
Characterization of the porcine reproductive and respiratory syndrome virus glycoprotein 5 (GP5) in...
6
Virus Research 104 (2004) 33–38 Characterization of the porcine reproductive and respiratory syndrome virus glycoprotein 5 (GP5) in stably expressing cells Changhee Lee a , Dragan Rogan b , Larry Erickson c , Jian Zhang c , Dongwan Yoo a,∗ a Department of Pathobiology, Ontario Veterinary College, University of Guelph, Guelph, Ontario, Canada N1G 2W1 b Bioniche Animal Health, Belleville, Ontario, Canada K8N 1E2 c Department of Plant Agriculture, University of Guelph, Guelph, Ontario, Canada N1G 2W1 Received 28 November 2003; received in revised form 1 March 2004; accepted 4 March 2004 Available online 23 April 2004 Abstract Porcine reproductive and respiratory syndrome virus (PRRSV) induces apoptosis in cultured cells and in infected tissues of pig, and the GP5 protein was reported to be responsible. To study the role of apoptosis during PRRSV infection, we established a subline of HeLa cells stably expressing the PRRSV GP5 protein under an inducible promoter to prevent cell death that may result from GP5 expression. The expression of GP5 was confirmed upon induction by immunofluorescence, Western blot, and immunoprecipitation. No reduction in cell numbers was observed in the GP5-expressing cells, and neither DNA fragmentation nor strand-breaks were detected in these cells. To examine if bystander cells underwent apoptosis, Marc-145 or HeLa cells were co-cultivated with GP5-expressing cells, and the apoptotic characteristics were examined in the bystanders. Neither DNA laddering, strand-breaks, nor reduction in cell numbers was observed in the co-cultivated cells. Gene expression profiles were examined for both GP5-expressing cells and bystanders by apoptosis-specific macroarrays and gene chip-based microarrays, but no genes related to apoptosis were specifically regulated. Our data suggest that the previously reported GP5-induced apoptosis may be an atypical observation. The GP5-expressing cells were immunoreactive with sera from pigs infected with the North American and European types of PRRSV, which implicates a potential usefulness of these cells as a diagnostic reagent for PRRSV. © 2004 Elsevier B.V. All rights reserved. Keywords: PRRS; GP5 protein; Apoptosis; Arterivirus Porcine reproductive and respiratory syndrome virus (PRRSV) is an emerged virus almost simultaneously in U.S.A. and Europe in the late 1980s and early 1990s, re- spectively (Keffaber, 1989; Wensvoort et al., 1991), and causes significant economic losses in the swine industry worldwide. PRRSV infection is characterized by abortions in sows and respiratory problems in pigs of all ages (Halbur et al., 1995). PRRSV is an enveloped virus containing a genome of single-stranded positive-sense RNA belonging to the family Arteriviridae, which has recently been re-grouped into the newly created order Nidovirales, together with the Coronaviridae family (Cavanagh, 1997). The PRRSV genome is approximately 15 kb in size and contains nine open reading frames (ORFs), designated ORF1a, ORFlb, and ORFs 2–7, in order starting from the 5 -end of the ∗ Corresponding author. Tel.: +519-824-4120x54729; fax: +519-767-0809. E-mail address: [email protected] (D. Yoo). genome. Both ORF1a and ORF1b code for non-structural proteins that are believed to be involved in genome replica- tion, and ORFs 2–7 code for viral structural proteins GP2a, GP2b, GP3, GP4, GP5, membrane (M) protein, and nucle- ocapsid (N) protein, respectively (Meulenberg et al., 1995). PRRSV has been reported to induce apoptosis in pig dur- ing infection as well as in cultured cells in vitro (Sirinarumitr et al., 1998; Sur et al., 1998). PRRSV-induced apoptosis occurs in alveolar macrophages and mononuclear cells in lungs and lymphoid tissues but is also observed in testic- ular germinal epithelial cells (Sur et al., 1997). It is note- worthy that most apoptotic cells do not co-localize with virus-infected cells in vivo, and PRRSV-induced apopto- sis has been mainly observed in uninfected bystander cells rather than infected cells (Sirinarumitr et al., 1998; Sur et al., 1998; Labarque et al., 2003). In 1996, Suárez et al. (1996) reported that the GP5 protein of PRRSV caused a strong cytotoxicity and that GP5-induced apoptosis was indepen- dent from the anti-apoptotic protein Bcl-2. Subsequently by 0168-1702/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.virusres.2004.03.001
-
Upload
changhee-lee -
Category
Documents
-
view
217 -
download
4
Transcript of Characterization of the porcine reproductive and respiratory syndrome virus glycoprotein 5 (GP5) in...

