
Cireadian Rhythms in - University of California, San...
6
Cireadian Rhythms in Man A self-sustained oscillator with an inherent frequency underlies human 24-hour periodicity. Jurgen Aschoff It is well known even to the layman that each day the body temperature reaches a highest value toward the evening and a low point early in the morning. Since this phenomenon was described by Gierse in 1842 (see 1), numerous clinical and physiological studies have shown that there is ap- parently no organ and no function in the body which does not exhibit a similar daily rhythmicity. Whether we measure, hour by hour, the number of dividing cells in any tissue, the volume of urine excreted, the reaction to a drug, or the accuracy and the speed with which arithmetical problems are solved, we usually find that there is a maximum value at one time of day and a minimum value at another. All these rhythms are expressions of a so- called "physiological clock" which we have to consider as a basic feature in nearly all living systems, including uni- cellular organisms (2). As a result of extensive work by zoologists and botanists, especially during the last 15 years, we now understand some of the mechanisms involved and have good reason to assume that the rhythm origi- nates primarily in the organism itself. The results of experiments with hu- man subjects which I report on here give support to this hypothesis. In or- der to introduce the problem, I shall briefly refer to an experiment with birds which may stand as an example for many similar findings. The Biological Background An easy technique for following the rhythmic behavior of an animal with- out disturbing it is to measure its loco- motor activity. Caged finches, kept in the laboratory in an artificial light- 11 JUNE 1965 dark cycle with 12 hours of light and 12 hours of darkness, display a clear 24-hour rhythm with a typical pattern, all activity being restricted to the 12 light hours (Fig. 1, left). If this rhyth- mic behavior were a mere passive reaction to the periodicity of the en- vironment, that is, to the light-dark cycle, it should cease in conditions of constant illumination. The two curves of Fig. 1, representing sections of much longer records, demonstrate that this is not the case. The activity of the birds continues to be cyclic in continuous dim light (1.8 lux), as well as in brighter light (120 lux). Further- more, there are differences which claim our attention. Besides changes in the total amount of activity-the birds are less active in dim light, and for shorter spans of time, than in bright light- the length of the period (T) iS of spe- cial interest. In a physical oscillation, we measure the period as the time interval between any two identical phases. In biological oscillations, be- cause of the variability in the shape of the curve and its unavoidable "4noise," the situation is more complex. In many cases the sharp onsets of activity have proven to be useful phases from which to measure the period. In light of 1.8 lux, the bird starts to be active each day at about 5 a.m.; its period is very near to, but not exactly, 24 hours. Contrary to this, in light of 120 lux the bird becomes active each day 2 hours earlier than on the preceding day; its period is 22 hours. These two facts, the undamped con- tinuation of the rhythm and the devia- tion of the period from 24 hours, al- low only one plausible explanation: that this rhythm is not imposed on the organism by the environment but is truly endogenous. We are dealing with a self-sustained oscillation which is free-running under constant conditions and has its own inherent frequency. An increasing number of experimental results demonstrated that we are right in using this technical terminology and that the "clock" on which the overt rhythms are based has all the characteristics of an active oscillator, as was first outlined by Pittendrigh (3). [The opposing hypothesis, that in constant conditions the rhythm is con- trolled by an unknown exogenous 24- hour input of, possibly, cosmic origin, has been criticized several times (4) with sufficient thoroughness; I there- fore refrain from discussing it again.] To signify that under constant con- ditions the frequency of the biological oscillator deviates more or less from 24 hours, Halberg (5) introduced the term "circadian," derived from the Latin circa and dies. Under natural conditions, the circadian period is syn- chronized with (or entrained to) the period of the earth's rotation by means of periodic factors in the environment, called Zeitgebers. The day-night changes in illumination and tempera- ture are the most important of these. To illustrate this point, in Fig. 2 I have graphed the results of a second experiment in another way. The hori- zontal black bars represent the times when the bird is active (corresponding to a in Fig. 1). The experiment starts with 5 days of entrainment by an artificial light-dark cycle. Thereafter the bird is free-running in constant illumination of 0.4 lux. On the 33rd day, the Zeitgeber is reintroduced for 15 days, entraining the bird immedi- ately. Finally, the bird is again placed under constant conditions, but in light of 120 lux, instead of the 0.4 lux used earlier. It is evident from the graph (Fig. 2) that the free-running period is longer than 24 hours in dim light and shorter in bright light. This type of behavior is not unique to the bird. Experiments with many species in several laboratories gave rise to the following "circadian" rule: With in- creasing intensities of illumination, the circadian period is shortened in diurnal (light-active) animals anid lengthened in nocturnal (dark-active) animals. Re- cently, Hoffmann reviewed the facts on which the rule is based (6). The author is professor of physio1ogy and di- rector at the Max-Planck-Institut fuir Verhaltens- physiologie, 8131 Erling-Andechs, Germany. 1427
Transcript of Cireadian Rhythms in - University of California, San...

