
Cloning, expression and applicability of thermo-alkali-stable xylanase of Geobacillus...
6
Cloning, expression and applicability of thermo-alkali-stable xylanase of Geobacillus thermoleovorans in generating xylooligosaccharides from agro-residues Digvijay Verma, T. Satyanarayana ⇑ Department of Microbiology, University of Delhi South Campus, New Delhi 110 021, India article info Article history: Received 10 November 2011 Received in revised form 11 December 2011 Accepted 12 December 2011 Available online 21 December 2011 Keywords: Thermo-alkali-stable xylanase Recombinant xyl-gt Geobacillus thermoleovorans Xylooligosaccharides Pre-bleaching of paper pulp abstract A xylanase gene (xyl-gt) of 1.224 kbp was cloned from the extremely thermophilic bacterium Geobacillus thermoleovorans that encodes a protein containing 408 amino acid residues. Eight conserved regions (sig- nature sequences) of GH family 10 xylanases have been found in the xylanase. When the xylanase gene was cloned and expressed in Escherichia coli BL21 (DE3), the recombinant strain produced xylanase titer of 270 U mg 1 which is 27-fold higher than the wild strain. It is optimally active at 80 °C and pH 8.5 with a high thermostability over broad range of pH (6–12) and temperature (40–100 °C). The end products of the hydrolysis of birch wood xylan and agro-residues included xylobiose, xylotriose, xylotetraose and xylopentaose. The xylanase of G. thermoleovorans is one of the rare xylanases that exhibits thermo- alkali-stability, and thus, it is a suitable candidate for pre-bleaching of paper pulps and generating xylool- igosaccharides from agro-residues for use as prebiotics. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Xylans are hetero- and homo-polysaccharides which are main constituents of hemicellulosic biomass of terrestrial plant cell wall matrix and second most abundant polysaccharide on earth. Its core comprises b-1,4 linked xylosyl residues, and the attachment of this core chain with various groups (4-O-methyl-D-glucuronosyl, a-arabinofuranosylresidues and others) makes it a heteropolysac- charide. Consequently the complete hydrolysis of xylan requires a group of enzymes including endo-b-D-xylanase, b-xylosidase, arabinofuranosidase, glucuronidase, acetylxylan esterase, and feru- lic acid and p-coumaric acid esterases. Among these, xylanases have attracted considerable attention due to their catalytic activity for releasing lower xylooligosaccharides, and their potential appli- cation in textile, feed and biofuel industries. Xylanases attracted attention further after Viikari et al. (1986) reported their applica- bility in bleaching of pulp in the paper industry. The inclusion of xylanase treatment step reduces the chlorine consumption up to 25–35% in bleaching kraft pulp to make the process environment friendly. Although several xylanases have been reported from a plethora of microorganisms, most of them do not tolerate the harsh conditions employed in pulp processing. This process requires thermo-alkali-stable and cellulase-free xylanase, because these processes are usually operated at higher temperature and alkaline conditions. An alkali-thermo-stable and cellulase-free xylanase suitable for pre-bleaching of paper pulps has been reported from Geobacillus thermoleovorans (Sharma et al., 2007). The xylanase produced by the bacterium exhibits optimum activity at 80 °C and pH 8.5. The cloning and expression of xylanase encoding gene (xyl-gt) of G. thermoleovorans is crucial for enhancing the enzyme production in order to make its production cost effective. In this investigation, cloning and expression of xylanase gene (xyl-gt) from G. thermo- leovorans and its applicability in saccharifying agro-residues to produce xylooligosaccharides (XOs) are presented. 2. Methods 2.1. Bacterial strains and plasmids G. thermoleovorans was isolated from the pulp samples collected from Century Paper Mills, Lal Kuan, Uttaranchal (India) (Sharma et al., 2007). Escherichia coli DH5a and E. coli BL21 (DE3) were used as hosts for DNA manipulations. The vectors pGEM-T Easy (Prome- ga) and pET 28a (+) (Novagen, Madison, USA) were used for sequencing of the amplified gene and construction of the expres- sion vector, respectively. 2.2. Genomic DNA extraction, amplification and sequencing The genomic DNA of G. thermoleovorans was extracted accord- ing to Bazzicalupo and Fani (1995) and processed for amplifying 0960-8524/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.biortech.2011.12.055 ⇑ Corresponding author. Address: Department of Microbiology, University of Delhi South Campus, Bentio Juarez Road, New Delhi 110 021, India. Tel.: +91 11 24112008; fax: +91 11 24115270. E-mail address: [email protected] (T. Satyanarayana). Bioresource Technology 107 (2012) 333–338 Contents lists available at SciVerse ScienceDirect Bioresource Technology journal homepage: www.elsevier.com/locate/biortech
-
Upload
digvijay-verma -
Category
Documents
-
view
214 -
download
1
Transcript of Cloning, expression and applicability of thermo-alkali-stable xylanase of Geobacillus...

Bioresource Technology 107 (2012) 333–338
Contents lists available at SciVerse ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Cloning, expression and applicability of thermo-alkali-stable xylanase of Geobacillusthermoleovorans in generating xylooligosaccharides from agro-residues
Digvijay Verma, T. Satyanarayana ⇑Department of Microbiology, University of Delhi South Campus, New Delhi 110 021, India
a r t i c l e i n f o
Article history:Received 10 November 2011Received in revised form 11 December 2011Accepted 12 December 2011Available online 21 December 2011
Keywords:Thermo-alkali-stable xylanaseRecombinant xyl-gtGeobacillus thermoleovoransXylooligosaccharidesPre-bleaching of paper pulp
0960-8524/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.biortech.2011.12.055
⇑ Corresponding author. Address: Department ofDelhi South Campus, Bentio Juarez Road, New Delhi24112008; fax: +91 11 24115270.