Virus Research 104 (2004) 33–38
Characterization of the porcine reproductive and respiratory syndromevirus glycoprotein 5 (GP5) in stably expressing cells
Changhee Leea, Dragan Roganb, Larry Ericksonc, Jian Zhangc, Dongwan Yooa,∗a Department of Pathobiology, Ontario Veterinary College, University of Guelph, Guelph, Ontario, Canada N1G 2W1
b Bioniche Animal Health, Belleville, Ontario, Canada K8N 1E2c Department of Plant Agriculture, University of Guelph, Guelph, Ontario, Canada N1G 2W1
Received 28 November 2003; received in revised form 1 March 2004; accepted 4 March 2004
Available online 23 April 2004
Abstract
Porcine reproductive and respiratory syndrome virus (PRRSV) induces apoptosis in cultured cells and in infected tissues of pig, and the GP5protein was reported to be responsible. To study the role of apoptosis during PRRSV infection, we established a subline of HeLa cells stablyexpressing the PRRSV GP5 protein under an inducible promoter to prevent cell death that may result from GP5 expression. The expressionof GP5 was confirmed upon induction by immunofluorescence, Western blot, and immunoprecipitation. No reduction in cell numbers wasobserved in the GP5-expressing cells, and neither DNA fragmentation nor strand-breaks were detected in these cells. To examine if bystandercells underwent apoptosis, Marc-145 or HeLa cells were co-cultivated with GP5-expressing cells, and the apoptotic characteristics wereexamined in the bystanders. Neither DNA laddering, strand-breaks, nor reduction in cell numbers was observed in the co-cultivated cells.Gene expression profiles were examined for both GP5-expressing cells and bystanders by apoptosis-specific macroarrays and gene chip-basedmicroarrays, but no genes related to apoptosis were specifically regulated. Our data suggest that the previously reported GP5-induced apoptosismay be an atypical observation. The GP5-expressing cells were immunoreactive with sera from pigs infected with the North American andEuropean types of PRRSV, which implicates a potential usefulness of these cells as a diagnostic reagent for PRRSV.© 2004 Elsevier B.V. All rights reserved.
Keywords: PRRS; GP5 protein; Apoptosis; Arterivirus
Porcine reproductive and respiratory syndrome virus(PRRSV) is an emerged virus almost simultaneously inU.S.A. and Europe in the late 1980s and early 1990s, re-spectively (Keffaber, 1989; Wensvoort et al., 1991), andcauses significant economic losses in the swine industryworldwide. PRRSV infection is characterized by abortionsin sows and respiratory problems in pigs of all ages (Halburet al., 1995). PRRSV is an enveloped virus containing agenome of single-stranded positive-sense RNA belonging tothe family Arteriviridae, which has recently been re-groupedinto the newly created order Nidovirales, together withthe Coronaviridae family (Cavanagh, 1997). The PRRSVgenome is approximately 15 kb in size and contains nineopen reading frames (ORFs), designated ORF1a, ORFlb,and ORFs 2–7, in order starting from the 5′-end of the
∗ Corresponding author. Tel.:+519-824-4120x54729;fax: +519-767-0809.
E-mail address: [email protected] (D. Yoo).
genome. Both ORF1a and ORF1b code for non-structuralproteins that are believed to be involved in genome replica-tion, and ORFs 2–7 code for viral structural proteins GP2a,GP2b, GP3, GP4, GP5, membrane (M) protein, and nucle-ocapsid (N) protein, respectively (Meulenberg et al., 1995).
