
Cloning, Sequencing, and Expression of the Gene · PDF fileCloning, Sequencing, and Expression...
12
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 265. No. 28, Issue of October 5, pp. 17307-17317,lSSO C 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S A. Cloning, Sequencing, and Expression of the Gene Encoding the Porcine cw2-Adrenergic Receptor ALLOSTERIC MODULATION BY Na+, H’, AND AMILORIDE ANALOGS* (Received for publication, April 17, 1990) Cheryl A. Guyer, Debra A. Horstman, Amy L. Wilson& J. David Clark& Edward J. Cragoe, Jr.ll, and Lee E. LimbirdII From the Department of Pharmacology, Vanderbilt University School of Medicine, Nashville, Tennessee 37232-6600 and VP. 0. iox 63158, kacogdoches, Teza.s 75963 The gene for an az-adrenergic receptor has been cloned from a porcine genomic library, using as a probe a 0.95-kilobase Pst fragment of the gene for the human platelet az-adrenergic receptor. The identity of the cloned porcine gene was confirmed initially on the basis of partial amino acid sequence information ob- tained following cyanogen bromide digestion of ho- mogeneous preparations of porcine brain az-adrener- gic receptors. The deduced amino acid sequence for the porcine receptor, when compared to other members of the family of guanine nucleotide-binding protein-cou- pled receptors, shares the same overall structural char- acteristics and most closely resembles the human plate- let Cl0 a2-adrenergic receptor (>93% homology). The putative porcine az-receptor gene was expressed in the CO&M6 cell line. Transfected cells display saturable [3H]yohimbine binding. The K. for [3H]yohimbine, de- termined in digitonin-solubilized preparations, is 5.8 IIM. The selectivity of agonists and antagonists in com- peting for [3H]yohimbine binding to membranes pre- pared from the transfected cells is characteristic of the azA subtype of adrenergic receptors. The porcine as- receptor also was expressed permanently in LLC-PKl porcine kidney cells at a level of 100 pmol/mg protein. The u2-agonist UK14304 is able to attenuate forskolin- or vasopressin-stimulated CAMP accumulation by at least 50% in these cells. Allosteric modulation of [3H] yohimbine binding by Na+, H+, and 5-amino-substi- tuted analogs of amiloride also was demonstrated for the a*-receptor expressed in CO&M6 cells. Moreover, these modulatory effects were quantitatively similar to those observed for homogeneous preparations of the a2-receptor purified from porcine brain cortex. Reten- tion of the effects of cations and amiloride analogs in transiently expressed a2-receptors supports the inter- pretation that the allosteric sites for these agents reside in the a2-receptor molecule itself. * This research was supported by National Institutes of Health Grants HL25182 and HL43671, and Fellowship Awards from the American Heart Association (to C. A. G. and D. A. H.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduer- tisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. The nucleotide sequence(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) 505652. $ Supported by NIH Training Grant GM07628. 3 Received support from NIH Training Grant GM07347. II To whom correspondence should be addressed. a2-Adrenergic receptors are one of a large family of cell surface receptors that interact with G-proteins as a means of transducing regulatory signals to target cells. In the case of cY2-receptors, G-proteins couple receptor occupancy to inhi- bition of adenylyl cyclase activity as well as to activation of K’ channels and suppression of voltage-sensitive Ca2+ chan- nel activity (1, 2). Subpopulations of o12-adrenergicreceptors are known to exist based on pharmacological as well as radioligand binding evidence (3), and the human genes for at least three cuz-receptorsubtypes have been cloned (4-7). Adrenergic ligand binding to the a*-adrenergic receptor has been shown to be allosterically modulated by Na+, H’, and 5- amino-substituted analogs of amiloride (8-10). The reciprocal effects of Na+ on cYz-receptor-agonist versus receptor-antago- nist interactions appear to be a property of all receptor populations linked to inhibition of adenylyl cyclase activity (11) and also have been demonstrated for /3-adrenergic recep- tors, which are linked to activation of adenylyl cyclase (12). In an effort to understand the molecular basis for these allosteric effects, the porcine brain o12-receptor was purified to apparent homogeneity (13). Unexpectedly, the homogene- ous receptor preparations retained allosteric modulation of adrenergic ligand binding by Na’ and amiloride analogs that was quantitatively indistinguishable from regulatory effects noted in crude membrane (9) and digitonin-solubilized (10) preparations of o12-receptors. The present manuscript describes cloning of the gene that codes for the porcine brain az-receptor previously purified to homogeneity. Transient expression of this gene in COS-M6 cells results in concomitant expression of adrenergic ligand binding and allosteric modulation of that binding by cations and 5-amino-substituted analogs of amiloride. EXPERIMENTAL PROCEDURES Materials Reagents-[[y-“*P]ATP (-6000 Ci/mmol), [a-32P]dCTP (-3000 Ci/ mmol) nick translation kit, [o(-““S]dATP (500 Ci/mmol), [‘*‘I]iodo- para-aminoclonidine (-2000 Ci/mmol), and [3H]yohimbine (70-90 Ci/mmol) were from Du Pont-New England Nuclear. Acetonitrile and methanol were purchased from Burdick and Jackson. HPLC’ ’ The abbreviations used are: HPLC, high performance liquid chro- - _ matography; HEPES, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid; EGTA, [ethylenebis(oxyethylenenitrilo)]tetraacetic acid; NMDG-Cl, N-methyl-D-glucamine chloride; WGA, wheat germ agglutinin; ““I-Rau-AzPEC, 17a-hydroxy-20a-yohimban-16@-[N-(4- azido-3-[““I]iodo)phenethyl]carboxamide; MOPS, 3-(N-morpho- 1ino)propanesulfonic acid; SDS, sodium dodecyl sulfate; kb, kilo- base(s); PBS, phosphate-buffered saline; GppNHp, guanyl-5’-yl imidodiphosphate. 17307 by guest on May 12, 2018 http://www.jbc.org/ Downloaded from
Transcript of Cloning, Sequencing, and Expression of the Gene · PDF fileCloning, Sequencing, and Expression...

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 265. No. 28, Issue of October 5, pp. 17307-17317,lSSO C 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S A.
Cloning, Sequencing, and Expression of the Gene Encoding the Porcine cw2-Adrenergic Receptor ALLOSTERIC MODULATION BY Na+, H’, AND AMILORIDE ANALOGS*
(Received for publication, April 17, 1990)
Cheryl A. Guyer, Debra A. Horstman, Amy L. Wilson& J. David Clark& Edward J. Cragoe, Jr.ll, and Lee E. LimbirdII From the Department of Pharmacology, Vanderbilt University School of Medicine, Nashville, Tennessee 37232-6600 and VP. 0. iox 63158, kacogdoches, Teza.s 75963
The gene for an az-adrenergic receptor has been cloned from a porcine genomic library, using as a probe a 0.95-kilobase Pst fragment of the gene for the human platelet az-adrenergic receptor. The identity of the cloned porcine gene was confirmed initially on the basis of partial amino acid sequence information ob- tained following cyanogen bromide digestion of ho- mogeneous preparations of porcine brain az-adrener- gic receptors. The deduced amino acid sequence for the porcine receptor, when compared to other members of the family of guanine nucleotide-binding protein-cou- pled receptors, shares the same overall structural char- acteristics and most closely resembles the human plate- let Cl0 a2-adrenergic receptor (>93% homology). The putative porcine az-receptor gene was expressed in the CO&M6 cell line. Transfected cells display saturable [3H]yohimbine binding. The K. for [3H]yohimbine, de- termined in digitonin-solubilized preparations, is 5.8 IIM. The selectivity of agonists and antagonists in com- peting for [3H]yohimbine binding to membranes pre- pared from the transfected cells is characteristic of the azA subtype of adrenergic receptors. The porcine as- receptor also was expressed permanently in LLC-PKl porcine kidney cells at a level of 100 pmol/mg protein. The u2-agonist UK14304 is able to attenuate forskolin- or vasopressin-stimulated CAMP accumulation by at least 50% in these cells. Allosteric modulation of [3H] yohimbine binding by Na+, H+, and 5-amino-substi- tuted analogs of amiloride also was demonstrated for the a*-receptor expressed in CO&M6 cells. Moreover, these modulatory effects were quantitatively similar to those observed for homogeneous preparations of the a2-receptor purified from porcine brain cortex. Reten- tion of the effects of cations and amiloride analogs in transiently expressed a2-receptors supports the inter- pretation that the allosteric sites for these agents reside in the a2-receptor molecule itself.
* This research was supported by National Institutes of Health Grants HL25182 and HL43671, and Fellowship Awards from the American Heart Association (to C. A. G. and D. A. H.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduer- tisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) 505652.
$ Supported by NIH Training Grant GM07628. 3 Received support from NIH Training Grant GM07347. II To whom correspondence should be addressed.
a2-Adrenergic receptors are one of a large family of cell surface receptors that interact with G-proteins as a means of transducing regulatory signals to target cells. In the case of cY2-receptors, G-proteins couple receptor occupancy to inhi- bition of adenylyl cyclase activity as well as to activation of K’ channels and suppression of voltage-sensitive Ca2+ chan- nel activity (1, 2). Subpopulations of o12-adrenergic receptors are known to exist based on pharmacological as well as radioligand binding evidence (3), and the human genes for at least three cuz-receptor subtypes have been cloned (4-7).
Adrenergic ligand binding to the a*-adrenergic receptor has been shown to be allosterically modulated by Na+, H’, and 5- amino-substituted analogs of amiloride (8-10). The reciprocal effects of Na+ on cYz-receptor-agonist versus receptor-antago- nist interactions appear to be a property of all receptor populations linked to inhibition of adenylyl cyclase activity (11) and also have been demonstrated for /3-adrenergic recep- tors, which are linked to activation of adenylyl cyclase (12). In an effort to understand the molecular basis for these allosteric effects, the porcine brain o12-receptor was purified to apparent homogeneity (13). Unexpectedly, the homogene- ous receptor preparations retained allosteric modulation of adrenergic ligand binding by Na’ and amiloride analogs that was quantitatively indistinguishable from regulatory effects noted in crude membrane (9) and digitonin-solubilized (10) preparations of o12-receptors.
