
College 4. Coordination interaction A dipolar bond, or coordinate covalent bond, is a description of...
16
College 4
-
date post
20-Dec-2015 -
Category
Documents
-
view
215 -
download
0
Transcript of College 4. Coordination interaction A dipolar bond, or coordinate covalent bond, is a description of...

College 4

Coordination interaction• A dipolar bond, or coordinate covalent bond, is a description of covalent
bonding between two atoms in which both electrons shared in the bond come from the same atom.
• Once such a bond has been formed, its strength and description is no different from that of other polar covalent bonds.
• They may involve metal ions. In such complexes, a molecule "donates" its"free" pairs of electrons to an otherwise naked metal cation, that "accepts" the electrons.
• For example O2 or CO to the Fe in porphyrins.

Protein folding
Proteins fold into a unique 3-D structure with a unique biological function. Even a mutation of one single amino acid may perturb this process in the sense that no longer a stable structure can be formed, or if it is formed it has no, or a much reduced, biological activity.
Folded state → minimum in energy
‘Simple’ rules:
1. Two atoms can never be in the same place. Steric hindering is one of the major driving forces for the formation of secondary structure
Protein folding
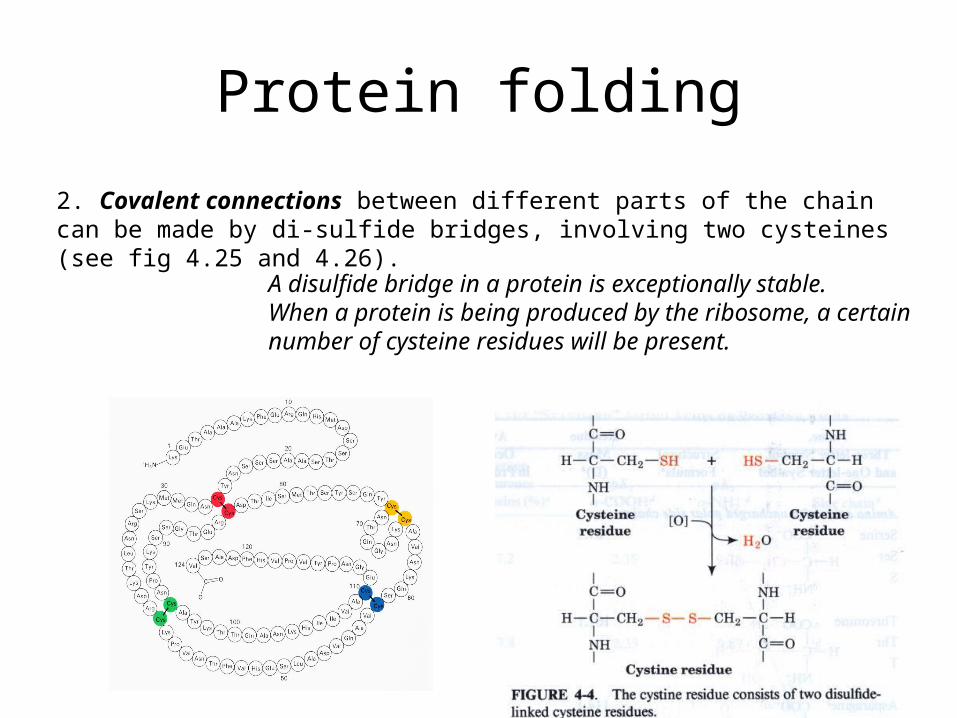
2. Covalent connections between different parts of the chain can be made by di-sulfide bridges, involving two cysteines (see fig 4.25 and 4.26).
A disulfide bridge in a protein is exceptionally stable. When a protein is being produced by the ribosome, a certain number of cysteine residues will be present.

Protein folding3. Non-covalent interactions.As a result of ionic interactions, Van der Waals forces and hydrogen bonds, each type of protein has a particular three dimensional structure, which is determined by the order of the amino acids in the chain.

Folded proteins
The final folded structure, or conformation, is the one in which the free energy is minimized (this minimum may not be unique for a system with so many degrees of freedom).
For globular, water soluble proteins this is generally a compact conformation with a hydrophobic core and polar/charged residues (such as arginine, glutamine, histidine etc.) exposed to the aqueous phase
Water → driving force

Folded proteins
When polar residues are buried in the inside of a protein they are generally hydrogen-bonded to other polar amino acids or to the polypeptide backbone

A distribution of unfolded species slide on the same energy surface to reach the minimal free energyThey meet at the saddle point where key residues have formed their native like contact.
Protein folding
Fig 4.31 Free energy of a folding protein plotted as a function of the number of contacts between residues (not all are favourable) and the number of native contacts (meaning contacts that also occur in the functional protein). The potential energy drives the system to a conformation where a certain number of native contacts has been established, but the chain is not yet folded. Note that there are many possible pathways. Once this point is reached, the chain folds rapidly.