PRRSV has been reported to induce apoptosis in pig dur-ing infection as well as in cultured cells in vitro (Sirinarumitret al., 1998; Sur et al., 1998). PRRSV-induced apoptosisoccurs in alveolar macrophages and mononuclear cells inlungs and lymphoid tissues but is also observed in testic-ular germinal epithelial cells (Sur et al., 1997). It is note-worthy that most apoptotic cells do not co-localize withvirus-infected cells in vivo, and PRRSV-induced apopto-sis has been mainly observed in uninfected bystander cellsrather than infected cells (Sirinarumitr et al., 1998; Sur et al.,1998; Labarque et al., 2003). In 1996,Suárez et al. (1996)reported that the GP5 protein of PRRSV caused a strongcytotoxicity and that GP5-induced apoptosis was indepen-dent from the anti-apoptotic protein Bcl-2. Subsequently by
0168-1702/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.virusres.2004.03.001

34 C. Lee et al. / Virus Research 104 (2004) 33–38
the same investigators, the domain responsible for apoptosiswas mapped to the first 119 amino acids of the GP5 protein(Fernandez et al., 2002). The biological relevance of apop-tosis to PRRSV pathogenesis is unknown.
As a first step towards studying the effects of thePRRSV GP5 protein on cell function, a sub-line of HeLacells was established to express GP5 under the controlof an inducible promoter, the tetracycline resistance op-erator of E. coli. The ORF5 gene was inserted into thetetracycline-responsive plasmid pTRE-hyg and the expres-sion plasmid pTRE-ORF5-hyg was constructed. For trans-formation with the expression plasmid, HeLa-Tet-off cells(Clontech) were chosen. HeLa-Tet-off cells are previouslytransformed cells with the regulatory plasmid pTet-off andtherefore express the tetracycline transactivator (tTA). Thepresence of tetracycline or its derivative doxycycline in theculture medium prevents binding of tTA to the promoter,while the removal of tetracycline induces gene transcrip-tion placed under control of the promoter (Gossen andBujard, 1992). HeLa-Tet-off cells were transfected withpTRE-ORF5-hyg to introduce the ORF5 gene to the cellsand to confer resistance to hygromycin selection. Trans-fected cells were maintained in the presence of 300�g/mlof hygromycin and 1�g/ml of doxycycline in order toprevent the expression of GP5 during selection.
Twenty hygromycin-resistant cell clones were initiallyobtained and screened by PCR and RT-PCR to determinethe ORF5 gene integration and the corresponding mRNA,respectively. Six clones were confirmed to be positive (datanot shown). The selected clones were further examinedfor protein expression by immunofluorescence. The cy-toplasmic fluorescence was evident in the cloned HeLacells incubated with PRRSV-specific hyperimmune serum(Fig. 1A), whereas no fluorescence was observed in normalHeLa and HeLa-Tet-off cells, confirming that the clonedcells expressed the GP5 protein. Every cell expressed GP5,showing a homogenous population of cells expressingGP5. The established cells also reacted well with the hy-perimmune pig serum raised against the European strainof PRRSV Lelystad virus (Fig. 1A). The parental HeLaTet-off cells and uninfected Marc-145 cells did not displayimmunofluorescence. One of the six cell clones was chosenfor subsequent studies and designated HeLa-GP5.
To further confirm the GP5 expression, immunoprecip-itation and Western blot were conducted. HeLa-GP5 cellswere radiolabeled at 48 h of induction for 6 h and subjectedto immunoprecipitation with pig anti-PRRSV hyperimmuneserum (Fig. 1B). A specific band of 25 kDa was identified inthe induced cells (lane 2), and its migration rate was similaras the authentic GP5 protein seen in PRRSV-infected cells(lane 3). This band was absent in the parental HeLa-Tet-offcells (lane 4). A band migrating at the same rate was ob-served at low levels in the uninduced condition of HeLa-GP5(lane 1), and it was considered a leaky expression. Westernblot was carried out to examine the accumulated level ofGP5 upon induction (Fig. 1C). HeLa-GP5 cells were culti-
vated in the absence of doxycycline, and the GP5 expressionwas monitored at various times post-induction. GP5 expres-sion was evident at 24 h post-induction (lane 2) and con-tinued to accumulate (lane 5). Because of the leaky expres-sion of GP5 under uninduced conditions, the non-transfectedparental HeLa-Tet-off cells were used as a negative controlin the subsequent studies.
