
Communication JOURNAL OF Vol. No. of 262, 22, Issue · PDF fileCommunication Vol. 262, No. 22,...
4
Communication Vol. 262, No. 22, Issue of August 5, pp. 10422-10425.1987 THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1987 by The American Society of Biological Chemists, Inc. Printed in U.S.A. Inactivation of Isocitrate Dehydrogenase by Phosphorylation Is Mediated by the Negative Charge of the Phosphate* (Received for publication, March 12, 1987) Peter E. Thorsness and Daniel E. Koshland, Jr.S From the Department of Biochemistry, University of California, Berkeley, California 94720 The control of isocitrate dehydrogenase through phosphorylation is necessary for growth of Esche- richia coli on acetate as the sole carbon source. To understand the mechanism by which phosphorylation inactivates isocitrate dehydrogenase, the sequence of icd, the isocitrate dehydrogenase structural gene, was determined and this information was used to construct mutants at the site of phosphorylation. Introduction of a negatively charged aspartate for the serine that is phosphorylated completely inactivates isocitrate de- hydrogenase. Substitution of the serine with other amino acids results in a partially active enzyme in which both maximal velocity and interaction with sub- strates has been altered. Neither threonine nor tyro- sine, when substituted for the serine at the phospho- rylation site, is detectably phosphorylatedby isocitrate dehydrogenase kinase. Protein phosphorylation has proven to be a common means by which the activity of a protein is regulated (1). Phospho- rylation of cell surface receptors and cytosolic proteins has been implicated in the control of cell growth and differentia- tion (2). The phosphorylation of structural proteins has been linked to the dynamic processes of cell shape determination (3). The regulation of major metabolic enzymes by protein phosphorylation has also been established (4). A well documented case of regulatory phosphorylation oc- curs in Escherichia coli during growth on acetate. When enteric bacteria use acetate as their sole carbon source, they must induce the enzymes of the glyoxylate bypass in order to grow. The phosphorylation of isocitrate dehydrogenase (5) controls the branchpoint between the glyoxylate bypass and the Krebs cycle and allows the effective operation of the glyoxylate bypass (6). The bypass is needed to accumulate carbon for biosynthetic purposes (7). The transfer of the y-phosphate from ATP to a serine residue on isocitrate dehydrogenase is particularly interesting because it causes complete inhibition of isocitrate dehydro- ~ * This work was supported by the National Institutes of Health and the National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solelyto indicate this fact. The nucleotide sequence(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) 502799. $ To whom correspondence should be addressed. genase activity (8). Inhibition of the enzyme activity by phos- phorylation is thought to act by decreasing the binding of the cofactor NADP (9). The covalent addition of a phosphate could cause this by several means. The serine that is phos- phorylated maybe necessary for binding of NADP or for catalysis. The phosphate could also mask other essential residues or sterically block a binding site. The conformation of the enzyme may be altered by the covalent addition of the bulky phosphate or negative charge. Using the cloned struc- tural gene for isocitrate dehydrogenase, icd, we have used site- directed mutagenesis to systematically alter the site of phos- phorylation. These experiments allow us to draw conclusions about the means by which the presence of a covalently linked phosphate affects the enzyme activity. MATERIALS AND METHODS Strains-The E. coli strain used for the generation of single- stranded DNA was DIH101, which harbors an F’ bearing the gene for kanamycin resistance. Uridine-labeled template was generated in the E. coli strain RZ1032. The analysis of icd-bearing plasmids was done in DEK2004, a trp, icd, recA strain. Growth and Harvesting of CeUs”DEK2004 containing various plasmids were grown overnight in LB; 10 g/liter