
Common principles in protein folding and antigen presentation
5
238 reviews 26 Nencioni, L., Vo~Fiai, G., Peppoloni, S., De Magistris, M. T., Marsili, i. and Rappuoli, R. (1991) hzfect, hnmun. 59, 625-630 27 Sekura, R. R. D., Zhang, Y., Roberson, R., Acton, D., Trollfors, B., Tolson, N., Shiloach, J., Bryla, D., Muir-Nash, J., Koeller, D., Scheerson, R. and Robbins, J. B. (1988) J. Pediatr. 113, 806-813 28 Winberry, L., Walker, R., Cohen, N., Todd, C., Sentissi, A., Siber, G. (1988)Abstracts ofhzternational Workshop of B. pertussis (Keith, J., ed.), Hamilton Rocky Mountains Laboratory 29 Edwards, K. M., Bradley, R. B., Decker, M. D., Palmer, S. P., Van Saraga, T., Taylor, J. C., Dupont, W. D., Hager, C. C. and Wright, P. F. (1989).]. hzfect. Dis. 160, 832-837 30 Rutter, D. A., Ashworth, L. A. E., Day, A., Funnel, S., Lovell, F. and Robinson, A. (1987) Vaccine 6, 29-32 Common principles in protein folding and antigen presentation Shan Lu, Victor E. Reyes, Rochelle R. Torgerson, Robert A. Lew and Robert E. Humphreys The regular recurrence of hydrophobic amino acid residues along a peptide sequence determines the formation of a longitudinal hydrophobic strip when the peptide forms an o~-helix. An understanding of the ways this may affect both folding of nascent proteins and antigen presentation should facilitate vaccine and therapeutics design. Formation of an 0c-helix in a protein situated against a hydrophobic surface is determined by the existence of hydrophobic amino acids recur- ring regularly at specific positions along the peptide chain. These residues form a longitudinal hydrophobic strip when the sequence is coiled as an 0~-helix. This principle appears to govern both formation of helices in folding intenaediates of nascent proteins and scavenging of T-cell-- presented epitopes in proteolytic fiagments of antigenic protein. The mechanisms which regu- late digestion of antigenic proteins into specific fragments, and selection of only some fragments for transfer to major histocompatibility complex (MHC) molecules are unclear. We hypothesize that consensus sequences (in or around the sequence which actually fits into the antigen- binding site of MHC molecules) are involved. If such consensus sequences are found, an under- standing of the biophysical or chemical mechan- isms by which they act will permit rational S. Lu, V. E. Reyes, R. R. Torgerson, R. A. Lew and R. E. Humphreys are at the Department of Pharmacology, University of Massachusetts Medical School, 55 Lake Avenue North, Worcester, MA 01655, USA. engineering of such sequences. One might exploit this idea to engineer potency and range of MHC restriction of vaccines, or to alter immunogenicity of therapeutic proteins. Helical folding of antigenic peptides DeLisi and Berzofsky noted that many T-cell- presented peptides are amphipathic helices 1. Kaiser and K~zdy had observed that amphiphilic peptides can coil on contact with lipid mem- branes 2. The progressive anchoring of hydro- phobic side chains into a lipid bilayer at successive turns of a peptide coiling into a helix might co-operate with hydrogen bonding along the pepddyl backbone to stabilize the growing helix (Fig. 1). Such membrane adsorption could be a step in transferring peptides to MHC molecules3. Such recurrent hydrophobic residues would form a longitudinal hydrophobic strip along the helix4-7. During antigen processing, binding of such pep- tides to the wall of a vesicle could lead to selective transfer of some digested peptides to a second compartment after expulsior~ of soluble contents to a lysosome-bound vesicles;9. Alternatively, the hydrophobic strip of such an adsorbed helix could promote its binding to the hydrophobic floor of either a scavenging/transfer molecule or a MHC TIBTECH JULY 1991 (VOL 9) ~) 1991, ElsevierScience Publist~ers Ltd (U;q 0167-9430/91/$2,00
Transcript of Common principles in protein folding and antigen presentation

238
reviews
26 Nencioni, L., Vo~Fiai, G., Peppoloni, S., De Magistris, M. T., Marsili, i. and Rappuoli, R. (1991) hzfect, hnmun. 59, 625-630
27 Sekura, R. R. D., Zhang, Y., Roberson, R., Acton, D., Trollfors, B., Tolson, N., Shiloach, J., Bryla, D., Muir-Nash, J., Koeller, D., Scheerson, R. and Robbins, J. B. (1988) J. Pediatr. 113, 806-813
28 Winberry, L., Walker, R., Cohen, N., Todd, C., Sentissi, A., Siber, G. (1988)Abstracts ofhzternational Workshop of B. pertussis (Keith, J., ed.), Hamilton Rocky Mountains Laboratory