Cireadian Rhythms in Man
A self-sustained oscillator with an inherent frequencyunderlies human 24-hour periodicity.
Jurgen Aschoff
It is well known even to the laymanthat each day the body temperaturereaches a highest value toward theevening and a low point early in themorning. Since this phenomenon was
described by Gierse in 1842 (see 1),numerous clinical and physiologicalstudies have shown that there is ap-
parently no organ and no functionin the body which does not exhibit a
similar daily rhythmicity. Whether we
measure, hour by hour, the number ofdividing cells in any tissue, the volumeof urine excreted, the reaction to a
drug, or the accuracy and the speedwith which arithmetical problems are
solved, we usually find that there is a
maximum value at one time of dayand a minimum value at another. Allthese rhythms are expressions of a so-
called "physiological clock" which we
have to consider as a basic feature innearly all living systems, including uni-
cellular organisms (2). As a result ofextensive work by zoologists andbotanists, especially during the last 15years, we now understand some of themechanisms involved and have goodreason to assume that the rhythm origi-nates primarily in the organism itself.The results of experiments with hu-man subjects which I report on heregive support to this hypothesis. In or-der to introduce the problem, I shallbriefly refer to an experiment withbirds which may stand as an examplefor many similar findings.
The Biological Background
An easy technique for following therhythmic behavior of an animal with-out disturbing it is to measure its loco-motor activity. Caged finches, kept inthe laboratory in an artificial light-11 JUNE 1965
dark cycle with 12 hours of light and12 hours of darkness, display a clear24-hour rhythm with a typical pattern,all activity being restricted to the 12light hours (Fig. 1, left). If this rhyth-mic behavior were a mere passivereaction to the periodicity of the en-vironment, that is, to the light-darkcycle, it should cease in conditions ofconstant illumination. The two curvesof Fig. 1, representing sections ofmuch longer records, demonstrate thatthis is not the case. The activity ofthe birds continues to be cyclic incontinuous dim light (1.8 lux), as wellas in brighter light (120 lux). Further-more, there are differences which claimour attention. Besides changes in thetotal amount of activity-the birds areless active in dim light, and for shorterspans of time, than in bright light-the length of the period (T) iS of spe-cial interest. In a physical oscillation,we measure the period as the timeinterval between any two identicalphases. In biological oscillations, be-cause of the variability in the shapeof the curve and its unavoidable"4noise," the situation is more complex.In many cases the sharp onsets ofactivity have proven to be usefulphases from which to measure theperiod. In light of 1.8 lux, the birdstarts to be active each day at about5 a.m.; its period is very near to, butnot exactly, 24 hours. Contrary to this,in light of 120 lux the bird becomesactive each day 2 hours earlier thanon the preceding day; its period is 22hours.
These two facts, the undamped con-tinuation of the rhythm and the devia-tion of the period from 24 hours, al-low only one plausible explanation:that this rhythm is not imposed on theorganism by the environment but is
truly endogenous. We are dealing witha self-sustained oscillation which isfree-running under constant conditionsand has its own inherent frequency.An increasing number of experimentalresults demonstrated that we are rightin using this technical terminologyand that the "clock" on which theovert rhythms are based has all thecharacteristics of an active oscillator,as was first outlined by Pittendrigh(3). [The opposing hypothesis, that inconstant conditions the rhythm is con-trolled by an unknown exogenous 24-hour input of, possibly, cosmic origin,has been criticized several times (4)with sufficient thoroughness; I there-fore refrain from discussing it again.]To signify that under constant con-
ditions the frequency of the biologicaloscillator deviates more or less from24 hours, Halberg (5) introduced theterm "circadian," derived from theLatin circa and dies. Under naturalconditions, the circadian period is syn-chronized with (or entrained to) theperiod of the earth's rotation by meansof periodic factors in the environment,called Zeitgebers. The day-nightchanges in illumination and tempera-ture are the most important of these.To illustrate this point, in Fig. 2 Ihave graphed the results of a secondexperiment in another way. The hori-zontal black bars represent the timeswhen the bird is active (correspondingto a in Fig. 1). The experiment startswith 5 days of entrainment by anartificial light-dark cycle. Thereafterthe bird is free-running in constantillumination of 0.4 lux. On the 33rdday, the Zeitgeber is reintroduced for15 days, entraining the bird immedi-ately. Finally, the bird is again placedunder constant conditions, but in lightof 120 lux, instead of the 0.4 luxused earlier. It is evident from thegraph (Fig. 2) that the free-runningperiod is longer than 24 hours in dimlight and shorter in bright light. Thistype of behavior is not unique to thebird. Experiments with many speciesin several laboratories gave rise to thefollowing "circadian" rule: With in-creasing intensities of illumination, thecircadian period is shortened in diurnal(light-active) animals anid lengthenedin nocturnal (dark-active) animals. Re-cently, Hoffmann reviewed the factson which the rule is based (6).