E-mail address: [email protected] (T. Satyana
a b s t r a c t
A xylanase gene (xyl-gt) of 1.224 kbp was cloned from the extremely thermophilic bacterium Geobacillusthermoleovorans that encodes a protein containing 408 amino acid residues. Eight conserved regions (sig-nature sequences) of GH family 10 xylanases have been found in the xylanase. When the xylanase genewas cloned and expressed in Escherichia coli BL21 (DE3), the recombinant strain produced xylanase titerof 270 U mg�1 which is 27-fold higher than the wild strain. It is optimally active at 80 �C and pH 8.5 witha high thermostability over broad range of pH (6–12) and temperature (40–100 �C). The end products ofthe hydrolysis of birch wood xylan and agro-residues included xylobiose, xylotriose, xylotetraose andxylopentaose. The xylanase of G. thermoleovorans is one of the rare xylanases that exhibits thermo-alkali-stability, and thus, it is a suitable candidate for pre-bleaching of paper pulps and generating xylool-igosaccharides from agro-residues for use as prebiotics.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Xylans are hetero- and homo-polysaccharides which are mainconstituents of hemicellulosic biomass of terrestrial plant cell wallmatrix and second most abundant polysaccharide on earth. Its corecomprises b-1,4 linked xylosyl residues, and the attachment of thiscore chain with various groups (4-O-methyl-D-glucuronosyl,a-arabinofuranosylresidues and others) makes it a heteropolysac-charide. Consequently the complete hydrolysis of xylan requiresa group of enzymes including endo-b-D-xylanase, b-xylosidase,arabinofuranosidase, glucuronidase, acetylxylan esterase, and feru-lic acid and p-coumaric acid esterases. Among these, xylanaseshave attracted considerable attention due to their catalytic activityfor releasing lower xylooligosaccharides, and their potential appli-cation in textile, feed and biofuel industries. Xylanases attractedattention further after Viikari et al. (1986) reported their applica-bility in bleaching of pulp in the paper industry. The inclusion ofxylanase treatment step reduces the chlorine consumption up to25–35% in bleaching kraft pulp to make the process environmentfriendly. Although several xylanases have been reported from aplethora of microorganisms, most of them do not tolerate the harshconditions employed in pulp processing. This process requiresthermo-alkali-stable and cellulase-free xylanase, because these
ll rights reserved.
Microbiology, University of110 021, India. Tel.: +91 11
rayana).
processes are usually operated at higher temperature and alkalineconditions.
An alkali-thermo-stable and cellulase-free xylanase suitable forpre-bleaching of paper pulps has been reported from Geobacillusthermoleovorans (Sharma et al., 2007). The xylanase produced bythe bacterium exhibits optimum activity at 80 �C and pH 8.5. Thecloning and expression of xylanase encoding gene (xyl-gt) ofG. thermoleovorans is crucial for enhancing the enzyme productionin order to make its production cost effective. In this investigation,cloning and expression of xylanase gene (xyl-gt) from G. thermo-leovorans and its applicability in saccharifying agro-residues toproduce xylooligosaccharides (XOs) are presented.
2. Methods
2.1. Bacterial strains and plasmids
G. thermoleovorans was isolated from the pulp samples collectedfrom Century Paper Mills, Lal Kuan, Uttaranchal (India) (Sharmaet al., 2007). Escherichia coli DH5a and E. coli BL21 (DE3) were usedas hosts for DNA manipulations. The vectors pGEM-T Easy (Prome-ga) and pET 28a (+) (Novagen, Madison, USA) were used forsequencing of the amplified gene and construction of the expres-sion vector, respectively.
2.2. Genomic DNA extraction, amplification and sequencing
The genomic DNA of G. thermoleovorans was extracted accord-ing to Bazzicalupo and Fani (1995) and processed for amplifying

334 D. Verma, T. Satyanarayana / Bioresource Technology 107 (2012) 333–338
the xylanase gene. One set of internal degenerate primer (Geo-xyl-int1F1/R1) was designed from the highly conserved regions ob-tained from multiple sequence alignment (MSA) of the amino acidsequences of xylanase protein of various Geobacillus and Bacillusspp. PCR amplification was carried out in a Thermocycler (Biorad,USA), and the amplicon was cloned into pGEM-T Easy vector forsequencing of the cloned fragment. The resulted sequence revealedstrong homology with xylanases of Geobacillus sp. WB1 and Geoba-cillus stearothermophilus. Therefore, new set of primers (Geo-xyl F/R) was designed from the end regions of xylanase encoding genesof Geobacillus WB1 in order to obtain the complete ORF. The full-length xylanase gene was amplified under defined PCR conditions(initial denaturation at 95 �C for 5 min followed by 29 cycles of94 �C for 50 s, 58 �C for 30 s and 72 �C for 1 min 20 s) in 50 lL reac-tion with final extension of 10 min at 72 �C using Geo-xyl F/R prim-ers in a Thermocyler (Biorad, USA). The primers used in this studyare listed in Table 1.
2.3. Construction and sequencing of the expression vector
To construct the recombinant vector, xyl-gt of G. thermoleovo-rans was amplified with the primers Geo-BamF and Geo-SacRprimers (Geo-xyl F/R primers with the added compatible restric-tion sites of Bam HI and Sac I, respectively). The digested PCR prod-uct was purified by Qiagen PCR purification kit and ligated intoBam HI and Sac I digested pET 28a (+) vector. The ligation mixturewas transformed into E. coli DH5a competent cells. The cloneswere screened for the positive recombinants using colony PCR fol-lowed by double digestion of the recombinant vectors with Bam HIand Sac I. Five positive clones were sequenced at the Nucleic AcidSequencing Facility, University of Delhi South Campus, New Delhifor obtaining the error free clone of xylanase gene.
