
Classification, natural history, and evolution of the ... SB 99 2... · Classification, natural...
90
5 Acta Musei Moraviae, Scientiae biologicae (Brno) 99(2): 594, 2014 Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae) Part XI. Generic taxonomy, intergeneric phylogeny, and catalogue of the subfamily WESTON OPITZ Kansas Wesleyan University, Department of Biology; 100 East Claflin Avenue, Salina, Kansas 67401-6196; USA; e-mail: [email protected] OPITZ W. 2014: Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). Part XI. Generic taxonomy, intergeneric phylogeny, and catalogue of the subfamily. Acta Musei Moraviae, Scientiae biologicae (Brno) 99(2): 594. This treatise is the authors culmination efforts with the checkered beetle subfamily Epiphloeinae Kuwert. The work contains a summation of the natural history of epiphloeine species, discussion of characters and character states found useful for the discernment of epiphloeine genera and species, description of the subfamily, key to genera, synoptic description of the genera, intergeneric hypothesis of phylogeny, catalogue of the species with each species represented by a color photograph, and index of genera, their species, and their synonyms. Keywords. Coleoptera, Cleridae, Epiphloeinae, genera, phylogeny, catalogue Introduction Epiphloeines are New World arbophilic beetles that frequent tree trunks infested with wood decomposing insects such as bark beetles. Within this group of checkered beetles one finds considerable intrageneric diversity of integumental color and body form varies extensively across generic lines. The species of some genera blend beautifully with the dark scabrous characteristic of tree bark; as in members of Plocamocera Spinola and Silverasia NemØsio. In other epiphloeines the elytra are brightly colored, as one finds among species of the mimetic Epiphloeus Spinola and Acanthocollum Opitz. Commonly, body form varies from suboval, rectangulate, or narrow triangular, with Batesian lycid- like body shapes among members of Ichnea Laporte and cantharoid shapes in Opitzia NemØsio and Hapsidopteris Opitz. As defined herein, Epiphloeinae contains 271 species classified into 25 genera whose combined distribution extends from eastern Canada to northern Argentina. The genera, and their species and type localities, are listed in the taxonomic catalogue that forms the last component of this treatise. Three synapotypic characteristics define the monophyly of Epiphloeinae. One, there are two fully-developed pairs of pronotal trichobothria, one pair is situated paralaterally on the pronotal disc, the second is located on the pronotal sides. Two, the antenna are inserted along the lower frontal margin of the eyes. Three, the metendosternite lacks a furcal lamina. ISSN 1211-8788
Transcript of Classification, natural history, and evolution of the ... SB 99 2... · Classification, natural...

5
Acta Musei Moraviae, Scientiae biologicae (Brno)99(2): 5�94, 2014
Classification, natural history, and evolution of the Epiphloeinae(Coleoptera: Cleridae)
Part XI. Generic taxonomy, intergeneric phylogeny, and catalogue of the subfamily
WESTON OPITZ
Kansas Wesleyan University, Department of Biology; 100 East Claflin Avenue, Salina, Kansas 67401-6196; USA; e-mail: [email protected]
OPITZ W. 2014: Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartXI. Generic taxonomy, intergeneric phylogeny, and catalogue of the subfamily. Acta Musei Moraviae, Scientiaebiologicae (Brno) 99(2): 5�94. � This treatise is the author�s culmination efforts with the checkered beetlesubfamily Epiphloeinae Kuwert. The work contains a summation of the natural history of epiphloeine species,discussion of characters and character states found useful for the discernment of epiphloeine genera and species,description of the subfamily, key to genera, synoptic description of the genera, intergeneric hypothesis ofphylogeny, catalogue of the species with each species represented by a color photograph, and index of genera,their species, and their synonyms.
Keywords. Coleoptera, Cleridae, Epiphloeinae, genera, phylogeny, catalogue
IntroductionEpiphloeines are New World arbophilic beetles that frequent tree trunks infested
with wood decomposing insects such as bark beetles. Within this group of checkeredbeetles one finds considerable intrageneric diversity of integumental color and body formvaries extensively across generic lines. The species of some genera blend beautifully withthe dark scabrous characteristic of tree bark; as in members of Plocamocera Spinola andSilverasia Nemésio. In other epiphloeines the elytra are brightly colored, as one findsamong species of the mimetic Epiphloeus Spinola and Acanthocollum Opitz. Commonly,body form varies from suboval, rectangulate, or narrow triangular, with Batesian lycid-like body shapes among members of Ichnea Laporte and cantharoid shapes in OpitziaNemésio and Hapsidopteris Opitz.
As defined herein, Epiphloeinae contains 271 species classified into 25 generawhose combined distribution extends from eastern Canada to northern Argentina.
The genera, and their species and type localities, are listed in the taxonomiccatalogue that forms the last component of this treatise. Three synapotypic characteristicsdefine the monophyly of Epiphloeinae. One, there are two fully-developed pairs ofpronotal trichobothria, one pair is situated paralaterally on the pronotal disc, the secondis located on the pronotal sides. Two, the antenna are inserted along the lower frontalmargin of the eyes. Three, the metendosternite lacks a furcal lamina.
ISSN 1211-8788

W. OPITZ
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 20146
Cleridae subfamily classification issues �The systematics of the Cleridae should be revised to reflect molecular data�
GUNTER et al. 2013: 8�. The authors of this statement may be correct when, and onlywhen, adequate representations of species, of an adequate representation of genera, havebeen investigated from a molecular basis; and when it is possible to determine theapotypic or plesiotypic evolutionary condition of a set of genes. Commonality in geneticbase pairs does not necessarily translate into phylogenetic relationship; no more thansimilarity in morphological characteristics translates into sister group relationships. Onlysynapotypic base pairs or synapotypic structural characteristics qualify for modernthoughts about phylogenetic kinships.
In the abovementioned molecular work 148 species and 70 genera [out of about3,629 species and 334 genera (OPITZ 2010: 55)] were considered. While the molecularstudy in question is an important augment to Cleridae systematic work, I submit that thestudy contains insufficient representation of Cleridae taxa for sustaining issues ofCleridae higher classification. Moreover, the molecular analysis bestows monophyly onsome taxa, but their definition of monophyly is based on a computer program that doesnot distinguish between the apotypic and plesiotypic state of a gene or sets of genes. Themolecular study does not define which base pairs are derived and which are primitive.Having a similar set of genes does not necessarily mean phylogenetic relationships. Thehuman genome shares base pairs with the genome of bacteria, which does not mean thatthe two taxa in question are closely related. The sharing of similar base pairs couldsimply mean that two sets of organisms have evolved similar sets of genes to solve acommon problem for survival; nuances in unproven techniques of phylogenetic analysisshould not trump carefully prepared morphological analysis in accordance with theproven phylogenetic principles of HENNIG (1966). The exclusion of apotypic andplesiotypis parameters in molecular phylogenetics is akin to the idea the commonality ofcharacteristics translates into evolutionary kinships. Such methodology is similar the thedicredited canons of Numerical Taxomy. These considerations becomes particularlyrelevant when results from a molecular study are used to minimize the significance oftaxa-rich morphological analyses.
In a recent contribution BARTLETT (2013: 412) writes...� The subfamilies of OPITZ�S(2010) �split� classification (of Korynetinae, sensu latu), though satisfying the need for amore detailed classification, are not all well-defined in terms of synapomorphies andmost, with the exception of Epiphloeinae, are not well-supported by molecularevidence.� I presume that this statement relates to Bartlett�s belief that the synapotypiesgiven in OPITZ�S (2010) for subfamilies Enopliinae, Tarsosteninae, Korynetinae,Neorthopleurinae and Peloniinae are manifestations of the use of �continuous charactersas defined by SMITH & HENDRICKS (2013: 367). If this assumption is correct I vehementlydisagree with Bartlett�s assessment. For example, Enopliinae is clearly defined by apartial commissure (a result of a meeting of the pronotal hem with the dorsolateral carinapartially mesad to the hind corner of the pronotum), or in Tarsosteninae where thecapitulum is consistently shorter than the combined length of funicular antennomeres. In

7
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
the taxa that I have examined, there is no in-between of these characteristic among taxawith a reduced 4th tarsomere. The meeting of the pronotal hem with the dorsolateralcarina either meets at the corner or it does not. Moreover, elsewhere (OPITZ 2011:142) Ihave addressed the issue of �homoplastic apotypy� being a viable consideration inestablishing phylogenetic relationships (on that basis... �homoplastic apotypies may be amanifestation of selection of (distantly related) different ancestral genomes to solve acommon (functional) problem�. Similarity of morphological character states based onfunctional parameters would of course assure similar nucleic acid base pairs. However,similar structure, and their corresponding similar base pairs, does not necessarilytranslate into close evolutionary kinships.
The molecular study of GUNTER et al. (2013: 634), and the works of KOLIBÁČ (1997:358; 2010: 260), LESCHEN (2010: 5, 257), BOUCHARD et al. (2011: 348), and LAWRENCEet al. (2010: 5; 2011: 8) place a high value on the phylogenetic significance of thereduction of the 4th tarsomere in the Cleridae. The result of such an interpretation hasprompted some of the above authors to place OPITZ�s (2010: 117) Epiphloeinae,Enopliinae, Neorthopleurinae, Peloniinae, Tarsosteninae, and Korynetinae under onesubfamily, the Korynetinae. I found the length, and therefore expression, of thistarsomere variable within the family; some of this variation can be seen in illustrationsby OPITZ (1998: Fig. 68; 2006: Fig. 39a; 2010: Fig. 92). Moreover, the completereduction of the metatarsus to four tarsomeres in Anthicoclerinae [synonymized underClerinae by BOUCHARD et al. (2011: 348) but resurrected herein] although a significantevolutionary change, represents, and gives an indication of, a magnitude of evolutionaryplasticity in the expression of the tarsomere structure. Moreover, the molecular surfacingof the Clerinae (sensu latu) � Korynetinae (sensu latu) gap is bound to become apparentin any comprehensive morphologic or molecular study. This has always been consideredan obvious break in the family, but the real issue is what manner of discontinuity(characteristic gap) is worthy of subfamily status. If we regard the moleculardiscontinuities, exhibited in GUNTER et al. (2013), worthy of subfamily status then wemight suggest that in the Cleridae we have only two subfamilies, Clerinae andKorynetinae; an idea that would produce a very unbalanced classification in the family.
Epiphloeinae taxonomic historyThe history of the Epiphloeinae taxon begins with Thomas Say who in 1825
described Enoplium dislocatum (SAY 1825: 176). Then, Laporte described Ichnealycoides (LAPORTE 1836: 55). Laporte was followed by SPINOLA (1841: 75) who classifiedIchnea Laporte and Epiphloeus Spinola under the informal name Ichnoïdes. AGASSIZ(1846: 193) Latinized Ichnoïdes to Ichneoidea and credited the name to Spinola. Thisaction by Agassiz made Ichneoidea available for subfamily nomenclatural consideration;however as pointed out by OPITZ & HERMAN (2009: 183), the correct name for thesubfamily under consideration is Epiphloeinae Kuwert as governed by the provisions ofArticles 23.9.1 and 23.9.2 of the International Commission on Zoological Nomenclature(1999: 115).
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

8
W. OPITZ
Subsequent proliferations of epiphloeine species description were generated byKLUG (1842: 356), SPINOLA (1844b: 38), ERICHSON (1847: 86), THOMSON (1860: 60),CHEVROLAT (1874: 323), GORHAM (1877a: 246; 1882: 166), KUWERT (1893: 492), HORN(1896: 374), and SCHENKLING (1900: 397). During more modern times species and generawere added to Epiphloeinae by WOLCOTT (1927: 92), CHAPIN (1927: 5), and OPITZ (1997:51, 2004: 1, 2006: 97, 2007: 77, 2008a: 1, 2008b: 1, 2008c: 1, 2010a: 1, 2010b: 1, 2011a:63, 2011b: 133).
MaterialsSince 1997, when part I of this series of works was published (OPITZ 1997), I
examined more than a thousand epiphloeine specimens. The majority of my studyanimals were borrowed from various national and international collections. These arelisted in my generic revisions as noted in the section of references. Special efforts weremade by some colleagues, named in the above mentioned revisions, to collect and liquidpreserve epiphloeines [primarily in Pampel�s fixative as described in EKIS (1977: 6); nowOpitz]. This made possible exploration of the taxonomic value of the mesodermal organs.Various collecting expeditions throughout the new world facilitated procurement of fluidpreserved specimens, information about beetle-floral associations, and information aboutbeetle behavior.
MethodsThe optical equipment used in the preparation of habitus illustrations involved a
Nikon DXM1200 digital camera attached to a Leica MZ microscope. Illustrations of themorphology of the various genera that comprise Epiphloeinae were detailed in revisionsthat specifically dealt with these genera. The publications in which such illustrations arepresent are noted in the reference section.
Sources for entomological orismology involved NICHOLS (1989) and CALAHAN(1975: 390) whose efforts contributed greatly to my understanding terminology foridentifying antennal sensilla. Institutional acronyms were taken from ARNETT Jr., et al.(1993). To formulate taxa epithets I relied on BROWN (1956) and various Latindictionaries. In the catalogue portion of this treatise only post-CORPORAAL (1950a)citations are included under the species listings.
My hypotheses concerning species status are based on the canons of the biologicalspecies as advocated by STANDFUSS (1896: 115), DOBZHANSKY (1937: 312), and MAYR(1963: 19), and hypotheses of supraspecific relationships stem from the concepts ofHENNIG (1966: 88). Elsewhere (OPITZ 2010c: 48) I discuss characters, and their variousstates, that seem to provide clues of �generic level� morphological gaps; fullyrecognizing the subjectivity in traditional considerations in the assignment of rank tosupraspecific taxa. For example, in my experience, the tarsal spur formula and the tarsalpulvillar formula have been shown to be very consistent in predictions of intragenericspecies congruity within Cleridae.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

9
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Revisionary systematists are experiencing fortunate times in light of the availablesoftware options for phylogenetic analyses. These can be useful tools to augment diligentyears of mental phylogenetic thoughts. Moreover, manually prepared phylogenies can beas useful as computer generated schemes, as attested by often similar results (OPITZ 2006:155; 2008a: 15; 2008b: 28). In some of my generic revisions hypotheses of phylogenywere prepared manually. I also implemented the computer programs Hennig86 and, mostrecently, NONA in combination with Winclada version 1.00.08.
The following codons indicate the repository of types and they generally follow thesuggestions of ARNETT Jr. et al. (1993).
AMNH . . . . . . . . . American Museum of Natural History, Department of Entomology, Central ParkWest at 79th Street, New York, NY 10024-5192 (Lee Herman; [email protected])
BMNH . . . . . . . . . . . . . British Museum of Natural History, Department of Entomology, SW 5BD,London, England (Beulah Garner; [email protected]
Maxwell V. L. Barclay; [email protected])CASC . . . . . . . . . California Academy of Sciences, Department of Entomology, Golden Gate Park,
San Francisco, California 94118 (David H. Kavanaugh [email protected] D. Penny; [email protected])
CMNC . . . . . . . . Canadian Museum of Nature, Insect Collection, Post Office Box 3443, Station D,Ottawa, Ontario, Canada K1P 6P4 (Robert S. Anderson; randerson@mus-natur-ca. Francois
Genier; [email protected])CMNH . . . . . . . . Carnegie Museum of Natural History, Invertebrate Zoology, 4400 Forbes Avenue,
Pittsburgh, Pennsylvania 15213 (Robert L. Davidson: [email protected])CNCI . . . . . . . . . . . Agriculture-Food Canada, K.W. Neatby Building, 960 Carling Avenue, Ottawa,
K1A OC6, Canada (Patrice Bouchard: [email protected])CNIN . . . . . . . . Instituto de Biología, UNAM, Departamento de Zoología, Aparado Postal 70-153,
Ciudad Universitaria 04510 México D. F., México (Santiago Zaragoza: [email protected])
DEIG . . . . . . . . . . Deutsches Entomologisches Institute, Leibniz-Zentrum für Agrarlandschaffs-undLandnutzungsforschung e. V. Ebersvalde Str. 84, D-15374 Müncheberg, Germany
(Lutz Behne: [email protected])DZUP . . . . . . . . . . . . . . . . Universidade Federal do Paraná, Departamento de Zoologia, Colecão de
Entomologia, Caixa Postal 19020, 81531-990, Curitiba, Paraná, Brazil (Germano H. Rosado-Neto)
EMEC . . . . . . . . . . Essig Museum of Entomology, University of California, College of Agriculture,Division of Entomology and Parasitology, California Insect Survey, Berkeley,
California 94720 (Pete Oboyski; [email protected])FDZC . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Fernando de Zayas Collection, Havana, CubaFMNH . . . . . . . . Field Museum of Natural History, Department of Entomology, Roosevelt Road at
Lake Shore Drive, Chicago, Illinois 60605 (James H. Boone; [email protected])FSCA . . . . . . . . . . . . . . . Florida State Collection of Arthropods, Division of Plant Industry, Florida
Department of Agriculture, P. O. Box 147100, Gainesville, Florida 32614-7100 (Mike Thomas; [email protected]. Paul E. Skelley; [email protected])
IAVH . . . . . . . . . Istituto de Investigación de Recursos Biológicos Alexander von Humbolt, Carrera7 No. 35-20, Bogotá D. C., Colombia (José Enrique Castillo; [email protected])
IMLA . . . . . . . . . . Fundacion Miguel Lillo, Dirección de Zoologia, Miguel Lillo 251, Entomologia,,4000 San Miguel de Tucumán, Argentina
(Virginia Colomo de Correa; [email protected])INBC . . . . . . . . . . . Instituto Nacional de Bioversidad. Santo Domingo de Heredia, Apartado Postal
22-3100, Heredia, Costa Rica (Angel Solis; [email protected])INHS . . . . . . . . . Illinois Natural History Survey, Center for Biodiversity, 607 East Peabody Drive,
Champaign, Illinois 61820-6970 (Kathleen R. Zeider; [email protected])
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

10
W. OPITZ
IZAV . . . . . . . . . . . . . . Universidad Central de Venezuela, Facultad de Agronomia, Departamento eInstituto de Zoologia Agricola, Apartado Postal 4579, Maracay 2101-A, Venezuela
(José Clavijo; [email protected]) LACM . . . . . . . . . . . . . Natural History Museum of Los Angeles County, Entomology Section, 900
Exposition Boulevard, Los Angeles, California 90007 (Brian V. Brown; [email protected])MCMC . . . . . . . Museo de Historia Natural de la Ciudad de México, Apartado 18845, México, D.FMCNZ . . . . . . . . . Fundacão Zoobotãnica do Rio Grande do Sur, Museo de Ciências Naturais, Rua
Dr. Salvador Franca, 1427 Caixa Postal 1188, 90001-970, Porto Alegre, RS, Brasil (M.H. M. Galileo; [email protected])
MCZC . . . . . . . . . Museum of Comparative Zoology, Harvard University, Entomology, Cambridge,Massachusetts 02138 (Philip D. Perkins; [email protected])
MIUP . . . . . . . . . . Universidad de Panamá, Museo de Invertebrados G. B. Fairchild, Departamentode Zoología, Estafeta Universitaria, Panamá, Panamá
(Roberto Cambra T. Diomedes Quintero; [email protected]) MLPA . . . . . . . . . . . . . Universidad Nacional de la Plata, Facultad de Ciencias Naturales Y Museo,
Division Entomologia, 1900 Paseo del Bosque, La Plata, Argentina (Liliana A. Fernández;[email protected])
MNHN . . . . . . . . . . Museum d�Histoire Naturelle, Entomologie, 45 bis, Rue de Buffon, Paris (Ve),France (Antoine Mantilleri; [email protected])
MRSN . . . . . . . . . . . . . . Museo Regionale di Scienze Naturali, Via Giolitti 36, 10123, Torino, Italy (Luca Piccian; [email protected])
MZSP . . . . . . . . . . Museu de Zoologia Universidade de Sao Paulo, Caixa Postal 42.694 01064-970,Sao Paulo, Brazil (Cleide Costa; [email protected])
QCAZ . . . . . . . . . Pontifica Universidad Catolica del Ecuador, Departamento de Biologia, Avenida12 de Octubre, entre Patria y Beintilla, Apartado 17-01-2184, Quito, Ecuador
(Gionanni Onore; [email protected])SEAN . . . . . . . . . . . Museo Entomologico. S. E. A., A.P. 527, Leon, Nicaragua (Jean-Michel Maes;
[email protected])SEMC . . . . . . . . . . . The University of Kansas, Snow Entomological Division, The Natural History
Museum of the University of Kansas, Lawrence, Kansas 66045-2454 (Zachary Falin; [email protected])
TAMU . . . . . . . . Texas A & M University, College of Agriculture and Life Sciences, Department ofEntomology, Minnie Belle Heep Building, College Station, Texas 77843-7029
(Edward G. Riley; [email protected])UASC . . . . . . . . . Museo Historio Natural, Noel Kempff Mercado, Santa Cruz de la Sierra, Bolivia
(Julieta Ledzema; [email protected])USNM . . . . . . . . United States Department of Agriculture. Systematic Entomology Laboratory, c/o
National Museum of Natural History MRC 168, Washington, D.C. 20560-0165 (Natalia J. Vandenberg; [email protected])
ZMHB . . . . . . . . . . . . . . . . . . . . . . . Museum für Naturkunde, Institute für Systematische Zoologie, Invalidenstrasse 43, D-10115, Berlin, Germany
(Bernd Jaeger; [email protected])
Natural historyAs adults, many species of the Cleridae have made an art out of evolving color,
shape, and behavioral characteristics that minimize their vulnerability to predators duringtheir own exposed predatory activities. Mimicry is abound in this beetle family(HESPENHEIDE 1973: 51; EKIS 1977: 4; MENIER 1985: 1071; MAWDSLEY 1994: 115, andOPITZ 2005: 13). Among the Epiphloeinae we may add cryptic coloration to the deceptiongenre. Based on field observations of various authors it can be said that most, if not all
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

11
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
species of epiphloeines, spend most of their life in association with bark of recently felledtrees or of standing diseased trees (GORHAM 1886: 341; CHAMPLAIN 1920: 637; BALDUF1935: 108; BEAL & MASSEY 1945: 76; CRAIGHEAD 1950; HESPENHEIDE 1973: 51; GRUNER1974: 120; OPITZ 2004: 13; 2008b: 5).
The evolution towards a bark niche existence has in some genera resulted insubstantial structural homogeneity, to an extent that interspecific relationships could notbe discerned via adult morphology (OPITZ 2004: 61; 2007: 143). In fact, my hypothesesof evolution involved almost exclusively morphological apotypies relevant to species-group levels. In several genera, such as Epiphloeus Spinola and Ichnea Laporte, bodyform and integumental coloration have evolved towards a Batesian mimetic plan referredas Batesian polymorphism (JORON & MALLET 1998: 461). Moreover, in the latter genusBatesian mimicry not only involves forebody color characteristics, but also the catapultescape behavior of lignicolous flies (HESPENHEIDE 1973: 51).
Published information about epiphloeine immature stages suggests that larvaeundergo several instars and that the mature larva �overwinters� during adverseenvironmental conditions (CHAMPLAIN 1920: 637). Larval instars consume the immaturestages of lignicolous insects with bark beetles being the predominant prey. The numberof generations of these checkered beetle predators appears to be codependent on thegeneration frequency of prey species (GRUNER 1974). Further, epiphloeines have beenassociated with a great variety of beetle infested hardwoods with oaks and mahoganybeing frequently mentioned, in the temperate and tropical zones, respectively. Lastly,details about epiphloeine/floral associations are included in the individual genericrevisions of the subfamily (OPITZ 2004: 13; 2006: 107; 2007: 88; 2008a: 16; 2008b: 5;2008c: 16).
Character and character states found useful for the discernment of genera within Epiphleinae
I began this project with the resolve to assign generic status to a taxon only when Ilearned of at least one synapotypy for that taxon. In time, I discovered that it was notpossible to follow this strict edict of Hennigian methodology without sacrificing somelikely monophyletism, and possibly distort reasonable balance in morphological gaps inthe preparation of classifications. Therefore, part of the consideration when assessingrank to morphological gaps involved evaluation of gap magnitude to encouragetaxonomic stability, increase classification balance, and maximize the utilitarian andheuristic value in categorization of species within Epiphloeinae.
The following set of adult morphological characteristics was found useful towardsfinding generic-level synapotypies and morphological gaps to discriminate Epiphloeinaegenera. Antenna: In general, epiphloeine antennae are capitate with variations of antennalcomponents involving length of the scape, shape of the pedicel, shape of the funicularantennomeres, relative combined length of the funicle, shape of the capitulumantennomeres, and relative length of the antennal setae.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

12
W. OPITZ
Mouthparts: The terminal palpomeres of the labium and maxilla are almost alwaysdigitiform; however, the maxillary terminal palpomere may be subsecuriform. Themandible although usually broad triangular may be falciform. Pronotum: The pronotum may be transverse, quadrate, oblong rectangulate, ortrapezoidal.Metafemur: Usually the metafemur is narrowing elongate, but can be robustly broadened.Pygidium: The posterior margin of the pygidium is usually fully pigmented, rarely setinward by a triangular membranous region.Aedeagus: The phallic plates usually coalesce posteriorly to form an uncomplicatedphallic apex; however, the phallic apex may show spring-like acuminate projections.Male mesodermal reproductive organs: Most commonly, there are two pairs of maleaccessory glands, but a reduction to one pair is also found in the subfamily.Female mesodermal reproductive organs: The female organs feature a well-developedbursa copulatrix, spermathecal capsule that is slightly sclerotized, and a spermathecalgland that originates near the apex of the capsule.
Subfamily Epiphloeinae Kuwert, 1893: 492Type genus. Epiphloeus Spinola, 1841: 75.
KUWERT 1893, OPITZ (1997: 51, 2004: 1, 2006: 97, 2007: 77, 2008a: 1, 2008b: 1, 2008c: 1, 2010b: 1, 2010c:76, 2011a: 63, 2011b: 133), OPITZ & HERMAN 2009: 183.
Differential diagnosis. The most convenient characteristics to identify epiphloeinespecimens are to observe the attachment of the antenna to the cranium, at the lowerportion of the eye margin, and the presence of four trichobothria on the pronotum.Description. Integument: Integument vested profusely with erect and suberect setae onthe cranium, eyes, pronotum, elytra, legs, and abdomen.
Head: Strongly deflexed; as wide or wider than the anterior margin of the pronotum;cranium most often finely punctate; eyes finely grained by very small ommatidia;antenna comprised of 8 to 11 antennomeres, capitate and with funicular antennomeresslightly expanded, triangular, or narrowly transverse, funicular antennomeres sometimesstrongly compressed as group; cranium moderately punctate or deeply punctate,sometimes with setal aggregates near eyes or on vertex; clypeus distinct, very narrow;labrum distinct, incised anteriorly; mandibles triangular-arcuate, robust, rarely falciform,usually with well-developed dens; maxilla prominent, with laterolacinia, terminalpalpomere usually digitiform, rarely subsecuriform; labium prominent, terminalpalpomere usually digitiform, rarely curved-rectangular; gula broadly triangular, suturesarcuate and converging; two setose postgular processes present.
Thorax: Pronotal form quadrate, transverse, elongate, or trapezoidal, moderatelyconvex; lateral tubercle well developed, faintly visible, or absent; anterior transversedepression present or not, surface finely punctate, coarsely punctate, or cribrate; discaland lateral pair of trichobothria well developed; procoxal cavities open behind, pronotal
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

