because of their physiological characteristics and short generation time.
Arabidopsis has also been a good source of information on flavonoid enzyme
complexes (Burbulis and Winkel-Shirley, 1999). However, little is known about
the “phytoalexin response” in desert-adapted plants such as “old man” cactus. The
details of the surprising formation of the B-ring-deoxyflavonoids (Paré et al.,
1991) remain unclarified and the relationship of this culture pathway to whole
plant pathways in both “old man” cactus and related species has not been
determined. The 4´-deoxy phenomenon is unusual in flavonoid formation as most
flavonoids contain the hydroxyl group at the 4´ -position in the B-ring in most
plant systems.
F. OUTLINE OF THE PRESENT DISSERTATION
The present dissertation includes the investigation of the molecular and
cellular processes involved in the production of 4´-deoxyflavonoids in chitin-
elicited cultures of “old man” cactus. This research is summarized as follows:
13

1). Cephalocerone, a phytoalexin-type 4´-deoxy aurone was isolated from
the elicited cultures and is responsible for the red pigmentation (Paré et al.,
1991). Metabolic profile studies of cell cultures in elicited “old man”
cactus were made to compare the observed pigment responses. Profile
analyses also provide evidence whether or not the 4´-deoxy phenomenon
in the “old man” cactus is a result of the flavonoid pathway induction in
general or whether such unusual flavonoids are a specific response to
chitin-elicitation and microbial infection.
2). Most of the investigations on the biosynthetic control of the
production of 4´-deoxyflavonoids in elicited “old man” cactus have dealt
with induction and substrate specificity of the phenylpropanoid enzymes.
Incorporation of radiolabeled L-phenylalanine and unlabeled precursors
cinnamic and coumaric acids in a pulse-chase experiment were conducted
to follow their levels of incorporation into the phenylpropanoid pathway
and to investigate the cellular and metabolic system that leads to B-ring
deoxyflavonoids. Metabolic profile studies of these isotope dilution
experiments allow the testing of the 4´-hydroxy “bypass” and the 4´-
dehydroxylation hypotheses.
14

LITERATURE CITED
Allwood, E. G., Davies, D. R., Gerrish, C., Ellis, B. E. and Bolwell, G. P., 1999.
Phosphorylation of phenylalanine ammonia-lyase: evidence for a novel protein kinase and identification of the phosphorylated residue. Febs Letters 457, 47-52.
Bonness, M. S. and Mabry, T. J., 1992. Tissue-culture of endod (Phytolacca dodecandra Lherit) - Growth and production of ribosome-inactivating proteins. Plant Cell Reports 11, 66-70.
Bucciarelli, B., Jung, H. G., Ostry, M. E., Anderson, N. A. and Vance, C. P., 1998. Wound response characteristics as related to phenylpropanoid enzyme activity and lignin deposition in resistant and susceptible Populus tremuloides inoculated with Entoleuca mammata (Hypoxylon mammatum). Canadian Journal of Botany-Revue Canadienne De Botanique 76, 1282-1289.
Burbulis, I. E. and Winkel-Shirley, B., 1999. Interactions among enzymes of the Arabidopsis flavonoid biosynthetic pathway. Proceedings of the National Academy of Sciences of the United States of America 96, 12929-12934.
Chen, C. H., Huang, L. L. H., Huang, C. C., Lin, C. C., Lee, Y. and Lu, F. J., 2000. Baicalein, a novel apoptotic agent for hepatoma cell lines: A potential medicine for hepatoma. Nutrition and Cancer-an International Journal 38, 287-295.
Czichi, U. and Kindl, H., 1975. Formation of p-coumaric acid and o-coumaric acid from L- phenylalanine by microsomal membrane-fractions from potato - evidence of membrane-bound enzyme complexes. Planta 125, 115-125.
Czichi, U. and Kindl, H., 1975. Model of closely assembled consecutive enzymes on membranes - formation of hydroxycinnamic acids from L-phenylalanine on thylakoids of Dunaliella marina. Hoppe-Seylers Zeitschrift Fur Physiologische Chemie 356, 475-485.
Czichi, U. and Kindl, H., 1977. Phenylalanine ammonia-lyase and cinnamic acid hydroxylases as assembled consecutive enzymes on microsomal-
15

membranes of cucumber cotyledons - cooperation and subcellular-distribution. Planta 134, 133-143.
Dalkin, K., Edwards, R., Edington, B. and Dixon, R. A., 1990. Stress responses in alfalfa (Medicago sativa L) .1. Induction of phenylpropanoid biosynthesis and hydrolytic enzymes in elicitor-treated cell-suspension cultures. Plant Physiology 92, 440-446.
Dixon, R. A., 1999. Plant natural products: the molecular genetic basis of biosynthetic diversity. Current Opinion in Biotechnology 10, 192-197.
Dixon, R. A., 2001. Natural products and plant disease resistance. Nature 411, 843-847.
Dixon, R. A., Chen, F., Guo, D. J. and Parvathi, K., 2001. The biosynthesis of monolignols: a "metabolic grid", or independent pathways to guaiacyl and syringyl units? Phytochemistry 57, 1069-1084.
Dixon, R. A., Harrison, M. J. and Paiva, N. L., 1995. The isoflavonoid phytoalexin pathway - from enzymes to genes to transcription factors. Physiologia Plantarum 93, 385-392.
Dixon, R. A., Howles P.A., Lamb C., He X.Z., and Reddy, J.T., 1998. Prospects for the metabolic engineering of bioactive flavonoids and related phenylpropanoid compounds. Advances in Experimental Medicine and Biology 439, 55-66.
Essenberg, M., 2001. Prospects for strengthening plant defenses through phytoalexin engineering. Physiological and Molecular Plant Pathology 59, 71-81.
Fliegmann, J., Schroder, G., Schanz, S., Britsch, L. and Schroder, J., 1992. Molecular analysis of chalcone and dihydropinosylvin synthase from scots pine (Pinus sylvestris), and differential regulation of these and related enzyme-activities in stressed plants. Plant Molecular Biology 18, 489-503.
Gao, Z. H., Huang, K. X. and Xu, H. B., 2001. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in HS-SY5Y cells. Pharmacological Research 43, 173-178.
Goodwin, T. W. and Mercer, E. I., 1983. Introduction to Plant Biochemistry. Pergamon Press, New York.
16

Habereder, H., Schroder, G. and Ebel, J., 1989. Rapid induction of phenylalanine ammonia-lyase and chalcone synthase messenger-Rnas during fungus infection of soybean (Glycine max L) roots or elicitor treatment of soybean cell- cultures at the onset of phytoalexin synthesis. Planta 177, 58-65.
Hahlbrock, K. and Scheel, D., 1989. Physiology and molecular-biology of phenylpropanoid metabolism. Annual Review of Plant Physiology and Plant Molecular Biology 40, 347-369.
Hammerschmidt, R., 1999. Induced disease resistance: how do induced plants stop pathogens? Physiological and Molecular Plant Pathology 55, 77-84.
Hammerschmidt, R., 1999. Phytoalexins: What have we learned after 60 years? Annual Review of Phytopathology 37, 285-306.
Hartmann, T., 1996. Diversity and variability of plant secondary metabolism: a mechanistic view. Entomologia Experimentalis et Applicata 80, 177-188.
Hartmann, T., 1999. Chemical ecology of pyrrolizidine alkaloids. Planta 207, 483-495.
Hrazdina, G. and Jensen, R. A., 1992). Compartmentation in aromatic metabolism.In: H. Stanford and R. K. Ibrahim (Eds), Phenolic Metabolism in Plants. Plenum, New York, pp. 1-23.
Hrazdina, G. and Wagner, G. J., 1985. Metabolic pathways as enzyme complexes: evidence for the synthesis of phenylpropanoids and flavonoids on membrane-associated enzyme complexes. Archives of Biochemistry and Biophysics 237, 88-100.
Kimura, Y., Aoki, T. and Ayabe, S., 2001. Chalcone isomerase isozymes with different substrate specificities towards 6 '-hydroxy- and 6 '-deoxychalcones in cultured cells of Glycyrrhiza echinata, a leguminous plant producing 5-deoxyflavonoids. Plant and Cell Physiology 42, 1169-1173.
Kuzovkina, I. N., Guseva, A. V., Alterman, I. E. and Karnachuk, R. A., 2001. Flavonoid production in transformed Scutellaria baicalensis roots and ways of its regulation. Russian Journal of Plant Physiology 48, 448-452.
17

Leon, J., Yalpani, N., Raskin, I. and Lawton, M. A., 1993. Induction of benzoic-acid 2-hydroxylase in virus-inoculated tobacco. Plant Physiology 103, 323-328.
Liu, Q. (1994). Chemical and enzymological investigations of the phenylpropanoid pathway in elicited cultures of Cephalocereus senilis ("Old man" cactus). Ph.D. Thesis. Botany. Austin, University of Texas at Austin: 1-15.
Liu, Q., Bonness, M. S., Liu, M., Seradge, E., Dixon, R. A. and Mabry, T. J., 1995. Enzymes of B-ring-deoxy flavonoid biosynthesis in elicited cell-cultures of old man cactus (Cephalocereus senilis). Archives of Biochemistry and Biophysics 321, 397-404.
Liu, Q., Liu, M., Mabry, T. J. and Dixon, R. A., 1994. Flavonol glycosides from Cephalocereus senilis. Phytochemistry 31, 229-331.
Liu, Q., Markham, K. R., Paré, P. W., Dixon, R. A. and Mabry, T. J., 1993. Flavonoids from elicitor-treated cell-suspension cultures of Cephalocereus senilis. Phytochemistry 32, 925-928.
Markham, K. R. and Mabry, T. J., 1968. The identification of twenty-three 5-deoxy- and ten 5-hydroxy-flavonoids from Baptisia lecontei (Leguminosae). Phytochemistry 7, 791-801.
Markham, K. R., Mabry, T. J. and Swift III, W. T., 1968. New isoflavones from the genus Baptisia (Leguminosae). Phytochemistry 7, 803-808.
Markham, K. R., Mabry, T. J. and Swift III, W. T., 1970. Distribution of flavonoids in the genus Baptisia (Leguminosae). Phytochemistry 9, 2359-2364.
Markham, K. R., Swift III, W. T. and Mabry, T. J., 1968. A new isoflavone glycoside from Baptisia australis. Journal of Organic Chemistry 33, 462-4.
Ni, W. T., Fahrendorf, T., Ballance, G. M., Lamb, C. J. and Dixon, R. A., 1996. Stress responses in alfalfa (Medicago sativa L) .20. Transcriptional activation of phenylpropanoid pathway genes in elicitor-induced cell suspension cultures. Plant Molecular Biology 30, 427-438.
18

O'Neal, D. and Keller, C. J., 1970. Partial purification and some properties of phenylalanine ammonia-lyase of tobacco (Nicotiana tabacum). Phytochemistry 9, 1373-1383.
Paré, P. W., Dmitrieva, N. and Mabry, T. J., 1991. Phytoalexin aurone induced in Cephalocereus senilis liquid suspension-culture. Phytochemistry 30, 1133-1135.
Paré, P. W., Mischke, C. F., Edwards, R., Dixon, R. A., Norman, H. A. and Mabry, T. J., 1992. Induction of phenylpropanoid pathway enzymes in elicitor- treated cultures of Cephalocereus senilis. Phytochemistry 31, 149-153.
Pillai, B. V. and Swarup, S., 2002. Elucidation of the flavonoid catabolism pathway in Pseudomonas putida PML2 by comparative metabolic profiling. Applied and Environmental Microbiology 68, 143-151.
Raskin, I., 1992. Salicylate, a new plant hormone. Plant Physiology 99, 799-803.
Rasmussen, S. and Dixon, R. A., 1997. Evidence for a microsomal enzyme complex of PAL and CA4H in tobacco plants. Plant Physiology 114, 1568-1568.
Rasmussen, S. and Dixon, R. A., 1999. Transgene-mediated and elicitor-induced perturbation of metabolic channeling at the entry point into the phenylpropanoid pathway. Plant Cell 11, 1537-1551.
Rossi, M., Meyer, R., Constantinou, P., Caruso, F., Castelbuono, D., O'Brien, M. and Narasimhan, V., 2001. Molecular structure and activity toward DNA of baicalein, a flavone constituent of the Asian herbal medicine "Sho-saiko-to". Journal of Natural Products 64, 26-31.
Shirley, B. W., 1996. Flavonoid biosynthesis: 'New' functions for an 'old' pathway. Trends in Plant Science 1, 377-382.
Shirley, B. W., 1999. Evidence for enzyme complexes in the phenylpropanoid and flavonoid pathways. Physiologia Plantarum 107, 142-149.
Smith, C. J., 1996. Accumulation of phytoalexins: Defence mechanism and stimulus response system. New Phytologist 132, 1-45.
Spivey, H. O. and Ovadi, J., 1999. Substrate channeling. methods- a Companion to Methods in Enzymology 19, 306-321.
19

Stafford, H. A., 1974. Possible multi-enzyme complexes regulating the formation of C6-C3 phenolic compounds and lignins in higher plants. Recent Advances in Phytochemistry 8, 53-79.
20
O
OHOOH
OHOH
O
OHOO
OH
O
OHOOH
OH
O
OHOOH
OH
O
OHOHO
OH
O
OHOH
OHOH
O
OHOH
OHOH
O
NH
OOH
OH
OH
O
NH
OOH
OH
O
NHOHOH
OH
OCH3 CH3 CH3
O O
n
O
NH2
OOH
OH
OH
O
NH2
OOH
OH
O
NH2
OHOH
OH
n
(a)
(b)
(c)
Figure 1.1. Representative Elicitors of Phytoalexin Synthesis.
(a) The hepat-β-glucoside from the β-glucan chain of Phytophthora megasperma; (b) an N-acetylchitooligosaccharide from chitin and its deacetylated derivative (c) Chitosan oligosaccharide. In both chitin and chitosan, oligosaccharides with elicitor activity have an n value of 5 or 6.
21
O
O
OH
O CH3
O
OO
O
OH
medicarpin
cephalocerone
O
O
O
OH
OH
CH3
CH3
glyceollin
baicalein
O
OOH
OH
OH
Figure 1.2. Examples of Phytoalexins.
Medicarpin, from Medicago sativa (alfalfa), glyceollin from Glycine max (soybean), baicalein from Scutellaria baicalensis (skullcap) and cephalocerone from Cephalocereus senilis (“old man” cactus). Baicalein and cephalocerone are 4´-deoxyflavonoids.
22

Figure 1.3. “Old Man” Cactus (Cephalocereus senilis).
(Photograph courtesy of Richard Ryon, owner of www.oldmancactus.com)
23

Figure 1.4. “Old Man” Cactus Callus and Cell Suspension Cultures.
Chitin-elicited (red) and normal (yellow).
24
COOH
NH2
COOH COOH
OH
COSCoA
OH
PAL C4H 4CL
phenylalanine trans -cinnamic acid para -coumaric acid
4-coumaroyl-CoA
+
COOH
CH2
COSCoA3
malonyl-CoA
CHS
OH
OH
OH
OH
O
tetrahydroxychalcone
CHIO
OH
OH
OH
Onaringenin (flavonone)
O
OH
OH
OH
O
aurone
Flavonoids, Isoflavonoids, Flavones,
Anthocyanins, condensed Tannins, etc.
B
B
B
B
B
peroxidase
A
A
A
Figure 1.5. The Phenylpropanoid Pathway.
Starting with general phenylpropanoid metabolism and leading to the major subgroups: the chalcones, aurones, flavonoids, anthocyanins, etc. Enzyme names are abbreviated as follows: phenylalanine ammonia lyase (PAL), cinnamate 4-hydroxylase, 4-coumaroyl-CoA ligase (4CL), chalcone synthase (CHS), and chalcone isomerase (CHI). All flavonoid derivatives contain the phenylpropanoid nucleus (B-ring) from phenylalanine and a second aromatic ring (A-ring) derived from the polyketide formed from three malonyl CoA molecules.
25
COOH
NH2
COOH COOH
OH
COSCoA
OH
PAL C4H 4CL
phenylalanine trans- cinnamic acid p-coumaric acid
4-coumaroyl-CoA
GENERAL PHENYLPROPANOID METABOLISM
O
O
OH
OH
OH
OH
O
CH3CH3
O
OH
OH
OH
OHO
OH
O
O
O
FLAVONOIDS ISOFLAVONOIDSAURONE
e.g. e.g. e.g.
kaempferol cephalocerone kievitone
OH
COOH
OH
OH
O
O
OH
OH
SOLUBLE ESTERS
e.g.
chlorogenic acid
O OO
COUMARINS
e.g.
psoralen
SUBERIN, LIGNIN, and other cell wall bound phenolics
Figure 1.6. Natural Products from the Phenylpropanoid Pathway.
Reaction scheme illustrating the production of different natural products leading from the core reactions of the phenylpropanoid pathway.
26

Figure 1.7. Mold-Infected Callus Cultures of “Old Man” Cactus.
27

O
OH
OCH3
GlcO
O
O
OH
OH
GlcO
O
O
OH
GlcO
O
O
GlcO
OH
OH
O
O
OH
OH
OH
O
O
OH
OH O
O
OH
OOH
OH
O
O
O
OH
O
O
O
OH
O
Figure 1.8. Structures of Major 4´-Deoxyflavonoids from Elicited “Old Man” Cactus Cell Suspension Cultures.
28
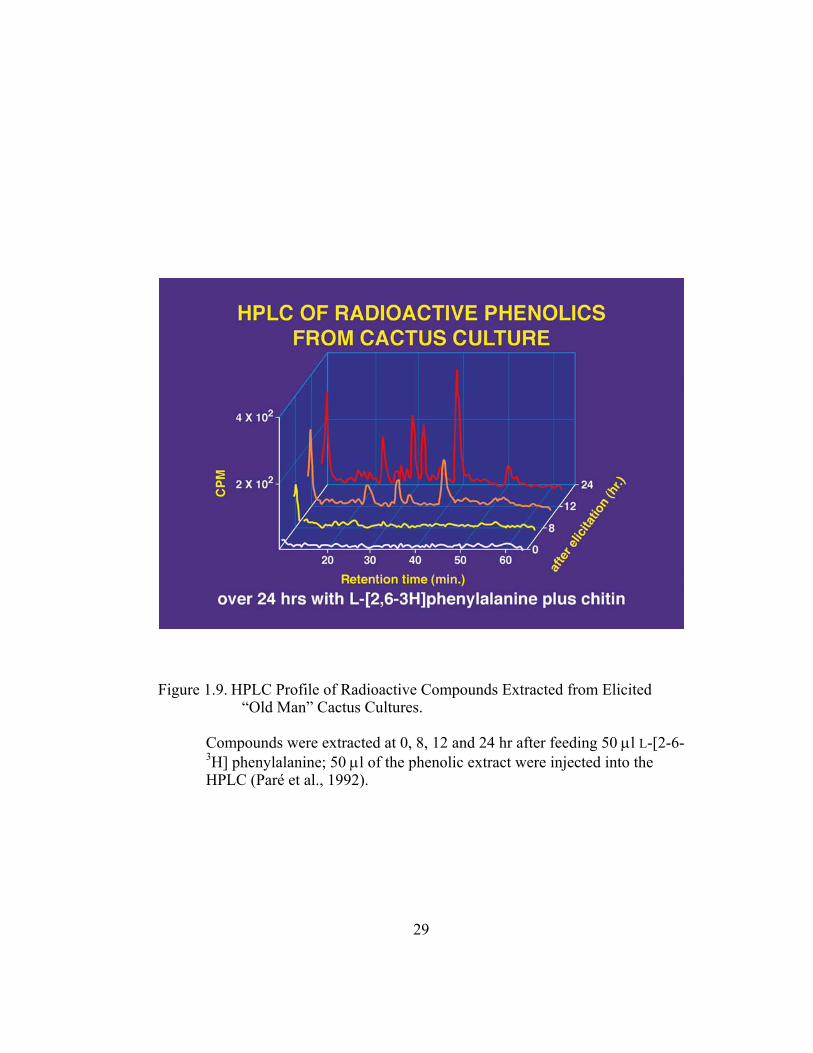
Figure 1.9. HPLC Profile of Radioactive Compounds Extracted from Elicited “Old Man” Cactus Cultures.
Compounds were extracted at 0, 8, 12 and 24 hr after feeding 50 µl L-[2-6- 3H] phenylalanine; 50 µl of the phenolic extract were injected into the HPLC (Paré et al., 1992).
29
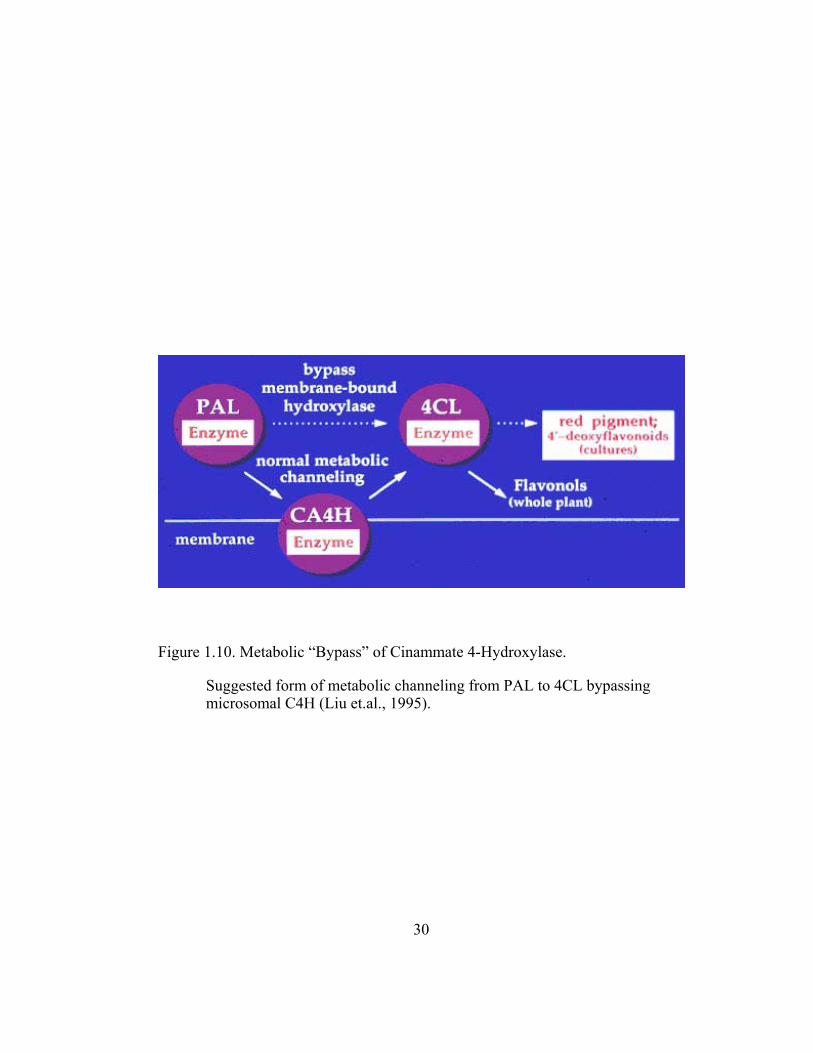
Figure 1.10. Metabolic “Bypass” of Cinammate 4-Hydroxylase.
Suggested form of metabolic channeling from PAL to 4CL bypassing microsomal C4H (Liu et.al., 1995).
30
OH
OH
OH
OH
O
4' 6'
2'
4
1
O
OH
OH
OH
O
78
5
4'
1'
6
1
2
3
4
O
OO
O
OH
4'
1'1
2
34
5
67
chalcone
flavone
4'-deoxyaurone (cephalocerone)
A
B
A
B
B
A
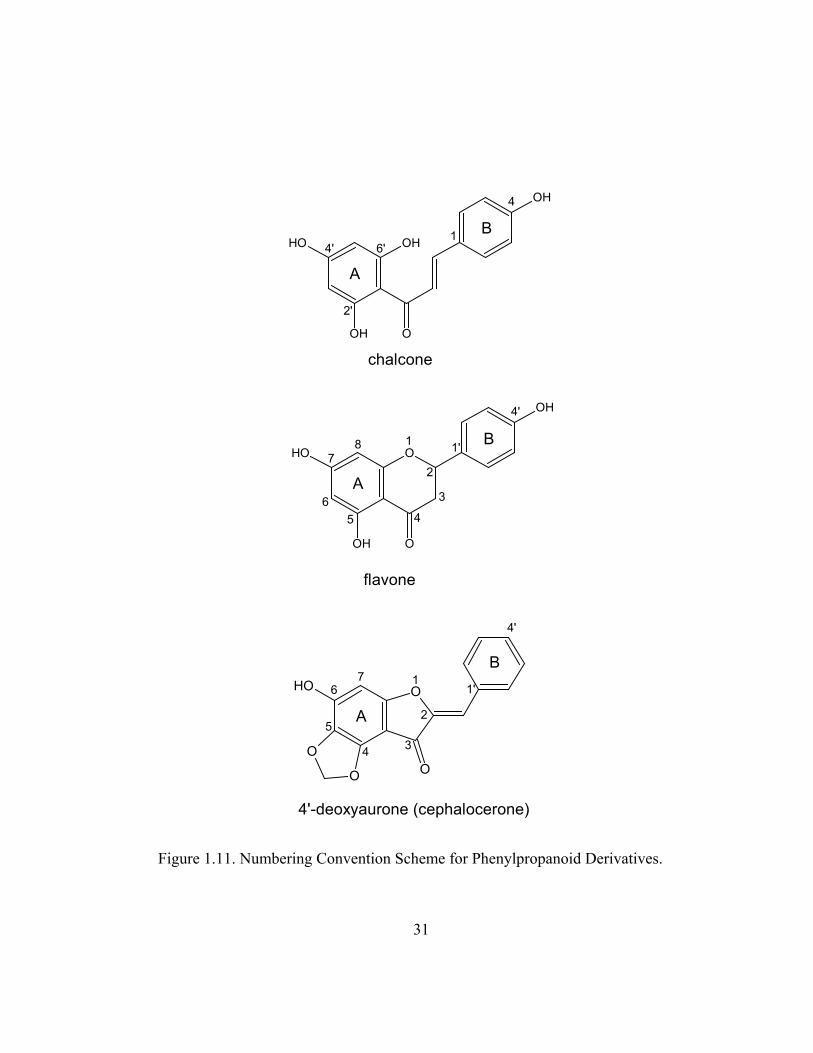
Figure 1.11. Numbering Convention Scheme for Phenylpropanoid Derivatives.
31
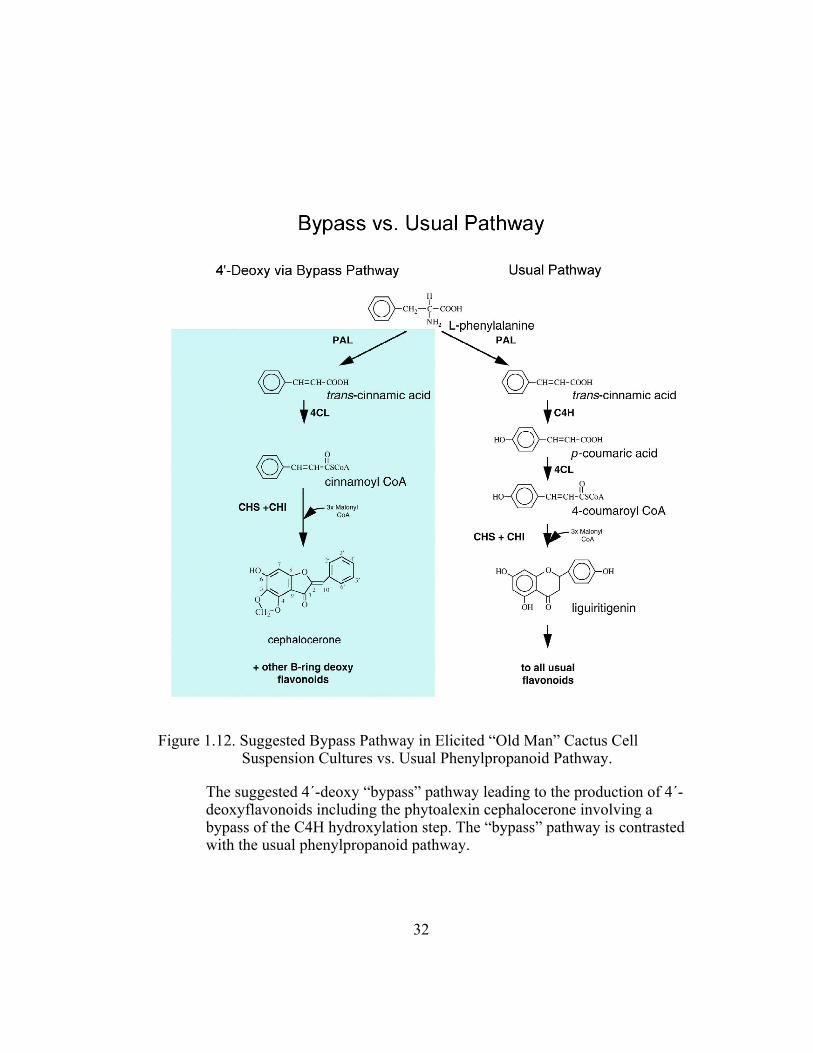
Figure 1.12. Suggested Bypass Pathway in Elicited “Old Man” Cactus Cell Suspension Cultures vs. Usual Phenylpropanoid Pathway.
The suggested 4´-deoxy “bypass” pathway leading to the production of 4´-deoxyflavonoids including the phytoalexin cephalocerone involving a bypass of the C4H hydroxylation step. The “bypass” pathway is contrasted with the usual phenylpropanoid pathway.
32
COOH
NH2
COOH COOH
OH
COSCoA
OH
COOH
CH2
COSCoA3 x
4'-dehydroxylase
O
O
OH
OO
cephalocerone (4'-deoxyaurone)
OH
OH
OH
O
4'-deoxychalcone
peroxidaseother
4'-deoxyflavonoids
PAL C4H 4CL
phenylalanine trans-cinnamic acid
para-coumaric acid
4-coumaroyl-CoA
+
malonyl-CoA
CHS
OH
OH
OH
OH
O
tetrahydroxychalcone
O
OH
OH
OH
O
naringenin
CHI
usual 4'-hydroxyflavonoids
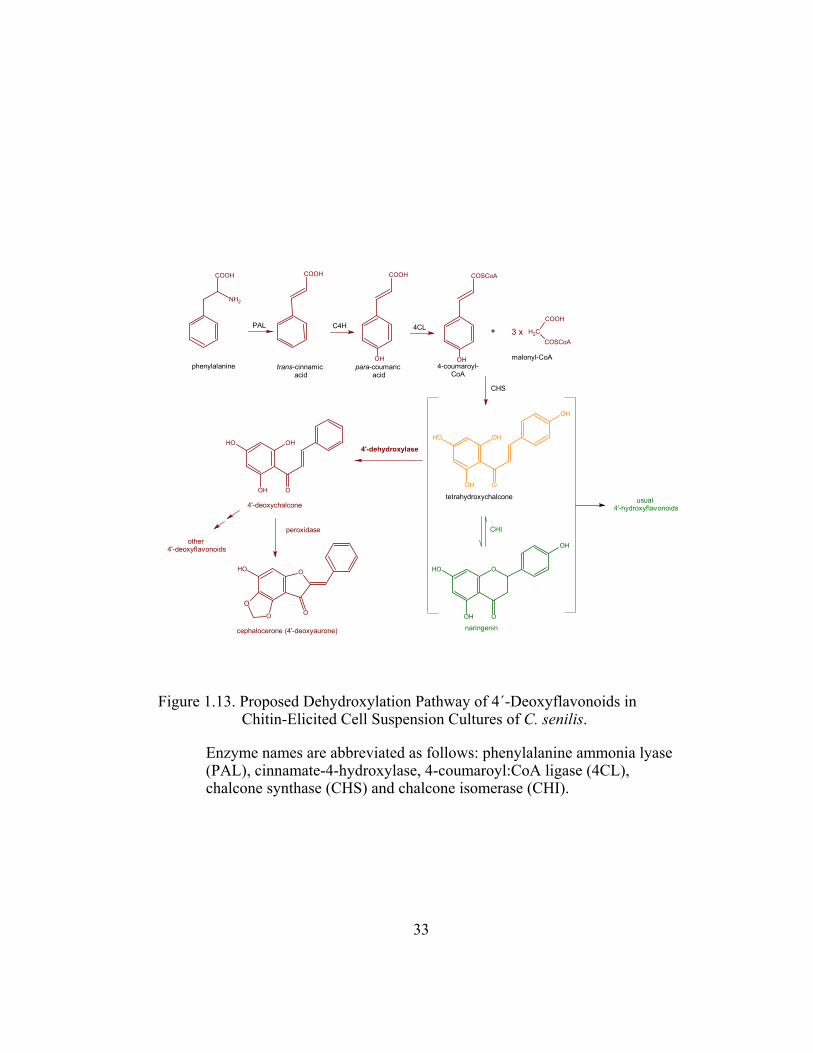
Figure 1.13. Proposed Dehydroxylation Pathway of 4´-Deoxyflavonoids in Chitin-Elicited Cell Suspension Cultures of C. senilis.
Enzyme names are abbreviated as follows: phenylalanine ammonia lyase (PAL), cinnamate-4-hydroxylase, 4-coumaroyl:CoA ligase (4CL), chalcone synthase (CHS) and chalcone isomerase (CHI).
33
4-coumaroyl-CoA
3 x Malonyl CoA
CHS PKR
O
OH
OH
OH
4' 6'
2'
4
1
OH
OH
O
OH4'
6'
2'
4
1
CHI (DCHI)
OH
OH
O
O H7
5
2
4'
1'
Isoliquiritigenin
2S-liquiritigenin (Note:flavone numbering)
OH
OCH3
O
O
OH
7
5
2
4'
1'
calycosin + other 5-deoxyisoflavonoids
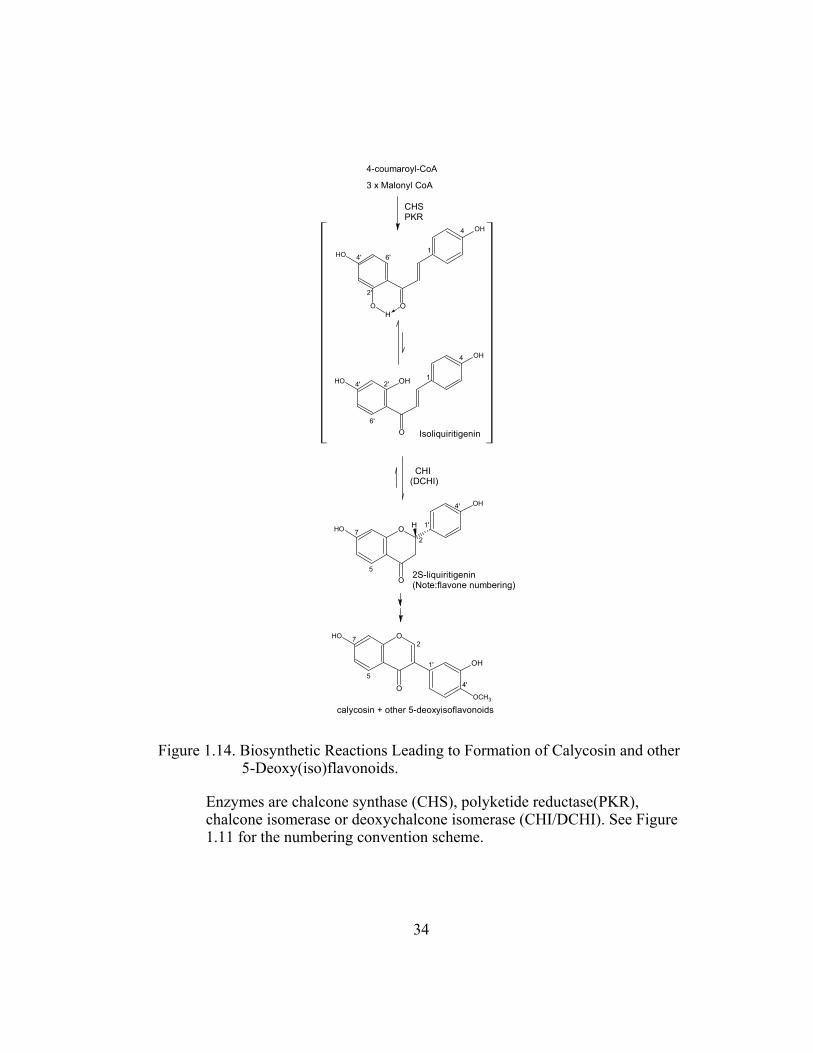
Figure 1.14. Biosynthetic Reactions Leading to Formation of Calycosin and other 5-Deoxy(iso)flavonoids.
Enzymes are chalcone synthase (CHS), polyketide reductase(PKR), chalcone isomerase or deoxychalcone isomerase (CHI/DCHI). See Figure 1.11 for the numbering convention scheme.
34