29 Edwards, K. M., Bradley, R. B., Decker, M. D., Palmer, S. P., Van Saraga, T., Taylor, J. C., Dupont, W. D., Hager, C. C. and Wright, P. F. (1989).]. hzfect. Dis. 160, 832-837
30 Rutter, D. A., Ashworth, L. A. E., Day, A., Funnel, S., Lovell, F. and Robinson, A. (1987) Vaccine 6, 29-32
Common principles in protein folding and antigen presentation
Shan Lu, Victor E. Reyes, Rochelle R. Torgerson, Robert A. Lew and Robert E. Humphreys
The regular recurrence of hydrophobic amino acid residues along a peptide
sequence determines the formation of a longitudinal hydrophobic strip when
the peptide forms an o~-helix. An understanding of the ways this may affect both
folding of nascent proteins and antigen presentation should facilitate vaccine
and therapeutics design.
Formation of an 0c-helix in a protein situated against a hydrophobic surface is determined by the existence of hydrophobic amino acids recur- ring regularly at specific positions along the peptide chain. These residues form a longitudinal hydrophobic strip when the sequence is coiled as an 0~-helix. This principle appears to govern both formation of helices in folding intenaediates of nascent proteins and scavenging of T-cell-- presented epitopes in proteolytic fiagments of antigenic protein. The mechanisms which regu- late digestion of antigenic proteins into specific fragments, and selection of only some fragments for transfer to major histocompatibility complex (MHC) molecules are unclear. We hypothesize that consensus sequences (in or around the sequence which actually fits into the antigen- binding site of MHC molecules) are involved. If such consensus sequences are found, an under- standing of the biophysical or chemical mechan- isms by which they act will permit rational
S. Lu, V. E. Reyes, R. R. Torgerson, R. A. Lew and R. E. Humphreys are at the Department of Pharmacology, University of Massachusetts Medical School, 55 Lake Avenue North, Worcester, MA 01655, USA.
engineering of such sequences. One might exploit this idea to engineer potency and range of MHC restriction of vaccines, or to alter immunogenicity of therapeutic proteins.
Helical folding of antigenic peptides DeLisi and Berzofsky noted that many T-cell-
presented peptides are amphipathic helices 1. Kaiser and K~zdy had observed that amphiphilic peptides can coil on contact with lipid mem- branes 2. The progressive anchoring of hydro- phobic side chains into a lipid bilayer at successive turns of a peptide coiling into a helix might co-operate with hydrogen bonding along the pepddyl backbone to stabilize the growing helix (Fig. 1). Such membrane adsorption could be a step in transferring peptides to MHC molecules 3. Such recurrent hydrophobic residues would form a longitudinal hydrophobic strip along the helix 4-7. During antigen processing, binding of such pep- tides to the wall of a vesicle could lead to selective transfer of some digested peptides to a second compartment after expulsior~ of soluble contents to a lysosome-bound vesicle s;9. Alternatively, the hydrophobic strip of such an adsorbed helix could promote its binding to the hydrophobic floor of either a scavenging/transfer molecule or a MHC
TIBTECH JULY 1991 (VOL 9) ~) 1991, Elsevier Science Publist~ers Ltd (U;q 0167-9430/91/$2,00

239
reviews
L Y Q E L Q K L T Q T L K
1 " " " 1 " " 1 " " " i ¢
~ d membrane
II
IT llll lll
figure 1 Selection of T-cell-presented epitopes. Helical folding on a lipid membrane of a peptide with recurrent hydrophobic residues (indicated by bold bars). These hydrophobic residues merge into a hydrophobic surface and stabilize hydrogen bonding between adjacent coils of an oc- helix. Such structures may be relatively protease resistant, physically scavenged as the contents of an endosome are extruded to a lysosome, and transferred by special proteins to antigen binding sites on MHC molecules.
molecule's antigen-binding site. Either as a helical configuration per se or within a protective struc- ture, the peptide might be relatively resistant to further proteolytic attack. The potential amphi- philic helical configuration of T-cell-presented peptides might, thus, reflect selection at a scaveng- ing/transfer step in antigen processing rather than coiling as a helix within the desetope (antigen binding site) itself.
