
Contribution of c-erbB-2and Topoisomerase IIato...
10
[CANCER RESEARCH 59, 3206 –3214, July 1, 1999] Contribution of c-erbB-2 and Topoisomerase IIa to Chemoresistance in Ovarian Cancer 1 Jan G. Hengstler, 2 Jost Lange, Alexandra Kett, Nadja Dornho ¨fer, Rolf Meinert, Michael Arand, Paul G. Knapstein, Roger Becker, Franz Oesch, and Berno Tanner Institute of Toxicology [J. G. H., J. L., N. D., M. A., R. B., F. O.], Department of Gynecology [A. K., P. G. K., B. T.], and Institute of Statistics and Documentation [R. M.], University of Mainz, 55131 Mainz, Germany ABSTRACT Overexpression of the c-erbB-2 (HER-2/neu) oncogene, which encodes a transmembrane receptor tyrosine kinase, has been shown to be associated with poor prognosis in ovarian and breast cancer. Recent studies indicate that c-erbB-2 may also be involved in determining the chemosensitivity of human cancers. In the present study, we examined the role of c-erbB-2 for chemoresistance in ovarian cancer. Overexpression of c-erbB-2 mRNA in tumor tissue was associated with a shorter survival of patients with primary ovarian cancer (P 5 0.0001; n 5 77) and was an independent prognostic factor in the proportional-hazard model adjusted for Interna- tional Federation of Gynecologists and Obstetricians stage, residual dis- ease, chemotherapy, and age (P 5 0.035). A significant association be- tween expression of c-erbB-2 mRNA and survival was obtained for the subgroup of patients who received a standard chemotherapy with carbo- platin or cisplatin and cyclophosphamide (P 5 0.0003), whereas only a nonsignificant trend was observed for patients who did not receive a standard chemotherapy (P 5 0.124). In addition, the application of a standard chemotherapy improved the survival of patients with relatively low c-erbB-2 expression (P 5 0.013) but not of patients with overexpres- sion of c-erbB-2 (P 5 0.359). Expression of c-erbB-2 mRNA correlated with expression of topoisomerase IIa mRNA determined by a reverse semiquantitative PCR technique (P 5 0.009), whereas expression of c- erbB-2 and topoisomerase IIb mRNA did not correlate (P 5 0.221). To examine the hypothesis that coamplified and/or coregulated topoisomer- ase IIa contributes to the resistance of c-erbB-2-overexpressing carcino- mas, we established a chemosensitivity assay using primary cells from an ovarian carcinoma that overexpressed both c-erbB-2 and topoisomerase IIa. The combination of carboplatin with nontoxic concentrations of the topoisomerase II inhibitors etoposide or novobiocin enhanced the toxicity of carboplatin. In contrast, the tyrosine kinase inhibitor emodin exhibited no chemosensitizing effect in cells of this individual carcinoma. In conclu- sion, overexpression of c-erbB-2 was associated with poor prognosis and poor response to chemotherapy. The data suggest that topoisomerase IIa, which correlates with c-erbB-2 expression, contributes to the resistance of c-erbB-2-overexpressing carcinomas. INTRODUCTION The c-erbB-2 (HER-2/neu) proto-oncogene encodes a M r 185,000 transmembrane receptor tyrosine kinase that is a member of the class I receptor tyrosine kinase family, which includes the epidermal growth factor, HER-2/neu (c-erbB-2), HER-3, and HER-4 (1). C- erbB-2 was originally identified as an oncogene in chemically induced rat neuroglioblastomas, in which a single point mutation in the trans- membrane domain of the molecule confers oncogenic activation (2, 3). In contrast, in humans, the c-erbB-2 proto-oncogene is not acti- vated by a point mutation but through amplification and overexpres- sion of the wild-type gene (4, 5). Amplification of the c-erbB-2 gene is observed in 20 –30% of many cancer types, and its overexpression is correlated with a poor prognosis for breast and ovarian cancer patients (6 –9). Consistent with these clinical observations, overex- pression of c-erbB-2 in human ovarian and breast cancer cell lines has been shown to promote cell growth and cause tumorigenicity in nude mice xenograft models (1, 10 –12). Thus, the poor prognosis of ovarian and breast cancer patients associated with c-erbB-2 overex- pression seemed to be sufficiently explained by a c-erbB-2-mediated high rate of tumor cell proliferation. However, recently evidence was presented suggesting that c-erbB-2 overexpression may also be associated with cytotoxic drug resistance because a dose-response effect with regard to survival was observed in patients with c-erbB-2-negative ovarian tumors but not in patients with c-erbB-2-positive tumors (7). Consistent with these observations, a monoclonal antibody specific to an extracellular epitope of c-erbB-2 (13) and the tyrosine kinase inhibitor emodin (14), which suppresses c-erbB-2 tyrosine kinase activity, both enhanced the cytotoxicity of cisplatin against c-erbB-2-overexpressing cell lines. However, the role of c-erbB-2 for chemoresistance still remains inconclusive because expression of a full-length c-erbB-2 cDNA into several human breast and ovarian cancer cell lines was not sufficient to induce resistance to cisplatin across all of the cell lines tested (15). Although the sensi- tivity to cisplatin decreased significantly in three cell lines transfected with c-erbB-2 cDNA (MCF7, BT-20, and 2008 cells), expression of the human c-erbB-2 cDNA did not significantly influence cisplatin sensitivity in the cell lines MDA-MB-435, MDA-MD-231, and Caov-3 (15). Thus, the influence of c-erbB-2 on chemosensitivity was cell-type-specific and not generic across the cell lines tested. In the present study, we examine the role of c-erbB-2 on the success of chemotherapy with carboplatin and cyclophosphamide for patients with primary epithelial ovarian cancer. We report that (a) chemother- apy significantly improves the survival time of patients with low c-erbB-2 expression but not of patients with overexpression of c- erbB-2;(b) mRNA expression of c-erbB-2 significantly correlates with mRNA expression of topoisomerase IIa; and (c) the inhibition of topoisomerase II but not the antagonization of c-erbB-2 function increased the carboplatin chemosensitivity of primary cells obtained from an ovarian tumor overexpressing c-erbB-2 and topoisomerase IIa. Thus, the consideration of topoisomerase IIa may help to under- stand the observed association between c-erbB-2 overexpression and chemoresistance. MATERIALS AND METHODS Patients, Tissue Specimens, and Pathological Data. Tumor tissue was collected from 77 patients who underwent surgery from 1986 to 1995 at the Department of Gynecology, University of Mainz, Mainz, Germany. Tissue specimens were taken intraoperatively, immediately frozen in liquid nitrogen, and stored at 280°C. From all of the tumor specimens, 20-mm-thick cryosec- tions were prepared using a cryostat (System Dittes Duspiva, Heidelberg, Germany). After 20 cryosections (20 mm thick), a 7-mm-thick cryosection was prepared that was reviewed by a pathologist. Only those tissue samples with Received 10/20/98; accepted 5/3/99. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 This study was supported by the Stiftung Rheinland Pfalz fu ¨r Innovation, the Deutsche Forschungsgemeinschaft (SFB 519, project no. B5) and the European Commu- nity. 2 To whom requests for reprints should be addressed, Institute of Toxicology, Obere Zahlbacher Str. 67, University of Mainz, 55131 Mainz, Germany. Phone: 49-6131-173318 or 173115; Fax: 49-6131-230506; E-mail: [email protected]. 3206 Research. on September 8, 2018. © 1999 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of Contribution of c-erbB-2and Topoisomerase IIato...

