
Composition & Architecture of plant viruses
46
Composition & Architecture of plant viruses P.N. Sharma Department of Plant Pathology, CSK HPKV, Palampur (H.P.)
Transcript of Composition & Architecture of plant viruses

Composition & Architecture
of plant viruses
P.N. Sharma
Department of Plant Pathology,
CSK HPKV, Palampur (H.P.)

Plant Viruses
Classification, Morphology, Genome,
and Structure

Importance
Detailed knowledge of virus structure is
important to understand different aspects of
virology e.g. how virus survive, infect, spread,
replicate and how they are related with one
other.
Knowledge of virus architecture has increased
greatly with the invention of EM, optical
defraction, X-ray crystallography procedures;
Molecular techniques
Chemical information about viruses

Morphology of Viruses
About ½ of all known plant viruses are
elongate (flexuous threads or rigid rods).
About ½ of all known plant viruses are
spherical (isometric or polyhedral).
A few viruses are cylindrical bacillus-like
rods.

Chemical composition of plant viruses
Protein( Capsid)
Capsomere
Nucleic acids
RNA
+ve strand RNA
-ve strand RNA
ssRNA
dsRNA
DNA
ssDNA
dsDNA

Viral Composition
Proteins 60-95% of the virion
Repeating subunits, identical for each virus type but varies from virus to virus and even from strain to strain TMV subunits - 158 amino acids with a mass of 17,600
Daltons (17.6 kDa, kd or K)
TYMV – 20,600 Dalton protein
Nucleic acid is 5-40% of the virion Spherical viruses: 20-40%
Helical viruses: 5-6%

Viral Composition
Nucleic acid (5-40%) represents the genetic material, indispensable for replication
Nucleic acid alone is sufficient for virus replication – Fraenkel-Conrat, Schramm
Protein (60-95%) protects virus genome from
degradation
facilitates movement through the host and
transmission from one host to another

A/a composition of capsid proteins of some viruses
1. Alanine
CMV: 17; PVY: 16
TMV: 14; PVX: 76
6. Glutamine
CMV: 20; PVY: 23
TMV: 16; PVX: 33
11. Leucine
CMV: 26; PVY: 10
TMV: 12; PVX: 19
16. Serine
CMV: 32; PVY: 10
TMV: 16; PVX: 31
2. Arginine
CMV: 24; PVY: 11
TMV: 11; PVX: 18
7. Glutamic acid 12. Lysine
CMV: 18; PVY: 13
TMV: 2; PVX: 22
17. Tryptophane
CMV: 1 ; PVY: 2
TMV: 3; PVX: 9
3. Asparatic acid
CMV: 30; PVY: 22
TMV: 18; PVX: 42
8. Glycine
CMV: 16; PVY: 13
TMV: 6 ; PVX: 23
13. Methionine
CMV: 8; PVY: 8
TMV: 0 ; PVX: 15
18. Tyrosine
CMV: 11; PVY: 6
TMV: 4; PVX: 4
4. Asparagines 9. histidine
CMV: 4; PVY: 4
TMV: - ; PVX: 4
14. Phenylalanine
CMV: 7 ; PVY: 5
TMV: 8; PVX: 22
19. Threonine
CMV: 17; PVY: 13
TMV: 16; PVX: 58
5. Cystein
CMV: 0; PVY: 1
TMV: 1; PVX: 5
10. Isoleucine
CMV: 16; PVY: 12
TMV: 9 ; PVX: 21
15. Proline
CMV: 18; PVY: 11
TMV: 8 ; PVX: 34
20. Valine
CMV: 22; PVY: 13
TMV: 14; PVX: 27
Total CMV: 287 PVY: 203 TMV: 158 PVX: 463

%age of protein & n/a in some viruses
%age of protein & n/a in some viruses
Virus n/a (%) Protein (%)
TMV 5 95
PVX 6 94
PVY 5 95
CpMV 31-33 67-69
CMV 18 82
TRSV 40 60

Viral Ultrastructure
Terminology for virus components
Capsid is the protein shell that encloses the nucleic acid
Capsomers are the morphological units seen on the surface of particles and represent clusters of structure units
Capsid and enclosed nucleic acid is called the nucleocapsid
The virion is the complete infectious virus particle
Caspar, D. L. D. and Klug, A. (1963) "Structure and Assembly of Regular Virus Particles." In Viruses, Nucleic Acids, and Cancer, 17th Annual Symposium on Fundamental Cancer Research, University of Texas, Williams and Wilkins, Baltimore, pp. 27-39.