The present manuscript describes cloning of the gene that codes for the porcine brain az-receptor previously purified to homogeneity. Transient expression of this gene in COS-M6 cells results in concomitant expression of adrenergic ligand binding and allosteric modulation of that binding by cations and 5-amino-substituted analogs of amiloride.
EXPERIMENTAL PROCEDURES
Materials
Reagents-[[y-“*P]ATP (-6000 Ci/mmol), [a-32P]dCTP (-3000 Ci/ mmol) nick translation kit, [o(-““S]dATP (500 Ci/mmol), [‘*‘I]iodo- para-aminoclonidine (-2000 Ci/mmol), and [3H]yohimbine (70-90 Ci/mmol) were from Du Pont-New England Nuclear. Acetonitrile and methanol were purchased from Burdick and Jackson. HPLC’
’ The abbreviations used are: HPLC, high performance liquid chro- - _ matography; HEPES, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid; EGTA, [ethylenebis(oxyethylenenitrilo)]tetraacetic acid; NMDG-Cl, N-methyl-D-glucamine chloride; WGA, wheat germ agglutinin; ““I-Rau-AzPEC, 17a-hydroxy-20a-yohimban-16@-[N-(4- azido-3-[““I]iodo)phenethyl]carboxamide; MOPS, 3-(N-morpho- 1ino)propanesulfonic acid; SDS, sodium dodecyl sulfate; kb, kilo- base(s); PBS, phosphate-buffered saline; GppNHp, guanyl-5’-yl imidodiphosphate.
17307
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17308 Cloning and Expression of cwAdrenergic Receptor Gene
grade trifluoracetic acid, cyanogen bromide, and RBS-35 detergent for cleaning glassware used for protein sequencing were from Pierce Chemical Co. &Mercaptoethanol (>99%) was obtained from Fluka Biochemika. Acrylamide, N,N’-methvlenebisacrvlamide, and sodium dodecyl sulfate were specially purified for electrophoresis by BDH Chemicals Ltd. (United Kingdom). Formic acid (98%. ACS erade) and phenol (99+% Gold Label) were from Aldrich. The EMBL-3 porcine genomic library was obtained from Clontech. Restriction endonucleases were purchased either from Boehringer Mannheim, New England Biolabs, International Biotechnologies, Inc., or Pro- mega. T4 polynucleotide kinase was from New England Biolabs and T4 DNA ligase was from Promega. Bacterial alkaline phosphatase and endonuclease-free bovine serum albumin were from Bethesda Research Laboratories. Sequenase DNA sequencing kits were ob- tained from United States Biochemical Corp. M13mplS/M13mp19 and Ficoll 400 were from Pharmacia LKB Biotechnology Inc. pBluescript SK(+) was purchased from Stratagene. Cetylpyridinium bromide, chloramphenicol, ampicillin, polyvinylpyrrolidone, salmon sperm DNA, (-)-epinephrine, (+)-epinephrine, yohimbine, and oxg- metazoline were from Sigma. Phentolamine was kindly provided by CIBA Pharmaceutical Co. Prazosin and UK-14304 were eifts from Pfizer. Forskolin was purchased from Calbiochem. Nylon Hybond-N filters (85 and 137 mm) were from Amersham, Biotrans nylon mem- branes were from ICN, and nitrocellulose HA filters (0.45 Mm, 137 mm) were from Millipore. Urea was purchased from J. T. Baker Chemical Co. and Sea Plaque agarose was from PMC Corporation. Bacto-agar, Bacto-tryptone, and yeast extract were purchased from Difco. The pCMV4 eukaryotic expression vector was kindly provided by Dr. David Russell (University of Texas Health Sciences, Dallas, TX). LLC-PKl (Cl,) cells were kindly provided by Dr. Carolyn Slavman (Yale). WGA-aaarose was purchased from Vector Labora- torl’es. The cuz-adrenergic-receptor-selective photoaffinity label, “‘I- Rau-AzPEC, was synthesized by the method of Lanier et al. (14). Amiloride analogs were purchased from Dr. E. J. Cragoe, Jr. All other chemicals were reagent grade or better and all other biochemicals were of molecular biology grade or better.
Methods
Purification and Electroelution of the Porcine Brain an-Adrenergic Receptor-ap-Adrenergic receptor was purified to homogeneity from porcine brain cortex by two sequential rounds of affinity chromatog- raphy on yohimbine-agarose as previously described (13).
To remove digitonin, which might otherwise interfere with the HPLC sequence analysis, ocx-adrenergic receptor preparations were subjected to SDS-polyacrylamide-gel electrophoresis and electroelu- tion. Approximately 1 nmol of homogeneous az-adrenergic receptor (265 ml) was concentrated in an Amicon stirred cell over a PM-30 membrane to a volume of 10 ml. This solution was further concen- trated in an Amicon CF50 filtration cone by centrifugation at 1000 rpm until the volume reached approximately 1.1 ml. SDS (20%) was added to the sample to a final concentration of 5%. This sample (1.4 ml) was exchanged into SDS sample buffer (0.125 M Tris, pH 6.8, 2% SDS, 10% glycerol, 7.5 mg/ml dithiothreitol) by chromatography over an 18 ml (0.9 x 28 cm) G-50 Sephadex column. The eluted sample (2.5 ml) was subjected to SDS-polyacrylamide gel electrophoresis (11% separating gel) as described by Laemmli (15) with the modifi- cations and protein visualization procedures recommended by Hun- kapillar et al. (16). Electroelution of a*-adrenergic receptor from finely minced gel was performed according to the protocol of Hunkapillar et al. (16).
Cyanogen Bromide Digestion and HPLC Separation of Resultant Peptides-The electroeluted a,-adrenergic receptor (10 ml) in elec- troelution dialysis buffer (10 mM ammonium bicarbonate, 0.02% SDS) was concentrated to 0.5 ml in a Savent Speed-Vat concentrator. To reduce any methionyl residues which may have been oxidized during the electrophoresis and electroelution procedures (17), 2- mercaptoethanol (25 ~1) was added to a final concentration of 5% and the reaction was allowed to stir 18 h at room temperature. The sample was then dialyzed extensively against 10 mM ammonium bicarbonate to remove SDS and Z-mercaptoethanol prior to cyanogen bromide digestion. Following evaporation to dryness in a l&ml Reactivial (Pierce Chemical Co.), the sample was redissolved in 70% formic acid (150 ~1) and flushed with dry nitrogen. Cyanogen bromide (50 ~1, 240 mg/ml in 70% formic acid) was added to a final concen- tration of 60 mg/ml and the reaction was allowed to stir for 24 h at room temperature in the dark (17). A second aliquot of cyanogen bromide (50 ~1, 300 mg/ml in 70% formic acid) was added and
digestion was continued for 24 h. Following evaporation to dryness as described (17), a crude separation of water-soluble from hydropho- bic peptides was affected by dissolution of the sample in 100 ~1 of water-saturated n-butanol and by extraction twice with an equal volume of water. Each phase was fractionated further by reverse- phase HPLC (Applied Biosystems 130A HPLC System) with a Brownlee (4 Frn, Aquapore) C-8 column (1.0 X 250 mm) using a gradient of 20-80% acetonitrile in 0.1% trifluoracetic acid, at a flow rate of 70 pl/min. Eluted material was detected by monitoring ab- sorbance at 220 nm. Fractions corresponding to peaks of interest, as determined by comparison to blank samples which had been treated identically, were pooled and subjected to amino acid sequence analysis on an Applied Biosystems 470A Protein Sequencer in conjunction with an Applied Biosystems 120A phenylthiohydantoin derivative analyzer.
DNA Probes for Library Screening and Sequencing-Oligonucleo- tides used for screening and sequencing primers were synthesized on an Applied Biosystems 380A DNA Synthesizer using cyanoethyl- phosphoramidite chemistry. Oligonucleotides were purified either by standard sodium acetate/ethanol precipitation or by polyacrylamide gel eiectrophoresis (1.5 mm thick, 15% gel) under denaturing condi- tions (18). Following electrophoresis, oligonucleotides were visualized by short wave UV illumination (UVS-54 Mineralight) and excised from the gel. The oligonucleotides were recovered by extraction in 0.5 M ammonium acetate, 1 mM EDTA, 10 mM magnesium acetate, followed by purification on a C-18 cartridge Sep-Pak (Waters Asso- ciates) and sodium acetate/ethanol precipitation (18).
Oligonucleotide probes used for screening purposes were 5’-end- labeled with [r-“*P]ATP using T4 polynucleotide kinase (IS), and purified from unincorporated [-y-“‘P]ATP by cetylpyridinium bro- mide nrecinitation (19). In addition. a 0.95-kb Pst fragment from the human CiO al-adrenergic receptor’gene (6, kindly provided by Dr. Claire Fraser. NIH) was radiolabeled with la-“*PldCTP bv nick translation using the materials and protocols ‘provided with fhe DU Pont-New England Nuclear nick translation kit and used as a probe for library screening. In general, the specific activity of radiolabeled probe was on the order of 1 X 10’ dpm/pg DNA.
Screening of the EMBL-3 Library-Recombinant phages from an amplified EMBL-3 porcine genomic library (Clontech, titer 1 x 10’ pfu/ml) were incubated with Escherichia coli NM538 cells for 20 min at 37 “C, mixed with 0.75% Luria broth soft agarose and plated on 1.5% Luria broth agar in 150-mm Petri dishes at a density of approx- imatelv 50,000 plaaue-forming units/dish. Plates were incubated at 37 “C until lytic plaques began to appear (-8-9 h). After cooling to 4 “C, phage were lifted onto duplicate nylon filters which were then prepared for screening as described (18).
Replicate filters were prehybridized for 12 h at 42 ‘C in a solution containing5 X SSC (1 X SSC = 150 mM NaCl, 15 mM sodium citrate), 0.1% SDS, 5 X Denhardt’s (1 X Denhardt’s = 0.02% Ficoll, 0.02% polyvinylpyrrolidone, and 0.02% bovine serum albumin), 100 rg/ml sheared salmon sperm DNA, 30% formamide. Hybridization was carried out for 18 h, at 42 “C in the same solution containing also 0.5 x 10” cpm/ml denatured “‘P-labeled 0.95.kb Pst fragment human ClO-a:! gene probe. Filters were washed initially with 6 X SSC at room temperature then with 1 x SSC at 65 “C until background radioactivity, as monitored with a hand-held Geiger counter, had decreased to acceptable levels (-45 min). Subsequently, filters were subjected to autoradiography on Kodak XAR-5 film with one DuPont Quanta III screen for 12-15 h at -70 “C. Phage plaques that hybrid- ized positively on replicate filters were identified and plaque-purified by secondary and tertiary screening procedures as described (18).