13
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
projections slightly extended to middle; dorsolateral carina not extended to anteriormargin of pronotum, carina confluent posteriorly with pronotal hem; metendosternitewith one exception without furcal lamina; prosternum short in front of coxae;metasternum moderately developed, broad; epimeron scarcely visible; femora usuallyslender and elongated, rarely robustly broad; tibial spur formula 0-1-1, rarely 0-0-1; tarsalpad formula 3-3-2 or 3-3-1; fourth metatarsomere minute; unguis with well-developeddenticle; mesoscutellum triangular or quadrate; elytral form oblong rectangular, oblongsubovoid; or narrow triangular, punctations vary in size and arranged in rows or not,elytra covering abdomen entirely; epipleural fold extended to elytra apex or not;epipleural margin with or without trichobothria.
Abdomen: With six visible sterna; sixth visible sternum of males usually incised indistal margin; pygidium usually entire, rarely set with triangular depigmentation;aedeagus tubular; tegmen lobed at posterior extremity; phallobasic apodeme welldeveloped; phallobasic rod usually prominent; ventral tegminal sinus usually moreprominent than dorsal tegminal sinus; phallus consists of two phallic plates that unite toform blunt phallic apex; intraspicular plate of spicular fork linear; female genitalia,paraprocts with long bacilli; valvifers with long bacilli; coxites nearly dividedtransversally into two subunits; stylus small and setose; dorsal and ventral laminavariously incised.
Mesodermal internal organs: Alimentary canal with ventricular crypts slightlydeveloped, bulbous at middle, and with long narrow recurviture; ileum very short,proximal half of colon narrow, remainder bulbous; rectum gradually decreasing indiameter posteriorly; proventriculus bulgy from exterior view, internally stomodaealintima with spines to anterior region of proventriculus; pharyngeal intima with fourprimary and two secondary folds; stomodaeal valve comprised of four primary lobes,dorsal and ventral lobes short, former broad; four cryptonephridial malpighian tubules;female organs comprised of well-developed but not encapsulated spermatheca;spermathecal gland attached to subapex of spermatheca; with well-developed saccularbursa copulatrix; ovary usually comprised of 12 follicles; male organs comprised of oneor two pairs of accessory glands; testis comprised of six to 22 follicles.
Description of the external morphology of known immature EpiphloeinaeThis description of epiphloeine larvae is based on three species of the subfamily:
Madoniella dislocata (Say), Pennasolis merkeli (Horn), and Pyticeroides laticornis(Say).Description. Form: Elongate vermiform; head slightly narrower or about as wide asthoracic segments, latter slightly wider or as wide as abdominal segments.
Head: Prognathus, transverse or subquadrate, subglobose or triangular in lateralview; frontal sutures prominent; two rows of stemmata present, front row of three nearbase of mandible, back row of two near frontal suture; antenna comprised of threeantennomeres; mandible falcate, apex unidentate, retinaculum distinct; maxillary cardoand stipes trapezoidal, palpifer feebly sclerotized, palpus comprised of three palpomeres;
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

14
W. OPITZ
labial mentum and submentum slightly sclerotized, palpus comprised of threepalpomeres.
Thorax: Prothorax slightly longer than wide, tergal shield slightly sclerotized;prosternal plate well developed, extended to coxae; legs short, comprised of fivesegments that taper to unguiform ungues.
Abdomen: With large intersegmental membranes, segments without sclerotizationexcept ninth segment with or without basal plate and cerci.
Key to the genera of EpiphloeinaeCheckered beetles belong to the subfamily Epiphloeinae if the eyes are finely
grained by very small ommatidia; the antenna are set into the cranium at the lower marginof the eye distad to the eye notch; the pronotum shows four highly developedtrichobothria, with two present on the disc near the posterior hind angles and two on thelateral deflection of the disc; with one exception, the metendosternite lacks a furcallamina; and the tibial spur formula is almost always 0-1-1.
1 Metatarsus showing two pulvilli. ............................................................ 21� Metatarsus showing one pulvillus. ......................................................... 82(1) Antenna comprised of 11 antennomeres. ................................................. 32� Antenna comprised of 10 antennomeres. ................................................ 73(2) Terminal maxillary palpomere subsecuriform, not digitiform. ................ 43� Terminal maxillary palpomere digitiform. .............................................. 54(3) Pronotum minutely roughened, punctations indistinct; width between
eyes about same as eye width in dorsal view (Brazil). ........ Opitzius Barr4� Pronotum not minutely roughened, punctations distinct; width between
eyes smaller than eye width in dorsal view. .............. Epiphloeus Spinola5(3�) Elytral disc not very roughly sculptured (México to Brazil). ......................
.................................................................................. Megaphloeus Opitz5� Elytral disc very roughly sculptured. ....................................................... 66(5�) Body form short and somewhat wedge shaped (Brazil). .........................
....................................................................................... Pteroferus Opitz6� Body form long and rectangular (Colombia to Bolivia). ............................
.................................................................................. Turbophloeus Opitz7(2�) Basal antennomere of antennal capitulum longer than combined length of
funicular antennomeres (French Guiana to Argentina). ................................................................................................................. Iontoclerus Opitz
7� Basal antennomere of antennal capitulum shorter than combined lengthof funicular antennomeres (Haiti). .................................. Pericales Opitz
8(1�) Antenna comprised of less than 10 antennomeres. ................................. 98� Antenna comprised of 10 antennomeres. .............................................. 10
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

15
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
9(8) Antenna comprised of 8 antennomeres; eyes nearly contiguous in dorsalview (Brazil). ............................................................ Diapromeces Opitz
9� Antenna comprised of 9 antennomeres; eyes not nearly contiguous indorsal view (USA to Argentina). ............................. Pyticeroides Kuwert
.10(8�) Funicular antennomeres from moderately triangular to strongly
transverse. ............................................................................................. 1110� Funicular antennomeres not strongly triangular, possible slightly
expanded but not triangular. ................................................................. 1711(10) Funicular antennomeres strongly compressed as a group. ..................... 1211� Funicular antennomeres not compressed as a group. ............................ 1412(11) Combined length of funicular antennomeres much shorter than basal
antennomere of antennal capitulum (Costa Rica to Brazil). .......................................................................................................... Stegnoclava Opitz
12� Combined length of funicular antennomeres about as long as basalantennomere of antennal capitulum. .................................................... 13
13(12�) Side margins of pronotum with dense ridge of yellow setae; elytral discwith strongly defined long ridges; pygidium without membranoustriangle (Brazil). ................................................... Acanthocollum Opitz
13� Side margins without ridge of yellow setae; elytral disk without stronglydefined long ridges; pygidium with membranous triangle (USA toArgentina). ..................................................................... Ichnea Laporte
14(11�) Combined length of funicular antennomeres much longer thanlength of basal antennomere of antennal capitulum (México toArgentina). ..................................................................... Amboakis Opitz
14� Combined length of funicular antennomeres about as long as basalantennomere of antennal capitulum. ..................................................... 15
15(14�) Antennal capitulum conspicuously slender (Panamá to Brazil). ................................................................................................. Parvochaetus Opitz
15� Antennal capitulum not conspicuously slender. .................................... 1616(15�) Sixth antennomere large, quadrate. ...................... Hapsidopteris Opitz16� Sixth antennomere small, narrow transverse (México to Honduras,
Bolivia). ........................................................................ Opitzia Nemésio17(10�) Pronotum conspicuously oblong. ........................................................ 1817� Pronotum not oblong. ........................................................................... 1918(17) Elytral disc dull in sheen, disc with several pale elevations (Nicaragua).
................................................................................... Katamyurus Opitz18� Elytra bright in sheen, disc without elevations (Costa Rica to Brazil). ......
.................................................................................... Ellipotoma Spinola19(17�) Pronotum very narrow transverse. ........................................................ 2019� Pronotum broad transverse, subquadrate. ............................................. 21
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

16
W. OPITZ
20(19) Antenna with long filamentous setae (México to Paraguay). ................................................................................................. Plocamocera Spinola
20� Antennal without filamentous setae (Bolivia, Brazil). .................................................................................................................. Decaphloeus Opitz
21(19�) Elytral disc with densely setose conate tubercles (México). ...................................................................................................... Megatrachys Opitz
21� Elytral disc without densely setose conate tubercles. ............................ 2222(21�) Dorsum vested with very stout black setae that are particularly prominent
on cranium and pronotum (Brazil). .......................... Silverasia Nemésio22� Dorsum not vested with long stout setae. ............................................. 2323(22)� Seventh row of elytral punctations (from suture to epipleuron) abruptly
terminates at midelytron (Dominican Republic). ............. Decorosa Opitz23� Seventh row of elytral punctations does not terminate at midelytron. ... 2424(23�) Elytral disc with pair of paralateral depressions (USA). ...........................
..................................................................................... Pennasolis Opitz24� Elytra not depressed at sides. ................................................................. 2525(24�) Eighth antennomere as long as combined length of funicular
antennomeres; pronotum mostly yellow, with narrow centrally locatedbrown line (Brazil). .................................................... Decaphloeus Opitz
25� Eighth antennomere much shorter than combined length of funicularantennomeres; pronotal coloration never as described above (Canada toArgentina). ........................................................................ Madoniella Pic
D e s c r i p t i o n o f g e n e r a
Acanthocollum Opitz, 2010a: 2. (Fig. 1)OPITZ 2010b: 8.Type species: Enoplium melanurum Klug, 1842: 376 (by original designation).
Number of species. 1.Differential diagnosis. The dense setal tuft on the upper sides of the pronotum and thedistinct elongate setal ridges on the elytral disc will conveniently distinguish themembers of this genus among other epiphloeines.Apotypies. Pronotum with setal ridges and elytral disc with pronounced carina.Description. Size: Large specimens, length about 11.0 mm, width about 3.0 mm.
Form: Somewhat triangular. Head: Head and pronotum vested densely with aggregates of yellow setae; vertex
between eyes wide; antenna comprised of 10 antennomeres, scape with distinct carina,pedicel and funicular antennomeres transverse; medial tormal processes transverse andcontiguous; mandible broad triangular, apex subacuminate; mandibular penicillus welldeveloped; maxillary laterolacinia present; pronotal form trapezoidal.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

17
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Thorax: Pronotal anterior transverse depression absent, pronotal bothria domed;elytral punctations small and diffusely distributed; elytra flared towards the back;protibial anterior margin with 16 spines; tarsal spur formula 0-1-1; tarsal pulvillarformula 3-3-1.
Abdomen: Aedeagus tubular; phallic apex consists of rounded knob.Distribution. Central and southeastern Brazil.
Amboakis Opitz, 2006: 118. (Figs 2�29)OPITZ 2008a: 4.Type species. Teutonia nova Opitz, 1997: 63 (by original designation).
Number of species. 29.Differential diagnosis. The triangular shape of the funicular antennomeres incombination with the distinctly rowed elytral punctations will separate the members ofthis genus from other epiphloeines.Apotypies. The triangular condition of the funicular antennomeres represents anintermediate condition between the subfiliform stage and the distinctly transverse stage.Therefore, triangular funicular antennomeres are considered apotypic in Amboakisspecies.Description. Size: Small specimens, length about 5.0 mm, width 1.5 mm.
Form: Variable, short oblong subovoid, short narrow oblong, long narrow oblong.Head: Subquadrate in frontal view, usually wider than width of pronotum; vertex betweeneyes wide; antenna comprised of 10 antennomeres, pedicel subglobose, funicularantennomeres triangular; mandible broad triangular, apex subacuminate, penicillus welldeveloped; maxillary laterolacinia well developed.
Thorax: Pronotal anterior transverse depression usually developed at sides only,rarely fully developed; pronotal bothrium partially domed; elytral punctations large orsmall, if large arranged into rows; elytral form spatulate; anterior margin of protibia with1 to 10 spines; tibial spur formula 0-1-1, tarsal pulvilar formula 3-3-1.
Abdomen: Aaedeagus tubular; phallobasic rod usually prominent; phallic apexconsists of rounded knob; two pairs of male accessory glands.Distribution. Central México to northern Argentina.
Decaphloeus Opitz, 2010a: 10. (Fig. 30)Type species. Epiphloeus vitticollis Schenkling, 1900: 397 (by original designation).
Number of species. 1.Diagnosis. From the superficially similar specimens of Megaphloeus, members ofDecaphloeus may be distinguished by the antenna, which is comprised of 10antennomeres, not 11 as is the case in Megaphloeus specimens.Apotypies. The pronotal disc is adorned with a linear dark line.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

18
W. OPITZ
Description. Size: From small to large specimens, length 5.0�10.0 mm, width 1.2�1.4.mm.
Form: Oblong, short subrectangulate. Head: Slightly wider than pronotum; vertex narrow; eyes bulgy; antenna comprised
of 10 antennomeres, pedicel oblong, funicular antennomeres subfiliform; mandible broadtriangular, penicillus well developed; maxillary laterolacinia well developed; lastpalpomere of maxilla and labium digitiform.
Thorax: Pronotal anterior transverse depression well developed, bothrium notdomed; elytral punctations not arranged in rows, elytra broad spatulate; tibial spurformula 0-1-1; tarsal pulvillar formula 3-3-1; protibial anterior margin with 8 spines.
Abdomen: Aedeagus, aedeagal information not available. Distribution. Known only from Bolivia and Brazil.
Decorosa Opitz, 2008a: 4. (Figs 31�34)Type species. Decorosa iviei Opitz, 2008: 8 (by original designation).
Number of species. 4.Differential diagnosis. The most convenient characteristic that distinguishes Decorosaspecimens from those of other epiphloeine genera is the absence of the 7th row ofpunctations from the anterior half of the elytral disc. Also, the labial palpomeres arecurved-rectangular.Apotypies. Two synapotypies have been found in this genus; one, the 7th row of elytralpunctations is absent in the anterior half of the elytral disc; and two, the labial terminalpalpomere is curvate-rectangular.Descriptions. Size: Small specimens, length about 4.0 mm, with about 1.2 mm.
Form: Narrow rectangulate. Head: Much wider than pronotum; vertex between eyes very wide; antenna
comprised of 10 antennomeres, pedicel oblong, funicular antennomeres subfiliform;mandible broad triangular, apex subacuminate, penicillus poorly developed; maxillarylaterolacinia well-developed; last maxillary palpomere curved-subconic; last labialpalpomere curved-rectangular.
Thorax: Pronotal anterior transverse depression absent; elytral punctations rowed;7th row of elytral punctations absent in basal half of elytral disc; elytra spatulate; anteriormargin of protibia with one to three spines; tibial spur formula 0-1-1; tarsal pulvillarformula 3-3-1.
Abdomen: Aedeagus tubular; phallobasic rod prominent; phallic plates very narrow;phallic apex consists of rounded knob. Distribution. Insular genus from the Dominican Republic.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

19
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Diapromeces Opitz, 1997: 58. (Fig. 35)Type species. Diapromeces aclydis Opitz, 1997: 59 (by original designation).
Number of species. 1.Differential diagnosis. These small, narrow, somewhat wedge-shaped beetles, can bemost conveniently separated from other epiphloeines by the number of antennomeres thatcomprise the antenna, which is eight.Apotypies. There are four uniquely derived characteristics; one, the antenna is comprisedof eight antennomeres; two, the eyes occupy the major portion of the head; three, thecranium is substantially extended behind the eyes; and four, the tarsal spur formula is 0-0-1.Description. Size: Small specimens, length about 6.0 mm, width about 1.2 mm.
Form: Narrow long rectangulate. Head: As wide as pronotum; eyes occupy most space of head, eyes nearly
contiguous when viewed from above; cranium extended substantially posteriorly behindeyes; antenna comprised of eight antennomeres, funicular antennomeres subfiliform;mandible narrow triangular, apex acuminate, penicillus highly reduced; maxillarylaterolacinia well developed; last palpomeres of maxilla and labium digitiform, tapereddistally.
Thorax: Pronotum narrow oblong, pronotal anterior transverse depression absent,bothrium domed; elytral punctations rowed, elytra narrow, but gradually widened toapex; protibial anterior margin with 5 to 7 spines; tibial spur formula 0-0-1; tarsalpulvillar formula 3-3-1.
Abdomen: Aedeagus tubular; phallobasic rod absent; phallic apex consists ofrounded knob.Distribution. This monotypic genus is known only from southern Brazil.
Ellipotoma Spinola, 1844b: 36. (Figs 36�37)OPITZ 1997: 60, 2006: 145.Type species. Ellipotoma tenuiformis Spinola 1844b: 38 (by monotypy).
Number of Species. 2.Differential diagnosis. These beetles may be distinguished, among epiphloeines, by theiruniformly slender body form, oblong narrow pronotum, shallow, not bulgy, eyes, narrowvertex to the point that the eyes are nearly contiguous, nine rows of punctations insteadof the typical 10; elytral interstitial spaces shiny smooth, and absence of 2° elytral setae.Apotypies. This taxon shows four uniquely derived characteristics: one, body formextraordinarily slender; two, elytral punctations arranged into nine rows; three, elytraldisc without 2° setae; and four, phallic struts with central rod.Description. Size: Small specimens, length about 5.0 mm, width about 1.2 mm.
Form: Long and uniformly slender.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

20
W. OPITZ
Head: About as wide as pronotum; vertex between eyes narrow; eyes not bulgy;antenna comprised of 10 antennomeres, pedicel oblong, funicular antennomeressubfiliform or slightly expanded; mandible narrow triangular, apex acuminate, penicilluspoorly developed; maxillary laterolacinia well developed; last maxillary palpomeredigitiform; last labial palpomere curvate digitiform.
Thorax: Pronotal anterior transverse depression absent, pronotal bothrium withdome; elytral punctations arranged into nine rows, 2° elytral setae absent, elytra narrowspatulate; protibial anterior margin with 10 spines; tibial spur formula 0-1-1; tarsalpulvilli formula 3-3-1.
Abdomen: Aedeagus tubular; phallobasic rod present; phallic struts with centralrods; phallic apex consists of rounded knob. Distribution. Honduras to Brazil.
Epiphloeus Spinola, 1841: 75. (Figs 38�45)OPITZ 1997: 53, 2008b: 5.Type species. Enoplium duodecimmaculatus Klug, 1842: 370 (subsequent designation by CORPORAAL 1950a:
253).
Number of species. 8.Differential diagnosis. In the members of this genus the maxillary terminal palpomeresare subsecuriform, a form of maxillary palpomere unique among the epiphloeines. Also,in these beetles the scape is as long as the combined length of the pedicel and funicularantennomeres combined, and there are large elytral punctations that abruptly diminish insize at about elytral middle.Apotypies. There are six uniquely derived characteristics: one, scape very long; two,ocular setal tuft present; three, antennal club abbreviated; four, last maxillary palpomeresubsecuriform; five, vertex with narrow furrow; and six, pygidium very large.Description. Size: Large specimens, length about 8.0 mm, width about 2.5 mm.
Form: Oblong subovate. Head: Wider than pronotum; vertex between eyes about fourth of width of eye; eyes
very bulgy; frontal and epicranial margins of eyes with white setal tufts; antennacomprised of 11 antennomeres; scape very long curvate; pedicel oblong; funicularantennomeres subfiliform; antennal capitulum reduced in size; mandible broad triangular,apex subacuminate, penicillus well developed; maxillary laterolacinia well developed;last maxillary palpomere subsecuriform; last labial palpomere digitiform.
Thorax: Pronotal anterior transverse depression well developed; pronotal archtransversely wrinkled or not, bothrium not domed; elytral punctations large or small,when large restricted to elytral basal half; elytra oblong ovate; anterior margin of protibiawith 7 to 13 spines; tibial spur formula 0-1-1; tarsal pulvillar formula 3-3-2.
Abdomen: Aedeagus tubular, phallobasic rod present; phallic apex consist ofrounded knob. Distribution. Nicaragua, south to Venezuela and east to French Guiana.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

21
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Hapsidopteris Opitz, 1997: 62. (Fig. 46)OPITZ 2008c: 5.Type species. Hapsidopteris diastenus Opitz, 1997: 63 (by original designation).
Number of species. 1.Differential diagnosis. Only in the members of this epiphloeine species is the antennalcapitulum apparently comprised of four antennomeres; the seventh antennomere iscryptic, diminutive.Apotypies. Sixth antennomere exceptionally large.Description. Size: Small specimens, length about 5.0 mm, width about 2 mm.
Form: Oblong rectangulate. Head: Wider than pronotum; vertex between eyes wide; eyes bulgy; antenna
comprised of 10 antennomeres, sixth antennomere extraordinarily large and seventhdiminutive, pedicel transverse, funicular antennomeres highly variable in size; mandiblenarrow triangular, apex acuminate; last maxillary and labial palpomere digitiform.
Thorax: Pronotal anterior depression absent; elytral punctations arranged into rows,elytra narrow spatulate; tibial spur formula 0-1-1; tarsal pulvillar formula 3-3-1; protibialanterior margin with two spines.
Abdomen: Aedeagus tubular; phallic apex consists of rounded knob.Distribution. México: Veracruz.
Ichnea Laporte, 1836: 55. (Figs 47�65)OPITZ 2010b: 17.Type species. Ichnea lycoides Laporte 1836: 55 (by monotypy).
Number of species. 19.Differential diagnosis. The most distinguishing characteristic of the members of thisepiphloeine genus is the triangular depigmented patch on the distal region of thepygidium.Apotypies. Triangular pygidial depigmented patch present. Description. Size: Medium to large-sized specimens, length about 8.0 mm, width 3.0mm.
Form. Narrow triangular or oblong subovoid. Head: About as wide as pronotum; vertex between eyes wide or very narrow; eyes
bulgy; antenna comprised of 10 antennomeres, pedicel transverse, funicularantennomeres transverse; mandible narrow triangular, apex acuminate, penicillus poorlydeveloped; maxillary laterolacinia well developed; last maxillary and labial palpomeresdigitiform.
Thorax: Pronotal anterior transverse depression absent, bothrium domed; elytralpunctations arranged into rows in anterior third of elytral disc but diffusely distributed inremainder, elytra flared posteriorly or oblong subovate; tibial spur formula 0-1-1; tarsalpulvillar formula 3-3-1; protibial anterior margin with 9 to 14 spines.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

22
W. OPITZ
Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consists ofrounded knob. Distribution. Southwestern United States to northeastern Argentina.
Iontoclerus Opitz, 1997: 56. (Figs 66�67)OPITZ 2008c: 8.Type species. Enoplium humerale Klug 1842: 373 (by original designation).
Number of species. 2.Differential diagnosis. Epiphloeines beetles belong to this genus if they have twometatarsal pulvilli, 10 antennomeres, and funicular length as long, or slightly shorter,than the length of the basal antennomere of the antennal capitulum.Apotypies. Elytra vested profusely with silvery 2° setae; mesoscutellum transverse-oval.Description. Size: Large specimens, length about 7.0 mm, width 2.5 mm.
Form: Oblong rectangulate. Head: About as wide as pronotum; vertex between eyes narrow; eyes bulgy; antenna
comprised of 10 antennomeres, pedicel globose, funicular antennomeres somewhatserrate; mandible broad triangular, apex subacuminate, penicillus well developed;maxillary laterolacinia well developed; maxillary and labial last palpomere digitiform.
Thorax: Pronotal anterior transverse depression feebly impressed, bothrium notdomed; elytral punctations not arranged in rows, very small and profusely randomlydistributed, elytra broad spatulate; tibial spur formula 0-1-1; tarsal pulvillar formula 3-3-2
Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consists ofrounded knob.Distribution. French Guiana south to northeastern Argentina.
Katamyurus Opitz, 1997: 60. (Figs 68�69)OPITZ 2008c: 15.Type species. Katamyurus paxillus Opitz, 1997: 97 (By original designation).
Number of species. 1.Differential diagnosis. Long slender epiphloeines with pale elevations on the elytral discbelong to this genus.Apotypies. Pale elevations on the elytral disc, phallic apex with spring-like processes,and spicular fork apodemes separated.Description. Large specimens, length about 6.0 mm; width 1.5 mm.
Form: Long and slender, pronotum particularly oblong and slender. Head: About as wide as pronotum; vertex between eyes narrow; eyes not bulgy;
antenna comprised of 10 antennomeres, pedicel oblong, funicular antennomeressubfiliform; mandible broad triangular, apex subacuminate, penicillus well developed;maxillary laterolacinia well developed; last maxillary and labial palpomere digitiform.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

23
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Thorax: Pronotal anterior transverse depression absent; elytral punctationsarranged in rows; elytra narrow spatulate; tibial spur formula 0-1-1; tarsal pulvillar
formula 3-3-1; protibial anterior margin with 10 spines. Abdomen: Aedeagus tubular; phallobasic rod present; phallic plates broad, apex
with spring-like processes.Distribution. Southern México.
Madoniella Pic, 1935: 10. (Figs 70�147)OPITZ 2011b: 143.Type species. Madoniella minor Pic 1935: 10 (subsequent designation by CORPORAAL 1950a: 306).
Number of species. 78.Differential diagnosis. The elytral insignia is the most convenient characteristic toidentify specimens of many species of this genus. The difficulty lies in that the insigniahas been reduced in a few species, and in one species lost entirely. However, the key togenera, provided herein, readily will isolate the members of this genus. Apotypies. Elytral insignia present.Description. Size: Small specimens, length about 5.0 mm, width 1.5 mm.
Form: Short oblong quadrate or rarely short oblong subovoid. Head: Slightly wider than pronotum; vertex between eyes wide; antenna comprised
of 10 antennomeres, pedicel oblong, funicular antennomeres subfiliform; mandible broadtriangular, apex subacuminate, penicillus well developed; maxillary laterolacinia welldeveloped; last maxillary and labial palpomere digitiform.
Thorax: Pronotal anterior transverse depression usually not present, bothria domed;elytral punctations usually large and arranged into rows, elytra short spatulate; tibial spurformula 0-1-1; tarsal pulvillar formula 3-3-1; protibial anterior margin with 1 to 7 spines.Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consists of roundedknob.Distribution. Eastern Canada to east of the Rocky Mountains in the USA, and south toArgentina.
Megaphloeus Opitz, 2010a: 16. (Figs 148�173)OPITZ 2011a: 63Type species. Epiphlaeus setulosus Thomson, 1860: 60 (by original designation).
Number of Species. 26.Differential diagnosis. Size: From superficially similar members of Epiphloeus,Megaphloeus specimens may be distinguished by the digitiform condition of the terminalpalpomere of the maxilla.Apotypies. Phallic apex large triangular.Description. Size: From small to large specimens, length 4.0�10.0 mm, width 1.3�3.0 mm.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

24
W. OPITZ
Form: Oblong subrectangulate. Head: Slightly wider than pronotum; vertex narrow; eyes bulgy; antenna comprised
of 11 antennomeres, pedicel oblong, funicular antennomeres subfiliform; mandible broadtriangular, penicillus well developed; maxillary laterolacinia well developed; lastpalpomere of maxilla and labium digitiform.
Thorax: Pronotal anterior transverse depression well developed, bothrium notdomed; elytral punctations not arranged in rows, elytra broad spatulate; tibial spurformula 0-1-1; tarsal pulvillar formula 3-3-2; protibial anterior margin with 3 to 11spines. Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consists ofrounded knob.Distribution. Southeastern México to southern Brazil.
Megatrachys Opitz, 1997: 61. (Figs 174�176)OPITZ 2008: 21.Type species. Megatrachys paniculus Opitz, 1997: 61 (by original designation).
Number of species. 3.Differential diagnosis. The members of this genus have an elytral surface that iscorrugated and set with setose tubercles.Apotypies. Elytral disc corrugated; elytral disc with setose tubercles.Description. Size: Large specimens, length about 8.0 mm, width about 2.0 mm.
Form: Oblong rectangulate. Head: About as wide as pronotum; vertex between eyes wide and set with setal tufts;
eyes bulgy; antenna comprised of 10 antennomeres, pedicel oblong, funicularantennomeres subfiliform; mandible broad triangular, apex subacuminate, penicillus welldeveloped; maxillary laterolacinia well developed; last palpomere of maxilla and labiumdigitiform.
Thorax: Pronotal anterior transverse depression absent, bothrium domed; elytralpunctations not discernable from above, elytra long spatulate, elytral disc corrugated andwith setose tubercles; tibial spur formula 0-1-1; tarsal pulvilli formula 3-3-1; protibialmargin with 4 spines.
Abdomen: Aedeagus tubular; phallic apex consist of subacuminate knob.Distribution. Known only from southern México.
Opitzia Nemésio, 2005: 77. (Figs 177�178)Type species. Arenaria chiapas Opitz, 1996: 57 (by original designation).
Number of species. 2.Differential diagnosis. Epiphloeines belong to this genus if they are of medium size(about 8.0 mm), have every funicular antennomere transverse, have a dorsum that ismostly yellow-red, or all yellow-red, whose venter is entirely dark brown, whose elytralsides are sharply deflexed, and who have more than 12 rows of elytral punctations.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