bactotryptone (Difco), 5 g/liter yeast extract (Difco), and 10 g/liter NaCl. The cells were collected by centrifugation and resuspended in 25 mM MOPS,’ 1 mM EDTA, 1 mM dithiothreitol. The OD6, of the cell suspension was measured, and the cells were subjected to sonic disruption as described previously (6). The glutamate auxotrophy of bacteria was tested by streaking the LB culture on 1.5% Bacto Agar plates. The media was the MOPS based recipe of Neidhardt et al. (IO), supple- mented with the appropriate amino acids (0.5 mM) and glucose (0.5%). DNA Sequence Analysis-The sequence of the 1.9-kilobase insert in a pEMBL vector (11) (plasmids pTK512 and pTK513) was deter- mined for both strands of DNA by the dideoxy method of Sanger (12). Nested deletions of pTK512 and pTK513 were generated by the exo III/S1 method of Henikoff (13). Site-specific Mutagenesis-Site-specific mutants in the icd gene were generated using the uridine-labeled template method of Kunkel (14). Potential mutants were screened by DNA sequencing. Mutant alleles were isolated and sequenced to assure that only the intended base changes were present. Assays-The isocitrate dehydrogenase assays used to determine the kinetic constants of the various isocitrate dehydrogenase species were 1 ml in volume and done in 25 mM MOPS, pH 7.5,5 mM MgC12, 100 mM NaCl, 1 mM dithiothreitol, 1 mM EDTA, and concentrations of dl-isocitrate ranging from 5 PM to 1 mM. The assays were performed at 37 ‘C, and the reduction of NADP was monitored at 340 nm. All other isocitrate dehydrogenase assays were done as described previ- ously (6). Protein was determined by the method of Lowry et al. (15). Phosphorylation of Zsocitrate Dehydrogenase-Zn vitro phosphoryl- ation of isocitrate dehydrogenase was done using purified isocitrate dehydrogenase kinase (16). 40 p1 of cellular extracts containing equiv- alent amounts of isocitrate dehydrogenase protein were incubated at 37 ‘C with 7.5 pCi of [y-32P]ATP, 0.1 pM ATP, 25 mM MOPS, 1 mM EDTA, 1 mM dithiothreitol, 5 mM MgClz, 100 mM NaCl, and 2 milliunits of isocitrate dehydrogenase kinase. When the activity of isocitrate dehydrogenase was to be monitored after the protein had been exposed to isocitrate dehydrogenase kinase and ATP, the radio- active ATP was replaced with unlabeled ATP at a concentration of 10 mM. The in vivo phosphorylation of isocitrate dehydrogenase was ana- lyzed in DEK2004 containing either wild type or mutant plasmids. The cells were grown overnight in LB containing 250 pg/ml ampicillin The abbreviations used are: MOPS, 3-(N-morpholino)propane- sulfonic acid; ODsso,optical density of a solution with a 1-cm path length at 650 nm; SDS, sodium dodecyl sulfate. 10422
Transcript of Communication JOURNAL OF Vol. No. of 262, 22, Issue · PDF fileCommunication Vol. 262, No. 22,...

Communication Vol. 262, No. 22, Issue of August 5, pp. 10422-10425.1987 THE JOURNAL OF BIOLOGICAL CHEMISTRY
0 1987 by The American Society of Biological Chemists, Inc. Printed in U.S.A.
Inactivation of Isocitrate Dehydrogenase by Phosphorylation Is Mediated by the Negative Charge of the Phosphate*
(Received for publication, March 12, 1987) Peter E. Thorsness and Daniel E. Koshland, Jr.S From the Department of Biochemistry, University of California, Berkeley, California 94720
The control of isocitrate dehydrogenase through phosphorylation is necessary for growth of Esche- richia coli on acetate as the sole carbon source. To understand the mechanism by which phosphorylation inactivates isocitrate dehydrogenase, the sequence of icd, the isocitrate dehydrogenase structural gene, was determined and this information was used to construct mutants at the site of phosphorylation. Introduction of a negatively charged aspartate for the serine that is phosphorylated completely inactivates isocitrate de- hydrogenase. Substitution of the serine with other amino acids results in a partially active enzyme in which both maximal velocity and interaction with sub- strates has been altered. Neither threonine nor tyro- sine, when substituted for the serine at the phospho- rylation site, is detectably phosphorylated by isocitrate dehydrogenase kinase.