Denaturation
Protein folding in a living cell is often assisted by special proteins called molecular chaperones. These proteins bind to partly folded polypeptide chains and help them progress along the energetically most favorable folding pathway. Chaperones are vital in the crowded conditions of the cytoplasm, since they prevent the temporarily exposed hydrophobic regions in newly synthesized proteins from associating with each other to form protein aggregates.

Structures
In vitro, X-ray
Highly organised ‘misfolded’ structure .Disease

Denaturation
Each protein normally folds up into a single stable configuration. However, this conformation often changes slightly when the protein interacts with other proteins in the cell. It is this modulation of the shape of proteins by its interaction with the ‘environment’ that is often crucial to the function of the protein.

Membranes
All cells are enclosed in a plasma membrane. This container acts as a selective barrier that enables the cell to concentrate nutrients gathered from its environment and retain the products it has synthesized for its own use, while excreting waste products. Without its plasma membrane the cell could not maintain its integrity as a coordinated chemical system.
Integral membrane proteins, like receptors and channels, cross the membrane. Receptors often have specific oligosaccharides attached to them that play a role in the recognition of specific compounds like hormones. Binding of a hormone then leads to a conformational change that is sensed at the cytosolic side of the membrane.
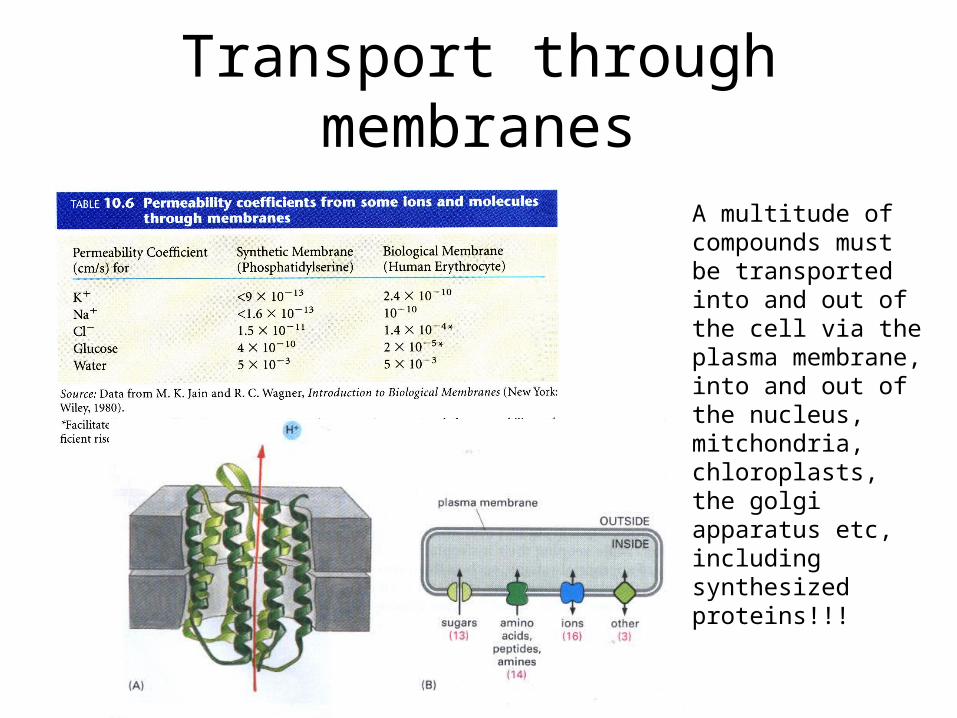
Transport through membranes
A multitude of compounds must be transported into and out of the cell via the plasma membrane, into and out of the nucleus, mitchondria, chloroplasts, the golgi apparatus etc, including synthesized proteins!!!
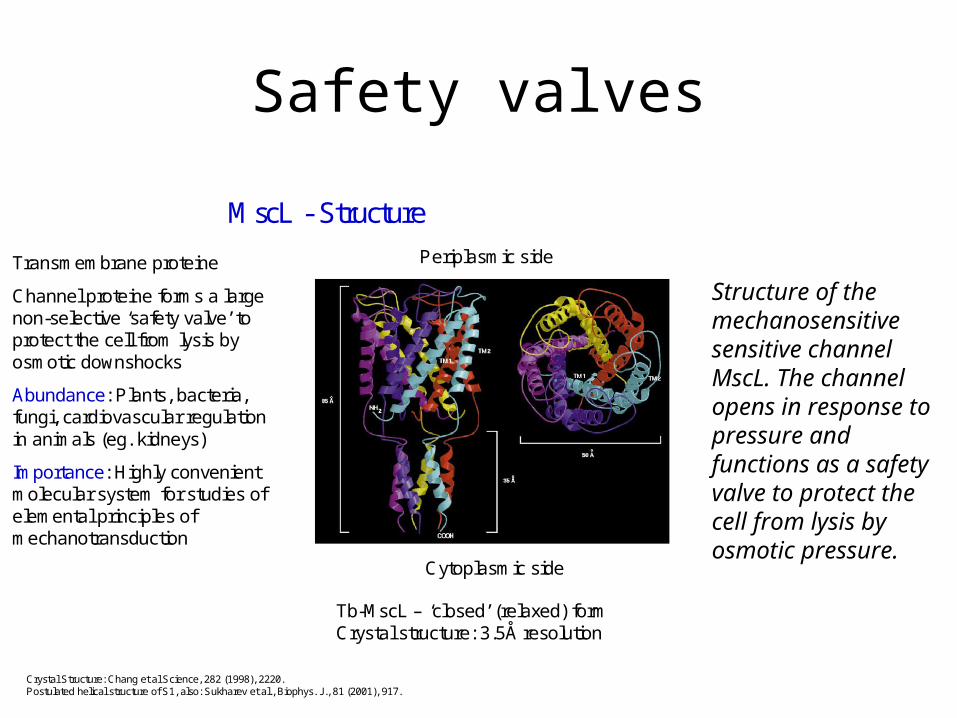
Safety valves
MscL - Structure
Transmembrane proteine
Channel proteine forms a large non-selective ‘safety valve’ to protect the cell from lysis by osmotic downshocks
Abundance: Plants, bacteria, fungi, cardiovascular regulation in animals (eg. kidneys)
Importance: Highly convenient molecular system for studies of elemental principles of mechanotransduction
Periplasmic side
Cytoplasmic side
Crystal Structure: Chang et al Science, 282 (1998), 2220.Postulated helical structure of S1, also: Sukharev et al., Biophys. J., 81 (2001), 917.
Tb-MscL – ‘closed’ (relaxed) form Crystal structure: 3.5Å resolution
Structure of the mechanosensitive sensitive channel MscL. The channel opens in response to pressure and functions as a safety valve to protect the cell from lysis by osmotic pressure.

