
Choline Acetyltransferase Uses a Non-AUG Initiation Codon and ...
7
THE JOURNAL OF BIOLOGICAL. CHEMISTRY 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 265, No. 35, Issue of December 15, PP. 21714~21719,199O Printed in U.S. A. DrosophiZa Choline Acetyltransferase Uses a Non-AUG Initiation Codon and Full Length RNA Is Inefficiently Translated* (Received for publication, June 4, 1990) Hidemitsu Sugiharas, Veronica Andrisani, and Paul M. Salvaterrae From the Division of Neurosciences, Beckman Research Institute of the City of Hope, Duarte, California 91010 RNA from a partial cDNA clone containing the entire protein coding sequence of Drosophila melanogaster acetyl-CoA:choline 0-acetyltransferase (EC 2.3.1.6; choline acetyltransferase) can be translated into active enzyme. This is unusual since this partial cDNA clone contains no appropriate ATG (AUG) initiation codon. In this study we use in vitro deletion and point mutants to identify GTG as the starting codon for protein trans- lation. We also report the sequence of a full length Drosophila choline acetyltransferase cDNA and dem- onstrate that RNA produced by this clone is translated into active choline acetyltransferase but at a signifi- cantly reduced efficiency when compared to the partial cDNA clone. These results indicate that translational control may be an important regulatory step in pro- duction of Drosophila choline acetyltransferase. Individual neurons use only a small subset of potential neurotransmitters by regulating the expression of genes cod- ing the appropriate neurotransmitter biosynthetic enzymes. The enzyme required for biosynthesis of the important central and peripheral neurotransmitter acetylcholine is choline ace- tyltransferase (acetyl-CoA:choline 0-acetyltransferase, EC 2.3.1.6). We have previously reported the complete purifica- tion (Slemmon et al., 1982), molecular characterization, (Slemmon et QL, 1984), and partial cDNA cloning of choline acetyltransferase from Drosophila (Itoh et al., 1986) and are interested in the molecular logic involved in regulation of this gene. Immunocytochemical (Buchner et al., 1986; Gorzyca and Hall, 1987; Ikeda and Salvaterra, 1989), as well as in situ hybridization (Barber et al., 1988) studies in Drosophila indi- cate that choline acetyltransferase protein and mRNA are expressed in only certain neurons thought to use acetylcholine as a neurotransmitter. A major feature of choline acetyltrans- ferase regulation is thus likely to be the spatial and temporal control of transcription. In addition to transcriptional control, regulation of expres- sion can involve RNA processing, RNA turnover, or transla- tional control. Drosophila choline acetyltransferase mRNA is * This work was supported by a grant from the National Institutes of Health, National Institute of Neurological and Communicative Disorders and Stroke. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solelv to indicate this fact. The nucleotide sequence(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) JO5 706. $ Present address: Nippon Kayaku Co., Ltd., 31-12 Shimo 3- Chome, Kita-Ku, Tokyo 115, Japan. § To whom correspondence should be addressed: Div. of Neurosci- ences. Beckman Research Institute of the Citv of HoDe, 1450 E. DuaAe Rd., Duarte, CA 91010. more than double the size required to code for a choline acetyltransferase polypeptide with a molecular mass of 75 kilodaltons (Itoh et al., 1986; Muiioz-Maines et al., 1988). The size of our original partial Drosophila choline acetyltransfer- ase cDNA clone is 2.4 kilobases and contains a long open reading frame which could theoretically code for a protein of approximately 81 kilodaltons. This clone lacks any in frame ATG (methionine) initiation codon upstream from the known protein coding regions. The first methionine codon is con- tained internally in an amino-terminal tryptic peptide deter- mined by direct amino acid sequencing and can thus not be the site of initiation in uivo. Surprisingly, in vitro synthesized RNA using this partial cDNA clone as a template can be efficiently translated into active choline acetyltransferase when injected into Xenopus oocytes (McCaman et al., 1988). The major protein product produced by the oocytes has a molecular size of 75 kilodaltons determined by Western blot- ting and appears indistinguishable from native Drosophila choline acetyltransferase. This protein is much larger than that which would be produced if the first in frame ATG is used as the site of protein translation initiation. Moreover, the same cDNA inserted into the pKK-223-3 plasmid expres- sion vector can produce active enzyme in transfected Esche- richia coli (Sugihara et al., 1988) which has an isoelectric point identical to native Drosophila choline acetyltransferase. Thus both a eucaryotic and a procaryotic translation system have the ability to properly translate Drosophila choline acetyl- transferase RNA into active enzyme. These unexpected results suggest that Drosophila choline acetyltransferase initiates protein translation at a non-ATG codon. While nearly all genes to date appear to contain ATG as the site of protein translation initiation (Kozak, 1987) it is known from in vitro studies that this is not an absolute requirement (Peabody, 1987). Several precedents exist for initiation of protein translation at a GTG codon in bacteria (Gold, 1988; Kozak, 1983) where this unusual feature is thought to mediate regulation of protein production in a polycystronic mRNA coding for two different enzymes in an amino acid biosynthetic pathway (Kozak, 1986b). Recently, precedents have also been established for natural eucaryotic genes which initiate from non-ATG codons (Hann et al., 1988; Florkiewicz and Sommer, 1989; Acland et al., 1990). An inspection of our partial cDNA sequence identified several potential in frame GTG codons which could serve as protein translation initiation sites. In this paper we present evidence for one particular GTG codon at position 33 in the original cDNA sequence as the most likely site of translation initiation. We have constructed several deletion and point mutations and tested the ability of the resulting in vitro synthesized RNA to direct the synthesis of active choline acetyltransferase of appropriate size following translation by rabbit reticulocyte lysates. We also report the sequence of a full length Drosophila choline acetyltransferase cDNA ex- 21714 by guest on February 12, 2018 http://www.jbc.org/ Downloaded from
Transcript of Choline Acetyltransferase Uses a Non-AUG Initiation Codon and ...