Protein phosphorylation has proven to be a common means by which the activity of a protein is regulated (1). Phospho- rylation of cell surface receptors and cytosolic proteins has been implicated in the control of cell growth and differentia- tion (2). The phosphorylation of structural proteins has been linked to the dynamic processes of cell shape determination (3). The regulation of major metabolic enzymes by protein phosphorylation has also been established (4).
A well documented case of regulatory phosphorylation oc- curs in Escherichia coli during growth on acetate. When enteric bacteria use acetate as their sole carbon source, they must induce the enzymes of the glyoxylate bypass in order to grow. The phosphorylation of isocitrate dehydrogenase ( 5 ) controls the branchpoint between the glyoxylate bypass and the Krebs cycle and allows the effective operation of the glyoxylate bypass (6). The bypass is needed to accumulate carbon for biosynthetic purposes (7).
The transfer of the y-phosphate from ATP to a serine residue on isocitrate dehydrogenase is particularly interesting because it causes complete inhibition of isocitrate dehydro- ~
* This work was supported by the National Institutes of Health and the National Science Foundation. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) 502799.
$ To whom correspondence should be addressed.
genase activity (8). Inhibition of the enzyme activity by phos- phorylation is thought to act by decreasing the binding of the cofactor NADP (9). The covalent addition of a phosphate could cause this by several means. The serine that is phos- phorylated may be necessary for binding of NADP or for catalysis. The phosphate could also mask other essential residues or sterically block a binding site. The conformation of the enzyme may be altered by the covalent addition of the bulky phosphate or negative charge. Using the cloned struc- tural gene for isocitrate dehydrogenase, icd, we have used site- directed mutagenesis to systematically alter the site of phos- phorylation. These experiments allow us to draw conclusions about the means by which the presence of a covalently linked phosphate affects the enzyme activity.
MATERIALS AND METHODS
Strains-The E. coli strain used for the generation of single- stranded DNA was DIH101, which harbors an F’ bearing the gene for kanamycin resistance. Uridine-labeled template was generated in the E. coli strain RZ1032. The analysis of icd-bearing plasmids was done in DEK2004, a trp, icd, recA strain.
Growth and Harvesting of CeUs”DEK2004 containing various plasmids were grown overnight in LB; 10 g/liter bactotryptone (Difco), 5 g/liter yeast extract (Difco), and 10 g/liter NaCl. The cells were collected by centrifugation and resuspended in 25 mM MOPS,’ 1 mM EDTA, 1 mM dithiothreitol. The OD6, of the cell suspension was measured, and the cells were subjected to sonic disruption as described previously (6). The glutamate auxotrophy of bacteria was tested by streaking the LB culture on 1.5% Bacto Agar plates. The media was the MOPS based recipe of Neidhardt et al. (IO), supple- mented with the appropriate amino acids (0.5 mM) and glucose (0.5%).
DNA Sequence Analysis-The sequence of the 1.9-kilobase insert in a pEMBL vector (11) (plasmids pTK512 and pTK513) was deter- mined for both strands of DNA by the dideoxy method of Sanger (12). Nested deletions of pTK512 and pTK513 were generated by the exo III/S1 method of Henikoff (13).
Site-specific Mutagenesis-Site-specific mutants in the icd gene were generated using the uridine-labeled template method of Kunkel (14). Potential mutants were screened by DNA sequencing. Mutant alleles were isolated and sequenced to assure that only the intended base changes were present.
Assays-The isocitrate dehydrogenase assays used to determine the kinetic constants of the various isocitrate dehydrogenase species were 1 ml in volume and done in 25 mM MOPS, pH 7.5,5 mM MgC12, 100 mM NaCl, 1 mM dithiothreitol, 1 mM EDTA, and concentrations of dl-isocitrate ranging from 5 PM to 1 mM. The assays were performed at 37 ‘C, and the reduction of NADP was monitored at 340 nm. All other isocitrate dehydrogenase assays were done as described previ- ously (6). Protein was determined by the method of Lowry et al. (15).