The author is professor of physio1ogy and di-rector at the Max-Planck-Institut fuir Verhaltens-physiologie, 8131 Erling-Andechs, Germany.
1427
on
Aug
ust 5
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

Human Subjects without Time Cues
L....-a 600a °
(o_
4o0
C 800
a. -l
* 400-11;
0-1 3 5 7 9
Tim e (days)
18 24 6 12 16 24 6 12 18
011
')
TI I I I16 24 6 12 16 24
Zone time (hours)6
I l12 18
Fig. 1 (top). Activity rhythms of two chaffinches (Fringilla coelebs). The birds wer3 days in an artificial light-dark cycle, and thereafter in continuous illuminatiorin light with an intensity of 1.8 lux (above), and the other in 120 lux (belo,Period; a activity time; p, rest time; ordinate, perch-hopping activity, recordmeans of print-out counters. Fig. 2 (bottom). Activity rhythm of a chaffinchgilla coelebs) in a light-dark cycle with 12 hours of light and 12 hours of da(LD 12:12) and in continuous illumination (LL) with an intensity of 0.4 lux a120 lux. Black bars, activity time; shaded area, darkness.
1428
The older hypothesis of exogenouscausation of diurnal rhythms wasbased, to some extent, on studies ofhuman subjects. The rhythm of nightworkers was found not to- be shiftedas compared to the rhythm of thoseworking in the daytime; scientists whohad traveled with sailboats along lati-tudes reported that the rhythm oftheir body temperature was always"in phase" with local time. These andother observations suggested an un-known (cosmic) control of the rhythmin man. When the circadian nature ofthe oscillation and its endogenousorigin were demonstrated in animals,my co-workers and I became eager toknow whether man also possesses a
24 circadian clock. In order to investigate24 this properly, it became necessary to
separate single isolated subjects from7' all possible factors which could act as
Zeitgebers. Therefore we had to ex-clude natural daylight, as well as anyother periodic input from the environ-ment which might contain informationon time. For some pilot experiments,a soundproof underground operationroom left over from the wartime wasused at first. Recently we were ableto build our own underground bunkerwith more sophisticated facilities. Herethe subjects live in complete isolationfor up to 3 or 4 weeks. Attached toa rather comfortable bed-sitting roomare a shower and a small kitchen. Theoccupant of the bunker prepares hisown meals. We ask him to lead a"regular" life, that is, to have threemeals in a normal sequence, not tonap after lunch, and to perform afew psychological tests. Otherwise, heis allowed to do what he wishes. Manysubjects are students who cram for anexamination. Listening to recordedmusic is their favored entertainment.
The setup for registration is out-side the bunker. We measure continu-ously the subject's body temperature(by means of a rectal probe), his ac-tivity pattern, his time estimation (of
_ intervals of 20 seconds and of hours),24 and his movements in the bed. The
subject also collects his urine at in-tervals of his own choice. There is a
e kept locked double door at the entrance.i, one In the small room between the twow). r, doors, an icebox serves for a firsted by storage of urine samples. Through therkness same channel we supply the subjectnd Of with fresh food and other necessities,
such as one bottle of Andechs beer
SCIENCE, VOL. 148
NoOL immmma_____im
i-l iill
t8 Lux
-A--.P
120 Lux
"k- i I
I - - . - - - ----
on
Aug
ust 5
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