25
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Apotypies. Eyes small; greater than 10 rows of elytral punctations; most distal spine ofanterior margin of protibia very large; body black at the flanks; 7th antennomereacuminate. Description. Size: Large specimens, length about 8.0 mm, with about 2.5 mm.
Form: Long rectangulate. Head: Wider than pronotum; vertex between eyes very wide; eyes small; antenna
comprised of 10 antennomeres, pedicel transverse, funicular antennomeres transverse;mandible slender triangular, penicillus well developed; maxillary laterolacinia welldeveloped; last palpomere of maxilla and labium digitiform.
Thorax: Pronotal anterior transverse depression absent, bothria slightly domed;elytral punctations arranged into 14 rows; elytra narrow spatulate; tibial spur formula 0-1-1; tarsal pulvillar formula 3-3-1; protibial anterior margin with 5 spines.
Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consists ofrounded knob.Distribution. Southern México.
Opitzius Barr, 2006: 215. (Fig. 179)OPITZ 2008b: 29.Type species. Opitzius thoracicus Barr 2006: 215 (by original designation).
Number of species. 1.Differential diagnosis. The known members of this genus have the pronotal discminutely roughened.Apotypies. Pronotal disc minutely roughened.Description. Size: Large specimens, length about 8.0 mm, width about 2.5 mm.
Form: Oblong, slightly subovate. Head: Head slightly wider than pronotum; vertex between eyes wide; eyes bulgy;
antenna comprised of 11 antennomeres, scape very long curvate, pedicel oblong,funicular antennomeres subfiliform; mandible broad triangular, apex subacuminate,penicillus well developed; maxillary laterolacinia well developed; last maxillarypalpomere subsecuriform; last labial palpomere digitiform.
Thorax: Pronotal anterior transverse depression present, bothria not domed; elytralpunctations large in elytral basal half, smaller in remainder, not rowed, elytra spatulate,side margin slightly swollen at middle; tibial spur formula 0-1-1; tarsal pulvilli formula3-3-2; protibial anterior margin with 10 spines.
Abdomen: Aedeagus tubular; phallic apex triangular.Distribution. Known only from Brazil.
Parvochaetus Opitz, 2006: 109. (Figs 180�184)Type species. Parvochaetus fucolatus Opitz, 2006: 113 (by original designation).
Number of species. 5.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

26
W. OPITZ
Differential diagnosis. The extraordinarily slender form of the antennal club and theswollen condition of the epipleural fold distinguish the members of this genus.Apotypies. Antennal club slender; epipleural fold swollen.Description. Size: Small specimens, length about 4.5 mm, width about 1.5 mm.
Form: Short rectangulate; head wider than pronotum; vertex between eyes broad;eyes bulgy; antenna comprised of 10 antennomeres, pedicel transverse, funicularantennomeres expanded; mandible broad triangular, apex subacuminate, penicillus welldeveloped; maxillary laterolacinia well developed; last maxillary and labial palpomeredigitiform.
Thorax: Pronotal anterior transverse depression absent; pronotal bothrium domed;elytral punctations rowed; elytra short spatulate; tibial spur formula 0-1-1; tarsal pulvillarformula 3-3-1; protibial anterior margin with 4 spines.
Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consists ofrounded knob.Distribution. From western Panamá to southern Brazil.
Pennasolis Opitz, 2008c: 27. (Figs 185�187)RIFKIND, TOLEDO & CORONA 2010: 54. Type species. Phyllobaenus merkeli Horn 1896: 374 (by original designation).
Number of species. 3.Differential diagnosis. The depressions on the sides of the elytral disc, particularly smallantennal club, and an extraordinary transverse head will conveniently distinguish thesebeetles from others in Epiphloeinae.Apotypies. There are four derived characteristics: one, pronotal setal bases elevated; two,elytral disc with elongated depressions; three, head extraordinarily transverse; and four,phallobasic apodeme very short.Description. Size: Medium size, length about 5.0 mm, width about 1.2 mm.
Form: Short rectangulate. Head: Wider than pronotum; vertex broad; eyes bulgy; antenna comprised of 10
antennomeres, pedicel globose, funicular antennomeres subfiliform; mandible broadtriangular, penicillus well developed; maxillary laterolacinia well developed; lastmaxillary and labial palpomere digitiform.
Thorax: Pronotal anterior transverse depression notable at sides, bothria not domed;elytral disc with distinct concavities at sides, subcorrugated, punctations variouslynotable on disc; elytra short spatulate; tibial spur formula 0-1-1; tarsal pulvillar formula3-3-1; protibial anterior margin with nine spines.
Abdomen: Aedeagus tubular; phallobasic rod present; phallobasic apodeme veryshort; phallic apex consists of rounded knob. Distribution. Western USA and south/central México.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

27
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Pericales Opitz, 2008c: 32. (Fig. 188)Type species. Pericales albogilvus Opitz, 2008c: 33 (by original designation).
Number of species. 1.Differential diagnosis. Only in specimens of this genus are there whorls of shallowridges on the pronotal disc.Apotypies. Pronotal disc with whorls of shallow ridges; funicular antennomeres serrate.Description. Size: Large specimens, length about 6 mm, width about 2.0 mm. Form:Long rectangulate.
Head: About as wide as pronotum; vertex between eyes wide; eyes bulgy; antennacomprised of 10 antennomeres, pedicel suboval, funicular antennomeres serrate;mandible broad triangular, apex subacute, penicillus well developed; maxillarylaterolacinia well developed; last maxillary and labial palpomere digitiform.
Thorax: Pronotal anterior transverse depression absent, bothria not domed, elytralpunctations small not rowed; tibial spur formula 0-1-1; tarsal pulvillar formula 3-3-2;protibial anterior margin with 4 spines.
Abdomen: Aedeagus tubular; phallobasic apodeme short; phallobasic rod present;phallic apex consist of rounded knob.Distribution. Known from Haiti and the Dominican Republic.
Plocamocera Spinola, 1844b: 17. (Figs 189�225)OPITZ 2004: 23.Type species. Plocamocera sericella Spinola 1844b: 19 (by monotypy).
Number of species. 37.Differential diagnosis. The most conspicuous characteristic that will identify themembers of this genus is the �plumose� condition of the antenna. Extended from theantennomeres are very long filamentous setae.Apotypies. There are five apotypies: one, antenna with very long filamentous setae; two,femora swollen; pronotum boldly transverse; three, metacoxa exceptionally bulbous; andfour, abdominal color sexually dimorphic.Description. Size: Medium specimens, length from 4.0 to 8.0 mm, width from 1.5 to 2.8mm.
Form: Elongate subovate. Head: Wider than pronotum; vertex between eyes narrow; eyes bulgy; antenna
comprised of 10 antennomeres, antennomeres vested with long filamentous setae, pedicelsuboval, funicular antennomeres short, slightly swollen; mandible broad triangular, apexsubacute, penicillus absent; maxillary laterolacinia well developed; last maxillary andlabial palpomere digitiform.
Thorax: Pronotal anterior transverse depression present, bothria not domed; elytraldisc sub corrugated, elytral punctations not easily seen; tibial spur formula 0-1-1; tarsalpulvillar formula 3-3-1; protibial anterior margin with five spines.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

28
W. OPITZ
Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consist ofrounded knob or triangular apex.Distribution. From México to Paraguay.
Pteroferus Opitz, 2008c: 33. (Fig. 226)Type species. Pteroferus zolnerowichi Opitz, 2008c: 34 (by original designation).
Number of species. 1.Differential diagnosis. The short triangular body form in combination with 11antennomeres and corrugated elytral disc will conveniently distinguish these beetles fromothers in Epiphloeinae.Apotypies. Short triangular body form; highly corrugated elytral disc.Description. Size: Medium size, length about 5.0 mm, width about 1.5 mm.
Form: Short triangular. Head: Wider than pronotum; vertex broad; eyes bulgy; antenna comprised of 11
antennomeres, pedicel ovoid, funicular antennomeres subfiliform, mandible broadtriangular, penicillus well developed; maxillary laterolacinia well developed; lastmaxillary and labial palpomere digitiform.
Thorax: Pronotal anterior transverse depression well developed, pronotal archelevated, pronotal disc highly undulated, bothria not domed; discal punctations not easilydiscernable; elytra short, epipleural margin diverging; tibial spur formula 0-1-1; tarsalpulvillar formula 3-3-2; protibial anterior margin with four spines.
Abdomen: Aedeagus very narrow and long, tubular; phallobasic rod present;phallobasic apodeme very long; phallic apex consists of rounded knob. Distribution. Southern Brazil.
Pyticeroides Kuwert, 1894: 7. (Figs 227�265)OPITZ 2007: 87.Type species. Pyticeroides arrogans Kuwert 1894: 9 (by monotypy).
Number of species. 39.Differential diagnosis. The most conspicuous characteristic that will identify themembers of this genus is that the antenna is comprised of nine antennomeres.Apotypies. Antenna comprised of nine antennomeres; cranium concave behind eyes.Description. Size: Medium specimens, length from 4.0 to 7.0 mm, width from 1.0 to 2.4mm.
Form: Long rectangulate, broad short rectangulate, or broad long rectangulate.Head: Wider than pronotum; vertex between eyes moderately wide; eyes bulgy;
antenna comprised of nine antennomeres, pedicel suboval, funicular antennomeresshort, transverse; mandible narrow triangular, apex acute, penicillus well developed;maxillary laterolacinia well developed; last maxillary and labial palpomere digitiform.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

29
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Thorax: Pronotal anterior transverse depression present, bothria not domed; elytralpunctations rowed; tibial spur formula 0-1-1; tarsal pulvillar formula 3-3-1; protibialanterior margin with four to nine spines.
Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consist ofrounded knob or triangular apex.Distribution. From eastern Canada to northern Argentina.
Silverasia Nemésio, 2005: 77. (Fig. 266)Type species. Chaetophloeus hispidus Opitz, 2004: 21 (by original designation).
Number of species. 1.Differential diagnosis. The exceptionally stout and profusely distributed primary elytralsetae will distinguish the members of this species from other epiphloeines. Apotypies. Exceptionally stout 1° setae on elytral disc.Description. Size: Small, length about 4.5 mm, width about 1.5 mm.
Form: Short oblong subovate. Head: Wider than pronotum; vertex broad; eyes bulgy; antenna comprised of 10
antennomeres, pedicel globose, funicular antennomeres subfiliform; mandible broadtriangular, penicillus not discernable; maxillary laterolacinia well developed; lastmaxillary and labial palpomere digitiform.
Thorax: Pronotal anterior transverse depression notable at pronotal sides, pronotalbothria not domed, large discal punctations; elytra short somewhat arcuate at epipleuralmargin; tibial spur formula 0-1-1; tarsal pulvillar formula 3-3-1; protibial anterior marginwith three spines.
Abdomen: Aedeagus tubular; phallobasic rod present; phallobasic apodeme long;phallic apex consists of rounded knob. Distribution. Southern Brazil.
Stegnoclava Opitz, 2010a: 25 (Figs 267�269)OPITZ 2010b: 9.Type species. Ichnea fumigata Gorham 1877b: 414 (by original designation).
Number of species. 3.Differential diagnosis. The drastic shortening of the funicular antennomeres and theblack mark on the frons will easily identify the members of this genus.Apotypies. Funicular antennomeres drastically shortened; frons with black mark; scapevery short. Description. Size: Large specimens, length about 8.0 mm, width 2.5 mm.
Form: Very slightly oblong triangular. Head: Wider than pronotum; vertex between eyes wide; eyes bulgy; antenna
comprised of 10 antennomeres, scape particularly short, pedicel transverse, funicular
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

30
W. OPITZ
antennomeres very shortened; mandible narrow triangular, apex acuminate, penicilluswell developed; maxillary laterolacinia well developed; last maxillary and labialpalpomeres digitiform.
Thorax: Pronotal anterior transverse depression noted at sides of pronotum,bothrium domed; elytral punctations not arranged into rows, diffusely distributed, elytraslightly flared posteriorly; tibial spur fur formula 0-1-1; tarsal pulvillar formula 3-3-1;protibial anterior margin with 11 to 14 spines.
Abdomen: Aedeagus tubular; phallobasic rod present; phallic apex consists ofrounded knob. Distribution. Northeastern Costa Rica to south-central Brazil.
Turbophloeus Opitz, 2008c: 35. (Fig. 270)Type species. Epiphloeus simplex Schenkling 1900: 397 (by original designation).
Number of species. 1.Differential diagnosis. Epiphloeine specimens that have 11 antennomeres, are longrectangulate, and have an extensively roughened elytral disc belong to this genus.Apotypies. Furcal lamina absent.Description. Size: Medium-sized specimens, length about 6.0 mm, width 1.6 mm.
Form: Oblong rectangulate. Head: Wider than pronotum; vertex between eyes wide; eyes bulgy; antenna
comprised of 11 antennomeres, pedicel oblong, funicular antennomeres subfiliform;mandible broad triangular, apex subacuminate, penicillus very small; maxillarylaterolacinia well developed; last maxillary and labial palpomeres digitiform.
Thorax: Pronotal anterior transverse depression well developed, bothrium notdomed; elytral punctations not arranged into rows, diffusely distributed, elytron longspatulate; tibial spur fur formula 0-1-1; tarsal pulvillar formula 3-3-2; protibial anteriormargin with six spines.
Abdomen: Aedeagus very long and slender, tubular; phallobasic rod present; phallicapex consists of rounded knob. Distribution. Colombia to Bolivia.
Characters selected for phylogenetic analysis of Epiphloeinae generaSeventy-seven morphological, distributional, or mimetic adult characters of
epiphloeine genera, and the outgroup genus Pilosirus Opitz (NIXON & CARPENTER 1993:423) were used to establish the evolutionary states of character variations. PilosirusOpitz, is removed herein from the subfamily Epiphloeinae and transferred to PeloniinaeOpitz, where it is linked evolutionarily to other genera within Peloniinae Opitz that arecharacterized by having a pair of incipient bothria on each side of the pronotum.Character states designated as �0� are considered plesiotypic whereas those assigned avalue of �1� are judged apotypic. The phylogeny (Fig. 271) was prepared by computer
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

31
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
via NONA (GOLOBOFF 2003) in concert with Winclada version 1.00.08, and involved 9steps, index of consistency 82, and an index of retention of 85. A heuristic analysis(maximum trees (hold) = 100, number of replication 1 (mult) = 100, multiple TBR = TBR(mult max used).
Character statesCharacter 0. Some level of bothrial development: (0) no; (1) yes.Character 1. Pronotal bothrium: (0) not incipient; (1) incipient.Character 2. Pronotal bothrium development: (0) incipient; (1) fully developed.Character 3. Insertion of antenna: (0) near middle of ocular notch; (1) lower limit of
ocular notch.Character 4. Furcal lamina: (0) present; (1) absent.Character 5. Number of metatarsal pulvilli: (0) 3; (1) 2.Character 6. Number of metatarsal pulvilli: (0) 2; (1) 1.Character 7. Number of antennomeres: (0) 11; (1) less than 11. Character 8. Shape of scape: (0) not boldly convex; (1) boldly convex.Character 9. Elytral trichobothria: (1) not present; (1) present.Character 10. Size of 2° setae: (0) not minute; (1) minute.Character 11. Abundance of 2° setae: (0) not very abundant; (1) very abundant.Character 12. Sculpture of pronotal arch: (0) not transversally wrinkled; (1) transversally
wrinkled.Character 13. Length of scape: (0) not very long; (1) very long.Character 14. Size of punctations on elytral basal half: (0) not very large; (1) very large.Character 15. Size of pygidium: (0) not extraordinarily large; (1) extraordinarily large.Character 16. Elytral disc: (0) without elevated diagonal ridges; (1) with elevated
diagonal ridges.Character 17. Ommatidia: (0) large; (1) small.Character 18. Number of antennomeres: (0) not 10; (1) 10.Character 19. Number of antennomeres: (0) not less than 10; (1) less than 10.Character 20. Shape of pedicel: (0) not globose; (1) globose.Character 21. Funicular antennomeres: (0) not very expanded; (1) very expanded.Character 22. Funicular antennomeres: (0) not compacted; (1) very compacted.Character 23. Model in mimicry: (0) not Lycidae; (1) Lycidae.Character 24. Shape of pedicel: (0) not transverse; (1) transverse.Character 25. Sixth antennomere: (0) not quadrate; (1) quadrate.Character 26. Elytral sides: (0) not strongly deflexed; (1) strongly deflexed. Character 27. Carina on scape: (0) absent; (1) present.Character 28. Funicular antennomeres: (0) not subfiliform; (1) subfiliform.Character 29. Elytral punctations near sutural margin: (0) not serially arranged; (1)
serially arranged.Character 30. First degree setae: (0) not very stout; (1) very stout.Character 31. Pronotal shape: (0) not oblong; (1) oblong. Character 32. Elytral disc coloration: (0) not ornate; (1) ornate.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

32
W. OPITZ
Character 33. Length of metatarsus: (0) not very long; (1) very long.Character 34. Number of accessory glands: (0) two pairs; (1) one pair.Character 35. Second degree setae: (0) not aggregated into setal tufts; (1) aggregated into
setal tufts.Character 36. Elytral interstitial spaces: (0) not smooth; (1) smooth.Character 37. Funicular antennomeres: (0) not expanded; (1) slightly expanded.Character 38. Elytral setal tuft: (0) not yellow; (1); yellow.Character 39. Elytral setal tuft: (0) not white; (1); white.Character 40. Ocular setal tuft: (0) not preset; (1) present.Character 41. Pronotal disc: (0) not asperous; (1) asperous.Character 42. Pronotal lateral carina: (0) not entire; (1) entire.Character 43. Elytral humeral inverted T: (0) absent; (1) present.Character 44. Funicular antennomeres: (0) not gradually and progressively serrate; (1)
gradually and progressively serrate.Character 45. Distribution: (0) not insular; (1) insular.Character 46. Body form: (0) not lanceolate; (1) lanceolate.Character 47. Phallic-strut rod: (0) absent; (1) present.Character 48. Frons: (0) wide; (1) narrow.Character 49. Eye size: (0) not very small; (1) very small.Character 50. Elytral disc: (0) without insignea; (1) with insignea.Character 51. Elytral punctations: (0) not large throughout elytral surface; (0) large
throughout elytral surface.Character 52. Eyes depth: (0) normal; (1) very shallow.Character 53. Antennal sensilla trichodea: (0) not longer than usual; (1) longer than
usual.Character 54. Antennal sensilla trichodea: (0) not filamentous; (1) filamentous.Character 55. Chaetosomes: (0) not profuse on dorsum; (1) profuse on dorsum.Character 56. Elytral disc: (0) without conic tumescences; (1) with conic tumescences.Character 57. Elytral disc: (0) without irregular ridges; (1) with irregular ridges.Character 58. Elytra: (0) not flattened laterally; (1) flattened laterally.Character 59. Vertex setal tuft: (0) not yellow; (1) yellow.Character 60. Epipleural fold: (0) not convex; (1) convex.Character 61. Funicular antennomeres: (0) not extensively compacted; (1) extensively
compacted.Character 62. Pronotal setal ridges: (0) absent; (1) present.Character 63. Pygidium: (0) not notched; (1) notched.Character 64. Pronotal sides: (0) with 2 angulations; (1) with 3 angulations.Character 65. Epipleural margin: (0) not infuscated; (1) infuscated.Character 66. Antennomere 5: (0) not acuminate; (1) acuminate.Character 67. Antennomere 6: (0) not exceptionally large; (1) exceptionally large.Character 68. Epipleural fold: (0) present; (1) absent.Character 69. Mesoscutellum: (0) not notched; (1) notched.Character 70. Cranial color: (0) without postocular dark streak; (1) with postocular dark
streak.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

33
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Character 71. Spicular fork apodeme: (0) not entirely bifid; (1) entirely bifid.Character 72. Number of antennomeres: (0) not 8; (1) 8.Character 73. Phallic apex: (0) not very large; (1) very large.Character 74. Furcal lamina: (0) not present by character reversal; (1) present by
character reversal.Character 75. Pronotum: (0) without central thin line; (1) with central thin line.Character 76. Phallic apex: (0) not spring-like; (1) spring-like.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
Characters 1 2 3 4 5 6 7Taxa 01234567890123456789012345678901234567890123456789012345678901234567890123456
Pilosirus 11000000000000000000000000000000000000000000000000000000000000000000000000000Turbophloeus 10110100110000001100000000000000000000100000000000000000000000000000000001000Pteroferus 10111100110000001100000000000000000000010000000000000000000000000000000000000Epiphloeus 10111100110011110100000000000000000000001000000000000000000000000000000000000Opitzius 10111100110011110100000000000000000000000110000000000000000000000000000000000Megaphloeus 10111100110010010100000000000000000000000001000000000000000000000000000000000Iontoclerus 10111100011100000100000000000000000000000000100000000000000000000000000000000Pricales 10111100011100000100000000000000000000000000010000000000000000000000000000000Ellipotoma 10111011010000000110100000001101000000000000001110001000000000000000000000000Katamyurus 10111011010000000110100000001101000000000000000000001000000000000000000000001Decorosa 10111011010000000110100000001100100000000000000001010000000000000000000000000Madoniella 10111011010000000110100000001100100000000000000000110000000000000000000000000Plocamocera 10111011010000000110100000001010011000000000000000000110000000000000000000000Silveirasia 10111011010000000110100000001010011000000000000000000010100000000000000000000Pennasolis 10111011010000000110100000001010000100000000000000000000011000000000000000000Megatrachys 10111011010000000110100000001010000100000000000000000000110000000000000000000Amboakis 10111011010000000110100000000000000011000000000000000000000100000000000000000Parvochaetus 10111011010000000110100000000000000011000000000000000000000110000000000000000Decaphloeus 10111011010000000110100000000000000010000001000000000000000000000000000000010Stegnoclava 10111011010000000110011110010000000000000000000000000000000001001000000000000Acanthocollum10111011010000000110010110010000000000000000000000000000000000101000000000000Ichnea 10111011010000000110010110000000000000000000000100010000000000010000000000000Opitzia 10111011010000000110010000100000000000000000000000001000000000001110000000000Hapsidopteris 10111011010000000110010000100000000000000000000000000000000000001101000000000Pyticeroides 10111011010000000101000000000000000000000000000000000000000000000000111000000Diapromeces 10111011010000000101000000000000000000000000001100000000000000000000100110000
Tab. 1. The character state matrix.

34
W. OPITZ
Evolutionary considerationsThe exclusively New World distribution of Epiphloeinae and its sister taxa suggests
that the foundation stock of the subfamily existed in New World terrains, after thebreakup of Gondwana during the Cretaceous, some 180 million years ago. The sistergroup of Epiphloeinae is undoubtedly a group of enopliine genera exampled by PylosirusOpitz. These enopliines exhibits the germinal state of the eventual fully-developedtrichobothrium of Epiphloeinae. The ancestor of this enopliine-epiphloeine stock(ancestor A) presents an incipient form of the trichobothrium. This progenitor yieldedseveral genera of Enopliinae and the basic stock of the Epiphloeinae (ancestor B), whichis characterized by having four fully developed trichobothria, insertion of the antenna atthe lower limits of the ocular notch, and the reduction and eventual absence of the furcallamina.
One line of progenitor B generated descendants that led to ancestor C in which themetathoracic pulvilli were reduced to two. From C there evolved a lineage with the 2°elytral setae becoming diminutive and profusely distributed. This line eventuallygenerated the genera Iontoclerus Opitz and Pericales Opitz. A portion of the C genomeled toward a line of evolution (ancestor E) in which the scape became highly convex. Onebranch of ancestor E evolved towards ancestor F characterized by having ridges on theelytral disc. Progenitor F diverged to produce the sister genera Turbophloeus Opitz andPteroferus Opitz. Ancestor E also derived progenitor G characterized by having apronotum transversally wrinkled and the pygidium became extraordinarily large. Thisprogenitor produced the genus Megaphloeus Opitz and ancestor H distinguished byhaving a long scape and very large punctations at elytral basal half. Ancestor Heventually evolved the genera Epiphloeus Spinola and Opitzius Barr.
Ancestral species B also produced a complementary stock that led to progenitor I,characterized by having one metatarsal pulvillus and less that 11 antennomeres. In oneline, from I, there evolved progenitor J in which the antennae were comprised of less than10 antennomeres and the mesoscutellum is notched. Progenitor J evolved the generaPyticeroides Kuwert and Diapromeces Opitz. Progenitor I also led to ancestor K whosedescendants inherited an antenna comprised of 10 antennomeres. Ancestor K divergedand led to progenitors L and P.
Ancestor L generated two lineages. One lineage led to progenitor M, characterizedby an elytron that is strongly deflexed at the sides, an epipleuron that is stronglyinfuscated, and antennomere 5 became acuminate. Ancestor M evolved the generaOpitzia Nemésio and Hapsidopteris Opitz. Ancestor L led to a lineage (ancestor N) inwhich the pedicel became transverse and the body form Lycid-like. Ancestor N evolvedinto the genus Ichnea Laporte and into a complementary stock that yielded, via ancestorO, the genera Stegnoclava Opitz and Acantocollum Opitz.
The complementary stock of ancestral species K, progenitor P, characterized by aglobose pedicel, generated ancestor Q and the complementary stock progenitor S. In oneline ancestor Q evolved the genus Decaphloeus Opitz and in the complementary stockthat first yielded progenitor R, characterized by having the funicular antennomeres
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

35
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
slightly expanded, which led to the sister genera Amboakis Opitz and ParvochaetusOpitz. Progenitor S dichotomized into ancestral species T and W. Progenitor T evolvedvery robust 1° elytral setae and produced two evolutionary lines that ended in ancestralspecies U and V. Progenitor U, characterized by having the 2° elytral setae aggregatedinto setal tufts, eventually produced the sister genera Pennasolis Opitz and MegatrachysOpitz. The metatarsus became elongated in progenitor V which eventually yielded thesister genera Plocamocera Spinola and Silverasia Nemésio. The complementary stock ofprogenitor S, characterized by having subfiliform funicular antennomeres, evolved intoancestral species W, in which the elytral punctations became serially arranged near thesutural margin. Progenitor W diverged to produce the sister lineages that eventuallyevolved into progenitor X and progenitor Y. In the former ancestor the elytral discbecame ornate and the elytral punctations large. This line yielded the sister generaDecorosa Opitz and Madoniella Pic. The complementary stock, leading to ancestor Y,characterized by having an oblong pronotum, yielded the sister genera EllipotomaSpinola and Katamyurus Opitz.
AcknowledgmentsI am grateful to Jaques Rifkind for his review of the manuscript and to Jiøí Kolibáč
for supporting the publication of this manuscript.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