[CANCER RESEARCH 59, 3206–3214, July 1, 1999]
Contribution of c-erbB-2and Topoisomerase IIa to Chemoresistance inOvarian Cancer1
Jan G. Hengstler,2 Jost Lange, Alexandra Kett, Nadja Dornhofer, Rolf Meinert, Michael Arand, Paul G. Knapstein,Roger Becker, Franz Oesch, and Berno TannerInstitute of Toxicology [J. G. H., J. L., N. D., M. A., R. B., F. O.], Department of Gynecology [A. K., P. G. K., B. T.], and Institute of Statistics and Documentation [R. M.],University of Mainz, 55131 Mainz, Germany
ABSTRACT
Overexpression of thec-erbB-2(HER-2/neu) oncogene, which encodes atransmembrane receptor tyrosine kinase, has been shown to be associatedwith poor prognosis in ovarian and breast cancer. Recent studies indicatethat c-erbB-2may also be involved in determining the chemosensitivity ofhuman cancers. In the present study, we examined the role ofc-erbB-2forchemoresistance in ovarian cancer. Overexpression ofc-erbB-2mRNA intumor tissue was associated with a shorter survival of patients withprimary ovarian cancer (P 5 0.0001;n 5 77) and was an independentprognostic factor in the proportional-hazard model adjusted for Interna-tional Federation of Gynecologists and Obstetricians stage, residual dis-ease, chemotherapy, and age (P 5 0.035). A significant association be-tween expression ofc-erbB-2 mRNA and survival was obtained for thesubgroup of patients who received a standard chemotherapy with carbo-platin or cisplatin and cyclophosphamide (P 5 0.0003), whereas only anonsignificant trend was observed for patients who did not receive astandard chemotherapy (P 5 0.124). In addition, the application of astandard chemotherapy improved the survival of patients with relativelylow c-erbB-2expression (P5 0.013) but not of patients with overexpres-sion of c-erbB-2 (P 5 0.359). Expression ofc-erbB-2 mRNA correlatedwith expression of topoisomerase IIa mRNA determined by a reversesemiquantitative PCR technique (P 5 0.009), whereas expression ofc-erbB-2 and topoisomerase IIb mRNA did not correlate (P 5 0.221). Toexamine the hypothesis that coamplified and/or coregulated topoisomer-ase IIa contributes to the resistance ofc-erbB-2-overexpressing carcino-mas, we established a chemosensitivity assay using primary cells from anovarian carcinoma that overexpressed bothc-erbB-2 and topoisomeraseII a. The combination of carboplatin with nontoxic concentrations of thetopoisomerase II inhibitors etoposide or novobiocin enhanced the toxicityof carboplatin. In contrast, the tyrosine kinase inhibitor emodin exhibitedno chemosensitizing effect in cells of this individual carcinoma. In conclu-sion, overexpression ofc-erbB-2was associated with poor prognosis andpoor response to chemotherapy. The data suggest that topoisomerase IIa,which correlates with c-erbB-2expression, contributes to the resistance ofc-erbB-2-overexpressing carcinomas.
INTRODUCTION
The c-erbB-2(HER-2/neu) proto-oncogene encodes aMr 185,000transmembrane receptor tyrosine kinase that is a member of the classI receptor tyrosine kinase family, which includes the epidermalgrowth factor, HER-2/neu (c-erbB-2), HER-3, and HER-4 (1).C-erbB-2was originally identified as an oncogene in chemically inducedrat neuroglioblastomas, in which a single point mutation in the trans-membrane domain of the molecule confers oncogenic activation (2,3). In contrast, in humans, thec-erbB-2proto-oncogene is not acti-vated by a point mutation but through amplification and overexpres-
sion of the wild-type gene (4, 5). Amplification of thec-erbB-2geneis observed in 20–30% of many cancer types, and its overexpressionis correlated with a poor prognosis for breast and ovarian cancerpatients (6–9). Consistent with these clinical observations, overex-pression ofc-erbB-2in human ovarian and breast cancer cell lines hasbeen shown to promote cell growth and cause tumorigenicity in nudemice xenograft models (1, 10–12). Thus, the poor prognosis ofovarian and breast cancer patients associated withc-erbB-2overex-pression seemed to be sufficiently explained by ac-erbB-2-mediatedhigh rate of tumor cell proliferation.
However, recently evidence was presented suggesting thatc-erbB-2overexpression may also be associated with cytotoxic drug resistancebecause a dose-response effect with regard to survival was observedin patients withc-erbB-2-negative ovarian tumors but not in patientswith c-erbB-2-positive tumors (7). Consistent with these observations,a monoclonal antibody specific to an extracellular epitope ofc-erbB-2(13) and the tyrosine kinase inhibitor emodin (14), which suppressesc-erbB-2 tyrosine kinase activity, both enhanced the cytotoxicity ofcisplatin againstc-erbB-2-overexpressing cell lines. However, the roleof c-erbB-2 for chemoresistance still remains inconclusive becauseexpression of a full-lengthc-erbB-2cDNA into several human breastand ovarian cancer cell lines was not sufficient to induce resistance tocisplatin across all of the cell lines tested (15). Although the sensi-tivity to cisplatin decreased significantly in three cell lines transfectedwith c-erbB-2cDNA (MCF7, BT-20, and 2008 cells), expression ofthe humanc-erbB-2cDNA did not significantly influence cisplatinsensitivity in the cell lines MDA-MB-435, MDA-MD-231, andCaov-3 (15). Thus, the influence ofc-erbB-2on chemosensitivity wascell-type-specific and not generic across the cell lines tested.
In the present study, we examine the role ofc-erbB-2on the successof chemotherapy with carboplatin and cyclophosphamide for patientswith primary epithelial ovarian cancer. We report that (a) chemother-apy significantly improves the survival time of patients with lowc-erbB-2 expression but not of patients with overexpression ofc-erbB-2; (b) mRNA expression ofc-erbB-2 significantly correlateswith mRNA expression of topoisomerase IIa; and (c) the inhibition oftopoisomerase II but not the antagonization ofc-erbB-2 functionincreased the carboplatin chemosensitivity of primary cells obtainedfrom an ovarian tumor overexpressingc-erbB-2and topoisomeraseIIa. Thus, the consideration of topoisomerase IIa may help to under-stand the observed association betweenc-erbB-2overexpression andchemoresistance.
MATERIALS AND METHODS
Patients, Tissue Specimens, and Pathological Data.Tumor tissue wascollected from 77 patients who underwent surgery from 1986 to 1995 at theDepartment of Gynecology, University of Mainz, Mainz, Germany. Tissuespecimens were taken intraoperatively, immediately frozen in liquid nitrogen,and stored at280°C. From all of the tumor specimens, 20-mm-thick cryosec-tions were prepared using a cryostat (System Dittes Duspiva, Heidelberg,Germany). After 20 cryosections (20mm thick), a 7-mm-thick cryosection wasprepared that was reviewed by a pathologist. Only those tissue samples with
Received 10/20/98; accepted 5/3/99.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 This study was supported by the Stiftung Rheinland Pfalz fur Innovation, theDeutsche Forschungsgemeinschaft (SFB 519, project no. B5) and the European Commu-nity.
2 To whom requests for reprints should be addressed, Institute of Toxicology, ObereZahlbacher Str. 67, University of Mainz, 55131 Mainz, Germany. Phone: 49-6131-173318or 173115; Fax: 49-6131-230506; E-mail: [email protected].
3206
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

more than 80% tumor cells in all of the 7-mm-thick cryosections were used forfurther analysis.
All of the patients examined in this study underwent surgery because ofprimary epithelial ovarian cancer. Patients with benign and nonepithelialtumors and borderline tumors were excluded from the study. Treatment ofpatients with ovarian carcinoma was primarily by surgery. Patients weretreated by total abdominal hysterectomy, bilateral salpingoophorectomy, andomentectomy. In the case of FIGO3 stages III and IV, in which canceroustissue could not be completely removed, a debulking operation was performedto reduce the remaining tumor to a diameter not to exceed 2 cm. In this study,patients with FIGO stages Ib, Ic, II, III, and IV usually received a postoperativechemotherapy with carboplatin (350 mg/m2) or cisplatin (50 mg/m2) andcyclophosphamide (1000 mg/m2) in six courses. The first course of chemo-therapy usually was performed 3–4 weeks after surgery. The following treat-ment regimens were defined as standard chemotherapy (n 5 44): (a) sixcourses of 50 mg/m2 cisplatin i.v. and 1000 mg/m2 cyclophosphamide i.v.(n 5 26); (b) six courses of 350 mg/m2 carboplatin i.v. and 1000 mg/m2
cyclophosphamide i.v. (n 5 16); and (c) six courses of 350 mg/m2 carboplatini.v. and 600 mg/m2 cyclophosphamide i.v. (n5 2). Because of myelotoxicityor nephrotoxicity, not all of the patients received a complete standard chem-otherapy. In addition, some patients refused to receive a chemotherapy. Thus,several treatment regimens contained no doses or much lower doses of plati-num compounds and cyclophosphamide. They were classified as “no standardchemotherapy” (n5 33): (a) no cytotoxic chemotherapy at all (n 5 19); (b)four courses of 1000 mg/m2 cyclophosphamide i.v. and 150 mg/m2 etoposidei.v. (n 5 1); (c) three courses of 1000 mg/m2 cyclophosphamide i.v. (n5 2);(d) two courses of 350 mg/m2 carboplatin i.v. and 1000 mg/m2 cyclophosph-amide i.v. (n 5 1); (e) medroxyprogesterone acetate, 500 mg/day p.o. (long-term therapy;n 5 1); (f) one course of 35 mg/m2 epirubicin i.v. and threecourses of treosulfan, 63 1500 mg/day p.o. (n 5 1); (g) tamoxifen, 30 mg/dayp.o. (long-term therapy;n 5 1); (h) cyclophosphamide, 23 50 mg/day p.o.(long-term therapy;n 5 4); (i) cyclophosphamide, 33 6 mg/day p.o. (long-term therapy;n 5 1); and (j) cyclophosphamide, 100 mg/day p.o., and5-fluorouracil, 750 mg/week i.v., for 12 weeks (n 5 2).