Helical nucleation in nascent proteins Helical folding of some segments of a growing
protein might occur against a lipid bilayer or within unfoldase enzymes which also catalyse interchange of amphiphilic helices until the most stable conformation is found. Such helices might bind to each other through their longitudinal hydrophobic strips or to hydrophobic regions of a nascent protein 6. This hypothesis is supported by studies using cassette mutagenesis along one seg- ment of the phage lambda repressor to determine which amino acids substituted at each position could be tolerated in the folding of the repressor without loss of function 1°. In the sequence (resi- dues 84--91):
I le-Tyr--Gh-Met-Tyr-Gh-Ala-Val
Box 1. Presentation of antigen epitopes
Potential therapeutics and vaccine design
Two structurally distinct types of antigenic epitopes exist in an immunizing protein. Superficial, secondary-structure-dependent epitopes are recog- nized by antibodies. Other epitopes are recognized by T cells even after denaturation or in peptides cleaved from the antigen. To boost antibody production, an antigen-presenting B cell binds and internalizes the antigen recognized by its surface-expressed immunoglobulin. Within endosomes in the cell, the antigen is digested into fragments, some of which are transferred to class II major histocompatibility complex (MHC) molecules to become surface-expressed as peptide fragments. T cells recognize such peptides only in association with certain alleles of class I or II MHC molecules and, thus, presentation of the epitope is said to be MHC- restricted. T cells recognizing MHC-presented peptides stimulate the B cell both directly and indirectly, through lymphokines, to mature and proliferate into antibody-producing plasma cells. Peptides from such T-cell-presented sites might be used as vaccines. Such T-cell-presented peptides could be conjugated to other peptides from the immunoglobulin-recognized sites to make bivalent vaccines to boost neutralizing antibody to the 'B' epitope. T epitopes in proteins of infectious organisms may also trigger autoimmune diseases if they mimic segments of self proteins. Finally, such T epitopes in therapeutic or diagnostic proteins (e.g. enzymes, routine antibodies, lympbokines) can augment patient antibodies which would preclude further use of those products.
I1e84 and Met87 could be substituted only with aliphatic hydrophobic residues (lie, Val, Met), and Va191 could be substituted only with these same residues or Ala, Cys, Ser, Thr. Each of the other positions accepted a wide range of amino acids, in some cases either positively or negatively charged residues were tolerated, with some degree of function being retained. The recurrent, aliphatic hydrophobic residues I1e84, Met87 and Va191, represented in bold type in this sequence, formed a longitudinal hydrophobic strip, if the sequence were coiled as a helix. In fact, in the crystallographic structure it, this segment did form an 0~-hehx with the hydrophobic strip pointing inwards and the more-freely mutable residues facing outwards, towards the solvent.
The strip-of-helix hydrophobicity index ( s o H I - n )
In order to rank and determine lengths of segments with longitudinal hydrophobic strips in a helical configuration, we calculate the SOHHI. The SOHHI for each position n in a protein is the mean hydrophobicity of amino acids at positions n, n+4, n+7, n+11, n+14, etc. for a chosen number of turns (usually 3 to 6), in a putative 0c-helix extending toward the C-terminus from position n. The SOHHI provides a method of quantifying the mean hydrophobicity of residues in a longitudinal strip along that helix. This index is incorporated in algorithms used to predict (1) T-cell-presented epitopes (the strip-of-helix hydrophobicity algorithm, SOHHA) 4's'7, and (2) structural helices in proteins (structural helices algorithm, SHA) 6.