36
W. OPITZ
C a t a l o g u e o f E p i p h l o e i n a e
Genus Acanthocollum Opitz, 2010a: 2.OPITZ 2010b: 8.Type species: Enoplium melanurum Klug, 1842: 376 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2010b: 8.Distribution: Brazil.Number of species: 1.Acanthocollum melanurum (Klug) (Ichnea), 1842: 376. Lectotype female (ZMHB).
Type locality: Brazil. Fig. 1 (nontype specimen). Distribution: Brazil: Mato Grosso;Rio de Janeiro.
Genus Amboakis Opitz, 2006: 118.OPITZ 2008a: 4. Type species: Teutonia nova Opitz 1997: 63 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2006: 118.Corrigendum: There are corrections in the abovementioned publication: Page 119,couplet 8(7�) the couplet should end with Amboakis epiomidia, n. sp.: Page 160 (clave),couplet 8(7�) the couplet should end with Amboakis epiomidia, n. sp.Distribution: Argentina: Belize, Bolivia, Brazil, Colombia, Costa Rica, Cuba, Honduras,México, Panamá, Peru, and Venezuela. Number of species: 29.Amboakis ampla Opitz, 2010a: 3. Holotype female (USNM). Type locality: Ecuador:
Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 2 (holotype).Distribution: Ecuador: Orellana.
Amboakis anapsis Opitz, 2006: 120. Holotype female (MNHN). Type locality:Venezuela: Apure: Sarare River. Fig. 3 (holotype). Distribution: Venezuela: Apure.
Amboakis antegalba Opitz, 2010a: 5. Holotype male (USNM). Type locality: Ecuador:Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 4 (holotype).Distribution: Ecuador: Orellana.
Amboakis atra Opitz, 2006: 134. Holotype male (IMLA). Type locality: Argentina:Salta: Campo Quijano. Fig. 5 (paratype). Distribution: Argentina: Salta.
Amboakis barinas Opitz, 2006: 122. Holotype female (IZAV). Type locality: Venezuela:Barinas: Rio Socopo. Fig. 6 (holotype). Distribution: Venezuela: Barinas. Colombia:Amazonas.
Amboakis binotonis Opitz, 2006: 124. Type locality: Cuba: Cienfuegos: Soledad.Holotype female (MCZC). Fig. 7 (holotype). Distribution: Cuba: Cienfuegos.
Amboakis capitata (Gorham) (Epiphloeus), 1877a: 248. Lectotype female (BMNH).Type locality: Brazil: Amazonas: Ega. Fig. 8 (paralectotype). Corporaal 1950: 25(Phlogistosternus). Distribution: Brazil: Amazonas; Mato Grosso.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

37
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Amboakis cauca Opitz, 2006: 140. Holotype female (IAVH). Type locality: Colombia:Valle del Cauca: PNN Farallones de Cali, Anchcaya. Fig. 9 (holotype). Distribution:Colombia: Valle del Cauca.
Amboakis charis Opitz, 2006: 126. Holotype male (FSCA).Type locality: Honduras:Olancho: Parque Nacional La Muralla. Fig. 10 (holotype). Distribution: Honduras:Olancho. Panamá: Chiriquí.
Amboakis demagna Opitz, 2010a: 7. Holotype male (USNM). Type locality: Ecuador:Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 11(holotype). Distribution: Ecuador: Orellana.
Amboakis diffusa Opitz, 2010a: 6. Holotype male (FSCA). Type locality: Ecuador:Tungurahua, 38 km E Baños. Fig. 12 (holotype). Distribution: Ecuador: Tugurahua.
Amboakis epiomidia Opitz, 2006: 141. Holotype female (BMNH). Type locality:Panamá: Chiriquí: Tolé. Fig. 13 (paratype). Distribution: Panamá: Chiriquí.
Amboakis erythrohapsis Opitz, 2006: 129. Holotype male (FSCA). Type locality:México: Guerrero: Highway 134, 6.9 km NE junction 200. Fig. 14 (holotype).Distribution: México: Guerrero.
Amboakis flavicollis (Zayas) (Phlogistosternus), 1988: 61. Syntype, gender not known(FDZC). Type locality: Cuba: Oriente: Cabo Cruz. Fig. 15 (nontype specimen).Distribution: Cuba: Cienfuegos; Granma; Oriente; Las Villas.
Amboakis funebris Opitz, 2006: 124. Holotype female (MZSP). Type locality: Brazil:Amazonas: Reserva Ducke. Fig. 16 (holotype). Distribution: Brazil: Amazonas.
Amboakis incondita Opitz, 2006: 134. Holotype male (MNHN). Type locality: Bolivia:Cochabamba: Cochabamba. Fig. 17 (holotype). Distribution: Bolivia: Cochabamba.
Amboakis katatonis Opitz, 2006: 130. Holotype female (FSCA). Type locality: México:Michoacán. Uruapán. Fig. 18 (paratype). Distribution: México: Jalisco: Michoacán.
Amboakis linitis Opitz, 2006: 132. Holotype female (MCZC). Type locality: DominicanRepublic: Mount Diego de Ocampo. Fig. 19 (holotype). Distribution: DominicanRepublic: San Pedro de Macoris
Amboakis mica Opitz, 2006: 135. Holotype female (DZUP). Type locality: Brazil: Pará:Jacareacanga. Fig. 20 (paratype). Distribution: Brazil: Pará.
Amboakis micula Opitz, 2006: 135. Holotype female (AMNH). Type locality: Brazil:Mato Grosso: Diamantino, Facenda Rio Arenos. Fig. 21 (holotype). Distribution:Perú: Huanuco. Bolivia: Cochabamba: Santa Cruz. Brazil: Mato Grosso.
Amboakis nitida (Gorham) (Epiphloeus), 1877a: 248. Type locality: Brazil: Amazonas:Ega. Lectotype female (BMNH). Fig. 22 (paralectotype). Corporaal 1950a: 252(Phlogistosternus). Distribution: Brazil: Amazonas.
Amboakis nova (Opitz), 1997: 63. Holotype male (MZSP). Type locality: Brazil: NovaTeutonia: Santa Catarina. Fig. Habitus (homotype). Distribution: Brazil: Goias;Paraná; Pernambuco; Rio de Janeiro; Santa Catarina; São Paulo.
Amboakis prolata Opitz, 2006: 136. Holotype female (IZAV). Type localiy: Brazil:Paraná: Rondon. Fig. 23 (holotype). Distribution: Brazil: Paraná.
Amboakis rudis Opitz, 2006: 142. Holotype male (MNHN). Type locality: Bolivia:Cochabamba: Cochabamba. Fig. 24 (holotype). Distribution: Bolivia: Cochabamba.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

38
W. OPITZ
Amboakis selva Opitz, 2006: 143. Holotype male (INBC). Type locality: Costa Rica:Heredia: Estación Biologica La Selva. Fig. 25 (paratype). Distribution: Costa Rica:Heredia.
Amboakis stenosis Opitz, 2006: 145. Holotype female (LACM). Type locality: México:Oaxaca: Sierra de Zempoaltepetl, 15.2 km S Benito Juarez. Fig. 26 (holotype).Distribution: México: Oaxaca.
Amboakis taruma Opitz, 2006: 138. Holotype male (MCNZ). Type locality: Brazil:Amazonas: 1 km W Taruma Falls. Fig. 27 (holotype). Distribution: Brazil: Amazonas.
Amboakis vesca Opitz, 2006: 143. Holotype female (MNHN). Type locality: Bolivia:Cochabamba: Cochabamba. Fig. 28 (holotype). Distribution: Bolivia: Cochabamba.
Amboakis waodani Opitz, 2010a: 9. Holotype male (USNM).Type locality: Ecuador:Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 29(holotype). Distribution: Ecuador: Orellana.
Genus Decaphloeus Opitz, 2010a: 10.Type species: Epiphloeus vitticollis Schenkling, 1906: 299 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2010a: 10.Distribution: Bolivia, Brazil.Number of species: 1.Decaphloeus vitticollis (Schenkling) (Epiphloeus), 1906: 299. Lectotype, gender not
known (DEIG). Type locality: Amazon. Fig. 30 (lectotype). Corporaal 1950: 255(Epiphloeus). Distribution: Amazon.
Genus Decorosa Opitz, 2008a: 4.Type species: Decorosa iviei Opitz, 2008a: 8 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008a: 8.Corrigendum: There is a correction in the abovementioned publication: Page 8. UnderDecorosa limatula it should read HOLOTYPE: Male.Distribution: Dominican Republic.Number of species: 4.Decorosa aladecoris Opitz, 2008a: 6. Holotype female (USNM). Type locality:
Dominican Republic: La Vega: Constanza. Fig. 31 (holotype). Distribution:Dominican Republic: La Vega.
Decorosa iviei Opitz, 2008a: 8. Holotype female (USNM). Type locality: DominicanRepublic: San Juan: Pico Duarte weather station. Fig. 32 (holotype). Distribution:Dominican Republic: San Juan, La Vega,
Decorosa limatula Opitz, 2008a: 8. Holotype male (MCZC). Type locality: DominicanRepublic: La Vega: Loma Vieja. Fig. 33 (holotype). Distribution: DominicanRepublic: La Vega.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

39
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Decorosa neiba Opitz, 2008a: 11. Holotype female (CMNH). Type locality: Dominican.Republic: San Juan: Cierra de Neiba, 9.1 km WSW Hondo Valle. Fig. 34 (holotype).Distribution: Dominican Republic: San Juan.
Genus Diapromeces Opitz, 1997: 58.
OPITZ 2007: 138.Type species: Diapromeces aclydis Opitz, 1997: 59 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2007: 138.Distribution: Brazil.Number of species: 1.Diapromeces aclydis Opitz, 1997: 59. Holotype male (AMNH). Type locality: Brazil:
Santa Catarina: Nova Teutonia. Opitz 2007: 138. Fig. 35 (holotype). Distribution:Brazil: Santa Catarina; São Paulo.
Genus Ellipotoma Spinola, 1844b: 36.OPITZ 1997: 60; 2006: 145.Type species: Ellipotoma tenuiformis Spinola, 1844b: 38 (by monotypy).
Generic synonyms: None.Most recent generic revision: OPITZ 2006: 145. Distribution: Honduras. Costa Rica. Panamá. Guyana. Ecuador. Peru. Brazil.Number of species: 2.Ellipotoma tenuiformis Spinola, 1844b: 38. Holotype female (MNHN). Type locality:
Colombia. Fig. 36 (holotype). Corporaal 1950a: 253. Ekis (now Opitz) 1975: 50,Opitz 2006: 150. Distribution: Honduras: Olancho. Costa Rica: Heredia; Puntarenas.Panamá: Chiriquí; Colón; Panamá. Guyana: Mazaruni-Potaro. Ecuador: Tungurahua.Peru: Amazonas, Huánuco. Brazil: Amazonas, Nova Teutonia. Pará.
Ellipotoma turmalis Opitz, 2006: 152. Holotype male (MNHN). Type locality: Bolivia:Cochabamba. Fig. 37 (holotype). Distribution: Bolivia: Cochabamba. Argentina:Salta.
Genus Epiphloeus Spinola, 1841: 75.OPITZ 1997: 53; 2008: 5. OPITZ & HERMAN 2009: 183.Type species: Enoplium duodecimmaculatus Klug, 1842: 370 (subsequent designation by CORPORAAL 1950a:
253).
Generic synonyms: None.Most recent generic revision: OPITZ 2008b.Distribution: Colombia to Brazil.Number of species: 8.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

40
W. OPITZ
Epiphloeus adonis Opitz, 2008b: 21. Holotype female (MNHN). Type locality: FrenchGuiana: Guyane. Fig. 38 (holotype). Distribution: French Guiana: Guyane. Regina.
Epiphloeus duodecimmaculatus Klug, 1842: 370. Holotype female (ZMHB). Typelocality: Brazil: Pará. Fig. 39 (nontype specimen). Corporaal 1950a: 253. Ekis (nowOpitz) 1975: 48, Opitz 2008b: 11. Distribution: Panamá: Panamá. Colombia:Amazonas, Caquetá. Venezuela: Bolivar. Guyana: East Demerara-West CoastBerbice. Peru: Junín. Brazil: Amazonas, Mato Grosso, Pará, Rondônia..
Enoplium pantherinus Chevrolat, 1843: 35.Epiphloeus erwini Opitz, 2010a: 11. Holotype male (USNM). Type locality: Ecuador:
Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 40(paratype). Distribution: Ecuador: Orellana.
Epiphloeus fundurufus Opitz, 2008b: 20. Holotype female (INBC). Type locality: CostaRica: Heredia: Biological Station La Selva. Fig. 41 (paratype). Distribution: CostaRica: Alajuela, Guanacaste, Heredia. Panamá: Panamá. Colombia: Amazonas, Chocó.
Epiphloeus princeps Gorham, 1886: 340. Lectotype female (MNHN). Type locality:Panamá: Chiriquí: Bugaba. Fig. 42 (paratype). Corporaal 1950a: 254. Opitz 2008b:20. Distribution: Panamá: Chiriquí. Ecuador: Orellana.
Epiphloeus pulcherrimus Gorham, 1877a: 246. Lectotype male (BMNH). Typelocality: Brazil: Amazonas: Ega. Fig. 43 (lectotype). Corporaal 1950: 254. Opitz2008b: 21. Distribution: Brazil: Amazonas.
Epiphloeus quattuordecimmaculatus Chevrolat, 1876: 27. Lectotype male (MNHN).Type locality: Brazil. Fig. 44 (homotype). Corporaal 1950a: 254. Opitz 2008b: 13.Distribution: Brazil: Goiás; Guanabara; Santa Catarina. Argentina: Misiones.
Epiphloeus tigrinus Opitz, 2008b: 15. Holotype male (FSCA). Type locality: Panamá:Colón: Fort Sherman. Fig. 45 (paratype). Distribution: Nicaragua: Matagalpa. CostaRica: Alajuela. Panamá: Colón: Panamá.
Genus Hapsidopteris Opitz, 1997: 62.OPITZ 2008c: 5.Type species: Hapsidopteris diastenus Opitz, 1997: 62 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008c: 5. Distribution: México: Veracruz.Number of species: 1.Hapsidopteris diastenus Opitz, 1997: 62. Holotype male (MNHN). Type locality:
México: Veracruz: Jalapa. Fig. 46 (holotype). Opitz, 2008c: 6. Distribution: México:Veracruz.
Genus Ichnea Laporte, 1863: 55.OPITZ 1997: 57; 2010b: 17.Type species: Ichnea lycoides Laporte, 1836: 55 (by monotypy).
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

41
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Generic synonyms: None.Most recent generic revision: OPITZ 2010b.Distribution: Argentina, Bolivia, Brazil, Colombia, Ecuador, French Guiana, Guyana,Honduras, México, Panamá, Paraguay, Peru, United States of America, Venezuela.Number of species: 19.Ichnea acanthomelina Opitz, 2010b: 24. Holotype female. (MZSP). Type locality:
Brazil: Rondônia, 62 km SW Ariquemes, near Fazenda Rancho Grande. Fig. 47(holotype). Distribution: Brazil: Mato Grosso; Pará; Rondônia.
Ichnea aequinoctialis Spinola, 1844b: 22. Neotype male (MIUP). Type locality:Panamá: Colón: Madden forest. Fig. 48 (neotype). Corporaal 1950a: 270. Ekis (nowOpitz), 1975: 49. Opitz 2010a: 29. Distribution: Honduras: Atlántida; Cortés;Olancho; Yoro. Panamá: Boca del Toro; Chiriquí; Colón; Panamá; Darien. Colombia:Boyacá; Chocó; Meta; Magdalena; Valle del Cauca. Venezuela: Amazonas; Aragua;Bolivar; Tachira.Falcón. Guyana: Bartica. French Guiana: Cayenne. Ecuador: Napo.Peru: Huanuco; Junin; Loreto. Bolivia: Cochabamba; La Paz; Santa Cruz. Brazil:Alagoas; Amazonas; Bahia; Espiritu Santo; Goás; Pará; Rio de Janeiro; Rio Grandedo Sul; Rondônia; Santa Catarina; Tocantins. Paraguay: Caazapá, Itapúa. Argentina:Misiones.
Ichnea confluence Kuwert 1894: 12Ichnea disjuncta Gorham 1877b: 411.Ichnea funesta Gorham 1877b: 410.Ichnea histrica Gorham 1883: 178.Ichnea spinolai Corporaal 1949: 398.
Ichnea aterrima (Klug) (Enoplium), 1842: 378. Holotype male (ZMHB). Type locality:México. Fig. 49 (homotype). Corporaal 1950a: 270. Opitz 2010b: 33. Distribution:United States of America: Arizona; New Mexico. México: Chiapas; Jalisco; Morelos;Nayarit; Oaxaca; Tamaulipas; Veracruz. Guatemala: Baja Verapaz; Chiquimula; ElProgresso; Huehuetenango; Izabal; Sacatepéquez; Zacapa. Belize: Orange Walk.Honduras: Atlántida, Cortés, El Paraiso, Francisco Morazán, Olancho. Nicaragua:Granada; Matagalpa. Costa Rica: Cartago; Guanacaste; San José. Panamá: Chiriquí;Panamá. Colombia: Cauca. Venezuela: Falcón. Ecuador: Carchí.
Ichnea kuwerti Lohde 1899: 305.Ichnea elongata Knull 1939: 27.Ichnea mexicana Thomson 1860: 65.Ichnea religiosa Chevrolat 1874: 323.Ichnea vitticollis Gorham 1877b: 415.
Ichnea atra Opitz, 2010b: 26. Holotype male (MZSP).Type locality: Brazil: Amazonas:1 km W Taruma Falls. Fig. 50 (holotype). Distribution: Guyana: Rupunnuni.Suriname: Commewijne. Brazil: Amazonas.
Ichnea callanga Opitz, 2010b: 33. Holotype female (ZMHB). Type locality: Peru:Callanga. Fig. 51 (paratype). Distribution: Colombia: Huila. Bolivia: La Paz. Peru;Cuzco. Brazil: Mato Grosso.
Ichnea digna Wolcott, 1927: 93. Holotype, gender not known (USNM). Type locality:Costa Rica: Alajuela: San Carlos (= Ciudad Quesada). Fig. 52 (nontype specimen).Corporaal 1950a: 270. Opitz 2010b: 38. Distribution: Costa Rica: Alajuela;
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

42
W. OPITZ
Guanacaste; Heredia; Limón. Panamá: Bocas del Toro; Chiriquí; Panamá. Colombia:Cauca.
Ichnea dimidiatipennis Spinola, 1844b: 162. Lectotype female (MRSN). Type locality:Colombia. Fig. 53 (notype specimen). Corporaal 1950a: 270. Ekis (now Opitz) 1975:49. Opitz 2010b: 36. Distribution: Colombia.
Ichnea divisa (Chevrolat) (Enoplium), 1843: 37. Lectotype female (MNHN). Typelocality: Brazil. Fig. 54 (nontype specimen). Corporaal 1950: 272. Opitz 2010b: 40.Distribution: Panamá: Colón, Panamá. Colombia Magdalena.Venezuela: Bolivar.French Guiana: Cayenne. Ecuador: Napo. Peru: Loreto. Bolivia: Santa Cruz. Brazil:Goiás; Mato Grosso; Rondônia; Santa Catarina.
Ichnea enoplioides Spinola, 1844: 25. Ichnea frenata (Erichson) (Enoplium), 1847: 86, Lectotype male (ZMHB). Type
locality: Peru. Fig. 55 (nontype specimen). Corporaal 1950a: 270. Opitz 2010b: 42.Distribution: Nicaragua: Rio San Juan. Costa Rica: Heredia; Puntarenas. Panamá:Chiriquí; Colón; Darien; Panamá. Colombia: Amazonas; Cauca; Meta. Venezuela:Amazonas; Bolivar; Tachira. Peru: Huánuco; Junín; Loreto; Madre de Dios; SanMartín; Tambopata. Ecuador: Esmeraldas; Napo; Sucumbios. Bolivia: Cochabamba;La Paz; Santa Cruz. Brazil: Amazonas; Goiás; Mato Grosso; Pará.
Ichnea confluence Kuwert 1894: 12.Ichnea disjuncta Gorham 1877b: 411.Ichnea femoralis Schenkling 1900: 401.Ichnea mitella Gorham 1877b: 411.Ichnea panamensis Gorham 1883: 179.Ichnea striaticollis Kuwert 1894: 11.Ichnea subfasciata Gorham 1877b: 410.
Ichnea gregata Opitz, 2010b: 36. Holotype male (AMNH). Type locality: Peru: Ancash:Utcuyacu, Tarma Junin. Fig. 56 (holotype). Distribution: Peru: Ancash.
Ichnea gremia Opitz, 2010b: 39. Holotype male (FSCA). Type locality: Ecuador:Esmeraldas, vicinity Bisly. Fig. 57 (holotype). Distribution: Ecuador: Esmeraldas.
Ichnea incerta Gorham, 1877b: 413. Lectotype female (BMNH). Type locality:Amazon. Fig. 65 (non-type specimen). Corporaal 1950a: 271. Distribution: Brazil:Amazonas.
Ichnea lycoides Laporte, 1836: 55. Neotype male (CMNH). Type locality: Brazil:Bahia: Encruzilhada. Fig. 58 (neotype). Corporaal 1950a: 271. Ekis (now Opitz)1975: 49. Menier 1985: 1071. Opitz 2010b: 45. Distribution: Bolivia: La Paz. Brazil:Bahia; Espiritu Santo; São Paulo.
Ichnea marginella (Klug) (Enoplium), 1842: 376. Holotype female (ZMHB). Typelocality: Brazil: Pará. Fig. 59 (homotype). Corporaal 1950a: 271. Ekis (now Opitz)1975: 49. Opitz 2010b: 48. Distribution: México: Sinaloa; Oaxaca; Veracruz.Guatemala: Baja Verapaz. Belize: Cayo. Honduras: Atlántida; Olancho; SantaBárbara. Costa Rica: Cartago; Puntarenas. Panamá: Chiriquí; Colón, Panamá, Darien,Veraguas. Choco. Colombia: Amazonas, Cauca, Magdalena, Meta, Norte deSandaner, Putumayo.Venezuela: Amazonas, Aragua, Bolivar, Monagas, Sucre,Tachira. Trinidad: Arima Borough, Saint Andrew. Guyana: Masaruni-Potaro.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

43
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Surinam: Surinam. French Guiana: Cayenne, Guyane. Ecuador: Napo, Sucumbios.Peru: Huanuco. Bolivia: Cochabamba, Santa Cruz. Brazil: Amazonas, Espírito Santo,Paraná. Paraguay: Alto Paraná.
Hylecoetus cylindricus Germar 1912: 719. Ichnea circumcincta Chevrolat 1874: 324.Ichnea laterale Klug 1842: 377.Ichnea pubescens Spinola 1844b: 25.
Ichnea mimica Gorham, 1877b: 412. Lectotype male (BMNH). Type locality: Brazil:Amazonas: Pará: Pará. Fig. 60 (homotype). Corporaal 1950a: 272. Opitz 2010b: 48.Distribution: Brazil: Pará.
Ichnea opaca (Klug) (Enoplium), 1842: 377. Lectotype male (ZMHB). Type locality:French Guiana: Cayenne. Fig. 61 (homotype). Corporaal 1950a: 272. Ekis (nowOpitz) 1975: 50, Opitz 2010b: 41. Distribution: Guyana: Masaruni-Potaro; Rupununi.Bartica. French Guiana: Cayenne; Régina. Brazil: Amapá.
Ichnea plumbea Gorham, 1877b: 413. Lectotype female (BMNH). Type locality:Brazil: Amazonas. Fig. 62 (nontype specimen). Corporaal 1950a: 272. Opitz 2010b:53. Distribution: Panamá: Panamá. Trinidad: Arima Borpugh. Colombia: Amazonas;Caquetá; Putumayo. Ecuador: Napo. Peru: Loreto. Bolivia: La Paz. Brazil:Amazonas; Mato Grosso; Pará; Rondônia.
Ichnea helvolicollis Corporaal, 1950b: 94.Ichnea praeusta (Klug) (Enoplium), 1842: 376. Holotype male (ZMHB). Type locality:
Brazil. Fig. 63 (nontype specimen). Corporaal 1950a: 272. Opitz 2010b: 37.Distribution: Brazil: Amazonas; Mato Grosso; Bahia; Espíritu Santo; Minas Gerais;Rio Grande do Sul; Santa Catarina. Argentina: Misiones.
Ichnea procera Schenkling, 1900: 399. Lectotype male (DEIG). Fig. 64 (paralectotype).Type locality: Peru: Cuzco: Callanga. Corporaal 1950a: 272. Opitz 2010b: 37.Distribution: Peru: Cuzco. Bolivia: Cochabamba; Santa Cruz.
Genus Iontoclerus Opitz, 1997: 56.OPITZ 2008c: 8.Type species: Enoplium humerale Klug, 1842: 373 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008c: 8. Distribution: Colombia to Argentina.Number of species: 2.Iontoclerus humeralis (Klug) (Enoplium), 1842: 373. Lectotype female (ZMHB). Type
locality: Brazil: Pará. Fig. 66 (homotype). Corporaal 1950a: 254 (Epiphloeus). Ekis(now Opitz) 1975: 48, Opitz 1997: 56. Opitz 2008c: 10. Distribution: Colombia:Putumayo. Guyana: Rupununi. Ecuador: Napo. Brazil: Amazonas: Mato Grosso;Pará; Paraná; Rio Grande do Sul; Rondônia. Peru: Chambireyacú; Madre de Dios.Bolivia: Santa Cruz. Argentina: Misiones.
Apolopha vittate Pic 1936: 127.Epiphloeus marginellus Spinola 1844 b: 15.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

44
W. OPITZ
Iontoclerus sericeus (Klug) (Enoplium), 1842: 373. Lectotype male (ZMHB). Typelocality: Brazil. Fig. 67 (homotype). Corporaal 1950a: 272 (Ichnea). Opitz 2008c: 12.Distribution: Colombia: Putumayo. French Guiana: Cayenne: Ret De L�Amse. Brazil:Amazonas: Mato Grosso; Pará; Rondônia. Ecuador: Napo; Sucumbíos. Peru:Huallaga; Huánuco: Madre de Dios. Bolivia: Cochabamba; La Paz.
Genus Katamyurus Opitz, 1997: 60.OPITZ 2008c: 15Type species: Katamyurus paxillus Opitz, 1997: 60 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008c: 15.Distribution: México to Nicaragua.Number of species: 2.Katamyurus albopaniculus Opitz, 2008c: 16. Holotype female (FSCA). Type locality:
México: Sinaloa: 14 km NE La Cap. De Taxte (= Taste). Fig. 68 (holotype).Distribution: México: Durango; México, D.F.; Sinaloa.
Katamyurus paxillus Opitz, 1997: 60. Holotype female (USNM). Type locality:Nicaragua: Cerro Cimborazo. Fig. 69 (holotype). Opitz 2008c: 16. Distribution:Guatemala: Huehuetenango. Honduras: Olancho. Nicaragua: Matagalpa.
Genus Madoniella Pic, 1935: 10.OPITZ 1997: 62; 2002: 278, 2008a: 4. Type species: Madoniella minor Pic, 1935: 10 (subsequent designation by Corporaal, 1950: 306).
Generic synonyms: Phlogistosternus Wolcott, 1944: 124.Most recent generic revision: OPITZ 2011b: 143.Distribution: Canada, United States of America, Andros Island, México, Guatemala,Cuba, Dominican Republic, Jamaica, Puerto Rico, Dominica, Montserrat, Guana Island,Guadeloupe, Belize, Honduras, Nicaragua, Costa Rica, Panamá, Colombia, Venezuela,French Guiana, Guyana, Brazil, Argentina.Number of species: 78.Madoniella abacula Opitz, 2011b: 55. Holotype male (CMNC). Type locality: México:
Oaxaca: 14 km NW Diaz Ordaz. Fig. 70 (holotype). Distribution: México: Oaxaca. Madoniella adona Opitz, 2011b: 156. Holotype female (MCZC). Type locality:
Dominican Republic: San Pedro de Macoris: Mt. Diego de Ocampo. Fig.71(holotype). Distribution: Dominican Republic: Hato Mayor; San Pedro de Macoris;Santiago.
Madoniella aktis Opitz, 2011b: 179. Holotype female (EMEC). Type locality: México:Chiapas: Portugal. 11.2 km S E Simojovel. Fig. 72 (paratype). Distribution: México:Chiapas.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