Histological typing was performed according to the WHO criteria. Theepithelial tumors were subdivided into serous and nonserous carcinomas. Thehistological grade of malignancy ranged from GI (differentiated) to GIII(undifferentiated). Grading was performed by the following criteria: tumorarchitecture, amount of solid tumor, nuclear pleomorphism, nucleus cyto-plasma ratio, number of nucleoli, and mitoses (16). Tumor staging was doneaccording to the guidelines of FIGO. Because residual disease is a strongprognostic factor, we differentiated between patients with macroscopicallycomplete removal of all of the tumor tissue and patients with# 2 cm3 and.2cm3 of tumor tissue left after surgery.
Reverse Semiquantitaive PCR.Total RNA from frozen tissue specimensof about 10 mm3 were isolated with a commercially available kit according tothe supplier’s instructions (Rneasy Mini Kit 59, Qiagen, Hilden, Germany).The RNA concentration was determined spectrophotometrically at 260 nm.The quality of the isolated RNA was checked by agarose gel electrophoresis bymeans of the presence of 28S, and 18S rRNA. Reverse transcription of mRNAcDNA was performed using a kit (Ready-To-Go, Pharmacia), including 500 ngof total RNA (dissolved in RNase free water) into a total reaction volume of50 ml. The PCR mixture of 50ml contained 7.5 mM (NH4)2SO4, 30 mM
Tris-HCl (pH 8.5), 1.75 mM MgCl2 (pH 8.5), all of the four deoxynucleotidetriphosphates (250mM each), 25 pmol each of the primers (Table 1), 2.5 unitsof Taq polymerase, and 2ml of cDNA (undiluted and, if required, in appro-priate serial dilution). The PCR conditions included an initial denaturation—2min at 95°C, 29 cycles consisting of (a) 1 min denaturation at 94°C; (b) 1 minprimer annealing at 54°C for topoisomerase IIa, 60°C for topoisomerase IIb,60°C for c-erbB-2, and 64°C for GAPDH, respectively; (c) 1 min elongationat 72°C; and (d) one final step of 5 min at 72°C. The resulting DNA fragmentswere separated by electrophoresis on a 1.5% agarose gel containing 0.4mg ofethidium bromide/ml, visualized by UV, and quantified densitometrically. Ifsingle bands were too intensive for densitometric determination (out of the
linear range), they were diluted serially down to the linear range, and theintensity of the original band was calculated by multiplication with the dilutionfactor. To avoid artificial differences between individual gels, aliquots of thesame DNA standard (a PCR product of GAPDH) were included in each gel,and the ratio of the relative intensities of individual samples:this standard werecalculated. The latter values obtained for topoisomerase IIa, b, andc-erbB-2were divided by the value of GAPDH of the respective tumor, to obtain therelative mRNA expression factor. For instance a relative mRNA expressionfactor of 0.5 forc-erbB-2means that the intensity of the amplification productof c-erbB-2 was one-half of the intensity obtained for the housekeepingenzyme GAPDH. The coefficients of variation of relative mRNA expressionfactors ofc-erbB-2were#20% if the same RNA preparation was analyzed in6 independent experiments. If RNA was isolated from 6 different sites (eachapproximately 1 mm3), the analyzed mRNA (topoisomerase IIa, b, or c-erbB-2) was consistently either detectable or not detectable at all of the sixsites. However, if mRNA expression was detectable, the variability wasrelatively high (coefficients of variation from six different sites of each tumorfor topoisomerase IIa andb, 58 and 51% in tumor 1 and 51 and 73% for tumor2, respectively). Because of this relatively high degree of heterogeneity, tumorswere classified as either positive or negative for mRNA expression of topoi-somerase IIa, b, or c-erbB-2. A typical result of semiquantitative PCR withdifferent volumes of cDNA given to the amplification reaction shows asufficient quantifiability (Fig. 1).
Chemosensitivity Assay with Primary Tumor Cells. The chemosensitiv-ity assay described by Andreottiet al. and Kurbacheret al. (17, 18) was usedwith modifications. Tissue specimens (approximately 0.5 cm3) from ovariantumors were taken intraoperatively, immediately transferred into cold (4°C)serum-free DMEM (Life Technologies, Inc.) containing 300 units/ml penicillinand 300mg/ml streptomycin, and were processed within 2 h after surgery. Thetissue specimens were minced into approximately 1-mm3 fragments understerile conditions. Fragments were then dissociated into a cell suspension ofsingle cells and small aggregates of single cells by incubation in 10 ml of CAM(Complete Assay Medium; DCS Innovative Diagnostik Systeme, Hamburg,Germany) with 1.5 mg/ml collagenase (Type: CLS II, 375 units/mg, Seromed,Berlin, Germany) at 37°C for 4 h on a gently shaking platform. Afterward thesuspension was centrifuged at 4003 g for 5 min (room temperature), resus-pended in PBS, and centrifuged and resuspended for a second time. Large cellaggregates in the latter suspension were allowed to sedimentate for 1 min, andthe supernatant was further purified by density-gradient centrifugation usingFicoll-Separating-Solution (Seromed, Berlin), to remove erythrocytes and deadcells. The cells from the interphase were washed twice in PBS and resuspendedin a chemically defined serum-free medium (Complete Assay Medium; DCSInnovative Diagnostik Systeme, Hamburg, Germany). Several collagenasebatches had to be tested to identify an appropriate batch that allowed anaverage cell harvest of at least 15 million cells (trypan blue exclusion,$95%)using ovarian tumor tissue specimens of approximately 0.5 cm2. Cultures of20,000 cells in a total volume of 200ml of CAM per well were incubated withthe test substances in 96-well round-bottomed polypropylene microplates(Costar 3790). Stock solutions of carboplatin (10 mg/liter; Sigma) and novo-biocin (254ml/ml; Sigma) were prepared in CAM, whereas the stock solutionof emodin (6480mg/ml; Sigma) was prepared in ethanol. Etoposide (VePeside;20 mg/ml) was obtained from Bristol (Munich, Germany). The stock solutionswere further diluted in CAM. All of the incubations with test substances wereperformed in triplicate, with the exception of the negative and positive controls(the latter incubated with 0.1 ml of Maximum-ATP-Inhibitor; DCS InnovativeDiagnostik Systeme, Hamburg, Germany), which were incubated in six wells.
3 The abbreviations used are: FIGO, International Federation of Gynecologists andObstetricians; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; SHC,src homologycollagen; GRB2, growth factor receptor-bound protein 2; MAP kinase, mitogen-activatedprotein kinase.
Table 1 Primer sequences
Primerdesignation Sequence
Productlength
Annealingtemperature
Topoisomerase Forward:GCCCTCCTGCTACACATTTC 596bp 54°CIIa Reverse:AACACTTGGGCTTTACTTCA CTT
Topoisomerase Forward:ATCAAAAGCCACTCCAGAAA AATC508bp 60°CIIb Reverse:AGAAGGTGGCTCAGTAGGGA AGTC
C-erbB-2 Forward: TGCGGCTCGTACACAGGGAC TT420bp 60°CReverse:TGCGGGAGAATTCAGACACC AACT
GAPDH Forward:GTCTTCACCACCATGGAGAA GGCT393bp 64°CReverse:CATGCCAGTGAGCTTCCCGT TCA
3207
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

After incubation of the cells for 7 days at 37°C in a.98% humidified, 95%air-5% CO2 atmosphere, cellular ATP was extracted and stabilized by mixing50 ml of Tumor-Cell-Extraction-Reagent (DCS Innovative DiagnostikSysteme, Hamburg, Germany) into each well. Twenty min after the addition ofTumor-Cell-Extraction-Reagent, the ATP content was measured in a BertholdLB-953 luminometer using 50ml of culture extract injected with 50ml ofLuciferin-Luciferase-Reagent (DCS Innovative Diagnostik Systeme, Ham-burg, Germany). An ATP standard curve was performed using 50-ml aliquotsof a 250-ng/ml ATP standard serially diluted 1:3 in dilution buffer. ATPcontent of solvent-treated control cells was defined as 100%. The additivity ofsubstances (inhibitors) with carboplatin was determined as follows: the meanATP value was defined as the mean ATP content (% of controls) of cellsincubated with 1, 2, 4, 8, 16, and 32mg/ml of carboplatin. The effect of thecombination of carboplatin with an inhibitor was defined as “less than addi-tive” if the difference between the mean ATP value of carboplatin alone andthe mean ATP value of carboplatin combined with inhibitor was smaller thanthe reduction in ATP content by the inhibitor alone.