Watson and Crick
In 1956 proposed:
Amount of the virus nucleic acid was
insufficient to code for more than a few
proteins of limited size
Therefore the protein shell must be of identical
subunits
Subunits had to be arranged to provide
each with an identical environment, i.e.,
symmetrical packing

Virus Architecture
Detailed knowledge of virus structure is
important to understand different aspects
of virology
Knowledge of virus architecture has
increased greatly with the innovation like
EM, optical defraction, X-Ray
crystallography procedures, mol.
techniques and chemical nature of the
virus.

Various feature of viruses can be
estimated by studying:
Chemical & biochemical studies
Size of particles Hydrodynamics
Laser scattering has been used to determine the radii of spherical viruses
E.M.
X-ray crystallography it gives accurate estimates of radius of icosahedral
viruses but condition is that the virus should be able to form stable crystals.

Electron microscopy
In 1924 L. de BROGLIE discovered the wave-character
of electron rays thus giving the prerequisite for the
construction of the electron microscope.
Invented by M. KNOLL and E. RUSKA (Technische
Universität Berlin, 1932).
One of the first biological objects observed was the
tobacco mosaic virus (TMV).
The first picture of a cell was published in 1945 by K. R.
PORTER, A. CLAUDE and E. F. FULLAM (Rockefeller
Institute, New York).
The Transmission Electron Microscope (TEM)
The Scanning electron microscope (SEM)
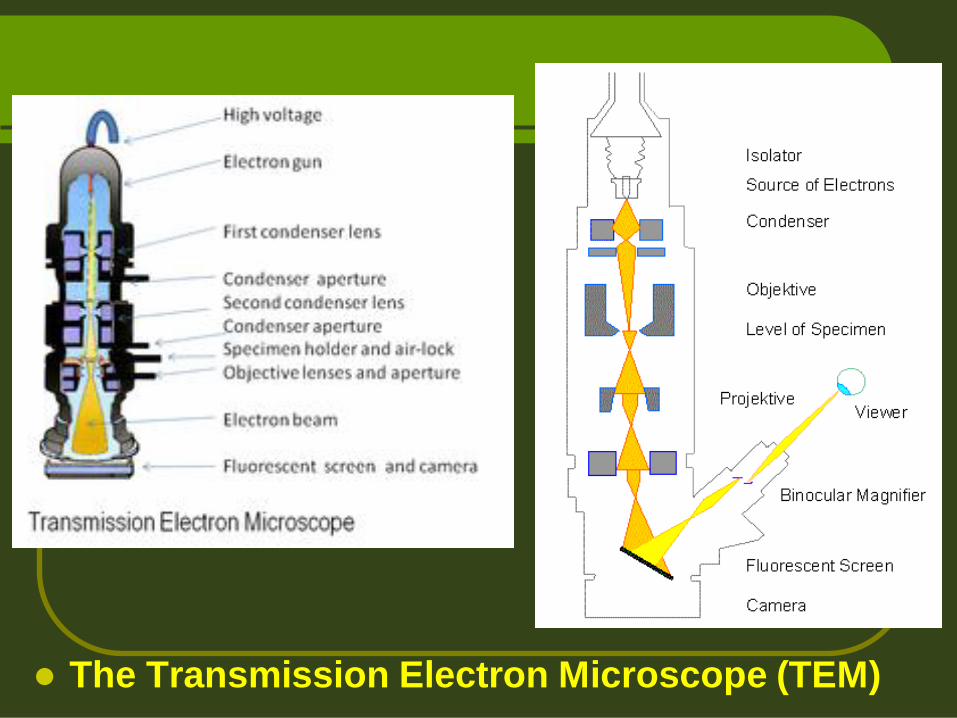
The Transmission Electron Microscope (TEM)

A 1973 Siemens electron microscope, EM developed by E. Ruska 1933
The Transmission Electron Microscope (TEM)

Fine structures determination
E.M.
Metal shadow preparations: using heavy metals, it enhances the contrast of particles
Freeze drying: useful about surface details particularly with lipid protein bilayer mambranes (Large viruses)
Negative staining: the use of electron dense stains is more important than heavy metals shadowing for morphological details. Such stains may be +ve or –ve