DNA Subcloning and Sequencing-Genomic DNA restriction endonuclease fragments of interest were identified by agarose-gel electrophoresis and Southern hybridization analysis (18) with either ““P-labeled 0.95.kb Pst fragment or annronriate “‘P-labeled oliaonu- __ _ cleotide probes. These fragments were subcloned by standard recom- binant DNA techniques into M13mp19 (Sac1 fragments), into M13mp18 and 19 (SacI/ScaI fragments), or into Stratagene pBlue&ript SK(+) (Pst fragment). DNA sequencing was performed bv the dideoxv chain termination method of Sanger et al. (20) with [&-3”S]dATP and T7 DNA polymerase (Sequenase) using the proto- cols appropriate for Ml3 single-stranded templates or for double- stranded templates included in the United States Biochemical Corp. Sequenase DNA kit. Electrophoresis and autoradiography of reaction products was carried out in 6% polyacrylamide gels containing 42% urea (w/v), 45 mM Tris borate, 1.25 mM EDTA followed by autora- diography on Kodak XAR film. In cases of extreme G-C compression, nucleotide mixtures containing 7-deaza-dGTP were employed and
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cloning and Expression of cwAdrenergic Receptor Gene 17309
reaction products were separated by electrophoresis as described previously (20) except that formamide was substituted for water in the gel mix. Sequencing was initiated with Ml3 universal primer, oligonucleotides used for screening, or, as porcine sequence became available, with appropriately designed complimentary oligonucleo- tides. Using this primer-walk strategy, the subcloned inserts were sequenced as depicted in Fig. 1A.
Construction of the Eukaryotic Expression Vector fpCMV4-aJR) Containing the Porcine cu,-Adrenergic Receptor Gene-A 6-kb KpnI fragment containing the porcine a*-receptor was excised from the EMBL-3 clone and subcloned into the KpnI site of pBluescript SK(+) (Stratagene). From this subclone, a 1.5-kb NcoI-ScaI fragment cor- responding to the coding region of the cu2-receptor and 177 base pairs of 3’-noncoding sequence was purified and converted to a blunt-end fragment using the Klenow fragment of DNA polymerase I. Following ligation to KpnI linkers and digestion to produce cohesive ends, this fragment was subcloned into the KpnI site of the eukaryotic expres- sion vector pCMV4.
Growth of COS-h46 Cells and Transfection with DCMV~-CWIR- COS-M6 cells (a subclonal line of COS-7 cells obtained from Edith Womack in the laboratory of J. L. Goldstein, University of Texas Health Sciences, Dallas) were grown in monolayer cultures at 37 “C in a 5% CO, incubator in Dulbecco’s modified Eagle’s medium sup- plemented with 10% fetal calf serum (Sigma), 20 mM HEPES, pH 7.4, 100 units/ml penicillin, and 100 pg/ml streptomycin (“complete medium”). Transient expression of cu2-receptor gene in the COS-M6 cells was mediated by DEAE-dextran (21). Complete medium was supplemented to contain the expression plasmid, pCMV4-a2AR (10 rg/8 ml), 100 mM Tris-HCl, pH 7.3, and 0.25 mg/ml DEAE-dextran (Pharmacia, M, = 5 x 10”). COS-M6 cells were plated at 2 X lo6 cells/lOO-mm dish. The following day the cells were rinsed with phosphate-buffered saline (PBS, 137 mM NaCl, 2.68 mM KCl, 1.47 mM KH,PO,, and 8.2 mM NaH,PO+ pH 7.4) and were incubated with the DNA-containing DEAE-dextran medium (8 ml/lOO-mm dish) for 6 h at 37 “C. After rinsing once with PBS, the cells were incubated for 3 min with 10% dimethyl sulfoxide (in PBS) at room temperature. The dimethyl sulfoxide solution was aspirated and 0.1 mM chloro- quine in complete medium (8 ml/lOO-mm dish) was added for 2 h at 37 “C. Following two rinses with PBS, the cells were grown in com- plete medium (10 ml/lOO-mm dish) and as-receptor binding assayed 42-72 h after transfection.
Establishing Permanent Transformants Expressing the Porcine 012- Adrenergic Receptor Gene-To assess cu2-adrenergic receptor coupling to GTP-binding proteins and distal signaling systems, permanent transformants expressing the oc2-receptor were established. LLC-PKl porcine kidney cells plated at 50% confluence in loo-mm dishes were transfected with 20 fig of the pCMV4-u2AR plasmid and 2 pg of the RSVneo plasmid using standard CaPO,, precipitation techniques (22). Individual clones were selected and expanded. Those clones express- ing a2-adrenergic receptor binding activity were identified using the radiolabeled partial agonist [““I]iodo-para-aminoclonidine (23). Thus, confluent loo-mm dishes (-2 x 10’ cells) were washed twice with phosphate-buffered saline at room temperature and then placed on ice. Five ml of ice-cold lvsis buffer containing 5 mM Tris-HCl, pH 7.6, and 5 mM EGTA was added to each dish. The cells were detached from the dish with a rubber policeman and lysed by 4 up and down passes through a 21-gauge needle attached to a lo-ml syringe. This lysate was transferred to a 12-ml tube. The dish was washed with an additional 5 ml of ice-cold lysis buffer and this wash was pooled with the 5 ml of lysate. The cell lysate was centrifuged at 4 “C for 10 min at 18,000 rpm in an SS-34 rotor in a Sorvall RC5B centrifuge. The pellet obtained was washed two additional times by resuspension with a plastic pipette into the ice-cold lysis buffer. The final pellet was dislodged into 500 ~1 of 50 mM Tris-HCl, 10 mM MgCl*, 5 mM EGTA, pH 8.0, transferred to a 1.5-ml Eppendorf tube and resuspended using a 25-gauge needle attached to a 3-ml syringe.
The assay for [““Iliodo-para-aminoclonidine binding was per- formed in 12 X 75-mm polypropylene tubes containing 80 ~1 of LLC- PKl cell particulate preparations, prepared as described above, 10 ~1 of [““Iliodo-para-aminoclonidine (160,000 cpm, 0.5 nM final concen- tration of radioligand in the incubation), and 10 ~1 of H20 or lo-” M yohimbine (to define nonspecific binding). Incubations were for 30 min at 25 “C, and were ter-minated by the addition of 4.5 ml of ice- cold 25 mM glvcvlglvcine buffer, pH 7.6, followed bv vacuum filtration through GF/B glass fibre filters (Whatman). The filters were washed twice with 5 ml of ice-cold glycylglycine buffer and then counted in a Beckman 4000 y counter. Clones expressing [““I]iodo-para-amino- clonidine binding were characterized further for their properties of
[“Hlyohimbine binding and receptor-mediated inhibition of CAMP accumulation.
Preparation of Cell Membranes and Digitonin-solubilized Prepara- tions from pCMV4-aAR-transfected COS-M6 Cells-At various times after transfection, membranes were prepared from COS-M6 cells transfected with the pCMV2-crsAR plasmid. The cells were rinsed once with PBS then removed from the dish into PBS containing 1 mM EDTA (5 ml/dish) bv scrauing with a rubber uoliceman. The cells were collected by cenirifugation at 1000 rpm ina Sorvall GLC- tabletop centrifuge and subsequently were lysed in hypotonic buffer (10 ml) containing 15 mM HEPES, 5 mM EDTA, and 5 mM EGTA, pH 8.0, by passage through a 25-gauge needle 10 times. Membranes were collected by recentrifugation at 39,000 X g, 10 min, 4 “C. The membrane pellet was resuspended in the hypotonic buffer (10 ml), rehomogenized, and collected by recentrifugation. The pellet was resuspended in assay buffer (20 ml) containing 25 mM glycylglycine, 20 mM HEPES, 40 mM NMDG-Cl, 5 mM EGTA, and 10 mM MgCl*, pH 8.0, by homogenization, and a preliminary binding assay was performed to determine total and nonspecific [“Hlyohimbine binding for each membrane preparation. The membrane protein concentra- tion was adjusted so-that each assay contained between 5 and 10 X 10” cnm of 1”Hlvohimbine bindine. assaved at 7.5 nM 1”Hlvohimbine. Typically, one’iOO-mm dish seeded at 2 x lo6 cells/dish-and trans- fected with 10 pg of ~CMV-LU~AR plasmid generated 1.2 X lo6 cpm (15 pmol) of [“Hlyohimbine binding. Membranes were used immedi- ately for ligand binding assays or stored at -70 “C until needed.
To prepare digitonin-solubilized preparations of the a*-receptor, 5 ml of membrane preparations containing -1.5 x lo6 cpm (18.8 pmol) of [“Hlyohimbine binding were washed by resuspension in 15 ml of buffercontaining 15 mM- HEPES, 5 mM.EDTA, 5 mM EGTA, pH 8.0, and centrifuged at 39.000 x P for 30 min at 4 “C. Solubilization of the cup-receptors was accomplished by homogenization into 10 ml of ice-cold buffer containing 1% digitonin, 20 mM HEPES, 25 mM glycylglycine, 100 mM NaCl, 5 mM EGTA, pH 8.0 (solubilization buffer), using 10 up and down strokes in a tight-fitting, motor-driven Teflon glass homogenizer held in ice. After dilution of this lo-ml suspension to 28 ml with 18 ml of additional solubilization buffer, the preparation was sonicated in a bath sonifier at 4 “C for 30 min, followed by centrifugation at 100,000 X g for 60 min at 4 “C. Efficiency of receptor extraction was routinely 50-55%, yielding 30,000 cpm of [“Hlyohimbine binding/ml of digitonin-solubilized preparation, as- sayed at 7.5 nM [“Hlyohimbine. The digitonin-solubilized preparation was used immediately or stored at -70 “C until needed.