daily. A magnetic catch permits onlyone of the two doors to be opened ata time. The only way in which thesubject can communicate with the out-side world is by sending and receivingletters. The times at which experi-menters enter the double door fromoutside to deliver supplies and so forthare randomized. Usually we allow thesubjects to turn the light off when theygo to bed and to turn it on when theyget up. The intensity of illumination,however, is controlled from outside.The temperature in the room can beset by the subject himself.The curves in Fig. 3 give an im-
pression of the behavior of a subjectunder those conditions (7). There is aclear cycle of sleep and wakefulnesswhich is reflected also in the rhythmsof body temperature and of urine ex-cretion. The maxima of the four func-tions measured are not always exactly"in phase" with each other and withthe activity cycle, but they all have,on an average, the same free-runningperiod of about 25.0 hours. Since inthis case I myself was the subject, Ican add a few remarks on personalfeelings. After a great curiosity about"true" time during the first 2 daysof bunker life, I lost all interest inthis matter and felt perfectly comfort-able to live "timeless." From theknowledge of animal experiments Iwas convinced that I had a periodshorter than 24 hours; when I wasreleased on day 10, I was thereforehighly surprised to be told that mylast waking-up time was 3 p.m. In the"mornings," I had difficulty in decid-ing whether I had slept long enough.On day 8, I got up after only 3 hoursof sleep (see Fig. 3). Shortly afterbreakfast I wrote in my diary: "Some-thing must be wrong. I feel as if I amon dogwatch." I went to bed againand started the day anew after threemore hours of sleep. Judging from thecurve of body temperature, my first starthappened to coincide with the worstphase of the circadian period, that is,with the low point of temperature. Iwas mistaken by an effort of will andput to order by my physiological clock.The regularity of the circadian
rhythm and its steady drift against lo-cal time become more evident fromFig. 4. Corresponding to the activitytimes of the bird in Fig. 2, the blackbars here represent the times when thesubject is awake, the span being mea-sured from the moment when he turnsthe lights on to the moment when he11 JUNE 1965
q)
200- 100
14Time (days)
Fig. 3. Circadian rhythm of urine excretion (calcium, potassium, and water), bodytemperature, and sleep-wake cycle in a human subject kept without timepiece incomplete isolation from the outside world.
turns them off. Initially, the subjectlives in contact with the outside worldand keeps his normal phase relation-ship. After the beginning of isolation,some irregularities appear; two ex-tremely long activity times (more than20 hours of wakefulness) are followedby a very short one. Thereafter, therhythm becomes remarkably stable,with a mean period of 25.9 hours. Themaxima of urine excretion (obtainedby averaging the hours at which the
0 8 16 24 o
105
40~%10_ ii
maxima of water, potassium, calcium,and sodium occur) follow the sametrend; the mean period differs onlyby 0.3 hour from that of the activitycycle. After 18 physiological days inconfinement, the subject wakes up at8 p.m.; he has "lost" 1.5 real days.Readjustment to the correct phase re-lationship with local time occurs witha "jump," produced by an interposedactivity time of more than 30 hoursduration.
0 6 16 24 8 16 24 8 16 24 8Zone time (hours)