Cells & membranes
• Nucleus: Genome management• Mitochondria & chloroplasts: Energy generation• EP & Golgi complex: protein synthesis and modification
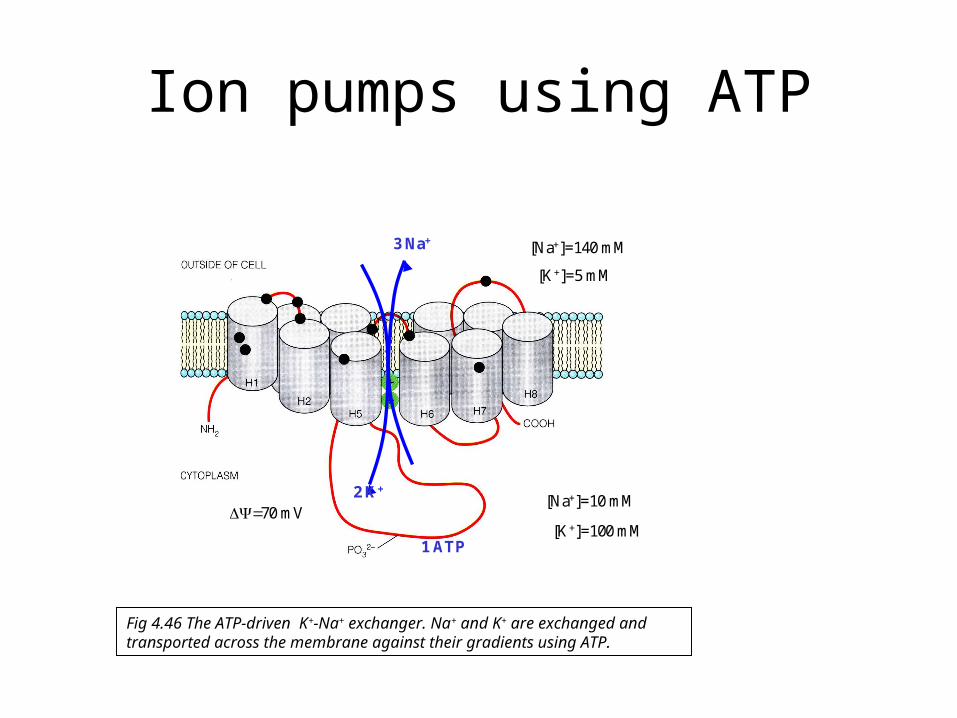
Ion pumps using ATP
[Na+]=140 mM
[Na+]=10 mM
[K+]=100 mM
[K+]=5 mM
70 mV
3 Na+
2 K+
1 ATP
Fig 4.46 The ATP-driven K+-Na+ exchanger. Na+ and K+ are exchanged and transported across the membrane against their gradients using ATP.


