45
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Madoniella anapsis Opitz, 2011b: 160. Holotype female (FSCA). Type locality:Dominica: St. Peter: Dublanc. Fig. 73 (paratype). Distribution: Dominica: St. Peter;St. Paul. Guadeloupe: Basse Terre.
Madoniella antennatra Opitz, 2011b: 165. Holotype male (FSCA). Type locality: Honduras: Yoro, Parque Nacional Pico Pijol. Fig. 74 (holotype). Distribution: Honduras:
Yoro.Madoniella apotoma Opitz, 2011b: 180. Holotype male (FSCA). Type locality:
Honduras: Comayagua: 7 km E Siguatepeque. Fig. 75 (holotype). Distribution:Honduras: Comayagua.
Madoniella apsis Opitz, 2011b: 158. Holotype male (FSCA). Type locality: México:Chiapas: El Aguacero. Fig.76 (paratype). Distribution: México: Chiapas.
Madoniella aspera Opitz, 2010: 14. Holotype female (IAVH). Type locality: Colombia:Boyaca: SFF Iquaque Canana Mamaramus. Fig. 77 (holotype). Distribution:Colombia: Boyaca.
Madoniella avina Opitz, 2011b: 199. Holotype male (CMNC). Type locality: Argentina:Salta: El Rey National Park, Rio La Selva. Fig. 78 (holotype). Distribution:Argentina: Salta.
Madoniella basilaris Opitz, 2011b: 161. Holotype female (FSCA). Type locality:Dominican Republic: La Vega, 1 km NW Manaboa. Fig. 79 (paratype). Distribution:Dominican Republic: La Vega; Independencia; Pedernales.
Madoniella basilia Opitz, 2011b: 180. Holotype male (INHS).Type locality: Cuba:Granma: Loma del Gato, Sierra Maestra. Fig. 80 (holotype). Distribution: Cuba:Granma.
Madoniella bilineata (Chevrolat) (Aulicus), 1874: 300. Type locality: Cuba. Primarytype: holotype male (MNHN). Fig. 81 (nontype specimen). Corporaal 1950a: 197(Aulicus). Opitz 2011b: 225. Distribution: Cuba: Granma.
Madoniella bullalis Opitz, 2011b: 164. Holotype female (FSCA). Type locality:Dominican Republic: Monte Cristi, 8.6 km N Villa Elisa. Fig. 82 (holotype).Distribution: Dominican Republic: Monte Cristi; Pedernales.
Madoniella cardinalis Opitz, 2011b: 166. Holotype female (INBC). Type locality: CostaRica: Heredia: Estación Biologica La Selva.Fig. 83 (holotype). Distribution: CostaRica: Heredia; Cartago; Guanacaste; Puntarenas. Panamá: Panamá.
Madoniella careorita Opitz, 2011b: 168. Holotype male (INBC). Type locality: CostaRica: Heredia: Estación Biologica La Selva. Fig. 84 (holotype). Distribution: CostaRica: Alajuela; Heredia; Cartago; Guanacaste.
Madoniella cavina Opitz, 2011b: 204. Holotype male (CMNH). Type locality:Dominican Republic: Pedernales: 23.5 N Cabo Rojo. Fig. 85 (holotype). Distribution:Dominican Republic: Pedernales.
Madoniella cerviculina Opitz, 2011b: 206. Holotype female (CMNH). Type locality:Dominican Republic: Pedernales: 3.3 km NE Los Arroyos. Fig. 86 (holotype).Distribution: Dominican Republic: Pedernales.
Madoniella chiricahua Opitz, 2011b: 212. Holotype male (FMNH). Type locality:Arizona: Cochise County, Chiricahua Mountains. Fig. 87 (paratype). Distribution:United States of America: Arizona. México: Chiricahua; Durango; Sinaloa.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

46
W. OPITZ
Madoniella collata Opitz, 2011b: 200. Holotype female (CMNH). Type locality: Brazil:Goiás: Chapada. Fig. 88 (holotype). Distribution: Brazil: Goiás.
Madoniella corporaali Pic, 1935: 11. Lectoype female (MNHN). Type locality:Guadeloupe: Guadeloupe, Trois-Rivers. Fig. 89 (lectotype). Corporaal 1950a: 306.Opitz 2011b: 157. Distribution: Guadeloupe: Basse Terre; Guadeloupe. Dominica: St.Peter; St. Paul.
Madoniella cracentis Opitz, 2011b: 178. Holotype male (MCNZ). Type locality: Brazil:Amazonas: 1 km W Taruma Falls. Fig. 90 (holotype). Distribution: Colombia:Putumayo. Venezuela: Bolivar. Peru: Huánuco. Brazil: Amazonas.
Madoniella crinis Opitz, 2011b: 213. Holotype male (FSCA). Type locality: México:Tlaxcala: Temetzontla. Fig. 91 (holotype). Distribution: México: Michoacán;Morelos; Tlaxcala.
Madoniella cymatilis Opitz, 2011b: 207. Type locality: Haiti: Nord: Cap du Nord.Holotype female (INHS). Fig. 92 (holotype). Distribution: Haiti: Nord.
Madoniella dariensis Opitz, 2011b: 169. Holotype female (MIUP). Type locality:Panamá: Darien: Pirre, Estación Rancho Frio. Fig. 93 (paratype). Distribution:Panamá: Darien.
Madoniella darlingtoni Opitz, 2010: 15. Holotype, gender not known (MCZC). Typelocality: Puerto Rico, Maricao Forest. Fig. 94 (holotype). Distribution: Puerto Rico:Maricao Forest
Madoniella disjuga Opitz, 2011b: 182. Holotype female (CMNC). Type locality:México: San Luis Potosi: 40 km W Xilitla. Fig. 95 (holotype). Distribution: México:San Luis Potosi.
Madoniella dislocata (Say) (Enoplium), 1825: 176. Neotype male (MCZC). Typelocality: United States of America: Georgia: Clark County, Whitehall Forest. Fig. 96(nontype specimen). Corporaal 1950a: 251 (Phlogistosternus). Ekis [(now Opitz)1971: 61 (Phlogistosternus)], 1975: 47 (Phlogistosternus), Opitz 1997: 62, 2002: 278;Opitz 2011b: 182. Distribution: Canada: New Brunswick; Nova Scotia; Quebec;Ontario. United States of America: Alabama; Connecticut; Florida; Georgia; Illinois;Indiana; Iowa; Kansas; Kentucky; Lousiana; Maine; Maryland; Massachusetts;Michigan; Mississippi; Missouri; New Hampshire; New Jersey; New York; NorthCarolina; Ohio; Oklahoma; Pennsylvania; Rhode Island; Tennessee; Texas; Virginia;West Virginia.
Enoplium distrophum Klug 1842: 374.Phyllobaenus transversalis Spinola 1844b: 4.
Madoniella displicata Opitz, 2011b: 196. Lectotype female (FSCA). Type locality:Venezuela: Aragua: Ocumare. Fig. 97 (holotype). Distribution: Venezuela: Aragua.
Madoniella divida Opitz, 2010: 15. Holotype male (IAVH). Type locality: Colombia:Bolivar, SFF Los Colorados, Alto de Mirador: Fig. 98 (holotype). Distribution:Colombia: Bolivar.
Madoniella ebena Opitz, 2011b: 208. Lectotype female (CMNH). Type locality:Dominican Republic: Hato Major: Parque Loa Haitises, 3 km W Cueva de arena. Fig.99 (holotype). Distribution: Venezuela: Aragua.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

47
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Madoniella emblema Opitz, 2011b: 195. Type locality: Costa Rica: Heredia: 11 km SELa Virgen. Holotype female (INBC). Fig. 100 (holotype). Distribution: Costa Rica:Heredia.
Madoniella erythrocephala (Gorham) (Phlogistosternus), 1882: 167. Type locality:Panamá: Chiriquí: Volcán de Chiriquí. Lectotype female (INBC). Fig. 101(homotype). Corporaal 1950a: 251 (Phlogistosternus). Opitz 2011b: 170.Distribution: Panamá: Chiriquí.
Madoniella extensiva Opitz, 2011b: 197. Type locality: Puerto Rico: Maricao:MaricaoForest. Holotype female (MCZC). Fig. 102 (holotype). Distribution:Dominican Republic: Pedernalis. Puerto Rico: Maricao.
Madoniella facis Opitz, 2011b: 201. Holotype male (CASC). Type locality: Guyana:Rupununi: Iwokrama Research Forest, 6.4 km N of Kurupukari. Fig. 103 (holotype).Distribution: Guyana: Rupunui.
Madoniella fonteboa Opitz, 2011b: 202. Holotype male (ZMHB). Type locality: Brazil:Amazonas: Fonteboa. Fig. 104 (holotype). Distribution: Brazil: Amazonas.
Madoniella gonia Opitz, 2011b: 188. Holotype female (FSCA). Type locality: México:Durango: 4.8 km E El Salto. Fig. 105 (holotype). Distribution: México: Durango.
Madoniella guana Opitz, 2011b. 158. Holotype female (USNM). Type locality: GuanaIsland. Fig. 106 (holotype). Distribution: British Virgin Island: Guana Island. PuertoRico: Guánica; San Juan.
Madoniella howdenorum Opitz, 2011b: 188. Holotype female (FSCA). Type locality:México: Chiapas: 2.6�6 km S La Trinitaria. Fig. 107 (paratype). Distribution:México: Chiapas; Oaxaca.
Madoniella ignis Opitz, 2011b: 204. Holotype female (FSCA). Type locality:Guatemala: Zacapa: San Lorenzo. Fig. 108 (holotype). Distribution: Guatemala:Zacapa.
Madoniella infula Opitz, 2011b: 222. Holotype, gender not known (CMNH). Typelocality: Dominican Republic: La Vega: Cordillera Central, 4.1 km SW El Convento.Fig. 109 (holotype). Dominican Republic: La Vega. Distribution: DominicanRepublic: La Vega.
Madoniella insignis Opitz, 2011b: 202. Holotype male (ZMHB). Type locality: Brazil:Bahia: Santo Antonio de Barra. Fig. 110 (holotype). Distribution: Brazil: Bahia.
Madoniella knullorum Opitz, 2011b: 230. Holotype male (FMNH). Type locality:United States of America: Texas: Jeff Davis Co., Davis Mountains. Fig. 111(paratype). Distribution: United States of America: Texas.
Madoniella kuehlorum Opitz, 2011b: 172 . Holotype female (FSCA). Type locality:Nicaragua: Matagalpa. Fig. 112 (paratype). Distribution: México: Chiapas.
Belize: Cayo. Honduras: Francisco Morazán; Olancho; Comayagua. Nicaragua:Granada; Matagalpa. Costa Rica: San José.
Madoniella latinopsis Opitz, 2011b: 203. Holotype female (IZAV). Type locality:Venezuela: Aragua: El Limón. Fig. 113 (holotype). Distribution: Venezuela: Aragua.
Madoniella leona Opitz, 2011b: 189. Holotype female (TAMU). Type locality: México:Nuevo Leon: 14.4 km west Iturbide. Fig. 114 (holotype). Distribution: México:Nuevo Leon.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

48
W. OPITZ
Madoniella linea Opitz, 2011b: 208. Holotype female (CNCI). Type locality: Jamaica:Portland: Hardwar Gap. Fig. 115 (paratype). Distribution: Jamaica: Saint Andrew;Saint Thomas
Madoniella lineola Opitz, 2011b: 190. Holotype female (FSCA). Type locality:Guatemala: Zacapa: 2 km S San Lorenzo. Fig. 116 (paratype). Distribution: México:Chiapas. Guatemala: Baja Verapaz; El Progresso, Sacatepéquez; Zacapa. Honduras:Comayagua.
Madoniella lurida Opitz, 2011b: 209. Holotype female (CMNH) Type locality:Dominican Republic: Pedernales, La Abeja, 38 km NNW Cabo Rojo. Fig. 117(holotype). Distribution: Dominican Republic: Pedernales.
Madoniella magdalena Opitz, 2011b: 197. Holotype male (IAVH). Type locality:Colombia: Magdalena: PNN Tayrona Pueblito. Fig. 118 (holotype). Distribution:Colombia: Magdalena.
Madoniella maxicornis Opitz, 2011b: 210. Holotype female (USNM). Type locality:México: Vera Cruz: Fig. 119 (holotype). Distribution: México: San Luis Potosi; VeraCruz. Guatemala: Alta Vera Paz.
Madoniella melina Opitz, 2011b: 172. Holotype female (FSCA). Type locality:Honduras: Yoro: 8 km N La Habana. Fig. 120 (holotype). Distribution: Honduras:Yoro.
Madoniella merga Opitz, 2011b: 213. Holotype female (CNCI). Type locality: México:Durango: 32 km W El Salto. Figure 121 (holotype). Distribution: México: Durango;Sinaloa
Madoniella minor Pic, 1935: 10. Lectotype female (MNHN). Type locality: Guadeloupe.Fig. 122 (nontype specimens). Lepesme, 1947: 169. Corporaal, 1950: 306. Opitz,1997: 62; 2002: 278, 2011b: 218. Distribution: Guadeloupe: Basse-Terre. Montserrat.Dominica: St. Peter.
Madoniella nana Opitz, 2011b: 231. Holotype female (FSCA). Type locality: UnitedStates of America: Texas: Cameron Co., Sabal Palm Grove. Fig. 123 (paratype).Distribution: United States of America: Texas.
Madoniella nebulosa (Chevrolat) (Epiphlaeus), 1876: 28. Lectotype, gender unknown(MNHN). Type locality: Cuba. Fig. 125 (homotype). Corporaal 1950a: 254(Epiphloeus). Peck 2005: 124 (Epiphloeus). Opitz 2011b: 223. Distribution: Cuba.
Madoniella orientalis Zayas, 1988: 60. Syntype series not available (FDZC). Typelocality: Cuba. Fig. 126 (nontype specimen). Peck 2005: 124. Opitz 2011b: 226.Distribution: Cuba: Cienfuego; Granma; Guantánamo; Pinar del Rio.
Madoniella orosiensis Opitz, 2011b: 191. Holotype female (INBC). Type locality: CostaRica: Guanacaste: Estación Maritza, eastern slope of Volcán Orosi. Fig. 127(homotype). Distribution: México: Oaxaca; Chiapas; Comayagua; Quintana Roo;Yucatán. Honduras: Atlantida; El Paraíso; Francisco Morazán; Lampira; Olancho;Santa Barbara. Nicaragua: Granada; Matagalpa. Costa Rica: Guanacaste; Heredia;Puntarenas; San José. Panamá: Chiriquí; Panamá.
Madoniella patula Opitz, 2011b: 192. Holotype female (FSCA). Type locality: México:Chiapas: 54 km S Ocosingo. Fig. 124 (paratype). Distribution: México: Chiapas;
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

49
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Oaxaca; Queretaro; Veracruz; Yacatán; Guatemala: Suchitipéquez. Belize: Belize;Cayo. Honduras: Comayagua; Copán; Cortez; Ocotepeque.
Madoniella pedalis Opitz, 2011b: 227. Holotype male (USNM). Type locality: Cuba: LaHabana, Cayamas. Fig. 128 (holotype). Distribution: Cuba: La Habana.
Madoniella pellis Opitz, 2011b: 162. Holotype male (USNM). Type locality: Cuba:Holguin: Sierra de Nipe, 23 km S Mayari, Pinares de Mayari. Fig. 129 (paratype).Distribution: Cuba: Holguin; La Habana.
Madoniella penninsularis Opitz, 2011b: 155. Holotype female (FSCA). Type locality:México: Oaxaca: Baja California, Cabo San Lucas. Fig. 130 (paratype). Distribution:México: Oaxaca.
Madoniella pici Lepesme. 1947: 169. Lectotype male (MNHN). Type locality:Guadeloupe: Guadeloupe: Trois Rivieres. Fig. 131 (lectotype). Corporaal 1950a: 306.Opitz 2011b: 219. Distribution: Guadeloupe: Guadeloupe.
Madoniella pinicola Opitz, 2011b: 215. Holotype female (FMNH). Type locality:United States of America: Arizona: Cochise Co., Chiricahua Mountains. Fig. 132(paratype). Distribution: United States of America: Arizona. México: México.
Madoniella plenita Opitz, 2011b: 174. Holotype female (INBC). Type locality: CostaRica: Cartago: Turrialba. Fig. 133 (holotype). Distribution: Costa Rica: Cartago.
Madoniella pumilis Opitz, 2011b: 228. Holotype male (IAVH).Type locality: Colombia:Magdalena: Pueblito. Fig. 134 (holotype). Distribution: Colombia: Magdalena.
Madoniella punctata Opitz, 2011b: 216. Holotype female (BMNH). Type locality:Guatemala: El Quiché: Quiché Mountains. Fig. 135 (nontype specimen).Distribution: México: Chiapas. Guatemala: El Quiché.
Madoniella quintana Opitz, 2011b: 194. Holotype female (FSCA). Type locality:México: Quintana Roo, 66 km E Xpujil. Fig. 136 (paratype). Distribution: México:Quintana Roo.
Madoniella rectangularis Opitz, 2011b: 232. Holotype female (FMNH). Type locality:United States of America: Texas: Hidalgo Co. Fig. 137 (paratype). Distribution:United States of America: Texas. México: Coahila; Nuevo Leon; Tamaulipas.
Madoniella redacta Opitz, 2011b: 174. Holotype male (INBC). Type locality: CostaRica: Heredia: Estación Biologica La Selva, 3 km S Pto. Viejo. Fig. 138 (holotype).Distribution: Costa Rica: Cartago.
Madoniella rubidia Opitz, 2011b: 175. Holotype female (MNHN). Type locality: Brazil:Bahia. Fig. 139 (holotype). Distribution: Brazil: Bahia.
Madoniella spilota Opitz, 2010a: 16. Holotype female (IAVH). Type locality: Colombia:Norte de Santander, Los Estoraques, Bosque Piritama. Fig. 140 (holotype).Distribution: Colombia: Norte de Santander.
Madoniella storea Opitz, 2011b: 163. Holotype female (CNCI). Type locality: Jamaica:Portland: Hardwar Gap. Fig. 141 (paratype). Distribution: Jamaica: Portland; SaintAndrew; Saint Thomas; Trelawney; Saint Catherine.
Madoniella tegetis Opitz, 2011b: 228. Holotype male (FSCA). Type locality: Honduras:Copán: 6 km NW San Agustin. Fig. 142 (holotype). Distribution: Honduras: Copán.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

50
W. OPITZ
Madoniella texis Opitz, 2011b: 175. Holotype female (INBC). Type locality: Costa Rica:Cartago: Turrialba. Fig. 143 (paraype). Distribution: Costa Rica: Alajuela; Cartago;Coclé; Guanacaste; Heredia; Puntarenas. Panamá: Chiriquí; Panamá.
Madoniella thomasi Opitz, 2011b: 198. Holotype female (FSCA). Type locality:Bahamas: Andros Island, Maidenhair Coppica. Fig. 144 (holotype). Distribution:Bahamas: Andros Island. In Opitz (2011b: 248: map 18) thomasi was erroneouslylisted as thomsoni. Also, in that work, Figs 112 and 235 involve this species and noteximia..
Madoniella vogti Opitz, 2011b: 234. Holotype female (FMNH). Type locality: UnitedStates of America: Texas: South-east Hidalgo Co. Fig. 145 (holotype). Distribution:United States of America: Texas. México: San Luis Potosi; Tamaulipas.
Madoniella welderi Opitz, 2011b: 235. Holotype female (FMNH). Type locality: UnitedStates of America: Texas: San Patricio Co., Welder Wildlife Refuge. Fig. 146(holotype). Distribution: United States of America: Texas.
Madoniella zonula Opitz, 2011b: 177. Holotype female (MIUP). Type locality: Panamá:Colón. Fig. 147 (holotype). Distribution: Panamá: Colón.
Genus Megaphloeus Opitz, 2011a: 71EKIS (now OPITZ) 1975: 47 (Epiphloeus). MAWDSLEY 1992: 129 (Epiphloeus). OPITZ 1997: 53 (Epiphloeus);
2011a: 71. Type species: Epiphloeus setulosus Thomson, 1860: 60 (by original designation).
Most recent generic revision: OPITZ 2011a.Distribution: México to Argentina.Number of species: 26.Megaphloeus absentis Opitz, 2011a: 91. Holotype male (CMNC). Type locality:
México: Puebla: 24 km E Teziutlan. Fig. 148 (paratype). Distribution: México:Puebla: Veracruz.
Megaphloeus animosus (Wolcott), 1927: 92 (Epiphloeus). Holotype female (FMNH).Type locality: Brazil: Costa Rica: Cartago. Fig. 149 (nontype specimen). Corporaal1950a: 253. (Epiphloeus). Opitz 2011a: 106. Distribution: México: Veracruz.Honduras: Atlántida. Nicaragua: Zelaya. Costa Rica: Cartago; Guanacaste; Heredia;Limón; Puntarenas. Panamá: Bocas del Toro; Chiriquí; Colón; Darien; Veraguas.Colombia: Amazonas; Chocó; Caquetá. Venezuela: Tachira. Ecuador: Napo. Peru:Junín. Brazil: Rondônia.
Megaphloeus bulatus Opitz, 2011a: 80. Holotype male (INBC). Type locality: CostaRica: Puntarenas, Estación Agujas. Fig. 150 (paratype). Distribution: Nicaragua:Chontales. Costa Rica: Alajuela. Heredia; Limón; Puntarenas. Panamá: Colón;Panamá.
Megaphloeus cartus Opitz, 2011a: 91. Holotype male (FSCA). Type locality: Belize:Orange Walk, Rio Bravo, La Milpa. Fig. 151 (holotype). Distribution: Belize: OrangeWalk.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

51
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Megaphloeus circinus Opitz, 2011a: 109. Holotype male (IAVH). Type locality:Colombia: Vaupés, R. N. Mosiro-Itajura (caparú), Centro Abiental. Fig. 152(paratype). Distribution: Colombia: Amazonas; Vaupés.
Megaphloeus fucoaquilus Opitz, 2011a: 82. Holotype male (FSCA). Type locality:Brazil: Rondônia: 62 km SE Ariquemes. Fig. 153 (holotype). Distribution: Brazil:Rondônia.
Megaphloeus lividipes (Chevrolat) (Epiphlaeus), 1874: 320. Lectotype female(MNHN). Type locality: Venezuela. Fig. 154 (homotype). Corporaal 1950: 254(Epiphloeus). Opitz 2011a: 110. Distribution: Venezuela: Aragua, Carabobo.
Megaphloeus longius Opitz, 2011a: 92. Holotype male (CMNC). Type locality:Honduras: Olancho: Parque Nacional La Muralla. Fig. 155 (paratype). Distribution:Honduras: Cortés; Olancho.
Megaphloeus marginipes (Chevrolat) (Epiphlaeus), 1874: 320. Lectotype male(MNHN).Type locality: México: Veracruz. Fig. 156 (nontype specimen). Corporaal1950a: 254 (Epiphloeus). Opitz 2011a: 93. Distribution: México: Veracruz. Belize:Cayo; Orange Walk; Toledo. Guatemala: Izabal. Honduras: Atlántida; Olancho. CostaRica: Alajuela; Guanacaste; Heredia; Puntarena. Panáma: Chiriquí; Colón; Panamá.
Megaphloeus megasensibilis Opitz, 2011a: 82. Holotype male (FSCA). Type locality:Brazil: Rondônia: 62 km SE Ariquemes. Fig. 157 (paratype). Distribution: Brazil:Mato Grosso; Rondônia.
Megaphloeus mucoreus (Klug) (Enoplium), 1842: 371. Holotype male (FSCA). Typelocality: Brazil. Fig. 158 (homotype). Ekis (now Opitz) 1975: 48 (Epiphloeus), Opitz1997: 68 (Epiphloeus). Opitz 2011a: 84. Distribution: Corporaal 1950a: 254(Epiphloeus). Colombia: Amazonas; Magdalena. Venezuela: Bolívar. Guyana: Grossodo Sul; Potaro. Ecuador: Napo; Santiago Morona. Peru: Huanuco; Madre de Dios;San Martin. Brazil: Acre; Alagôas; Amazonas; Bahia; Seará; Espírito Santo; Goiás;Guanabara; São Paulo; Pará; Mato Grosso; Paraná; Rio Grande do Sul; Rondônia;Santa Catarina. Paraguay: Itapua; San Pedro. Argentina: Entre Ríos; Misiones.
Megaphloeus nubilus (Klug) (Enoplium), 1842: 370. Type locality: Brazil. Holotypemale (FSCA). Fig. 159 (homotype). Corporaal 1950a: 254 (Epiphloeus). Opitz 2011a:94. Distribution: French Guiana: Cayenne. Brazil: Paraná; Espítu Santo.
Megaphloeus parvulus (Schenkling) (Epiphloeus), 1900: 398. Lectotype male (DEIG).Type locality: Brazil: Goiás. Fig. 160 (holotype). Corporaal 1950a: 254 (Epiphloeus).Opitz, 2011a: 95. Distribution: Brazil: Bahia; Goiás; Mato Grosso; Mato Grosso doSul; Paraná; Rio de Janeiro; Rondônia; Santa Catarina; São Paulo.
Megaphloeus pectilus Opitz, 2011a: 96. Holotype male (SEMC). Type locality: Guyana:Demerara: 1 km W Kurupukari, Iwokrama Field Station. Fig. 161 (paratype).Distribution: Guyana: Demerara. Brazil: Pará.
Megaphloeus platyglenus Opitz, 2011a: 77. Holotype female (USNM). Type locality:Ecuador Orellana: Ethnica, Waorani, 1 km S Onko, Gare camp. Fig. 162 (holotype).Distribution: Ecuador: Orellana.
Megaphloeus rictocaliginus Opitz, 2011a: 112. Holotype female (FSCA). Type locality:Brazil: Rondônia, 62 km SE Ariquemes. Fig. 163 (paratype). Distribution: Costa
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