Ligand Binding Assays-Membranes were incubated for 30 min at 25 “C with 7.5 nM [“Hlyohimbine in the absence or presence of various concentrations of competing ligands in assay buffer containing 25 mM glycylglycine, 10 mM HEPES, 40 mM NMDG-Cl, 5 mM EGTA, and 10 mM MgC12, pH 8.0. Nonspecific binding was defined as that binding obtained in the presence of 10 PM phentolamine (typically 10% of total binding). Assays were terminated by addition of 5 ml of ice-cold 25 mM glycylglycine, pH 7.6, followed by filtration through GF/B glass fibre filters (Whatman), and washing twice with 5 ml of ice-cold 25 mM glycylglycine, pH 7.6. The filters were dried for 30 min in a 60 “C oven and counted in 10 ml of NEN-963 scintillation fluor at an efficiency of 45%.
To obtain saturation binding isotherms and to investigate the effects of amiloride analogs and cations on receptor-ligand interac- tions, digitonin-solubilized preparations of transfected COS-M6 cells were used rather than membrane vesicles to reduce further variability arising potentially from the nonuniform distribution of ao-receptors in membrane vesicles. The effects of receptor density on measuring ligand binding are most clearly evidenced in studies assessing the rate of dissociation of radiolabeled ligand. To interpret effects of amiloride analogs or cations on the rate of [“Hlyohimbine dissociation as resulting from interaction at sites “other” (i.e. allosteric) than the adrenergic ligand-binding pocket, it is essential to first establish that the concentration of unlabeled yohimbine added to allow monitoring of the dissociation rate is sufficient to fully occupy all adrenergic ligand sites. Thus, the inclusion of additional yohimbine in the incubation should not itself accelerate the rate of [“Hlyohimbine dissociation. Presumably because of the non-uniform density of crp- receptors in membrane vesicles prepared from COS-M6 cells tran- siently expressing the cuZ-receptor gene, the rate of [“Hlyohimbine dissociation in the presence of 50 FM yohimbine was further acceler- ated by adding unlabeled yohimbine up to 200 pM. In contrast, once the a,-receptor was solubilized from the membranes and presumably monodispersed in solution, 50 pM yohimbine was sufficient to fully occupy all cup-receptors during the dissociation phase and no further
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cloning and Expression of cu2-Adrenergic Receptor Gene
CTCC*AGCC‘ TCCGCTGFC* GCTGGGITTG hGGGGCGCGT CThTGTIGTC CTTRGCCCTC TGAllhCCCGG
FIG. 1. Sequencing strategy, nucleotide and deduced amino acid sequence of the porcine cu2-adrenergic receptor gene. A, sequencing strategy of various restriction fragments derived from positively hybridizing porcine genomic EMBL-3 recombinant phage and subcloned as described under “Experimental Procedures.” Open boxes indicate the coding region and arrows indicate the direction and extent of sequencing. B, the nucleotide and deduced amino acid
acceleration of the dissociation rate occurred if an additional 200 flM yohimbine was introduced in the incubation.
Assays measuring [“Hlyohimbine binding to soluble cuz-receptor preparations were performed at 15 “C using digitonin-solubilized 01~- receptor preparations which had been exchanged by Sephadex G-50 chromatography (13) into “Na’-free” buffer containing 0.2% digi- tonin, 25 mM glycylglycine, 40 mM HEPES, 100 mM NMDG-Cl, 5 mM EGTA, pH 8.0. The rate of [“Hlyohimbine dissociation in the presence of 50 FM yohimbine and in the absence or presence of allosteric modulators was monitored as described previously for (Ye- receptor preparations digitonin-solubilized from porcine brain cortex (10). Separation of bound from free [SH]yohimbine was accomplished using Sephadex G-50 chromatography on 4-ml columns, as described previously (13). Details pertinent to each experiment are provided in the figure legends.
WGA-Agarose Chromatography of Digitonin-solubilized Prepara- tions of pCVM4-&AR-transfected COS-M6 Cells-To 10 ml of digi- tonin-solubilized preparation was added 1 M MgC12 to a final concen- tration of 10 mM to stabilize receptor-wheat germ agglutinin inter- actions and phentolamine to a final concentration of 10 pM to stabilize the receptor during chromatography. The receptor preparation was then batch-adsorbed overnight by rotating end over end at 4 “C with 1 ml of WGA-agarose previously equilibrated in 0.2% digitonin, 25 mM glycylglycine, 100 mM NaCl, 5 mM EGTA, 10 mM MgC12, pH 7.6. Following five rapid washes with 10 ml/wash of the above equilibra- tion buffer, the resin (as a slurry) was quickly packed (25 ml/h) into a 0.9 X 1.5-cm column. The packed column was washed with 1 ml of Mg’+-free wash buffer (0.1% digitonin, 25 mM glycylglycine, 100 mM NaCl, 5 mM EGTA, pH 7.6) at a flow rate of 10 ml/h. The receptor binding activity subsequently was eluted from the column at a rate of 4 ml/h in 0.4-ml fractions using a gradient of O-500 mM N- acetylglucosamine in Mg’+-free wash buffer. The a2-receptor binding activity recovered in the pooled fractions represented 57% of the total binding activity in the digitonin-solubilized preparation used as start- ing material.
Photoaffinity Labeling of Expressed Porcine aa-Adrenergic Recep- tor-WGA-agarose eluates of expressed porcine as-receptor prepara- tions were incubated with 2 nM of the oc,-adrenergic receptor-selective photoaffinity label, ““I-Rau-AzPEC, in 0.2% digitonin, 25 mM gly- cylglycine, 100 mM NaCl, 5 mM EGTA, 400 nM N-acetylglucosamine, pH 7.6, for 3 h at 15 “C in the dark in the absence and presence of various adrenergic ligands. To reduce nonspecific labeling, a final concentration of 1 mM reduced glutathione was added to each reaction just prior to photolysis in order to scavenge any reactive intermediates that dissociate from the ligand binding site. Photolysis was carried out in quartz tubes by irradiation at 300 nM for 3 min at 4 “C in a Rayonet photochemical minireactor, as described previously (13, 24). Samples were quenched by the addition of SDS sample buffer and subjected to electrophoresis in 7.5-20% SDS-polyacrylamide gradient gels according to the method of Scherer et al. (25), except for the omission of urea. Following Coomassie Blue staining to identify the migration of molecular weight markers, the gel was dried and sub- jected to autoradiography.
Radioimmunoassay of CAMP Content in LLC-PKI Cells-LLC- PKl cells grown to confluence on Costar 24-well plates were serum- deprived for 18 h prior to assay. The a-modified Eagle’s medium in which they were grown was aspirated, replaced with MOPS-buffered saline (10 mM MOPS, 134 mM NaCl, 5 mM KCl, 2 mM CaCl,, 2 mM MgCl*, 5.6 mM glucose, 5 mM alanine, pH 7.4) containing 500 gM methylisobutylxanthine to serve as an inhibitor of CAMP-dependent phosphodiesterase and hand-swirled for 3 min on a Titretek warming plate, set at 37 “C. This buffer was then aspirated and replaced with fresh methylisobutylxanthine-containing MOPS-buffered saline to which was added no drugs (basal), or to which 10 FM forskolin or 4.6 pM arginine vasopressin (0.01 unit/ml) was added to stimulate CAMP accumulation. The ability of ru2-adrenergic receptors to attenuate CAMP accumulation was assessed by the simultaneous addition of 10 pM of UK14304, a selective a,-adrenergic receptor agonist. The cul- ture plate was then placed in a gently shaking waterbath at 37 “C for 10 min and the assay terminated by the addition of ice-cold trichlo- roacetic acid to a final concentration of 6.6%. Samples were then subjected to sequential alumina/Dowex chromatography to purify
sequence for the porcine cus-adrenergic receptor. The nucleotide se- quence is numbered from the ATG initiator codon. Potential sites of N-linked glycosylation are indicated by A, and the Thr within the consensus site for phosphorylation by CAMP-dependent protein ki- nase is marked by a 0.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cloning and Expression of cu2-Adrenergic Receptor Gene 17311
CAMP and subsequently assayed for CAMP content by radioimmuno- assay, as described previously (26).
RESULTS
Cloning and Sequencing of the Porcine wAdrenergic Recep- tor Gene-The 0.95-kb PstI fragment derived from the coding region of the human platelet Cl0 a*-adrenergic receptor gene (4, 6) was used to screen 1.2 x lo6 recombinants of a porcine genomic EMBL-3 library by plaque hybridization analysis. Of five positive clones isolated, restriction endonuclease map- ping and Southern blot analysis indicated that four were identical and contained a 5.5-kb KpnI fragment that hybrid- ized under conditions of high stringency (0.1 X SSC, 0.1% SDS, 65 “C) to the human ClO-an probe. Two of these clones were subcloned as described under “Experimental Proce- dures” and subjected to DNA sequence analysis. Fig. 1 outlines the strategy used for subcloning and DNA sequencing and presents the nucleotide and derived amino acid sequence for the porcine a2-adrenergic receptor gene.
The identity of the cloned gene was confirmed on the basis of protein sequence information and by comparison to the DNA and protein sequence published for the human platelet Cl0 cY2-receptor (4, 6). To obtain protein sequence, porcine brain cortex crz-adrenergic receptors were purified to homo- geneity and subjected to CNBr digestion. Of the peptides separated by reverse-phase HPLC, one yielded the following amino acid sequence; (Met)-Ile-Leu-Val-Tyr-Val-Arg-Ile- Tyr-Gln-Ile-Ala-Lys-Arg-Arg-Thr-Arg-Val-Pro-Pro. This se- quence corresponds exactly to residues 212-231 of the deduced amino acid sequence for the porcine as well as for the human Cl0 a2-receptor gene. As demonstrated for other members of the family of G-protein coupled receptors (27-29), including the human Cl0 a*-receptor, hydropathy analysis of the amino acid sequence of the porcine az-receptor indicates the presence of seven hydrophobic stretches of amino acids approximately 20-24 residues in length that probably represent transmem- brane-spanning domains.