Positive stains React chemically with and are bound to virus
surface e.g. various Osmium, lead and uranyl compounds and phosphotungustic acid (PTA) are used under appropriate conditions. However, the chemical reaction may alter or disintegrate the virus so –ve stains are more important
Negative stains: They do not react with the virus but penetrate
available spaces on the surfaces or with in virus particle e.g. Uranylacetate or Potassium phosphotungstate (KPT) are used near pH 5.0
Fine structures determination

Thin sections
Cryo EM
X-ray crystallography analysis
Neutron small angle scattering: neutron scattering by virus solution is a method by which
low resolution information can be obtained about structure of virus. E.g. important for radii of isometric particles
Mass spectrography
Serological method's
Gel diffusion
ELISA
ISEM
Fine structures determination

Methods for studying stabilizing
bonds
The primary structure of viral CP & n/a depends upon covalent bonds.
Three kinds of interactions are involved in viruses : Protein : protein
Protein : RNA
RNA : RNA
In addition, small molecules e.g. divalent metal ions (CA2+ in particular) have marked effects on the stability of some viruses.
These interactions determine how much the virus is stable
How it might be assembled during virus synthesis
How viral n/a is released following infection of cell
These help the CP and n/a
to be held together
precisely

The stabilizing interactions are hydrophobic bonds, H= bonds, salt linkage etc. these interactions cab be studied by: X-ray crystallography
Stability to chemicals and physical agents: e.g. Phenol, urea, temperature and detergents etc.
Chemical modification of CP: a/a changes
Removal of ions: in viruses whose structure are stabilized by Ca2+ ions can be affected by their removal e.g. in isometric particles, CA2+ ions removal by EDTA causes swelling of the particles. So this phenomenon can give information about the kind of bond important fro virus stability.
Methods for studying stabilizing bonds

Circular dichroism: Spectra can be used
to obtain estimates of the extent of a-
helix and B- structure in a viral protein
subunit.
n/a tests
Methods for studying stabilizing bonds

Architecture of rod shaped viruses
Crick & Watson (1956) put forwarded a
hypothesis regarding structures of small viruses
(TYMV & TMV) that:
Viral RNA enclosed in CP
Naked RNA is infectious
Basic requirement is protein shell to protect n/a etc.
In rod shaped viruses, the protein subunits are
arranged in a helical manner regardless of protein
subunit number into a helical array.

X-ray crystallography
X-ray crystallography is a method of determining the
arrangement of atoms within a crystal, in which a
beam of X-rays strikes a crystal and diffracts into
many specific directions.
From the angles and intensities of these diffracted
beams, a crystallographer can produce a three-
dimensional picture of the density of electrons within
the crystal.
From this electron density we can determined:
the mean positions of the atoms in the crystal, as well as
their chemical bonds,
their disorder and various other information.

X-ray sources
The brightest and most
useful X-ray sources
are synchrotrons
Workflow for solving the
structure of a molecule by X-
ray crystallography.
A protein crystal seen under amicroscope.
Crystals used in X-ray crystallography may
be smaller than a millimeter across.

Diffractometer A Diffractometer is a measuring
instrument for analyzing the structure of a
material from the scattering pattern
produced when a beam of radiation or
particles (as X rays or neutrons) interacts
with it.
Principle Because it is relatively easy to
use electrons or neutrons having wavelengths smalle
r than a nanometer, electrons and neutrons may be
used to study crystal structure in a manner very
similar to X-ray diffraction. Electrons do not penetrate
as deeply into matter as X-rays, hence electron
diffraction reveals structure near the surface;
neutrons do penetrate easily and have an advantage
that they possess an intrinsic magnetic moment that
causes them to interact differently with atoms having
different alignments of their magnetic moments.
An X-ray diffraction pattern of a
crystallized enzyme. The pattern of
spots (called reflections) can be used to
determine the structure of the enzyme.

TMV
TMV particles are:
Rigid helical rods
300 nm long X 18 nm dia
95% protein & ~5% n/a (RNA)
ssRNA
Extremely stable structure
Retain infectivity at room temp. for ~50 years
Naked RNA is highly unstable like others.