Fig. 4. Circadian rhythm of activity and urine excretion in a human subject keptfor 3 days under normal conditions, then for 18 days in isolation, and finally againunder normal conditions. Black bars, times of being awake; circles, maxima of urineexcretion; T, mean values of period for onset and end of activity and for urine maxima.
1429
on
Aug
ust 5
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
0 6 12 17I I I I I A I I
241 1
6 12l l I
-+ I . . 1w
2.-
4.-
%so
6.-
.-
10.-
12.-
14.-
r' 25(hour.
at)
+ \- \~~~\ 25\ 24.5
\ rkClIt'
?:=24.4(hours)*bodyxtemperatureminimum
O0 6
24.0
I-
7.I 24.1 23.6
excret7i
- 1--'II--I] I --I I I I -I12 16 24 6
40Lux
1.--.9g; :2
200Lux
7
12
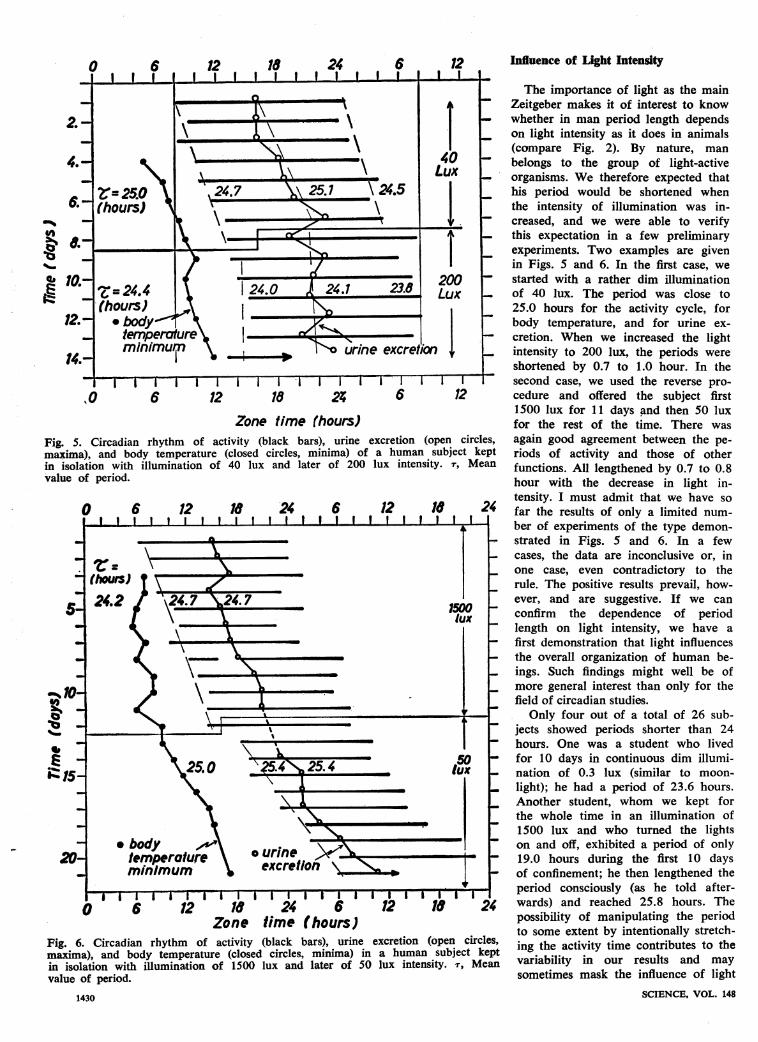
Zone time thours)Fig. 5. Circadian rhythm of activity (black bars), urine excretion (open circlimaxima), and body temperature (closed circles, minima) of a human subject k4in isolation with illumination of 40 lux and later of 200 lux intensity. 7, Mevalue of period.
18 24 6Zone time (hours)
Fig. 6. Circadian rhythm of activity (black bars), urine excretion (open circmaxima), and body temperature (closed circles, minima) in a human subject kin isolation with illumination of 1500 lux and later of 50 lux intensity. r, Mvalue of period.
1430
Influence of Light Intensity
The importance of light as the main- Zeitgeber makes it of interest to know- whether in man period length depends- on light intensity as it does in animals
(compare Fig. 2). By nature, manbelongs to the group of light-active
_ organisms. We therefore expected thathis period would be shortened whenthe intensity of illumination was in-creased, and we were able to verifythis expectation in a few preliminaryexperiments. Two examples are givenin Figs. 5 and 6. In the first case, westarted with a rather dim illuminationof 40 lux. The period was close to25.0 hours for the activity cycle, forbody temperature, and for urine ex-cretion. When we increased the lightintensity to 200 lux, the periods wereshortened by 0.7 to 1.0 hour. In thesecond case, we used the reverse pro-cedure and offered the subject first1500 lux for 11 days and then 50 luxfor the rest of the time. There was
Ces, again good agreement between the pe-ept riods of activity and those of otherban functions. All lengthened by 0.7 to 0.8
hour with the decrease in light in-tensity. I must admit that we have sofar the results of only a limited num-ber of experiments of the type demon-strated in Figs. 5 and 6. In a fewcases, the data are inconclusive or, inone case, even contradictory to therule. The positive results prevail, how-ever, and are suggestive. If we canconfirm the dependence of periodlength on light intensity, we have afirst demonstration that light influencesthe overall organization of human be-ings. Such findings might well be ofmore general interest than only for thefield of circadian studies.
Only four out of a total of 26 sub-jects showed periods shorter than 24hours. One was a student who livedfor 10 days in continuous dim illumi-nation of 0.3 lux (similar to moon-light); he had a period of 23.6 hours.Another student, whom we kept forthe whole time in an illumination of1500 lux and who turned the lightson and off, exhibited a period of only19.0 hours during the first 10 daysof confinement; he then lengthened theperiod consciously (as he told after-
24 wards) and reached 25.8 hours. Thepossibility of manipulating the periodto some extent by intentionally stretch-
Jesp ing the activity time contributes to theePn variability in our results and may
sometimes mask the influence of lightSCIENCE, VOL. 148
I I I
A II -9%-~~~~~~~Nr11
24.7x T71
---
Lie
rlr -,%.
N
urine oAlb.