52
W. OPITZ
Rica: Cartago; Guanacaste. Panamá: Colón. Venezuela: Bolivar; Barinas. Guyana:Essequibo. French Guiana. Bolivia: Chaparé. Brazil: Pará; Rondônia.
Megaphloeus setulosus (Thomson) (Epiphlaeus), 1860: 60. Lectotype female (MNHN).Type locality: México. Fig. 164 (lectotype). Corporaal 1950a: 254 (Epiphloeus).Opitz 1997: 66 (Epiphloeus); 2010a: 16, 2011a: 97. Distribution: México: Chiapas;Guerrero; Jalisco; Nayarit; Oaxaca; Quintana Roo; Sinaloa; Tabasco; Veracruz;Yucatán. Guatemala: Alta Verapaz; Baja Verapaz; Quetzaltenango. Belize: Belize.Honduras: Atlantida; El Paraiso; Olancho. El Salvador: Morazán. Nicaragua:Chontales. Costa Rica: Alajuela; Cartago; Guanacaste; Heredia; Limón; San José;Puntarenas. Panamá: Chiriquí; Colón; Darién; Panamá; Colombia: Cundinamarca;Magdalena; Putumayo. Ecuador: Napo; Tachira. Bolivia: Cavinas; Santa Cruz. Peru:Cuzco. Brazil: Mato Grosso; Rondônia.
Megaphloeus sexplagiatus (Kuwert) (Epiphlöus), 1893: 494. Holotype, gender notknown (MNHN).Type locality: Peru: Amazones. Fig. 165 (homotype). Corporaal1950a: 254 (Epiphloeus). Opitz 2011a: 100. Distribution: Belize: Stann Creek.Panamá: Colon; Darien. Colombia: Magdalena. Venezuela: Bolivar; Tachira. Guyana:Demerara. Ecuador: Napo. Peru: Huánuco; San Martín; Madre de Dios. Brazil:Amazonas; Pará; Rondônia; Tocatins.
Megaphloeus terzonatus (Gorham) (Epiphloeus), 1877a: 242. Lectotype male(BMNH). Type locality: Brazil: Amazon: Tefé. Fig. 166 (homotype). Corporaal1950a: 254 (Epiphloeus). Opitz 2011a: 88. Distribution: Ecuador: Esmeraldas;Pichincha. Peru: Junín; Madre de Dios. Bolivia: Cochabamba. Brazil: Amazonas;Chapare; Mato Grosso; Pará; Rondônia.
Megaphloeus tigrinus Opitz, 2011a: 102. Holtotype male (IAVH). Type locality:Colombia: Putumayo, PNN La Paya, Finca Charapa. Fig. 167 (paratype).Distribution: Colombia: Putumayo; Caquetá; Magdalena.
Megaphloeus tricolor (Kuwert) (Epiphlöus), 1893: 493. Lectotype, gender not known(MNHN). Type locality: Brazil: Amazonas. Fig. 168 (lectotype). Corporaal 1950a:255 (Epiphloeus). Opitz 2011a: 113. Distribution: Costa Rica: Cartago; Heredia;Puntarenas. Panamá: Colón; Darien; Panamá. Colombia: Amazonas; Caquetá;Putumayo; Vaupés. Venezuela: Aragua. Ecuador: Zemora-Chinchipe, Napo. Bolivia:Beni; Cochabamba; La Paz; Santa Cruz. Brazil: Amazonas; Mato Grosso; Rondônia.
Megaphloeus ustafinis Opitz, 2011a: 104. Holotype male (FSCA). Type locality: Brazil:Mato Grosso. Fig. 169 (paratype). Distribution: Brazil: Amazonas; Mato Grosso;Rondônia.
Megaphloeus ustus Opitz, 2011a: 79. Holotype male (FSCA). Type locality: Brazil:Santa Catarina: Nova Teutonia. Fig. 170 (paratype). Distribution: Brazil: SantaCatarina.
Megaphloeus variegatus (Klug) (Enoplium), 1842: 371. Lectotype male (ZMHB). Typelocality: Brazil: Pará. Fig. 171 (homotype). Corporaal 1950a: 255 (Epiphloeus). Ekis(now Opitz) 1975: 48, 49 (Epiphloeus). Opitz 2011a: 89. Distribution: Colombia:Amazonas; Magdalena; Meta; Nariño; Putumayo. Venezuela: Delta Amacura.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

53
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Guyana: Demerara. Ecuador: Napo. Peru: Huanuco; Lareto; San Martín; Santipo.Brazil: Amazonas; Mato Grosso; Pará; Paranhâo.
Megaphloeus velutinus (Gorham) (Epiphloeus), 1877a: 247. Type locality: Brazil:Amazonas. Lectotype, male (BMNH). Fig. 172 (homotype). Corporaal 1950a: 255(Epiphloeus). Opitz 2011a: 118. Distribution: Panamá: Colón; Darien; Panamá.Colombia, Amazonas; Caquetá; Putumayo. Suriname: Saramacca. Peru: Lareto.Bolivia: Chaparé. Brazil: Amazonas; Mato Grosso; Pará; Rondônia.
Megaphloeus vitellinus Opitz, 2011a: 78. Type locality: Brazil: Bahia: Encruzilhada.Holotype male (FSCA). Fig. 173 (holotype). Distribution: Brazil: Bahia.
Genus Megatrachys Opitz, 1997: 61.OPITZ 2008c: 21.Type species: Megatrachys paniculus Opitz, 1997: 61 (by original designation).
Most recent generic revision: OPITZ 2008c: 21.Distribution: México to Guatemala.Number of species: 3.Megatrachys bibara Opitz, 2008c: 21. Holotype female (FSCA). Type locality:
Guatemala: Zacapa: 2 km San Lorenzo. Fig. 174 (holotype). Distribution: Guatemala:Zacapa.
Megatrachys paniculus Opitz, 1997: 61. Holotype male. (CNCI). Type locality: México:Chiapas: 8 km W San Cristóbal (= San Cristobal de Las Casas). Fig. 175 (holotype).Opitz 2008c: 23. Distribution: México: Chiapas.
Megatrachys truncatia Opitz, 2008c: 23. Holotype female (FSCA). Type locality:México: Chiapas: 47.5 km NW Comitán. Fig. 176 (holotype). Distribution: México:Chiapas.
Genus Opitzia Nemésio, 2006: 77.OPITZ 2008c: 24.Type species: Arenaria chiapas Opitz, 1997: 57 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008c. Distribution: México and Bolivia.Number of species: 2.Opitzia apicula Opitz, 2008c: 25. Holotype female (UASC). Type locality: Bolivia:
Santa Cruz: Amboro road, above Achira Campo. Fig. 177 (holotype). Distribution:Bolivia: Santa Cruz.
Opitzia chiapas (Opitz) (Arenaria), 1997: 57. Type locality: México: Chiapas: 39 kmNW Comitán. Holotype male (MCMC). Fig. 178 (holotype). Nemésio 2006: 77(Opitzia). Opitz 2008c: 25 (Opitzia). Distribution: México: Chiapas. Guatemala: ElProgresso: Zacapa.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

54
W. OPITZ
Genus Opitzius Barr, 2006: 215.OPITZ 2008b: 29.Type species: Opitzius thoracicus Barr, 2006: 215 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008b: 29. Distribution: Brazil.Number of species: 1.Opitzius thoracicus Barr, 2006: 215. Holotype female (MZSP).Type locality: Brazil:
Bahia: Encruzilhada. Fig. 179 (holotype). Opitz 2008b: 29. Distribution: Brazil:Bahia; Espírito Santo; Guanabara; Paraná; Rio de Janeiro.
Genus Parvochaetus Opitz, 2006: 109.Type species: Parvochaetus fucolatus Opitz, 2006: 113 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2006: 109. Distribution: Panamá to Brazil.Number of species: 5.Parvochaetus albicornis Opitz, 2006: 112. Holotype male (MCNZ). Type locality:
Brazil: Mato Grosso: Sinop, Coordenadas. Fig. 180 (holotype). Distribution: Brazil:Mato Grosso.
Parvochaetus froeschneri Opitz, 2006: 112. Holotype female (DZUP). Type locality:Brazil: Rio de Janeiro: Rio Grande (= Rio Grandina). Fig. 181 (holotype).Distribution: Brazil: Mato Grosso; Rio de Janeiro.
Parvochaetus fucolatus Opitz, 2006: 113. Holotype male (IZAV). Type locality: Brazil:Rio de Janeiro: Rio Grande (= Riograndina). Fig. 182 (holotype). Distribution: Brazil:Rio de Janeiro
Parvochaetus linearis (Gorham) (Apolopha), 1883: 182. Type locality: Panamá:Bugaba. Holotype male (BMNH). Fig. 183 (holotype). Corporaal 1950a: 252(Phlogistosternus). Opitz 2006: 116. Distribution: Panamá: Bugaba.
Parvochaetus sandaracus Opitz, 2006: 117. Holotype male (AMNH).Type locality:Brazil: Nova Teutonia: Santa Catarina. Fig. 184 (paratype). Distribution: Brazil: NovaTeutonia; Paraná.
Genus Pennasolis Opitz, 2008c: 27.Type species: Phyllobaenus merkeli Horn, 1896: 374 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008c: 27.Distribution: USA and México.Number of species: 3.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

55
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Pennasolis californica (Van Dyke), 1923: 47 (Phyllobaenus). Holotype female (CASC).Type locality: Arizona. Corporaal 1950a: 251 (Phlogistosternus). Fig. 185 (nontypespecimen). Opitz 2008c: 29 (Pennasolis). Rifkind, Toledo, & Corona 2010: 55.Distribution: USA: Arizona: California.
Pennasolis merkeli (Horn), 1896: 374 (Phyllobaenus). Lectotype male (CASC). Typelocality: USA: California: El Dorado County; Nevada County. Yosemite Valley. Fig.186 (nontype specimen). Corporaal 1950a: 252 (Phlogistosternus). Opitz, 2008c: 29(Pennasolis). Rifkind, Toledo, & Corona 2010: 55. Distribution: USA: Arizona;California; Colorado; New Mexico; Texas; Utah.
Pennasolis opitzi Rifkind, Toledo, & Corona, 2010: 54 Holotype male (CNIN). Typelocality: México, Morelos, Tepalcingo, S El Limón. Fig. 187 (paratype).
Genus Pericales Opitz, 2008c: 32.Type species: Pericales albogilvus Opitz, 2008c: 32 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008c.Distribution: Haiti and the Dominican Republic.Number of species: 1.Pericales albogilvus Opitz, 2008c: 32. Holotype male (FSCA). Type locality: Haiti: Sud-
Oueste (=Sud-Est), Massif de La Selle, Morne d� Enfer. Fig. 188 (holotype).Distribution: Haiti: Sud-Est. Dominican Republic: Pedernales
Genus Plocamocera Spinola, 1844b: 17.OPITZ W. 1997: 55; 2004: 1. Type species: Plocamocera sericella Spinola, 1844b: 19 (by monotypy).
Generic synonyms: None.Most recent generic revision: OPITZ 2004: 1. Corrigendum: There are corrections in the abovementioned publication: Page 5, thelegends should read as follows: Figs 139�169. Antenna or antennal club. 139.Plocamocera salasis. 140. P. manausensis. 141. P. minima. 142. P. prolixa. 143. P. lucis.144. P. castanea. 145. P. pupula. 146. P. quadrula. 147. P. baria. 148. P. bolivari. 149.P. iota. 150. P. aliguantula. 151. P. sesquipedalis. 152. P. confrata. 153. P. aspera. 154.P. bispina. 155. P. ambra. 156. P. sericella. 157. P. cericellopsis. 158. P. similis. 159. P.procera. 160. P. santa. 161. P. aura. 162. P. taruma. 163. P. coactilis. 164. P. paris. 165.P. onorei. 166. P. auratilis. 167. P. insula. 168. P. carnegei. 169. P. selva: Page 60, indescription of Plocamocera ambra, Head: antennal club as in figure 155.Distribution: México to Brazil.Number of species: 37.Plocamocera aliguantula Opitz, 2004: 38. Holotype male (USNM). Type locality: Costa
Rica: Limón: Revantazón, Hamburg Farm. Fig. 189 (paratype). Distribution: CostaRica: Limón.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

56
W. OPITZ
Plocamocera ambra Opitz, 2004: 60. Holotype male (INBC). Type locality: Costa Rica:Puntarenas: Estación Agujas. Fig. 190 (paratype). Distribution: Costa Rica: Alajuela;Puntarenas.
Plocamocera argentea Opitz, 2004: 55. Holotype male (CMNH). Type locality: Bolivia:Santa Cruz: Provincia del Sara. Fig. 191 (paratype). Distribution: Bolivia: Santa Cruz.
Plocamocera aspera Opitz, 2004: 52. Holotype female (MZSP). Type locality: Brazil:Amazonas: Paraque, 30 km E of Manaus. Fig 192 (holotype). Distribution: Brazil:Amazonas.
Plocamocera aura Opitz, 2004: 46. Holotype female (USNM). Type locality: Bolivia:La Paz: Tumupasa. Fig. 193 (holotype). Distribution: Bolivia: La Paz.
Plocamocera auratilis Opitz, 2004: 55. Holotype male (AMNH). Type locality: Panamá:Panamá: Pan American Highway, 30 km E of Canita. Fig. 194 (paratype).Distribution: Costa Rica: Alajuela. Panamá: Panamá.
Plocamocera baria Opitz, 2004: 46. Holotype female (IZAV). Type locality: Venezuela:Amazonas: Rio Negro, Rio Baria. Fig. 195 (holotype). Distribution: Venezuela:Amazonas.
Plocamocera bispina Opitz, 2004: 53. Holotype female (FMNH). Type locality: Brazil:Mato Grosso: Rio Caraguata. Fig. 196 (holotype). Distribution: Brazil: Mato Grosso.
Plocamocera bolivari Opitz, 2004: 59. Holotype male (IZAV). Type locality: Venezuela:Trujillo: Betijoque. Fig. 197 (holotype). Distribution: Venezuela: Trujillo.
Plocamocera buenavista Opitz, 2004: 46. Holotype male (FSCA). Type locality:Bolivia: Santa Cruz: 3.7 km SSE Buenavista, Hotel Flora & Fauna. Fig. 198(holotype). Distribution: Bolivia: Santa Cruz.
Plocamocera carnegei Opitz, 2004: 54. Type locality: Brazil: Guanabara: Rio deJaneiro. Holotype female (CMNH). Fig. 199 (holotype). Distribution: Brazil:Guanabara.
Plocamocera castanea Opitz, 2004: 42. Holotype female (MZSP). Type locality: Brazil:Goías: Jatai Goías. Fig. 200 (paratype). Distribution: French Guiana: Cayenne;Guyane; Saint-Georges. Ecuador: Napo. Bolivia: Santa Cruz; Cochabamba. Brazil:Goías.
Plocamocera clinata Opitz, 2010: 17. Holotype female (FSCA). Type locality: Bolivia:Santa Cruz: 3.7 km SSE Buena Vista, Hotel Flora & Fauna. Fig. 201 (holotype).Distribution: Bolivia: Santa Cruz
Plocamocera coactilis Opitz, 2004: 51. Holotype male (MZSP). Type locality: Brazil:Mato Grosso: Mato Grosso. Fig. 202 (paratype). Distribution: Brazil: Amazonas;Goias; Mato Grosso; Pará.
Plocamocera confrater Kuwert, 1893: 496. Lectotype female (MNHN).Type locality:Peru: Amazon. Fig. 203 (nontype specimen). Corporaal 1950a: 255. Opitz 2004: 48.Distribution: Colombia: Caquetá; Meta; Putumayo. Guyana: Bartica. French Guiana:Cayenne. Ecuador: Napo. Peru: Madre de Dios. Bolivia: Beni. Brazil: Amazonas;Goiás; Mato Grosso; Pará; Rondônia.
Plocamocera insula Opitz, 2004: 60. Holotype male (FSCA). Type locality: Trinidad:Saint George Parsh: Arima. Fig. 204 (holotype). Distribution: Trinidad.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

57
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Plocamocera iota Opitz, 2004: 38. Holotype female (FMNH). Type locality: Brazil:Mato Grosso: Rio Caraguata. Fig. 205 (holotype). Distribution: Brazil: Mato Grosso.
Plocamocera jayhawkalis Opitz, 2004: 45. Holotype, gender not known (SEMC). Typelocality: Suriname: Saramaca: 175 km WSW Zanderij Airport. Fig. 206 (holotype).Distribution: Suriname: Saramaca.
Plocamocera lena Opitz, 2010: 18. Holotype male (USNM). Type locality: Ecuador:Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 207(holotype). Distribution: Ecuador: Orellana.
Plocamocera lucis Opitz, 2004: 40. Holotype male (MZSP). Type locality: Brazil:Coordenadas: Mato Grosso: Sinop. Fig. 208 (paratype). Distribution: Brazil: MatoGrosso.
Plocamocera manausensis Opitz, 2004: 33. Holotype male (MZSP). Type locality:Brazil; Amazonas: Manaus: 1 km W Taruma Falls. Fig. 209 (paratype). Distribution:Ecuador: Orellana. Brazil: Amazonas.
Plocamocera minima Opitz, 2004: 38. Holotype male (AMNH). Type locality: Panamá:Panamá: Barro Colorado Island. Fig. 210 (paratype). Distribution: Panamá: BarroColorado Island; Canal Zone.
Plocamocera onorei Opitz, 2004: 53. Holotype female (FSCA). Fig. 211 (holotype).Type locality: Ecuador: Napo: Yasuni Research Station. Distribution: Ecuador: Napo.
Plocamocera paris Opitz, 2004: 52. Holotype female (CNCI). Type locality: Brazil:Distrito Federal: Parque Nacional. Fig. 212 (holotype). Distribution: Brazil: DistritoFederal.
Plocamocera procera Opitz, 2004: 51. Holotype male (LACM). Type locality:Colombia: Amazonas: P.N.N. Amacayacu. Fig. 213 (holotype). Distribution:Colombia: Amazonas.
Plocamocera prolixa Opitz, 2004: 39. Holotype male (INBC). Type locality: Alajuela:20 km S Upsula. Fig. 214 (holotype). Distribution: Costa Rica: Alajuela.
Plocamocera pupula Opitz, 2004: 42. Holotype male (MZSP). Type locality: Brazil:Mato Grosso: Sinop. Fig. 215 (paratype). Distribution: Brazil: Mato Grosso.
Plocamocera quadrula Opitz, 2004: 44. Holotype male (AMNH). Type locality: Brazil:Mato Grosso: Sinop. Fig. 216 (holotype). Distribution: Brazil: Mato Grosso.
Plocamocera salasis Opitz, 2004: 33. Holotype male (MZSP). Type locality: Brazil;Amazonas: Manaus: 1 km W Taruma Falls. Fig. 217 (holotype). Distribution:Brazil: Amazonas.
Plocamocera santa Opitz, 2004: 54. Holotype male (FSCA). Type locality: Brazil: SantaCatarina: Nova Teutonia. Fig. 218 (holotype). Distribution: Brazil: Santa Catharina.
Plocamocera selva Opitz, 2004: 61. Holotype male (FSCA). Type locality: Bolivia:Santa Cruz: 5 km ESE Warnes, Hotel Rio Selva. Fig. 219 (holotype). Distribution:Bolivia: Santa Cruz.
Plocamocera sericella Spinola, 1844b: 19. Neotype male (AMNH). Type locality:Panamá: Panamá: Madden Forest. Fig. 220 (nontype specimen). Corporaal 1950a:255. Opitz 2004: 57. Distribution: México: Campeche; Chiapas; Nayarit; Oaxaca;Quintana Roo; Tamaulipas; Veracruz. Belize: Belize. Honduras: Comayagua. Costa
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

58
W. OPITZ
Rica: Alajuela; Guanacaste; Limón. Panamá: Barro Colorado Island; Canal Zone;Cólon. Colombia: Cundilamarca; Magdalena; Putumayo. Trinidad. Venezuela:Aragua; Bolivar. French Guiana: Nouvea Chantier. Guyana: Kourou PisteSoumourou. Ecuador: Napo; Pichincha. Peru: Loretto; Sam Martin. Bolivia:Cochabamba; Santa Cruz. Brazil: Amazonas; Goiás; Mato Grosso; Rondônia.
Plocamocera sericellopsis Opitz, 2004: 59. Holotype male (MZSP). Type locality:Brazil: Rondônia: 62 km SE Ariquenes. Fig. 221 (holotype). Distribution: Brazil:Rondônia.
Plocamocera sesquipedalis Opitz, 2004: 40. Holotype male (BMNH). Type locality:Guyana: Demerara: Ikuribisi. Fig. 222 (paratype). Distribution: Guyana: Bartica;Demerara. Peru: Loreto. Brazil: Rondônia.
Plocamocera similis Opitz. 2004: 41. Holotype male (QCAZ). Type locality: Ecuador:Pichincha: 18 km S Tinalandia. Fig. 223 (holotype). Distribution: Ecuador: Pichincha.
Plocamocera specula Klug, 1842: 372. Holotype male (ZMHB). Type locality: Brazil.Fig. 224 (holotype). Corporaal 1950a: 254. Opitz 2004: 45. Distribution: Brazil.
Plocamocera taruma Opitz, 2004: 47. Holotype male (MZSP). Type locality: Brazil:Amazonas: Manaus: 1 km W Taruma Falls. Fig. 225 (holotype). Distribution: Brazil:Amazonas.
Genus Pteroferus Opitz, 2008c: 33.Type species: Pteroferus zolnerowichi Opitz, 2008c: 33 (by original designation).
Most recent generic revision: OPITZ 2008cDistribution: Brazil.Number of species: 1.Pteroferus zolnerowichi Opitz, 2008c: 33. Type locality: Brazil: Santa Catarina: Nova
Holotype female (FSCA). Fig. 226 (holotype). Distribution: Brazil: Santa Catarina.
Genus Pyticeroides Kuwert, 1894: 7, 9.OPITZ 1997: 59; 2002: 278; 2007: 87. Type species: Pyticeroides arrogans Kuwert, 1894: 7 (by monotypy).
Generic synonyms: Neichnea Wolcott & Chapin 1918: 103.Most recent generic revision: OPITZ 2007. Corrigendum: There are corrections in the abovementioned publication: Page 103.Couplet 19(18) should read... Pronotum moderately transverse, width more than 1.3xlength (Fig. 62). Couplet 19� should read... Pronotum very transverse, width 1.6x length(Fig. 65). Couplet 21(20�) should end in P. vichadium sp. nov.: Page 104. Couplet 26(25)should end in P. arrogans Kuwert. Couplet 31(30�) should end in P. trilineatum(Chevrolat). Page 106. I the legend of figure 73 is P. rifkindi and figure 83 is P. hansoni.Page 136. Other material examined for P. trilineatum México should be followed byJalisco.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

59
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Distribution: Canada to Argentina.Number of species: 39.Pyticeroides acaris Opitz, 2007: 113. Holotype female (IAVH). Type locality: Colombia:
Magdalena: PNN Tyrona Palangana. Fig. 227 (holotype). Distribution: Colombia:Magdalena.
Pyticeroides arrogans Kuwert, 1894: 7. Lectotype female (MNHN). Type locality: Peru:Amazonas. Fig. 228 (holotype). Corporaal 1950a: 253. Opitz 2007:130. Distribution:Peru: Amazonas.
Pyticeroides caecoatus Opitz. 2007: 135. Holotype male (AMNH). Type locality:México: Chiapas: 2.6�6 km S La Trinitaria. Fig. 229 (holotype). Distribution:México: Chiapas.
Pyticeroides caligneus Opitz, 2007: 115. Holotype male (FMNH). Type locality: Brazil:Paraná: Rondon. Fig. 230 (paratype). Distribution: Brazil: Paraná.
Pyticeroides chiriquianum (Gorham) (Apolopha), 1883: 182. Lectotype male (BMNH).Type locality: Panamá: Chiriquí: Volcan de Chiriquí. Fig. 231 (homotype). Corporaal1950a: 252. Opitz 2007: 122. Distribution: Costa Rica: Guanacate; Puntarenas.Panamá: Chiriquí; Colón: Panamá. Colombia: Bolivar; Magdalena. Ecuador: Guayas.
Pyticeroides cinericius Opitz, 2007: 119. Holotype male (FMNH). Type locality: Brazil:Mato Grosso: Rio Caraguata. Fig. 232 (paratype). Distribution: Brazil: Mato Grosso.
Pyticeroides decurialis Opitz, 2007: 111. Holotype male (MCNZ). Type locality: Brazil:Espírito Santo, Linhares. Fig. 233 (holotype). Distribution: Brazil: Espírito Santo.
Pyticeroides enormis Opitz, 2007: 105. Type locality: Bolivia: Cochabamba. Holotypefemale (MNHN). Fig. 234 (holotype). Distribution: Bolivia: Cochabamba.
Pyticeroides eurides Opitz, 2007: 116. Holotype male (MCNZ). Type locality: Brazil:Mato Grosso: Diamantino, Fazenda Rio Arinos. Fig. 235 (holotype). Distribution:Brazil: Mato Grosso.
Pyticeroides fustis Opitz, 2007: 125. Holotype male (MCNZ). Type locality: Brazil:Mato Grosso: Sinop, Coordenadas. Fig. 236 (holotype). Distribution: Brazil: MatoGrosso; Espíritu Santo.
Pyticeroides hansoni Opitz, 2007:120. Holotype male (FSCA). Type locality: Ecuador:Napo: Yasuni Research Station. Fig. 237 (holotype). Distribution: Ecuador: Napo.
Pyticeroides ichnopsis Opitz, 2007: 129. Holotype female (MNHN). Type locality:Brazil: Bahia: Salobro. Fig. 238 (holotype). Distribution: Brazil: Bahia.
Pyticeroides inconscriptus Opitz, 2007: 134. Holotype male (AMNH). Type locality:México: Chiapas: Parque Nacional Montebello. Fig. 239 (holotype). Distribution:México: Chiapas.
Pyticeroides inexilis Opitz, 2007: 107. Holotype male (MZSP). Type locality: Brazil:São Paulo: Fazenda de Alho Itu. Fig. 240 (holotype). Distribution: Brazil: São Paulo:Paraná.
Pyticeroides iscus Opitz, 2007: 108. Holotype male (MCNZ). Type locality: Brazil:Pará: Tucurui. Fig. 241 (holotype). Distribution: Brazil: Pará.
Pyticeroides jubaris Opitz, 2007: 109. Holotype male (FMNH). Type locality: Brazil:São Paulo: Rio Caraguata. Fig. 242 (holotype). Distribution: Brazil: São Paulo.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

60
W. OPITZ
Pyticeroides laticornis (Say) (Enoplium), 1835: 164. Lectotype, gender not known(MCZC). Type locality: USA, North Carolina. Fig. 243 (nontype). Corporaal 1950a:252 (Neichnea). Opitz 1997: 59; 2002: 278; 2007: 87. Fig. 145 (holotype).Distribution: Canada: Ontario. USA: Alabama; Arkansas; Colorado; Connecticut;District of Columbia; Florida; Georgia; Illinois; Indiana; Kansas; Kentucky;Maryland; Massachusetts; Michigan; Missouri; Mississippi; New Jersey; New York;North Carolina; Oklahoma; Ohio; Pennsylvania; Tennessee; Texas; Virginia; WestVirginia. México: San Luis Potosi.
Pyticeroides latisentis Opitz, 2010: 19. Holotype female (USNM). Type locality:Ecuador: Orellana, Tiputini Biodiversity Station. Fig. 244 (holotype). Distribution:Ecuador: Orellana.
Pyticeroides luteolus Opitz, 2007: 121. Holotype male (FMNH). Type locality: Brazil:Santa Catarina: Nova Teutonia. Fig. 245 (paratype). Distribution: Brazil: SantaCatarina.
Pyticeroides maesi Opitz, 2007: 124. Holotype male (SEAN). Type locality: Nicaragua:Granada: Volcán Mombacho, Finca San Joaquin. Fig. 246 (paratype). Distribution:Nicaragua: Granada. Costa Rica: Guanacaste.
Pyticeroides manni Chapin, 1927: 8. Holotype, gender not known (USNM). Typelocality: Bolivia: Beni: Cavinas, Rio Beni. Fig. 247 (nontype specimen). Corporaal1950a: 253. Opitz 2007: 112. Distribution: Bolivia: Beni: Cochabamba. Brazil:Rondônia.
Pyticeroides moraguesi Opitz, 2010: 20. Holotype female (FSCA). Type locality: FrenchGuiana: Guyane: Kourou Piste Soumourou. Fig. 248 (holotype). Primary type:Distribution: French Guiana: Guyane.
Pyticeroides notialis Opitz, 2007: 126. Holotype male (MLPA). Type locality:Argentina: Tucumán: Las Tipas. Fig. 249 (paratype). Distribution: Argentina: Salta;Tucumán; Santa del Estero; Misiones.
Pyticeroides parvoporis Opitz, 2010: 21. Holotype male (USNM). Type locality:Ecuador: Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 250(holotype). Distribution: Ecuador: Orellana.
Pyticeroides petilus Opitz, 2007: 108. Holotype female (MNHN). Type locality: Bolivia:Cochabamba. Fig. 251 (holotype). Distribution: Boilivia: Cochabamba.
Pyticeroides pinnacerinis Opitz, 2010: 22: Holotype male (USNM). Type locality:Ecuador: Orellana: Reserva Ethnica Waorani, 1 km S of Onkone Gare Camp. Fig. 252(holotype). Distribution: Ecuador: Orellana.
Pyticeroides plautus Opitz, 2007: 135. Holotype male (CNCI). Type locality: México:Oaxaca: highway 131, 75 km S Oaxaca. Fig. 253 (paratype). Distribution: México:Oaxaca.
Pyticeroides pullis Opitz, 2010: 22. Holotype male (FSCA). Type locality: Brazil, 62 kmSW Ariquemes, Fazenda Rancho Grande. Fig. 254 (holotype). Distribution: Ecuador:Orellana.
Pyticeroides quadratus Opitz, 2007: 132. Holotype female (DEIG). Type locality:Brazil: Goias: Jatai. Primary type: Fig. 255 (holotype): Distribution: Brazil: Goias.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