TIBTECH JUlY 1991 (VOL 9)
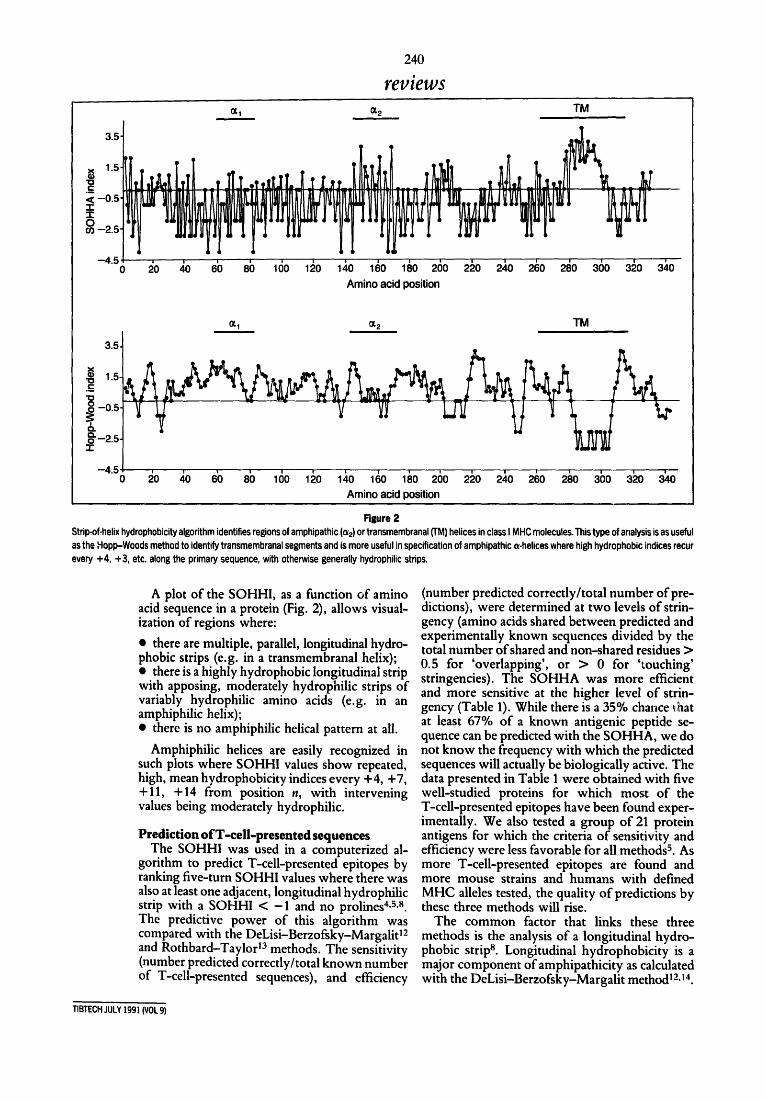
240
reviews (xl ~-2 TM
¢n -2.5
-450 20 ~0 6b 8b 100 1~0 laO 1~0 1~0 260 240 210 2~0 2~0 360 a~o a~o Amino acid position
1.5
1 0
o¢ 1 o¢ 2 TM
20 ~,0 6'0 80 160 120 1~,0 1(30 1130 260 220 2a,0 2(~0 280 3C)0 320 3~lO Amino acid position
Figure 2 Strip-of-helix hydrophobicity algorithm identifies regions of amphipathic (o~2) or transmembranal (TM) helices in class I MHC molecules. This type of analysis is as useful as the Hopp-Woods method to identify transmembranal segments and is more useful in specification of amphipathic o<-helices where high hydrophobic indices recur every +4, +3, etc. along the primary sequence, with otherwise generally hydrophilic strips.
A plot of the SOHHI, as a function of amino acid sequence in a protein (Fig. 2), allows visual- ization of regions where:
• there are multiple, parallel, longitudinal hydro- phobic strips (e.g. in a transmembranal helix); • there is a highly hydrophobic longitudinal strip with apposing, moderately hydrophilic strips of variably hydrophilic amino acids (e.g. in an amphiphilic helix); • there is no amphiphilic helical pattern at all.
Amphiphilic helices are easily recognized in such plots where SOHHI values show repeated, high, mean hydrophobicity indices every +4, +7, +11, +14 from position n, with intervening values being moderately hydrophilic.