Isolation of RNA and S1 Nuclease Assay.Total RNA was isolated by asingle-step method using the guanidinium thiocyanate-phenol-chloroform ex-
traction technique (19). The concentration of RNA was determined spectro-photometrically at 260 nm. After phenol extraction and ethanol precipitation,all of the RNA samples were stored as aliquots of 20mg at 280°C until use.All of the total RNAs prepared from cryopreserved tumor samples werechecked for integrity by conventional agarose gel electrophoresis before the S1nuclease assay, by means of the presence of 28S and 18S rRNA. DegradedRNA samples were excluded from the study. The S1 nuclease protectionassays were performed as described previously (8, 20). Briefly, theEcoRI/NcoIfragment of thec-erbB-2was isolated from the plasmid pMAC-14–1, whichcontains the full-lenghthc-erbB-2cDNA (21) and subcloned into theEcoRI/SmaI site of pSP65 involving first ligation at theEcoRI site and then filling intheNcoI site by T4-DNA-polymerase in the presence of dNTPs followed by asecond ligation of the remaining blunt ends. Radioactively labeled antisenseRNA of the c-erbB-2cDNA fragment (EcoRI/NcoI) was prepared byin vitrotranscription of the resulting construct (pSP-erbB-2) using SP6-polymerase(Boehringer Mannheim, Germany) andPvuII-linearized pSP-erbB-2-DNA.The transcript was purified by phenol extraction and ethanol precipitation. Atotal of 105 dpm of radioactively labeled transcript (specific activity 107
dpm/mg RNA) were mixed with 5mg of total tissue RNA and 100mg of
Fig. 1. Semiquantitative PCR with primers forc-erbB-2and topoisomerase IIa andb using different volumes of cDNA obtained from the incubation of isolated RNA with reversetranscriptase.
Table 2 Correlation between c-erbB-2 and topoisomerase IIa and IIb mRNA expression
Expression of topoisomerase IIa mRNA (%) a
No. of cases (%)Expression of topoisomerase IIb mRNAb
No. of cases (%)
Not detectable Detectable Total Not detectable Detectable Total
FIGO stages I–IV (n 5 70)Expression ofc-erbB-2mRNANo. of cases
Not detectable 27 (56.2) 21 (43.8) 48 (100) 25 (52.1) 23 (47.9) 48 (100)Detectable 5 (22.7) 17 (77.3) 22 (100) 8 (36.4) 14 (63.6) 22 (100)
FIGO stages I, II (n 5 20)Expression ofc-erbB-2mRNANo. of cases
Not detectable 13 (76.5) 4 (23.5) 17 (100) 12 (70.6) 5 (29.4) 17 (100)Detectable 0 (0) 3 (100) 3 (100) 0 (0) 3 (100) 3 (100)
FIGO stages III, IV (n 5 50)Expression ofc-erbB-2mRNANo. of cases
Not detectable 14 (45.2) 17 (54.8) 31 (100) 13 (41.9) 18 (58.1) 31 (100)Detectable 5 (26.3) 14 (73.7) 19 (100) 8 (42.1) 11 (57.9) 19 (100)
a Correlation betweenc-erbB-2 and topoisomerase IIa mRNA expression in tumor tissue specimens of 70 patients with primary ovarian cancer determined by reverse PCR(P 5 0.009,x2 test).
b No significant correlation was obtained betweenc-erbB-2 and topoisomerase IIb mRNA expression in tumor tissue specimens of 70 patients with primary ovarian cancerdetermined by reverse PCR (P 5 0.221,x2 test).
3208
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
Escherichia colitRNA, dried in a speed vac, and resuspended in hybridizationbuffer (40 mM PIPES (pH 6.4), 1 mM EDTA, 400 mM NaCl, and 80%formamide). The RNA mix was denatured by incubation at 90°C for 10 min,and subsequent hybridization was carried out at 52°C for 3 h. Fifty units S1nuclease (Pharmacia) in 300mg S1-buffer (30 mM sodium acetate (pH 4.6), 50mM NaCl, and 1 mM ZnCl) was added and incubation was continued at 37°Cfor 60 min. After phenol-chloroform extraction and ethanol precipitation, thehybrids were dissolved in loading buffer, heat-denatured at 95°C for 15 min,and separated by denaturing PAGE (containing 8M urea). Quantitation ofmRNA expression was carried out by densitometry (Densitometer Elscript 400from Kirschmann). For quantification, two standards were included in eachgel. Because of densitometric determination, the intensity that the band ob-tained from standard 1 was defined as 0.05 (relativec-erbB-2mRNA expres-sion, arbitrary unit). Compared with standard 1, the intensity of standard 2 was0.5. Both of the standards were used to calculate the intensity of the bands ofeach tumor sample. The problem of false negative results as a consequence ofmRNA degradation was excluded by analyzing the samples for expressionGAPDH mRNA. The coefficient of variation of five determinations of thesame RNA preparation was smaller than 20% (three tumors tested). In thepresent study, mean values of two determinations (of a single RNA prepera-tion) were calculated for each tumor.
Statistical Analysis. Kaplan-Meier curves were plotted to assess overallsurvival (22). Different survival curves were compared using the log-rank test. Theproportional hazard modal (22) was applied to examine whetherc-erbB-2was anindependent prognostic factor (univariate and multivariate analysis). A possibleassociation between expression ofc-erbB-2and topoisomerase IIa or IIb wasinvestigated using thex2 test. To examine whether the correlation betweenc-erbB-2 and topoisomerase IIa is independent from FIGO stage, the Cochran-Mantel-Haenszel test adjusted for FIGO stage I, IIversusIII, IV was applied. Allof the two-sidedP-values smaller than 0.05 were considered as statisticallysignificant. Statistical analysis was performed using the SAS software (Cary, NC).
RESULTS
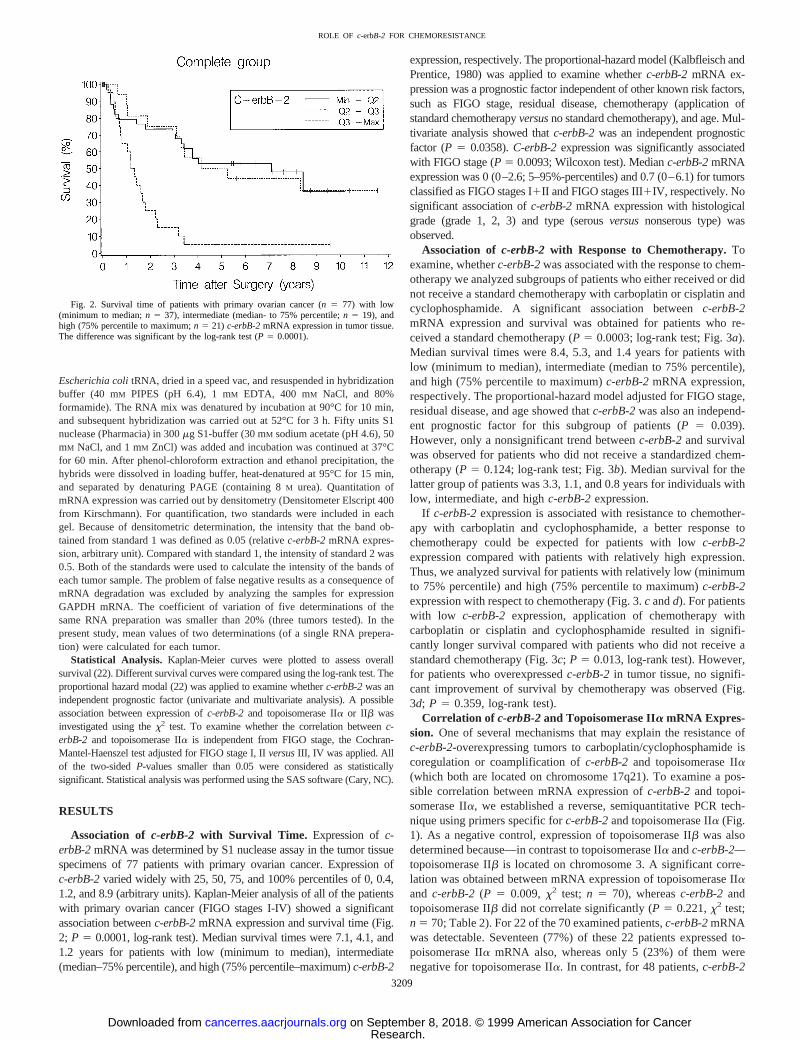
Association of c-erbB-2 with Survival Time. Expression ofc-erbB-2mRNA was determined by S1 nuclease assay in the tumor tissuespecimens of 77 patients with primary ovarian cancer. Expression ofc-erbB-2varied widely with 25, 50, 75, and 100% percentiles of 0, 0.4,1.2, and 8.9 (arbitrary units). Kaplan-Meier analysis of all of the patientswith primary ovarian cancer (FIGO stages I-IV) showed a significantassociation betweenc-erbB-2mRNA expression and survival time (Fig.2; P 5 0.0001, log-rank test). Median survival times were 7.1, 4.1, and1.2 years for patients with low (minimum to median), intermediate(median–75% percentile), and high (75% percentile–maximum)c-erbB-2
expression, respectively. The proportional-hazard model (Kalbfleisch andPrentice, 1980) was applied to examine whetherc-erbB-2mRNA ex-pression was a prognostic factor independent of other known risk factors,such as FIGO stage, residual disease, chemotherapy (application ofstandard chemotherapyversusno standard chemotherapy), and age. Mul-tivariate analysis showed thatc-erbB-2was an independent prognosticfactor (P 5 0.0358).C-erbB-2expression was significantly associatedwith FIGO stage (P 5 0.0093; Wilcoxon test). Medianc-erbB-2mRNAexpression was 0 (0–2.6; 5–95%-percentiles) and 0.7 (0–6.1) for tumorsclassified as FIGO stages I1II and FIGO stages III1IV, respectively. Nosignificant association ofc-erbB-2mRNA expression with histologicalgrade (grade 1, 2, 3) and type (serousversusnonserous type) wasobserved.