61
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Pyticeroides rifkindi Opitz, 2007: 127. Holotype male (LACM). Type locality: CostaRica: San José: Cerro de Escazu, Mtinilla. Fig. 256 (holotype). Distribution: CostaRica: San José.
Pyticeroides roseicollis Opitz, 2007: 123. Holotype male (INBC). Type locality: CostaRica: Guanacaste: 3 km SE Rio Naranjo. Fig. 257 (holotype). Distribution: CostaRica: Guanacaste. Panamá: Panamá.
Pyticeroides similis Opitz, 2007: 113. Holotype male (MCNZ). Type locality: Brazil:Rondonia: 62 km SE Ariquemes. Fig. 258 (holotype). Distribution: Brazil: Rondônia.
Pyticeroides stenotes Opitz, 2007: 128. Holotype female (IZAV). Type locality:Venezuela: Aragua: El Limón. Fig. 259 (holotype). Distribution: Venezuela: Aragua.
Pyticeroides strictus Opitz, 2007: 117. Holotype male (MNHN). Type locality: Bolivia:Cochabamba. Fig. 260 (holotype). Distribution: Colombia: Meta. Bolivia:Cochabamba. Peru: Madre de Dios.
Pyticeroides tessares Opitz, 2007: 126. Holotype female (FSCA). Type locality:Honduras: Olancho: La Muralla. Fig. 261 (holotype). Distribution: Honduras:Olancho. Guatemala: Izabal.
Pyticeroides trilineatum (Chevrolat) (Ichnea), 1874: 324. Type locality: México:Jalisco. Lectotype male (MNHN). Fig. 262 (lectotype). Corporaal 1950: 253. Opitz2007: 136. Distribution: México: Jalisco: Veracruz.
Pyticeroides turbosiris Opitz, 2010: 23. Holotype female (USNM). Type locality:Ecuador: Orellana: Tiputini Biodiversity Station. Fig. 263 (holotype). Distribution:Ecuador: Orellana.
Pyticeroides ustulatis Opitz, 2010: 24. Holotype male (USNM). Type locality: Ecuador:Orellana: 1 Km S Okone Camp. Fig. 264 (holotype). Distribution: Ecuador: Orellana.
Pyticeroides vichadium Opitz, 2007: 117. Holotype male (IAVH). Type locality:Colombia: Vichada: PNN Tuparro, Cerro Tomas. Fig. 265 (holotype). Distribution:Colombia: Vichada.
Genus Silveirasia Nemésio, 2006: 77.Type species: Chaetophloeus hispidus (Opitz), 2004: 21 (by monotypy).
Generic synonyms: None.Most recent generic revision: NEMÉSIO 2006. Distribution: Brazil: Rio Grande do Sul.Number of species: 1.Silveirasia hispida (Opitz) (Chaetophloeus), 2004: 23. Holotype male (MCNZ). Type
locality: Brazil: Rio Grande do Sul: São Francisco de Paula. Fig. 266 (paratype).Mecke, et al, 2001: 119 (misidentified as Plocamocera sp.). Distribution: Brazil: RioGrande do Sul: São Francisco de Paula, forest reserve Pró-Mata.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

62
W. OPITZ
Genus Stegnoclava Opitz, 2010b: 9.Type species: Ichnea fumigata Gorham, 1877b: 414 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2010b.Distribution: Argentina, Costa Rica, Peru, Brazil.Number of species: 3.Stegnoclava fumigata (Gorham) (Ichnea), 1877b: 414. Lectotype female (BMNH).
Type locality: Brazil: Fig. 267 (homotype). Corporaal 1950a: 270 (Ichnea). Opitz2010b: 14. Distribution: Peru: Loreto, Madre de Dios. Brazil: Amazonia; Pará.
Stegnoclava velatis Opitz, 2010b: 14. Holotype male (FSCA). Type locality: Brazil:Mato Grosso: Sinop. Fig. 268 (holotype). Distribution: Brazil: Mato Grosso.
Stegnoclava zorropsis Opitz, 2010b: 17. Holotype female (INBC). Type locality: CostaRica: Heredia. Fig. 269 (holotype). Distribution: Costa Rica: Heredia
Genus Turbophloeus Opitz, 2008c: 35.Type species: Epiphloeus simplex Schenkling, 1900: 397 (by original designation).
Generic synonyms: None.Most recent generic revision: OPITZ 2008c. Distribution: Columbia, Bolivia.Number of species: 1.Turbophloeus simplex (Schenkling) (Epiphloeus), 1900: 397. Lectotype male (DEIG).
Type locality: Bolivia. Fig. 270 (homotype). Corporaal 1950a: 254 (Epiphloeus).Distribution: Colombia: Boyacá; Caquetá; Huila; Norte de Santander; Risaralda.Bolivia: Cochabamba; Santa Cruz.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

63
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
ReferencesAGASSIZ L. 1846. Nomenclatoris zoologici. Index universalis, continens nomina systematica classium, ordinum,
familiarum et generum animalium omnium, tam viventium quam fossilium, secundum ordinemalphabeticum unicum disposita, adjectis homonymiis plantarum, nec non variis adnotationibus etemendationibus. Fasciculus XII. Jent and Gassmann, Soloduri.
ARNETT JR. R.H., G.A. SAMUELSON & G.M. NISHIDA 1993. The insect and spider collections of the world. Floraand fauna handbook no. 11, 2nd ed. Gainesville, Fl: Sandhill Crane Press.
BALDUF W.V. 1935. Cleridae. Checkered Beetles. In: W.V. Balduf (editor), The bionomics of entomophagousColeoptera. pp. 109�111. John S. Swift Co. New York.
BARR W.F. 2006. A new genus and species of Epiphloeinae from Brazil (Coleoptera: Cleridae). Insecta Mundi20 (3�4): 215�26.
BARTLETT J.S. 2013. A revised taxonomy for Australian Teneropsis Chapin and Cregya LeConte (Coleoptera:Cleridae: Korynetinae). Memoirs of the Queensland Museum-Nature 58: 411�420.
BEAL A.B., & C.L. MASSEY 1945. Bark beetles and ambrosia beetles (Coleoptera: Scolytoidea): With specialreference to species occurring in North Carolina. Duke University School of Forestry Bulletin 10: 1�178.
BOUCHARD, P., Y. BOUSQUET, A.E. DAVIES, M.A. ALONSO-ZARAZAGA, J.F. LAWRENCE, C.H. LYAL, A.F. NEWTON,C.A.M. REID, M. SCHMITT, S.A. �LIPIÑSKI & A.B.T. SMITH 2011: Family-group names in Coleoptera(Insecta). Zookeys 88: 1�972.
BROWN R.W. 1956. Composition of scientific words. 2nd ed. Baltimore, MD. Reese Press.CALAHAN P.S. 1975. Insect antenna with special reference to the mechanism of scent detection and the evolution
of the sensilla. International Journal of Insect Morphology and Embryology 4(5): 381�430.CHAMPLAIN A.B. 1920. Larvae of North American beetles of the family Cleridae. Proceedings of the United
States National Museum 57(2323): 575�649.CHAPIN E.A. 1927. The beetles of the family Cleridae collected on the Mulford Biological Exploration of the
Amazon Basin 1921�1922. Proceedings of the United States National Museum 71(2): 1�10.CHEVROLAT M.A. 1843. Description de vingt-quatre nouvelles espéces de terediles, pour faire suite a la
monographie das clairones, de M. le docteur Klug. Annales de la Société Entomologique de France. Series2. 1: 31�42.
CHEVROLAT M.A. 1874. Catalogue des Clerides de la collecion de M.A. Chevrolat. Revue et Magasin deZoologie (Series 3) 2 (6): 252�329.
CHEVROLAT M.A. 1876. Mémoire sur la famille des Clérites. Buquet: Paris.CORPORAAL J.B. 1949. Sixth series of notes on systematics and synonymy. Entomologische Berichten 12:
398�399.CORPORAAL J.B. 1950. Cleridae. In: W. D. Hincks (editor): Coleopterorum Catalogus Supplementa, Pars 23
(editio secunda). 373 pp. Gravenhage: W. Junk�s. CORPORAAL J.B. 1950b. On some primary homonyms. Entomologische Berichten 13: 93�94.CRAIGHEAD F.C. 1950. Insect and enemies of eastern forests. Family Cleridae. United States Department of
Agriculture Miscellaneous Publications, no. 657: 197�203.DOBZHANSKY T. 1937. Genetics and the origin of species. New York: Columbia University Press.EKIS G. (now OPITZ W.) 1975. Taxonomic and nomenclatural status of clerid taxa described by Massimiliano
Spinola (1780�1857) (Coleoptera, Cleridae). Bollettino del Museo di Zoologia dell�Università di Torino 1:1�80.
EKIS G. 1977 Classification, phylogeny, and zoogeography of the genus Perilypus (Coleoptera: Cleridae).Smithsonian Contribution to Zoology 227: 1�138.
EKIS G. & A.P. GUPTA 1971. Digestive system of Cleridae (Coleoptera). International Journal of InsectMorphology and Embryology 1(1):51�86.
ERICKSON W.F. 1847. Conspectus Insectorus Coleopterorum quae in republica Peruana observata sunt. XII.Fam. Clerii. Archiv fuer Naturgeschichte 13: 85�86.
GERMAR F. 1912. Untersuchungen über den Bau und die Lebensweisse der Lymexyloniden, speziell desHylecoetus dermestoides L. In: C.T. SIEBOLD & A. KÖLLIKER (editors): Zeitschrift für WissenschafflicheZoologie. 1912, Engleman, Leipzig.
GOLOBOFF P.A. 2003. NONA version 2.0. A tree searching program. Distributed by the author.GORHAM H.S. 1877a. Descriptions of new species of Cleridae. Transactions of the Royal Entomological Society
of London 18(3): 245�263.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

64
W. OPITZ
GORHAM H.S. 1877b. Descriptions of new species of Cleridae, with notes on the genera and corrections ofsynonymy. Transactions of the Royal Entomological Society of London 1877(4): 401�426.
GORHAM H.S. 1882. Cleridae. Biologia Centrali-Americana. Insecta, Coleoptera. 3: 129�168. GORHAM H.S. 1883. Cleridae. Biologia Centrali-Americana. Insecta, Coleoptera. 3: 169�192. GORHAM H.S. 1986. Cleridae. Biologia Centrali-Americana, Insecta, Coleoptera,. 3(2): Supplement page 341.GRUNER L. 1974. Biologie et dégats d�Hexacolus guyanensis Schedl. Dans les plantations d�acajou rouge
(Swietenia macrophylla King., Méliacée) en Guadeloupe (Coleoptera: Scolytidae). Annales des Sciencesforestiéres, Institute Nacional de la Recherche Agronomique 31(2): 111�128.
GUNTER N.L., J.M. LEAVENGOOD, J.S. BARTLETT, E.G. CHAPMAN & S.L. CAMERON 2013. A molecular phylogenyof the checkered beetles and a description of Epiclininae a new subfamily (Coleoptera: Cleroidea:Cleridae). Systematic Entomology 38(3): 626�636.
HENNIG W. 1966. Phylogenetic systematics. Urbana, Illinois: University Press.HESPENHEIDE H.A. 1973. A novel mimicry complex: Beetles and flies. Journal of Entomology 48(1): 49�56.HORN G.H. 1896. Coleoptera of Baja California (Supplement II). Proceedings of the California Academy of
Sciences. (Series 2). 6: 367�381. ICZN (International Commission on Zoological Nomenclature) 1999. International Code of Zoological
Nomenclature. Fourth edition. The International Trust for Zoological Nomenclature, London.JORON M.J. & MALLET L.B. 1998. Diversity in mimicry: paradox or paradigm? Trends in Ecology and Evolution
13(11): 461�466.KLUG J.C.F. 1842. Versuch einer systematischen Bestimmung und Auseinandersetzung der Gattunger und
Auseinandersetzung der Gattungen und Arten der Clerii, einer Insectenfamilie aus der Ordung derColeopteren. Abhandlungen Berlin Akademie der Berlin Wissenschaften. Pp. 259�397.
KNULL J.N. 1939. Four new Arizona Coleoptera (Cleridae, Corynetidae and Buprestidae). Annals of theEntomological Siciety of America 32: 27�31.
KNULL J.N. 1951. The checkered beetles of Ohio (Coleoptera: Cleridae). Ohio Biological Survey 8(2): 269�350.KOLIBÁČ J. 1997. Classification of the subfamilies of Cleridae (Coleoptera: Cleroidea). Acta Musei Moraviae
Scientiae Naturales 81: 307�361.KOLIBÁČ J. 2010. Cleridae Latreille. p. 260. In: R.A.B. LESCHEN, R.G. BEUTEL and J.F. LAWRENCE. Handbook
of Zoology, Coleoptera, Beetles, Vol 2. Walter de Gruyter: Berlin.KUWERT A. 1893. Die Epiphloinen Gattungen der Cleriden und einige neue Arten derselben. Annales de la
Societe Entomologique de Belgique 37: 492�497.KUWERT A. 1894. Die Epiphloinen Gattungen der Cleriden und einige neue Arten derselben. Annales de la
Societe Entomologique de Belgique 38: 6�13.LAPORTE F.L. 1836. Etudes entomologiques, ou descriptions d�insectes nouveaux et observations sur la
synonymie. Review Entomologique (Published by G. Silbermann) 4: 5�60.LAWRENCE F.J., R.G. BEUTEL, R.A.B. LESCHEN & S.A. �LIPIÑSKI. 2010. Changes in classification and list of
families and subfamilies. p. 5. In: R.A.B. LESCHEN, R.G. BEUTEL & J.F. LAWRENCE: Handbook of Zoology,Coleoptera, Beetles, Vol 2. Walter de Gruyter: Berlin.
LAWRENCE F.J., S.A. �LIPIÑSKI, A.E. SEAGO, M.K. THAYER, A.F. NEWTON & A.E. MARVALDI 2011. Phylogeny ofthe Coleoptera based on morphological characters of adults and larvae. Annales Zoologicy (Warszawa)61(1): 1�217.
LEPESME P. 1947. Famille des Cleridae. In E. FLEUTIAX, C. LEGROS, P. LEPESME et R. PAULIAN: Faune de L�Empire Français. VII. Colèoptéres des Antilles. Volume 1. Paris: Office de la Recherche ScientifiqueColoniale.
LOHDE R. 1899. Umtaufe einiger Cleridaen-Arten. Entomologischen Nachrichten 25: 305.MAWDSLEY J. 1992. A re-evaluation of the checkered beetles from the upper Miocene of Florissant, Colorado
(Coleoptera: Cleridae). Psyche 99(2�3): 129�139.MAYR E. 1963. Populations, Species and Evolution. An abridgment of Animal Species and Evolution.
Cambridge: The Belknap Press of Harvard University Press.MECKE R., M.H.M. GALILEO & W. ENGELS 2001. New records of insects associated with Araucaria Trees:
Phytophagous Coleoptera and Hymenoptera and their natural enemies. Studies on Neotropical Fauna andEnvironment 36(2): 113�124.
MENIER J.J. 1985. Quelques aspects du �mimétisme� chez les Cleridae. Bulletin de la Société entomologique deFrance 90: 1071�1083.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

65
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
NEMÉSIO A. 2005. Two homonomies in Epiphloeinae (Coleoptera: Cleridae): new generic names for ArenariaOpitz and Chaetophloeus Opitz. Lundiana 7(1): 77.
NICHOLS S.W. 1989. The Torre-Bueno Glossary of Entomology. New York: Entomological Society, AmericanMuseum of Natural History.
NIXON K. C. 2002. Winclada ver. 1.00.08. Published by the author.NIXON C.N. & Carpenter J.A. 1993. On outgroups. Cladistics 9: 413�426.OPITZ W. 1997. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). Part I.
The genera of Epiphloeinae. Insecta Mundi 11(1): 51�96.OPITZ W. 2002. Family 73. Cleridae Latreille 1804. Pp. 267�280. In: R.H. ARNETT, Jr., M.C. THOMAS, P.E.
SKELLEY & J.H. FRANK (eds): American Beetles, vol. 2. Boca Raton: CRC Press.OPITZ W. 2004. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). Part
II. The genera Chaetophloeus Opitz and Plocamocera Spinola. Bulletin of the American Museum of NaturalHistory 280: 1�82.
OPITZ W. 2005. Classification, natural history, and evolution of the genus Aphelocerus Kirsch (Coleoptera:Cleridae: Clerinae). Bulletin of the American Museum of Natural History. 293: 1�128.
OPITZ W. 2006. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartIII. The genera Parvochaetus, n. gen., Amboakis, n. gen, and Ellipotoma Spinola. Insecta Mundi 20(3�4):97�164.
OPITZ W. 2007. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartIV. The genera Pyticeroides Kuwert and Diapromeces Opitz. Entomologica Basiliensia et Collectionis Frey29: 77�166.
OPITZ W. 2008a. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartV. Decorosa Opitz, a new genus of checkered beetles from Hispaniola with description of its four newspecies. American Museum Novitates 3628: 1�19.
OPITZ W. 2008b. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartVI. The genera Epiphlaeus Spinola and Opitzius Barr. Annales Zoologoci (Warszawa) 58: 1�34.
OPITZ W. 2008c. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartVII. The genera Hapsidopteris Opitz, Iontoclerus Opitz, Katamyurus Opitz, Megatrachys Opitz, OpitziaNemésio, Pennasolis Opitz, new genus, Pericales Opitz, new genus, Pteroferus Opitz, new genus, andTurbophloeus, new genus. Zootaxa 1754: 1�40.
OPITZ W. & L. HERMAN 2009. Epiphloeinae Kuwert and Ichneinae, 1841: Two names for the same subfamilyof checkered beetles (Coleoptera: Cleridae). The Coleopterists Bulletin 63(2): 183�189.
OPITZ W. 2010a. New taxa of Epiphloeinae Kuwert (Cleridae) and Chaetosomatidae Crowson (Cleroidea).Insecta Mundi 0123: 1�28.
OPITZ W. 2010b. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartVIII. The genera Acanthocollum Opitz, Stegnoclava Opitz, and Ichnea Laporte. Coleopterists Bulletin 64(4) Supplement Monograph 9: 1�65.
OPITZ W. 2010c. Classification, evolution, and subfamily composition of the Cleridae, and generic content andkey of the subfamilies (Coleoptera: Cleroidea). Entomologica Basiliensia et Collectionis Frey 32: 31�128.
OPITZ W. 2011a. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartIX. The genus Megaphloeus Opitz). Entomologica Basiliensia et Collectionis Frey 33: 63�132.
OPITZ W. 2011b. Classification, natural history, and evolution of the Epiphloeinae (Coleoptera: Cleridae). PartIX. The genus Madoniella Pic. Entomologica Basiliensia et Collectionis Frey 33: 133�248.
PECK S.B. 2005. A checklist of the beetles of Cuba with data on distributions and bionomics (Insecta:Coleoptera). Arthropods of Florida and Neighboring Land Areas 18: 1�241.
PIC M. 1935. Noveautés diverses. Mélanges Exotico-Entomologiques 65: 1�36.PIC M. 1936. Nouveaux colèoptéres exotiques. Bulletin de la Société Zoologique de France 61: 126�128.PIC M. 1942. Opuscula martialis. 7: 3.RIFKIND J., TOLEDO V.H., & CORONA A.M.A 2010. New species of Cleridae (Coleoptera) from Morelos,
Mexico. Zootaxa 2659: 53�59.SAY T. 1825. Description of new species of coleopterous insects inhabiting the United States. Journal of the
Academy of Natural Sciences of Philadelphia 5(1): 160�204.SCHENKLING S. 1900. Neue amerikanische Cleriden nebst Bemerkungen zu schon beschriebenen Arten
Deutchen. Entomologische Zeitschrift 1900: 385�409.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

66
W. OPITZ
SCHENKLING S. 1906. Die Cleridaen des Deutschen Entomologischen National-Museums, nebstBeschreibungen neuer Arten. Deutshe Entomologische Zeitschrift 1: 241�320.
SPINOLA M. 1841. Monographie des Térédiles. Revue Zoologique, par La Société Cuvierienne 4: 70�76.SPINOLA M. 1844a. Essai monographique sur les Clerites: insects Coléoptéres. 1, i�ix, 1�386. Genes.SPINOLA M. 1844b. Essai monographique sur les Clerites: insects Coléoptéres. 2, 1�216, pls. 1�xlvii. Genes.STANDFUSS M. 1896. Handbuch der paläarktischen Gross-Scmetterlinge für Forscher und Sammler. Jena:
Gustav FischerTHOMSON J.M. 1860. Matériaux pur servir a une monographie nouvelle de la famille des clerids. Musée
Scientifique 1860: 46�67.VANDYKE E.C. 1923. New species of Coleoptera from California. Bulletin of the Brooklyn Entomological
Society 18(2): 37�53.WOLCOTT A.B. 1927. A review of the Cleridae of Costa Rica. Coleopterological Contributions 1(10): 1�104. WOLCOTT A.B. 1944. A generic review of the subfamily Phyllobaeninae (olim Hydnocerinae) (Col.). Journal of
the New York Entomological Society 52(2): 121�152.WOLCOTT A.B. & E.A. CHAPIN 1918. Notes on Cleridae. Bulletin of the Brooklyn Entomological Society 13(5):
107�108.ZAYAS F. 1988. Entomofauna Cubana, orden Coleoptera, seperata descripcion de nuevas especies. La Habana:
Editioral Cientifico-Técnica.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

67
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 1�12. Habitus: 1, Acanthocollum melanurum; 2, Amboakis ampla; 3, A. anapsis; 4, A. antegalba; 5, A.atra; 6, A. barinas; 7, A. binotonis; 8, A. capitata; 9, A. cauca; 10, A. charis; 11, A. demagna; 12, A. diffusa.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

68
W. OPITZ
Figs 13�24. Habitus: 13, Amboakis epiomedia; 14, A. erythrohapsis; 15, A. flavicollis; 16, A. funebris; 17, A.incondita; 18, A. katatonis; 19, A. linitis; 20, A. mica; 21, A. micula; 22, A. nitida; 23, A. prolata; 24, A.rudis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

69
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 25�36. Habitus: 25, Amboakis selva; 26, A. stenosis; 27, A. taruma; 28, A. vesca; 29, A. vaodani; 30,Decaphloeus vitticollis; 31, Decorosa aladecoris; 32, D. iviei; 33, D. limatula; 34, D. neiba; 35.Diapromeces aclydis; 36. Ellipotoma tenuiformis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

70
W. OPITZ
Figs 37�48. Habitus: 37, Ellipotoma turmalis; 38, Epiphloeus adonis; 39, E. duodecimmaculatus; 40, E. erwini;41, E. fundurufus; 42, E. princeps; 43, E. pulcherrimus; 44, E. quattuordecimmaculatus; 45, E. tigrinus;46, Hapsidopteris diastenus; 47, Ichnea acanthomelina; 48, I. aequinoctialis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

71
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 49�60. Habitus: 49, Ichnea aterrima; 50, I. atra; 51, I. callanga; 52, I. digna; 53, I. dimidiatipennis; 54, I.divisa; 55, I. frenata; 56, I. gregata; 57, I. gremia; 58, I. lycoides; 59, I. marginella; 60, I. mimica.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

72
W. OPITZ
Figs 61�72. Habitus: 61, Ichnea opaca; 62, I. plumbea; 63, I. praeusta; 64, I. procera; 65, I. incerta; 66,Iontoclerus humeralis; 67, I. sericeus; 68, Katamyurus albopaniculus; 69, K. paxillus; 70, Madoniellaabacula; 71, M. adona; 72, M. aktis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