Prediction ofT-ceil-presented sequences The SOHHI was used in a computerized al-
gorithm to predict T-cell-presented epitopes by ranking five-turn SOHHI values where there was also at least one adjacent, longitudinal hydrophilic strip with a SOHHI < -1 and no prolines 4,5,s. The predictive power of this algorithm was compared with the DeLisi-Berzofsky-Margalid 2 and Rothbard-Taylor 13 methods. The sensitivity (number predicted correctly/total known number of T-cell-presented sequences), and efficiency
(number predicted correctly/total number of pre- dictions), were determined at two levels of strin- gency (amino acids shared between predicted and experimentally known sequences divided by the total number of shared and non-shared residues > 0.5 for 'overlapping', or > 0 for 'touching' stringencies). The SOHHA was more efficient and more sensitive at the higher level of strin- gency (Table 1). While there is a 35% chance ~hat at least 67% of a known antigenic peptide se- quence can be predicted with the SOHHA, we do not know the frequency with which the predicted sequences will actually be biologically active. The data presented in Table 1 were obtained with five well-studied proteins for which most of the T-cell-presented epitopes have been found exper- imentally. We also tested a group of 21 protein antigens for which the criteria of sensitivity and efficiency were less favorable for all methods s. As more T-cell-presented epitopes are found and more mouse strains and humans with defined MHC alleles tested, the quality of predictions by these three methods will rise.
The common factor that links these three methods is the analysis of a longitudinal hydro- phobic strip s. Longitudinal hydrophobicity is a major component ofamphipathicity as calculated with the DeLisi-Berzofsky-Margalit method 12a4.
TIB'rECH JULY 199] (VOL 9)

241
reviews
The Rothbard-Taylor method identifies T-cell epitopes on the basis of motifs of four or five amino acids centered on hydrophobic amino acids 13.
Prediction o f helices in proteins The structural helices algorithm (SHA) was
generated to identify eight amino acid segments with potential longitudinal hydrophobic strips with mean SOHHI > 1.0, to merge overlapping predictions, and to shorten them if N-terminal Asn or Pro, or C-terminal Gly or Pro were present 6. The predictive power of this algorithm, in terms of sensitivity and efficiency at three levels of stringency, was equal to, or better than, the Chou-Fasman t5 and Garnier-Robson 16 methods for prediction of structural helices.
Analysis of the distribution of amino acids in longitudinal quadrants (i.e. a quadrant of the end- on view of the protein helix) of 247 helices of 55 proteins showed that Leu, lie, Val and Phe were strictly placed in, but Asp, Glu, Lys and Arg excluded from, the longitudinal quadrant with the greatest SOHHI (p < 0.001) along one side of the helix 7. In many instances, however, "I~cell- presented sequences were predicted by both SOHHA and SHA methods, indicating that helical folding of a polypeptide or protein seg- ment could be stabilized by a longitudinal hydro- phobic strip. In four instances, T-cell-presented segments which were not helices in crystallo- graphically determined protein structures were predicted with the SOHHA 6. Perhaps after ex- cision from a protein antigen, recurrent, aliphatic, hydrophobic residues lead to coiling and, thus, selection of the excised segment.
Testing the theory Catalysis afhelix formatian by peptides with longitudinal hydrophobic strips in th.,: presence of lipid vesicles
We tested the hypothesis that potential helices with longitudinal hydrophobic strips lead to structural helices in proteins and T-cell-presented epitopes by analysis of helical nucleation of syn- thetic peptides in the presence of lipid vesides. A series of peptides (including not only those known to be T-cell-presented but also prototypic helices [PH-1] constructed from a few different amino acids and having hydrophobic residues only in one longitudinal quadrant [Fig. 3]) was tested for helical coiling in phosphate buffer, with and without the presence of vesicles 9. Peptides without Pro frequently formed helices in tri- fluoroethanol (TFE), which decreased the di- electric constant of the solvent and enhanced hydrogen bonding between cydes of the peptidyl backbone when coiled as an o~-helix. More im- portantly, however, in a physiological phosphate buffer, helix formation was observed only with lipid vesicles (i.e. in the presence ofa hydrophobic
Table 1. Comparisons of methods to predict T-cell-presented peptides, evaluated with five well-studied proteins"
Known Amphi- Motifs Motifs sequences SOHHA pathicity a 4 e 5 e
Peptides 14 17 29 48 16
Sensitivity b Overlapping 0.43 0.29 0.00 0.00 Touching 0.57 0.71 0.79 0.43
Efficiency c Overlapping 0.35 0.14 0,00 0.00 Touching 0.47 0.36 0.25 0.40
a Chicken ovalbumin, chicken lysozyme, horse cytochrome, sperm whale myoglobin and staphylococcal nuclease. b Correct predictions/number of known T-cell-presented peptides. c Correct predictions/number of predictions. a Predictions from core positions with the DeLisi-Berzofsky-Margalit method=L e,Motifs 4' and "Motifs 5' are two methods incorporating predictions generated by the Rothbard-Taylor method, selecting for motifs of either 4 or 5 amino acids, respectively.