Chapter 2
Results: Novel Dehydroxylation Pathway in Elicited Suspension Cultures of Cephalocereus senilis
A. COMMENTS ON THE TEST SYSTEM: CHITIN-ELICITED C. SENILIS CELL SUSPENSION CULTURES
A novel system with the production of B-ring 4´-deoxyflavonoids in
chitin-elicited cell suspension cultures of Cephalocereus senilis (“old man”
cactus) suggests an unusual form of metabolic activity (Liu et al., 1993). The
attachment of the B-ring 4´-hydroxyl group is catalyzed by C4H prior to the
formation of the C15 flavonoid skeleton in the phenylpropanoid pathway. The
usual B-ring hydroxylation pattern is found in flavonoids isolated from healthy
whole plants of “old man” cactus. Some of the elicited compounds so far
identified were shown to be derived from L-[2,6-3H] phenylalanine as determined
by HPLC with a radioisotope detector (Paré et al., 1992) (Figure 2.1). In
collaboration with the group of Dr. Richard Dixon, it has been shown that the
elicited suspension cultures have all the necessary enzymatic activities needed for
the conversion of L-phenylalanine to the B-ring deoxyflavonoids, a single form of
CoA ligase that is unusually active with cinnamic acid, a chalcone synthase
exhibiting activity towards cinnamoyl CoA, and two isoforms of chalcone
isomerase showing activities to the 4´-hydroxychalcone and 4´-deoxychalcone
(Liu et al., 1995; Paré et al., 1992). The simplest explanation would be a reduction
in the activity of C4H, but enzymatic studies in Dr. Mabry’s group has shown that
35

C4H activity is not significantly reduced in elicited cactus cultures (Liu et al.,
1995). This result suggests metabolic channeling, specifically a “bypass” of
microsomal C4H in the biosynthetic pathway from PAL to CoA ligase with the
possibility that a different isoform of PAL is induced, that is, an isoform that
interacts directly with CoA ligase rather than with C4H. It is also a possibility that
a novel B-ring dehydroxylation step after 4CL is induced in the phenylpropanoid
pathway for B-ring deoxyflavonoid biosynthesis in chitin-elicited C. senilis cell
suspension cultures.
B. RESULTS AND DISCUSSION
Here we describe the biochemical methods used to provide evidence, from
in vivo labeling experiments, for the lack of metabolic channeling among the
early enzymes in the phenylpropanoid biosynthetic pathway of C. senilis and
support for a dehydroxylation step in the initial stages of flavonoid biosynthesis.
Application of unlabeled precursors, cinnamic and coumaric acids, in a pulse-
chase experiment were conducted and their levels of incorporation with respect to
the deoxyaurone were followed into the phenylpropanoid pathway and the cellular
and metabolic system that leads to B-ring deoxyflavonoids was investigated.
These labeling studies show a novel dehydroxylase activity for the synthesis of
the phytoalexin 4´-deoxyaurone in the elicited C. senilis system.
36

1. Analysis of the Phytoalexin Red Pigment
Analysis by HPLC photodiode array of extracts from cell suspension
cultures of C. senilis sampled 24 hr after elicitation, revealed one major peak
absorbing in the UV range 200-400 nm. This compound had a retention time of
20.7 min and its UV spectra generated by the photodiode array showed
absorbance peaks at 280 and 315, consistent with the UV spectra of the previously
isolated red phytoalexin cephalocerone (Paré et al., 1991) (Figure 2.2). The
identity of the peak at 20.7 min was further confirmed by LC/ESI-MS analysis of
the extracts which showed a major ion with a molecular mass of 283, also as
expected for the previously identified 4´-deoxyaurone. Accumulation of the red
pigment cephalocerone is not observed in non-elicited cell suspension cultures
and the aurone peak does not appear in its extracts (Figure 2.3). HPLC metabolic
profile analysis of both elicited and non-elicited cell cultures show that the
production of the 4´-deoxyaurone is a very specific response to chitin or fungal
elicitation, and its accumulation is not just a biochemical artifact of the usual
phenylpropanoid pathway induction. Cephalocerone has phytoalexin properties
and may be produced in whole plants as a defense mechanism against pathogen
infection (Paré et al., 1991).
2. Metabolic Fate of trans-Cinnamic Acid in “Old man” Cactus Cell Suspension Cultures
Enzyme organization into membrane-bound macromolecular complexes,
instead of free-floating cytosolic proteins in metabolic pathways, is the central
37

feature of primary metabolism and has now become evident in secondary
metabolic pathways (Burbulis and Winkel-Shirley, 1999; Ovadi and Srere, 1996,
2000). Flavonoid biosynthesis competes directly with a number of pathways
branching from the phenylpropanoid core reactions and different environmental
cues “switch” the branches to favor one metabolite from another. This
phenomenon, which was suggested by H. Stafford as early as 1974, is referred to
as metabolic channeling, the direct transfer of metabolites in active sites, via
access to a common substrate pool or through the formation of multienzyme
complexes (Hrazdina and Wagner, 1985).
Substrate channeling occurs when reaction intermediates do not
equilibrate freely with exogenously applied intermediates. Different intermediate
pools may be formed as a result of different environmental cues although these
pools may or may not result from metabolic channeling. However, if externally
applied intermediates equilibrate freely with biosynthetic intermediate pools,
there would be a strong possibility that metabolic channeling does not occur.
Therefore we use elicited “old man” cactus cell suspension cultures to study the
existence of different phenylpropanoid intermediate pools that constitute the
metabolic grid leading to the synthesis of cephalocerone, a B-ring deoxyaurone. A
metabolic profile using 3H-L-Phe showed that elicited and non-elicited cultures
accumulate various labeled phenolic compounds with 15% of the total extractable
label belonging to the cephalocerone peak at 20.7 min (Figure 2.4).
Because of its unusual B-ring deoxy functionality, the production of the
4´-deoxyaurone may require selective substrate channeling of trans-cinnamic acid
38

to give a chalcone lacking the B-ring hydroxyl group. Studies have shown that the
downstream enzymes of cactus cell suspension cultures exhibit an unusually high
activity towards B-ring deoxy intermediates (Liu et al., 1995). These enzymes,
4CL, CHI and CHS, also show no selectivity between 4´-deoxy and 4´-hydroxy
substrates. Therefore, a bypass in the C4H hydroxylation step in the 4´-deoxy
pathway may account for the selective formation of 4´- deoxyflavonoids (Figure
2.5). This means that trans-cinnamic acid is accessed directly by 4CL instead of
p-coumaric acid, which is its usual substrate. If “old man” cactus cell suspension
cultures possessed only a single pool of trans-cinnamic acid by which 4CL
derives its substrates from, then the presence of excess unlabeled trans-cinnamic
acid in 3H-L-Phe fed cell suspension cultures would result in a lower specific
activity of tritium in cephalocerone as compared to the values of tritium activity
in the aurone in the absence of the unlabeled trans-cinnamic acid.
To examine the metabolic fate of trans-cinnamic acid in the above 4´-
deoxy hypothesis, chitin-elicited “old man” cactus cell suspension cultures were
simultaneously fed 3H-L-Phe and two different concentrations (10-4 and 10-5 M) of
unlabeled trans-cinnamic acid. The exogenous substrates were fed at 12 hr and
16 hr post elicitation, at the start and at the height of (hydroxyl)-cinnamate:CoA
ligase (4CL) activity, respectively, as determined by Liu (1996; Figure 2.6).
Cephalocerone accumulation also starts at 12 hr and maximizes at 16 hr (Paré et
al., 1991). Results are expressed as the ratio of incorporation of 3H-L-Phe
(determined as counts per minute; CPM) in the phenolic product (Iproduct; in this
39

case, cephalocerone) to its absorbance (Aproduct(280nm); milliabsorbance units; mAu)
per g fresh weight of culture collected.
Figure 2.7 shows the results from such series of isotope dilution
experiments in chitin-elicited “old man” cactus cell suspension cultures. In the
absence of externally applied unlabeled trans-cinnamic acid, the cephalocerone
Iproduct/Aproduct ratio (ratios are in units of CPM/mAu·g fresh wt) in elicited cell
cultures is relatively high at around 0.006 for precursors added at 12 hr which is at
the start of the increase in 4CL induction. Non-elicited cell cultures have a zero
value for their ratio because no red pigment cephalocerone is produced.
Significant ratio reduction by about 47% is observed when 10-4 M unlabeled
trans-cinnamic acid is added to the elicited cell cultures. Ratio reduction,
although present, is less significant at a lower concentration (10-5 M) of
externally added trans-cinnamic acid. A higher ratio of about 0.016-0.018 is
observed in cell cultures where precursors are added at 16 hrs, at the height of
4CL activity, when more incorporation of 3H-L-Phe in elicited cell cultures is
expected. The ratio reductions are more pronounced at the 16 hr and 3H- labeling
of cephalocerone is significantly higher (Figure 2.7).
These results on the 3H-L-Phe pulse-chase experiments support time
induction studies (Liu et al., 1995) on phenylpropanoid enzymes in elicited “old
man” cell suspension cultures that show the start of increased specific activities
12 hr after elicitation and leveling off after 16 hr. These results also indicate that
externally added unlabeled trans-cinnamic acid dilutes isotope incorporation into
the red pigment 4´-deoxyaurone, suggesting that the phenylpropanoid enzymes
40

have access to external cytoplasmic pools. However, this does not directly prove
the lack of metabolic channeling nor does it indicate the presence of a single
endogenous pool of trans-cinnamic acid in the absence of an externally fed
compound.
3. Metabolic Fate of p-Coumaric Acid in “Old Man” Cactus Cell Suspension Cultures
To increase the sensitivity and specificity of the isotope dilution profile
studies, the above-mentioned in-vivo labeling experiments were repeated with
cell suspension cultures elicited by chitin 16 hr before simultaneous addition of 3H-L-Phe and unlabeled trans-cinnamic or p-coumaric acid. Figure 2.8 (raw data
in Appendix B and C) shows the incorporation ratios of these dilution
experiments (for this experiment, labeling measurements were done in
disintegrations per minute, DPM; ratio is expressed as DPM/mAu·g fresh wt).
Both precursors are able to dilute the isotopic label on the 4´-deoxyaurone.
Isotopic dilution by the two concentrations of trans-cinnamic acid shows similar
results to earlier studies (Figure 2.7) as expected. Figure 2.8 also shows that even
if the amount of 3H-specific activity of cephalocerone is reduced through the
addition of unlabeled intermediates, a significant amount of 3H-L-Phe is still
channeled through the pathway. However, if B-ring deoxyflavonoid production is
a result of the selective substrate channeling of trans-cinnamic acid, assuming that
the phenylpropanoid pathway has only one cytoplasmic pool of each precursor
present, the externally added unlabeled p-coumaric acid should not dilute the 3H
isotopic label significantly on the 4´-deoxyaurone. If unlabeled p-coumaric acid is
41

disrupting the pathway then the amount of 4´-deoxyaurone produced by the
elicited cultures should be lower because a 4´-hydroxyl phenyl precursor such as
p-coumaric acid is being accessed by the downstream phenylpropanoid enzymes
in the production of the 4´-deoxyaurone. Figure 2.9 (raw data in Appendix B and
C) shows that the amount of 4´-deoxyaurone does not change significantly with
the addition of unlabeled trans-cinnamic and p-coumaric acids. Incorporation of
unlabeled trans-cinnamic acid is not expected to change the amount of 4´-
deoxyaurone produced since it is a 4´-deoxy precursor. However, the addition of
unlabeled p-coumaric acid, if it is indeed incorporated into the 4´-deoxy bypass
pathway, should lower the amount of 4´-deoxyaurone accumulated because it is a
4´-hydroxylated precursor. But the amount of 4´-deoxyaurone produced by the
elicited “old man” cactus cell cultures is not reduced with the addition of
unlabeled p-coumaric acid (Figure 2.9). Figure 2.8 and 2.9 also show a greater
efficiency of incorporation of unlabeled p-coumaric acid into the 4´-deoxyaurone
compared to unlabeled trans-cinnamic acid, without changing the amount of 4´-
deoxyaurone produced. The similarity in the isotope dilution behavior of p-
coumaric acid and trans-cinnamic acid show that the synthesis of the 4´-
deoxyaurone requires a 4´-hydroxylated precursor (p-coumaric acid). This
implicates a novel dehydroxylase activity for the 4´-deoxyflavonoid biosynthesis
in the elicited “old man” cactus cell culture system.
42

4. The Proposed 4´-Deoxyflavonoid Bypass Theory
The formation of secondary compounds is known to be characterized by
low steady-state dynamics and often do not undergo rapid turnover (Hartmann,
1996). This means that synthesis of secondary metabolites is regulated differently
compared to the straightforward and feedback regulated primary metabolism (i.e.
glycolysis and Kreb’s cycle; Figure 2.10). The existence of a “metabolic grid” in
flavonoid metabolism seems reasonable since constitutive secondary metabolism
intermediates must be synthesized and stored to be available to the cell when
necessary (e.g. phytoalexin production in plant defense; Hartmann, 1996).
Therefore, the exact metabolic fate of a particular intermediate is not easily
determined. The disappearance of a metabolite in a certain part of the pathway is
not strong evidence for its degradation as the compound can be converted to
another intact compound in different cytoplasmic pools.
If strict metabolic channeling such as the phenomenon defined by
Hrazdina and Wagner (1985) occurs, incorporation of externally added unlabeled
precursors into the final product should not take place. The above dilution effects
indicate that free equilibration of substrate pools occur. It is possible that there
exists more than one pool of each precursor which the different phenylpropanoid
pathway enzymes can “tap”. The relative stability of reaction intermediates in the
phenylpropanoid pathway allows for the equilibration of these intermediates with
other cytosolic compartments or even for the possibility to form redundant
substrate pools. The fact that the level of 4´-deoxyaurone accumulation is not
lowered (Figure 2.9) may mean that all of the enzymes in the 4´-deoxy pathway
43

have the ability to access the different reaction intermediate pools by which they
can maximize the production of cephalocerone. Phenylpropanoid intermediates in
the “old man” cactus chitin-elicitor induced 4´-deoxypathway either have several
pools or one pool that is accessible to the bulk compartment of the cell. These
pools are assumed to be present even before 3H-L-Phe is introduced. Upon
addition of 3H-L-Phe, a new labeled pool of L-Phe may be created or the
externally added 3H-L-Phe may equilibrate with the initial pool of L-Phe already
present in the cell suspension cultures. The assumption is since L-Phe is the first
substrate of the core phenylpropanoid pathway only a single pool of L-Phe is
present. However, it is known that different isoforms of PAL exist (Shirley, 1999)
and it is possible that these isoforms have their own pools of L-Phe. Multiple
genes encode the early enzymes and lack of gene duplication for the more
downstream enzymes are observed in some plants (Shirley, 1999). Gene
multiplicity for certain enzymes may confer plasticity in the phenylpropanoid
pathway because different isoforms of the same enzyme (e.g. PAL) may occur
depending on environmental cues (Dixon, 1997; Dixon et al., 1995; Ozeki et al.,
2000; Shirley, 1996; Shirley, 1999).
In elicited “old man” cactus cell cultures, PAL and C4H activity is
relatively high and downstream enzymes 4CL, CHS and CHI do not discriminate
between 4´-deoxy and 4´-hydroxy precursors (Liu et al., 1995; Paré et al., 1992).
A different PAL isoform may exist in “old man” cactus elicited cell cultures and
this isoform may be the enzyme that preferentially passes its trans-cinnamic acid
product to 4CL instead of giving it to C4H for hydroxylation. Data from the
44

isotope dilution studies show the incorporation of the cinnamate. However, higher
isotope dilution ratios from the addition of a more downstream 4´-hydroxylated
intermediate p-coumaric acid in the pathway without affecting the amount of 4´-
deoxyaurone that is produced do not support the bypass of C4H.
The observed isotopic dilution from the addition of a 4´-hydroxylated
intermediate, p-coumaric acid may be due to the disruption of 4CL’s pool of 3H-
trans-cinnamic acid which forces 4CL to find an alternate cinnamic pool that
happens to be unlabeled. It is possible that the external addition of p-coumaric
acid dilutes the 3H-labeled cinnamic pools and forces 4CL to find its cinnamic
acid source from a different pool. This “switch” may dilute the 3H-label in
cephalocerone but should not affect the amount of cephalocerone produced in the
cell. However, it is known that accumulation of trans-cinnamic acid is highly
unlikely because it is known to inhibit PAL (O'Neal and Keller, 1970). (Note,
however that the addition of 10-4 M trans-cinnamic acid in our isotope dilution
studies did not appear to inhibit metabolic flux via the PAL reaction in vivo, as the
tritium label from externally added 3H-L-Phe was observed to be incorporated into
various phenylpropanoid compounds including the 4´-deoxyaurone.) Earlier
studies provide evidence that trans-cinnamic acid is metabolized quickly to
prevent its accumulation in the phenylpropanoid pathway (Noe et al., 1980; Noe
and Seitz, 1983). It is therefore highly unlikely that multiple trans-cinnamic acid
pools exist to allow substrate pool “switching” of 4CL as a compensation for
dilution.
45