Phosphorylation of Zsocitrate Dehydrogenase-Zn vitro phosphoryl- ation of isocitrate dehydrogenase was done using purified isocitrate dehydrogenase kinase (16). 40 p1 of cellular extracts containing equiv- alent amounts of isocitrate dehydrogenase protein were incubated at 37 ‘C with 7.5 pCi of [y-32P]ATP, 0.1 pM ATP, 25 mM MOPS, 1 mM EDTA, 1 mM dithiothreitol, 5 mM MgClz, 100 mM NaCl, and 2 milliunits of isocitrate dehydrogenase kinase. When the activity of isocitrate dehydrogenase was to be monitored after the protein had been exposed to isocitrate dehydrogenase kinase and ATP, the radio- active ATP was replaced with unlabeled ATP at a concentration of 10 mM.
The in vivo phosphorylation of isocitrate dehydrogenase was ana- lyzed in DEK2004 containing either wild type or mutant plasmids. The cells were grown overnight in LB containing 250 pg/ml ampicillin
The abbreviations used are: MOPS, 3-(N-morpholino)propane- sulfonic acid; ODsso, optical density of a solution with a 1-cm path length at 650 nm; SDS, sodium dodecyl sulfate.
10422

Mechanism of Inactivation of Isocitrate Dehydrogenase by Phosphorylation 10423
FIG. 1. Nucleotide sequence of the icd open reading frame and pre- dicted amino acid sequence of isocit- rate dehydrogenase. The 14 amino acid residues previously identified as the phosphorylated peptide obtained from proteolytic digestion of phosphorylated isocitrate dehydrogenase (17) are under- lined. The phosphorylated serine is boxed.
and resuspended at an OD, of 0.5 in 1 ml of phosphate-free, 2% sodium acetate, MOPS-based media (10). The cells were incubated with 80 pCi of carrier-free 32P04 for 20 min at 37 "C. The cells were pelleted, resuspended in SDS-polyacrylamide gel electrophoresis sam- ple buffer, subjected to SDS-polyacrylamide gel electrophoresis, elec- troblotted to nitrocellulose, and exposed at -70 "C to x-ray film for 4 h with an intensifying screen. Electroblotting the samples was found to reduce the radioactive background that occurs when whole cells labeled with 32P0, are subjected to SDS-polyacrylamide gel electrophoresis and autoradiography.
RESULTS
The complete DNA sequence of icd was determined by dideoxy sequencing both strands of a 1.9-kilobase fragment carrying the gene. The plasmid containing this fragment was called pTK512 and was derived from a clone previously iso- lated from the Clarke-Carbon plasmid bank (6). The plasmid produces a functional and full sized isocitrate dehydrogenase protein as determined by activity assays and SDS-polyacryl- amide gel electrophoresis. The nucleotide and inferred amino acid sequences obtained from this clone and related deletion constructs are shown in Fig. 1.
The 416-amino acid protein encoded by icd has a calculated molecular mass of 45764 g/mol. The region from residues 104 through 117 comprises the amino acid sequence of a peptide reported by Borthwick et al. (17) which contains the phos- phorylated serine residue.
E. coli isocitrate dehydrogenase does not have any signifi- cant sequence homology with other dehydrogenases or nu- cleotide binding proteins. Using the FASTP algorithm with ktup = 1 (18) for a homology search of the proteins in the Dayhoff data base, isocitrate dehydrogenase shows the most homology to 3-isopropylmalate dehydrogenase from Thermus aquaticus (19). In a 94-amino acid overlap occurring in the C- terminal portion of both proteins (residues 274 through 368 for isocitrate dehydrogenase) 35% of the amino acid residues are identical and 44% are conservative changes. This is more than twice the homology shown to any other protein in the
data base. The lack of homology and absence of specific sequence patterns characteristic of nucleotide binding pro- teins does not allow the assignment of a nucleotide binding domain to any segment of the primary sequence.