surface). With vesicles only, peptides with a strongly hydrophobic longitudinal strip formed helices. For example, the prototypic helix PH-1.0 (Leu-Tyr-Gln-Glu-Leu-Gln-Lys-Leu-Thr-Gln- Thr-Leu-Lys) which was not helical in phosphate buffer, became 43% helical with the addition of lipid vesicles. Analogs of PH-1.0 in which one or two Leu residues were replaced with Thr, coiled as helices on lipid residues only when three adjacent Leu residues were present, or when Leu5 and Leu8 were present on loops also joined by a salt bridge 17. While one can hypothesize that the longitudinal hydrophobic strip of a foreign peptide helix dissolves into the hydrophobic floor of the class II MHC desetope, such peptides might actually be in extended conformations in
PH-1 l G i n G l n G i n ZZ G l u Lys Thr l l l ~ L e u Leu Leu Leu ZV ~ Tyr Thr Lys
le~ I ~ ' a o l u ~ : g A s n T h : T : p I Z ~ A s n H i s Thr Lys I I I ~ L e u Leu N e t I l e V a l I V ~ L y s G l u Asp
Rgure 3 Sheet projections: the sequences of PH-] and I,-3 are presented in the format of an cx-helical sheet projection 4.s with the axial, hydrophobic strip arbitrarily placed in longitudinal quadrant II1. The sequence coils through the illustration as does the stripe on the barber's pole. This projection is better than the Edmundson wheel method for evaluating long helices and interactions between residues on adjacent loops of the helices. Optimal fits which are predicted with a computer closely approximate crystallographic structures 7, I,-3 is the 'prototypic antigen' sequence in I, which associates with class II MHC molecules until cleavage and release may permit peptide binding s. PH-1 is a synthetic peptide we have studied for correlations between structure and biophysical and biological function 9a7.
TIBI'ECH JULY 1991 {VOL 9)

242
reviews
desetopes. The amphiphilic helical nature of such peptides could reflect instead a step in processing such fragments that leads to protease protection as a tight, helical structure and/or to scavenging and transfer via the wall of a veside which extrudes non-adherent contents to a lysosome-bound sac. Such peptides might also fold on a transfer molecule, as hypothesized by Pierce et al. ts.
Future directions The strip-of-helix hydrophobicity index,
which quantifies the mean hydrophobicity of residues in a longitudinal strip along sequences coiled as a helix, reflects a basic principle that governs formation (and prediction) of structural helices in proteins and T-cell-presented epitopes. Stabilization of adjacent loops of a helix by the hydrophobic, usually aliphatic, residues leads to helix formation against hydrophobic regions of proteins or lipid membranes. Engineering of such hydrophobic strips might alter their biological properties in clinically useful ways.
One long-range goal is to determine the feasi- bility of exploiting these principles in altering the potency and MHC range of restriction ofpeptide vaccines and in altering the immunogenicity of therapeutic proteins. For example, by increasing the hydrophobicity of residues in the longitudinal hydrophobic strip, the potency of a peptide vaccine might be increased. There is usually room to do this because Nature does not engineer perfect strips tg. Patients who do not respond well might be immunized against an engineered pep- tide so that when an infection presents the wild- type sequence, enough MHC molecules will bind and present the wild-type epitope, such that in the presence of a primed T-cell response, the result is clinical protection. Such an approach could effect- ively expand the MHC range of response to the wild-type peptide.