2.4. Expression of the recombinant xylanase
One of the three clones having exactly the same gene sequencewas selected for expressing the xylanase protein. Recombinantplasmid was isolated and transformed into E. coli BL21 (DE3). A16–18 h old seed culture was prepared by growing the clone inLuria–Bertani (LB) broth containing kanamycin (50 lg/mL). Fiftymilliliter of LB medium containing kanamycin (50 lg/mL) wasinoculated with 1% (v/v) seed culture followed by further cultiva-tion at 37 �C to achieve an optical density of 0.5–0.7 at 600 nm.Thereafter the expression of xylanase was induced by adding1 mM isopropyl-b-D-1-thiogalactopyranoside (IPTG). The cell bio-mass was harvested periodically to study the growth profile as wellas xylanase synthesis.
2.5. Bioinformatics analysis
Homology analysis of the xyl-gt nucleotide sequence and itspredicted amino acid sequence with other known xylanases was
Table 1Primers used in this investigation.
Primersa Oligomeric sequences (50–30)
Geo-xyl-intF1 CACACACTCGTTTGGCACARCCAAGeo-xyl-intR1 NATGCTCACATCAAGCTCAGTNADNTGGeo-xylF CCCGGATCCATGCGGAACGTCGTGeo-xylR CCCGAGCTCCTATTTGTGGTCGATAATAGC
N: A/C/G/T; R: A/G; Y: C/T.a Internal degenerate primer (Geo-xyl-int1F1/R1) was used to obtain partial
region of xylanase gene from G. thermoleovorans. This region showed stronghomology with Geobacillus sp. WB1 and Geobacillus stearoethermophilus. Therefore,new set of primer (Geo-xylF/R) was picked from the end regions of xylanaseencoding genes of Geobacillus WB1 in order to obtain the full length gene.
carried out by using the database BLASTn (Basic Local AlignmentSearch Tool for Nucleotides) and BLASTp (Basic Local AlignmentSearch Tool for Protein), respectively. The CLUSTALW program(http://www.ebi.ac.uk/clustalW) and MEGA 4 (with minimum evo-lution) were used for the multiple sequence alignment (MSA) ofamino acid sequences and preparing the dendrogram, respectively.
2.6. Analysis and localization of recombinant xylanase (xyl-gt)
Localization of recombinant xylanase was done for confirmingthe cytoplasmic, periplasmic or extracellular nature of xylanase.The induced culture was harvested by centrifugation after 5 h ofinduction. The supernatant was collected as extracellular fractionwhile the pellet was suspended in 20 mM TE buffer (pH 8.0) with25% (w/v) sucrose and 1 mM EDTA. The cell suspension was shakenin an incubation shaker at 200 rpm for 15 min and cells were sed-imented by centrifugation at 4 �C, and subjected to osmotic treat-ment for 10 min by suspending the pellet into 5 mM chilledMgSO4 solution to release the periplasmic fraction of the cellswhich was collected as supernatant on centrifugation. Finally theintracellular fraction was collected on homogenizing the cells inchilled sonication buffer (25 mM Tris–HCl, 100 mM NaCl, 10 mMMgCl2, 6 M urea and 1 mg/mL lysozyme). The intracellular proteinwas released on sonication with 10 cycles of 1 min pulse (2 s on/off). The supernatant was separated from the cell debris after cen-trifugation and stored at �20 �C for assaying the enzyme in theintracellular fraction.
2.7. Xylanase assay
The activity of the recombinant xylanase was determinedaccording to Archana and Satyanarayana (1997). The reaction mix-ture consisting of 500 lL of 1% (w/v) birchwood xylan (Sigma, St.Louis, MO) prepared 0.1 M glycine–NaOH buffer (pH 8.5) and500 lL of appropriately diluted xylanase enzyme sample was incu-bated at 80 �C for 10 min. The reducing sugars liberated by the ac-tion on xylan were determined using 3, 5-dinitrosalicylic acidreagent (DNSA) (Miller, 1959). One unit of xylanase is defined asthe amount of enzyme that liberates 1 lmol of reducing sugarsas xylose under the assay conditions.
2.8. Purification of recombinant xylanase and zymogram analysis
The recombinant xylanase was purified by affinity chromatog-raphy using Ni2+-NTA agarose resins with minor alterations (Singhet al., 2004). Supernatant was collected after sonication of the in-duced cells in lysis buffer [6 M Urea, 100 mM NaCl, 25 mM TrisHCl (pH 8.0), 10 mM MgCl2 and 1 mg/mL lysozyme]. The columnhaving 5 mL of Ni2+-NTA resin were washed with sterile MQ (MilliQ water) and equilibrated with five column volume of buffer A(6 M urea in 100 mM phosphate buffer, pH 8, containing 5 mM b-mercaptoethanol) and buffer B [buffer A containing 10 mM TrisHCl (pH 8.0)] followed by buffer C [buffer B containing 20 mM TrisHCl (pH 8.0)]. Thereafter, supernatant having recombinant xylan-ase was passed five times through the column to bind the histi-dine-tagged protein. The column was washed with buffer C–D(100 mM phosphate buffer containing 10 mM Tris HCl pH 8.0,20% v/v glycerol and 5 mM b-mercaptoethanol) in gradient mannerfollowed by buffer E (buffer D containing 300 mM NaCl) to removethe unbound, non-specific proteins as well as other impurities. Fi-nally, the bounded protein was eluted from the column by passingbuffer D having a range of 100–500 mM imidazole. The fractionswere collected and analyzed on 15% SDS–PAGE for the purity ofthe recombinant protein. The activity of the pure protein was con-firmed by zymogram analysis on native PAGE. The gel was overlaidonto a 0.3% (w/v) xylan containing agar plate and incubated at

Fig. 1. Nucleotide and deuced amino acid sequences of the G.thermoleovoransxylanase gene. Underlined regions I (I V A E N V M K), II(R F H T L V W H), III (D V V N E), IV (L Y I N D Y N), V(I G H Q S H I), VI (I T E L D V), VII (T F W G I A D N H T W)and VIII (D Y I K V A F Q T A) denote highly conserved GH10 xylanase.Highlighted amino acids (Glu187; Asp230 and Glu293) are expected crucialcatalytic residues.