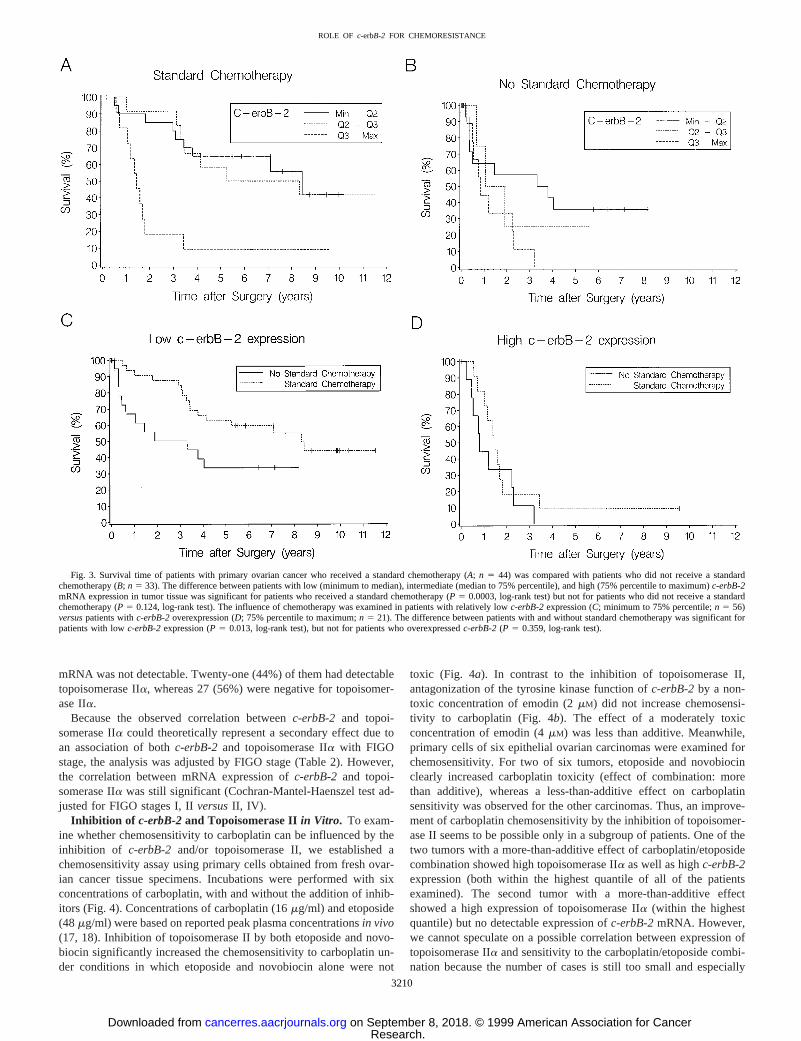
Association of c-erbB-2 with Response to Chemotherapy.Toexamine, whetherc-erbB-2was associated with the response to chem-otherapy we analyzed subgroups of patients who either received or didnot receive a standard chemotherapy with carboplatin or cisplatin andcyclophosphamide. A significant association betweenc-erbB-2mRNA expression and survival was obtained for patients who re-ceived a standard chemotherapy (P5 0.0003; log-rank test; Fig. 3a).Median survival times were 8.4, 5.3, and 1.4 years for patients withlow (minimum to median), intermediate (median to 75% percentile),and high (75% percentile to maximum)c-erbB-2mRNA expression,respectively. The proportional-hazard model adjusted for FIGO stage,residual disease, and age showed thatc-erbB-2was also an independ-ent prognostic factor for this subgroup of patients (P5 0.039).However, only a nonsignificant trend betweenc-erbB-2and survivalwas observed for patients who did not receive a standardized chem-otherapy (P5 0.124; log-rank test; Fig. 3b). Median survival for thelatter group of patients was 3.3, 1.1, and 0.8 years for individuals withlow, intermediate, and highc-erbB-2expression.
If c-erbB-2expression is associated with resistance to chemother-apy with carboplatin and cyclophosphamide, a better response tochemotherapy could be expected for patients with lowc-erbB-2expression compared with patients with relatively high expression.Thus, we analyzed survival for patients with relatively low (minimumto 75% percentile) and high (75% percentile to maximum)c-erbB-2expression with respect to chemotherapy (Fig. 3.c andd). For patientswith low c-erbB-2 expression, application of chemotherapy withcarboplatin or cisplatin and cyclophosphamide resulted in signifi-cantly longer survival compared with patients who did not receive astandard chemotherapy (Fig. 3c;P 5 0.013, log-rank test). However,for patients who overexpressedc-erbB-2 in tumor tissue, no signifi-cant improvement of survival by chemotherapy was observed (Fig.3d; P 5 0.359, log-rank test).
Correlation of c-erbB-2and Topoisomerase IIa mRNA Expres-sion. One of several mechanisms that may explain the resistance ofc-erbB-2-overexpressing tumors to carboplatin/cyclophosphamide iscoregulation or coamplification ofc-erbB-2 and topoisomerase IIa(which both are located on chromosome 17q21). To examine a pos-sible correlation between mRNA expression ofc-erbB-2and topoi-somerase IIa, we established a reverse, semiquantitative PCR tech-nique using primers specific forc-erbB-2and topoisomerase IIa (Fig.1). As a negative control, expression of topoisomerase IIb was alsodetermined because—in contrast to topoisomerase IIa andc-erbB-2—topoisomerase IIb is located on chromosome 3. A significant corre-lation was obtained between mRNA expression of topoisomerase IIaand c-erbB-2 (P 5 0.009,x2 test; n 5 70), whereasc-erbB-2andtopoisomerase IIb did not correlate significantly (P5 0.221,x2 test;n 5 70; Table 2). For 22 of the 70 examined patients,c-erbB-2mRNAwas detectable. Seventeen (77%) of these 22 patients expressed to-poisomerase IIa mRNA also, whereas only 5 (23%) of them werenegative for topoisomerase IIa. In contrast, for 48 patients,c-erbB-2
Fig. 2. Survival time of patients with primary ovarian cancer (n 5 77) with low(minimum to median;n 5 37), intermediate (median- to 75% percentile;n 5 19), andhigh (75% percentile to maximum;n 5 21) c-erbB-2mRNA expression in tumor tissue.The difference was significant by the log-rank test (P 5 0.0001).
3209
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
mRNA was not detectable. Twenty-one (44%) of them had detectabletopoisomerase IIa, whereas 27 (56%) were negative for topoisomer-ase IIa.
Because the observed correlation betweenc-erbB-2 and topoi-somerase IIa could theoretically represent a secondary effect due toan association of bothc-erbB-2 and topoisomerase IIa with FIGOstage, the analysis was adjusted by FIGO stage (Table 2). However,the correlation between mRNA expression ofc-erbB-2 and topoi-somerase IIa was still significant (Cochran-Mantel-Haenszel test ad-justed for FIGO stages I, IIversusII, IV).