Detailed worked by using
X-ray defraction gave details of arrangement of protein subunits and RNA in rod.
The particles comprises ~2130 subunits that are closely packed in a helical array.
The pitch of helix is 2.3 (fig.) and the RNA chain is compactly coiled in a helix following that of the protein subunits
There are 49 nt. & 161/3 protein subunits per turn
The PO4 of the RNA are at about 4nm from the rod axis.
The helix of TMV is right handed (Finch, 1972)

TMV architecture
Negatively stained particles revealed that :
One end of the rod can be seen as concave
The other end is convex
3’end of the RNA is at the convex end & 5’ at
concave end (Wilson wt al. 1976; Butler et al.,
1977)
A central canal with a radius of ~2nm becomes
filled with stain in –vely stained preparations
Short Rods: of variable length & <300nm,
causes problem of end to end aggregation
etc.

SYMPTOMS OF TMV

Rod shaped particles
Helix (rod)
e.g., TMV
TMV rod is 18 nanometers (nm) X 300 nm

PARTICLE STRUCTURE
Tobacco mosaic virus is typical, well-studied example
Each particle contains only a single molecule of RNA (6395 nt) and 2130 copies of the coat protein subunit (158 aa; 17.3 kDa)
3 nt/subunit
16.33 subunits/turn
49 subunits/3 turns
TMV protein subunits + nucleic acid will self-assemble in vitro in an energy-independent fashion
Self-assembly also occurs in the absence of RNA
TMV rod is 18 nanometers (nm) X 300 nm

Tobacco mosaic virus

Properties of coat proteins
CP consists of 158 amino acid with a mol. Wt of ~17-18 KDa.
Fibre defraction have determined the structure to 2.0oA resolution (Namba et al., 1989)
The protein has high proportion of secondary structures with 50%of the residues form four a- helices and 10% of residues in B-turns.
The four closely parallel and antiparallel a- helices (residues 20-32, 38-48, 74-88 & 114-134) make up the core of the subunits.
And the distal end of the four helices are connected transversely by a narrow and twisted strip of b-sheet.

The central part of the subunit distal to the b-sheet is a cluster aromatic residues (Phe12, Trp17, Phe62, Tyr70, Tyr139, Phe144) forming a hydrophobic patch.
The N- & C- termini of the protein are to the outside of the particle
The polypeptide chain is in a flexible or disordered state below a radius in t particle of about 4nm so that no structure is revealed in this region.
Properties of coat proteins

One of the reassembly product of TMV
protein subunit is a double disk containing two
rings of 17 protein subunits and in this region
the details of the inter subunit contacts can be
determined (by X-ray crystallography) (Klug et
al.; Bloomer et al., 1978).
The subunits of the upper ring in the disk are
flat and in the lower ring are tilted down ward
toward the centre of the disk with three regions
of contact between the subunits.
Properties of coat proteins

Plant viruses are
diverse, but not as
diverse as animal
viruses – probably
because of size
constraints imposed by
requirement to move
cell-to-cell through
plasmodesmata of host
plants

Viral Morphological Groups
Cubic (icosahedral)
Helical
Horne, R. W. & Wildy, P.
(1961). Symmetry in virus
architecture. Virology 15,
348–373

Icosahedral arrangement is typical
in virus structure An icosahedron has 20
triangular (equilateral) faces (facets), 12 vertices, and a 5:3:2 axes of rotational symmetry

Isometric viruses
Icosahedron
(sphere) e.g., BMV

Tobacco necrosis virus, 26 nm in diameter

BROME MOSAIC VIRUS
• Type member of the
Bromovirus genus, family
Bromoviridae
• Virions are nonenveloped
icosohedrals (T=3), 26 nm in
diameter, contain 22% nucleic
acid and 78% protein
• BMV genome is composed
of three positive sense RNAs
separately encapsidated
RNA1 (3.2 kb), RNA2 (2.9
kb), RNA3 (2.1 kb), RNA4
(0.9 kb)
RNA1 RNA2 RNA3 RNA4

Francki, Milne & Hatta. 1985 Atlas of Plant Viruses, vol. I.
Three-dimensional image of Turnip yellow mosaic virus (TYMV)
reconstructed from EM

Tobacco mosaic virus
First virus crystallized (1946 Stanley was awarded the Nobel prize)
First demonstration of infectious RNA (1950s)
First virus to be shown to consist of RNA and protein
First virus characterized by X-ray crystallography to show a helical structure
First virus genome to be completely sequenced

Tobacco mosaic virus (TMV), 300 nm
Potato virus Y (PVY), 740 nm

Maize streak virus,
Geminiviridae
Cocoa swollen shoot virus,
Badnavirus


