D. Verma, T. Satyanarayana / Bioresource Technology 107 (2012) 333–338 335
80 �C for 2 h. The xylan hydrolysis zone was confirmed stainingwith Congo-red.
2.9. Biochemical characterization of recombinant xylanase
The physical and biochemical properties of pure xylanase havebeen studied. A range of buffers (pH 3–6, 0.1 M citrate buffer; 7–8,0.1 M citrate buffer and for 9–12, 0.1 M glycine–NaOH buffer) wereused for determining optimum pH. Similarly, optimum tempera-ture was determined by assaying the enzyme at different temper-atures (40–100 �C). Thermostability and pH stability weremeasured by incubating the recombinant enzyme at different tem-peratures (60–90 �C) and pH (8–10), respectively for varying timeintervals followed by determining the enzyme activities. The ef-fects of various metal ions, inhibitors, solvents and detergents onthe recombinant enzyme activity have been assessed by includingthese in the reaction mixtures. In addition, the specificity of recom-binant xylanase for different substrates was studied using xylansfrom different sources and non-xylan substrates (starch, cellulose,pullulan and chitin) in glycine–NaOH buffer (pH 8.5) and assayedat 80 �C. The Km and Vmax were determined at 80 �C by recordingthe effect of various concentrations of birchwood xylan and plot-ting Lineweaver–Burk curve.
2.10. Analysis of hydrolysis product
Two hundred fifty microgram of xylan (from birchwood) wasincubated with 5 U of recombinant xylanase in 200 lL glycine–NaOH buffer (pH 8.5) at 80 �C for 3 h. The samples were collectedat different time intervals and sugars were analyzed on silica plateby Thin layer chromatography (TLC). The sugars were identified bycomparing the Rf values of the standards of xylose, xylobiose, xylo-triose, xylotetraose and xylopentaose (Sigma, USA).
2.11. Saccharification of agro-residues
The saccharification of various natural agro-residues was car-ried out at 80 �C and pH 8.5. In order to optimize xylanase dosage,different titers of xylanase have been used in saccharifying wheatbran. The samples were withdrawn at the desired intervals (1, 2,4, 8 and 12 h) and analyzed by HPLC using carbohydrate column(3.9 � 300 mm, Nova-Pak spherical silica bonded with trifunction-al aminopropylsilane, Waters) with acetonitrile/water (70:30, v/v)as the mobile phase at 0.5 mL min�1, and injection volume of10 lL. The peaks of sugars were identified and quantified using dif-ferential refractive index detector and analyzed by Empower 2154software (Waters). The reducing sugars liberated by the enzymeaction were determined using DNSA reagent.
2.12. Nucleotide sequence accession numbers
The amplified xylanase gene of G. thermoleovorans was se-quenced and has been submitted in GenBank (Accession No.JN680872).
3. Results and discussion
3.1. PCR amplification, construction, sequencing and bioinformaticanalysis of xylanase gene
When the fragment of xylanase gene was amplified using Geo-xyl-intF1/R1, 500 bp amplicon was obtained. The full-length xyl-gtgene (1224 bp) was amplified using Geo-xyl F/R primers with com-patible restriction sites at 58 �C and cloned into pET 28a (+) vector.The presence of the cloned xyl-gt in the recombinant construct
(Gxyl1-pET28) was confirmed by colony PCR followed by doubledigestion with Bam HI and Sac I and sequencing of the cloned frag-ment. The complete ORF of recombinant xylanase gene starts withATG as initiation and ends in TAG as stop codon (Fig. 1). The size ofamplified gene was 1224 bp, with an overall G + C content of 46%.This ORF encodes 408 amino acid residue protein with 53 stronglybasic and 51 strongly acidic amino acids. The deduced amino acidsequence showed 99% and 97% identity with xylanase of G. stearo-thermophilus and Geobacillus sp. WBI, respectively. It also exhibits65% identity with the alkalistable and thermostable endoxylanaseof Bacillus sp. NG-27, Bacillus firmus and Bacillus halodurans, andwith the other family 10 xylanases of Dictyoglomus turgidumDSM 6724 (44%) and Anoxybacillus sp. E2 (209) (43%), respectively.
Its predicted molecular weight is �45 kDa. The phylogenetictree of the sequence of amino acids of this xylanase with otherxylanases in the protein databases revealed that it belongs to thefamily 10 xylanases (Fig. 2). The higher molecular mass(�45 kDa) of this enzyme also supports its categorization in GHfamily 10 (Guo et al., 2009; Mamo et al., 2006). When the aminoacid sequence of the xylanase was aligned with various GH10

Xyl anase (Geobacillus sp. WBI)
Xylanase (Geobacillus stearothermophilus)
endo-14-beta-xylanase (Geobacillus sp. C56-T3)
Xylanase (Geobacillus thermoleovorans)
Xylanase (Geobacillus sp. TC-W7)
Xylanase T-6 (Geobacillus thermodenitrificans)
alkaline thermostable endoxylanase (Bacillus sp. NG27)
Xylanase 10A (Bacillus firmus)
Xylanase (Bacillus sp. N16-5)
Xylanase (Anoxybacillus sp. E2(2009))
endo-1,4-beta-xylanase (Geobacillus sp.(2)
endoxylanase (Paenibacillus sp. E18)
endo-beta-1,4-xylanase (Bacillus sp.)
79
48
100
99
100100
100
78
64
92
0.1
Fig. 2. Neighbour Joining (NJ) Tree (MEGA 4 software) constructed by using xylanase sequence of G. thermoleovorans along with xylanases retrieved from NCBI to show thephylogenetic relationship. Predicted amino acid sequence shared highest homology with xylanase of Geobacillus sp. WBI followed by G. stearothermophilus.