Synthetic oligonucleotide-directed mutagenesis was used to replace the serine a t residue 113 with a threonine, tyrosine, cysteine, alanine, or aspartate. Extracts made from cells con- taining these altered isocitrate dehydrogenase proteins were subjected to SDS-polyacrylamide gel electrophoresis and stained with Coomassie Blue (Fig. 2). Isocitrate dehydrogen- ase protein in all of these extracts was on the order of 50% of the total protein present in the cell. The amount of isocitrate dehydrogenase protein was constant both with cell number (optical density) and protein as judged by intensity of staining of SDS-polyacrylamide gels with Coomassie Blue. The amount of isocitrate dehydrogenase protein in a given assay was normalized to the optical density of the cells at harvest or to the protein concentration.
lane a b C d e f g h
FIG. 2. SDS-polyacrylamide gel electrophoresis of E. coli cells containing plasmids bearing wild type and mutant icd genes. Lane a, vector; lane b, Thr-113 mutant; lane c, Tyr-113 mutant; lane d, Cys-113 mutant; lane e, Ala-113 mutant; l a n e / , Asp- 113 mutant; lane g, Ser-113 wild type; lane h, purified isocitrate dehydrogenase.
10424 Mechanism of Inactivation of Isocitrate Dehydrogenase by Phosphorylation TABLE I
Kinetic constants of rwrmal and altered isocitrate dehydrogenase proteins
Residue 113 VIM=a NADP iswitrate K,
lane a b c d e f g " - -
f l M f l M
Serine 100 7 8 Threonine 66 68 26 Tyrosine 12 72 124 Cysteine 8 65 71 Alanine 40 31 18 Aspartate Ob
"The specific activity of the wild type isocitrate dehydrogenase extract has been set at 100 and the mutant proteins normalized to that value.
Background levels of iswitrate dehydrogenase activity were de- termined in extracts that had been grown with pEMBL vector as the control plasmid. This activity had a Vma., of 0.2. The Vmnx of the Asp- 113 mutant never exceeded that of the vector alone extracts and therefore was given a Vmax of 0.
A
lane a b c d e l g h
c
B
I . . . . . . . . . . . I 0 0 0 7 0 4 0 6 O B 1'0 1'2
. . .
F R ~ I I O ~ 1% ~CIIVIIY rernmnp an.. tnatnunt wnn IDH ma..
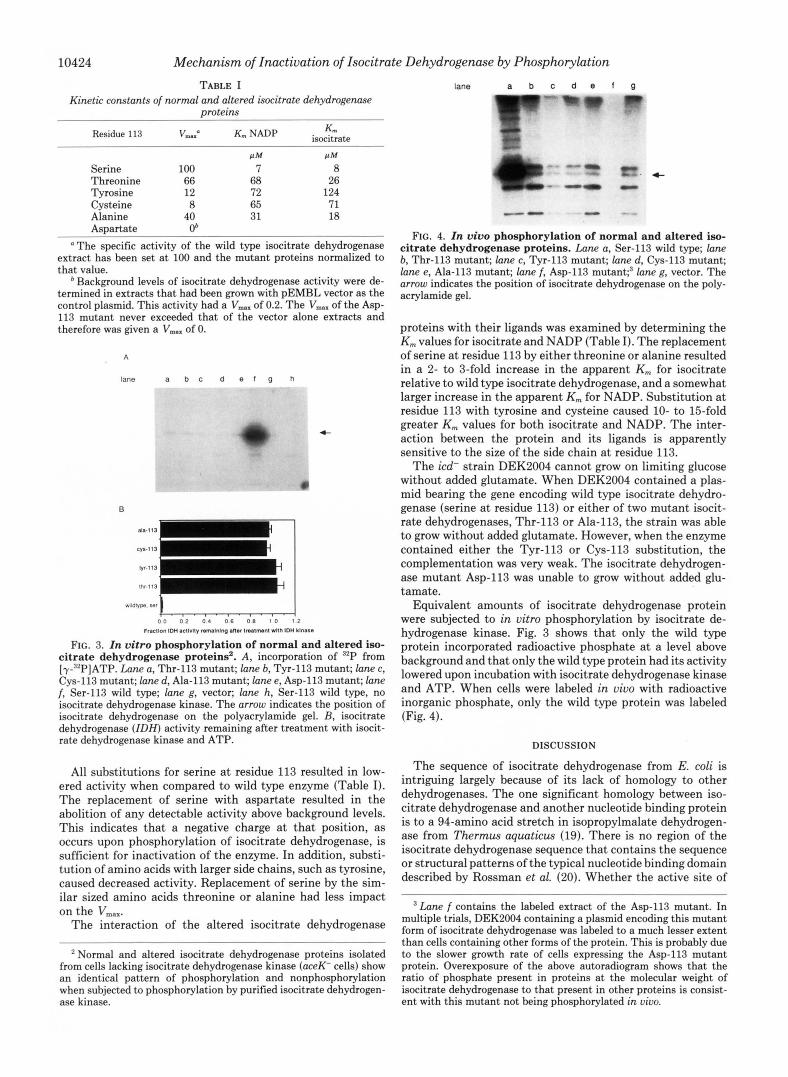