Inhibition of c-erbB-2and Topoisomerase IIin Vitro. To exam-ine whether chemosensitivity to carboplatin can be influenced by theinhibition of c-erbB-2 and/or topoisomerase II, we established achemosensitivity assay using primary cells obtained from fresh ovar-ian cancer tissue specimens. Incubations were performed with sixconcentrations of carboplatin, with and without the addition of inhib-itors (Fig. 4). Concentrations of carboplatin (16mg/ml) and etoposide(48mg/ml) were based on reported peak plasma concentrationsin vivo(17, 18). Inhibition of topoisomerase II by both etoposide and novo-biocin significantly increased the chemosensitivity to carboplatin un-der conditions in which etoposide and novobiocin alone were not
toxic (Fig. 4a). In contrast to the inhibition of topoisomerase II,antagonization of the tyrosine kinase function ofc-erbB-2by a non-toxic concentration of emodin (2mM) did not increase chemosensi-tivity to carboplatin (Fig. 4b). The effect of a moderately toxicconcentration of emodin (4mM) was less than additive. Meanwhile,primary cells of six epithelial ovarian carcinomas were examined forchemosensitivity. For two of six tumors, etoposide and novobiocinclearly increased carboplatin toxicity (effect of combination: morethan additive), whereas a less-than-additive effect on carboplatinsensitivity was observed for the other carcinomas. Thus, an improve-ment of carboplatin chemosensitivity by the inhibition of topoisomer-ase II seems to be possible only in a subgroup of patients. One of thetwo tumors with a more-than-additive effect of carboplatin/etoposidecombination showed high topoisomerase IIa as well as highc-erbB-2expression (both within the highest quantile of all of the patientsexamined). The second tumor with a more-than-additive effectshowed a high expression of topoisomerase IIa (within the highestquantile) but no detectable expression ofc-erbB-2mRNA. However,we cannot speculate on a possible correlation between expression oftopoisomerase IIa and sensitivity to the carboplatin/etoposide combi-nation because the number of cases is still too small and especially
Fig. 3. Survival time of patients with primary ovarian cancer who received a standard chemotherapy (A; n 5 44) was compared with patients who did not receive a standardchemotherapy (B;n 5 33). The difference between patients with low (minimum to median), intermediate (median to 75% percentile), and high (75% percentile to maximum)c-erbB-2mRNA expression in tumor tissue was significant for patients who received a standard chemotherapy (P 5 0.0003, log-rank test) but not for patients who did not receive a standardchemotherapy (P5 0.124, log-rank test). The influence of chemotherapy was examined in patients with relatively lowc-erbB-2expression (C; minimum to 75% percentile;n 5 56)versuspatients withc-erbB-2overexpression (D; 75% percentile to maximum;n 5 21). The difference between patients with and without standard chemotherapy was significant forpatients with lowc-erbB-2expression (P5 0.013, log-rank test), but not for patients who overexpressedc-erbB-2(P 5 0.359, log-rank test).
3210
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

because one of the tumors with a less-than-additive effect for thecarboplatin/etoposide combination showed a high expression of to-poisomerase IIa mRNA.
DISCUSSION
A large number of studies have suggested the involvement of someoncogenes in the development of cytotoxic drug resistance. For in-stance, transfection ofc-myc into several cell lines was shown toincrease resistance to cisplatin (23, 24), whereas transfection with
antisensec-myc increased sensitivity to cisplatin (25). Similarly,transfection ofH-ras into MCF-7cells (26) or into NIH 3T3 cells (27)markedly increased resistance to cisplatin. However, the relationshipbetween oncogene expression and drug resistance may be indirectbecause the transfectionof v-H-ras or v-raf into rat liver epithelialcells led to an increased expression ofP-glycoprotein (mdr-1) andglutathioneS-transferasep, which may cause resistance (28). Thus,some oncogenes are unlikely to modify drug sensitivity directly butmay influence expression of other genes that are responsible for drugresistance.
In the present study, we show that overexpression of the oncogenec-erbB-2 is associated with shorter survival time of ovarian cancerpatients, confirming results of earlier investigations (7–9, 29). Inaddition, expression ofc-erbB-2mRNA was significantly associatedwith FIGO stage, which gives a measure of tumor extension. Thus,overexpression ofc-erbB-2seems to increase with tumor progression.Despite its association with FIGO stage,c-erbB-2mRNA expressionwas also an independent prognostic factor in the proportional-hazardmodel (Cox model) adjusted for FIGO stage, residual disease, appli-cation of standard chemotherapy, and age, which represent the mostimportant classical prognostic factors of ovarian cancer.
To examine a possible involvement ofc-erbB-2in the developmentof cytotoxic drug resistance, we compared survival times of patientswho received a standard chemotherapy with those of patients who didnot receive a standard chemotherapy. A highly significant associationbetween survival and expression ofc-erbB-2mRNA was obtained forpatients with standard chemotherapy, whereas only a nonsignificanttrend toward longer survival was observed for patients without stand-ard chemotherapy. In addition, the application of a standard chemo-therapy increased the survival time of patients with lowc-erbB-2expression but not of patients with c-erbB-2 overexpression. Thesedata suggest thatc-erbB-2 overexpression may be involved in thedevelopment of resistance against chemotherapy with cisplatin orcarboplatin and cyclophosphamide. However, this conclusion still hasto be treated with caution because the selection of patients withoutstandard chemotherapy may cause a bias. Patients without standardchemotherapy received no doses or only very low doses of platinumcompounds and cyclophosphamide because of nephrotoxicity or my-elotoxicity or because patients refused to receive chemotherapy. Thefirst two reasons (nephrotoxicity and myelotoxicity) may represent aselection criterion that possibly exerts an influence on prognosisindependent of chemotherapy. Nevertheless, the evidence thatc-erbB-2 may be involved in the development of drug resistance wasused as a working hypothesis and was further investigated.
If c-erbB-2is involved in the development of drug resistance, twopossibilities seem to be plausible: (a) a mechanism mediated by thetyrosine kinase function ofc-erbB-2; and (b) coamplification and/orcoregulation ofc-erbB-2with topoisomerase IIa (Fig. 5). The firstmechanism includes heterodimerization ofc-erbB-2with other mem-bers of the class I family of receptor tyrosine kinases, such as theepidermal growth factor receptor, HER-3, or HER-4 (1), which gen-erates a high affinity receptor for a family of ligands termed neuregu-lins (Fig. 5; Ref. 30). Activation of thec-erbB-2 receptor inducestyrosine phosphorylation on specific tyrosine residues in the COOHterminus ofc-erbB-2 (31, 32). After phosphorylation, downstreamsignaling proteins such as SHC and/or GRB2 bind to the COOHterminus of c-erbB-2 through src homology 2 domains (33, 34).Interaction of the guanine nucleotide exchange factor (SOS) withSHC/GRB2 brings the complex to the cytoplasmic surface of theplasma membrane, in which SOS activatesras-GDP toras-GTP (35),which leads to the activation of MAP kinase (36) and culminates inthe transcription of nuclear transcription factors, such asc-myc,c-fos,andc-jun. The effects of the latter products may explain not only an
Fig. 4. Primary cells obtained from an epithelial ovarian carcinoma were incubatedwith carboplatin for 7 days with and without etoposide, novobiocin, or emodin. Bothtopoisomerase II inhibitors (etoposide and novobiocin);A, enhanced carboplatin cytotox-icity in nontoxic concentrations, whereas a nontoxic concentration of the tyrosine kinaseinhibitor emodin (2mM); B, showed no effect.
3211
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

increased mitogenic activity but possibly also chemoresistance be-cause transfection ofc-myc into Friend erythroleukemia cells wasshown to decrease chemosensitivity to cisplatin (24). However, re-sistance to cisplatin may also be caused by MAP kinase-mediatedtranscription of yet unknown other genes involved in DNA excisionrepair or in inactivation of cisplatin. Similarly, thec-erbB-2-mediatedphosphodiesterase phospholipase C-g or phosphatidylinositol 3-ki-nase pathways may induce the transcription of resistance-associatedgenes, although there is no current experimental evidence for this.
The second mechanism by whichc-erbB-2may be associated withresistance to platinum compounds is coamplification or coregulationwith topoisomerase IIa. The chromosomal location of both,c-erbB-2and topoisomerase IIa is 17q21–22. Amplifications occur over largesegments of the chromosome and may comprise many mega bases(37). Topoisomerase IIa was found to be coamplified in 12% ofc-erbB-2amplified cases (38) and similar results were obtained byother investigators (39). Thus, a subgroup of patients withc-erbB-2overexpression is likely to have a high topoisomerase IIa activity.High activity of topoisomerase IIa is known to be associated withhigh sensitivity toward topoisomerase II-inhibitors such as anthracy-clines and epipodophyllotoxines. However, for platinum compoundsand alkylating antineoplastic agents, the opposite constellation hasbeen shown. Topoisomerase IIa-transfected cell lines were;5- to10-fold more resistant to cisplatin and the alkylating agent mechlor-ethamine (40). In addition, several types of topoisomerase IIa inhib-itors enhanced the toxicity of platinum compounds and alkylatingagents in topoisomerase IIa-expressing cellsin vitro (41–43). Be-cause the inhibition of topoisomerase IIa significantly decreased the
repair of DNA interstrand cross-links with a corresponding decreasein the clonogenic survival of the cells (44–45), topoisomerase IIapossibly influences chemosensitivity by altering the accessibility ofDNA sequences for alkylation and for subsequent repair, although thishas not yet been shown directly.