5. Evidence for the 4´-Dehydroxylation Step Leading to the Biosynthesis of Cephalocerone
The synthesis of 4´-deoxyflavonoids is highly unusual and exists only in a
very few plants. Baicalein, a 4´-deoxyflavonoid from skullcap (Scutellaria
baicalensis), and also one of the 4´-deoxyflavonoid compounds isolated from
elicited “old man” cactus cell cultures, is widely studied for its biochemical
effects on certain types of cancer but its biosynthesis is not well studied (Hsu et
al., 2001; Lazinsky et al., 2001). It has been shown recently that elicitation with
methyl jasmonate increased baicalein production in skullcap root cultures but
details of the pathway induction are not known (Kuzovkina et al., 2001). A group
of unusual 3-deoxyflavonoids is also found in apple (Malus domestica Borkh) and
pear (Pyrus communis L.) and is known to confer both plants’ resistance to
fireblight (Erwinia amylovora Burrill) (Rommelt et al., 1999; Treutter, 2001). A
chemosystematic study of Greyiaceae discusses the lack of B-ring subsititution in
some flavonols isolated from the leaf surface of three species of Greyia (Bohm
and Chan, 1992). However, lack of hydroxy-substitution in the different positions
of the B-ring other than the 4´-position is not uncommon because the B-ring is
mostly only 4´-hydroxylated at the beginning of flavonoid biosynthesis (Goodwin
and Mercer, 1983). While there are specific enzyme activities that remove
hydroxyl groups in the A and C ring, no activity has been studied extensively for
the 4´-dehydroxylation of the B-ring (Goodwin and Mercer, 1983; Kimura et al.,
2001).
The production of B-ring deoxyflavonoids in elicited C. senilis cultures
may not be due to the strict preference of the enzymes for 4´-deoxy substrates in
46

the elicited phenylpropanoid pathway. Our results strongly suggest that the
metabolic flux involved in the selective production of 4´-deoxyflavonoids may be
controlled by a specific enzyme activity that dehydroxylates the 4´-position of the
tetrahydroxychalcone B-ring. The 4´-dehydroxylation step appears to be induced
by elicitation because 4´-deoxyaurone accumulation is not observed on non-
elicited cultures (Figure 2.3). The isotope dilution experiments clearly establish
the metabolic fate of externally added p-coumaric acid in chitin-elicited cultures.
The incorporation of exogenous unlabeled p-coumaric acid leading to the dilution
of 3H-isotopic label in cephalocerone suggests the presence of a 4´-
dehydroxylation step within the phenylpropanoid pathway downstream of 4CL.
The addition of unlabeled compounds in pulse-chase experiments may not
provide direct evidence whether or not metabolic channeling occurs because
external addition of the precursor may affect metabolic flux. However, we show
that the amount of extracted cephalocerone from the elicited cell cultures with or
without addition of unlabeled precursors remains the same.
A dehydroxylation step within the flavonoid pathway would still be
consistent with the observed heightened activity of the phenylpropanoid enzymes
in elicited “old man” cactus cultures (Liu et al., 1995; Paré et al., 1992). It has
been determined from earlier enzymatic studies that all the phenylpropanoid
enzymes can activate both 4´-deoxy and 4´-hydroxy precursors (Liu et al., 1995).
This means that the usual phenylpropanoid biosynthesis can proceed up to the
production of the tetrahydroxychalcones. The tetrahydroxychalcone may then be
reacted on by a “dehydroxylase” thus producing a trihydroxychalcone lacking a
47

hydroxyl group at the 4´-position of the B-ring. Figure 2.11 suggests a model for
the chitin-induced biosynthesis of the 4´-deoxyaurone in elicited cell suspension
cultures of C. senilis.
General dehydroxylation activity is not unique to “old man” cactus cell
cultures. Flavonoids lacking the usual hydroxyl substitution pattern at the 5-
position of the A-ring were isolated from several species of Baptisia
(Leguminosea) (Markham and Mabry, 1968; Markham et al., 1968,1970;
Markham et al., 1968). The dehydroxylation activity at the C-5 position (C-6´,
chalcone numbering) probably occurs at the aromatic ring formation during the
polyketide stage or, at the latest, at the chalcone stage because the presence of the
step controlling the removal of the hydroxyl group results in all flavonoids being
equally affected (Goodwin and Mercer, 1983; Kimura et al., 2001). Baicalein, a
major 4´-deoxyflavonoid from skullcap roots (Scutellaria baicalensis), is also one
of the 4´-deoxyflavonoid compounds isolated from elicited “old man” cactus cell
cultures (Liu et al., 1993; Rossi et al., 2001). Baicalein has been shown to exhibit
biochemical activity against certain types of cancer cells (Gao et al., 2001; Rossi
et al., 2001), however the biosynthesis of baicalein in plants is not well
established (Kuzovkina et al., 2001). The conversion of purine and pyrimidine
nucleotides by ribonucleotide reductases to deoxynucleotides in DNA replication
and repair is an example of a well characterized dehydroxylation step that is
present in all organisms (Stubbe et al., 2001). The existence of different types of
dehydroxylation activity for the production of essential deoxy-compounds (Figure
48

2.12) in some biosynthetic pathways is evidence that the same activity such as the
proposed 4´-dehydroxylation step in “old man” cactus can occur.
6. Comments on the 4´-Deoxyflavonoid Phenomenon in Cephalocereus senilis
Biochemical and enzymological studies on the “old man” cell suspension
cultures began in the early 1990’s upon a serendipitous discovery in Mabry’s lab
that the cell cultures turn red upon fungal infection. Paul Paré conducted the
initial biochemical studies and more detailed enzymological investigations were
continued by Qin Liu in collaboration with Richard Dixon’s lab at the Noble
Foundation. The 4´- deoxyflavonoid phenomenon continues to draw two schools
of thought, the 4´- hydroxylation “bypass” pathway and the 4´-dehydroxylation
route. Below are the comments contributed by Mabry and Paré:
Mabry’s comments
I have favored the “bypass” theory (bypassing a hydroxylase in the phenylpropanoid pathway) to form the 4’-deoxyflavonoids in chitin-elicited C. senilis cultures because of the simplicity of requiring only a modification at the cellular-membrane transport system to execute the “bypass”. In contrast, the removal of the 4’-hydroxyl group from a flavonoid is almost not known presumably because the 4’-hydroxyl is one of the most acidic groups in flavonoids playing a significant role in most of the metabolic reaction of these compounds. To generate a dehydroxylase seems to me to be highly unlikely because it probably requires activation of a dehydroxylase gene not previously reported.
Gani’s findings support the dehydroxylase theory and set the stage for a new round of labeling experiments which we hope to jointly conduct with Paul Paré, Texas Tech University.
49

Paré’s comments
The cactus aurone, cephalocerone is devoid of the usual 4' hydroxyl group in the B-ring. Gani’s experimental data with radiolabeled precursors indicates that the p-coumaryl CoA serves as the precursor for deoxy-aurone synthesis. Such a dehydroxylation step is not unprecedented. In the case of ribonucleotide reductase both purine and pyrimidine nucleotides are converted to deoxynucleotides via a dehydroxylation step. Three radical reaction mechanisms have been proposed for the separate classes of reductases that generate the monomeric precursor for DNA replication and repair. Each of the enzyme reactions catalyzes the dehydroxylation of the 2' hydroxy in the ribose ring to form the deoxyribose DNA backbone (Stubbe et al., 2001). A substantive distinction between ribonucleotide reductase and the proposed dehydroxylase reactions is that the reductase is converting a preformed precursor while aurone formation requires almost a back to back hydroxylation/dehydroxylation set of reactions. The loss of methyl units can also occur during the "building" of molecules. In the case of cholesterol synthesis, the A-ring methyl groups at position 4 are removed by a 3-step process involving the successive intermediates 4"-hydroxymethyl and 4"-formyl sterols (Nes and McKean, 1977).
C. EXPERIMENTAL
1. Plant Material
Callus cultures of Cephalocereus senilis were initiated from inner stem
tissue of greenhouse-grown plants and cultured on a medium containing MS
minimal organic salts, 3% (w/v) sucrose, 0.2% (w/v) Phytagel, 4.5 µM 2,4-
dichlorophenoxyacetic acid, 4.4 µM N-6-benzyladenine, with the pH adjusted to
5.7 (Figure 2.13). Suspension cultures were initiated from callus material and
grown on a rotary shaker at 118 rpm in the dark in the same medium but in the
absence of gelrite (Figure 2.13). Cultures were subcultured every fourth day by
mixing 150 ml of suspension culture with an equal amount of fresh medium.
50

Four days after subculturing, dark-grown “old man” cactus cell suspension
cultures (25 ml batches) were added to 5 ml of fresh media and treated with 5 ml
of a sterile chitin suspension (8 mg chitin ml-1 fresh media). The chitin-elicited
cells were harvested at various times after elicitation and control cells were
treated with the same amount of fresh media.
2. Chemicals
3H-L- 2,3,4,5,6-Phe (124 Ci/mmol) was supplied by Amersham (Little
Chalfont, United Kingdom). Unlabeled trans-cinammic acid and p-coumaric acid
were obtained from Sigma.
3. Precusor Dilution Experiments
“Old man cactus” cell suspension cultures (25-ml batches) were incubated
4 days after subculturing with 50 µl of 3H-L- 2,3,4,5,6-Phe (124 Ci/mmol) with or
without unlabeled trans-cinnamic or 4-coumaric acid (10-4 and 10-5). After 24 hr,
the cells were filtered through a nylon mesh and cooled in liquid N2. The soluble
phenolics were extracted with 25 ml of chilled acetonitrile by sonication for 10
min and centrifuged at 14,000 rpm for 18 min. The residue was re-extracted with
25 ml of 50% (v/v) aqueous acetonitrile and centrifuged at 16,000 rpm for 20 min.
The combined extracts were evaporated to dryness in vacuo. The concentrate was
redissolved in 5.0 ml of 0.01 N HCl, and the solution was acidified with 20 drops
of 1 N HCl.
51

The following solid phase extraction steps were performed using a
Supelco Visiprep vacuum manifold (Supelco, Bellefonte, PA). The acidic solution
was applied to C-18 Sep Pak Plus cartridge (Waters Associates, Milford, MA; the
C-18 Sep Pak was equilibrated first with 1 ml of acetonitrile followed by 1 ml
H2O). After washing the cartridge with 4 ml of 0.01 N HCl, the phenolic fraction
was recovered by eluting the Sep Pak with 3 ml of H2O:acetonitrile (6:4).
Aliquots (50 µl) were separated by HPLC (as described below) and radioactivity
was monitored with a Beckman 171 Radioisotope detector (32 Karat system
software, Beckman-Coulter Inc., Fullerton, CA). The radioactive peaks were
identified by co-migration with isolated standard samples and authentic standards
(Sigma). A quench curve with 25% acetonitrile in pH 3.0 water (pH adjusted
with glacial acetic acid), spiked with tritium labeled phenylalanine, showed
optimal counts at 4:1 cocktail: effluent ratio with ReadyFlow III scintillation fluid
(Beckman-Coulter Inc., Fullerton, California). Aliquots of 3H-labeled
phenylalanine were injected into the HPLC system to calculate the total amount of
radioactivity (DPM) administered to cultures.
4. Separation of Phenolics by Reverse Phase HPLC
HPLC analysis was performed by using a modification of the method of
Burbulis (1996). The Beckman chromatography system (Beckman-Coulter Inc.,
Fullerton, California) is composed of a model 126 solvent module/dual pump
system, model 507e autosampler, model 168 photodiode array detector and a
model 171 radioisotope detector (Figure 2.14). System operation and data
52

collection were managed by Beckman 32 Karat® system software on an IBM
300PL (IBM Corp., White Plains, NY) with Windows NT (Microsoft, Redmond,
WA). Plant or cell suspension phenolic fractions (50 µl each) were applied to an
Allsphere ODS-2 reverse phase HPLC column (5 micron particle size, 4.6 x 250
mm; Alltech, Deerfield, IL) and eluted in HPLC-grade water (Solvent A; pH 3.0
adjusted with glacial acetic acid) with an increasing acetonitrile (Solvent B)
concentration gradient (starting with 5% [v/v] B; 0-2 min, 0-10% B; 2-27 min,
10-50% B; 27-29 min, 50-100% B; 29-31 min, 100% B; 31-33 min, 100-5% B) at
a constant flowrate of 1.0 ml min-1 (Burbulis et al., 1996). Elution was monitored
at 280 nm, based on absorbance maxima of the cephalocerone, although similar
profiles were obtained at 255 and 330 nm.
5. Liquid Chromatography Electrospray Ionisation Mass Spectrometry (LC/ESI-MS) analysis
ESI-MS has been used for routine mass spectrometric analysis of a wide
range of biomolecules including flavonoids. ESI-MS is considered a soft
ionization technique that allows an accurate molecular weight measurement of
both fragile and non-volatile biomolecules. Cell suspension extracts were
analyzed at the University of Texas at Austin Mass Spectrometry Laboratory with
a Finnigan LCQ, an ion trap mass spectrometer equipped with electrospray
ionization (ESI) and atmospheric pressure chemical ionization (APCI) ion sources
(Figure 2.15; Thermo Finnigan, San Jose, CA).
53

6. Other Experimental Activities
It should be noted that only the final experimental procedures are
presented; the development of these procedures each required dozens of trial runs
and refinement over several years. Also not included here were numerous
investigations not relevant to this dissertation:
(a). Further characterization of the red pigment.
Attempts were made to re-isolate the red pigment for use in metabolic
profiling as a standard and for further molecular studies. Light microscopy
was also used to determine localization of the red pigment in the cactus
cultures. An organic synthesis procedure for cephalocerone was developed
with Didier Villemin (Ecole Nationale Superieure d'Ingenieurs de Caen
ISMRA, Universite de Caen, France) but has not been carried out due to
time and budget constraints.
(b). Development of cell suspension cultures from Opuntia and
Mammalaria callus cultures.
Numerous attempts were made to develop cell suspension cultures from
other cacti species to survey the 4´-deoxyflavonoid phenomenon. Time
and budget constraints did not allow for a long term maintenance of the
new cultures because metabolic profiles were being developed for the C.
senilis system.
54

(c). HPLC- radiolabeling experiments using 14C-labeled trans-cinnamic
and p-coumaric acid.
Synthesis of trans-cinnamic acid and p-coumaric acid were successful but
the yield from each synthesis procedure was so low that the radioactivity
levels were undetectable in isotope experiments using these labeled
precursors (see Chapter 3).
LITERATURE CITED
Bohm, B. A. and Chan, J., 1992. Flavonoids and affinities of Greyiaceae with a
discussion of the occurrence of B-ring deoxyflavonoids in dicotyledonous families. Systematic Botany 17, 272-281.
Burbulis, I. E., Iacobucci, M. and Shirley, B. W., 1996. A null mutation in the first enzyme of flavonoid biosynthesis does not affect male fertility in Arabidopsis. Plant Cell 8, 1013-1025.
Burbulis, I. E. and Winkel-Shirley, B., 1999. Interactions among enzymes of the Arabidopsis flavonoid biosynthetic pathway. Proceedings of the National Academy of Sciences of the United States of America 96, 12929-12934.
Dixon, R. A., 1997. Plant secondary metabolism: Control points and prospects for genetic manipulation. Abstracts of Papers of the American Chemical Society 213, 8-AGFD.
Dixon, R. A., Harrison, M. J. and Paiva, N. L., 1995. The isoflavonoid phytoalexin pathway - from enzymes to genes to transcription factors. Physiologia Plantarum 93, 385-392.
Gao, Z. H., Huang, K. X. and Xu, H. B., 2001. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in HS-SY5Y cells. Pharmacological Research 43, 173-178.
55

Goodwin, T. W. and Mercer, E. I., 1983. Introduction to Plant Biochemistry. Pergamon Press, New York.
Hartmann, T., 1996. Diversity and variability of plant secondary metabolism: a mechanistic view. Entomologia Experimentalis et Applicata 80, 177-188.
Hrazdina, G. and Wagner, G. J., 1985. Metabolic pathways as enzyme complexes: evidence for the synthesis of phenylpropanoids and flavonoids on membrane-associated enzyme complexes. Archives of Biochemistry and Biophysics 237, 88-100.
Hsu, S. L., Hsieh, Y. C., Hsieh, W. C. and Chou, C. J., 2001. Baicalein induces a dual growth arrest by modulating multiple cell cycle regulatory molecules. European Journal of Pharmacology 425, 165-171.
Kimura, Y., Aoki, T. and Ayabe, S., 2001. Chalcone isomerase isozymes with different substrate specificities towards 6 '-hydroxy- and 6 '-deoxychalcones in cultured cells of Glycyrrhiza echinata, a leguminous plant producing 5-deoxyflavonoids. Plant and Cell Physiology 42, 1169-1173.
Kuzovkina, I. N., Guseva, A. V., Alterman, I. E. and Karnachuk, R. A., 2001. Flavonoid production in transformed Scutellaria baicalensis roots and ways of its regulation. Russian Journal of Plant Physiology 48, 448-452.
Lazinsky, A., Lu, Y., Qiao, W. and Wei, H., 2001. Anti-photocarcinogenic effects of baicalein. Journal of Investigative Dermatology 117, 591.
Liu, Q., Bonness, M. S., Liu, M., Seradge, E., Dixon, R. A. and Mabry, T. J., 1995. Enzymes of B-ring-deoxy flavonoid biosynthesis in elicited cell-cultures of old man cactus (Cephalocereus senilis). Archives of Biochemistry and Biophysics 321, 397-404.
Liu, Q., Dixon, R. A. and Marby, T. J., 1993. Additional flavonoids from elicitor-treated cell-cultures of Cephalocereus senilis. Phytochemistry 34, 167-170.
Liu, Q., Markham, K. R., Paré, P. W., Dixon, R. A. and Mabry, T. J., 1993. Flavonoids from elicitor-treated cell-suspension cultures of Cephalocereus senilis. Phytochemistry 32, 925-928.
56

Markham, K. R. and Mabry, T. J., 1968. The identification of twenty-three 5-deoxy- and ten 5-hydroxy-flavonoids from Baptisia lecontei (Leguminosae). Phytochemistry 7, 791-801.
Markham, K. R., Mabry, T. J. and Swift III, W. T., 1968. New isoflavones from the genus Baptisia (Leguminosae). Phytochemistry 7, 803-808.
Markham, K. R., Mabry, T. J. and Swift III, W. T., 1970. Distribution of flavonoids in the genus Baptisia (Leguminosae). Phytochemistry 9, 2359-2364.
Markham, K. R., Swift III, W. T. and Mabry, T. J., 1968. A new isoflavone glycoside from Baptisia australis. Journal of Organic Chemistry 33, 462-464.
Nes, W. R. and McKean, M. L., 1977. Biochemistry of Steroids and Other Isopentenoids. University Park Press, Baltimore.
Noe, W., Langebartels, C. and Seitz, H. U., 1980. Anthocyanin accumulation and Pal activity in a suspension- culture of Daucus carota L - Inhibition by L-alpha-aminooxy- beta-phenylpropionic acid and t-cinnamic acid. Planta 149, 283-287.
Noe, W. and Seitz, H. U., 1983. Studies on the regulatory role of trans-cinnamic acid on the activity of the phenylalanine ammonia-lyase (Pal) in suspension-cultures of Daucus carota L. Zeitschrift Fur Naturforschung C-a Journal of Biosciences 38, 408-412.
O'Neal, D. and Keller, C. J., 1970. Partial purification and some properties of phenylalanine ammonia-lyase of tobacco (Nicotiana tabacum). Phytochemistry 9, 1373-1383.
Ovadi, J. and Srere, P. A., 1996. Metabolic consequences of enzyme interactions. Cell Biochemistry and Function 14, 249-258.
Ovadi, J. and Srere, P. A., 2000. Macromolecular compartmentation and channeling. International Review of Cytology - a Survey of Cell Biology, 192, 255-280.
Ozeki, Y., Ito, Y., Sasaki, N., Oyanagi, M., Akimoto, H., Chikagawa, Y. and Takeda, J., 2000. Phenylalanine ammonia-lyase genes involved in anthocyanin synthesis and the regulation of its expression in suspension cultured carrot cells. Journal of Plant Research 113, 319-326.
57