THE JOURNAL OF BIOLOGICAL. CHEMISTRY 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 35, Issue of December 15, PP. 21714~21719,199O Printed in U.S. A.
DrosophiZa Choline Acetyltransferase Uses a Non-AUG Initiation Codon and Full Length RNA Is Inefficiently Translated*
(Received for publication, June 4, 1990)
Hidemitsu Sugiharas, Veronica Andrisani, and Paul M. Salvaterrae From the Division of Neurosciences, Beckman Research Institute of the City of Hope, Duarte, California 91010
RNA from a partial cDNA clone containing the entire protein coding sequence of Drosophila melanogaster acetyl-CoA:choline 0-acetyltransferase (EC 2.3.1.6; choline acetyltransferase) can be translated into active enzyme. This is unusual since this partial cDNA clone contains no appropriate ATG (AUG) initiation codon. In this study we use in vitro deletion and point mutants to identify GTG as the starting codon for protein trans- lation. We also report the sequence of a full length Drosophila choline acetyltransferase cDNA and dem- onstrate that RNA produced by this clone is translated into active choline acetyltransferase but at a signifi- cantly reduced efficiency when compared to the partial cDNA clone. These results indicate that translational control may be an important regulatory step in pro- duction of Drosophila choline acetyltransferase.
Individual neurons use only a small subset of potential neurotransmitters by regulating the expression of genes cod- ing the appropriate neurotransmitter biosynthetic enzymes. The enzyme required for biosynthesis of the important central and peripheral neurotransmitter acetylcholine is choline ace- tyltransferase (acetyl-CoA:choline 0-acetyltransferase, EC 2.3.1.6). We have previously reported the complete purifica- tion (Slemmon et al., 1982), molecular characterization, (Slemmon et QL, 1984), and partial cDNA cloning of choline acetyltransferase from Drosophila (Itoh et al., 1986) and are interested in the molecular logic involved in regulation of this gene. Immunocytochemical (Buchner et al., 1986; Gorzyca and Hall, 1987; Ikeda and Salvaterra, 1989), as well as in situ hybridization (Barber et al., 1988) studies in Drosophila indi- cate that choline acetyltransferase protein and mRNA are expressed in only certain neurons thought to use acetylcholine as a neurotransmitter. A major feature of choline acetyltrans- ferase regulation is thus likely to be the spatial and temporal control of transcription.
In addition to transcriptional control, regulation of expres- sion can involve RNA processing, RNA turnover, or transla- tional control. Drosophila choline acetyltransferase mRNA is
* This work was supported by a grant from the National Institutes of Health, National Institute of Neurological and Communicative Disorders and Stroke. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solelv to indicate this fact.
The nucleotide sequence(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) JO5 706.
$ Present address: Nippon Kayaku Co., Ltd., 31-12 Shimo 3- Chome, Kita-Ku, Tokyo 115, Japan.
§ To whom correspondence should be addressed: Div. of Neurosci- ences. Beckman Research Institute of the Citv of HoDe, 1450 E. DuaAe Rd., Duarte, CA 91010.
more than double the size required to code for a choline acetyltransferase polypeptide with a molecular mass of 75 kilodaltons (Itoh et al., 1986; Muiioz-Maines et al., 1988). The size of our original partial Drosophila choline acetyltransfer- ase cDNA clone is 2.4 kilobases and contains a long open reading frame which could theoretically code for a protein of approximately 81 kilodaltons. This clone lacks any in frame ATG (methionine) initiation codon upstream from the known protein coding regions. The first methionine codon is con- tained internally in an amino-terminal tryptic peptide deter- mined by direct amino acid sequencing and can thus not be the site of initiation in uivo. Surprisingly, in vitro synthesized RNA using this partial cDNA clone as a template can be efficiently translated into active choline acetyltransferase when injected into Xenopus oocytes (McCaman et al., 1988). The major protein product produced by the oocytes has a molecular size of 75 kilodaltons determined by Western blot- ting and appears indistinguishable from native Drosophila choline acetyltransferase. This protein is much larger than that which would be produced if the first in frame ATG is used as the site of protein translation initiation. Moreover, the same cDNA inserted into the pKK-223-3 plasmid expres- sion vector can produce active enzyme in transfected Esche- richia coli (Sugihara et al., 1988) which has an isoelectric point identical to native Drosophila choline acetyltransferase. Thus both a eucaryotic and a procaryotic translation system have the ability to properly translate Drosophila choline acetyl- transferase RNA into active enzyme.