73
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 73�84. Habitus: 73, Madoniella anapsis; 74, M. antennatra; 75, M. apotoma; 76, M. apsis; 77, M. aspera;78, M. avina; 79, M. basilaris; 80, M. basilia; 81, M. bilineata; 82, M. bullalis; 83, M. cardinalis; 84, M.careorita.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
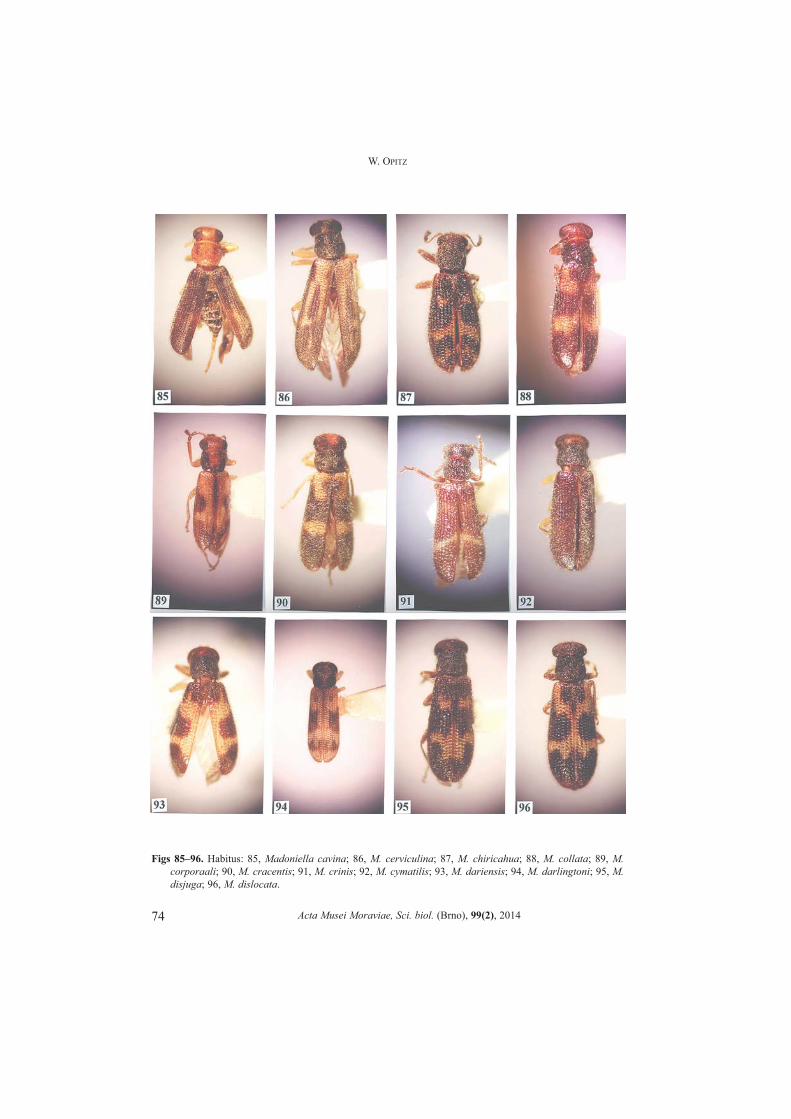
74
W. OPITZ
Figs 85�96. Habitus: 85, Madoniella cavina; 86, M. cerviculina; 87, M. chiricahua; 88, M. collata; 89, M.corporaali; 90, M. cracentis; 91, M. crinis; 92, M. cymatilis; 93, M. dariensis; 94, M. darlingtoni; 95, M.disjuga; 96, M. dislocata.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
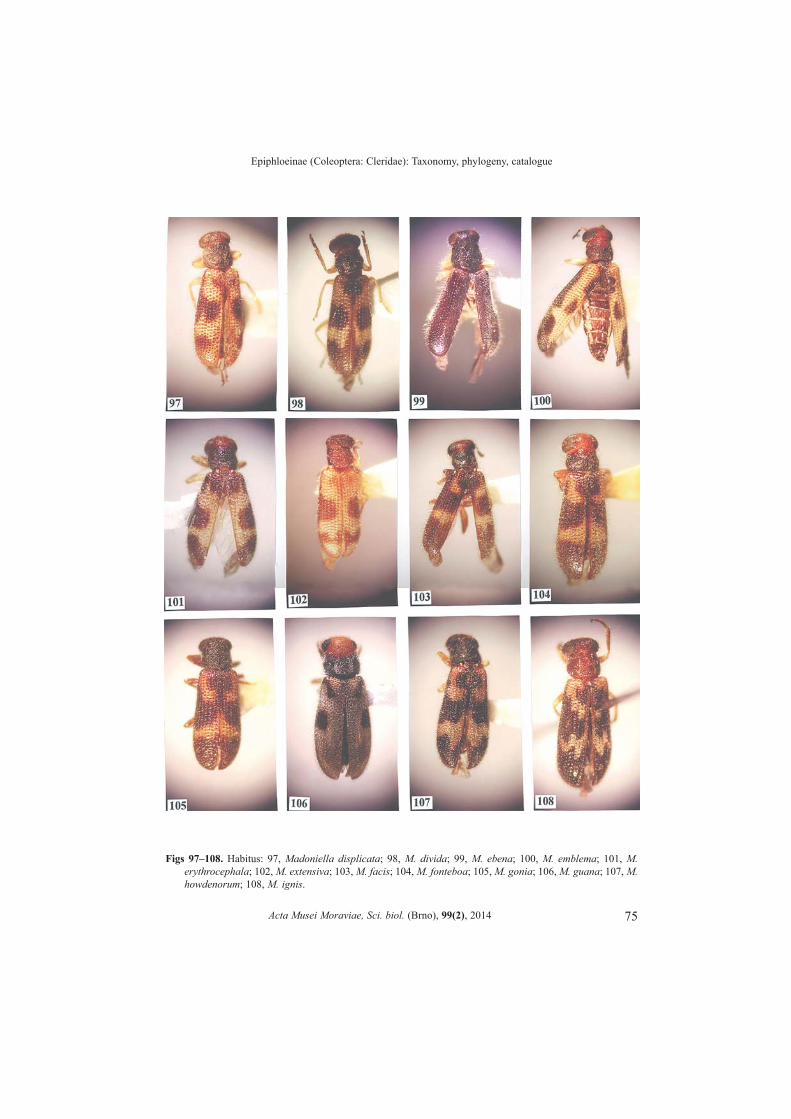
75
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 97�108. Habitus: 97, Madoniella displicata; 98, M. divida; 99, M. ebena; 100, M. emblema; 101, M.erythrocephala; 102, M. extensiva; 103, M. facis; 104, M. fonteboa; 105, M. gonia; 106, M. guana; 107, M.howdenorum; 108, M. ignis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

76
W. OPITZ
Figs 109�120. Habitus: 109, Madoniella infula; 110, M. insignis; 111, M. knullorum; 112, M. kuehlorum; 113,M. latinopsis; 114, M. leona; 115, M. linea; 116, M. lineola; 117, M. lurida; 118, M. magdalena; 119, M.maxicornis; 120, M. melina.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

77
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 121�132. Habitus: 121, Madoniella merga; 122, M. minor; 123, M. nana; 124, M. patula; 125, M.nebulosa; 126, M. orientalis; 127, M. orosiensis; 128, M. pedalis; 129, M. pellis; 130, M. peninsularis; 131,M. picis; 132, M. pinicola.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

78
W. OPITZ
Figs 133�144. Habitus: 133, Madoniella plenita; 134, M. pumilis; 135, M. punctata; 136, M. quintana; 137, M.rectangularis; 138, M. redacta; 139, M. rubida; 140. M. spilota; 141, M. storea; 142, M. tegetis; 143, M.taxis; 144, M. thomasi.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

79
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 145�156. Habitus: 145, Madoniella vogti; 146, M. welderi; 147, M. zonula; 148. Megaphloeus absentis;149, M. animosus; 150, M. bulatus; 151, M. cartus; 152, M. circinus; 153, M. fucoaquilus; 154, M.lividipes; 155, M. longius; 156, M. marginipes.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
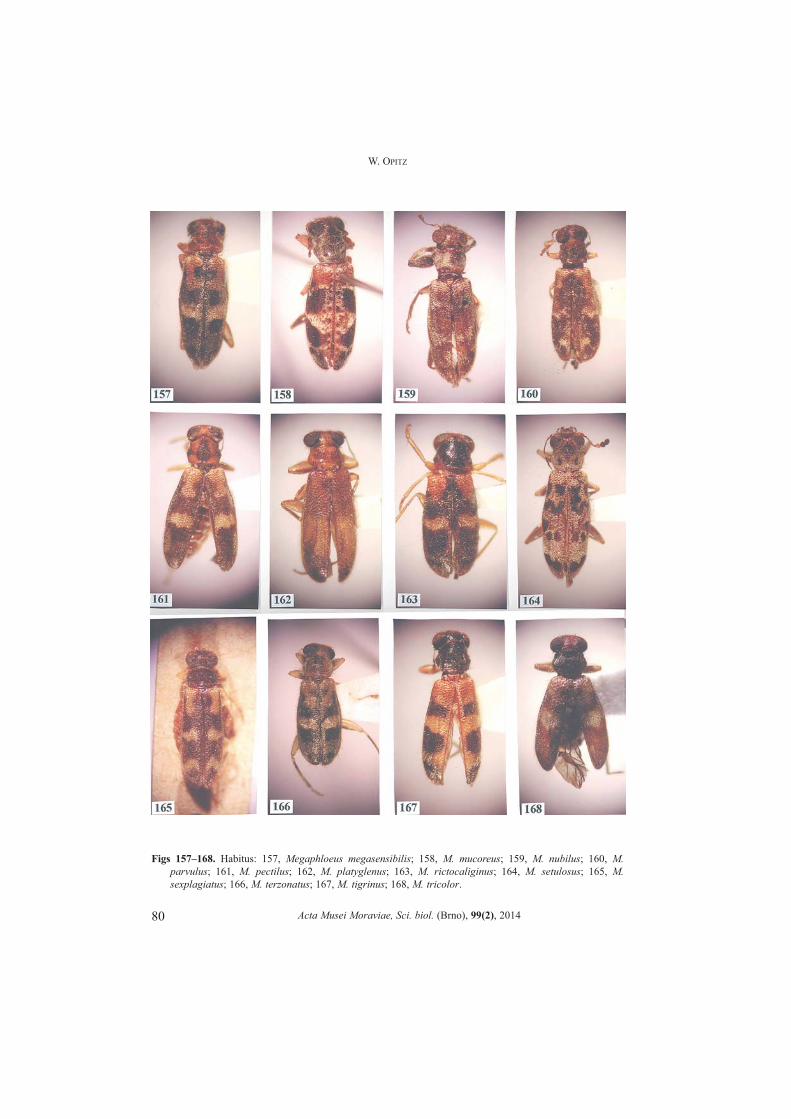
80
W. OPITZ
Figs 157�168. Habitus: 157, Megaphloeus megasensibilis; 158, M. mucoreus; 159, M. nubilus; 160, M.parvulus; 161, M. pectilus; 162, M. platyglenus; 163, M. rictocaliginus; 164, M. setulosus; 165, M.sexplagiatus; 166, M. terzonatus; 167, M. tigrinus; 168, M. tricolor.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

81
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 169�180. Habitus: 169, Megaphloeus ustafinis; 170, M. ustus; 171, M. variegatus; 172, M. velutinus; 173,M. vitellinus; 174, Megatrachys bibara; 175, M. paniculus; 176, M. truncatia; 177, Opitzia apicula; 178,O. chiapas; 179, Opitzius thoracicus; 180, Parvochaetus albicornis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

82
W. OPITZ
Figs 181�192. Habitus: 181, Parvochaetus froeschneri; 182, P. fucolatus; 183, P. linearis; 184, P. sandaracus;185, Pennasolis californica; 186, P. merkeli; 187, P. opitzi; 188, Pericales albogilvus; 189, Plocamoceraaliguantula; 190, P. ambra; 191, P. argentea; 192, P. aspera.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

83
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 193�204. Habitus: 193, Plocamocera aura; 194, P. auratilis; 195, P. baria; 196, P. bispina; 197, P.bolivari; 198, P. buenavista; 199, P. carnegei; 200, P. castanea; 201, P. clinata; 202, P. coactilis; 203, P.confrater; 204, P. insula.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

84
W. OPITZ
Figs 205�216. Habitus: 205, Plocamocera iota; 206, P. jayhawkalis; 207, P. lena; 208, P. lucis; 209, P.manausensis; 210, P. minima; 211, P. onorei; 212, P. paris; 213, P. procera; 214, P. prolixa; 215, P. pupula;216, P. quadrula.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
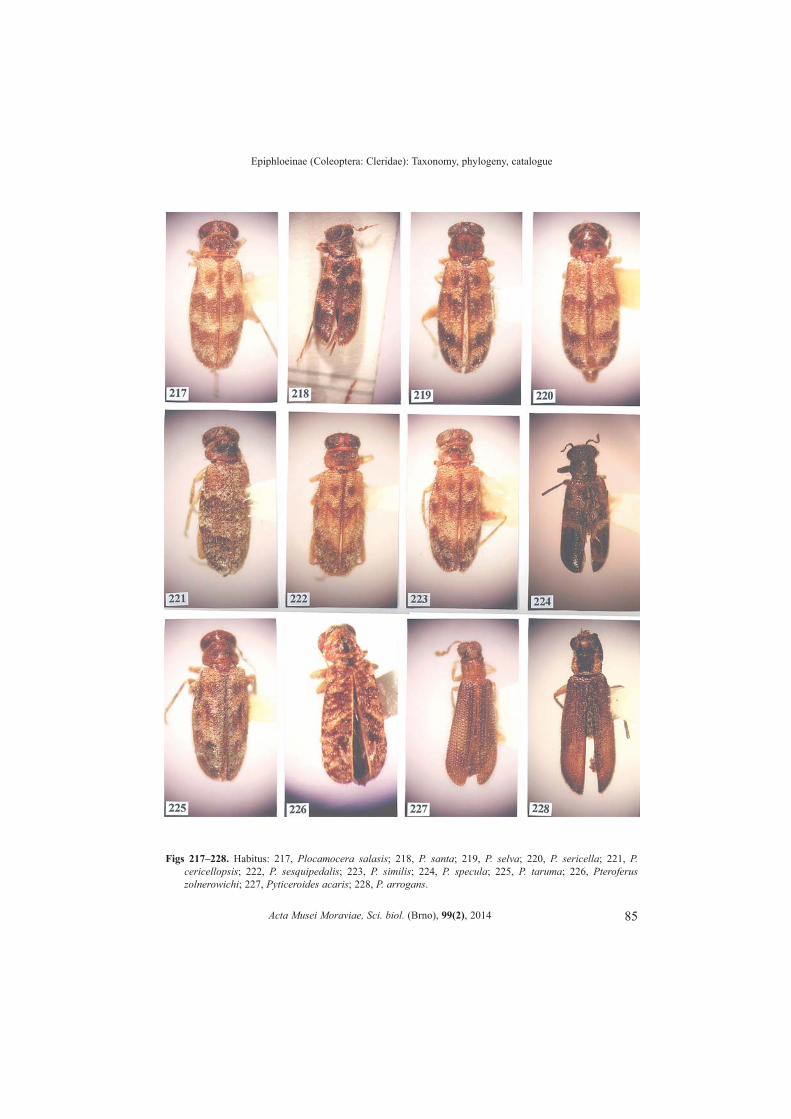
85
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 217�228. Habitus: 217, Plocamocera salasis; 218, P. santa; 219, P. selva; 220, P. sericella; 221, P.cericellopsis; 222, P. sesquipedalis; 223, P. similis; 224, P. specula; 225, P. taruma; 226, Pteroferuszolnerowichi; 227, Pyticeroides acaris; 228, P. arrogans.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

86
W. OPITZ
Figs 229�240. Habitus: 229, Pyticeroides caecoatus; 230, P. caligneus; 231, P. chiriquianum; 232, P.cinericeus; 233, P. decurialis; 234, P. enormis; 235, P. eurides; 236, P. fustis; 237, P. hansoni; 238, P.ichnopsis; 239, P. inconscriptus; 240, P. inexilis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

87
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 241�252. Habitus: 241, Pyticeroides iscus; 242, P. jubaris; 243, P. laticornis; 244, P. laticentis; 245, P.luteolus; 246, P. maesi; 247, P. manni; 248, P. moraguesi; 249, P. notialis; 250, P. parvoporis; 251, P.petilus; 252, P. pinnacerinis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

88
W. OPITZ
Figs 253�264. Habitus: 253, Pyticeroides plautus; 254, P. pullis; 255, P. quadratus; 256, P. rifkindi; 257, P.roseicollis; 258, P. similis; 259. P. stenotes; 260, P. strictus; 261, P. tessares; 262, P. trilineatum; 263, P.turbosiris; 264, P. ustulatis.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

89
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Figs 265�270. Habitus: 265, Pyticeroides vichadium; 266, Silverasia hispida; 267, Stegnoclava fumigata; 268,S. velatis; 269, S. zorropsis; 270, Turbophloeus simplex.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
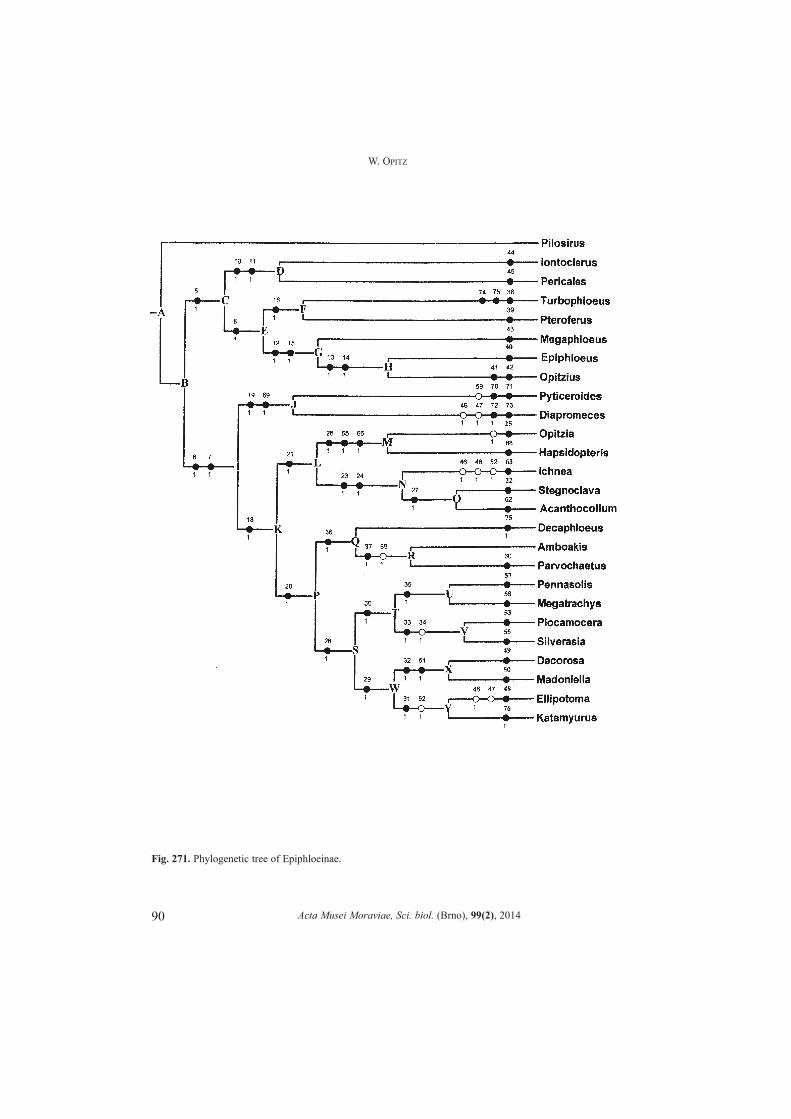
90
W. OPITZ
Fig. 271. Phylogenetic tree of Epiphloeinae.
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014

91
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
abacula (Madoniella) absentis (Megaphloeus)acanthomelina (Ichnea)acaris (Pyticeroides)adona (Madoniella)adonis (Epiphloeus)aequinoctialis (Ichnea)
funesta (Ichnea)disjuncta (Ichnea)histrica (Ichnea)confluence (Ichnea)spinolai (Ichnea)
aktis (Madoniella)aladecoris (Decorosa)albicornis (Parvochaetus)albogilvus (Pericalesalbopaniculus (Katamyurus)aliguantula (Plocamocera)ambra (Plocamocera)ampla (Amboakis)anapsis (Amboakis)anapsis (Madoniella)animosus (Megaphloeus)antegalba (Amboakis)antenatra (Madoniella)apicula (Opitzia)apotoma (Madoniella)apsis (Madoniella)argentea (Plocamocera)arogans (Pyticeroides)
Megatrachys OpitzOpitsia NemésioOpitzius BarrParvochaetus OpitzPennasolis OpitzPericales OpitzPlocamocera SpinolaPteroferus OpitzPyticeroides Kuwert
Neichnea Wolcott & ChapinSilveirasia NemésioStegnoclava OpitzTurbophloeus Opitz
Acanthocollum OpitzAmboakis OpitzDecaphloeus OpitzDecorosa OpitzDiapromeces OpitzEllipotoma SpinolaEpiphloeus SpinolaHapsidopteris OpitzIchnea CastelnauIontoclerus OpitzKatamyurus OpitzMadoniella Pic
Phlogistosternus WolcottMegaphloeus Opitz
Index of genera and generic synonyms
Index of species and specific synonyms
aspera (Madoniella)aspera (Plocamocera)aterrima (Ichnea)
mexicana (Ichnea)religiosa (Ichnea)vitticollis (Ichnea)kuwerti (Ichnea)elongate (Ichnea)
atra (Ichnea)atra (Amboakis)aura (Plocamocera)auratilis (Plocamocera)avina (Madoniella)baria (Plocamocera)barinas (Amboakis)basilaris (Madoniella)basilia (Madoniella)bibara (Megatrachys)bilineata (Madoniella)binotonis (Amboakis)bispina (Plocamocera)bolivari (Plocamocera)buenavista (Plocamocera)bulatus (Megaphloeus)bullalis (Madoniella)callanga (Ichnea)californica (Pennasolis)caligneus (Pyticeroides)capitata (Amboakis)cardinalis (Madoniella)

92
W. OPITZ
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
careorita (Madoniella)carnegei (Plocamocera)cartus (Megaphloeus)castanea (Plocamocera)cauca (Amboakis)caecoatus (Pyticeroides)cavina (Madoniella)cerviculina (Madoniella)charis (Amboakis)chiapas (Opitzia)chiricahua (Madoniella)chiriquianum (Pyticeroides)cinericeus (Pyticeroides)circinus (Megaphloeus)clinata (Plocamocera)coactitilis (Plocamocera)collata (Madoniella)confrater (Plocamocera)
impressicollis (Plocamocera)corporaali (Madoniella)cracentis (Madoniella)crinis (Madoniella)cymatilis (Madoniella)dariensis (Madoniella)darlingtoni (Madoniella)decurialis (Pyticeroides)diastenus (Hapsidopteris)diffusa (Amboakis)digna (Ichnea)dimidiatipennis (Ichnea)disjuga (Madoniella)dislocata (Madoniella)
distrophum (Enoplium)transversalis (Phyllobaenus)
displicata (Madoniella)divida (Madoniella)divisa (Ichnea)
enoplioides (Ichnea)duodecimmaculatus (Epiphloeus)
pantherinus (Enoplium)ebena (Madoniella)emagna (Amboakis)emblema (Madoniella) enormis (Pyticeroides)epiomidia (Amboakis)erwini (Epiphloeus)erythrocephala (Madoniella)erythrohapsis (Amboakis)extensiva (Madoniella) eurides (Pyticeroides)facis (Madoniella)flavicollis (Amboakis)fonteboa (Madoniella)
frenata (Ichnea)confluence (Ichnea)
disjuncta (Ichnea)femoralis (Ichnea)
mitella (Ichnea)panamensis (Ichnea)
striaticollis (Ichnea)subfasciata (Ichnea)froeschneri (Parvochaetus)fucoaquilus (Megaphloeus)fucolatus (Parvochaetus)fumigata (Stegnoclava) fundurufus (Epiphloeus)funebris (Amboakis)fustis (Pyticeroides)gonia (Madoniella)gregata (Ichnea)gremia (Ichnea)guana (Madoniella)hansoni (Pyticeroides)hispidus (Silveirasia)howdenorum (Madoniella)humeralis (Iontoclerus)ichnopsis (Pyticeroides)ignis (Madoniella)incerta (Ichnea)incondita (Amboakis)inconscriptus (Pyticeroides)inexilis (Pyticeroides)infula (Madoniella)insignis (Madoniella)insula (Plocamocera)iota (Plocamocera)iscus (Pyticeroides)iviei (Decorosa)jayhawkalis (Plocamocera)jubaris (Pyticeroides)katatonis (Amboakis)knullorum (Madoniella)kuehlorum (Madoniella)laticentis (Pyticeroides)laticornis (Pyticeroides)latinopsis (Madoniella)lena (Plocamocera)leona (Madoniella)limatula (Decorosa)linea (Madoniella)linearis (Parvochaetus)lineola (Madoniella)linitis (Amboakis)lividipes (Megaphloeus)longius (Megaphloeus)lucis (Plocamocera)

93
Epiphloeinae (Coleoptera: Cleridae): Taxonomy, phylogeny, catalogue
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
lurida (Madoniella)luteolus (Pyticeroides)lycoides (Ichnea)maesi (Pyticeroides)magdalena (Madoniella)manausensis (Plocamocera)manni (Pyticeroides)marginella (Ichnea)
laterale (Ichnea)circumcincta (Ichnea)bubescens (Ichnea)cylindricus (Hylecoetus)
marginipes (Megaphloeus)maxicornis (Madoniella)megasensibilis (Megaphloeus)melanurum (Acanthocollum)melina (Madoniella)merga (Madoniella)merkeli (Pennasolis)mica (Amboakis)micula (Amboakis)mimica (Ichnea)minima (Plocamocera)minor (Madoniella)moraquesi (Pyticeroides)mucoreus (Megaphloeus)
bakeri (Epiphloeus)balteatus (Epiphloeus) chevrolati Epiphloeus)fasciatum (Enoplium)tomentosus (Epiphloeus)
nana (Madoniella)nebulosa (Madoniella)neiba (Decorosa)nitida (Amboakis)notialis (Pyticeroides)nova (Amboakis)nubilus (Megaphloeus)onorei (Plocamocera)opaca (Ichnea)opitzi (Pennasolis)orientalis (Madoniella)orosiensis (Madoniella)paniculus (Megatrachys)paris (Plocamocera)parvoporis (Pyticeroides)parvulus (Megaphloeus)patula (Madoniella)paxillus (Katamyurus)pectilus (Megaphloeus)pedalis (Madoniella)pellis (Madoniella)peninsularis (Madoniella)
petilus (Pyticeroides)pici (Madoniella)pinicola (Madoniella)pinnacerinis (Pyticeroides)platyglenus (Megaphloeus)plautus (Pyticeroides)plenita (Madoniella)plumbea (Ichnea)
helvolicollis (Ichnea)praeusta (Ichnea)princeps (Epiphloeus)procera (Ichnea)procera (Plocamocera)prolata (Amboakis)prolixa (Plocamocera)pulcherrimus (Epiphloeus)pullis (Pyticeroides)pumilis (Madoniella)punctata (Madoniella)pupula (Plocamocera)quadratus (Pyticeroides)quadrula (Plocamocera)quattuodecimmaculatus (Epiphloeus)quintana (Madoniella)rectangularis (Madoniella)redacta (Madoniella)rictocaliginus (Megaphloeus)rifkindi (Pyticeroides)roseicollis (Pyticeroides)rubidia (Madoniella)rudis (Amboakis)salasis (Plocamocera)sandaracus (Parvochaetus)santa (Plocamocera)selva (Amboakis)selva (Plocamocera)setulosus (Megaphloeus)
debilis (Epiphloeus)obscurus (Epiphlöus)
sericella (Plocamocera)byssinus (Epiphloeus)latefasciata (Plocamocera)
sericellopsis (Plocamocera)sericeus (Iontoclerus)sesquipedalis (Plocamocera)sexplagiatus (Megaphloeus)
iracundus (Epiphloeus)similis (Plocamocera)similis (Pyticeroides)simplex (Turbophloeus)specula (Plocamocera)spilota (Madoniella)stenosis (Amboakis)

94
W. OPITZ
Acta Musei Moraviae, Sci. biol. (Brno), 99(2), 2014
stenotes (Pyticeroides)storea (Madoniella)strictus (Pyticeroides)taruma (Amboakis)taruma (Plocamocera)tegetis (Madoniella)tenuiformis (Ellipotoma)terzonatus (Megaphloeus)tessares (Pyticeroides)texis (Madoniella)thomasi (Madoniella)thoracicus (Opitzius)tigrinus (Megaphloeus)tigrinus (Epiphloeus)tricolor (Megaphloeus)
micaceus (Epiphloeus)trilineatum (Pyticeroides)truncatia (Megatrachys)turbosiris (Pyticeroides)turmalis (Ellipotoma)
ustafinis (Megaphloeus)ustulatis (Pyticeroides)ustus (Megaphloeus)variegatus (Megaphloeus)
buquetti (Epiphloeus)manni (Phyllobaenus)ornatus (Epiphloeus)velatis (Stegnoclava)velutinus (Megaphloeus)
ruficeps (Epiphlöeus)tibialis (Epiphlöeus)
vesca (Amboakis)vichadium (Pyticeroides)vitellinus (Megaphloeus)vogti (Madoniella)waodani (Amboakis)welderi (Madoniella)zolnerowichi (Pteroferus)zonula (Madoniella)zorropsis (Stegnoclava)


