Paré, P. W., Dmitrieva, N. and Mabry, T. J., 1991. Phytoalexin aurone induced in Cephalocereus senilis liquid suspension-culture. Phytochemistry 30, 1133-1135.
Paré, P. W., Mischke, C. F., Edwards, R., Dixon, R. A., Norman, H. A. and Mabry, T. J., 1992. Induction of phenylpropanoid pathway enzymes in elicitor- treated cultures of Cephalocereus senilis. Phytochemistry 31, 149-153.
Rommelt, S., Treutter, D., Speakman, J. B. and W., R., 1999. Effects of prohexadione-Ca on the flavonoid metabolism of apple with respect to plant resistance against fireblight. Acta Horticulturae 489, 359-363.
Rossi, M., Meyer, R., Constantinou, P., Caruso, F., Castelbuono, D., O'Brien, M. and Narasimhan, V., 2001. Molecular structure and activity toward DNA of baicalein, a flavone constituent of the Asian herbal medicine "Sho-saiko-to". Journal of Natural Products 64, 26-31.
Shirley, B. W., 1996. Flavonoid biosynthesis: 'New' functions for an 'old' pathway. Trends in Plant Science 1, 377-382.
Shirley, B. W., 1999. Evidence for enzyme complexes in the phenylpropanoid and flavonoid pathways. Physiologia Plantarum 107, 142-149.
Stubbe, J., Ge, J. and Yee, C. S., 2001. The evolution of ribonucleotide reduction revisited. Trends in Biochem Sci 26, 93-99.
Treutter, D., 2001. Biosynthesis of phenolic compounds and its regulation in apple. Plant Growth Regulation 34, 71-89.
58
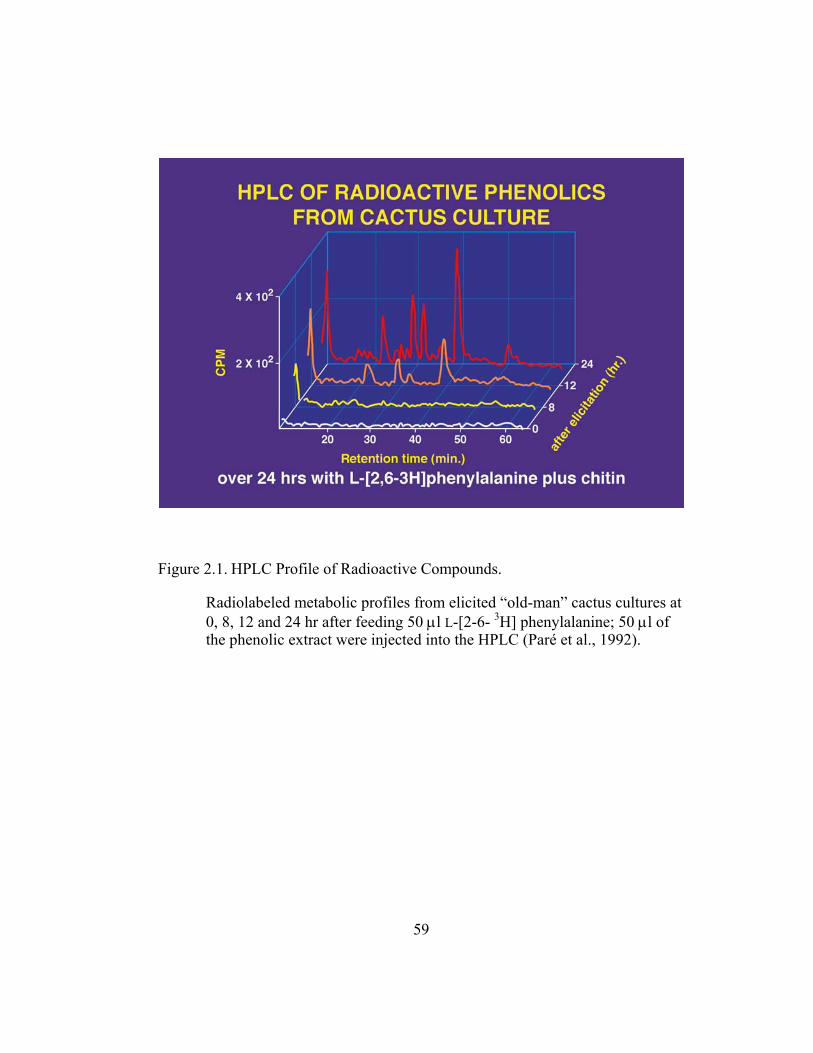
Figure 2.1. HPLC Profile of Radioactive Compounds.
Radiolabeled metabolic profiles from elicited “old-man” cactus cultures at 0, 8, 12 and 24 hr after feeding 50 µl L-[2-6- 3H] phenylalanine; 50 µl of the phenolic extract were injected into the HPLC (Paré et al., 1992).
59
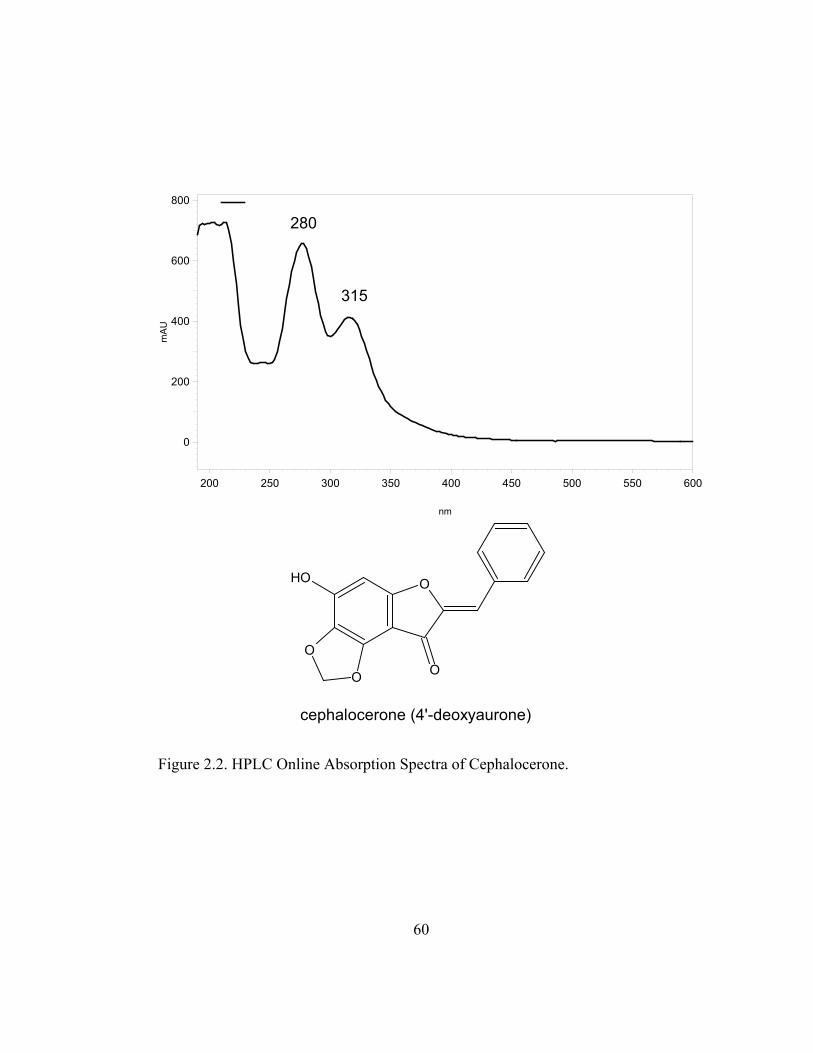
Spectrum at time 20.50 min.
nm
200 250 300 350 400 450 500 550 600
mAU
0
200
400
600
800 20.50 min
280
315
O
O
OH
OO
cephalocerone (4'-deoxyaurone)
Figure 2.2. HPLC Online Absorption Spectra of Cephalocerone.
60
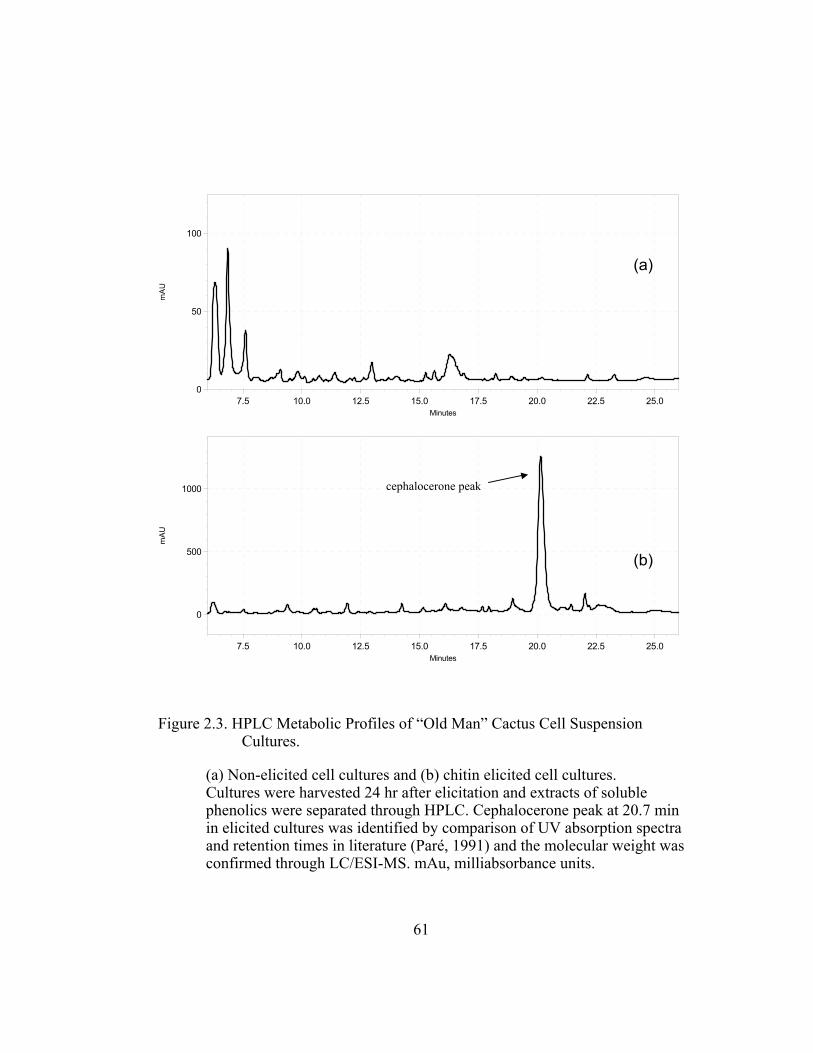
Minutes7.5 10.0 12.5 15.0 17.5 20.0 22.5 25.0
mAU
0
50
100
Minutes7.5 10.0 12.5 15.0 17.5 20.0 22.5 25.0
mAU
0
500
1000
(a)
cephalocerone peak
(b)
Figure 2.3. HPLC Metabolic Profiles of “Old Man” Cactus Cell Suspension Cultures.
(a) Non-elicited cell cultures and (b) chitin elicited cell cultures. Cultures were harvested 24 hr after elicitation and extracts of soluble phenolics were separated through HPLC. Cephalocerone peak at 20.7 min in elicited cultures was identified by comparison of UV absorption spectra and retention times in literature (Paré, 1991) and the molecular weight was confirmed through LC/ESI-MS. mAu, milliabsorbance units.
61
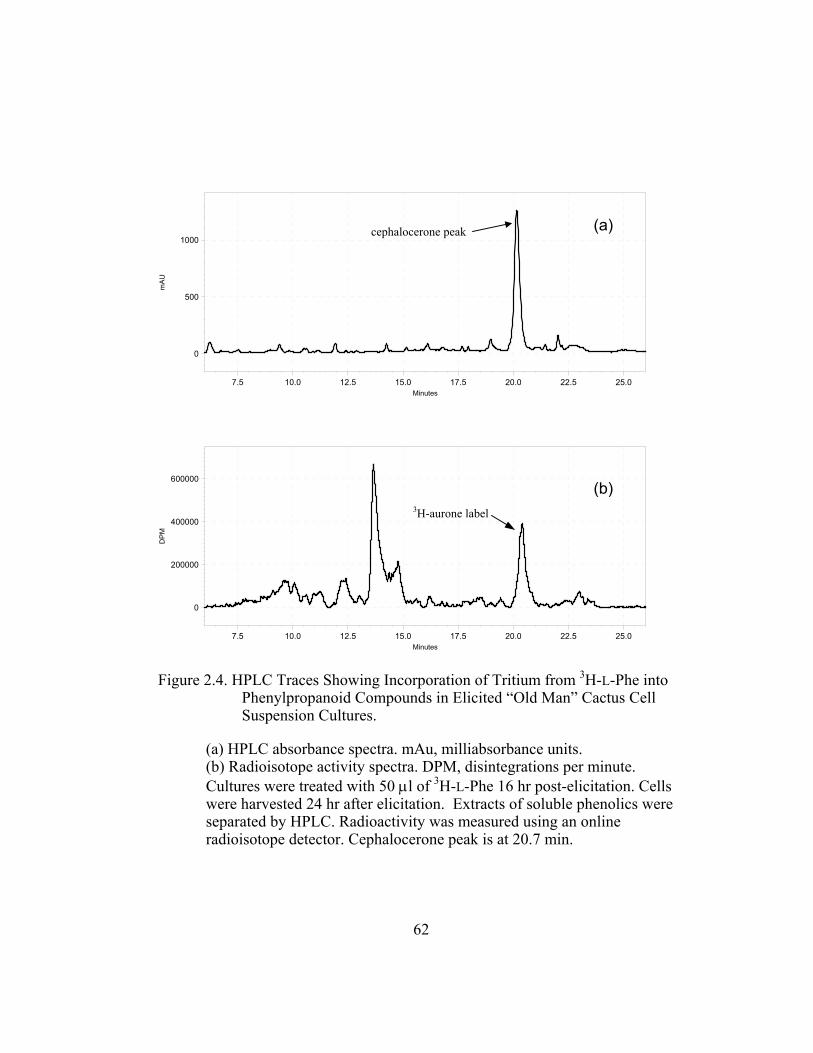
Minutes7.5 10.0 12.5 15.0 17.5 20.0 22.5 25.0
mAU
0
500
1000
(a) cephalocerone peak
Minutes7.5 10.0 12.5 15.0 17.5 20.0 22.5 25.0
DPM
0
200000
400000
600000
(b) 3H-aurone label
Figure 2.4. HPLC Traces Showing Incorporation of Tritium from 3H-L-Phe into Phenylpropanoid Compounds in Elicited “Old Man” Cactus Cell Suspension Cultures.
(a) HPLC absorbance spectra. mAu, milliabsorbance units. (b) Radioisotope activity spectra. DPM, disintegrations per minute. Cultures were treated with 50 µl of 3H-L-Phe 16 hr post-elicitation. Cells were harvested 24 hr after elicitation. Extracts of soluble phenolics were separated by HPLC. Radioactivity was measured using an online radioisotope detector. Cephalocerone peak is at 20.7 min.
62
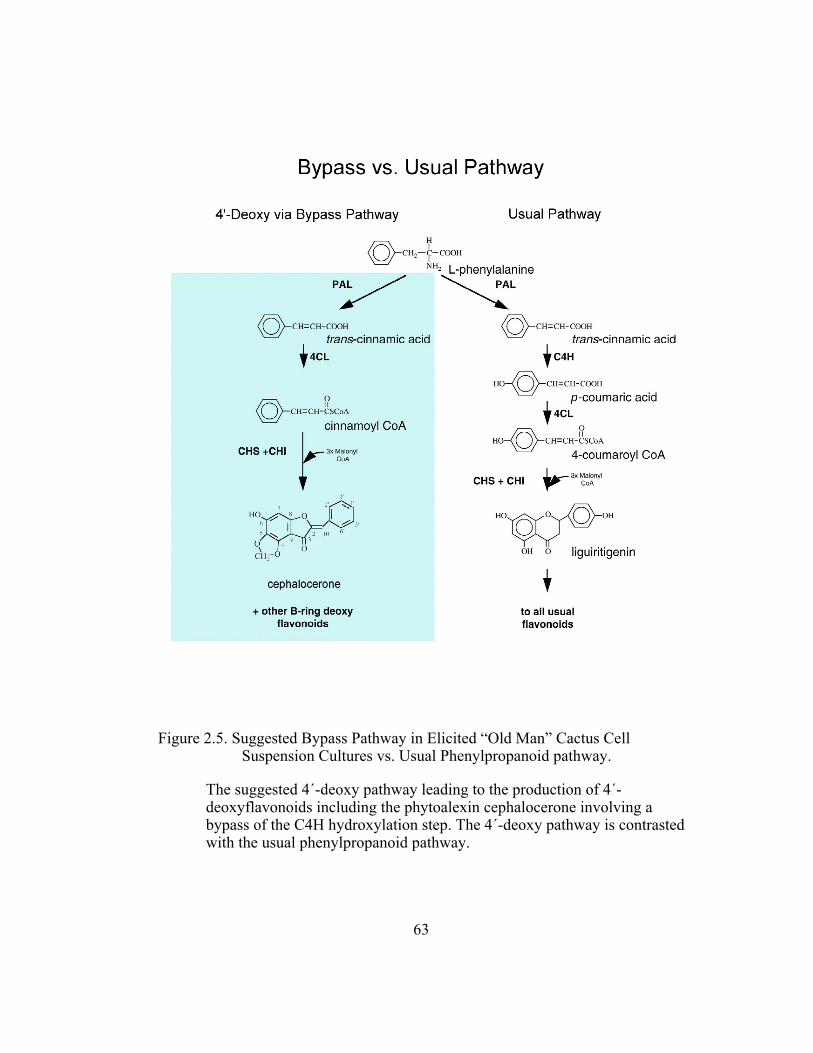
Figure 2.5. Suggested Bypass Pathway in Elicited “Old Man” Cactus Cell Suspension Cultures vs. Usual Phenylpropanoid pathway.
The suggested 4´-deoxy pathway leading to the production of 4´-deoxyflavonoids including the phytoalexin cephalocerone involving a bypass of the C4H hydroxylation step. The 4´-deoxy pathway is contrasted with the usual phenylpropanoid pathway.
63
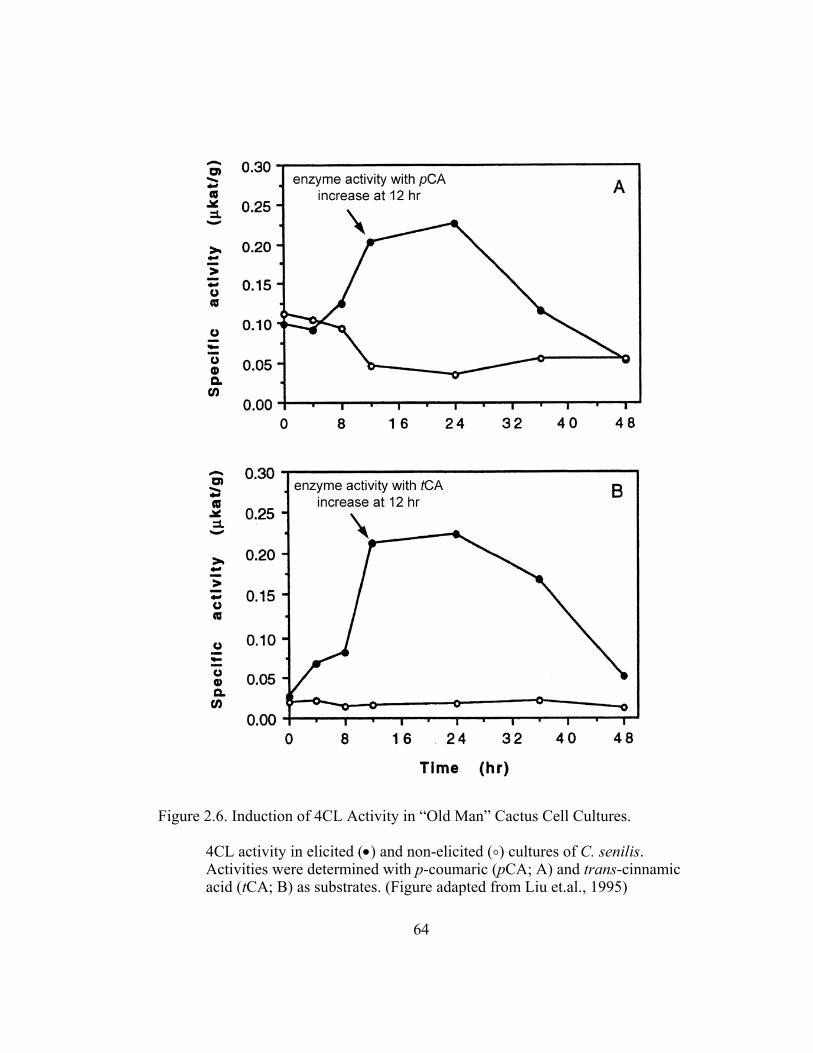
Figure 2.6. Induction of 4CL Activity in “Old Man” Cactus Cell Cultures.
4CL activity in elicited (•) and non-elicited (o) cultures of C. senilis. Activities were determined with p-coumaric (pCA; A) and trans-cinnamic acid (tCA; B) as substrates. (Figure adapted from Liu et.al., 1995)
64
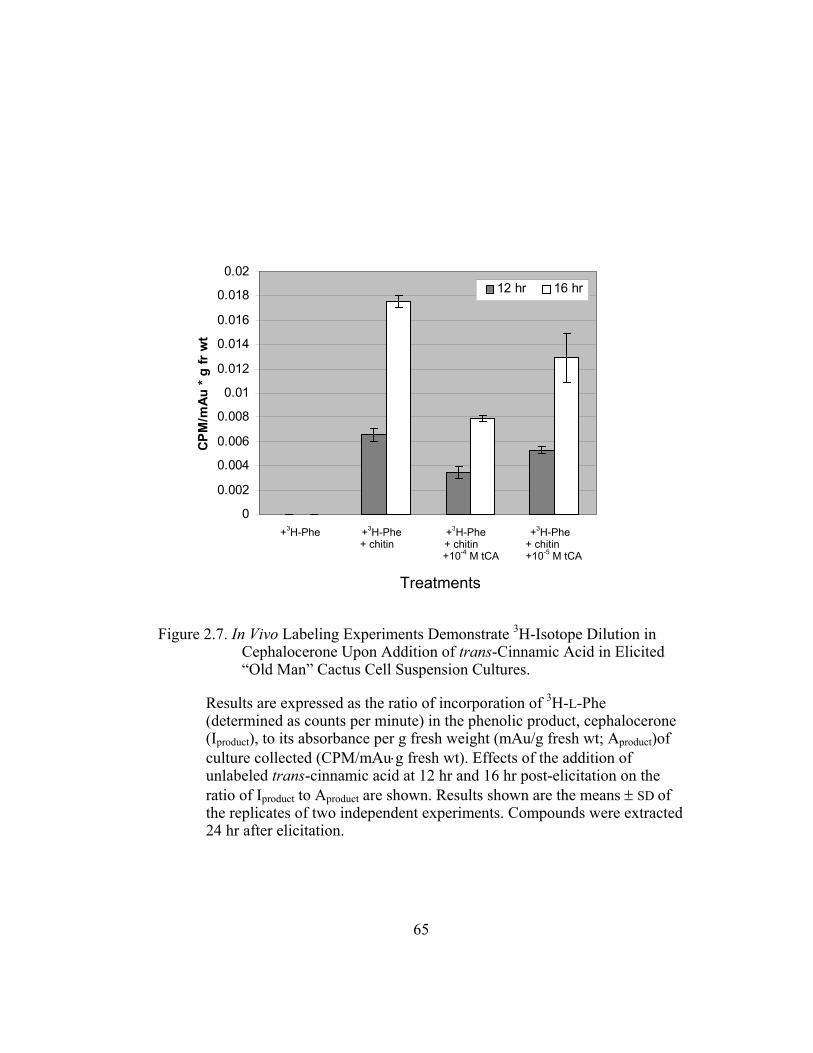
0
0.002
0.004
0.006
0.008
0.01
0.012
0.014
0.016
0.018
0.02
CPM
/mAu
* g
fr w
t
12 hr 16 hr
+3H-Phe +3H-Phe +3H-Phe +3H-Phe + chitin + chitin + chitin +10-4 M tCA +10-5 M tCA
Treatments
Figure 2.7. In Vivo Labeling Experiments Demonstrate 3H-Isotope Dilution in Cephalocerone Upon Addition of trans-Cinnamic Acid in Elicited “Old Man” Cactus Cell Suspension Cultures.
Results are expressed as the ratio of incorporation of 3H-L-Phe (determined as counts per minute) in the phenolic product, cephalocerone (Iproduct), to its absorbance per g fresh weight (mAu/g fresh wt; Aproduct)of culture collected (CPM/mAu⋅g fresh wt). Effects of the addition of unlabeled trans-cinnamic acid at 12 hr and 16 hr post-elicitation on the ratio of Iproduct to Aproduct are shown. Results shown are the means ± SD of the replicates of two independent experiments. Compounds were extracted 24 hr after elicitation.
65
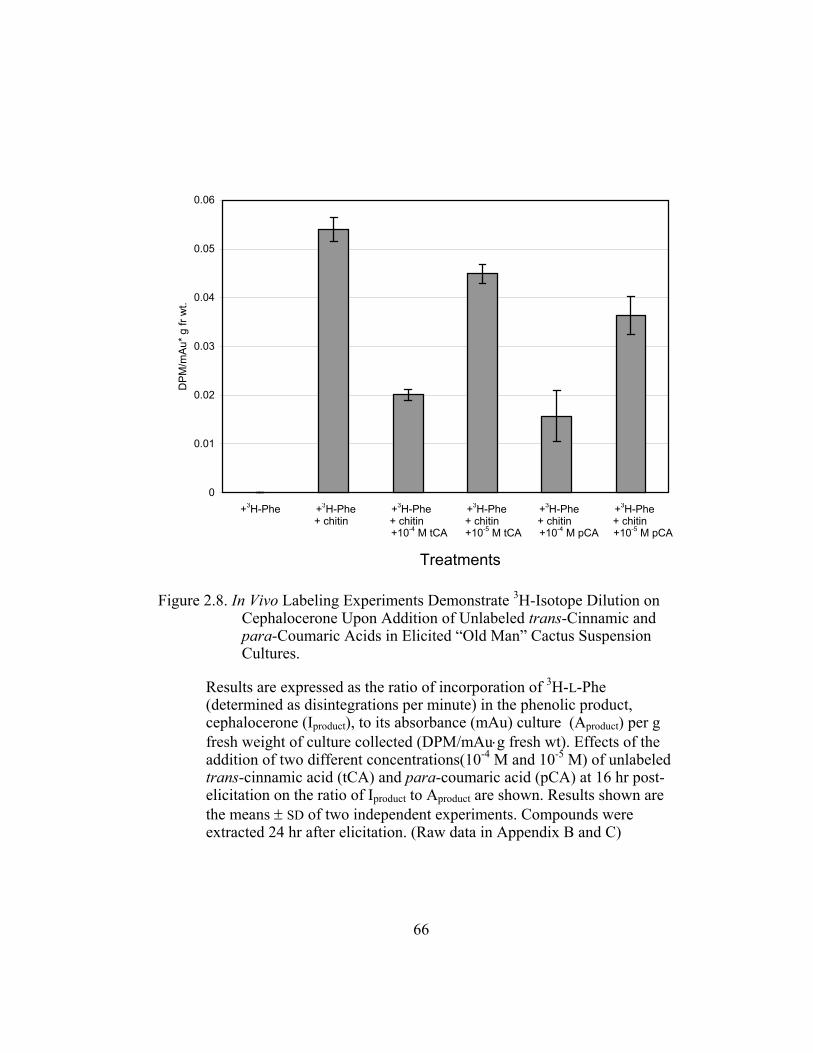
0
0.01
0.02
0.03
0.04
0.05
0.06D
PM/m
Au* g
fr w
t.
+3H-Phe +3H-Phe +3H-Phe +3H-Phe +3H-Phe +3H-Phe + chitin + chitin + chitin + chitin + chitin +10-4 M tCA +10-5 M tCA +10-4 M pCA +10-5 M pCA
Treatments
Figure 2.8. In Vivo Labeling Experiments Demonstrate 3H-Isotope Dilution on Cephalocerone Upon Addition of Unlabeled trans-Cinnamic and para-Coumaric Acids in Elicited “Old Man” Cactus Suspension Cultures.
Results are expressed as the ratio of incorporation of 3H-L-Phe (determined as disintegrations per minute) in the phenolic product, cephalocerone (Iproduct), to its absorbance (mAu) culture (Aproduct) per g fresh weight of culture collected (DPM/mAu⋅g fresh wt). Effects of the addition of two different concentrations(10-4 M and 10-5 M) of unlabeled trans-cinnamic acid (tCA) and para-coumaric acid (pCA) at 16 hr post-elicitation on the ratio of Iproduct to Aproduct are shown. Results shown are the means ± SD of two independent experiments. Compounds were extracted 24 hr after elicitation. (Raw data in Appendix B and C)
66
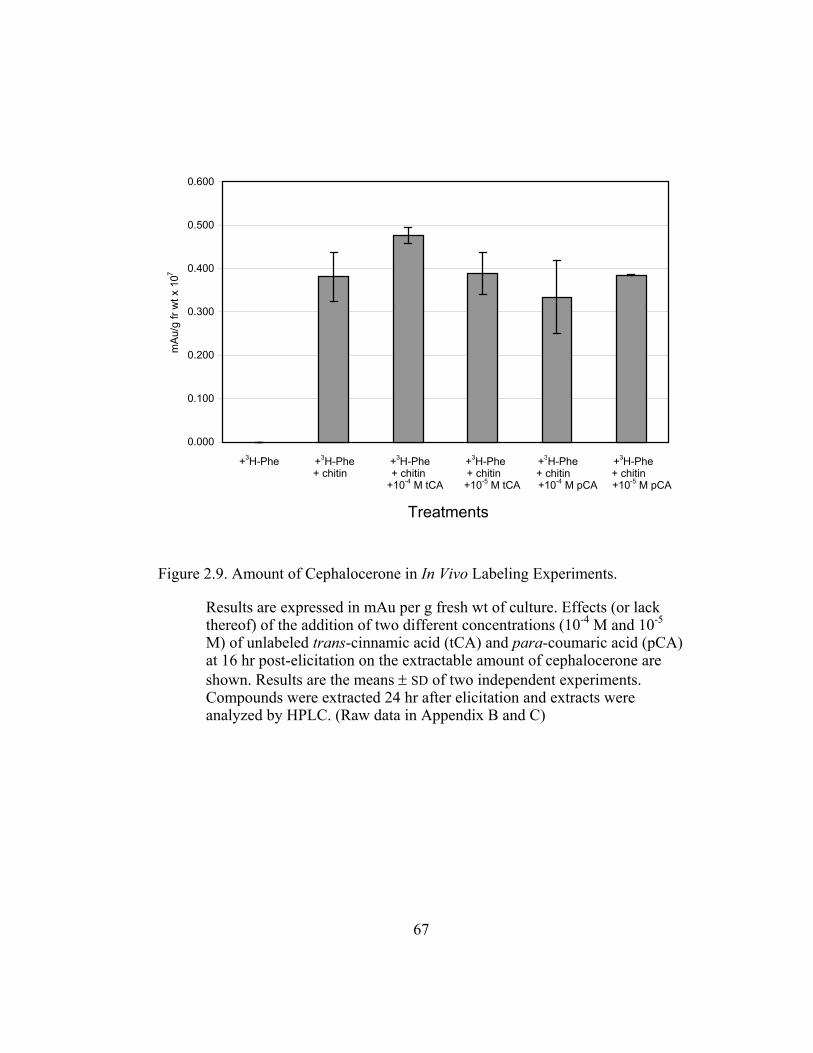
0.000
0.100
0.200
0.300
0.400
0.500
0.600m
Au/g
fr w
t x 1
07
+3H-Phe +3H-Phe +3H-Phe +3H-Phe +3H-Phe +3H-Phe + chitin + chitin + chitin + chitin + chitin +10-4 M tCA +10-5 M tCA +10-4 M pCA +10-5 M pCA
Treatments
Figure 2.9. Amount of Cephalocerone in In Vivo Labeling Experiments.
Results are expressed in mAu per g fresh wt of culture. Effects (or lack thereof) of the addition of two different concentrations (10-4 M and 10-5 M) of unlabeled trans-cinnamic acid (tCA) and para-coumaric acid (pCA) at 16 hr post-elicitation on the extractable amount of cephalocerone are shown. Results are the means ± SD of two independent experiments. Compounds were extracted 24 hr after elicitation and extracts were analyzed by HPLC. (Raw data in Appendix B and C)
67
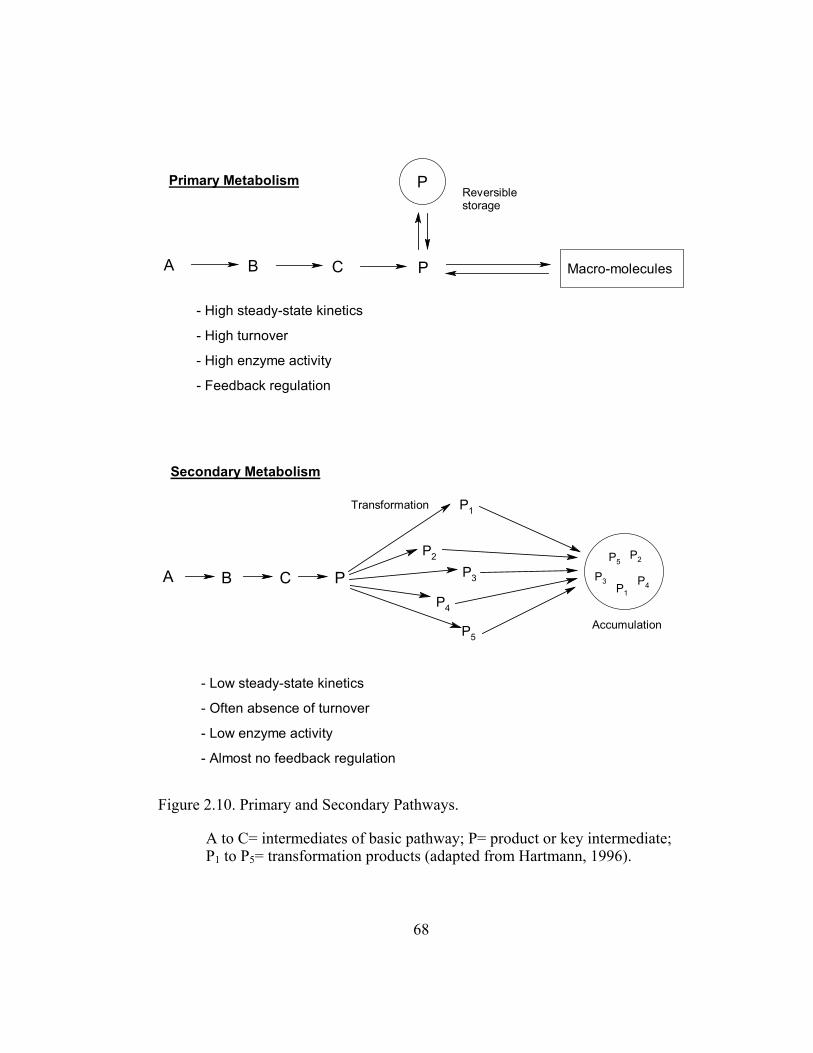
Primary Metabolism
A B C P
P
Macro-molecules
- High steady-state kinetics
- High turnover
- High enzyme activity
- Feedback regulation
Secondary Metabolism
A B C P
P1
P2
P3
P4
P5
P5P2
P3P1
P4
- Low steady-state kinetics
- Often absence of turnover
- Low enzyme activity
- Almost no feedback regulation
Transformation
Accumulation
Reversible storage
Figure 2.10. Primary and Secondary Pathways.
A to C= intermediates of basic pathway; P= product or key intermediate; P1 to P5= transformation products (adapted from Hartmann, 1996).
68
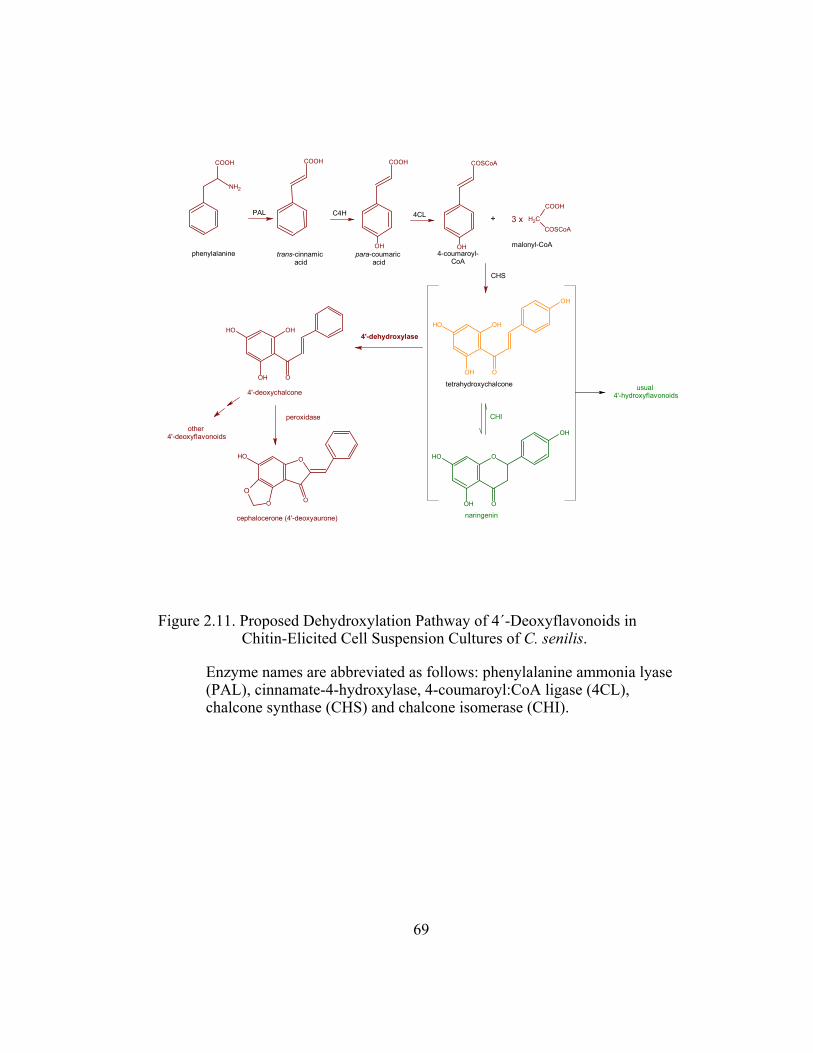
COOH
NH2
COOH COOH
OH
COSCoA
OH
COOH
CH2
COSCoA3 x
4'-dehydroxylase
O
O
OH
OO
cephalocerone (4'-deoxyaurone)
OH
OH
OH
O
4'-deoxychalcone
peroxidaseother
4'-deoxyflavonoids
PAL C4H 4CL
phenylalanine trans-cinnamic acid
para-coumaric acid
4-coumaroyl-CoA
+
malonyl-CoA
CHS
OH
OH
OH
OH
O
tetrahydroxychalcone
O
OH
OH
OH
O
naringenin
CHI
usual 4'-hydroxyflavonoids
Figure 2.11. Proposed Dehydroxylation Pathway of 4´-Deoxyflavonoids in Chitin-Elicited Cell Suspension Cultures of C. senilis.
Enzyme names are abbreviated as follows: phenylalanine ammonia lyase (PAL), cinnamate-4-hydroxylase, 4-coumaroyl:CoA ligase (4CL), chalcone synthase (CHS) and chalcone isomerase (CHI).
69
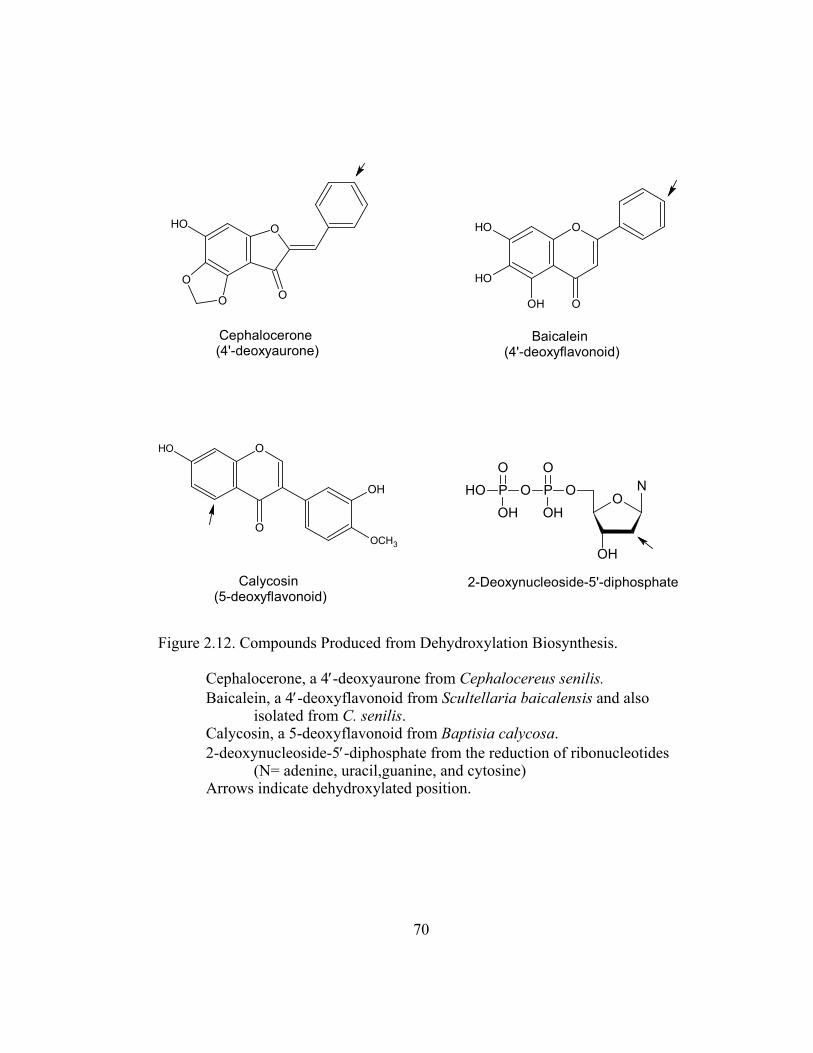
O
OO
O
OH
Cephalocerone (4'-deoxyaurone)
Baicalein (4'-deoxyflavonoid)
O
OOH
OH
OH
OH
OCH3
O
O
OH
Calycosin (5-deoxyflavonoid)
NOO
OH
PO
OHOH
OHO
OP
2-Deoxynucleoside-5'-diphosphate
Figure 2.12. Compounds Produced from Dehydroxylation Biosynthesis.
Cephalocerone, a 4′-deoxyaurone from Cephalocereus senilis. Baicalein, a 4′-deoxyflavonoid from Scultellaria baicalensis and also
isolated from C. senilis. Calycosin, a 5-deoxyflavonoid from Baptisia calycosa.
2-deoxynucleoside-5′-diphosphate from the reduction of ribonucleotides (N= adenine, uracil,guanine, and cytosine)
Arrows indicate dehydroxylated position.
70