These unexpected results suggest that Drosophila choline acetyltransferase initiates protein translation at a non-ATG codon. While nearly all genes to date appear to contain ATG as the site of protein translation initiation (Kozak, 1987) it is known from in vitro studies that this is not an absolute requirement (Peabody, 1987). Several precedents exist for initiation of protein translation at a GTG codon in bacteria (Gold, 1988; Kozak, 1983) where this unusual feature is thought to mediate regulation of protein production in a polycystronic mRNA coding for two different enzymes in an amino acid biosynthetic pathway (Kozak, 1986b). Recently, precedents have also been established for natural eucaryotic genes which initiate from non-ATG codons (Hann et al., 1988; Florkiewicz and Sommer, 1989; Acland et al., 1990).
An inspection of our partial cDNA sequence identified several potential in frame GTG codons which could serve as protein translation initiation sites. In this paper we present evidence for one particular GTG codon at position 33 in the original cDNA sequence as the most likely site of translation initiation. We have constructed several deletion and point mutations and tested the ability of the resulting in vitro synthesized RNA to direct the synthesis of active choline acetyltransferase of appropriate size following translation by rabbit reticulocyte lysates. We also report the sequence of a full length Drosophila choline acetyltransferase cDNA ex-
21714
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Choline Acetyltransferase Translation
tending our original clone in both the 5’ and 3’ directions. In vitro synthesized RNA from this full length clone is translated into the same size protein as the partial cDNA clone but at a significantly reduced efficiency. Taken together, our results indicate that translational control may play an important regulatory role in production of active choline acetyltransfer- 898.
EXPERIMENTAL PROCEDURES’
General Methods-Details of the full length cDNA isolation (pCha- JV), in vitro RNA synthesis, in vitro translation, choline acetyltrans- ferase enzyme assay, and SDS-gel electrophoresis are provided in the Miniprint section of this paper.
In Vitro Mutagenesis-Both deletion mutants and point mutants were constructed to identify the initiation codon used by the partial choline acetyltransferase cDNA (pCha-12). The deletion mutants were designed to cause a shift in reading frame if translation initiation occurs upstream from the deletion site, while initiation downstream should have little or no effect on translation. The unique ApaI site at position 131 or the XhoI site at position 379 on the pCha-12 plasmid was cut with the appropriate restriction endonuclease and the line- arized plasmids were digested with mung bean nuclease to remove overhanging DNA. The plasmids were then religated with T4 DNA ligase and the sequence of the resulting mutants was determined using the Sequenase kit (United States Biochemical Corp.). The SP6 sequencing primer (Promega) was used as a primer. Fig. 1 shows the diagramatic position of each deletion mutant along with the con- firmed sequence.
Oligonucleotide directed point mutations were constructed from an EcoRI I-PstI-digested fragment (857 base pairs) from pCha-12 which was first subcloned into M13mp18 phage. Single bases were changed using the Mutagene Ml3 kit from Bio-Rad. Two oligonucle- otides were purchased from Research Genetics: 5’-CGTTGGAUG- CCGCGTTCGCT-3’ and 5’-CGTTGGAUGCCATGTTCGCTC-3’. The resulting mutants (see Fig. 1) had the normal GTG triplet starting at position 33 of pCha-12 changed to GCG or ATG, respec- tively. The nucleotide sequences of point mutants were confirmed using Sequenase and the Ml3 universal primer. The confirmed mu- tated sequences were removed by digestion with EcoRI and XhoI and replaced in pCha-12 after removal of the EcoRI-XhoI fragment.
RESULTS
Deletion Mutations-Inspection of the 5’ sequence of pCha- JV (see Fig. 1 in the Miniprint section) or pCha-12 reveals a number of potential protein translation initiation sites. The positions of these sites are indicated in Fig. 1 for pCha-12. Obviously, the first in frame ATG codon at position 329 in this partial cDNA clone cannot be the initiation codon for native Drosophila choline acetyltransferase since it is posi- tioned internally in a sequenced tryptic peptide (Itoh et al., 1986). It is also unlikely to be used during in uitro translation since it would result in a protein product of only 64 kilodaltons while the major active protein produced by Xenopus oocytes had a size of 75 kilodaltons (McCaman et al., 1988). The initiation codon should therefore be positioned somewhere between position 1 and 329 and most likely in the 5’ part of this region to result in an active protein with the same molecular size and p1 as native choline acetyltransferase.
The polypeptides resulting from rabbit reticulocyte lysate in vitro translation of the RNA from pCha-12 and the two deletion mutants are shown in Fig. 2. The major 3”S-labeled protein translated from the pCha-12 RNA has a molecular mass of 75 kilodaltons. This polypeptide size is identical to native Drosophila choline acetyltransferase (Muiioz-Maines
’ Portions of this paper (including part of “Experimental Proce- dures” and Fig. 1) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press.
’ The abbreviation used is: SDS, sodium dodecyl sulfate.