FIG. 3. In vitro phosphorylation of normal and altered iso- citrate dehydrogenase proteins'. A, incorporation of '*P from [y-'*P]ATP. Lane a, Thr-113 mutant; lane b, Tyr-113 mutant; lane c, Cys-113 mutant; lane d, Ala-113 mutant; lane e, Asp-113 mutant; lane f, Ser-113 wild type; lane g, vector; lane h, Ser-113 wild type, no iswitrate dehydrogenase kinase. The arrow indicates the position of isocitrate dehydrogenase on the polyacrylamide gel. B, isocitrate dehydrogenase ( I D H ) activity remaining after treatment with isocit- rate dehydrogenase kinase and ATP.
All substitutions for serine a t residue 113 resulted in low- ered activity when compared to wild type enzyme (Table I). The replacement of serine with aspartate resulted in the abolition of any detectable activity above background levels. This indicates that a negative charge at that position, as occurs upon phosphorylation of isocitrate dehydrogenase, is sufficient for inactivation of the enzyme. In addition, substi- tution of amino acids with larger side chains, such as tyrosine, caused decreased activity. Replacement of serine by the sim- ilar sized amino acids threonine or alanine had less impact on the VmaX.
The interaction of the altered isocitrate dehydrogenase
Normal and altered isocitrate dehydrogenase proteins isolated from cells lacking isocitrate dehydrogenase kinase (aceK- cells) show an identical pattern of phosphorylation and nonphosphorylation when subjected to phosphorylation by purified isocitrate dehydrogen- ase kinase.
4-
FIG. 4. In vivo phosphorylation of normal and altered iso- citrate dehydrogenase proteins. Lane a, Ser-113 wild type; lane b, Thr-113 mutant; lane c, Tyr-113 mutant; lane d, Cys-113 mutant; lane e, Ala-113 mutant; lane f, Asp-113 r n ~ t a n t ; ~ lane g, vector. The arrow indicates the position of isocitrate dehydrogenase on the poly- acrylamide gel.
proteins with their ligands was examined by determining the K,,, values for isocitrate and NADP (Table I). The replacement of serine at residue 113 by either threonine or alanine resulted in a 2- to 3-fold increase in the apparent K,,, for isocitrate relative to wild type isocitrate dehydrogenase, and a somewhat larger increase in the apparent K,,, for NADP. Substitution at residue 113 with tyrosine and cysteine caused 10- to 15-fold greater K,,, values for both isocitrate and NADP. The inter- action between the protein and its ligands is apparently sensitive to the size of the side chain at residue 113.