A direct comparison of the porcine cup-amino acid sequence with that of the published human Cl0 a2-receptor revealed that both the porcine and human Cl0 genes (4, 6) code for proteins 450 amino acids in length. Striking homology ap- peared to exist between the receptors except within a short region (residues 332-370) contained in the putative third cytoplasmic loop. Within this region, the published human Cl0 a2-receptor sequence (4, 6) possesses a consensus site (residues 339-343) at Ser342 for CAMP-dependent protein kinase phosphorylation (30) that is not present in the porcine Lu2-receptor sequence. However, both receptor sequences con- tain a consensus site for CAMP-dependent protein kinase phosphorylation (residues 370-374) at Thr373. Alignment of the nucleotide sequences for the porcine cy2- and human Cl0 genes revealed that the porcine gene contained an insertion (C) at nucleotide 997 followed by a deletion after nucleotide 1089. These changes give rise to a “transient” shift in the reading frame and thus result in significant differences in the deduced amino acid sequences between the porcine as com- pared to the human receptor between nucleotides 997-1089. It should be noted that some difficulty in nucleotide sequenc- ing within this region was encountered, possibly due to the high GC content and thus increased probability for compres- sion and hairpin loop formation within the DNA sequence. Given the potential importance for differential regulation by CAMP-dependent protein kinase between the porcine and human receptors, appropriate subclones of the human (de- rived from the pUC18 a2AR clone available from the ATCC) and porcine genes were resequenced in both directions in order to verify the observed differences between the nucleotide
and derived amino acid sequences. The following notable differences in nucleotide sequence were detected between the previously published human ClO, the resequenced ATCC human ClO, and the porcine genomic cy2-receptor DNA se- quences within this region: Published human Cl0 970 AGCCAGGTGAAGCCGGGCGACAGCCTG Resequenced Cl0 970 AGCCAGGTGAAGCCGGGCGACAGCCTG Porcik
997 997 997
1042 1043 1043
1089 1089 1089
970 AGCCAGGTAAAGCCCGGGGACAGCCTG
CGCGGCGCGGGCCGGGGGCGACGGGGATCGGGACGCCGGCTGCAG CCGCGGCGCGGGCCGGGGGCGACGGGGATCGGGACGCCGGCTGCAG CCACGGCGCGGGCCGGGGGCGCCCGGGCCGGGGGCGCCCGCGACTG
GGCCGGGGGAGGAGCGCGTCGGGGCTGCCAAGGCGTCGCGCTGGCG GGCCGGGGGAGGAGCGCGTCGGGGCTGCCAAGGCGTCGCGCTGGCG GGGCCGGGGAGGAGCGCGGCGGGGTCGCCAAGGCGTCGCGCTGGCG
CGGGCGGGCAGAACCTCGAGAAGCGCTKACGTTCGTGCTGGCCGT CGGGC GGCAGAACCGCGAGAAGCGCTTCACGTTCGTGCTGGCCGT C GGAAGGCAGAACCGCGAGAAGCGCTTCACTTTCGTGCTGGCGGT
It should be noted that the observed change at nucleotide 997 (insertion of a C) in the human Cl0 sequence introduces a unique Sac11 restriction site into the gene. Restriction diges- tion of pUC18 a2AR Cl0 preparations with Sac11 (in combi- nation with several other diagnostic restriction endonucle- ases) generated fragments of appropriate size, further verify- ing the observed differences. Thus, the published sequence (4, 6) appears to be in error in this region. The derived amino acid sequence for the human Cl0 receptor that results from the nucleotide sequence that we obtain within this region displays greater overall homology to the porcine receptor (94% identity in amino acid sequence; 91% identity at the nucleo- tide level). Moreover, the previous consensus site (residues 339-343) identified as a potential substrate for CAMP-de- pendent protein kinase phosphorylation is absent, leaving Thr373 as the single consensus site common to both the human and porcine receptors. It recently has been demonstrated that the porcine a2-adrenergic receptor can be phosphorylated by the catalytic subunit of CAMP-dependent protein kinase and this phosphorylation can be blocked by a peptide containing the consensus phosphorylation site.* Studies are in progress to identify the phosphorylated residue.
Alignments of the deduced amino acid sequences for the porcine a2-adrenergic receptor and the human Cl0 and C4 a*-adrenergic receptor are presented in Fig. 2. The human & adrenergic receptor (31) and the porcine MP-cardiac musca- rinic receptor (32), which stimulate and inhibit adenylyl cy- clase activity, respectively, were included as representative members of the burgeoning family of G-protein-coupled receptors. As noted earlier, the porcine a2-adrenergic receptor shares the same general structure, consisting of an extracy- toplasmic amino terminus, followed by seven potential trans- membrane-spanning regions and a cytoplasmic carboxy tail, that has been described previously in detail by various inves- tigators (27, 29). Features of particular interest that are conserved in the porcine a2-receptor include two potential sites for N-linked glycosylation (Asn”’ and Asni4), aspartic acid residues present in transmembrane domains 2 and 3 (AspY and Aspu3) that have been postulated as being involved in adrenergic ligand binding, cysteine residues in exofacial loops 2 and 3 (Cys”’ and Cys’@‘) that potentially form a disulfide bridge, proline residues within each of the transmem- brane domains 4-7 (Pro167, Pro*“, Pro3*‘, and Pro423) that may play crucial roles in helix arrangement and transmem- brane signaling, and a cysteine in the cytoplasmic tail (CYSTS*) that could undergo fatty acyl esterification and thus serve to anchor the tail to the plasma membrane.
‘Wilson, A. L., Guyer, C. A., Cragoe, E. J., Jr., and Limbird, L. E. (1990) J. Biol. Chem. 265, 17318-17322.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
17312 Cloning and Expression of oyAdrenergic Receptor Gene
‘2 L c L 0 L P L 1 , 0 / Y ” /, ” ,/ I Y ” , 2 I/ I 2 i P c P li c 406
‘2 I 1 N 0 5 L \ 111
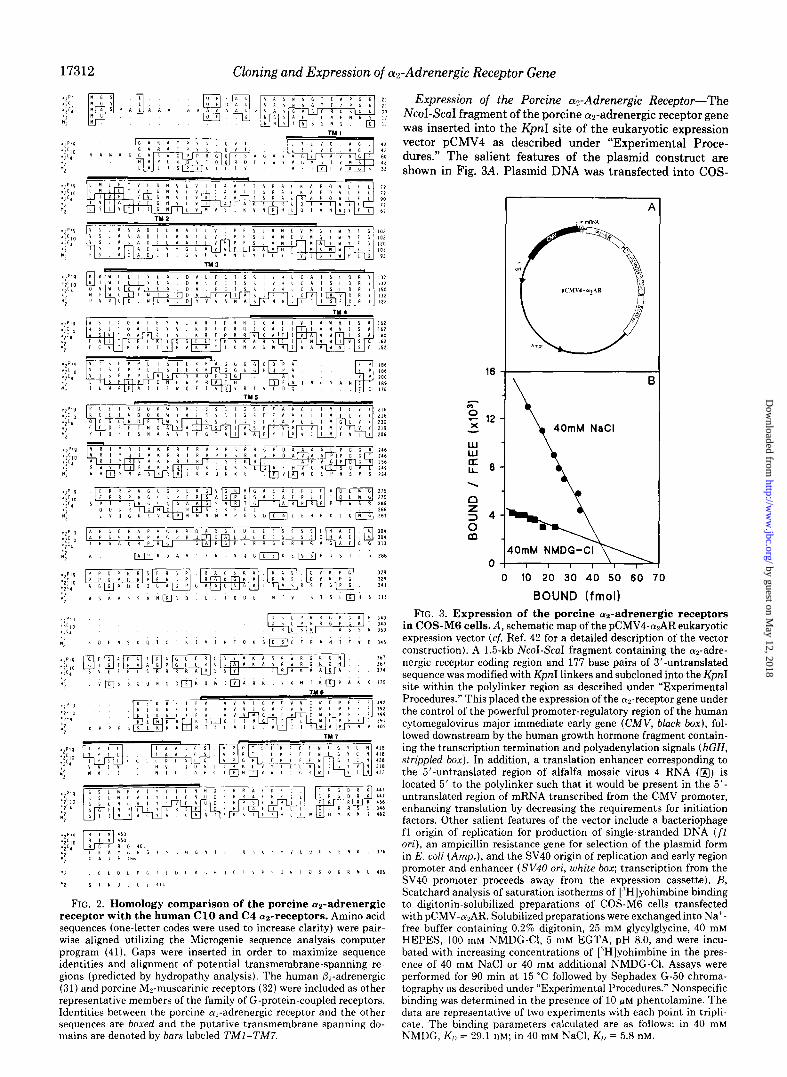
FIG. 2. Homology comparison of the porcine cxz-adrenergic receptor with the human Cl0 and C4 cuz-receptors. Amino acid sequences (one-letter codes were used to increase clarity) were pair- wise aligned utilizing the Microgenie sequence analysis computer program (41). Gaps were inserted in order to maximize sequence identities and alignment of potential transmembrane-spanning re- gions (predicted by hydropathy analysis). The human &adrenergic (31) and porcine M,-muscarinic receptors (32) were included as other representative members of the family of G-protein-coupled receptors. Identities between the porcine cus-adrenergic receptor and the other sequences are boxed and the putative transmembrane spanning do- mains are denoted by bars labeled 7’MI-TM7.
Expression of the Porcine a2-Adrenergic Receptor-The NcoI-ScaI fragment of the porcine a2-adrenergic receptor gene was inserted into the KpnI site of the eukaryotic expression vector pCMV4 as described under “Experimental Proce- dures.” The salient features of the plasmid construct are shown in Fig. 3A. Plasmid DNA was transfected into COS-
40mM NaCl
.