FIG. 1. Partial sequence of the 5’ end of pCha-12 cDNA. The position of the proposed GTG start codon is indicated by the solid black box. We are adopting the convention of using T rather than U since all sequences were determined from cDNA. Underneath this codon the base changes for the two point mutations are shown with a solid black bon indicating the G”” to A transition (ATG mutant) and the T:j4 to C transition (GCG mutant). The positions of the frame shift deletion mutants are shown by the shaded boxes enclosing the respective restriction sites (ApaI and XhoI) along with the actual bases removed by mung bean nuclease digestion. The position of the first in frame ATG codon is boxed and the 3 out of frame ATG codons are indicated by underlining with three small boxes. All mu- tations were confirmed by dideoxy sequencing. The original number- ing was taken from Itoh et al. (1986) starting at the first 5’ base in the Drosophila choline acetyltransferase cDNA clone.
200-
113-
67-
45-
dlud-I-- 12 3 4
FIG. 2. Polypeptides produced by rabbit reticulocyte lysate translation of in vivo synthesized RNA from pCha-12 and the ApaI and XhoI deletion mutants. The figure is an autoradiograph of [““Slmethionine labeled proteins produced by in vitro translation of approximately 1 rg of RNA according to the manufacturer’s protocol (Stratagene). Polypeptides were separated on a 7.5% SDS- polyacrylamide gel electrophoresis gel as described in the Miniprint section. The position of molecular weight markers electrophoresed on the same gel and stained with Coomassie Blue is indicated on the left hand side of the figure (myosin, E. coli fi-galactosidase, bovine serum albumin, and hen egg white ovalbumen). Lane 1, pCha-12 RNA; lane 2, ApaI deletion mutant; lane 3, XhoI deletion mutant; lane 4, no added RNA. All translation mixtures except lane 4 had the same amount of input RNA as determined by quantitative video densitometry of a slot blot as described in Miniprint section (data not shown). When RNA from pCha 8 in the antisense orientation was tested, no polypeptides or choline acetyltransferase activity was detected (data not shown). See Fig. 1 for details of the deletion mutants.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
21716 Choline Acetyltransferase Translation
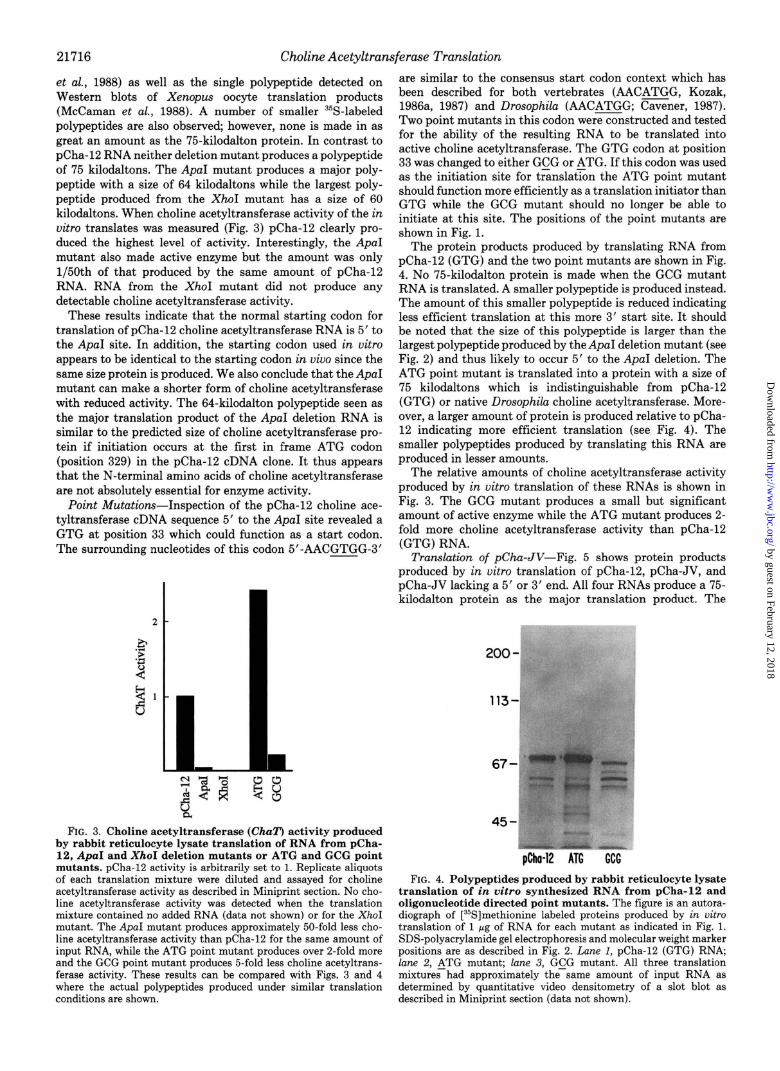
et al., 1988) as well as the single polypeptide detected on Western blots of Xenopus oocyte translation products (McCaman et al., 1988). A number of smaller 35S-labeled polypeptides are also observed, however, none is made in as great an amount as the 75-kilodalton protein. In contrast to pCha-12 RNA neither deletion mutant produces a polypeptide of 75 kilodaltons. The ApaI mutant produces a major poly- peptide with a size of 64 kilodaltons while the largest poly- peptide produced from the XhoI mutant has a size of 60 kilodaltons. When choline acetyltransferase activity of the in vitro translates was measured (Fig. 3) pCha-12 clearly pro- duced the highest level of activity. Interestingly, the ApaI mutant also made active enzyme but the amount was only 1/50th of that produced by the same amount of pCha-12 RNA. RNA from the XhoI mutant did not produce any detectable choline acetyltransferase activity.