The icd- strain DEK2004 cannot grow on limiting glucose without added glutamate. When DEK2004 contained a plas- mid bearing the gene encoding wild type isocitrate dehydro- genase (serine a t residue 113) or either of two mutant isocit- rate dehydrogenases, Thr-113 or Ala-113, the strain was able to grow without added glutamate. However, when the enzyme contained either the Tyr-113 or Cys-113 substitution, the complementation was very weak. The isocitrate dehydrogen- ase mutant Asp-113 was unable to grow without added glu- tamate.
Equivalent amounts of isocitrate dehydrogenase protein were subjected to in vitro phosphorylation by isocitrate de- hydrogenase kinase. Fig. 3 shows that only the wild type protein incorporated radioactive phosphate a t a level above background and that only the wild type protein had its activity lowered upon incubation with isocitrate dehydrogenase kinase and ATP. When cells were labeled in uiuo with radioactive inorganic phosphate, only the wild type protein was labeled (Fig. 4).
DISCUSSION
The sequence of isocitrate dehydrogenase from E. coli is intriguing largely because of its lack of homology to other dehydrogenases. The one significant homology between iso- citrate dehydrogenase and another nucleotide binding protein is to a 94-amino acid stretch in isopropylmalate dehydrogen- ase from Thermus aquaticus (19). There is no region of the isocitrate dehydrogenase sequence that contains the sequence or structural patterns of the typical nucleotide binding domain described by Rossman et al. (20). Whether the active site of
Lane f contains the labeled extract of the Asp-113 mutant. In multiple trials, DEK2004 containing a plasmid encoding this mutant form of isocitrate dehydrogenase was labeled to a much lesser extent than cells containing other forms of the protein. This is probably due to the slower growth rate of cells expressing the Asp-I13 mutant protein. Overexposure of the above autoradiogram shows that the ratio of phosphate present in proteins at the molecular weight of isocitrate dehydrogenase to that present in other proteins is consist- ent with this mutant not being phosphorylated in uiuo.

Mechanism of Inactivation of Isocitrate Dehydrogenase by Phosphorylation 10425
isocitrate dehydrogenase will actually be similar to other dehydrogenases in three-dimensional space is thus an inter- esting question.
The phosphorylation site of isocitrate dehydrogenase is unremarkable in terms of the amino acid sequence immedi- ately surrounding serine 113. Like the substrate phosphoryl- ation sites examined for CAMP-dependent protein kinase or phosphorylase kinase (21, 22), the phosphorylated serine in isocitrate dehydrogenase is flanked by a basic residue, arginine 112. Elucidation of the sequence specificity of isocitrate de- hydrogenase kinase will require studies such as those which used synthetic peptides and natural variants of substrates to examine the specificity of other protein kinases (21, 23, 24). Clearly, the serine at residue 113 is a necessary element for recognition by isocitrate dehydrogenase kinase, as neither the threonine 113 nor tyrosine 113 mutants were phosphorylated (Figs. 3 and 4). Presumably, other amino acids in the primary sequence about serine 113 are also critical for the interaction with isocitrate dehydrogenase kinase. As with other kinases, a complete understanding of the substrate requirements will require knowledge of the tertiary structure of the protein substrate.
The complete inhibition of isocitrate dehydrogenase activ- ity by phosphorylation is more dramatic than the effect phos- phorylation has upon either muscle phosphofructokinase or glycogen phosphorylase. Phosphorylation of phosphofructo- kinase increases sensitivity to ATP inhibition by allowing increased binding of ATP to the inhibitory site of the enzyme (25). For glycogen phosphorylase, possibly the most thor- oughly studied case of regulation by covalent modification, phosphorylation of the enzyme shifts the equilibrium of the protein conformation to the active form (26). This form of phosphorylase, phosphorylase a, is less sensitive to inactiva- tion by changing effector concentrations. The effect of phos- phorylation on the activity of these three enzymes is different, but the result of the process is the same: control of metabolism in response to the needs of the cell.