The hypothesis that coamplified and/or coregulated topoisomeraseIIa is responsible for the chemoresistance ofc-erbB-2-overexpressingcarcinomas, to our knowledge, has not yet been investigated. Weobserved a significant correlation betweenc-erbB-2and topoisomer-ase IIa mRNA expression in primary ovarian cancer tissue specimensthat was independent of the FIGO stage. In contrast, topoisomeraseIIb, which is located on chromosome 3, did not correlate withc-erbB-2. Because we examined mRNA expression, which was chosenbecause it is a step closer to the functional protein than DNA analyzedby Southern blotting, it is not possible to differentiate whether thiscorrelation is due to coamplification or coregulation, and it was notthe aim of the present study to analyze the molecular mechanism ofthe correlation betweenc-erbB-2 and topoisomerase IIa. Besidescoamplification, a likely explanation for the correlation betweenc-erbB-2and topoisomerase IIa may be ac-erbB-2-mediated stimula-tion of cell division (1, 10–12), which may cause an increase intopoisomerase IIa expression because topoisomerase IIa is requiredfor relieving torsional stress during DNA replication, is up-regulatedin actively dividing cells, and is down-regulated by the end of mitosis(38). The hypothesis that coamplified and/or coregulated topoisomer-ase IIa contributes to the resistance ofc-erbB-2-overexpressing car-cinomas was additionally supported by an examination of primarycells obtained from an ovarian carcinoma that overexpressed both
Fig. 5. Possible mechanisms ofc-erbB-2-induced chemoresistance.GRB2, growth factor receptor-bound protein 2;LD, ligand-binding domain;n, neuregulins;SHC,src-homologycollagen;SOS, guanine nucleotide exchange factor;TK, tyrosine kinase domain;Y, auto-phosphorylation domain. (The signal transduction pathways were adapted from Reese andSlamon (1).
3212
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

c-erbB-2and topoisomerase IIa mRNA. The combination of carbo-platin with nontoxic concentrations of the topoisomerase II-inhibitorsetoposide or novobiocin significantly enhanced toxicity in the primaryovarian cancer cells. Using the pharmacological inhibitors etoposideand novobiocin, we cannot exclude nonspecific effects. However, theenhanced sensitivity caused by etoposide or novobiocin is unlikely tobe caused by a decreased capacity of the primary tumor cells (Fig. 4)to repair a certain threshold quantity of DNA damage in generalbecause these cells were relatively resistant to carboplatin. The ATPcontent of the cells shown in Fig. 4 was 69% of controls afterincubation with 16mg of carboplatin/ml compared with the meanATP-content of 416 19% (mean6 SD) of 14 examined ovariancancer tissue specimens. In contrast to the topoisomerase II inhibitors,the combination of carboplatin with the tyrosine kinase inhibitoremodin under conditions known to antagonizec-erbB-2function (14)did not enhance carboplatin toxicity. Obviously, the sensitivity of theexamined primary ovarian cancer cells can be improved by the inhi-bition of topoisomerase II but not by the antagonization ofc-erbB-2function, which shows thatc-erbB-2is not necessarily associated withsusceptibility to carboplatin. Of course, this observation does notexclude the possibility thatc-erbB-2may be involved in the devel-opment of drug resistance in other cell types.
There have been clinical trials of topoisomerase inhibitors andplatinum complexes in ovarian cancer patients. For instance, i.p.application of cisplatin (or carboplatin) and etoposide as a consolida-tion therapy for advanced epithelial ovarian cancer resulted in asignificant—although not very large—increase in disease-free sur-vival time (46, 47). In our study with primary ovarian cancer cells, aclear improvement of carboplatin chemosensitivity by etoposide wasobserved only in a subgroup of patients. Thus, identification of thissubgroup could result in a more specific and more effective applica-tion of carboplatin/etoposide combination chemotherapy. Presently,we are investigating whether this subgroup can be identified by thedetermination of topoisomerase IIa expression.
In conclusion, overexpression ofc-erbB-2was associated with poorprognosis and poor response to platinum compounds and cyclophos-phamide in patients with primary ovarian cancer. Topoisomerase IIa,which correlates withc-erbB-2expression, is likely to contribute tothe resistance ofc-erbB-2-overexpressing carcinomas.
ACKNOWLEDGMENTS
We thank S. Gebhard for excellent organization and documentation of ourovarian cancer tumor bank and H. Hall for suggestions on the manuscript.
REFERENCES
1. Reese, D. M., and Slamon, D. J. HER-2/neu signal transduction in human breast andovarian cancer. Stem Cells,15: 1–8, 1997.
2. Shih, C., Padhy, L. C., Murray, M., and Weinberg, R. A. Transforming genes ofcarcinomas and neuroblastomas introduced into mouse fibroblasts. Nature (Lond.),290: 261–264, 1981.
3. Bargmann, C. I., Hung, M. C., and Weinberg, R. A. Multiple independent activationsof theneuoncogene by a point mutation altering the transmembrane domain of p185.Cell, 45: 649–657, 1986.
4. Slamon, D. J., Godolphin, W., Jones, L. A., Holt, J. A., Wong, S. G., Keith, D. E.,Levin, W. J, Stuart, S. G., Udove, J., Ullrich, A., Press, M. F. Studies of theHER-2/neuproto-oncogene in human breast and ovarian cancer. Science (WashingtonDC), 244: 707–712, 1989.
5. Venter, D. J., Tuzi, N. L., Kumar, S., and Gullick, W. J. Overexpression of thec-erbB-2 oncoprotein in human breast carcinomas: immunohistological assessmentcorrelates with gene amplification. Lancet,2: 69–72, 1987.
6. Slamon, D. J., Clark, G. M., Wong, S. G., Levin, W. J., Ullrich, A., and McGuire,W. L. Human breast cancer: correlation of relapse and survival with amplification ofthe HER-2/neuoncogene. Science (Washington DC),235: 177–182, 1987.
7. Meden, H., Marx, D., Roegglen, T., Schauer, A., and Kuhn, W. Overexpression of theoncogenec-erbB-2 (HER2/neu) and response to chemotherapy in patients withovarian cancer. Int. J. Gynecol. Pathol.,17: 61–65, 1998.
8. Tanner, B., Kreutz, E., Weikel, W., Meinert, R., Oesch, F., Knapstein, P. G., andBecker, R. Prognostic significance of c-erB-2 mRNA in ovarian carcinoma. Gynecol.Oncol.,62: 268–277, 1996.
9. Meden, H., and Kuhn, W., Overexpression of the oncogenec-erbB-2(HER2/neu) inovarian cancer: a new prognostic factor. Eur. J. Obstet. Gynecol. Reprod. Biol.,71:173–179, 1997.
10. Hudziak, R. M., Schlessinger, J., and Ullrich, A. Increased expression of the putativegrowth factor receptor p185HER2 causes transformation and tumorigenesis of NIH3T3 cells. Proc. Natl. Acad. Sci. USA,84: 7159–7163, 1987.
11. Di-Fiore, P. P., Pierce, J. H., Kraus, M. H., Segatto, O., King, C. R., Aaronson, S. A.erbB-2 is a potent oncogene when overexpressed in NIH/3T3 cells. Science (Wash-ington DC),237: 178–182, 1987.
12. Pietras, R. J., Arboleda, J., Reese, D. M., Wongvipat, N., Pegram, M. D., Ramos, L.,Gorman, C. M., Parker, M. G., Sliwkowski, M. X., and Slamon, D. J. HER-2 tyrosinekinase pathway targets estrogen receptor and promotes hormone-independent growthin human breast cancer cells. Oncogene,10: 2435–2446, 1995.
13. Hancock, M. C., Langton, B. C., Chan, T., Toy P., Monahan, J. J., Mischak, R. P., andShawver, L. K. A monoclonal antibody against thec-erbB-2protein enhances thecytotoxicity of cis-diamminedichloroplatinum against human breast and ovariantumor cell lines. Cancer Res.,51: 4575–4580, 1991.
14. Zhang, L., and Hung, M. C. Sensitization of HER-2/neu-overexpressing non-smallcell lung cancer cells to chemotherapeutic drugs by tyrosine kinase inhibitor emodin.Oncogene,12: 571–176, 1996.
15. Pegram, M. D., Finn, R. S., Arzoo, K., Beryt, M., Pietras, R. J., and Slamon, D. J. Theeffect of HER-2/neu overexpression on chemotherapeutic drug sensitivity in humanbreast and ovarian cancer cells. Oncogene15: 537–547, 1997.
16. Czernobilski, B. Blaustein’s Pathology of the Female Genital Tract, pp. 560–607.New York: Springer, 1987.
17. Andreotti, P. E., Cree, I. A., Kurbacher, C. M., Hartmann, D. M., Linder, D., Harel,G., Gleiberman, I., Caruso, P. A., Ricks, S. H., Untch, M., Sartori, C., and Bruckner,H. W. Chemosensitivity testing of human tumors using a microplate adenosinetriphosphate luminescence assay: clinical correlation for cisplatin resistance of ovar-ian carcinoma. Cancer Res.,55: 5276–5282, 1995.
18. Kurbacher, C. M., Cree, I. A., Bruckner, H. W., Brenne, U., Kurbacher, J. A., Muller,K., Ackermann, T., Gilster, T. J., Wilhelm, L. M., Engel, H., Mallmann, P. K., andAndreotti, P. E. Use of anex vivoATP luminescence assay to direct chemotherapy forrecurrent ovarian cancer. Anticancer Drugs,9: 51–57, 1998.