Since apoptosis was one of the reported functions of GP5,an attempt was made to examine whether the newly estab-lished cells expressing GP5 would undergo cell death. Ifthese cells underwent apoptotic death, cell numbers woulddecline. For this experiment, HeLa-GP5 cells were grownin the presence or absence of doxycycline and the numberof cells was counted daily. The parental HeLa-Tet-off cellswere included as a negative control, and three repeated ex-periments were conducted to presentFig. 2A. The GP5 ex-pression did not affect cell growth, although the number ofHeLa-GP5 cells was slightly lower than HeLa-Tet-off cells,starting 2 days post-induction. Considering the homogenouspopulation of GP5-expressing cells, the difference in cellnumbers did not reflect cell death, and thus it was concludedthat the GP5 protein expression did not cause cytotoxicityin HeLa-GP5 cells.
PRRSV has been reported to induce apoptosis mainlyin uninfected bystander cells in cell culture and in tis-sues of infected animals. Since it was unable to detectapoptosis-related cell death in GP5-expressing cells, by-stander cells were examined for possible apoptosis. Forthis, Marc-145 cells were co-cultivated with HeLa-GP5 orHeLa-Tet-off cells at a ratio of 1:1. Marc-145 is the cellsubline derived from MA-104 cells (Kim et al., 1993) andis the only available cell line so far that supports PRRSVinfection in culture. The co-cultivation acts to simulate by-stander cells that neighbor GP5-expressing cells. As shownin Fig. 2B, the growth pattern of co-cultivated cells wasalmost identical to those co-cultivated with HeLa-Tet-offcontrol cells, suggesting the absence of cell death in thebystander cells.
To further determine the possible cell death which mightoccur unnoticeably, inter-nucleosomal DNA fragmentationwas determined. Cellular DNA was extracted at differenttimes post-induction and the extracted DNA was resolvedby agarose gel electrophoresis. The characteristic DNA frag-mentation was observed in PRRSV-infected Marc-145 cells24 and 48 h post-infection (Fig. 3A, lanes 2, 3). In contrast,DNA fragmentation was continually absent in HeLa-GP5cells up to 96 h post-induction (Fig. 3B, lanes 2–5). DNAcleavage was also examined by a terminal deoxynucleotidyltransferase-mediated dUTP nick-end labeling (TUNEL) as-say using the ApoAlert DNA fragmentation kit (Clontech).HeLa-GP5 cells were grown on microscope coverslips upto 4 days post-induction to allow sufficient expression ofGP5 prior to the TUNEL assay. Marc-145 cells infectedwith PRRSV showed distinct staining patterns typical forDNA strand-breaks, indicating that apoptosis is mediated byPRRSV infection (Fig. 3D). HeLa-GP5 cells, however, did

C. Lee et al. / Virus Research 104 (2004) 33–38 35
Fig. 1. Stable expression of the PRRSV GP5 protein in HeLa cells. (A) Immunofluorescence staining of HeLa cells expressing the GP5 protein. Proteinexpression was induced by removal of doxycycline. Cells were fixed and incubated with the anti-PRRSV pig serum followed by FITC-conjugated goatanti-swine antibody. The fluorescence was visualized by a fluorescent microscope at 40× magnification. Panels: (a) Marc-145 cells at 24 h post-infectionwith PRRSV; (b) HeLa-GP5 cells at 48 h post-induction incubated with anti-PA8 virus (North American strain of PRRSV) porcine serum as a primaryantibody; (c) HeLa-GP5 cells at 48 h post-induction without doxycycline incubated with anti-Lelystad virus (European strain of PRRSV) porcine serum.(B) Radioimmunoprecipitation of the GP5 protein. HeLa-GP5 cells were induced by growing in absence of doxycycline for 48 h and radiolabeled with[35S]methionine for 5 h. The cell lysates were immunoprecipitated using the porcine anti-PRRSV serum. The immuncomplex was precipitated by Sepharosebeads and resolved by SDS–12% PAGE followed by autoradiography. Lanes: 1, HeLa-GP5 cell in the presence of doxycycline; 2, HeLa-GP5 cells in theabsence of doxycycline; 3, Marc-145 cells infected with PRRSV; 4, the parental HeLa-Tet-off cells. In virus-infected Marc-145 cells (lane 3), threemajorviral proteins are seen evidently (arrows), representing the nucleocapsid (N), membrane (M), and GP5 proteins. (C) Immunoblot of GP5. Cells were grownin the absence of doxycycline for 24, 48, 72, and 96 h. Cell lysates were prepared and subjected to Western blot using the porcine anti-PRRSV antibody.