0 0 10 20 30 40 50 60 70
BOUND (fmol) FIG. 3. Expression of the porcine az-adrenergic receptors
in COS-MG cells. A, schematic map of the pCMV4-WAR eukaryotic expression vector (cf. Ref. 42 for a detailed description of the vector construction). A 1.5-kb NcoI-ScaI fragment containing the cuz-adre- nergic receptor coding region and 177 base pairs of 3’-untranslated sequence was modified with KpnI linkers and subcloned into the KpnI site within the polylinker region as described under “Experimental Procedures.” This placed the expression of the az-receptor gene under the control of the powerful promoter-regulatory region of the human cytomegalovirus major immediate early gene (CMV, black box), fol- lowed downstream by the human growth hormone fragment contain- ing the transcription termination and polyadenylation signals (hGH, strippled box). In addition, a translation enhancer corresponding to the 5’-untranslated region of alfalfa mosaic virus 4 RNA (@) is located 5’ to the polylinker such that it would be present in the 5’- untranslated region of mRNA transcribed from the CMV promoter, enhancing translation by decreasing the requirements for initiation factors. Other salient features of the vector include a bacteriophage fl origin of replication for production of single-stranded DNA (f1 ori), an ampicillin resistance gene for selection of the plasmid form in E. coli (Amp,), and the SV40 origin of replication and early region promoter and enhancer (S V40 ori, white box; transcription from the SV40 promoter proceeds away from the expression cassette). B, Scatchard analysis of saturation isotherms of [“Hlyohimbine binding to digitonin-solubilized preparations of COS-M6 cells transfected with pCMV-n2AR. Solubilized preparations were exchanged into Na’- free buffer containing 0.2% digitonin, 25 mM glycylglycine, 40 mM HEPES, 100 mM NMDG-Cl, 5 mM EGTA, pH 8.0, and were incu- bated with increasing concentrations of [“Hlyohimbine in the pres- ence of 40 mM NaCl or 40 mM additional NMDG-Cl. Assays were performed for 90 min at 15 “C followed by Sephadex G-50 chroma- tography as described under “Experimental Procedures.” Nonspecific binding was determined in the presence of 10 GM phentolamine. The data are representative of two experiments with each point in tripli- cate. The binding parameters calculated are as follows: in 40 mM NMDG, K,, = 29.1 nM; in 40 mM NaCl, KI, = 5.8 nM.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cloning and Expression of cwAdrenergic Receptor Gene 17313
M6 cells by the DEAE-dextran method (21). Membranes prepared 48-72 h after transfection and assayed for the pres- ence of a?-adrenergic receptors displayed saturable binding of the ap-adrenergic antagonist [3H]yohimbine. Membranes pre- pared from COS-M6 cells transfected with either salmon sperm DNA or pCMV4 lacking porcine az-gene insert dis- played no specific [“Hlyohimbine binding (data not shown). Digitonin-solubilized preparations of transfected COS-M6 cells were utilized in subsequent saturation analysis experi- ments to determine the dissociation constant of the expressed a2-receptor for yohimbine and to initially investigate the effect of Na+ on the cuz-receptor’s affinity for antagonist. Solubilization of the receptor eliminated any effects that nonuniform distribution of the expressed receptors might exert on binding measurements as previously described under “Experimental Procedures.” Scatchard analysis of saturation binding of [“Hlyohimbine in the presence and absence of Na’ is presented in Fig. 3B. The specific [3H]yohimbine binding yielded a calculated KD of 5.8 nM in the presence of Na’. In the absence of Na+, however, the expressed oc2-receptor dis- plays a decreased affinity for [3H]yohimbine with a calculated KD of 29.1 nM. These results are comparable to findings obtained for binding of [3H]yohimbine to purified cut-receptor preparations from porcine brain cortex (10, 13) or human platelets (9). Porcine cyn-adrenergic receptors were expressed in COS-M6 cells at a density of 26 pmol/mg membrane protein, which is over 300-fold higher than that observed for particulate fractions from porcine brain cortex. Since it is likely that only 5-10% of COS-M6 cells express the gene during transient expression assays, then receptor expression in those COS-M6 cells expressing the a2-receptor gene must be 3000-6000-fold enriched over typical expression in target membranes. Similar binding parameters were obtained for the transient expression of the human Cl0 an-receptor in COS-7 cells (4-6).
Competition by antagonists and agonists for binding of [“Hlyohimbine to membranes prepared from pCMV4-a2AR transfected COS-M6 cells is illustrated in Fig. 4. The rank order of potency for antagonist competition (Fig. 4A) was yohimbine (EC& = 7.5 nM) > idazoxan (EC& = 56 nM) >
phenotamine (EC&, = 86 nM) >> prazosin (EC& = 7000 nM)
>> propranolol, which parallels that observed for particulate and solubilized porcine Lu,-receptor preparations (13). Fig. 4B shows competition curve data for agonist binding with the LY*- selective agonist oxymetazoline displaying the greatest po- tency (EC&, = 35 nM) followed by UK-14304 (EC& = 250 nM) > (-)-epinephrine (EC& = 1,700 nM)) > (-)-norepinephrine (EC,” = 10,000 nM) > (+)-epinephrine (ECsO = 30,000 nM) >> the P-adrenergic selective agonist isoproterenol. These affin- ities are characteristic of the porcine brain a*-receptor and are similar to those reported for the cloned human Cl0 (Ye- receptor expressed in COS-7 cells (4,5) or CHO cells (6). The order of potency for antagonists and agonists, including the stereoselective preference for (-)-epinephrine versus (+)-ep- inephrine, indicates that the porcine gene expressed in COS- M6 cells indeed codes for a receptor of the anA-subtype, analogous to the purified porcine brain and human platelet Cl0 receptors.
An initial biochemical characterization of the expressed porcine cu,-receptor further confirmed the identification of the receptor subtype as (YEA. Partial purification of the expressed n2-receptor from digitonin-solubilized preparations using WGA-agarose chromatography indicates that the receptor is glycosylated. The a2-adrenergic-selective photoaffinity ligand, ‘2”I-Rau-AzPEC, labels a protein that migrates in SDS-poly- acrylamide gels with an apparent M, = 71,000 (Fig. 5), which is significantly greater than the mass expected on the basis of
140
s 2 120
m w 100 I
$ 60
z T 60 CL 0 40 iz G 2 20 (0 w 0
v r , I + *: I f 9 6 7 6 5 4 3
- LOG [ ANTAGONIST]
-B
9 6 7 6 5 4 3
-LOG [AGONIST]
FIG. 4. Antagonist and agonist binding to porcine cYz-adre- nergic receptor expressed in COS-M6 cells. Membranes pre- pared from transfected CO&M6 cells were incubated with 7.5 nM [“Hlyohimbine and the indicated concentrations of competing unla- beled ligand in assay buffer containing 40 mM NaCl for 30 min at 25 “C as described under “Experimental Procedures.” A, competition by antagonists: n , yohimbine; A, idazoxan; 0, phentolamine; A, prazosin; 0, propranolol. B, competition by agonists: A, oxymetazo- line; A, UK14304; 0, (-)-epinephrine; 0, norepinephrine; n , (+)- epinephrine; 0, isoproterenol. Total specific [“Hlyohimbine binding in the absence of competing ligand was 10,000 cpm/incubation (-125 fmol). Data points are the mean + S.E. from at least five separate experiments with each point performed in duplicate.
the derived amino acid sequence (48,981 Da) implying further that the protein is glycosylated, a property characteristic of (Y?A but not ol,B receptor subtypes (33).2 The protein band labeled corresponds to the expressed receptor since labeling by ‘Y-Rau-AzPEC is totally eliminated by yohimbine but is not significantly effected by the al-antagonist prazosin. More- over, the stereoselectivity with which epinephrine is able to compete with the photoaffinity ligand for labeling further indicates that ““I-Rau-AzPEC is identifying a physiologically relevant adrenergic receptor.
One property of cu2-adrenergic receptor-ligand binding in- teractions is the modulation of receptor affinity for agonists by guanine nucleotides as a result of receptor coupling to G- proteins. One manifestation of this modulation of receptor- agonist interactions by guanine nucleotides is a shift to the right in the concentration-response curves of agonist compe- tition for radiolabeled antagonist binding, due to the decrease in receptor affinity for agonist (34). When the ability of guanine nucleotides to modulate agonist binding to mem- branes prepared from pCMV4-a2AR-transfected COS-M6
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17314 Cloning and Expression of cwAdrenergic Receptor Gene
71 kDa
Fro. 5. Photoaffinity labeling of expressed porcine a2-ad- renergic receptors. Expressed porcine cu?-receptors were partially purified from digitonin-solubilized preparations of transfected COS- M6 cells by WGA-agarose chromatography. Samples containing ap- proximately 0.1 pmol of cu?-receptor were photolabeled with 2 nM ““I- Rau-AzPEC in the absence (CONTROL) of competing ligand or in the presence of 1 X lo-” M yohimbine, 1 X 10-fi M prazosin, 5 X lo-” M (-)-epinephrine (EPI), or 5 X lo-” M (+)-epinephrine as described under “Experimental Procedures.”
cells was investigated, no apparent decrease in receptor affin- ity for epinephrine was observed (data not shown). Given the overexpression of the a?-receptor with respect to relevant G- proteins in the small percentage of COS-M6 cells which are transiently expressing the receptor gene, it is not surprising that cr2-adrenergic receptor-G-protein coupling could not be detected. Similar observations have been reported for the & adrenergic receptor transiently expressed in COS-7 cells (35, 36).