Figure 2.13. Tissue Culture Propagation of “Old Man” Cactus.
Callus cultures and (B) Cell suspension cultures.
71

Figure 2.14. HPLC with Radioisotope Detector.
72

Figure 2.15. Liquid Chromatography/Electrospray Ionisation- Mass Spectrometry.
(LC/ESI-MS; Department of Chemistry, University of Texas at Austin).
73

Chapter 3
Conclusions and Summary
A. STUDIES COMPLETED
The flavonoid biosynthetic pathway of Cephalocereus senilis, “old man”
cactus, serves as a useful model for studying metabolic regulation. The unusual
production of 4'-deoxyflavonoids in elicited liquid cultures of “old man” cactus
suggests a novel 4'-dehydroxylation step in the elicited phenylpropanoid pathway.
HPLC metabolic profiles and isotope labeling studies were developed, and the
mechanism leading to the 4'-deoxyaurone, cephalocerone, was proposed and the
“bypass” theory is considered to be highly unlikely (Figure 3.1).
1. Metabolic Profiling
Previous studies on C. senilis chitin-elicited cell suspension cultures
demonstrate that a group of 4'-deoxyphenolics was synthesized de novo through
the phenylpropanoid pathway (Paré et al., 1991). In addition to cephalocerone,
eight other 4'-deoxyflavonoids were isolated from chitin-elicited “old man” cactus
cell suspension cultures (Liu et al., 1993; Liu et al., 1994; Liu et al., 1993).
Consequently, only 4'-hydroxylated flavonoids were isolated from whole stems of
C. senilis (Liu et al., 1994). A method in extracting cell cultures was established
to reflect an accurate metabolic profile with cephalocerone identified as the major
peak. Cephalocerone, aside from being a 4'-deoxyflavonoid, also has phytoalexin
74

properties and may be abundantly produced by “old man” cactus cell suspension
cultures as a defense response to pathogen infection (Paré et al., 1991).
Identification of cephalocerone in the metabolic profiles was made through
known UV absorption spectra and retention times in literature (Paré et al., 1992)
and confirmed by LC/ESI-MS.
2. Evidence for the Novel Chitin- Elicited 4´-Dehydroxylation Step
It has been shown that the elicited suspension cultures have all the
necessary enzymatic activities needed for the conversion of L-phenylalanine to
the B-ring deoxyflavonoids, a single form of CoA ligase that is unusually active
with cinnamic acid, a chalcone synthase exhibiting activity towards cinnamoyl
CoA, and two isoforms of chalcone isomerase showing activities to the 4´-
hydroxyl- and 4´-deoxychalcones (Liu et al., 1995; Paré et al., 1992). The
simplest explanation would be a reduction in the activity of C4H, but enzymatic
studies in Dr. Mabry’s group has shown that C4H activity is not significantly
reduced in elicited cactus cultures (Liu et al., 1995). This result suggests
metabolic channeling, specifically a “bypass” of microsomal C4H in the
biosynthetic pathway from PAL to CoA ligase (Figure 3.1). Another possibility
could involve the normal biosynthesis of the 2´,4,4´,6´-tetrahydroxychalcone with
subsequent induction of a specific 4´-dehydroxylase to remove the 4´-hydroxyl
group thereby allowing selective formation of the B-ring deoxyflavonoids upon
elicitation (Figure 3.1).
75

Although dehydroxylation reactions in different biosynthetic pathways are
not unprecedented, the 4´-dehydroxylation step in the flavonoid pathway is almost
not known. A 4´-hydroxylation “bypass” seems much simpler (see Mabry’s
comments in Chapter 2) but it has more extensive implications because of the role
of the 4´-hydroxylation by C4H in the formation of lignin. Conversion of
cinnamic acid and other phenolic acid derivatives to the alcohol monomers that
incorporate into lignin (Figures 3.2 and 3.3), is a process crucial to lignin
formation since the 4´-hydroxyl groups are directly involved in the linkages of
lignin (Freudenberg and Neish, 1968; Whetten and Sederoff, 1995). It appears
more unlikely that the 4´-hydroxylation step in the phenylpropanoid pathway is
bypassed as this bypass would probably affect lignin formation as well. From an
evolutionary point of view, the 4´-hydroxylation step may be genetically
constrained. For a plant to accumulate a set of defense compounds, it would make
more sense to have back to back hydroxylation/dehydroxylation reactions (see
Paré’s comments in Chapter 2) than to bypass an important hydroxylation step
that may affect and downgrade an important function such as lignin biosynthesis.
A detailed investigation on the biosynthesis of 4´-deoxyflavonoids was
conducted using isotope dilution experiments on elicited “old man” cactus cell
cultures. Timed isotope dilution studies with simultaneous addition of 3H-L-Phe
and unlabeled trans-cinnamic acid at 12 hr and 16 hr post-elicitation show more
incorporation of unlabeled trans-cinnamic acid at 16 hr. This confirms past
enzymological studies that indicate an increased enzyme activity starting at 12 hr
and maximizing at 16 hr after elicitation with chitin. There is no metabolic
76

channeling between PAL and C4H, and PAL and 4CL since exogenous unlabeled
trans-cinnamic acid and para-coumaric acid equilibrated with the endogenous 3H-
labeled precursor pools. Addition of unlabeled para-coumaric acid diluted the 3H-
isotopic label on cephalocerone without changing the amount of cephalocerone
produced. This shows that the cinnamic acid-4-hydroxylase bypass does not occur
and metabolic channeling between PAL and 4CL does not exist. Our results show
evidence for the existence of a novel dehydroxylation activity downstream of
4CL, probably involving a dehydroxylase that removes the 4´-hydroxyl group
from a tetrahydroxychalcone before conversion to the 4´-deoxyaurone (Figure
3.1).
B. FUTURE RESEARCH
1. Isolation of 4´-Deoxyflavonoid Standards
A more accurate metabolic profile of elicited “old man” cactus cell
cultures will be more informative if most, if not all, HPLC metabolic profile
peaks are accounted for. A significant amount of flavonoid standards from both
elicited and non-elicited cultures will be isolated in order to characterize
metabolic profiles.
2. Metabolic Profiling with Other Labeled Precursors
While metabolic profiles of 3H-L-Phe fed elicited “old man” cactus cell
cultures were obtained for data presented in this dissertation, metabolic profile
experiments using 14C- labeled trans-cinnamic acid and p-coumaric acid were
77

unsuccessful. The synthesis procedures used for both compounds provided low
yield, not just in compound amounts but in the radioactive label obtained as well.
For example, the radioactivity of commercially available 3H-L-Phe used in this
project is at 124 Ci/mmole which is relatively high compared to 56 mCi/mmole of
the commercially available 2-14C-malonic acid used as a precursor in the
synthesis of p-coumaric acid. The literature values for the yield of the p-coumaric
acid synthesis procedure is at 1-14% and the values obtained in the lab are
calculated at 5%. In order for us to get close to the level of the 3H-L-Phe used, we
would need about 5 ml of the labeled malonic acid to do a complete experiment.
This will not be feasible because the cost of 2-14C-malonic acid is beyond our
budget at $1400/ml (ICN Pharmaceuticals, Costa Mesa, CA). In general, plants
incorporate labeled compounds very poorly, often as little as 10-2 to 10-4 percent.
This can be increased to 1 percent in cell suspension cultures and to as much as 10
percent in cell free in vitro experiments with the use of microsomes (Mann,
1987). Microsome isolation procedures for “old man” cactus cell cultures will be
developed and metabolic profile studies using labeled trans-cinnamic acid and p-
coumaric will be conducted using these microsomes. Microsome studies will need
lesser amount of labeled compounds compared to isotope experiments done on
cell suspension cultures.
3. Dual-Labeling Experiments
Metabolic flux through the phenylpropanoid pathway can be further
characterized in detail using co-administration of differentially labeled
78

compounds. Microsomes from “old man” cactus cell suspension cultures will be
used in dual-labeling studies and 3H/14C incorporation ratios from the various
metabolites will be obtained to investigate metabolic channeling in the system.
For example, cell suspension cultures will be treated with both 3H- trans-cinnamic
acid and 14C-p-coumaric acid. If 3H/14C incorporation ratios are less than one, the
preferential utilization of p-coumaric acid versus trans-cinnamic acid would be
indicated and provide more evidence for the 4´-dehydoxylation step.
5. 4´-Deoxy-Biosynthesis Enzymes
The specific enzymes for aurone biosynthesis are not known. Studies
show that chalcones are the precursor of aurones (Wong, 1966). It has been
suggested that a peroxidase-like enzyme converts chalcones into aurones but this
reaction step is not well characterized (Goodwin and Mercer, 1983). The
enzymatic details of the dehydroxylation step (whether a specific dehydroxylase-
like enzyme is induced) to the 4´-deoxyaurone also needs to be established.
Incorporation of labeled chalcones in metabolic profile studies will help provide
evidence for the location of the 4´-dehydroxylation step. Skullcap (Scutellaria
baicalensis) cell cultures and whole plants are known to produce 4´-
deoxyflavonoids (e.g. baicalein) (Kuzovkina et al., 2001). 4´-deoxy biosynthesis
studies will also be extended to skullcap for comparison with the “old man”
cactus 4´-dehydroxylation pathway.
79

6. 4´-Deoxyflavonoid Survey
It is not known whether the 4´-deoxyflavonoid biosynthesis is exclusive to
cell cultures of “old man” cactus. Opuntia is known to be susceptible to fungal
infection (Russell and Felker, 1987). Phytoalexin and 4´-deoxyflavonoid
biosynthesis research will be extended to Opuntia species and possibly to other
cactus species closely related to “old man” cactus.
7. Molecular and Genetic Studies
Signal transduction events underlying 4´-deoxyflavonoid biosynthesis will
be determined by using molecular techniques investigating the molecular
“switches” that are activated by chitin elicitation. For example, once the 4´-
dehydroxylation step is fully characterized, cDNA clones of the different
phenylpropanoid enzymes including the dehydroxylase will be obtained by
screening elicited cell culture libraries with heterologous probes. Once clones are
obtained, in vitro experiments on enzymatic activity can be executed and
compared to the enzyme activities in vivo. If substrate specificities and Km values
obtained from these molecular studies are different from those in vivo, a more
complex regulatory mechanism such as phosphorylation or metabolic
compartmentation of the enzymes may exist and will be investigated further.
80

LITERATURE CITED
Freudenberg, K. and Neish, A.C., 1968. Constitution and Biosynthesis of Lignin.
Springer-Verlag, New York.
Goodwin, T. W. and Mercer, E. I., 1983. Introduction to Plant Biochemistry. Pergamon Press, New York.
Kuzovkina, I. N., Guseva, A. V., Alterman, I. E. and Karnachuk, R. A., 2001. Flavonoid production in transformed Scutellaria baicalensis roots and ways of its regulation. Russian Journal of Plant Physiology 48, 448-452.
Liu, Q., Bonness, M. S., Liu, M., Seradge, E., Dixon, R. A. and Mabry, T. J., 1995. Enzymes of B-ring-deoxy flavonoid biosynthesis in elicited cell-cultures of old man cactus (Cephalocereus senilis). Archives of Biochemistry and Biophysics 321, 397-404.
Liu, Q., Dixon, R. A. and Marby, T. J., 1993. Additional flavonoids from elicitor-treated cell-cultures of Cephalocereus senilis. Phytochemistry 34, 167-170.
Liu, Q., Liu, M., Mabry, T. J. and Dixon, R. A., 1994. Flavonol glycosides from Cephalocereus senilis. Phytochemistry 31, 229-331.
Liu, Q., Markham, K. R., Paré, P. W., Dixon, R. A. and Mabry, T. J., 1993. Flavonoids from elicitor-treated cell-suspension cultures of Cephalocereus senilis. Phytochemistry 32, 925-928.
Mann, J., 1987. Secondary metabolism. Clarendon Press, Oxford.
Paré, P. W., Dmitrieva, N. and Mabry, T. J., 1991. Phytoalexin aurone induced in Cephalocereus senilis liquid suspension-culture. Phytochemistry 30, 1133-1135.
Paré, P. W., Mischke, C. F., Edwards, R., Dixon, R. A., Norman, H. A. and Mabry, T. J., 1992. Induction of phenylpropanoid pathway enzymes in elicitor- treated cultures of Cephalocereus senilis. Phytochemistry 31, 149-153.
Russell, C. E. and Felker, P., 1987. The prickly-pears (Opuntia Spp Cactaceae) - A source of human and animal food in semi-arid regions. Economic Botany 41, 433-445.
81

Whetten, R. and Sederoff, R., 1995. Lignin biosynthesis. Plant Cell 7, 1001-1013.
Wong, E., 1966. Occurence and biosynthesis of 4',6'-dihydroxyaurone in soybean. Phytochemistry 5, 463-467.
82
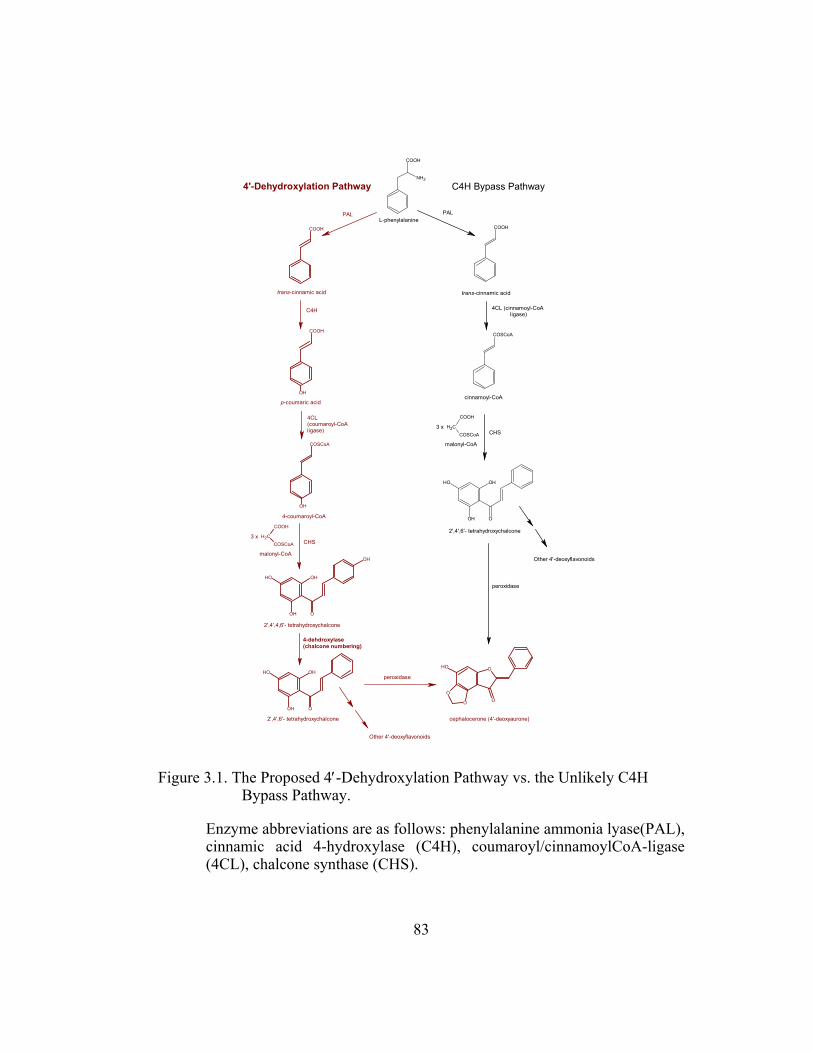
COOH
NH2
COOH
COSCoA
L-phenylalaninePAL
OH
OH
OH
O
trans-cinnamic acid
4CL (cinnamoyl-CoA ligase)
COOH
CH2COSCoA CHS
malonyl-CoA
3 x
2',4',6'- tetrahydroxychalcone
peroxidase
Other 4'-deoxyflavonoids
C4H Bypass Pathway
cinnamoyl-CoA
COOH
OH
COOH
CH2COSCoA
COOH
COSCoA
OH
OH
OH
OH
OH
O
OH
OH
OH
O
O
O
OH
OO
PAL
trans-cinnamic acid
C4H
4CL (coumaroyl-CoA ligase)
CHS
p-coumaric acid
malonyl-CoA
3 x
4-coumaroyl-CoA
2',4',4,6'- tetrahydroxychalcone
4-dehdroxylase (chalcone numbering)
2',4',6'- tetrahydroxychalcone
peroxidase
Other 4'-deoxyflavonoids
cephalocerone (4'-deoxyaurone)
4'-Dehydroxylation Pathway
Figure 3.1. The Proposed 4′-Dehydroxylation Pathway vs. the Unlikely C4H Bypass Pathway.
Enzyme abbreviations are as follows: phenylalanine ammonia lyase(PAL), cinnamic acid 4-hydroxylase (C4H), coumaroyl/cinnamoylCoA-ligase (4CL), chalcone synthase (CHS).
83

Figure 3.2. Synthesis Pathway of Lignin Monomers from Phenylpropanoid Precursors.
Enzyme abbreviations: C4H, cinnamate 4-hydroxylase; 4CL, 4-coumaroyl-CoA ligase.
84

Figure 3.3. Partial Structure of Spruce Lignin.
(Figure adapted from Freudenberg and Neish, 1968).
85