The effect phosphorylation has on the activity of a protein is often immediately obvious. What is not always clear is the means by which the addition of a phosphate achieves that result. Because the icd gene is cloned, unlike most of the mammalian genes encoding enzymes regulated by phospho- rylation, a systematic modification of the regulatory site was possible. The data in Table I demonstrate the importance of residue 113 to the structure and function of isocitrate dehy- drogenase. Since the alanine 113 mutant retains activity, the serine side chain at that site cannot be required as a catalytic entity in the actual chemistry of the enzymatic reaction. However, because isocitrate dehydrogenase activity is sensi- tive to substitution at residue 113, the serine at that site in
the wild type protein must be necessary for maintaining the proper structure at the active site. The finding that aspartate at residue 113, which has a negative charge but lacks the shape of phosphoserine, causes complete inactivation, whereas other changes merely lead to some loss of substrate binding or reduction of catalytic activity, suggests that the negative charge of the phosphate is the cause of the complete inactivation.
REFERENCES 1. Krebs, E. G. (1986) in The Enzymes (Boyer, P. D., ed) Vol. 17,
pp. 3-20, Academic Press, New York 2. Rozengurt, E. (1986) Science 2 3 4 , 161-166 3. Gard, D. L., and Kirschner, M. W. (1985) J. Cell Biol. 100 , 769-
4. Linn, T. C., Pettit, F. H., and Reed, L. J. (1969) Proc. Natl. Acad.
5. Garnak, M., and Reeves, H. C. (1979) Science 203, 1111-1112 6. LaPorte, D. C., Thorsness, P. E., and Koshland, D. E., Jr. (1985)
7. Kornberg, H. L. (1966) Biochem. J. 99 , l -11 8. LaPorte, D. C., and Koshland, D. E., Jr. (1983) Nature 305 ,
9. Garland, D., and Nimmo, H. G. (1984) FEBS Lett. 165,259-264 10. Neidhardt, F. C., Bloch, P. L., and Smith, D. F. (1974) J. Bucteriol.
11. Dente, L., Cesareni, G., and Cortese, R. (1983) Nucleic Acids Res.
12. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.
13. Henikoff, S. (1984) Gene (Amst.) 2 8 , 351-359 14. Kunkel, T. A. (1985) Proc. Nutl. Acad. Sci. U. S. A. 82,488-492 15. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
16. LaPorte, D. C., and Koshland, D. E., Jr. (1982) Nature 300,
17. Borthwick, A. C., Holms, W. H., and Nimmo, H. G. (1984) FEBS
18. Lipman, D. J., and Pearson, W. R. (1985) Science 227, 1435- 1441
19. Kagawa, Y., Nojima, H., Nukiwa, N., Ishizuka, M., Nakajima, T., Yasuhara, T., Tanaka, T., and Oshima, T. (1984) J. Biol. Chem.
20. Rossman, M. G., Moras, D., and Olsen, K. W. (1974) Nature
21. Kemp, B. E., Bylund, D. B., Huang, T.-S., and Krebs, E. G. (1975)
22. Nolan, C., Novoa, W. B., Krebs, E. G., and Fischer, E. H. (1964)
23. Kemp, B. E., Graves, D. J., Benjamini, E., and Krebs, E. G. (1977)
24. Chan, K.-F. J., Hurst, M. O., and Graves, D. J. (1982) J. Biol.
774
Sci. U. S. A. 62,234-241
J. Bwl. Chem. 260 , 10563-10568
286-290
119,736-747
11,1645-1655
Acad. Sci. U. S. A. 74, 5463-5467
(1951) J. Biol. Chem. 193,265-275
458-460
Lett. 174, 112-115
259,2956-2960
250,194-199
Proc. Nutl. Acud. Sci. U. S. A. 72 , 3448-3452
BiOChemktYy 3,542-551
J. Biol. Chem. 252,4888-4894
Chem. 257,3655-3659 25. Kitajima, S., Sakakibara, R., and Uyeda, K. (1983) J. Biol. Chem.
258, 13292-13298 26. Fletterick, R. J., and Sprang, S. R. (1982) Accts. Chem. Res. 15,
361-369


