not produce any nuclear staining (Fig. 3E). The absence ofcell death, measured through the TUNEL assay along withthe cell counting and DNA fragmentation, suggests that GP5may not be an apoptotic inducer. DNA fragmentation andTUNEL assays were conducted for bystander cells (Fig. 3Cand F), but no further evidence was obtained for apoptotic
Fig. 2. Growth characteristics of HeLa-GP5 and Marc-145 bystander cells co-cultivated with HeLa-GP5. (A) HeLa-GP5 cells grown in the absence orpresence of doxycycline. (B) Marc-145 cells co-cultivated with HeLa-GP5 or HeLa-Tet-off cells at 1:1 ratio. Number of cells were determined at theindicated times using a hemocytometer. Three independent experiments were performed and the average values were plotted.
cell death in bystanders, neither by DNA fragmentation norby TUNEL assays. Taken together, we concluded the ab-sence of apoptosis both in the GP5-expressing HeLa cellsas well as in the bystander Marc-145 cells.
To investigate whether cellular genes involved in theapoptotic pathway are affected by the GP5 protein ex-
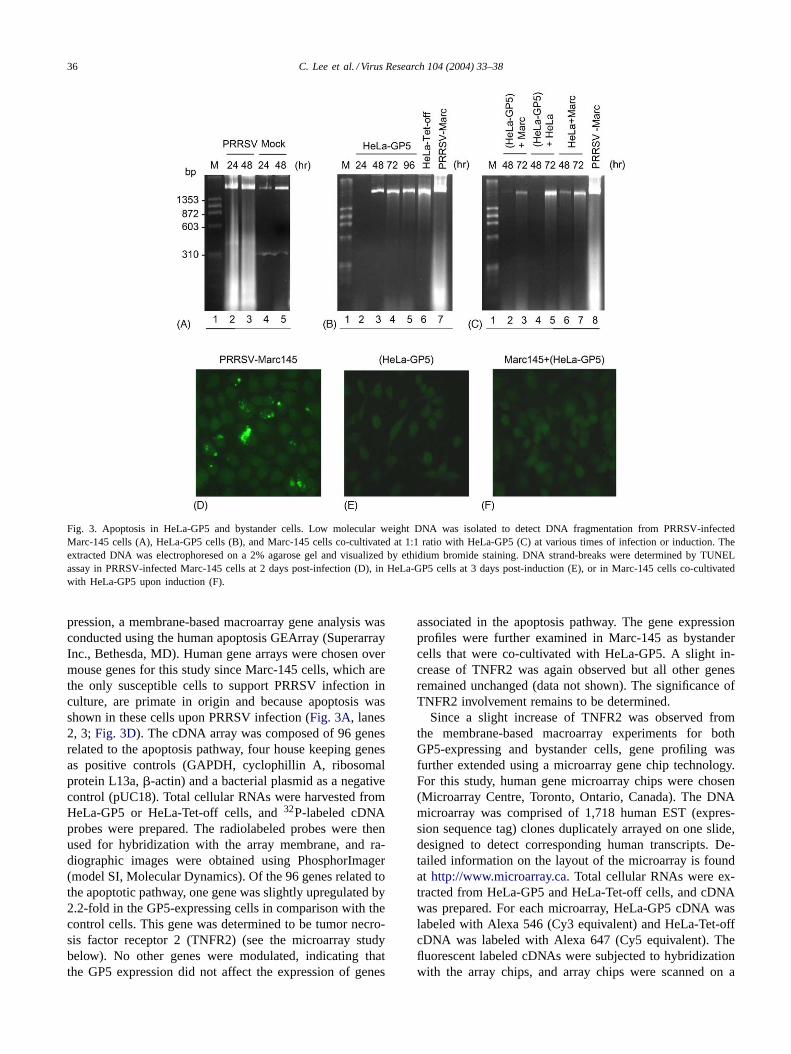
36 C. Lee et al. / Virus Research 104 (2004) 33–38
Fig. 3. Apoptosis in HeLa-GP5 and bystander cells. Low molecular weight DNA was isolated to detect DNA fragmentation from PRRSV-infectedMarc-145 cells (A), HeLa-GP5 cells (B), and Marc-145 cells co-cultivated at 1:1 ratio with HeLa-GP5 (C) at various times of infection or induction. Theextracted DNA was electrophoresed on a 2% agarose gel and visualized by ethidium bromide staining. DNA strand-breaks were determined by TUNELassay in PRRSV-infected Marc-145 cells at 2 days post-infection (D), in HeLa-GP5 cells at 3 days post-induction (E), or in Marc-145 cells co-cultivatedwith HeLa-GP5 upon induction (F).
pression, a membrane-based macroarray gene analysis wasconducted using the human apoptosis GEArray (SuperarrayInc., Bethesda, MD). Human gene arrays were chosen overmouse genes for this study since Marc-145 cells, which arethe only susceptible cells to support PRRSV infection inculture, are primate in origin and because apoptosis wasshown in these cells upon PRRSV infection (Fig. 3A, lanes2, 3; Fig. 3D). The cDNA array was composed of 96 genesrelated to the apoptosis pathway, four house keeping genesas positive controls (GAPDH, cyclophillin A, ribosomalprotein L13a,�-actin) and a bacterial plasmid as a negativecontrol (pUC18). Total cellular RNAs were harvested fromHeLa-GP5 or HeLa-Tet-off cells, and32P-labeled cDNAprobes were prepared. The radiolabeled probes were thenused for hybridization with the array membrane, and ra-diographic images were obtained using PhosphorImager(model SI, Molecular Dynamics). Of the 96 genes related tothe apoptotic pathway, one gene was slightly upregulated by2.2-fold in the GP5-expressing cells in comparison with thecontrol cells. This gene was determined to be tumor necro-sis factor receptor 2 (TNFR2) (see the microarray studybelow). No other genes were modulated, indicating thatthe GP5 expression did not affect the expression of genes
associated in the apoptosis pathway. The gene expressionprofiles were further examined in Marc-145 as bystandercells that were co-cultivated with HeLa-GP5. A slight in-crease of TNFR2 was again observed but all other genesremained unchanged (data not shown). The significance ofTNFR2 involvement remains to be determined.
Since a slight increase of TNFR2 was observed fromthe membrane-based macroarray experiments for bothGP5-expressing and bystander cells, gene profiling wasfurther extended using a microarray gene chip technology.For this study, human gene microarray chips were chosen(Microarray Centre, Toronto, Ontario, Canada). The DNAmicroarray was comprised of 1,718 human EST (expres-sion sequence tag) clones duplicately arrayed on one slide,designed to detect corresponding human transcripts. De-tailed information on the layout of the microarray is foundat http://www.microarray.ca. Total cellular RNAs were ex-tracted from HeLa-GP5 and HeLa-Tet-off cells, and cDNAwas prepared. For each microarray, HeLa-GP5 cDNA waslabeled with Alexa 546 (Cy3 equivalent) and HeLa-Tet-offcDNA was labeled with Alexa 647 (Cy5 equivalent). Thefluorescent labeled cDNAs were subjected to hybridizationwith the array chips, and array chips were scanned on a

C. Lee et al. / Virus Research 104 (2004) 33–38 37
GenPix 4000A scanner (Axon Instruments Inc., Union City,CA). The normalization and data analysis were performedusing GeneTraffic microarray data analysis software (Io-bion Informatics, La Jolla, CA). The log2 R/G (Cy5/Cy3)normalized ratio was selected as the value that correspondsto a fold change in regulation. When a fold-change wasconsistently 1.0 or greater in two independent experiments,the value was considered significant.