-- Ii, -I
--t
I
ITF- -"ll .,.
on
Aug
ust 5
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
intensity. There is, on the other side,the surprising fact that 85 percent ofall subjects studied so far had periodsconsiderably longer than 24 hours.This is in agreement with Siffre's rec-ords on his own activity cycle in acave, and with the data published re-cently by Mills (8). The following cir-cumstances might be causes of thelong periods: (i) The rather dim il-lumination, as compared with naturalconditions; in agreement with the cir-cadian rule, this could slow down theoscillator. (ii) The monotony of thesituation, those stimuli which we ex-perience under natural conditions,such as noise and social contacts, beingabsent; experiments with birds showthat grouped animals with mutual stim-ulation have a higher circadian fre-quency than those kept in isolation.(iii) The feedback between the sub-ject's endogenous activity cycle and theself-selected periodic stimuli-that is,turning the lights on and off. The lastexplanation is supported by resultsfrom experiments with birds which wekept in a situation similar to that ofthe bunker. By means of a special ap-paratus, bright light was turned on bythe bird itself shortly after it hadstarted to jump from perch to perch;when the bird went to rest, the brightlight was turned off and only a dimbackground illumination was left. Un-der those conditions, the period ofthe bird's activity cycle was longerthan under constant conditions witheither dim or bright light. More dataare necessary to allow a final decision.
Dissociation and Desynchronization
The adaptive significance of circa-dian rhythmicity is that it enables theorganism to master the changing con-ditions in a temporally programmedworld-that is, to do the right thingat the right time. This could beachieved, to some extent, by anexogenous rhythm. But by developinga self-sustained oscillation of approxi-mately the same frequency as that ofthe environment, the organism, in itsown organization, anticipates the re-spective states which will enable it toreact properly to the environmentalconditions which will ensue-it is pre-pared in advance. Another prerequisite,of course, is the entraining Zeitgeberwhich provides a distinct and species-specific phase relationship between theperiodicity of the organism and that
11 JUNE 1965
0 8 16 24 8 16 2 a8 16 24 0 16 2CI1-
5.-
, 10-
15:-
20-
25-
1I I I I I I II
II I
O 16 24 8 16 24 16 24 8 16 24Zone time (hours)
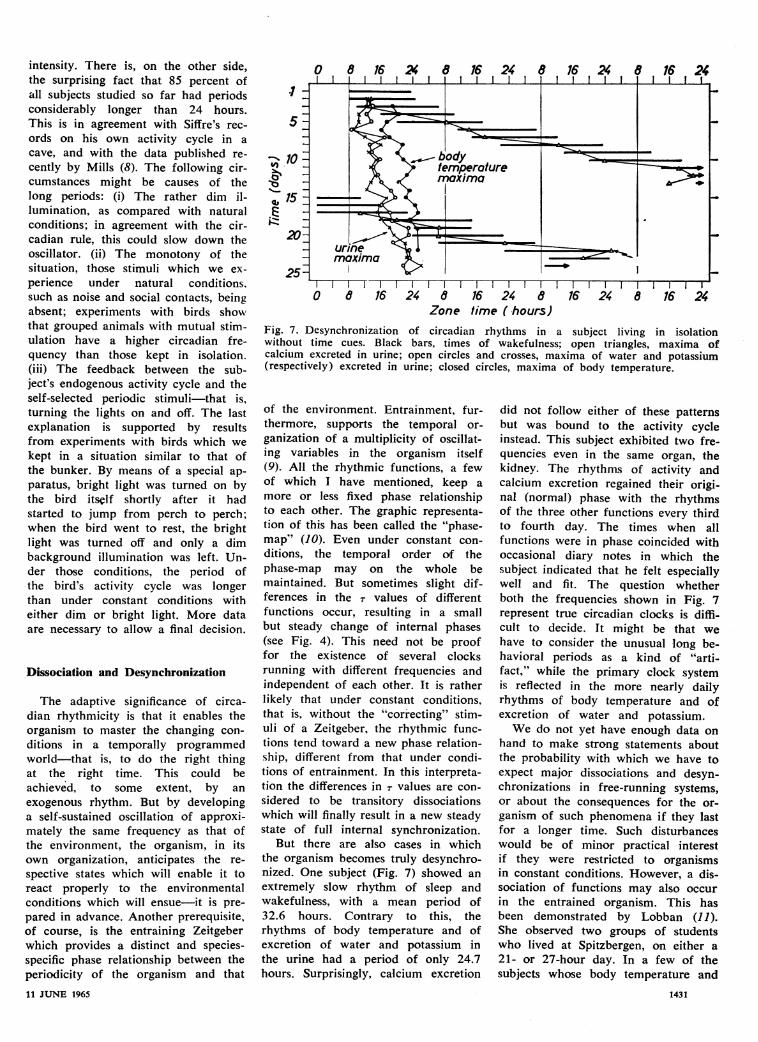
Fig. 7. Desynchronization of circadian rhythms in a subject living in isolationwithout time cues. Black bars, times of wakefulness; open triangles, maxima ofcalcium excreted in urine; open circles and crosses, maxima of water and potassium(respectively) excreted in urine; closed circles, maxima of body temperature.