These results indicate that the normal starting codon for translation of pCha-12 choline acetyltransferase RNA is 5’ to the ApaI site. In addition, the starting codon used in vitro appears to be identical to the starting codon in vivo since the same size protein is produced. We also conclude that the ApaI mutant can make a shorter form of choline acetyltransferase with reduced activity. The 64-kilodalton polypeptide seen as the major translation product of the ApaI deletion RNA is similar to the predicted size of choline acetyltransferase pro- tein if initiation occurs at the first in frame ATG codon (position 329) in the pCha-12 cDNA clone. It thus appears that the N-terminal amino acids of choline acetyltransferase are not absolutely essential for enzyme activity.
Point Mutations-Inspection of the pCha-12 choline ace- tyltransferase cDNA sequence 5’ to the ApaI site revealed a GTG at position 33 which could function as a start codon. The surrounding nucleotides of this codon 5’-AACGTGG-3’
2 -
.@ 2 9
FIG. 3. Choline acetyltransferase (ChaT) activity produced by rabbit reticulocyte lysate translation of RNA from pCha- 12, ApaI and XhoI deletion mutants or ATG and GCG point mutants. pCha-12 activity is arbitrarily set to 1. Replicate aliquots of each translation mixture were diluted and assayed for choline acetyltransferase activity as described in Miniprint section. No cho- line acetyltransferase activity was detected when the translation mixture contained no added RNA (data not shown) or for the XhoI mutant. The ApaI mutant produces approximately 50-fold less cho- line acetyltransferase activity than pCha-12 for the same amount of input RNA, while the ATG point mutant produces over 2-fold more and the GCG point mutant produces 5-fold less choline acetyltrans- ferase activity. These results can be compared with Figs. 3 and 4 where the actual polypeptides produced under similar translation conditions are shown.
are similar to the consensus start codon context which has been described for both vertebrates (AACATGG, Kozak, 1986a, 1987) and Drosophila (AACATGG; Cavener, 1987). Two point mutants in this codon were constructed and tested for the ability of the resulting RNA to be translated into active choline acetyltransferase. The GTG codon at position 33 was changed to either GCG or ATG. If this codon was used as the initiation site for t&slat%n the ATG point mutant should function more efficiently as a translation initiator than GTG while the GCG mutant should no longer be able to initiate at this site. The positions of the point mutants are shown in Fig. 1.
The protein products produced by translating RNA from pCha-12 (GTG) and the two point mutants are shown in Fig. 4. No 75-kilodalton protein is made when the GCG mutant RNA is translated. A smaller polypeptide is produced instead. The amount of this smaller polypeptide is reduced indicating less efficient translation at this more 3’ start site. It should be noted that the size of this polypeptide is larger than the largest polypeptide produced by the ApaI deletion mutant (see Fig. 2) and thus likely to occur 5’ to the ApaI deletion. The ATG point mutant is translated into a protein with a size of 75 kilodaltons which is indistinguishable from pCha-12 (GTG) or native Drosophila choline acetyltransferase. More- over, a larger amount of protein is produced relative to pCha- 12 indicating more efficient translation (see Fig. 4). The smaller polypeptides produced by translating this RNA are produced in lesser amounts.
The relative amounts of choline acetyltransferase activity produced by in uitro translation of these RNAs is shown in Fig. 3. The GCG mutant produces a small but significant amount of active enzyme while the ATG mutant produces 2- fold more choline acetyltransferase activity than pCha-12 (GTG) RNA.
Translation of pChn-JV-Fig. 5 shows protein products produced by in vitro translation of pCha-12, pCha-JV, and pCha-JV lacking a 5’ or 3’ end. All four RNAs produce a 75- kilodalton protein as the major translation product. The
200-
FIG. 4. Polypeptides produced by rabbit reticulocyte lysate translation of in vitro synthesized RNA from pCha-12 and oligonucleotide directed point mutants. The figure is an autora- diograph of [%]methionine labeled proteins produced by in vitro translation of 1 Fg of RNA for each mutant as indicated in Fig. 1. SDS-polyacrylamide gel electrophoresis and molecular weight marker positions are as described in Fig. 2. Lane I, pCha-12 (GTG) RNA; lane 2, ATG mutant; lane 3, GCG mutant. All three translation mixtures had approximately the same amount of input RNA as determined by quantitative video densitometry of a slot blot as described in Miniprint section (data not shown).
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Choline Acetyltransferase Translation 21717
200-
113- :v: $
67-, 'I. :* -
t ,.i
"
45-' ., .*,_.