Appendix A
Phytoalexin biosynthesis
The chemical structures of phytoalexins are diverse and no pathway is
known to be solely responsible for their synthesis. Most of the phytoalexins are
derived from the phenylpropanoid and terpenoid pathways with phytoalexin
pathway-specific enzymes branching out from the major pathways
(Hammerschmidt, 1999). The shikimic acid pathway yields essential vitamins,
cofactors and the aromatic amino acids, tyrosine, tryptophan and phenylalanine.
L-Phenylalanine is the first substrate of the phenylpropanoid pathway and is the
B-ring source for all flavonoids and its derivatives (Goodwin and Mercer, 1983).
Phytoalexins from the phenylpropanoid pathway are usually derivatives of the
isoflavonoids formed by chalcone synthase with malonyl CoA and coumaroyl
CoA as substrates (Hammerschmidt, 1999). Medicarpin, an anti-microbial
compound (Figure 1.3) from alfalfa (Medicago sativa L.) is produced via
branching from this pathway (Paiva et al., 1991).
A. ELICITOR SIGNAL
The phytoalexin defense response is induced by a variety of signals, most
of them divided into two groups, the abiotic and biotic elicitors.
Stresses with elicitor activity that are not derived from natural sources are
abiotic. This group of elicitors includes fungicides, detergents, heavy metal salts,
86

and DNA-intercalating agents (Smith, 1996). The production of phytoalexins can
also be observed in a variety of plant stress responses such as UV exposure,
starvation, and wounding.
Phytoalexin synthesis in plant-pathogen interactions is initiated by
compounds produced by the invading micro-organism. The most common of the
biotic elicitors are the components of the pathogen that are released exogenously;
these may represent parts of the cell wall or metabolites that secrete out of the cell
wall. Compounds belonging to the exogenous biotic elicitor class include
glycoproteins, lipids, and lipopolysaccharides, and the most studied class being
the poly- and oligosaccharides.
Fungal cell walls are composed mostly of polymers rich in glucan, chitin
and sugars such as galactose and mannose. Many exogenous elicitors are
derivatives of these saccharides (Hamdam and Dixon, 1986). The glucan elicitor
from the mycelial walls of Phytophthora megasperma Drenchs. var. sojae
Hildeb., a branched β - 1,3-1,6- linked glucan that has 6- and 3,6- linked glycosyl
residues (Figure 1.1), is the most studied among the polysaccharide elicitors
(Ayers et al., 1976). This hepta-β-glucoside elicitor system in soybean (Glycine
max L.) is very structure specific and it induces the synthesis of the phytoalexin
glyceollin (Figure 1.3). Variation of the heptaglucoside structure in various
elicitation studies show the importance of the β-1,6- linked backbone and the β-
1,3 side chain residues in achieving the appropriate response (Smith, 1996). Strict
structural requirements are equally critical in other carbohydrate elicitors such as
chitin and its deacetylated derivative, chitosan (Figure 1.1). Both elicitors are also
87

major components of many pathogens including fungi. The chitosan elicitor
system has been widely studied in peas while chitin is a major elicitor in rice (Ren
and West, 1992). Chitin and chitosan lack the branching complexity of the
heptaglucosides from P. megasperma var. sojae indicating that much of the
elicitor activity is determined by the degree of polymerization (DP) of the
glucosamine residues (Smith, 1996; Svitil and Kirchman, 1998; Walker-Simmons
et al., 1983). The minimum oligomer size of chitin critical for biological activity
in rice is the tetramer, with the N-acetylchitooctaose showing most potency
(Shibuya et al., 1993).
B. ELICITOR RELEASE
The release of lower molecular weight oligomer elicitors are facilitated by
cleavage enzymes present in both the host plant and the pathogen (Smith, 1996).
Increased activities of poly- and oligosaccharide degrading enzymes such as
chitinases and β-1,3-glucanases have been observed in many plants. Constitutive
expression of these enzymes appear to be a part of the overall defense strategy of
the infected plant (Smith, 1996). It has been shown that either singly or in
combination, the activities of chitinases and glucanases inhibit the growth of
pathogenic fungi (Broekaert et al., 1988). Degradation of the fungal cell wall to
inhibit pathogen growth would not be a difficult reason for the activities of the
cleavage enzymes. However it has been shown that the presence of the enzymes
show no toxic effect on the fungus unless plant tissue is present (Yoshikawa et al.,
1990).
88

The release of low molecular weight oligomers early in the infection
process suggest an early warning system in the overall defense strategy of the
plant. In order to avert further damage and pathogen growth to other tissues, the
immediate release of the elicitors signal the proximate plant tissues to produce the
appropriate defense compounds (Walker-Simmons et al., 1983; Yamaguchi et al.,
2000; Yamamoto et al., 1998).
C. SIGNAL TRANSDUCTION
Definite structural requirements for different elicitor systems imply that
specific cellular receptors play a significant role in plant-pathogen interactions.
This kind of specificity is manifested in many signal transduction systems.
Different pathogens may exhibit different levels of toxicity to the same plant and
a variety of responses may occur with exposure to different environmental
stresses. Studies have shown that it is most unlikely for the pathogen-derived
chemical stimuli to be internalized, therefore an intracellular signal must directly
induce phytoalexin release (Smith, 1996). Glucan binding sites on the surface of
soybean cells show very high specificity for fungal β-glucans including the well
studied hepta-β-glucosides from P. sojae. Similar β-glucan receptors also exist in
other legumes such as such as bean (Phaseolus vulgaris L.), lupine (Lupinus
albus L.), and pea (Pisum sativum L.).
Increased ion flux is highly correlated with the phytoalexin response in
elicited soybean cells (Ebel et al., 1995). Secondary messengers such as cytosolic
Ca2+ seem to be necessary in phytoalexin production when transgenic soybean
89

cell cultures expressing aequorin are treated with β-glucan and chitin elicitors
(Mithofer et al., 1999). Furthermore, inhibition of Ca2+-channels in soybean cells,
in the presence of elicitors, lower phytoalexin accumulation and chalcone
synthase activation (Stab and Ebel, 1987). Gene activation of the phytoalexin
defense pathway may involve Ca2+ regulation of an H+-ATPase and protein
kinases (Ebel and Mithofer, 1998).
Increased oxygen uptake or “oxidative burst” has also been observed as a
part of the early response system in elicited plant tissues (Figure 1.2). It has been
suggested that one of the critical elements in the trigger of the plant defense
response is the formation of reactive oxygen species such as •O2- and H2O2
(Mehdy, 1994). Application of H2O2 in bean cells (P. vulgaris) has been observed
to induce transcription of several major phenylpropanoid enzymes such as
phenylalanine ammonia lyase (PAL) and chalcone synthase (CHS) (Mehdy,
1994). However, the same accumulation of PAL and CHS is not observed in
soybean cells in the presence H2O2. Detailed studies in soybean suggest that H2O2
plays an indirect role in the elicitation process, either as an early inducer or as a
signal in a separate but related pathway (Tenhaken et al., 1995).
G-protein cascades, usually involved in growth and intracellular transport,
are also known to play an essential role in the defense signal pathway in rice
(Sano and Ohashi, 1995). Reversible protein phosphorylation patterns in the
cytoplasm and nucleus upon application and removal of elicitor in the presence of
Ca2+ were observed in parsley (Petroselinum crispum (Mill) AW Hill) cell-
suspension cultures (Dietrich et al., 1990). K-252a, a known inhibitor of animal
90

protein kinase blocks characteristic elicitation responses of tomato cells such as
ethylene biosynthesis and PAL activity at any time point in the elicitation process
(Felix et al., 1991). These studies show indirect evidence of a phosphorylation
cascade that needs to be continuous in order for the whole elicitation process to
take place (Figure 1.2).
LITERATURE CITED
Ayers, A. R., Valent, B., Ebel, J. and Albersheim, P., 1976. Host- pathogen
interactions. XI. Composition and structure of wall-released elicitor fractions. Plant Physiology 57, 766-774.
Broekaert, W. F., Vanparijs, J., Allen, A. K. and Peumans, W. J., 1988. Comparison of some molecular, enzymatic and antifungal properties of chitinases from thorn-apple, tobacco and wheat. Physiological and Molecular Plant Pathology 33, 319-331.
Dietrich, A., Mayer, J. E. and Hahlbrock, K., 1990. Fungal elicitor triggers rapid, transient, and specific protein-phosphorylation in parsley cell-suspension cultures. Journal of Biological Chemistry 265, 6360-6368.
Ebel, J., Bhagwat, A. A., Cosio, E. G., Feger, M., Kissel, U., Mithofer, A. and Waldmuller, T., 1995. Elicitor binding-proteins and signal-transduction in the activation of a phytoalexin defense response. Canadian Journal of Botany-Revue Canadienne De Botanique 73, S506-S510.
Ebel, J. and Mithofer, A., 1998. Early events in the elicitation of plant defence. Planta 206, 335-348.
Felix, G., Grosskopf, D. G., Regenass, M. and Boller, T., 1991. Rapid changes of protein-phosphorylation are involved in transduction of the elicitor signal in plant-cells. Proceedings of the National Academy of Sciences of the United States of America 88, 8831-8834.
91

Goodwin, T. W. and Mercer, E. I., 1983. Introduction to Plant Biochemistry. Pergamon Press, New York.
Hamdam, M. A. M. S. and Dixon, R. A., 1986. Fractionation and properties of elicitors of the phenylpropanoid pathway from Colletotrichum lindemuthianum. Physiological and Molecular Plant Pathology 31, 105-121.
Hammerschmidt, R., 1999. Phytoalexins: What have we learned after 60 years? Annual Review of Phytopathology 37, 285-306.
Mehdy, M. C., 1994. Active oxygen species in plant defense against pathogens. Plant Physiology 105, 467-472.
Mithofer, A., Ebel, J., Bhagwat, A. A., Boller, T. and Neuhaus-Url, G., 1999. Transgenic aequorin monitors cytosolic calcium transients in soybean cells challenged with beta-glucan or chitin elicitors. Planta 207, 566-574.
Paiva, N. L., Edwards, R., Sun, Y. J., Hrazdina, G. and Dixon, R. A., 1991. Stress responses in alfalfa (Medicago sativa L) .11. Molecular- cloning and expression of alfalfa isoflavone reductase, a key enzyme of isoflavonoid phytoalexin biosynthesis. Plant Molecular Biology 17, 653-667.
Ren, Y. Y. and West, C. A., 1992. Elicitation of diterpene biosynthesis in rice (Oryza sativa L) by chitin. Plant Physiology 99, 1169-1178.
Sano, H. and Ohashi, Y., 1995. Involvement of small GTP-binding proteins in defense signal-transduction pathways of higher plants. Proceedings of the National Academy of Sciences of the United States of America 92, 4138-4144.
Shibuya, N., Kaku, H., Kuchitsu, K. and Maliarik, M. J., 1993. Identification of a novel high-affinity binding-site for N- acetylchitooligosaccharide elicitor in the membrane-fraction from suspension-cultured rice cells. Febs Letters 329, 75-78.
Smith, C. J., 1996. Accumulation of phytoalexins: Defence mechanism and stimulus response system. New Phytologist 132, 1-45.
Stab, M. R. and Ebel, J., 1987. Effects of Ca2+ on phytoalexin induction by fungal elicitor in soybean cells. Archives of Biochemistry and Biophysics 257, 416-423.
92