336 D. Verma, T. Satyanarayana / Bioresource Technology 107 (2012) 333–338
xylanases of bacteria, fungi and plants, eight conserved regions (I–VIII) have been identified. The signature sequences (I–V) and VI–VIIof G. thermoleovorans have resemblance with those reported byBaba et al. (1994) and Fukumura et al. (1995). The region VIII isconsidered to be a fingerprint of GH10 xylanases (Fukumuraet al., 1995) (Fig. 1). The proposed 3D structure also revealed thatGlu187 and Glu239 are catalytically important amino acids (Guoet al., 2009; Mamo et al., 2006). The construct Gxyl1-pET28a wasfurther confirmed by expression in E. coli BL21 (DE3). The expres-sion of the recombinant xyl-gt was enhanced by induction with1 mM IPTG after 4 h. Most of the recombinant protein was in theintracellular fraction (�89%), while periplasmic and extracellularfractions accounted for 7% and 4% of the total, respectively, as inthe xylanases of B. halodurans (Guo et al., 2009) and Glaciecolamesophila KMM241 (Mamo et al., 2006).
3.2. Purification of recombinant xylanase, zymogram analysis andbiochemical characterization
The recombinant protein was eluted from Ni2+-NTA resins at200 mM of imidazole, and observed as single band on 15% SDS–PAGE. The molecular weight of recombinant protein was con-firmed against denaturing protein markers (Sigma, USA) (Fig. 3)and it was very close to the reported molecular weight (48 kDa)of the native xylanase of G. thermoleovorans (Sharma et al., 2007).The zymogram analysis further confirmed the presence of activexylanase protein on native PAGE. The recombinant xylanase exhib-
Fig. 3. SDS–PAGE analysis of recombinant xylanase expressed in pET 28a (+)systems (samples were resolved on 15% polyacrylamide gel and stained withCoomassie Brilliant Blue R-250 and by activity staining). (A) Lane 1–9: elutionprofile of recombinant xylanase obtained from Ni2+-NTA column using imidazole(50, 100, 200, 250, 300, 350, 400, 450, 500 mM). (B) Gel picture showing zymogramanalysis using Congo red. (C) Purified recombinant xylanase of 45 kDa.
ited activity over broad range of pH (6–12) and temperatures (40–100 �C) with optimum at pH 8.5 and 80 �C, respectively. The xylan-ase retained activity for 3 h at 60 and 70 �C with T1/2 of 60 and10 min at 80 and 90 �C, respectively and it is very similar to the na-tive xylanase of G. thermoleovorans (Table 2). Xylanases of Bacillusstearothermophilus T-6 and Geobacillus sp. 70PC53 were optimallyactive at 65 �C (Khasin et al., 1993; Son-Ng et al., 2009), while thatof B. firmus at 70 �C (Chang et al., 2004). Dictyoglomus thermolacti-cum and B. firmus xylanases are among the very few that are opti-mally active at 80 �C (Uchino and Fukuda, 1983; Mathrani andAhring, 1992). Similarly, xylanases of a few species of Bacillusand Geobacillus exhibit optimal activity in the alkaline pH range(Khasin et al., 1993; Chang et al., 2004; Archana and Satyanaraya-na, 1997). The stability of recombinant xylanase at higher pH andtemperatures makes it a candidate for pre-bleaching of paper pulps(Sharma et al., 2007; Archana and Satyanarayana, 1997).
Many metal ions did not show any significant effect on xylanaseactivity except Fe2+, Hg2+ and Cu2+. Iron slightly enhanced thexylanase activity as that of Bacillus subtilis AMX-4 (Hong, 2009).Copper inhibited the activity of xylanase of G. thermoleovorans asreported in various xylanases from Staphylococcus sp. SG-13 andPlectosphaerella cucumerina (Zhang et al., 2007; Gupta et al.,2000). The strong inhibitory effect of Hg2+ on xylanase suggestedthe significance and involvement of tryptophan residue in the cat-alytic activity of the enzyme, since Hg2+ oxidizes indole ring, it isconsidered to interact with aromatic ring present in tryptophan(Zhang et al., 2007; Liu et al., 2010). Among all the modulators,N-bromosuccinimide (N-BS) strongly inhibited xylanase activity
Table 2Comparison of the characteristics of native and recombinant xyl xylanases of G.thermoleovorans.
Property Native xylanase Recombinant xylanase(xyl gt)a
Specific enzymeactivity
10.2 U mg�1 270 U mg�1
Temperature optimum 80 �C 80 �CpH optimum 8.5 8.5Thermal stability (T1/2
80 �C)50–55 min 50–60 min
Molecular weight 48 kDa 45 kDaKm 2.6 mg mL�1 2.10 mg mL�1
Vmax 31.2 lmol mg�1 min�1 42.5 lmol mg�1 min�1
Vmax/Km 12.6 min�1 20.2 min�1
kcat 280.56 s�1 318.75 s�1
kcat/Km 107.90 mL mg�1 s�1 151.78 mL mg�1 s�1
a Recombinant xylanase exhibited properties similar to that of the native.