of the environment. Entrainment, fur-thermore, supports the temporal or-
ganization of a multiplicity of oscillat-ing variables in the organism itself(9). All the rhythmic functions, a fewof which I have mentioned, keep a
more or less fixed phase relationshipto each other. The graphic representa-tion of this has been called the "phase-map" (10). Even under constant con-ditions, the temporal order of thephase-map may on the whole bemaintained. But sometimes slight dif-ferences in the T values of differentfunctions occur, resulting in a smallbut steady change of internal phases(see Fig. 4). This need not be prooffor the existence of several clocksrunning with different frequencies andindependent of each other. It is ratherlikely that under constant conditions,that is, without the "correcting" stim-uli of a Zeitgeber, the rhythmic func-tions tend toward a new phase relation-ship, different from that under condi-tions of entrainment. In this interpreta-tion the differences in T values are con-sidered to be transitory dissociationswhich will finally result in a new steadystate of full internal synchronization.
But there are also cases in whichthe organism becomes truly desynchro-nized. One subject (Fig. 7) showed anextremely slow rhythm of sleep andwakefulness, with a mean period of32.6 hours. Contrary to this, therhythms of body temperature and ofexcretion of water and potassium inthe urine had a period of only 24.7hours. Surprisingly, calcium excretion
did not follow either of these patternsbut was bound to the activity cycleinstead. This subject exhibited two fre-quencies even in the same organ, thekidney. The rhythms of activity andcalcium excretion regained their origi-nal (normal) phase with the rhythmsof the three other functions every thirdto fourth day. The times when allfunctions were in phase coincided withoccasional diary notes in which thesubject indicated that he felt especiallywell and fit. The question whetherboth the frequencies shown in Fig. 7represent true circadian clocks is diffi-cult to decide. It might be that wehave to consider the unusual long be-havioral periods as a kind of "arti-fact," while the primary clock systemis reflected in the more nearly dailyrhythms of body temperature and ofexcretion of water and potassium.We do not yet have enough data on
hand to make strong statements aboutthe probability with which we have toexpect major dissociations and desyn-chronizations in free-running systems,or about the consequences for the or-
ganism of such phenomena if they lastfor a longer time. Such disturbanceswould be of minor practical interestif they were restricted to organismsin constant conditions. However, a dis-sociation of functions may also occurin the entrained organism. This hasbeen demonstrated by Lobban (11).She observed two groups of studentswho lived at Spitzbergen, on either a21- or 27-hour day. In a few of thesubjects whose body temperature and
1431
I I I I I I
A,4 rInxm
url_~epeatrmaxima . II
%F
I
---r. ~ ~~~ ~
on
Aug
ust 5
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

water excretion adapted perfectly tothe new routine, the potassium ex-cretion remained on a 24-hour rhythm.Similar investigations may be of majorinterest for future studies of circadianrhythm, especially with regard to prob-lems in applied physiology.
Applications
Industry makes allowance for thefact that workers have a circadianrhythm of efficiency. Accidents anderrors in tending machines or in read-ing indicator boards are most likelyto happen at 3 a.m. (12). There areonly slight phase differences in theserhythms between night workers andday workers. But night workers arein general less efficient and sufferpsychologically and physiologicallyduring night shifts. They live in asituation of conflict between two tend-encies in the circadian system: to re-main entrained by the social Zeitgebersof the normal environment, and to be-come adjusted to the shifted work-rest cycle. These unnatural conditionsare reflected in a narrowed range(amplitude) of the body temperaturecurve of nightworkers (13). Of course,in conditions without conflicting Zeit-gebers, that is, in more rigorously con-trolled environments of isolation, orafter a trip along latitudes in a jet air-plane (see below), a 180-degree shiftof the circadian rhythm will be accom-plished in about 6 days (14). It is stillan open question whether night shiftsshould last long to make use of someadaptation which may occur, or wheth-er frequent changes between night andday shifts, requiring repeated readjust-ments, are preferable.The situation becomes even more
complicated with a work-rest scheduleof 4 hours on duty and 4 hours off,as may be used for a crew in a space-craft. Under those conditions, thecircadian system could become synch-ronized in a 1-to-3 ratio with the
8-hour period of the artificial Zeit-geber. The few groups of subjectsstudied on those and similar schedulesshowed a clear circadian rhythm.There was, however, a continuousdrift in phase of the human rhythmagainst clock time. The experimentsconducted so far did not last longenough to determine whether the cir-cadian system was free running, orentrained with a tendency toward anew phase relationship to the Zeit-geber (15). The answer would be ofimportance for further decisions onwork-rest schedules and other livingconditions for crews.A common experience in the jet
century may serve as a third example.If we travel by air from Paris to NewYork, our circadian clock keeps ongoing with European time. Arriving atKennedy Airport, we are out of phaseby about 6 hours. Entrainment to lo-cal time takes 2 to 3 days, dur-ing which we feel tired and less effi-cient (16). As we could demonstratein birds, the time necessary for read-justment is not the same when weshorten the light-dark cycle once by 6hours as when we lengthen it by thesame amount. Applying these results toman, we have to expect different dura-tions of indisposition after long-distanceflights eastward and westward. Obvi-ously, it is of interest for civil purposesas well as for military use to know howwe can shorten the time required forresynchronization. One way seems tobe to influence the natural frequencyof the circadian oscillator; this fre-quency determines to some extentthe speed with which the systemregains its natural phase relationshipto the environment, at least in birds.Manipulation of artificial Zeitgebers-for example, the insertion of relativelyshort times of light and dark after ex-treme long flights-might be anotherpossibility. The discussion of theseproblems has only reached a state oftheoretical analysis and of preliminaryexperimentation with animals.