The T-cell-stimulating sequences of thera- peutic or diagnostic proteins (e.g. murine mono- clonal IgG used to image or treat malignancies or suppress T-lymphocyte subsets), might be made less potent by changing to threonines individual aliphatic residues in longitudinal hydrophobic strips. Overall folding of the molecule, at least at the antibody-combining site, presumably will not be greatly affected, while immunogenicity (in terms ofeliciting helper T-cell responses) could be suppressed or eliminated by weakening or de- stroying the dominant T-cell-presented epitopes.
We also seek to identify examples where a high SOHHI sequence, presumably with a scavenger function in antigen processing, occurs only near the T-cell-presented epitope. Genetic engineering of a cleavage site between those cpitopes might destroy antigenicity of the intact protein, while not changing the structure of the T-cell-presented sequence, the limits of which had been deter- mined with synthetic peptides.
Acknowledgements We thank the following colleagues for their
contributions to these studies: Christopher M. Bositis, Thomas Ciardelli, Elisabeth J. Fowlie, Thomas G. Goldschmidt, Larry Hardy, Valery Lam, Quoc V. Nguyen, Lisa Phillips, Markus Salomon, Christopher H. Sorli and Steven Vasquez.
References I DeLisi, C. and Berzofsky, J. A. (1985) Proc. NatlAcad. Sci. USA
82, 7048-7052 2 Kaiser, E. T. and K6zdy, F.J. (1984) Science 223, 24%255 3 Berzofsky, J. A., Cease, K. B., Comette, J. L., Spouge, J. L.,
Margalit, H., Berkower, !. J., Good, M. F., Miller, L. H. and DeLisi, C. (1987) Immunol. Reo. 98, %52
4 Elliott, W. L., Stille, C.J., Thomas, L.J. and Humphreys, R. E. (1987) J. lmmunol. 138, 2949-2952
5 Stille, C.J., Thomas, L.J., Reyes, V. E. and Humphreys, R. E. (198"]) z~io;, lmmunol. 24, 1021-1027
6 Reyes, V. E., Phillips, L., Humphreys, R. E. and Lew, R. A. (1989)J. Biol. Chem. 264, 12845-12858
7 Torgerson, R. R., Lew, R. A., Reyes, V. E., Hardy, L. and Humphreys, R. E. J. Biol. Chem. 266, 5521-5524
8 Reyes, V. E., Fowlie, E. J., Lu, S., Phillips, L., Chin, L. T., Humphreys, R. E. and Lew, R. A. (1990) Mol. lmmu,ol. 27, 1021-1027
9 Lu, S., Reyes, V. E., Lew, R. A., Anderson, J., Mole, J., Humphreys, R. E. and Ciardelli, T. (1990)J. lmmunol. 145, 899-904
10 Reidhaar-Olson, J. F. and Sauer, R. T. (1988) Scie,ce241, 53-57 11 Pabo, C. O. and Lewis, M. (1982) Nature 298, 443-447 12 Marga|it, H., Spouge, J. L., Comette, J. L., Cease, K. B.,
DeLisi, C. and Berzofsky, J. A. (1987) J. hnmunol. 138, 2213-2229
13 Rothbard, J. B. and Taylor, W. R. (1988) EMBOJ. 7, 93-100 14 Cornette, J. L., Margalit, H., DeLisi, C. and Berzofsky, J. A.
(1989) Methods EnzFmol. 178, 611-634 15 Chou, P. Y. and Fasman, G. D. (1978) Adv. Enzymol. 47, 45-148 16 Gamier, J., Osguthorpe, D. J. and Robson, B. (1978)J. Mol.
Biol. 120, 97-120 17 Lu, S., Ciardelli, T., Reyes, V. E. and Humphreys, R. E.J. Biol.
Chem. (in press) 18 Lakey, E. K., Margoliash, E. and Pierce, S. K. (1987) Doc. Natl
Acad. Sci. USA 84, 165%1663 19 Reyes, V. E., Chin, L. T. and Humphreys, R. E. (1988) Mol.
lmmunol. 25, 867-871
Erratum
In the letter from J. Mol et al. in the June issueof TIBTECH, part of a sentence was deleted. From column 2, line 16, the text should have read: "In the particular DNA construct used, the promoter driving the sense transgene conve~ed with the nos NPT II
promoters are in the same orientation.' (Deletion underlined.) We apologize if this error caused any confusion.
TIBTECH JULY 1991 (VOL 9)


