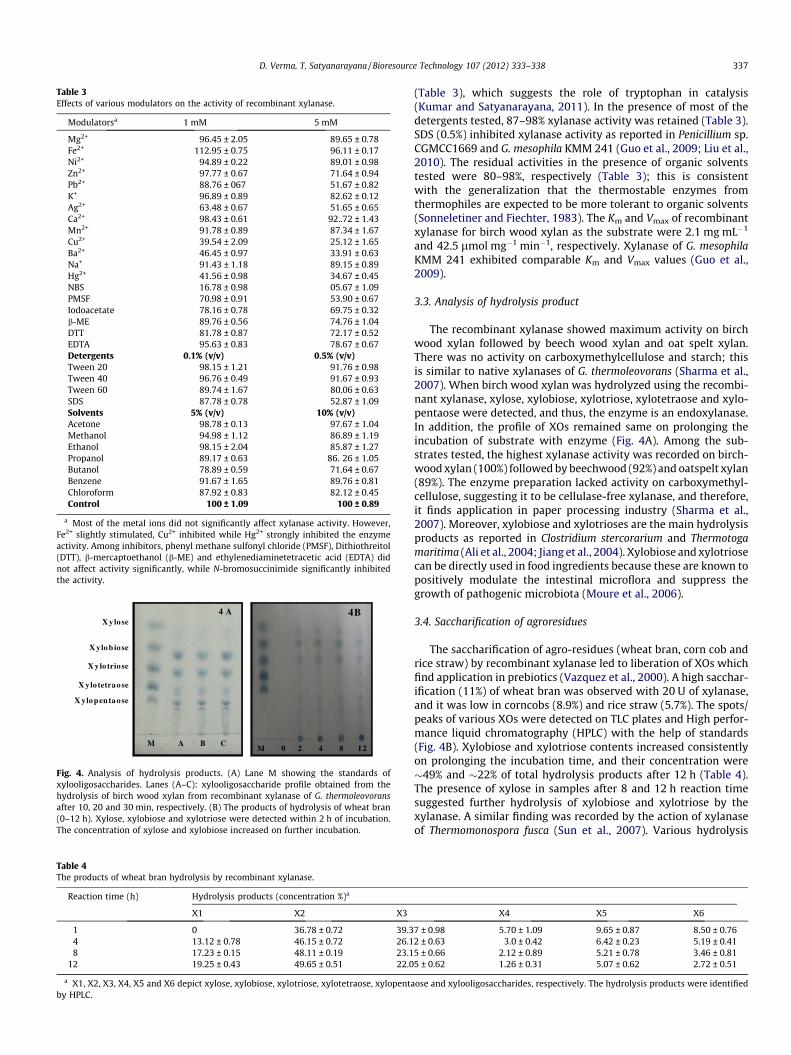
Table 3Effects of various modulators on the activity of recombinant xylanase.
Modulatorsa 1 mM 5 mM
Mg2+ 96.45 ± 2.05 89.65 ± 0.78Fe2+ 112.95 ± 0.75 96.11 ± 0.17Ni2+ 94.89 ± 0.22 89.01 ± 0.98Zn2+ 97.77 ± 0.67 71.64 ± 0.94Pb2+ 88.76 ± 067 51.67 ± 0.82K+ 96.89 ± 0.89 82.62 ± 0.12Ag2+ 63.48 ± 0.67 51.65 ± 0.65Ca2+ 98.43 ± 0.61 92..72 ± 1.43Mn2+ 91.78 ± 0.89 87.34 ± 1.67Cu2+ 39.54 ± 2.09 25.12 ± 1.65Ba2+ 46.45 ± 0.97 33.91 ± 0.63Na+ 91.43 ± 1.18 89.15 ± 0.89Hg2+ 41.56 ± 0.98 34.67 ± 0.45NBS 16.78 ± 0.98 05.67 ± 1.09PMSF 70.98 ± 0.91 53.90 ± 0.67Iodoacetate 78.16 ± 0.78 69.75 ± 0.32b-ME 89.76 ± 0.56 74.76 ± 1.04DTT 81.78 ± 0.87 72.17 ± 0.52EDTA 95.63 ± 0.83 78.67 ± 0.67Detergents 0.1% (v/v) 0.5% (v/v)Tween 20 98.15 ± 1.21 91.76 ± 0.98Tween 40 96.76 ± 0.49 91.67 ± 0.93Tween 60 89.74 ± 1.67 80.06 ± 0.63SDS 87.78 ± 0.78 52.87 ± 1.09Solvents 5% (v/v) 10% (v/v)Acetone 98.78 ± 0.13 97.67 ± 1.04Methanol 94.98 ± 1.12 86.89 ± 1.19Ethanol 98.15 ± 2.04 85.87 ± 1.27Propanol 89.17 ± 0.63 86. 26 ± 1.05Butanol 78.89 ± 0.59 71.64 ± 0.67Benzene 91.67 ± 1.65 89.76 ± 0.81Chloroform 87.92 ± 0.83 82.12 ± 0.45Control 100 ± 1.09 100 ± 0.89
a Most of the metal ions did not significantly affect xylanase activity. However,Fe2+ slightly stimulated, Cu2+ inhibited while Hg2+ strongly inhibited the enzymeactivity. Among inhibitors, phenyl methane sulfonyl chloride (PMSF), Dithiothreitol(DTT), b-mercaptoethanol (b-ME) and ethylenediaminetetracetic acid (EDTA) didnot affect activity significantly, while N-bromosuccinimide significantly inhibitedthe activity.
X ylose
X ylob iose
X ylo triose
X ylo tetraose
X ylopentaose
M A B CM 0 2 4 8 12
4 A 4B
Fig. 4. Analysis of hydrolysis products. (A) Lane M showing the standards ofxylooligosaccharides. Lanes (A–C): xylooligosaccharide profile obtained from thehydrolysis of birch wood xylan from recombinant xylanase of G. thermoleovoransafter 10, 20 and 30 min, respectively. (B) The products of hydrolysis of wheat bran(0–12 h). Xylose, xylobiose and xylotriose were detected within 2 h of incubation.The concentration of xylose and xylobiose increased on further incubation.
Table 4The products of wheat bran hydrolysis by recombinant xylanase.
Reaction time (h) Hydrolysis products (concentration %)a
X1 X2 X3
1 0 36.78 ± 0.72 39.34 13.12 ± 0.78 46.15 ± 0.72 26.18 17.23 ± 0.15 48.11 ± 0.19 23.1
12 19.25 ± 0.43 49.65 ± 0.51 22.0
a X1, X2, X3, X4, X5 and X6 depict xylose, xylobiose, xylotriose, xylotetraose, xylopentaby HPLC.