All three examples mentioned aboveshow that we still lack much necessaryinformation, but they indicate that theself-sustained circadian oscillator hasto be taken into account. Its mainproperties seem to be the same inhuman beings as in all other orga-nisms. We have to study them beforewe can discuss practical problemssuccessfully. As always in science, abetter understanding of the basic phe-nomena will be the first step towarda proper application in practice.
References and Notes
1. J. Aschoff, Klin. Wochschr. 33, 545 (1955);A. Gierse, "Quoniam sit ratio caloris or-ganici" (dissertation, Halle, 1842).
2. E. Bunning, The Physiological Clock (Spring-er, Heidelberg, 1964).
3. C. S. Pittendrigh and V. G. Bruce, inRhythmic and Syntthetic Processes in Growth,D. Rudnick, Ed. (Princeton Univ. Press,Princeton, N.J., 1957), p. 75.
4. J. Aschoff, Ann. Rev. Physiol. 25, 581 (1963);J. Enright, in Circadian Clocks, J. Aschoff,Ed. (North-Holland, Amsterdam, in press);A. Heusner, ibid.; K. Klotter, ibid.
5. F. Halberg, E. Halberg, C. P. Barnum, J. J.Bittner, in Photoperiodism and Related Phe-nomena in Plants and Animals, R. B. With-row, Ed. (AAAS, Washington, D.C., 1959),p. 803.
6. K. Hoffmann, in Ctrcadian Clocks, J.Aschoff, Ed. (North-Holland, Amsterdam, inpress).
7. J. Aschoff and R. Wever, Naturwissenschaften49, 337 (1962).
8. M. Siffre, Hors du Temps (Tuillard, Paris,1963); J. N. Mills, J. Physiol. 174, 217 (1964).
9. C. S. Pittendrigh, in The Harvey Lectures,1960-1961, Series 56 (Academic Press, NewYork, 1962), p. 93; J. Aschoff, in Biology ofSurvival, 0. G. Edholm, Ed. (Zoological So-ciety of London Symposium No. 13, 1964),p. 79.
10. J. Aschoff, Deut. Med. Wochschr. 88, 1930(1963); F. Halberg, Cold Spring Harbor Symp.Quant. Biol. 25, 289 (1960).
11. M. Lobban, Cold Sprinig Harbor Symp. Quant.Biol. 25, 325 (1960).
12. W. Menzel, Menschliche Tag-Nacht-Rhythmikund Schichtarbeit (Schwabe, Basel, 1962).
13. J. H. van Loon, Economics 6, 267 (1963).14. N. Kleitman, Sleep and Wakefulness (Univ.
of Chicago Press, Chicago, 1963); J. Aschoff,Naturwissenschaftent 42, 569 (1955).
15. E. A. Alluisi, T. J. Hall, G. R. Hawkes, W.D. Chiles, Final Report, Contract No. AF 33(616)-7607-M4 (1962); J. Aschoff, in "Pro-ceedings, Third International Symposium onBioastronautics and the Exploration of Space,"San Antonio, 1964 (in press).
16. H. Strughold, J. Aviation Med. 23, 464 (1952);G. T. Hauty, in Circadian Clocks, J.Aschoff, Ed. (North-Holland, Amsterdam, inpress).
17. Part of the experiments reported here weresupported by NASA research grant NsG-259-62. I thank Dr. Klaus Hoffmann for hisassistance with the translation.
SCIENCE, VOL. 1481432
on
Aug
ust 5
, 201
1w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from


