The levels of cellular mRNAs were significantly alteredfor 20 genes by the GP5 expression in HeLa cells. Ofthese, 11 genes were up-regulated and nine genes weredown-regulated (data not shown). The affected genes rep-resented those involved in the immune response and stresstoxicity, signal transduction pathways, cell cycle and cancersignals. Five genes were involved in synthesis, transport,replication, transcription, and translation. The microarrayprofiles were also examined in the bystander Marc-145cells co-cultivated with HeLa-GP5, and similar results wereobtained (data not shown), confirming that the gene reg-ulation patterns were consistent. No pro-apoptotic geneswere identified from the microarray experiments, includ-ing the TNFR2 gene detected as a slight increaser in themembrane array. This difference between the membranearray and microarray may be due to the sensitivity of bothtechniques, especially for those genes whose expressionis not significantly modulated. An alternative method toprecisely measure differential mRNA expression, such asreal-time quantitative RT-PCR, may be required to con-firm the significance of the genes that have been identifiedin the present study. From both the macroarray and mi-croarray gene profiling studies, it was concluded that nospecific apoptosis-related cellular genes were altered byGP5 expression in both GP5-expressing cells and bystandercells.
The present study was initiated to understand the roleof apoptosis during PRRSV infection and to investigate theGP5 function in this process. We established a cell line stablyexpressing the GP5 gene that was placed under the induciblepromoter using the Tet-on/Tet-off system. However, in bothGP5-expressing cells and bystander cells co-cultivated withthe GP5-expressing cells, it was not possible to observe anynoticeable evidence of cell death mediated by GP5 in ourexperimental conditions. The difference between the pre-vious reports by other investigators (Suárez et al., 1996;Fernandez et al., 2002) and the current study may reflect thefact that, unlike the authentic GP5 protein used in our study,GP5 fusion constructs were used in the previous studies suchthat additional amino acids representing the repeat of histi-dine tag or an antigenic tag were attached to the N-terminusof the ORF5 protein. These extra tags may have disruptedthe signal function of the N-terminus of the GP5 protein,since the first 31 amino acids at the N-terminus of GP5 arethought to be a putative endoplasmic reticulum (ER) translo-cational sequence (Mardassi et al., 1995; Meulenberg et al.,1995) that usually directs the protein into the ER. The mod-ification of the GP5 sequence such as in the previous studies
and thus the altered intracellular trafficking of the modifiedprotein may have caused the atypical apoptosis. Further-more, the recombinant vaccinia virus expressing T7 poly-merase was used in the previous studies to express the GP5fusion protein in cells, and thus the use of vaccinia virusmay also possibly have contributed to the unusual apopto-sis. Alternatively, a cell-specific factor may be involved inthe GP5-related apoptosis, since BSC-40 cells were used inthe previous studies, whereas HeLa and bystander Marc-145cells were used in the present study. It is noteworthy thatboth BSC-40 and Marc-145 cells are the same African greenmonkey (Cercopithecus aethiops) kidney in origin, but onlyMarc-145 cells are susceptible for PRRSV infection.
The immunofluorescence assay displayed strong cytoplas-mic fluorescence in GP5-expressing cells by anti-PRRSVhyperimmune pig serum raised against either the NorthAmerican or European type of PRRSV. Given this obser-vation, the established cell line expressing the GP5 proteinmay serve as an useful diagnostic reagent for detectingthe PRRSV antibodies in pigs or as a research tool forGP5-specific antibody production or vaccination studies.
Acknowledgements
The authors are grateful to Ryan Dowling for his techni-cal assistance in the microarray analysis. Funding for thisresearch was provided by Ontario Pork, the Ontario Min-istry of Agriculture and Food, and by the Canadian NationalScience and Engineering Research Councils Strategic Part-nership Program.