D. Verma, T. Satyanarayana / Bioresource Technology 107 (2012) 333–338 337
(Table 3), which suggests the role of tryptophan in catalysis(Kumar and Satyanarayana, 2011). In the presence of most of thedetergents tested, 87–98% xylanase activity was retained (Table 3).SDS (0.5%) inhibited xylanase activity as reported in Penicillium sp.CGMCC1669 and G. mesophila KMM 241 (Guo et al., 2009; Liu et al.,2010). The residual activities in the presence of organic solventstested were 80–98%, respectively (Table 3); this is consistentwith the generalization that the thermostable enzymes fromthermophiles are expected to be more tolerant to organic solvents(Sonneletiner and Fiechter, 1983). The Km and Vmax of recombinantxylanase for birch wood xylan as the substrate were 2.1 mg mL�1
and 42.5 lmol mg�1 min�1, respectively. Xylanase of G. mesophilaKMM 241 exhibited comparable Km and Vmax values (Guo et al.,2009).
3.3. Analysis of hydrolysis product
The recombinant xylanase showed maximum activity on birchwood xylan followed by beech wood xylan and oat spelt xylan.There was no activity on carboxymethylcellulose and starch; thisis similar to native xylanases of G. thermoleovorans (Sharma et al.,2007). When birch wood xylan was hydrolyzed using the recombi-nant xylanase, xylose, xylobiose, xylotriose, xylotetraose and xylo-pentaose were detected, and thus, the enzyme is an endoxylanase.In addition, the profile of XOs remained same on prolonging theincubation of substrate with enzyme (Fig. 4A). Among the sub-strates tested, the highest xylanase activity was recorded on birch-wood xylan (100%) followed by beechwood (92%) and oatspelt xylan(89%). The enzyme preparation lacked activity on carboxymethyl-cellulose, suggesting it to be cellulase-free xylanase, and therefore,it finds application in paper processing industry (Sharma et al.,2007). Moreover, xylobiose and xylotrioses are the main hydrolysisproducts as reported in Clostridium stercorarium and Thermotogamaritima (Ali et al., 2004; Jiang et al., 2004). Xylobiose and xylotriosecan be directly used in food ingredients because these are known topositively modulate the intestinal microflora and suppress thegrowth of pathogenic microbiota (Moure et al., 2006).
3.4. Saccharification of agroresidues
The saccharification of agro-residues (wheat bran, corn cob andrice straw) by recombinant xylanase led to liberation of XOs whichfind application in prebiotics (Vazquez et al., 2000). A high sacchar-ification (11%) of wheat bran was observed with 20 U of xylanase,and it was low in corncobs (8.9%) and rice straw (5.7%). The spots/peaks of various XOs were detected on TLC plates and High perfor-mance liquid chromatography (HPLC) with the help of standards(Fig. 4B). Xylobiose and xylotriose contents increased consistentlyon prolonging the incubation time, and their concentration were�49% and �22% of total hydrolysis products after 12 h (Table 4).The presence of xylose in samples after 8 and 12 h reaction timesuggested further hydrolysis of xylobiose and xylotriose by thexylanase. A similar finding was recorded by the action of xylanaseof Thermomonospora fusca (Sun et al., 2007). Various hydrolysis
X4 X5 X6
7 ± 0.98 5.70 ± 1.09 9.65 ± 0.87 8.50 ± 0.762 ± 0.63 3.0 ± 0.42 6.42 ± 0.23 5.19 ± 0.415 ± 0.66 2.12 ± 0.89 5.21 ± 0.78 3.46 ± 0.815 ± 0.62 1.26 ± 0.31 5.07 ± 0.62 2.72 ± 0.51
ose and xylooligosaccharides, respectively. The hydrolysis products were identified

338 D. Verma, T. Satyanarayana / Bioresource Technology 107 (2012) 333–338
products (X2–X5) were obtained at various time intervals with xy-lose (X1) as minimum hydrolysis product. Xylobiose and xylotriosewere detected as main XOs at 8 and 12 h of incubation. A similarprofile of hemicellulose hydrolysis end products has been reportedfor the xylanases of T. fusca and B. halodurans (Sun et al., 2007; Ku-mar and Satyanarayana, 2011). Since alkaline xylanases are knownto enhance the solubility of lignocellulosic materials (Mamo et al.,2007; Kumar and Satyanarayana, 2011), the alkalistable xylanaseof G. thermoleovorans appears to be a suitable candidate for gener-ating XOs from abundantly available agro-residues.
4. Conclusions
The thermo-alkali-stable and cellulase-free endo-b-1,4-xylan-ase of G. thermoleovorans is one of the rare xylanases because ofits stability at extreme process conditions prevailing in the paperindustry. The action of recombinant xylanase on birch wood xylanand agro-residues produced XOs which can be used as prebiotics.Further attempts are underway to clone and express xyl-gt in Bacil-lus megaterium extracellularly in order to simplify the recovery ofxylanase.
Acknowledgement
We gratefully acknowledge the financial assistance from theDepartment of Biotechnology and Department of Science & Tech-nology, Government of India for carrying out the work.
References
Ali, M.K., Rudolph, F.B., Bennett, G.N., 2004. Thermostable xylanase 10B fromClostridium acetobutylicum ATCC824. J. Ind. Microbiol. Biotechnol. 31, 229–234.
Archana, A., Satyanarayana, T., 1997. Xylanase production by thermophilic Bacilluslicheniformis A99 in solid state fermentation. Enzyme Microb. Technol. 21, 12–17.
Baba, T., Shinke, R., Nanmori, T., 1994. Identification and characterization ofclustered genes for thermostable xylan-degrading enzymes, b-xylosidase andxylanase, of Bacillus stearothermophilus 21. Appl. Environ. Microbiol. 60, 2252–2258.
Bazzicalupo, M., Fani, R., 1995. The use of RAPD for generating specific DNA probesfor microorganisms. In: Clapp, J.P. (Ed.), Methods in Molecular Biology, S-Species Diagnostic Protocols: PCR and Other Nucleic Acids Methods, vol. 50..Humana Press Inc., Totowa, NJ, pp. 155–177.