Svitil, A. L. and Kirchman, D. L., 1998. A chitin-binding domain in a marine bacterial chitinase and other microbial chitinases: implications for the ecology and evolution of 1,4-beta-glycanases. Microbiology-Uk 144, 1299-1308.
Tenhaken, R., Levine, A., Brisson, L. F., Dixon, R. A. and Lamb, C., 1995. Function of the oxidative burst in hypersensitive disease resistance. Proceedings of the National Academy of Sciences of the United States of America 92, 4158-4163.
Walker-Simmons, M., Hadwiger, L. and Ryan, C. A., 1983. Chitosans and pectic polysaccharides both induce the accumulation of the antifungal phytoalexin pisatin in pea pods and antinutrient proteinase inhibitors in tomato leaves. Biochemical & Biophysical Research Communications 110, 194-9.
Yamaguchi, T., Yamada, A., Hong, N., Ogawa, T., Ishii, T. and Shibuya, N., 2000. Differences in the recognition of glucan elicitor signals between rice and soybean: beta-glucan fragments from the rice blast disease fungus Pyricularia oryzae that elicit phytoalexin biosynthesis in suspension-cultured rice cells. Plant Cell 12, 817-826.
Yamamoto, H., Koga, T., Hayakawa, S., Ohgata, Y., Kurasaki, T. and Tohyama, K., 1998. Effect of chitin application on growth and yield of rice. Japanese Journal of Crop Science 67, 452-456.
Yoshikawa, M., Takeuchi, Y. and Horino, O., 1990. A mechanism for ethylene-induced disease resistance in soybean - Enhanced synthesis of an elicitor-releasing factor, beta-1,3- endoglucanase. Physiological and Molecular Plant Pathology 37, 367-376.
93
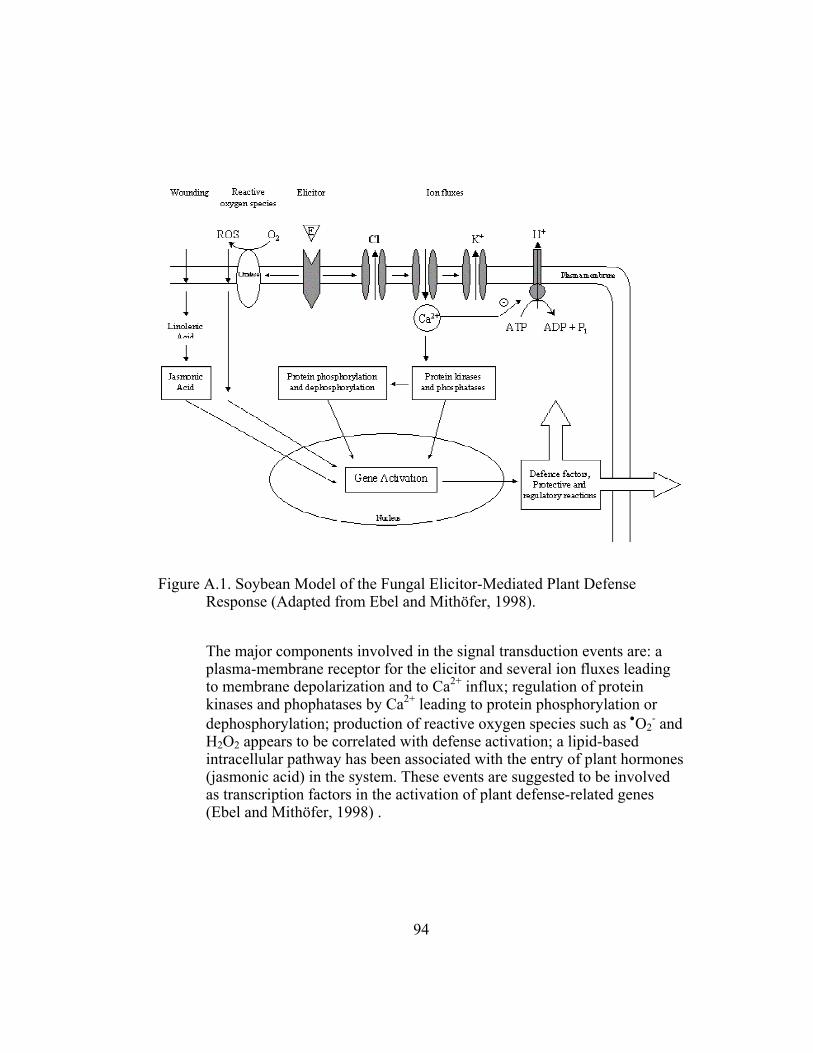
Figure A.1. Soybean Model of the Fungal Elicitor-Mediated Plant Defense
Response (Adapted from Ebel and Mithöfer, 1998).
The major components involved in the signal transduction events are: a plasma-membrane receptor for the elicitor and several ion fluxes leading to membrane depolarization and to Ca2+ influx; regulation of protein kinases and phophatases by Ca2+ leading to protein phosphorylation or dephosphorylation; production of reactive oxygen species such as •O2
- and H2O2 appears to be correlated with defense activation; a lipid-based intracellular pathway has been associated with the entry of plant hormones (jasmonic acid) in the system. These events are suggested to be involved as transcription factors in the activation of plant defense-related genes (Ebel and Mithöfer, 1998) .
94
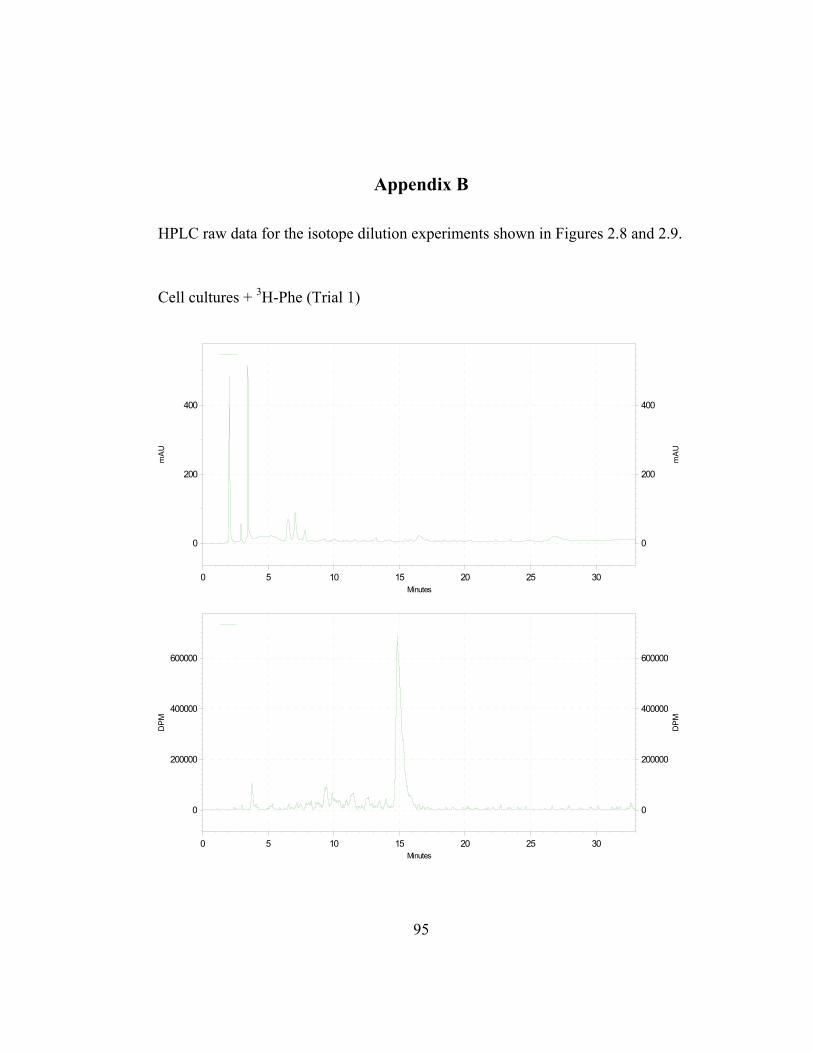
Appendix B
HPLC raw data for the isotope dilution experiments shown in Figures 2.8 and 2.9.
Cell cultures + 3H-Phe (Trial 1)
Minutes0 5 10 15 20 25 30
mAU
0
200
400
mAU
0
200
400
4: 280 nm, 4 nm3/28/02 6:57:19 PM_C161_DPM_1Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
200000
400000
600000
DPM
0
200000
400000
600000
171 A3/28/02 6:57:19 PM_C161_DPM_1Shirley171RadioIsoflavprofile_Bmethod.met
95
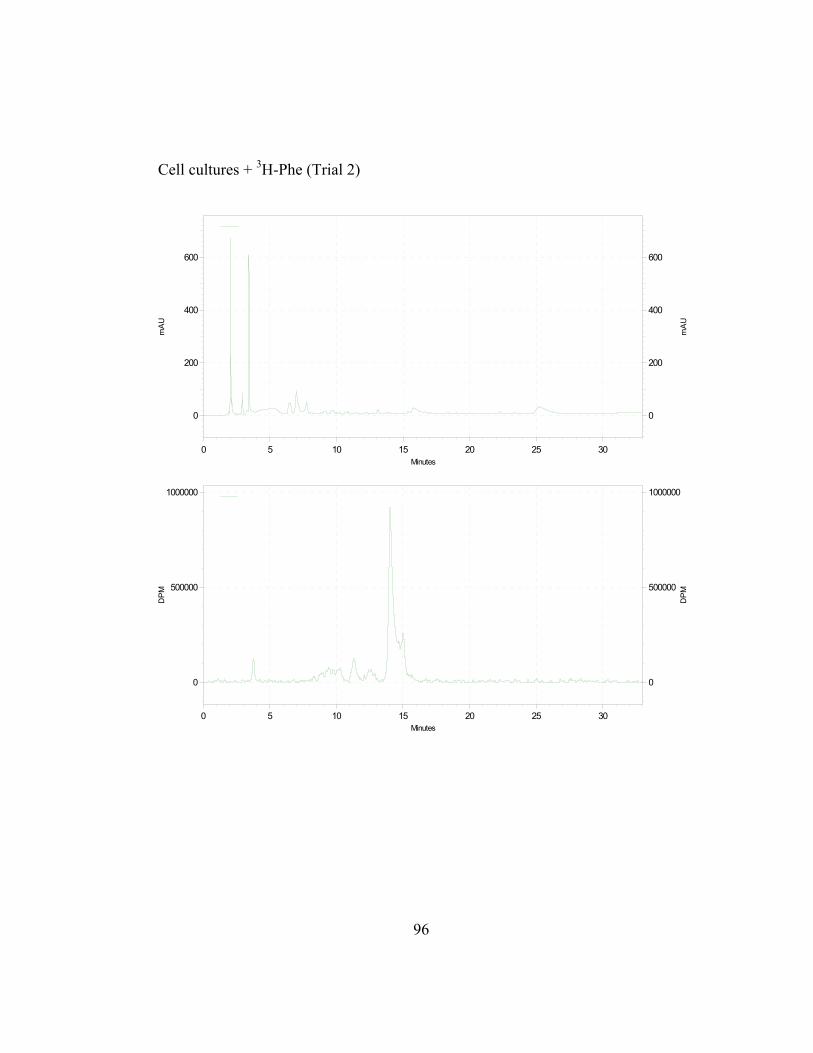
Cell cultures + 3H-Phe (Trial 2)
Minutes0 5 10 15 20 25 30
mAU
0
200
400
600
mAU
0
200
400
600
4: 280 nm, 4 nm3/28/02 7:37:51 PM_C162_DPM_2Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
500000
1000000
DPM
0
500000
1000000171 A3/28/02 7:37:51 PM_C162_DPM_2Shirley171RadioIsoflavprofile_Bmethod.met
96
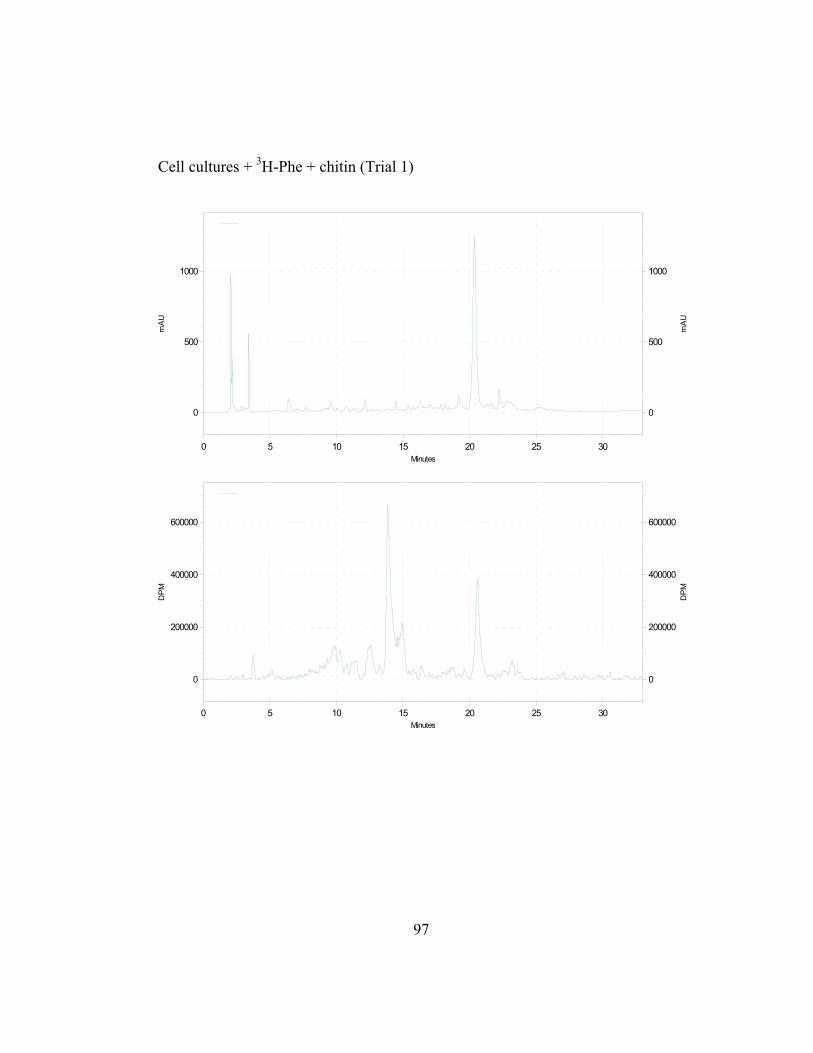
Cell cultures + 3H-Phe + chitin (Trial 1)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
mAU
0
500
1000
4: 280 nm, 4 nm3/28/02 8:18:18 PM_E161_DPM_3Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
200000
400000
600000
DPM
0
200000
400000
600000
171 A3/28/02 8:18:18 PM_E161_DPM_3Shirley171RadioIsoflavprofile_Bmethod.met
97
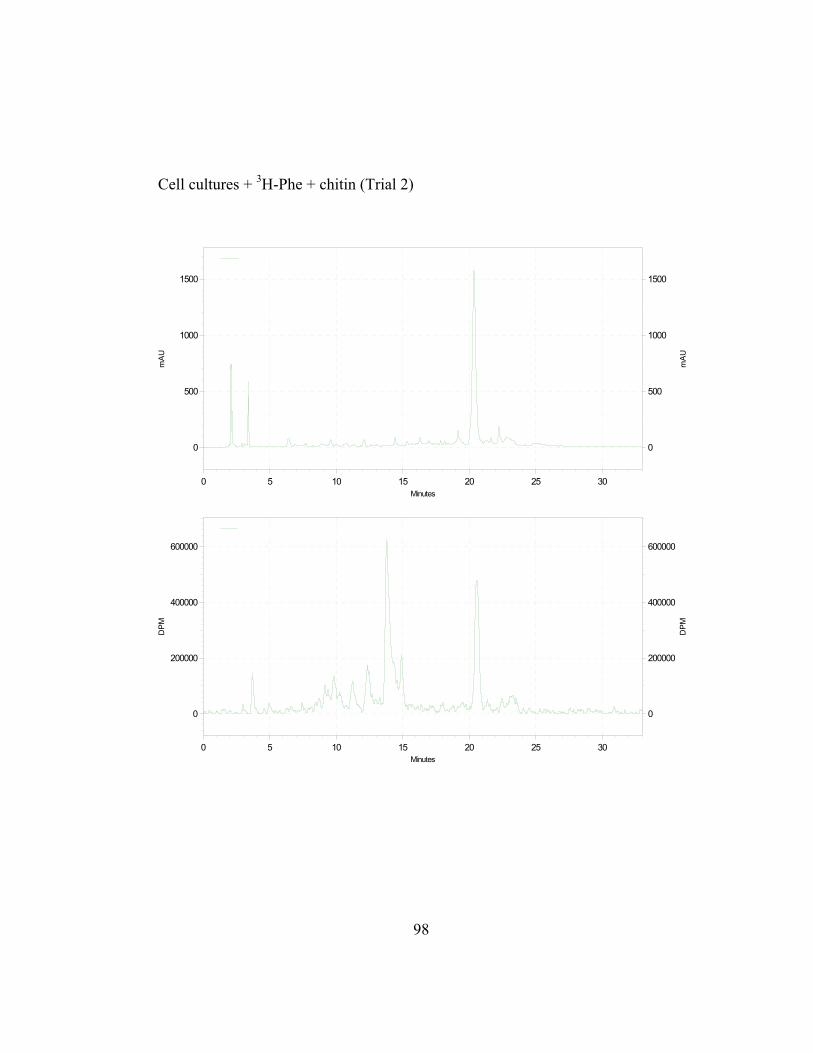
Cell cultures + 3H-Phe + chitin (Trial 2)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
1500
mAU
0
500
1000
1500
4: 280 nm, 4 nm3/28/02 8:58:41 PM_E162_DPM_4Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
200000
400000
600000
DPM
0
200000
400000
600000
171 A3/28/02 8:58:41 PM_E162_DPM_4Shirley171RadioIsoflavprofile_Bmethod.met
98
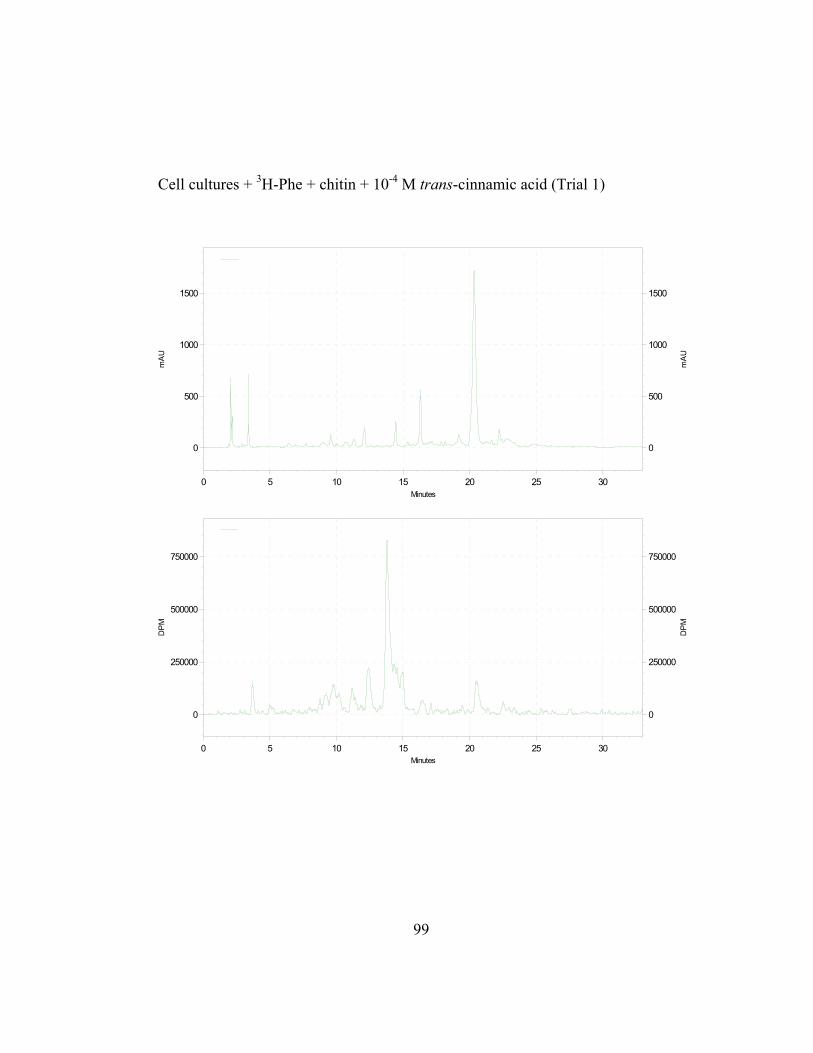
Cell cultures + 3H-Phe + chitin + 10-4 M trans-cinnamic acid (Trial 1)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
1500
mAU
0
500
1000
1500
4: 280 nm, 4 nm3/28/02 9:39:04 PM_E16AT1_DPM_5Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
250000
500000
750000
DPM
0
250000
500000
750000
171 A3/28/02 9:39:04 PM_E16AT1_DPM_5Shirley171RadioIsoflavprofile_Bmethod.met
99
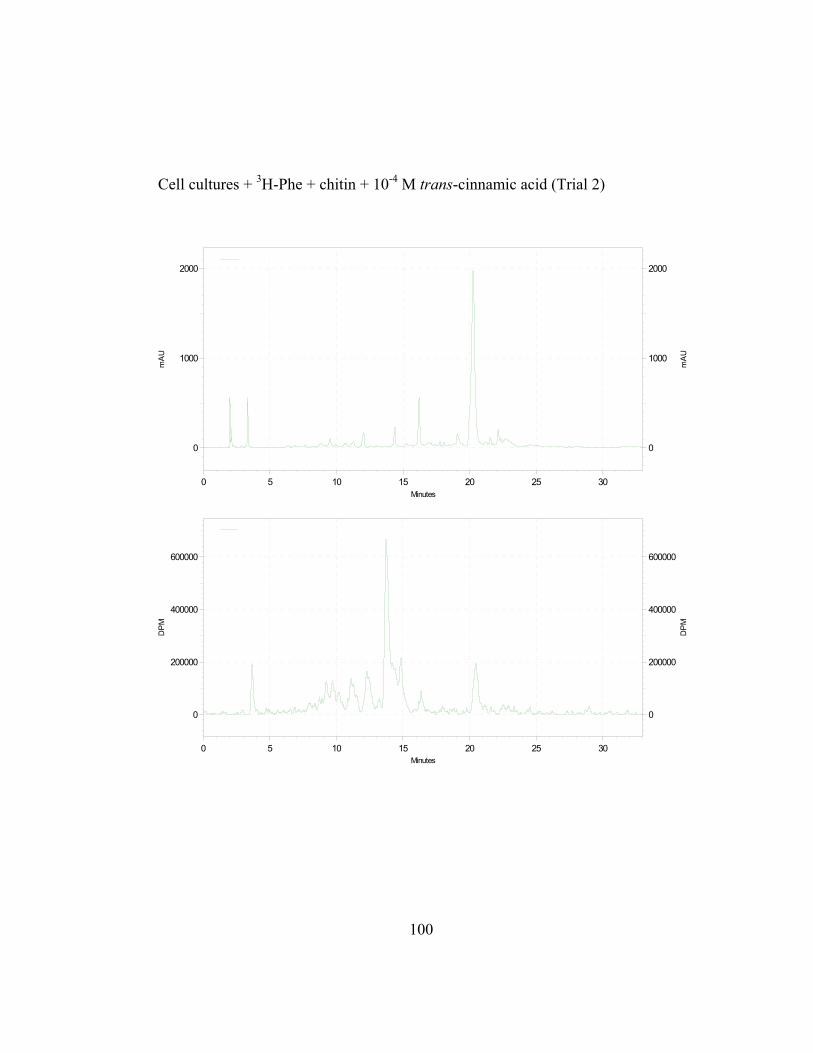
Cell cultures + 3H-Phe + chitin + 10-4 M trans-cinnamic acid (Trial 2)
Minutes0 5 10 15 20 25 30
mAU
0
1000
2000
mAU
0
1000
20004: 280 nm, 4 nm3/28/02 10:19:31 PM_E16AT2_DPM_6Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
200000
400000
600000
DPM
0
200000
400000
600000
171 A3/28/02 10:19:31 PM_E16AT2_DPM_6Shirley171RadioIsoflavprofile_Bmethod.met
100
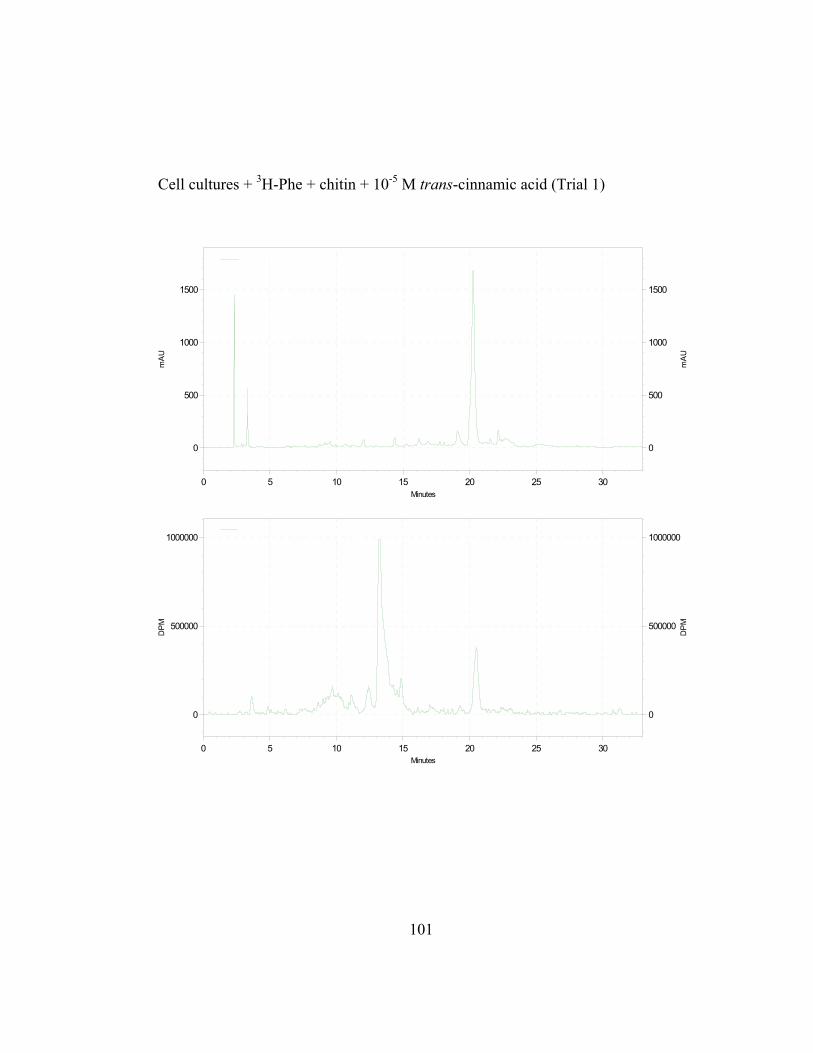
Cell cultures + 3H-Phe + chitin + 10-5 M trans-cinnamic acid (Trial 1)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
1500
mAU
0
500
1000
1500
4: 280 nm, 4 nm3/28/02 10:59:58 PM_E16DT1_DPM_7Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
500000
1000000
DPM
0
500000
1000000171 A3/28/02 10:59:58 PM_E16DT1_DPM_7Shirley171RadioIsoflavprofile_Bmethod.met
101
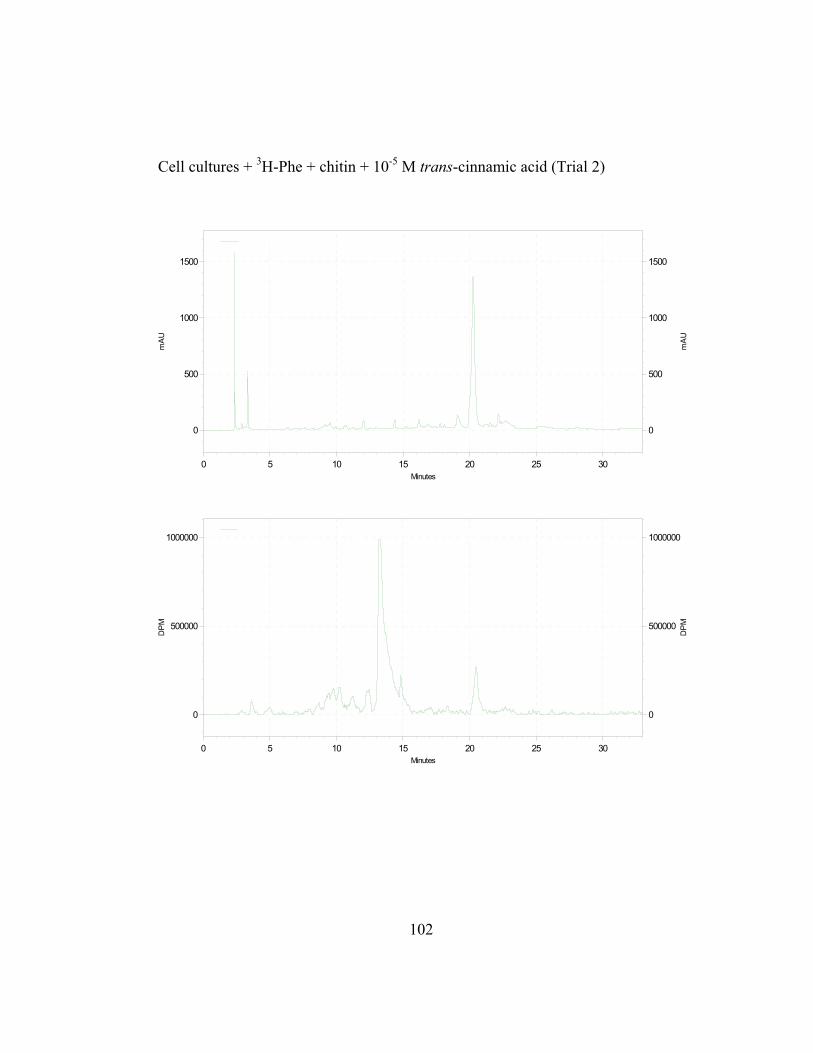
Cell cultures + 3H-Phe + chitin + 10-5 M trans-cinnamic acid (Trial 2)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
1500
mAU
0
500
1000
1500
4: 280 nm, 4 nm3/28/02 11:40:19 PM_E16DT2_DPM_8Shirley171RadioIsoflavprofile_Bmethod.met
Minutes0 5 10 15 20 25 30
DPM
0
500000
1000000
DPM
0
500000
1000000171 A3/28/02 11:40:19 PM_E16DT2_DPM_8Shirley171RadioIsoflavprofile_Bmethod.met
102
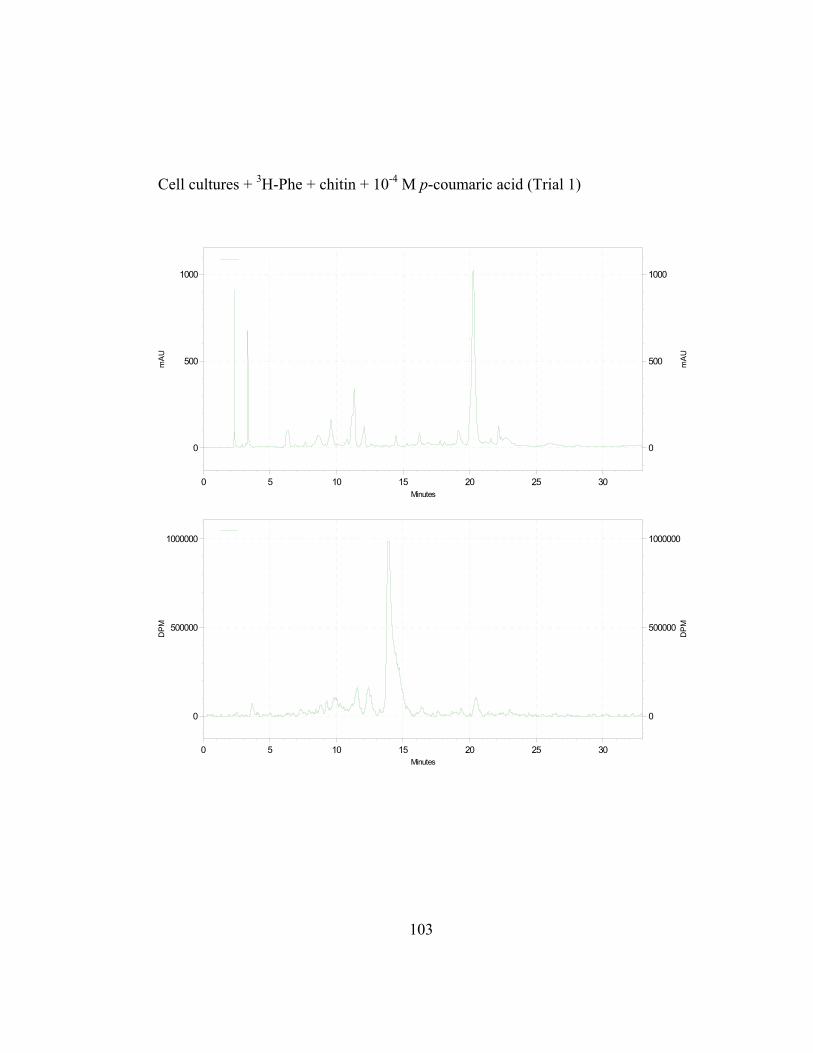
Cell cultures + 3H-Phe + chitin + 10-4 M p-coumaric acid (Trial 1)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
mAU
0
500
1000
4: 280 nm, 4 nm3/29/02 12:35:48 AM_E16AP1_1Shirley171RadioIsoflavprofile.met
Minutes0 5 10 15 20 25 30
DPM
0
500000
1000000
DPM
0
500000
1000000171 A3/29/02 12:35:48 AM_E16AP1_1Shirley171RadioIsoflavprofile.met
103
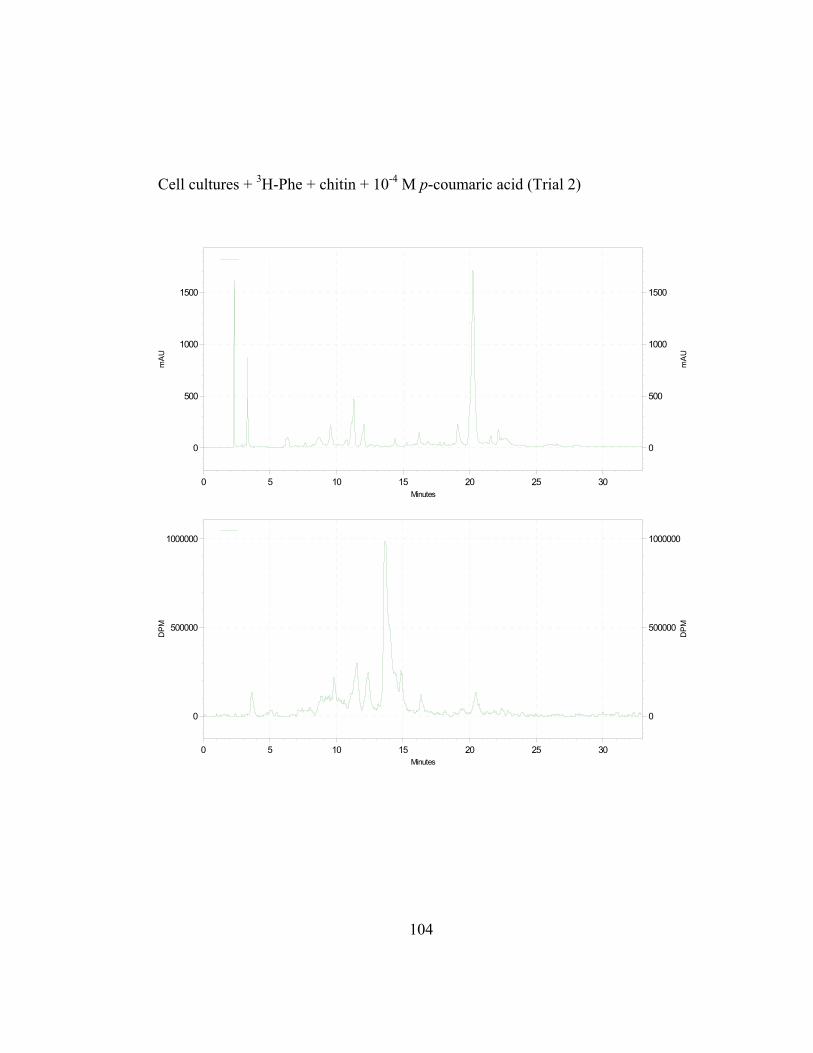
Cell cultures + 3H-Phe + chitin + 10-4 M p-coumaric acid (Trial 2)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
1500
mAU
0
500
1000
1500
4: 280 nm, 4 nm3/29/02 1:16:12 AM_E16AP2_2Shirley171RadioIsoflavprofile.met
Minutes0 5 10 15 20 25 30
DPM
0
500000
1000000
DPM
0
500000
1000000171 A3/29/02 1:16:12 AM_E16AP2_2Shirley171RadioIsoflavprofile.met
104