Given that Luz-adrenergic receptor-G-protein coupling could not be detected in COS-M6 cells, the ability of expressed aa- adrenergic receptors to attenuate CAMP accumulation was not assessed in this system. In order to determine whether the cloned porcine cus-receptor can regulate adenylyl cyclase activity, LLC-PKl cells which lack endogenous cu,-receptors were transfected using calcium phosphate precipitation with pCMV4-CQAR. Stable clones were selected for their ability to bind the cup-agonist [““Iliodo-para-aminoclonidine. Satura- tion analysis of [“Hlyohimbine binding to membranes from clonal cells (denoted SC-O) yielded a K,, of 5.2 nM (in the presence of Na’) for Lu,-receptors expressed at a density of 109 pmol/mg protein (data not shown). The expressed ~(a- receptors displayed modulation by guanine nucleotides as indicated by the ability of GppNHp to decrease high affinity [‘““Iliodo-para-aminoclonidine binding (data not shown) as previously described for a?-receptors from human platelets (23). To examine whether Luz-receptors expressed in LLC- PKl SC-O cells mediate inhibition of adenylyl cyclase activ- ity, the effect of the cu+pecific agonist, UK14304 on either arginine vasopressin- or forskolin-stimulated CAMP accu- mulation was measured. Arginine vasopressin increased intra- cellular CAMP levels to 1163 + 160 pmol of CAMP/well, approximately loo-fold higher compared to basal (97 + 33 pmol of CAMP/well). In the presence of 10 pM UKl4304, however, the CAMP levels produced in response to arginine vasopressin stimulation were reduced by 53% (557 + 71 pmol of CAMP/well). Similar results were observed for cells stim- ulated with forskolin (10 FM). Compared to basal (7 + 2 pmol of CAMP/well), forskolin caused a 30-fold increase (237 + 16 pmol of CAMP/well) in CAMP formation which was inhibited
A 1 Na,Cl added
0 30 60 90
TIME, MINUTES
bp 20 I I I I 0 10 20 30 40
SALT , mM
FIG. 6. Allosteric modulation of [3H]yohimbine binding by monovalent cations. Digitonin-solubilized cu2-adrenergic receptors from pCMV4-turAR transfected COS-M6 cells were exchanged into a Na+-free buffer containing 0.2% digitonin, 40 mM HEPES, 25 mM glycylglycine, 100 mM NMDG, 5 mM EGTA, pH 8.0. Association of 7.5 nM [ ‘Hlyohimbine with tu-receptors was accomplished by incu- bation at 15 “C for 60 min followed by 10 “C for 30 min. An aliquot was removed for determination of total and nonspecific binding. Nonspecific binding was defined as that obtained in the presence of 10 pM phentolamine and was never greater than 10% of total binding. Specific binding at time 0 was 2500 cpm/incubation. Unlabeled yohimbine was added at time 0 to a final concentration of 50 pM in order to initiate monitoring of the dissociation phase. At 15 min, an aliquot (500 ~1) was withdrawn, then monovalent cation was added to the incubation to evaluate its effect on the rate of [“Hlyohimbine dissociation. At the indicated times aliquots (500 ~1) of the reaction mixes were removed and bound radioligand was separated from free [.‘H]yohimbine by Sephadex G-50 chromatography (9). A, Na’ allo- sterically modulates (“Hlyohimbine binding: dissociation in the ab- sence (0, equiosmolar NMDG-Cl present) or the presence of 100 mM NaCl (A). The data shown are from one experiment representative of two independent experiments performed in triplicate. B, monova- lent cation selectivity for allosteric modulation of [“Hlyohimbine binding occurs with the order Na’ > Li’ > K’. The amount of [‘HI yohimbine remaining bound 60 min after the initiation of the disso- ciation phase was measured in the presence of increasing concentra- tions of the NaCl (A), LiCl (m), or KC1 (O), maintaining the total salt concentration at 100 mM using NMDG-Cl. The data shown are representative of one of two experiments performed in triplicate. The data are normalized to the percent [“Hlyohimbine dissociation ob- tained in the presence of 100 mM NaCl (100%).
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cloning and Expression of a2-Adrenergic Receptor Gene 17315
by 57% (101 f 28 pmol of CAMP/well) in the presence of 10 PM UK14304. Forskolin stimulation of untransfected LLC- PKl cells yielded similar levels of intracellular CAMP (227 f 32 pmol/well; basal = 13 f 7 pmol/well). However, UK14304 had no effect on forskolin-stimulated CAMP levels (234 f 11 pmol/well) in untransfected cells. These results imply that the porcine as-receptor when expressed in LLC-PKl cells can couple functionally to inhibition of adenylyl cyclase.
Allosteric Modulation of Receptor-Ligand Interactions- Previous studies have shown that Na+, H+, and &amino- substituted analogs of amiloride allosterically modulate li- gand-binding interactions in digitonin-solubilized and homo- geneous preparations of a2-receptors from porcine brain cor- tex (10). The allosteric effects of Na+, H’, and amiloride analogs on digitonin-solubilized a*-receptor preparations from pCMV4-cr2AR-transfected COS-M6 cells were investi- gated by analyzing the influence of each of these agents on the rate of dissociation of [3H]yohimbine from the solubilized receptor preparations. In these studies, [3H]yohimbine was incubated with the solubilized receptors until steady state binding was reached. To monitor the dissociation phase, unlabeled yohimbine was added to a final concentration of 50 PM to prevent the rebinding of dissociated [3H]yohimbine and to permit full occupancy of available adrenergic ligand-bind- ing sites. Later addition of more unlabeled yohimbine does not further accelerate the apparent rate of [3H]yohimbine dissociation. Thus, under these experimental conditions, all adrenergic sites are occupied and any effects of Na+, H’, or amiloride analogs on the rate of [3H]yohimbine dissociation would arise from interaction at sites other than the adrenergic ligand-binding site. Fig. 6A demonstrates that Na’ accelerates the rate of [3H]yohimbine dissociation from digitonin-solu- bilized as-receptor preparations from transfected COS-M6 cells. As noted previously, however, Na+ increases Lu2-receptor affinity for [3H]yohimbine. Although the ability of Na’ to lower the Ku for yohimbine may seem in opposition to the ability of Na’ to accelerate yohimbine dissociation, earlier studies (9) have indicated that the effect of Na’ on dissocia-
H+ added
&Q 20 1
0 30 60 90
TIME, MINUTES
FIG. 7. Protons accelerate the rate of [3H]yohimbine disso- ciation from expressed porcine cY*-adrenergic receptors. The rate of 7.5 nM ]“H]yohimbine dissociation from digitonin solubilized preparations of pdMV4-cusAR transfected COSM6 cells was moni- tored as described in the legend to Fig. 6. Binding at time 0 (defined as 100%) represents 2500 cpm specific binding. Dissociation was performed in Na’-free buffer at pH 8.0 (0) or pH 6.5 (0, sufficient 2 N HCl added at the time indicated by the arrow to reduce the pH to 6.5). The data are from one experiment representative of two inde- pendent experiments with each point performed in triplicate.
tion is offset by a greater than 3-fold increase in the rate of association, resulting in an overall decrease in the KD for yohimbine. Fig. 6B illustrates that the monovalent cation selectivity for acceleration of [3H]yohimbine dissociation was Na’ > Li’ > K’. This selectivity for monovalent cations is characteristic of that observed for digitonin-solubilized prep- arations of ocp-receptors from porcine brain cortex (10). In addition to effects on antagonist binding, Na+ caused a 5-9- fold decrease (n = 3) in the affinity of cut-adrenergic receptor from transfected COS-M6 cells for the crz-agonist, (-)-epi- nephrine (data not shown). These results are in good agree- ment with previous observations obtained using a2-receptor preparations from porcine brain cortex (10).
The ability of H’ to modulate ligand-binding interactions of cu*-receptors solubilized from transfected COS-M6 cells is demonstrated in Fig. 7. The rate of [3H]yohimbine dissocia- tion at pH 6.5 was accelerated as compared with the rate observed at pH 8.0 indicating that protons as well as Na+ allosterically modulate receptor-ligand binding.
Previous results on digitonin-solubilized and homogeneous preparations of Lup-receptors from porcine brain cortex have demonstrated that amiloride analogs substituted in the 5- amino position allosterically modulate oc2-receptor-ligand in- teractions with greater effectiveness than analogs modified in the guanidino position (lo), which parallels their ability to block Na’/H’ exchange in other systems (37). As shown in Fig. 8, the 5-amino-substituted amiloride analogs ethylisopro- pylamiloride and methylisobutylamiloride are able to accel- erate the rate of [3H]yohimbine dissociation from solubilized porcine o12-receptors expressed in COS-M6 cells. Conversely, dichlorobenzamil and chlorobenzyldimethylbenzamil, which are guanidino-substituted analogs of amiloride, when present at the same concentration during dissociation, do not signif-
analog or additional unlabeled YOH added
5-NH,-substituted:
0 MIA l EIA -. -.
guanidino-substituted: -.
‘.
A CBDMB ‘*
A DCB
I I I I 0 30 60 90 120
TIME. MINUTES
FIG. 8. The influence of amiloride analogs on transfected cr2-adrenergic receptor-ligand interactions. Digitonin-solubi- lized alp-receptors from pCMV4-cu,AR transfected COS-M6 cells were exchanged into Na’-free buffer, pH 8.0 (cf. “Experimental Proce- dures”) and the effect of various amiloride analogs on the rate of [“HI yohimbine dissociation was monitored as described in the legend to Fig. 5. Binding at time 0 (defined as 100%) was 2500 cpm specific binding. Additional yohimbine (50 FM, control, 0), 100 FM chloroben- zyldimethylbenzamil (CBDMB, A), 100 NM dichlorobenzamil (DCB, i), 100 /1M ethylisopropyl amiloride (HA, l ), or 100 PM methyl isobutvlamiloride (MIA. 0) were added 15 min after the addition of 50 PM-yohimbine at time 0. Compared to the control, the 5-amino- substituted amiloride analogs (ethylisopropylamiloride, methyl iso- butylamiloride), but not the guanidino-substituted analogs (chloro- benzyldimethylbenzamil, dichlorobenzamil) are able to accelerate the rate of [“Hlyohimbine ( YOH) dissociation from on-receptors. These data are from a single experiment repeated at least three times in triplicate.
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17316 Cloning and Expression of a2-Adrenergic Receptor Gene
icantly alter the rate of [“Hlyohimbine dissociation. These results indicate that the ligand-binding interactions of the cloned porcine a2-adrenergic receptor are preferentially ef- fected at an allosteric site by 5-amino-substituted amiloride analogs.
DISCUSSION
The existence of a number of a2-adrenergic receptor sub- types previously has been suggested on the basis of pharma- cological (3) and biochemical evidence (5, 13, 33, 35). Recent publications have described the isolation and expression of three DNA clones encoding a2-adrenergic receptors that can be distinguished by radioligand binding and by physical prop- erties. The human genomic clone, designated ClO-L-Q, encodes a protein of the CY~A subtype (4, 6). A cDNA clone isolated from a rat kidney library (7) has been shown to code for the LQB subtype, whereas a cDNA clone from a human kidney library encodes a protein which possesses the prazosin sensi- tivity characteristic of an o(*B subtype but, unlike the LY*B receptor subtype (7,33) possesses sites for N-linked glycosyl- ation (5).
The present studies report the genomic cloning of a gene encoding a porcine olz-adrenergic receptor. Expression of the clone in COS-M6 cells yields a gene product that displays the selectivity for binding agonists and antagonists that is char- acteristic of an o12-receptor of the (YEA subtype. In fact, the protein sequence obtained for the porcine brain cut-receptor which verified that the clone coded for the oc2-receptor occurs in a region that uniquely identifies each receptor subtype. Upon comparison, the porcine amino acid sequence is iden- tical to that of the CV*A human platelet Cl0 receptor in this region (amino acids 212-231). The glycosylation of the porcine a*-receptor gene product also is consistent with its classifi- cation as an asA receptor. Evidence for glycosylation of the expressed a2-receptor includes not only the adsorption to and elution from WGA-agarose of the digitonin-solubilized recep- tor but also the migration of the photoaffinity-labeled receptor in SDS-polyacrylamide gels with an apparent mass of 71,000 Da. This apparent mass is greater than that expected on the basis of the predicted amino acid sequence (48,981 Da) imply- ing that the expressed LQA receptor subtype, like the cr2- receptor obtained from porcine brain cortex, is glycosylated.