12 3 4
FIG. 5. Polypeptides produced by rabbit reticulocyte trans- lation of in vitro synthesized RNA from pCha-12, pCha-JV, pCha-JV-3’, and pCha-JV-5’. The figure is an autoradiograph of [“SS]methionine labeled proteins produced by in vitro translation of approximately 1 pg of RNA for each lane. All four RNAs produce a major protein at 75 kilodaltons. pCha-12 (lane 1) is translated about 20-fold more efficiently than pCha-JV (lane 2). pCha-JV-5’ (lane 3), and pCha-JV-3’ (lane 4) have 386 or 744 nucleotides removed from pCha-JV at the 5’ or 3’ end and show slightly better translation than pCha-JV. The sizes of the RNAs were confirmed by agarose gel electrophoresis and the amounts used for in uitro translation were determined by slot blot hybridization and video densitometry (data not shown) (see Fig. 2 for further details).
1.0 -
.2
.z
4
3 0.5 -
FIG. 6. Choline acetyltransferase (ChaZ’) activity produced by rabbit reticulocyte lysate translation of RNA from pCha- 12, pCha-JV, pCha-JV-3’, and pCha-JV-5’. pCha-JV RNA is translated into only l/20 the choline acetyltransferase activity of pCha-12. Removal of the 5’ end of pCha-JV results in a slight increase in choline acetyltransferase activity relative to pCha-JV while re- moval of the 3’ end results in either no change or a slight decrease as shown in the figure (see Miniprint section and Fig. 5 for further details).
translational efficiency is quite different, however. pCha-12 is translated most efficiently while pCha-JV is translated into 20-50-fold less 75kilodalton protein in a number of different experiments. When the 5’ end of pCha-JV is removed, a small increase in translational efficiency was observed while re- moval of the 3’ end of pCha-JV had little or no effect on translational efficiency.
The relative choline acetyltransferase activity produced by these four clones is shown in Fig. 6 and agrees well with the relative amounts of 75kDa protein product. These results indicate that a full length choline acetyltransferase RNA is less efficiently translated than the smaller size pCha-12 RNA.
DISCUSSION
Considerable experimental evidence and theoretical consid- erations have led to the conclusion that most eucaryotic
mRNAs initiate protein translation at the 5’ most ATG codon. Kozak (1986a, 1987) has proposed the widely accepted scanning model to account for the nonrandom initiation sites. Her extensive literature surveys of higher eucaryotic mRNAs (Kozak, 1987) along with a number of experimental studies (Curran and Yarus, 1987; Kozak, 1986a) have established not only the importance of an ATG initiator codon, but also the position of this codon in the proper context for translation initiation. Cavener (1987) has recently extended the analysis to include Drosophila mRNAs and reached similar conclusions about the importance of the ATG initiation codon as well as its context. It was thus a surprise when we observed that our partial Drosophila choline acetyltransferase cDNA clone could produce RNA which in turn could be translated into active enzyme by a number of experimental translation systems (McCaman et al., 1988), even though it lacked an in-frame ATG codon which was downstream to the known protein coding regions determined by direct amino acid sequencing of the purified enzyme (Itoh et al., 1986; Slemmon et al., 1982). Several possibilities could have accounted for translation of RNA into active choline acetyltransferase.
The RNA could have produced a protein with a truncated amino terminus, by initiating translation at the first in frame ATG codon, which retained some choline acetyltransferase activity. This possibility is unlikely since the present study, documenting rabbit reticulocyte lysate translation, as well as our previous study using Xenopus oocytes (McCaman et al., 1988) yielded a major protein product which is the same size as native Drosophila choline acetyltransferase. The largest polypeptide which could be produced from translation initi- ating at the first internal ATG codon at position 329 in the cDNA clone would be 64 kilodaltons which is considerably shorter than the 75-kilodalton product we have observed. Nevertheless, it is apparent from the present study that smaller polypeptides can be produced by reticulocyte lysate translation which retain some choline acetyltransferase activ- ity. The ApaI deletion mutant as well as the GCG point mutant (see Fig. 3) produce a small amount of active enzyme and no 75-kilodalton protein (see Figs. 3 and 4).
It is also possible that a reading frame shift could account for the in vitro translation of choline acetyltransferase RNA, as has been described for some bacterial systems (Craigen and Caskey, 1987). This possibility is unlikely for choline acetyl- transferase RNA since the two most 5’ out of frame ATG triplets in our clone are located downstream from the ApaI deletion and are not in a favorable context for translation initiation (see Fig. 1). We can also rule out sequencing errors or cloning artifacts which could be proposed to account for our results since we have repeatedly checked the sequence of the 5’ end of our clone and are confident that the sequence is accurate. In addition, we have isolated independent clones from two other cDNA libraries (one from the Oregon R strain), prepared independently of our original library, as well as other clones from our own cDNA library and two genomic libraries. The sequence in the 5’ region of all these clones was identical.
It appears likely therefore that Drosophila choline acetyl- transferase uses a non-ATG as the initiator codon. The stud- ies we have reported here indicate that the GTG starting at position 33 in pCha-12 is the most likely candidate for this function. The two frame shift deletion mutants we have constructed indicate that the initiator codon is 5’ to the ApaI site at position 131 in pCha-12. The most appealing codon in this region of the clone is the GTG at position 33 for two reasons: it is in frame with the known protein coding regions of the rest of the clone and it is in a near perfect context for
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

21718 Choline Acetyltransferase Translation
Drosophila initiation (Cavener, 1987). We have tested this hypothesis by constructing two point mutations in this codon and the results of in vitro translation indicate that this is the correct initiation site. Changing this codon to ATG results in more efficient translation of the mutant RNA as judged by increased choline acetyltransferase activity and the 75 kilo- dalton protein relative to the normal codon. A corresponding change of this codon to GCG results in a significant reduction of choline acetyltransferase activity and the complete absence of the 75 kilodalton protein. A smaller protein is made by this mutant RNA which also produces some choline acetyltrans- ferase activity.