Cell cultures + 3H-Phe + chitin + 10-5 M p-coumaric acid (Trial 1)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
1500
mAU
0
500
1000
1500
4: 280 nm, 4 nm3/29/02 1:56:40 AM_E16DP1_3Shirley171RadioIsoflavprofile.met
Minutes0 5 10 15 20 25 30
DPM
0
500000
1000000
DPM
0
500000
1000000171 A3/29/02 1:56:40 AM_E16DP1_3Shirley171RadioIsoflavprofile.met
105
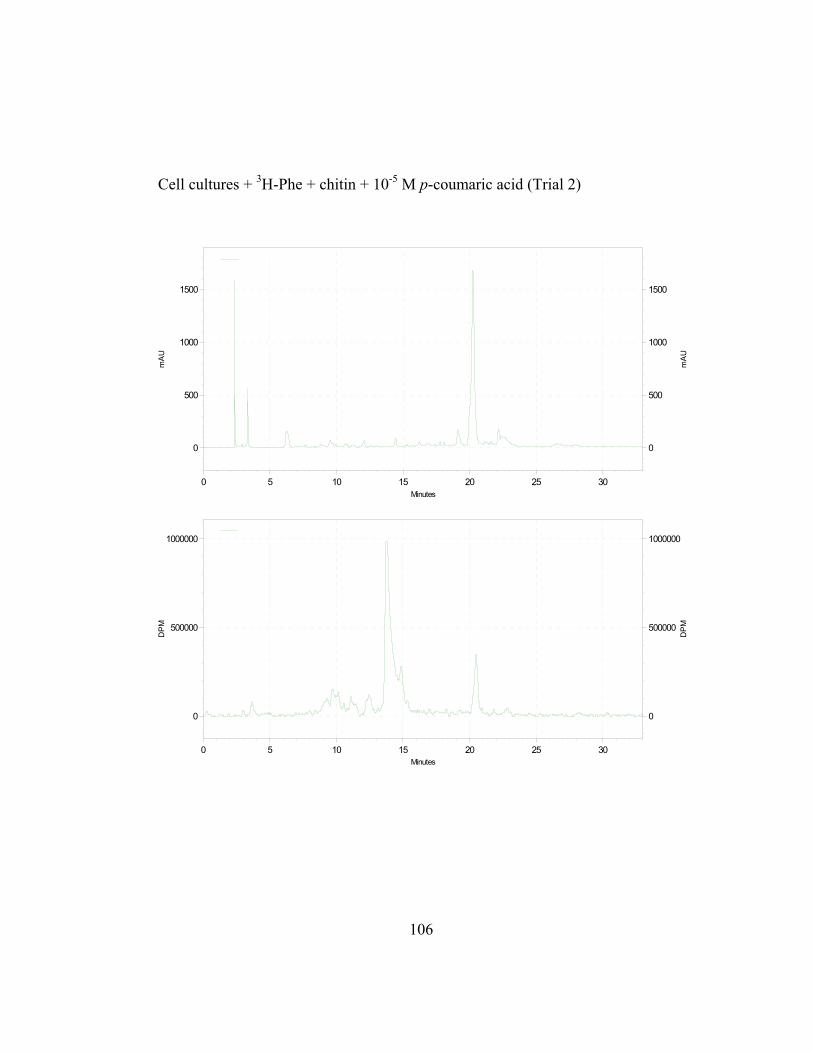
Cell cultures + 3H-Phe + chitin + 10-5 M p-coumaric acid (Trial 2)
Minutes0 5 10 15 20 25 30
mAU
0
500
1000
1500
mAU
0
500
1000
1500
4: 280 nm, 4 nm3/29/02 2:37:05 AM_E16DP2_4Shirley171RadioIsoflavprofile.met
Minutes0 5 10 15 20 25 30
DPM
0
500000
1000000
DPM
0
500000
1000000171 A3/29/02 2:37:05 AM_E16DP2_4Shirley171RadioIsoflavprofile.met
106
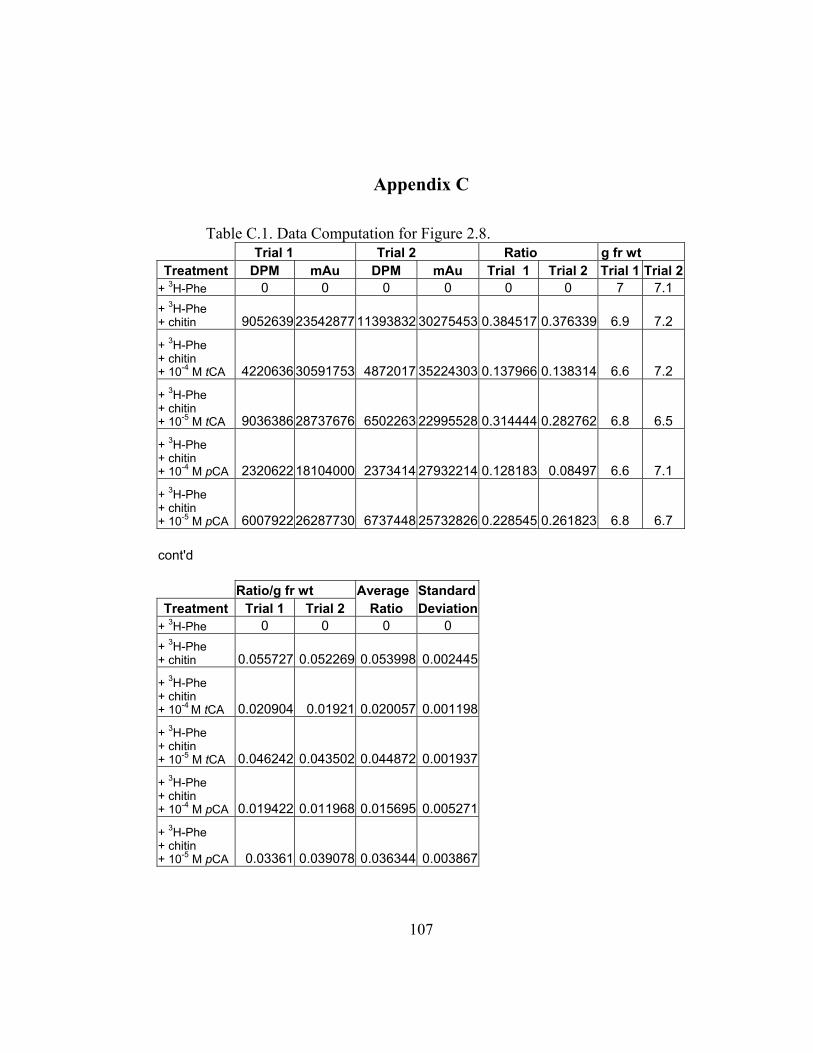
Appendix C
Table C.1. Data Computation for Figure 2.8. Trial 1 Trial 2 Ratio g fr wt Treatment DPM mAu DPM mAu Trial 1 Trial 2 Trial 1 Trial 2
+ 3H-Phe 0 0 0 0 0 0 7 7.1 + 3H-Phe + chitin 9052639 23542877 11393832 30275453 0.384517 0.376339 6.9 7.2
+ 3H-Phe + chitin + 10-4 M tCA 4220636 30591753 4872017 35224303 0.137966 0.138314 6.6 7.2
+ 3H-Phe + chitin + 10-5 M tCA 9036386 28737676 6502263 22995528 0.314444 0.282762 6.8 6.5
+ 3H-Phe + chitin + 10-4 M pCA 2320622 18104000 2373414 27932214 0.128183 0.08497 6.6 7.1
+ 3H-Phe + chitin + 10-5 M pCA 6007922 26287730 6737448 25732826 0.228545 0.261823 6.8 6.7 cont'd Ratio/g fr wt Average Standard Treatment Trial 1 Trial 2 Ratio Deviation
+ 3H-Phe 0 0 0 0 + 3H-Phe + chitin 0.055727 0.052269 0.053998 0.002445
+ 3H-Phe + chitin + 10-4 M tCA 0.020904 0.01921 0.020057 0.001198
+ 3H-Phe + chitin + 10-5 M tCA 0.046242 0.043502 0.044872 0.001937
+ 3H-Phe + chitin + 10-4 M pCA 0.019422 0.011968 0.015695 0.005271
+ 3H-Phe + chitin + 10-5 M pCA 0.03361 0.039078 0.036344 0.003867
107
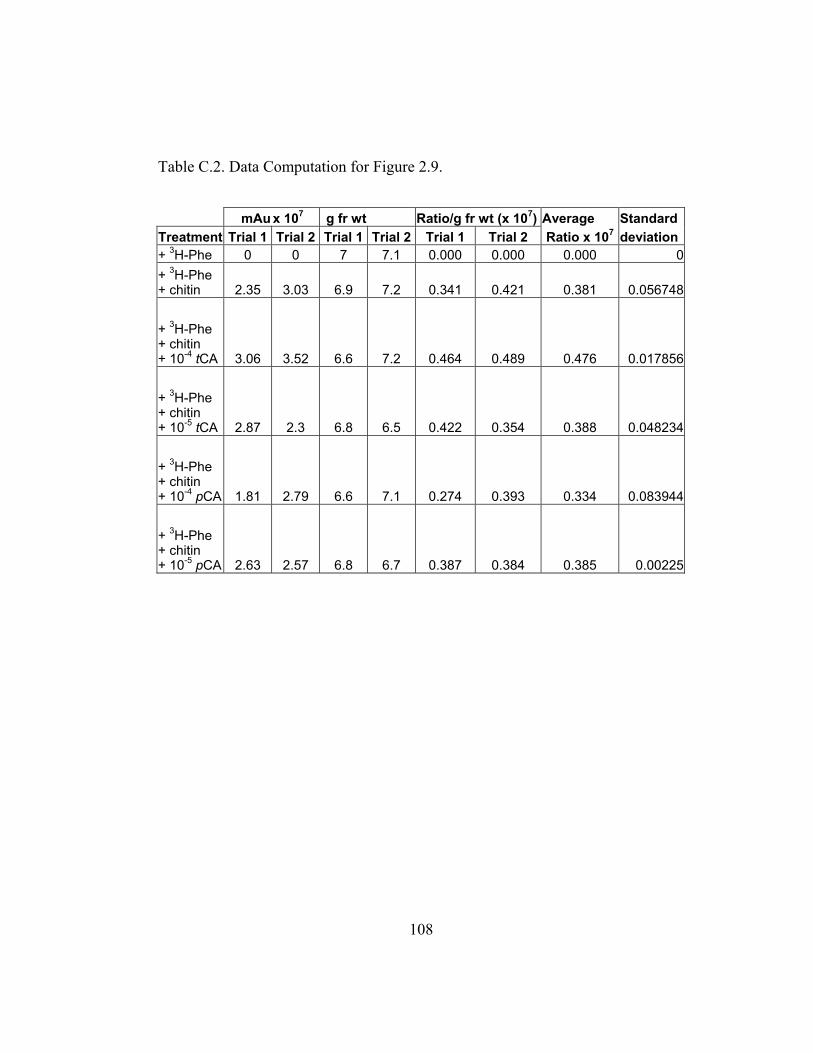
Table C.2. Data Computation for Figure 2.9.
mAu x 107 g fr wt Ratio/g fr wt (x 107) Average StandardTreatment Trial 1 Trial 2 Trial 1 Trial 2 Trial 1 Trial 2 Ratio x 107 deviation+ 3H-Phe 0 0 7 7.1 0.000 0.000 0.000 0+ 3H-Phe + chitin 2.35 3.03 6.9 7.2 0.341 0.421 0.381 0.056748
+ 3H-Phe + chitin + 10-4 tCA 3.06 3.52 6.6 7.2 0.464 0.489 0.476 0.017856
+ 3H-Phe + chitin + 10-5 tCA 2.87 2.3 6.8 6.5 0.422 0.354 0.388 0.048234
+ 3H-Phe + chitin + 10-4 pCA 1.81 2.79 6.6 7.1 0.274 0.393 0.334 0.083944
+ 3H-Phe + chitin + 10-5 pCA 2.63 2.57 6.8 6.7 0.387 0.384 0.385 0.00225
108

Bibliography
Allwood, E. G., Davies, D. R., Gerrish, C., Ellis, B. E. and Bolwell, G. P., 1999.
Phosphorylation of phenylalanine ammonia-lyase: evidence for a novel protein kinase and identification of the phosphorylated residue. Febs Letters 457, 47-52.
Ayers, A. R., Valent, B., Ebel, J. and Albersheim, P., 1976. Host- pathogen interactions. XI. Composition and structure of wall-released elicitor fractions. Plant Physiology 57, 766-774.
Bohm, B. A. and Chan, J., 1992. Flavonoids and affinities of Greyiaceae with a discussion of the occurrence of B-Ring deoxyflavonoids in dicotyledonous families. Systematic Botany 17, 272-281.
Bonness, M. S. and Mabry, T. J., 1992. Tissue-culture of Endod (Phytolacca dodecandra Lherit) - Growth and production of ribosome-inactivating proteins. Plant Cell Reports 11, 66-70.
Broekaert, W. F., Vanparijs, J., Allen, A. K. and Peumans, W. J., 1988. Comparison of some molecular, enzymatic and antifungal properties of chitinases from thorn-apple, tobacco and wheat. Physiological and Molecular Plant Pathology 33, 319-331.
Bucciarelli, B., Jung, H. G., Ostry, M. E., Anderson, N. A. and Vance, C. P., 1998. Wound response characteristics as related to phenylpropanoid enzyme activity and lignin deposition in resistant and susceptible Populus tremuloides inoculated with Entoleuca mammata (Hypoxylon mammatum). Canadian Journal of Botany-Revue Canadienne De Botanique 76, 1282-1289.
Burbulis, I. E., Iacobucci, M. and Shirley, B. W., 1996. A null mutation in the first enzyme of flavonoid biosynthesis does not affect male fertility in Arabidopsis. Plant Cell 8, 1013-1025.
Burbulis, I. E. and Winkel-Shirley, B., 1999. Interactions among enzymes of the Arabidopsis flavonoid biosynthetic pathway. Proceedings of the National Academy of Sciences of the United States of America 96, 12929-12934.
109

Chen, C. H., Huang, L. L. H., Huang, C. C., Lin, C. C., Lee, Y. and Lu, F. J., 2000. Baicalein, a novel apoptotic agent for hepatoma cell lines: A potential medicine for hepatoma. Nutrition and Cancer-an International Journal 38, 287-295.
Czichi, U. and Kindl, H., 1975. Formation of p-coumaric acid and o-coumaric acid from L- phenylalanine by microsomal membrane-fractions from potato - evidence of membrane-bound enzyme complexes. Planta 125, 115-125.
Czichi, U. and Kindl, H., 1975. Model of closely assembled consecutive enzymes on membranes - formation of hydroxycinnamic acids from L-phenylalanine on thylakoids of Dunaliella marina. Hoppe-Seylers Zeitschrift Fur Physiologische Chemie 356, 475-485.
Czichi, U. and Kindl, H., 1977. Phenylalanine ammonia-lyase and cinnamic acid hydroxylases as assembled consecutive enzymes on microsomal-membranes of cucumber cotyledons - cooperation and subcellular-distribution. Planta 134, 133-143.
Dalkin, K., Edwards, R., Edington, B. and Dixon, R. A., 1990. Stress responses in alfalfa (Medicago sativa L) .1. Induction of phenylpropanoid biosynthesis and hydrolytic enzymes in elicitor-treated cell-suspension cultures. Plant Physiology 92, 440-446.
Dietrich, A., Mayer, J. E. and Hahlbrock, K., 1990. Fungal elicitor triggers rapid, transient, and specific protein-phosphorylation in parsley cell-suspension cultures. Journal of Biological Chemistry 265, 6360-6368.
Dixon, R. A., 1997. Plant secondary metabolism: Control points and prospects for genetic manipulation. Abstracts of Papers of the American Chemical Society 213, 8-AGFD.
Dixon, R. A., 1999. Plant natural products: the molecular genetic basis of biosynthetic diversity. Current Opinion in Biotechnology 10, 192-197.
Dixon, R. A., 2001. Natural products and plant disease resistance. Nature 411, 843-847.
Dixon, R. A., Chen, F., Guo, D. J. and Parvathi, K., 2001. The biosynthesis of monolignols: a "metabolic grid", or independent pathways to guaiacyl and syringyl units? Phytochemistry 57, 1069-1084.
110

Dixon, R. A., Harrison, M. J. and Paiva, N. L., 1995. The isoflavonoid phytoalexin pathway - from enzymes to genes to transcription factors. Physiologia Plantarum 93, 385-392.
Dixon, R. A., Howles P.A., Lamb C., He X.Z., and Reddy, J.T., 1998. Prospects for the metabolic engineering of bioactive flavonoids and related phenylpropanoid compounds. Advances in Experimental Medicine and Biology 439, 55-66.
Ebel, J., Bhagwat, A. A., Cosio, E. G., Feger, M., Kissel, U., Mithofer, A. and Waldmuller, T., 1995. Elicitor binding-proteins and signal-transduction in the activation of a phytoalexin defense response. Canadian Journal of Botany-Revue Canadienne De Botanique 73, S506-S510.
Ebel, J. and Mithofer, A., 1998. Early events in the elicitation of plant defence. Planta 206, 335-348.
Essenberg, M., 2001. Prospects for strengthening plant defenses through phytoalexin engineering. Physiological and Molecular Plant Pathology 59, 71-81.
Felix, G., Grosskopf, D. G., Regenass, M. and Boller, T., 1991. Rapid changes of protein-phosphorylation are involved in transduction of the elicitor signal in plant-cells. Proceedings of the National Academy of Sciences of the United States of America 88, 8831-8834.
Fliegmann, J., Schroder, G., Schanz, S., Britsch, L. and Schroder, J., 1992. Molecular analysis of chalcone and dihydropinosylvin synthase from scots pine (Pinus sylvestris), and differential regulation of these and related enzyme-activities in stressed plants. Plant Molecular Biology 18, 489-503.
Freudenberg, K. and Neish, A.C., 1968. Constitution and Biosynthesis of Lignin. Springer-Verlag, New York.
Gao, Z. H., Huang, K. X. and Xu, H. B., 2001. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in HS-SY5Y cells. Pharmacological Research 43, 173-178.
Goodwin, T. W. and Mercer, E. I., 1983. Introduction to Plant Biochemistry. Pergamon Press, New York.
111

Habereder, H., Schroder, G. and Ebel, J., 1989. Rapid induction of phenylalanine ammonia-lyase and chalcone synthase messenger-Rnas during fungus infection of soybean (Glycine max L) roots or elicitor treatment of soybean cell- cultures at the onset of phytoalexin synthesis. Planta 177, 58-65.
Hahlbrock, K. and Scheel, D., 1989. Physiology and molecular-biology of phenylpropanoid metabolism. Annual Review of Plant Physiology and Plant Molecular Biology 40, 347-369.
Hamdam, M. A. M. S. and Dixon, R. A., 1986. Fractionation and properties of elicitors of the phenylpropanoid pathway from Colletotrichum lindemuthianum. Physiological and Molecular Plant Pathology 31, 105-121.
Hammerschmidt, R., 1999. Induced disease resistance: how do induced plants stop pathogens? Physiological and Molecular Plant Pathology 55, 77-84.
Hammerschmidt, R., 1999. Phytoalexins: What have we learned after 60 years? Annual Review of Phytopathology 37, 285-306.
Hartmann, T., 1996. Diversity and variability of plant secondary metabolism: a mechanistic view. Entomologia Experimentalis et Applicata 80, 177-188.
Hartmann, T., 1999. Chemical ecology of pyrrolizidine alkaloids. Planta 207, 483-495.
Hrazdina, G. and Jensen, R. A., 1992. Compartmentation in aromatic metabolism.In: H. Stanford and R. K. Ibrahim (Eds), Phenolic Metabolism in Plants. Plenum, New York, pp. 1-23.
Hrazdina, G. and Wagner, G. J., 1985. Metabolic pathways as enzyme complexes: evidence for the synthesis of phenylpropanoids and flavonoids on membrane-associated enzyme complexes. Archives of Biochemistry and Biophysics 237, 88-100.
Hsu, S. L., Hsieh, Y. C., Hsieh, W. C. and Chou, C. J., 2001. Baicalein induces a dual growth arrest by modulating multiple cell cycle regulatory molecules. European Journal of Pharmacology 425, 165-171.
Kimura, Y., Aoki, T. and Ayabe, S., 2001. Chalcone isomerase isozymes with different substrate specificities towards 6 '-hydroxy- and 6 '-deoxychalcones in cultured cells of Glycyrrhiza echinata, a leguminous
112

plant producing 5-deoxyflavonoids. Plant and Cell Physiology 42, 1169-1173.
Kuzovkina, I. N., Guseva, A. V., Alterman, I. E. and Karnachuk, R. A., 2001. Flavonoid production in transformed Scutellaria baicalensis roots and ways of its regulation. Russian Journal of Plant Physiology 48, 448-452.
Lazinsky, A., Lu, Y., Qiao, W. and Wei, H., 2001. Anti-photocarcinogenic effects of baicalein. Journal of Investigative Dermatology 117, 591.
Leon, J., Yalpani, N., Raskin, I. and Lawton, M. A., 1993. Induction of benzoic-acid 2-hydroxylase in virus-inoculated tobacco. Plant Physiology 103, 323-328.
Liu, Q. (1994). Chemical and enzymological investigations of the phenylpropanoid pathway in elicited cultures of Cephalocereus senilis ("Old man" cactus). Botany. Austin, University of Texas at Austin: 1-15.
Liu, Q., Bonness, M. S., Liu, M., Seradge, E., Dixon, R. A. and Mabry, T. J., 1995. Enzymes of B-ring-deoxy flavonoid biosynthesis in elicited cell-cultures of old man cactus (Cephalocereus senilis). Archives of Biochemistry and Biophysics 321, 397-404.
Liu, Q., Dixon, R. A. and Marby, T. J., 1993. Additional flavonoids from elicitor-treated cell-cultures of Cephalocereus senilis. Phytochemistry 34, 167-170.
Liu, Q., Liu, M., Mabry, T. J. and Dixon, R. A., 1994. Flavonol glycosides from Cephalocereus Senilis. Phytochemistry 31, 229-331.
Liu, Q., Markham, K. R., Paré, P. W., Dixon, R. A. and Mabry, T. J., 1993. Flavonoids from elicitor-treated cell-suspension cultures of Cephalocereus senilis. Phytochemistry 32, 925-928.
Mann, J., 1987. Secondary metabolism. Clarendon Press, Oxford.
Markham, K. R. and Mabry, T. J., 1968. The identification of twenty-three 5-deoxy- and ten 5-hydroxy-flavonoids from Baptisia lecontei (Leguminosae). Phytochemistry 7, 791-801.
Markham, K. R., Mabry, T. J. and Swift III, W. T., 1968. New isoflavones from the genus Baptisia (Leguminosae). Phytochemistry 7, 803-808.
113

Markham, K. R., Mabry, T. J. and Swift III, W. T., 1970. Distribution of flavonoids in the genus Baptisia (Leguminosae). Phytochemistry 9, 2359-2364.
Markham, K. R., Swift III, W. T. and Mabry, T. J., 1968. A new isoflavone glycoside from Baptisia australis. Journal of Organic Chemistry 33, 462-464.
Mehdy, M. C., 1994. Active Oxygen Species in Plant Defense against Pathogens. Plant Physiology 105, 467-472.
Mithofer, A., Ebel, J., Bhagwat, A. A., Boller, T. and Neuhaus-Url, G., 1999. Transgenic aequorin monitors cytosolic calcium transients in soybean cells challenged with beta-glucan or chitin elicitors. Planta 207, 566-574.
Nes, W. R. and McKean, M. L., 1977. Biochemistry of Steroids and Other Isopentenoids. University Park Press, Baltimore.
Ni, W. T., Fahrendorf, T., Ballance, G. M., Lamb, C. J. and Dixon, R. A., 1996. Stress responses in alfalfa (Medicago sativa L) .20. Transcriptional activation of phenylpropanoid pathway genes in elicitor-induced cell suspension cultures. Plant Molecular Biology 30, 427-438.
Noe, W., Langebartels, C. and Seitz, H. U., 1980. Anthocyanin accumulation and Pal activity in a suspension- culture of Daucus carota L - Inhibition by L-alpha-aminooxy- beta-phenylpropionic acid and t-cinnamic acid. Planta 149, 283-287.
Noe, W. and Seitz, H. U., 1983. Studies on the regulatory role of trans-cinnamic acid on the activity of the phenylalanine ammonia-lyase (Pal) in suspension-cultures of Daucus carota L. Zeitschrift Fur Naturforschung C-a Journal of Biosciences 38, 408-412.
O'Neal, D. and Keller, C. J., 1970. Partial purification and some properties of phenylalanine ammonia-lyase of tobacco (Nicotiana tabacum). Phytochemistry 9, 1373-1383.
Ovadi, J. and Srere, P. A., 1996. Metabolic consequences of enzyme interactions. Cell Biochemistry and Function 14, 249-258.
Ovadi, J. and Srere, P. A., 2000. Macromolecular compartmentation and channeling. International Review of Cytology - a Survey of Cell Biology, 192, 255-280.
114

Ozeki, Y., Ito, Y., Sasaki, N., Oyanagi, M., Akimoto, H., Chikagawa, Y. and Takeda, J., 2000. Phenylalanine ammonia-lyase genes involved in anthocyanin synthesis and the regulation of its expression in suspension cultured carrot cells. Journal of Plant Research 113, 319-326.
Paiva, N. L., Edwards, R., Sun, Y. J., Hrazdina, G. and Dixon, R. A., 1991. Stress responses in alfalfa (Medicago sativa L).11. Molecular- cloning and expression of alfalfa isoflavone reductase, a key enzyme of isoflavonoid phytoalexin biosynthesis. Plant Molecular Biology 17, 653-667.
Paré, P. W., Dmitrieva, N. and Mabry, T. J., 1991. Phytoalexin aurone induced in Cephalocereus senilis liquid suspension-culture. Phytochemistry 30, 1133-1135.
Paré, P. W., Mischke, C. F., Edwards, R., Dixon, R. A., Norman, H. A. and Mabry, T. J., 1992. Induction of phenylpropanoid pathway enzymes in elicitor- treated cultures of Cephalocereus senilis. Phytochemistry 31, 149-153.
Pillai, B. V. and Swarup, S., 2002. Elucidation of the flavonoid catabolism pathway in Pseudomonas putida PML2 by comparative metabolic profiling. Applied and environmental microbiology 68, 143-151.
Raskin, I., 1992. Salicylate, a new plant hormone. Plant Physiology 99, 799-803.
Rasmussen, S. and Dixon, R. A., 1997. Evidence for a microsomal enzyme complex of PAL and CA4H in tobacco plants. Plant Physiology 114, 1568-1568.
Rasmussen, S. and Dixon, R. A., 1999. Transgene-mediated and elicitor-induced perturbation of metabolic channeling at the entry point into the phenylpropanoid pathway. Plant Cell 11, 1537-1551.
Ren, Y. Y. and West, C. A., 1992. Elicitation of Diterpene Biosynthesis in Rice (Oryza sativa L) by Chitin. Plant Physiology 99, 1169-1178.
Rommelt, S., Treutter, D., Speakman, J. B. and W., R., 1999. Effects of prohexadione-Ca on the flavonoid metabolism of apple with respect to plant resistance against fireblight. Acta Horticulturae 489, 359-363.
Rossi, M., Meyer, R., Constantinou, P., Caruso, F., Castelbuono, D., O'Brien, M. and Narasimhan, V., 2001. Molecular structure and activity toward DNA
115

of baicalein, a flavone constituent of the Asian herbal medicine "Sho-saiko-to". Journal of Natural Products 64, 26-31.
Russell, C. E. and Felker, P., 1987. The prickly-pears (Opuntia Spp Cactaceae) - a source of human and animal food in semi-arid regions. Economic Botany 41, 433-445.
Sano, H. and Ohashi, Y., 1995. Involvement of small GTP-binding proteins in defense signal-transduction pathways of higher plants. Proceedings of the National Academy of Sciences of the United States of America 92, 4138-4144.
Shibuya, N., Kaku, H., Kuchitsu, K. and Maliarik, M. J., 1993. Identification of a novel high-affinity binding-site for N- acetylchitooligosaccharide elicitor in the membrane-fraction from suspension-cultured rice cells. Febs Letters 329, 75-78.
Shirley, B. W., 1996. Flavonoid biosynthesis: 'New' functions for an 'old' pathway. Trends in Plant Science 1, 377-382.
Shirley, B. W., 1999. Evidence for enzyme complexes in the phenylpropanoid and flavonoid pathways. Physiologia Plantarum 107, 142-149.
Smith, C. J., 1996. Accumulation of phytoalexins: Defence mechanism and stimulus response system. New Phytologist 132, 1-45.
Spivey, H. O. and Ovadi, J., 1999. Substrate channeling. Methods- a Companion to Methods in Enzymology 19, 306-321.
Stab, M. R. and Ebel, J., 1987. Effects of Ca2+ on phytoalexin induction by fungal elicitor in soybean cells. Archives of Biochemistry and Biophysics 257, 416-423.
Stafford, H. A., 1974. Possible multi-enzyme complexes regulating the formation of C6-C3 phenolic compounds and lignins in higher plants. Recent Advances in Phytochemistry 8, 53-79.
Stubbe, J., Ge, J. and Yee, C. S., 2001. The evolution of ribonucleotide reduction revisited. Trends in Biochemical Sciences 26, 93-99.
Svitil, A. L. and Kirchman, D. L., 1998. A chitin-binding domain in a marine bacterial chitinase and other microbial chitinases: implications for the
116

ecology and evolution of 1,4-beta-glycanases. Microbiology-Uk 144, 1299-1308.
Tenhaken, R., Levine, A., Brisson, L. F., Dixon, R. A. and Lamb, C., 1995. Function of the oxidative burst in hypersensitive disease resistance. Proceedings of the National Academy of Sciences of the United States of America 92, 4158-4163.
Treutter, D., 2001. Biosynthesis of phenolic compounds and its regulation in apple. Plant Growth Regulation 34, 71-89.
Walker-Simmons, M., Hadwiger, L. and Ryan, C. A., 1983. Chitosans and pectic polysaccharides both induce the accumulation of the antifungal phytoalexin pisatin in pea pods and antinutrient proteinase inhibitors in tomato leaves. Biochemical & Biophysical Research Communications 110, 194-9.
Whetten, R. and Sederoff, R., 1995. Lignin biosynthesis. Plant Cell 7, 1001-1013.
Wong, E., 1966. Occurence and biosynthesis of 4',6'-dihydroxyaurone in soybean. Phytochemistry 5, 463-467.
Yamaguchi, T., Yamada, A., Hong, N., Ogawa, T., Ishii, T. and Shibuya, N., 2000. Differences in the recognition of glucan elicitor signals between rice and soybean: beta-glucan fragments from the rice blast disease fungus Pyricularia oryzae that elicit phytoalexin biosynthesis in suspension-cultured rice cells. Plant Cell 12, 817-826.
Yamamoto, H., Koga, T., Hayakawa, S., Ohgata, Y., Kurasaki, T. and Tohyama, K., 1998. Effect of chitin application on growth and yield of rice. Japanese Journal of Crop Science 67, 452-456.
Yoshikawa, M., Takeuchi, Y. and Horino, O., 1990. A mechanism for ethylene-induced disease resistance in soybean - enhanced synthesis of an elicitor-releasing factor, Beta-1,3- endoglucanase. Physiological and Molecular Plant Pathology 37, 367-376.
117

Vita
The eldest of three children of William G. Padolina and Maria Cristina D.
Padolina, Isagani Damasco Padolina was born on January 22, 1974 in Quezon
City, Republic of the Philippines. He finished his elementary schooling at
Maquiling School Inc. and obtained his high school diploma from the University
of the Philippines Rural High School. In June 1990, he initiated his studies
towards a Bachelor of Science degree in Chemistry at the University of the
Philippines Los Baños and graduated in May 1994, completing his undergraduate
thesis under Prof. Juanita C. Mamaril. He was then employed by the International
Rice Research Institute (IRRI), Laguna, Philippines as a Research Assistant for
one year until he joined the San Miguel Corporation as a quality assurance
chemist in its Brewing Group. In August 1996, he attended the Graduate
Chemistry Program of the University of Texas at Dallas. He transferred to the
Plant Biology Graduate Program at the University of Texas at Austin in August
1997, and he was the recipient of the H.C. Bold Award for Teaching Excellence
for the academic year 2001-2002.
Permanent address: 2065 Sanggumay St., College, Laguna, Philippines 4031
This dissertation was typed by the author.


