19. Chomczynski, P., and Sacchi, N. Single-step method of RNA isolation by acidguanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem.,162: 156–159, 1987.
20. Tanner, B., Hengstler, J. G., Laubscher, S., Meinert, R., Oesch, F., Weikel, W.,Knapstein, P. G., and Becker, R. Mdm 2 mRNA expression is associated with survivalin ovarian cancer. Int. J. Cancer,74: 438–442, 1997.
21. Kraus, M. H., Popescu, N. C., Amsbaugh, S. C., and King, C. R. Overexpression ofthe EGF receptor-related proto-oncogeneerbB-2in human mammary tumor cell linesby different molecular mechanisms. EMBO J.,6: 605–610, 1987.
22. Kalbfleisch, J. D., and Prentice, R. C. The Statistical Analysis of Failure Data. NewYork: Wiley, 1980.
23. Warenius, H. M., Seabra, L. A., and Maw, P. Sensitivity tocis-diamminedichloro-platinum in human cancer cells is related to expression of cyclin D1 but not c-raf-1protein. Int. J. Cancer:67: 224–231, 1996.
24. Sklar, M. D., and Prochownik, E. V. Modulation ofcis-platinum resistance in Frienderythroleukemia cells byc-myc. Cancer Res.,51: 2118–2123, 1991.
25. Van-Waardenburg, R. C., Meijer, C., Burger, H., Nooter, K., De-Vries, E. G., Mulder,N. H., and De-Jong, S. Effects of an inducible anti-sensec-mycgene transfer in adrug-resistant human small-cell-lung-carcinoma cell line. Int. J. Cancer,73: 544–550,1997.
26. Fan, J., Banerjee, D., Stambrook, P. J., and Bertino, J. R. Modulation of cytotoxicityof chemotherapeutic drugs by activated H-ras. Biochem. Pharmacol.,53: 1203–1209,1997.
27. Isonishi, S., Hom, D. K., Thiebaut, F. B., Mann, S. C., Andrews, P. A., Basu, A.,Lazo, J. S., Eastman, A., and Howell, S. B. Expression of the c-Ha-ras oncogene inmouse NIH 3T3 cells induces resistance to cisplatin. Cancer Res.,51: 5903–5909,1991.
28. Burt, R. K., Garfield, S., Johnson, K., and Thorgeirsson, S. S. Transformation of ratliver epithelial cells with v-H-ras or v-raf causes expression of MDR-1, glutathione-S-transferase-p and increased resistance to cytotoxic chemicals. Carcinogenesis(Lond.), 9: 2329–2332, 1988.
29. Berchuck, A., Kamel, A., Whitaker, R., Kerns, B., Olt, G., Kinney, R., Soper, J. T.,Dodge, R., Clarke-Pearson, D. L., Marks, P., McKenzie, S., Yin, S., and Bast, R. C.,Jr. Overexpression ofHER-2/neu is associated with poor survival in advancedepithelial ovarian cancer. Cancer Res.,50: 4087–4091, 1990.
30. Lee, K. F., Simon, H., Chen, H., Bates, B., Hung, M. C., and Hauser, C. Requirementfor neuregulin receptor erbB2 in neural and cardiac development. Nature (Lond.),378: 394–398, 1995.
31. Ullrich, A., and Schlessinger, J. Signal transduction by receptors with tyrosine kinaseactivity. Cell, 61: 203–212, 1990.
32. Chazin, V. R., Kaleko, M., Miller, A. D., and Slamon, D. J. Transformation mediatedby the humanHER-2 gene independent of the epidermal growth factor receptor.Oncogene,7: 1859–1866, 1992.
33. Pelicci, G., Lanfrancone, L., Grignani, F., McGlade, J., Cavallo, F., Forni, G.,Nicoletti, I., Grignani, F., Pawson, T., and Pelicci, P. G. A novel transforming protein(SHC) with an SH2 domain is implicated in mitogenic signal transduction. Cell,70:93–104, 1992.
3213
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

34. Egan, S. E., Giddings, B. W., Brooks, M. W., Buday, L., Sizeland, A. M., and Weinberg,R. A. Association of Sos Ras exchange protein with Grb2 is implicated in tyrosine kinasesignal transduction and transformation. Nature (Lond.),363: 45–51, 1993.
35. Li, N., Batzer, A., Daly, R., Yajnik, V., Skolnik, E., Chardin, P., Bar-Sagi, D.,Margolis, B., and Schlessinger, J. Guanine-nucleotide-releasing factor hSos1 binds toGrb2 and links receptor tyrosine kinases to Ras signalling. Nature (Lond.),363:85–88, 1993.
36. Johnson, G. L., and Vaillancourt, R. R. Sequential protein kinase reactions controllingcell growth and differentiation. Curr. Opin. Cell. Biol.,6: 230–238, 1994.
37. Smith, K. A., Gorman, P. A., Stark, M. B., Groves, R. P., and Stark, G. R. Distinctivechromosomal structures are formed very early in the amplification ofCAD genes inSyrian hamster cells. Cell,63: 1219–1227, 1990.
38. Smith, K., Houlbrook, S., Greenall, M., Carmichael, J., and Harris, A. L. Topoi-somerase IIa co-amplification with erbB2 in human primary breast cancer and breastcancer cell lines: relationship to m-AMSA and mitoxantrone sensitivity. Oncogene,8:933–938, 1993.
39. Keith, W. N., Douglas, F., Wishart, G. C., McCallum, H. M., George, W. D., Kaye,S. B., and Brown, R. Co-amplification of erbB2, topoisomerase IIa and retinoic acidreceptora genes in breast cancer and allelic loss at topoisomerase I on chromosome20. Eur. J. Cancer,29A: 1469–1475, 1993.
40. Eder, J. P., Jr., Chan, V. T-W., Ng, S-W., Rizvi, N. A., Zacharoulis, S., Teicher, B. A.,and Schnipper, L. E. DNA topoisomerase IIa expression is associated with alkylatingagent resistance. Cancer Res.,55: 6109–6116, 1995.
41. De-Jong, S., Timmer-Bosscha, H., De-Vries, E. G., and Mulder, N. H. Effect ofnovobiocin on cisplatin cytotoxicity and DNA interstrand cross-link formation in acisplatin-resistant, small-cell lung carcinoma cell line. Int. J. Cancer,53: 110–117,1993.
42. Hashimoto, S., Jing, Y., Kawazoe, N., Masuda, Y., Nakajo, S., Yoshida, T., Kuroiwa,Y., and Nakaya, K. Bufalin reduces the level of topoisomerase II in human leukemiacells and affects the cytotoxicity of anticancer drugs. Leuk Res.21: 875–883, 1997.
43. Barret, J. M., Calsou, P., Larsen, A. K., and Salles, B. A cisplatin-resistant murineleukemia cell line exhibits increased topoisomerase II activity. Mol. Pharmacol.,46:431–346, 1994.
44. Ali-Osman, F., Berger, M. S., Rajagopal, S., Spence, A., and Livingston, R. B.Topoisomerase II inhibition and altered kinetics of formation and repair of nitrosoureaand cisplatin-induced DNA interstrand cross-links and cytotoxicity in human glio-blastoma cells. Cancer Res.,53: 5663–5668, 1993.
45. Stevnsner, T., and Bohr, V. A. Studies on the role of topoisomerases in general, gene-and strand-specific DNA repair. Carcinogenesis (Lond.),14: 1841–1850, 1993.
46. Barakat, R. R., Almadrones, L., Venkatraman, E. S., Aghajanian, C., Brown, C.,Shapiro, F., Curtin, J. P., and Spriggs, D. A Phase II trial of intraperitonealcisplatin and etoposide as consolidation therapy in patients with stage II-IVepithelial ovarian cancer following negative surgical assessment. Gynecol. On-col., 69: 17–22, 1998.
47. Aziz, Z., Zahid, M., Ud Din Ahmed, Z., and Arshad, T. Phase II trial of ifosfamideand cisplatinum in advanced ovarian cancer. Aust. N. Z. J. Med.,28: 403– 409,1998.
3214
ROLE OF c-erbB-2 FOR CHEMORESISTANCE
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

1999;59:3206-3214. Cancer Res Jan G. Hengstler, Jost Lange, Alexandra Kett, et al. Chemoresistance in Ovarian Cancer
toα and Topoisomerase IIc-erbB-2Contribution of
Updated version
http://cancerres.aacrjournals.org/content/59/13/3206
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/59/13/3206.full#ref-list-1
This article cites 45 articles, 12 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/59/13/3206.full#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/59/13/3206To request permission to re-use all or part of this article, use this link
Research. on September 8, 2018. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from


