References
Cavanagh, D., 1997. Nidovirales: a new order comprising Coronaviridaeand Arteriviridae. Arch. Virol. 142, 629–633.
Fernandez, A., Suárez, P., Castro, J.M., Tabares, E., Diaz-Guerra, M.,2002. Characterization of regions in the GP5 protein of porcine repro-ductive and respiratory syndrome virus required to induce apoptoticcell death. Virus Res. 83, 103–118.
Gossen, M., Bujard, H., 1992. Tight control of gene expression in mam-malian cells by tetracycline-responsive promoters. Proc. Natl. Acad.Sci. U.S.A. 89, 5547–5551.
Halbur, P.G., Paul, P.S., Frey, M.L., Landgraf, J., Eernisse, K., Meng,X.J., Lum, M.A., Andrews, J.J., Rathje, J.A., 1995. Comparison ofthe pathogenicity of two U.S. porcine reproductive and respiratorysyndrome virus isolates with that of Lelystad virus. Vet. Pathol. 32,648–660.
Keffaber, K.K., 1989. Reproductive failure of unknown etiology. Am.Assoc. Swine Practitioners Newsletter 1, 1–9.
Kim, H.S., Kwang, J., Yoon, I.J., Joo, H.S., Frey, M.L., 1993. Enhancedreplication of porcine reproductive and respiratory syndrome (PRRS)virus in a homogeneous subpopulation of MA-104 cell line. Arch.Virol. 133, 477–483.
Labarque, G., Van Gucht, S., Nauwynck, H., Van Reeth, K., Pensaert, M.,2003. Apoptosis in the lungs of pigs infected with porcine reproductiveand respiratory syndrome virus and associations with the productionof apoptogenic cytokines. Vet. Res. 34, 249–260.

38 C. Lee et al. / Virus Research 104 (2004) 33–38
Mardassi, H., Mounir, S., Dea, S., 1995. Molecular analysis of the ORFs3–7 of porcine reproductive and respiratory syndrome virus, Quebecreference strain. Arch. Virol. 140, 1405–1418.
Meulenberg, J.J.M., den Besten, A., de Kluyer, F.P., Moormann, R.J.M.,Schaaper, V.M.M., Wensvoort, G., 1995. Characterization of proteinsencoded by ORFs 2–7 of Lelystad virus. Virology 206, 155–163.
Sirinarumitr, T., Zhang, Y., Kluge, J.P., Halbur, P.G., Paul, P.S., 1998.A pneumo-virulent United States isolate of porcine reproductive andrespiratory syndrome virus induces apoptosis in bystander cells bothin vitro and in vivo. J. Gen. Virol. 79, 2989–2995.
Suárez, P., D́ıaz-Guerra, M., Prieto, C., Esteban, M., Castro, J.M., Nieto,A., Ort́ın, J., 1996. Open reading frame 5 of porcine reproductive and
respiratory syndrome virus as a cause of virus-induced apoptosis. J.Virol. 70, 2876–2882.
Sur, J.H., Doster, A.R., Christian, J.S., Galeota, J.A., Wills, R.W., Zim-merman, J.J., Osorio, F.A., 1997. Porcine reproductive and respiratorysyndrome virus replicates in testicular germ cells, alters spermatogene-sis, and induces germ cell death by apoptosis. J. Virol. 71, 9170–9179.
Sur, J.H., Doster, A.R., Osorio, F.A., 1998. Apoptosis induced in vivo dur-ing acute infection by porcine reproductive and respiratory syndromevirus. Vet. Pathol. 35, 506–512.
Wensvoort, G., Tepstra, C., Pol, J.M.A., ter Laak, E.A., 1991. Mysteryswine disease in the Netherlands: the isolation of Lelystad virus. Vet.Q 13, 121–130.



![Rational, unirational and stably rational varietiespirutka/survey.pdf · could be rational (resp. stably rational, resp. retract rational) [30, p.282]. Unirational nonrational varieties.](https://static.fdocuments.us/doc/165x107/5f8fad2d18211140cf6c6b61/rational-unirational-and-stably-rational-varieties-pirutka-could-be-rational.jpg)