Chang, W.S., Tsai, C.L., Tseng., M.J., 2004. Cloning and characterization of twothermostable xylanases from an alkaliphilic Bacillus firmus. Biochem. Biophys.Res. Commun. 319, 1017–1025.
Fukumura, M., Sakka, K., Shimada, K., Ohmiya, K., 1995. Nucleotide sequence of theClostridium stercorarium xynB gene encoding an extremely thermostablexylanase, and characterization of the translated product. Biosci. Biotechnol.Biochem. 59, 40–46.
Guo, B., Chen, X., Sun, C., Zhou, B., Zhang, Y., 2009. Gene cloning, expression andcharacterization of a new cold-active and salt tolerant endo-b-1,4-xylanase
from marine Glaciecola mesophila KMM 241. Appl. Microbiol. Biotechnol. 84,1107–1115.
Gupta, S., Bhushan, B., Hoondal, G.S., 2000. Isolation, purification andcharacterization of xylanase from Staphylococcus sp. SG-13 and its applicationin biobleaching of kraft pulp.. J. Appl. Microbiol. 88, 325–334.
Hong, Y., 2009. Cloning of the Bacillus subtilis AMX-4 xylanase gene andcharacterization of the gene product. J. Microbiol. Biotechnol. 19 (12), 1514–1519.
Jiang, Z.Q., Deng, W., Zhu, Y.P., Li, L.T., Sheng, Y.J., Hayashi, K., 2004. The recombinantxylanase B of Thermotoga maritima is highly xylan specific and producesexclusively xylobiose from xylans, a unique character for industrialapplications. J. Mol. catal. B Enzyme 27, 207–213.
Khasin, A., Alchanati, I., Shoham, Y., 1993. Purification and characterization of athermostable xylanase from Bacillus stearothermophilus T-6. Appl. Environ.Microbiol. 59, 1725–1730.
Kumar, V., Satyanarayana, T., 2011. Applicability of thermo-alkali-stable andcellulase-free xylanase from a novel thermo-halo-alkaliphilic Bacillushalodurans in producing xylooligosaccharides. Biotechnol. Lett. 33, 2279–2285.
Liu, W., Shi, P., Chen, Q., yang, P., wang, G., Wang, Y., Luo, H., Yao, B., 2010. Genecloning, over expression, characterization of a xylanase from Penicillium sp.CGMCC 1699. Appl. Biochem. Biotechnol. 162, 1–12.
Mamo, G., Delgado, O., Martinez, A., Mattiasson, B., Hatti-Kaul, R., 2006. Cloning,sequence analysis and expression of a gene encoding an endoxylanase fromBacillus halodurans S7. Mol. Biotechnol. 33, 149–160.
Mamo, G., Hatti-Kaul, R., Mattiasson, B., 2007. Fusion of carbohydrate bindingmodules from Thermotoga neapolitana with a family 10 xylanase from Bacillushalodurans S7. Extremophiles 11, 169–177.
Mathrani, I.M., Ahring, B.K., 1992. Thermophilic and alkaliphilic xylanase fromseveral Dictyoglomus isolates. Appl. Microbiol. Biotechnol. 38, 23–27.
Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducingsugars. Anal. Chem. 31, 426–428.
Moure, A., Gullon, P., Dominguez, H., Parajo, J.C., 2006. Advances in themanufacture, purification and applications of xylooligosaccharides as foodadditives and nutraceuticals. Process Biochem. 41, 1913–1923.
Sharma, S., Adhikari, S., Satyanarayana, T., 2007. Alkali-thermostable and cellulase-free xylanase production by an extreme thermophile Geobacillusthermoleovorans. World J. Microbiol. Biotechnol. 23, 483–490.
Singh, J., Joshi, M.C., Bhatnagar, R., 2004. Cloning and expression of mycobacterialglutamine synthetase gene in Escherichia coli. Biochem. Biophys. Res. Commun.317, 634–638.
Son-Ng, I., Li, C.W., Yeh, Y., Chen, P.T., Chir, J.L., Ma, C.H., Su-May Yu, S.M., Ho, T.D.,Tong, C.G., 2009. A novel endo-glucanase from the thermophilic bacteriumGeobacillus sp. 70PC53 with high activity and stability over a broad range oftemperatures. Extremophiles 13, 425–435.
Sun, J.Y., Liu, M.Q., Weng, X.Y., Qian, L.C., Gu, S.H., 2007. Expression of recombinantThermomonospora fusca xylanase A in Pichia pastoris and xylooligosaccharidesreleased from xylans by it. Food Chem. 104, 1055–1064.
Sonneletiner, B., Fiechter, A., 1983. Advantages of using thermophiles inbiotechnological processes: expectations and reality. Trend Biotechnol. 1 (3),74–80.
Uchino, F., Fukuda, O., 1983. Taxanomic characteristics of a acidophilic strain ofBacillus producing thermophilic acidophilic amylase and thermostablexylanase. Agric. Biol. Chem. 47, 965–967.
Vazquez, M.J., Alonso, J.L., Dominguez, H., Parajo, J.C., 2000. Xylooligosaccharides:manufacture and applications. Trends Food Sci. Technol. 11, 387–393.
Viikari, L., Ranua, M., Kantelinen, A., Sunduist, J., Linko, M. 1986. Bleaching withenzymes. In: Proceedings of the Third International Conference Biotechnologyin the Pulp and Paper Industry. STFI, Stockholm, Sweden, pp. 67–69.
Zhang, G.M., Huang, J., Huang, G.R., Ma, L.X., Zhang, X.E., 2007. Molecular cloningand heterologous expression of a new xylanase gene from Plectosphaerellacucumerina. Appl. Microbiol. Biotechnol. 74, 339–346.


