One of the goals of these studies following expression of the porcine a2-receptor was to investigate the effects of Na+, H’, and 5-amino-substituted analogs of amiloride on receptor- ligand interactions. For technical reasons, COS-M6 mem- branes containing expressed az-receptors were solubilized in digitonin in order to evaluate the allosteric modulation of ligand binding by these agents. The retention of the effects of Na+, H’, and 5-amino-substituted amiloride analogs on the rate of [“Hlyohimbine dissociation from the expressed (Ye- receptor was not entirely unexpected, since homogeneous preparations of porcine a,-receptors also exhibit allosteric modulation by these effecters. These data are thus consistent with the interpretation that the receptor itself possesses al- losteric binding sites for Na+, H’, and amiloride analogs. If these effects were due instead to coupling of the receptor to another protein, as in the case of guanine nucleotide regula- tion of a,-receptor-ligand interactions (39, 40), then the ef- fects quite likely would go undetected for the following rea- sons: 1) the detergent uncouples the receptor from the effector protein that confers allosteric modulation of binding to the receptor, or 2) the overexpression of the receptor in a small percentage of the total COS-M6 cell population would serve to severely limit or eliminate detection of allosteric-modula- tion by endogenous effector proteins. Instead, the effects of
Na+, H’ and amiloride analogs are quantitatively similar to those obtained from homogeneous preparations derived from porcine brain cortex, implying that the allosteric sites are in the receptor molecule itself. Site-directed mutagenesis studies on the gene encoding the porcine brain a*-receptor in concert with complementary biochemical studies using purified por- cine brain ap-receptor preparations will delineate further the molecular basis for the effects of Na+, H+ and B-amino- substituted amiloride analogs on receptor-ligand interactions and the role this allosteric modulation plays in receptor- mediated physiological functions.
Acknowledgments-We are grateful to Dr. Tom Lucas for his direction regarding protein sequencing and HPLC analysis, to Paul Matresian for his advice and efforts in obtaining the porcine peptide sequence, and to Dr. D. Martin Watterson for use of his Applied Biosystems HPLC and peptide sequencer. We would like to thank Dr. Claire Fraser for her generous gift of the human cyz-adrenergic receptor Cl0 clone used for library screening. We would like to express our appreciation to Dr. David Russell for his patient tutelage during L. E. L.‘s sabbatical in his laboratory and for providing us with the pCMV4 eukaryotic expression vector. We wish to thank Drs. Colin Funk, Steven Lloyd, and Brian Kobilka for their expert advice regarding the cloning of the porcine a2-receptor and to thank Drs. Mark Magnuson and Steven Lanier for their helpful suggestions concerning the expression of the cloned gene. We are grateful to Dr. Richard O’Brien for his invaluable assistance in establishing the stable clones and to Pat Adams for his careful maintenance of these clonal cell lines. We want to thank Dr. Jack Wells for use of his Rayonet photochemical mini-reactor. We are very appreciative of the technical assistance and encouragement given by Carol Ann Bonner and Suzanne Brandon. We also would like to acknowledge Mary Couey for her conscientious preparation of this manuscript.
REFERENCES
1. 2. 3. 4.
5.
6.
7.
8.
9.
10.
11. 12.
13.
14.
15. 16.
17.
18.
19. 20.
Ross, E. M. (1989) Neuron 3, 141-152 Limbird, L. E. (1988) FASEB J. 2, 2686-2695 Bvlund. D. B. (1988) Trends Pharmacol. Sci. 9. 356-361 Kbbilka, B. K:, Matsui, H., Kobilka, T. S., Yang-Feng, T. L.,
Francke, U., Caron, M. G., Lefkowitz, R. J., and Regan, J. W. (1987) Science 238.650-656
Regan, J. W., Kobilka, T. S., Yang-Feng, T. L., Caron, M. G., Lefkowitz. R. J.. and Kobilka. B. K. (1988) Proc. Natl. Acad. Sci. U s. i.85;6301-6305 ’
Fraser, C. M., Arakawa, S., McCombie, W. R., and Venter, J. C. (1989) J. Biol. Chem. 264, 11754-11761
Zeng, D., Harrison, J. K., D’Angelo, D. D., Barber, C. M., Tucker, A. L., Lee, Z., and Lynch, K. R. (1990) Proc. Natl. Acod. Sci. U. S. A. 87,3102-3106
Tsai, B. S., and Lefkowitz, R. J. (1978) Mol. Pharmacol. 14,540- 548
Limbird, L. E., Speck, J. L., and Smith, S. K. (1982) Mol. Pharmacol. 21,609-617
Nunnari, J. M., Repaske, M. G., Brandon, S., Cragoe, E. J., Jr., and Limbird, L. E. (1987) J. Biol. Chem. 262.12387-12392
Jacobs, K.-H. (1979) Mol. Cell. Endocrinol. 16, 147-156 Minuth. M.. and Jacobs. K.-H. (1986) Nuunvn-Schmeideberg’s
Arch.‘Pharmacol. 333,’ 124-129 ” Repaske, M. G., Nunnari, J. M., and Limbird, L. E. (1987) J.
Biol. Chem. 262,12381-12386 Lanier, S. M., Hess, H. J., Grodski, A., and Graham, R. M. (1986)
Mol. Pharmacol. 29. 219-227 Laemmli, U. K. (1970) Nature 227,680-685 Hunkanillar. M. W.. Luian. E.. Ostrander. F.. and Hood. L. E.
(1983)Methods Ekzy~ol.‘91; 227-236 Allen, G. (1983) in Laboratory Techniques in Biochemistry and
Molecular Biology: Sequencing of Proteins and Peptides (Work, T. S., and Burdon, R. H., eds) pp. 62-65, Elsevier, Amsterdam
Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Surina Harbor. NY
Geck, P.,-andNasz, 1.‘(1983) Anal. Biochem. 135, 264-268 Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.
Acad. Sci. U. S. A. 74, 5463-5467
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Cloning and Expression of cwddrenergic Receptor Gene 17317
21. Gorman, C. (1986) DNA Cloning: A Pratical Approach (Glover, D. M., ed) Vol. II, pp. 143-164, IRL Press, Oxford
22. Chen, C., and Okayama, H. (1987) Mol. Cell. Biol. 7,2745-2752 23. Van Dort, M., Neubig, R., and Counsell, R. E. (1987) J. Med.
Chem. 30, 1241-1244 24. Lanier, S. M., Graham, R. M., Hess, H. J., Grodski, A., Repaske,
M. G., Nunnari, J. M., Limbird, L. E., and Homcv, C. J. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,93&8-9362 -
25. Scherer. N. M.. Toro. M.-J.. Entman. M. L.. and Birnbaumer. L. (1987) Arch. ‘Biochem. Biiphys. 259, 431-440
26. Steiner, A. L., Kipnis, D. M., Utiger, R., and Parker, C. (1969) Proc. Natl. Acad. Sci. U. S. A. 64, 367-373
27. Dohlman, H. G., Caron, M. G., and Lefkowitz, R. J. (1987) Biochemistry 26, 2657-2664
28. Aanleburv. M. L.. and Harzrave. P. A. (1986) Vision Res. 26. ‘i881-1895
I
29. Liao, C.-F., Themmen, A. P. N., Joho, R., Barberis, C., Birn- baumer, M., and Birnbaumer, L. (1989) J. Biol. Chem. 264, 7328-7337
30. Kemp, B. E., Graves, D. J., Benjamini, E., and Krebs, E. G. (1977) J. Biol. Chem. 252,4888-4894
U. S. A. 84, 46-50 32. Peralta, E. G., Winslow, J. W., Peterson, G. L., Smith, D. H.,
Ashkenazi, A., Ramachandran, J., Schimerlik, M. I., and Ca- pon, D. J. (1987) Science 236,600-605
33. Lanier, S. M., Homey, C. J., Patenaude, C., and Graham, R. M. (1988) J. Biol. Chem. 263,14491-14496
34. Gilman, A. (1987) Annu. Rev. Biochem. 56.615649 35. Dixon, R. A. F., Sigal, I. S., Rands, E., Register, R. B., Candelore,
M. R., Blake, A. D., and Strader, C. D. (1987) Nature 326, 73- 77
36. Strader, C. D., Dixon, R. A. F., Cheung, A. H., Candelore, M. R., Blake, A. D., and Sigal, I. S. (1987) J. Biol. Chem. 262, 16439- 16443
37. Vigne, P., Frelin, C., Cragoe, E. J., Jr., and Lazdunski, M. (1984) Mol. Pharmacol. 25, 131-136
38. Regan, J. W., Nakata, H., DeMarinis, R. M., Caron, M. G., and Lefkowitz, R. J. (1986) J. Biol. Chem. 261, 3894-3900
39. Michel, T., Hoffman, B. B., Lefkowitz, R. J., and Caron, M. G. (1981) Biophys. Biochem. Res. Commun. 100, 1131-1136
40. Smith, S. K., and Limbird, L. E. (1982) Proc. Natl. Acod Sci. U. S. A. 78, 4026-4030
31. Kobilka, B. K., Dixon, R. A. F., Frielle, T., Dohlman, H. G., 41. Queen, C., and Korn, L. J. (1984) Nucleic Acids Res. 12,581-599 Bolanowski, M. A., Sigal, I. S., Yang-Feng, T. L., Francke, U., 42. Andersson, S., Davis, D. L., Dahlback, H., Jornvall, H., and Caron, M. G., and Lefkowitz, R. J. (1987) Proc. N&l. Acad. Sci. Russell, D. W. (1989) J. Biol. Chem. 264,8222-8229
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

C A Guyer, D A Horstman, A L Wilson, J D Clark, E J Cragoe, Jr and L E Limbird2-adrenergic receptor. Allosteric modulation by Na+, H+, and amiloride analogs.
Cloning, sequencing, and expression of the gene encoding the porcine alpha
1990, 265:17307-17317.J. Biol. Chem.
http://www.jbc.org/content/265/28/17307Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/28/17307.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