Naturally occurring non-ATG initiation codons are rare, but a few precedents have been described. Several procaryotic genes use an upstream GTG codon as well as a more 3’ ATG codon initiate synthesis of two different proteins from a polycystronic mRNA (Kozak, 1986b). The presence of the less efficient GTG initiator codon is thought to have metabolic regulatory significance for production of appropriate amounts of each enzyme from the polycystronic mRNA. Examples of naturally occurring higher eucaryotic non-ATG initiator co- dons are rare but a few have recently been described. The c- myc protein, thought to be involved in oncogenesis (Hann et al., 1988) results from an RNA which is capable of producing two proteins with different amino-terminal amino acids de- pending on whether initiation occurs at a CTG in exon 1 or an ATG in exon 2. Interestingly, initiation of c-myc protein translation at the non-ATG site was shown to be disrupted in Burkitt’s lymphomas. Human fibroblast growth factor mRNA can produce four polypeptides from a single RNA, three of which initiate at CTG codons (Florkiewicz and Som- mer, 1989). The int-2 gene, related to fibroblast growth factor, has also been shown to initiate partially at a CTG codon and the protein product is produced as either a nuclear or secretory form depending on the translation initiation site (Acland et al,, 1990). Several viral RNAs have also been shown to be capable of protein translation initiation at non-ATG codons (i.e. ACG) (Becerra et al., 1985; Gupta and Patwardhan, 1988). In addition to the rare examples of non-ATG initiation codons a number of in vitro mutagenesis studies have described the base sequences which can function both in vitro and in viva as initiator codons (Peabody, 1987,1989; Zitoner et al., 1984).
It is noteworthy that all cases of naturally occurring non- ATG initiation are thought to have profound regulatory con- sequences. In this regard it may be important to ask what the significance is for Drosophila choline acetyltransferase to incorporate this unusual feature. Choline acetyltransferase functions by producing the neurotransmitter acetylcholine used for synaptic transmission. In Drosophila, the enzyme protein appears to be concentrated in neutrophil regions rich in synaptic terminals (Buchner et al., 1986; Gorzyca and Hall, 1987; Ikeda and Salvaterra, 1989). In contrast, vertebrate choline acetyltransferase protein is detected easily in cell soma and only with more difficulty in synaptic terminals (Salvaterra and Vaughn, 1989). Perhaps the unusual trans- lation initiation codon used by Drosophila choline acetyltrans- ferase is important for routing the protein to its correct intracellular destination. Interestingly, in this regard, two partial vertebrate choline acetyltransferase cDNA clones ap- pear to use conventional ATG starting codons (Berrard et al., 1987; Brice et al., 1989). It should also be mentioned, however, that the predicted ATG initiation codons for vertebrate cho- line acetyltransferase are in a particularly unfavorable context for translation initiation (Berrard et al., 1987; Brice et al., 1989), perhaps indicating that they are used inefficiently in uiuo. It will thus be important to obtain more complete infor-
mation about the initiation site for protein translation from choline acetyltransferase in a number of species before the significance and generality of the non-ATG initiation site in Drosophila choline acetyltransferase can be assessed.
Nervous system genes in general often contain unusually long 5’-untranslated leader sequences and several examples are related to acetylcholine metabolism (Bossy et al., 1988). It is also more common for Drosophila mRNAs to contain ATG triplets upstream from known start codons when com- pared to their vertebrate counterparts (Cavener, 1987). The significance of these sequence motifs are unknown. The list of known mRNA sequences in different species is still rather selective and some vertebrate genes, such as oncogenes, seem to contain an unusually high number of upstream ATGs (Mueller and Hinnebusch, 1986).
Perhaps more significantly for in uiuo production of choline acetyltransferase protein, we have also noted a significant difference in the translational efficiency of full length RNA relative to the shorter pCha-12 RNA. The combination of a non-ATG start codon and the translational inefficiency con- tributed by the untranslated 5’ and 3’ sequences present in choline acetyltransferase mRNA should result in reduced translational efficiency in uiuo. Perhaps cholinergic neurons contain a factor which can stimulate more efficient transla- tion of choline acetyltransferase mRNA under conditions of increased demand. An analogous translational regulatory sys- tem has been elegantly described for ferritin mRNA whose translational efficiency is increased in response to hemin and repressed by a 90-kDa protein from liver (Brown et al., 1989; Lin et al., 1990). The existence of such a translational factor for choline acetyltransferase mRNA remains to be demon- strated.
Acknowledgments-We wish to thank K. Oghi for performing some of the initial experiments using rabbit reticulocyte lysate translation of choline acetyltransferase RNA, Sharyn Webb for preparing the manuscript, and our departmental colleagues for their encourage- ment, advice, and critical comments.
REFERENCES
Acland, P., Dixon, M., Peters, G., and Dickson, C. (1990) Nature 343, 662- 665
Barber, R. P., Sugihara, H., Lee, M., Vaughn, J. E., and Salvaterra, P. M. (1988) J. Camp. Neural. 280.533-543
Becerra, S. P., Rose, J. A., Hardy, M., Baroudy, B. M., and Anderson, C. W. (1985) Proc. N&l. Acad. Sci. U S. A. 82,7919-7923
Berrard, S., Brice, A., Lottspeich, F., Braun, A., Barde, Y.-A., and Mallet, J. (1987) Proc. Natl. Acad. Sci. U. S. A. 84,9280-9294
Bossy, B., Ballivet, M., and Spierer, P. (1988) EMBO J. ‘7,611-618 Brice, A., Berrad, S., Raynaud, B., Ansieau, S., Coppola, T., Weber, M. J., and
Mallet, J. (1989) J. Neurosci. Res. 23, 266-273 Brown, P. H., Daniel+McQueen, S., Walden, W. E., Patino, M. M., Gaffeld,
L., Bielser, D., and Thach, R. E. (1989) J. Biol. Chem. 264,13383-13386 Buchner, E., Buchner, S., Crawford, G., Mason, W. T., Salvaterra, P. M., and
Sattelle, D. B. (1986) Cell Tissue Res. 246, 57-62 Cavener, D. R. (1987) Nucleic Acids Res. 16, 1353-1361 Craigen, W. J., and Caskey, C. T. (1987) Cell 60, l-2 Curran, J. F., and Yarus, M. (1987) Science 238,1545-1550 Florkiewicz, R. A., and Sommer, A. (1989) Proc. NatL Acad. Sci. U. S. A. 86,
3978-3981 Fonnum, F. (1975) J. Neurochem. 24,407-409 Gold, L. (1988) Anna. Reo. Biochem. 67, 199-233 Gorzyca, M. G., and Hall, J. C. (1987) J. Neurosci. 7,1361-1369 Gubler, U., and Hoffman, B. J. (1983) Gene (Amst.) 26,263-269 Gupta, K. C., and Patwardhan, S. (1988) J. BioL Chem. 263,8553-8556 Hann. S. R.. Kine. M. W.. Bentlev. D. L.. Anderson, C. W., and Eisenman, R.
N. (1988)‘Cell ai, 185-i95 ” Ikeda, K., and Salvaterra, P. M. (1989) J. Camp. Neural. 280,283-290 Itoh, N., Salvaterra, P., and Itakura, K. (1985) Drosp. Inf. Ser. 61, 89 Itoh, N., Slemmon, J. E., Hawke, D. H., Williamson, R., Morita, E., Itakura,
D., Roberts, E., Shively, J., Crawford, G. D., and Salvaterra, P. M. (1986) Proc. Natl. Acad. Sci. U. S. A. 83.4081-4085
Kozak, M. (1983) Microbial. Rev. 49, l-45 Kozak, M. (1986a) Cell 44,283-292 Kozak, M. (1986b) Cell 47,481-483 Kozak, M. (1987) Nucleic Acids Res. 16.8125-8148 Laemmli, U. K. (1970) Nature 227,680-685 Lin, J.-J., Daniels-McQueen, S., Patino, M. M., GaffeId, L., Walden, W. E.,
and Thach, R. E. (1990) Science 247,74-77 Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Choline Acetyltransferase Translation
Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, Peabody, D. S. (1989) J. &I o-l.~-r__-- n \” .-A I,^.... 11. Chem. 264,5031-5035 NY DIIV~LOII~, r. IV,. L(I,U v aughn, J. E. (1989) Int. Reu. Neurobiol. 31, 81-143
McCaman, R. E., Carbini, L., Maines, V., and Salvaterra, P. M. (1988) Mol. 511 “‘emmon J. R. galvaterra P. M., Crawford, G. D., and Roberts, E. (1982) J.
Brain Res. 3, 107-114 DI^I m!.... “‘E” 9Q)*, “&2 UL”L. L,‘CIIC. a”, , 00-z I-UC
ElPmmnn .T R Sa,“.+mra Mueller, P. P., and Hinnebusch, A. G. (1986) Celf 45,201 -207
“Ib.I.III.U.L, “. .*., YY....“-.-.., P. M., and Roberts, E. (1984) Neurochem. Int. 6,
Muiioz-Maines, V. J., Selmmon, J. R., Panicker, M. M., Neighbo or, N., and Salvaterra, P. M. (1988) J. Neurochem. 50, 167-175
S$&!z5H. Andrisani V. and Salvaterra P. M. (1988) FASEB J. 2, 6266 Zitoner, k. $. Walthalf, D: A., Rymond, d. C., and Hollenberg, C. P. (1984)
Peabody, D. S. (1987) J. Biol. Chem. 262,11847-18851 Mol. Cell. B;ol. 4, 1191-1197
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

H Sugihara, V Andrisani and P M Salvaterralength RNA is inefficiently translated.
Drosophila choline acetyltransferase uses a non-AUG initiation codon and full
1990, 265:21714-21719.J. Biol. Chem.
http://www.jbc.org/content/265/35/21714Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/35/21714.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from


















