
Characterization of soybean peroxidase for the treatment of aqueous phenols
11
Characterization of soybean peroxidase for the treatment of aqueous phenols Harold Wright, James A. Nicell * Department of Civil Engineering & Applied Mechanics, McGill University, 817 Sherbrooke Street West, Montreal PQ,Canada H3A 2K6 Received 24 July 1998; revised 12 January 1999; accepted 18 January 1999 Abstract The application of soybean peroxidase (SBP) to catalyze the polymerization and precipitation of aqueous phenols by hydrogen peroxide is potentially promising because this peroxidase is less expensive than horseradish peroxidase (HRP), which has been the focus of most wastewater research. SBP can act on a broad range of compounds and retains its catalytic ability over wide ranges of temperature and pH. Activity was optimal at pH 6.4, with significant activity observed between pH 3 and 9. SBP was very stable at 25°C at neutral and alkaline conditions but experienced rapid inactivation below pH 3. SBP underwent biphasic inactivation by hydrogen peroxide in the absence of a reductant substrate. SBP was most eective when used to treat phenolic solutions between pH 6 and 9. In comparison with HRP, the activity of SBP was only slightly more sensitive to pH, was more stable at elevated tem- peratures, and was less susceptible to permanent inactivation by hydrogen peroxide. However, SBP was catalytically slower than HRP and a larger molar quantity of SBP was usually required to remove a given quantity of phenolic substrate. Ó 1999 Elsevier Science Ltd. All rights reserved. Keywords: Soybean peroxidase; Wastewater treatment; Inactivation; Stability; Temperature; pH 1. Introduction The catalytic removal of phenolic and other aromatic compounds from wastewaters using peroxidase enzyme and hydrogen peroxide has been the focus of extensive research since the initial work of Klibanov et al. (1980). Treatment involves the catalytic oxidation of the aro- matic contaminants by hydrogen peroxide to form ar- omatic free radicals that subsequently combine to create high molecular weight products. These products pre- cipitate from solution due to their low solubility. The method presents a possible alternative for the treatment of industrial wastewaters when conventional methods such as biological treatment, activated carbon and ad- vanced oxidation may be ineective due to the nature of the wastewater stream (Aitken, 1993; Nicell et al., 1993). While a number of peroxidase enzymes obtained from plant, animal and microbial sources have been investigated for their ability to catalyze the removal of aromatic compounds from wastewaters, the majority of studies have focused on using horseradish peroxidase (HRP). HRP is commercially available in purified form and has a proven ability to remove a variety of phenolic contaminants from aqueous solution over relatively wide ranges of pH and temperature (Nicell et al., 1993). However, one of the major challenges associated with peroxidase catalyzed phenol removal is the prohibitive cost of the enzyme (Cooper and Nicell, 1996). Enzyme costs are significant because the enzyme is susceptible to inactivation by various side reactions of the treatment process (Buchanan and Nicell, 1997). While recent work has shown that enzyme inactiva- tion can be reduced using chemical additives (Nakamoto and Machida, 1991), the problem of enzyme cost could be circumvented by using a less expensive source of enzyme. In 1991, the seed coat of the soybean was identified as a rich source of a single peroxidase isoen- zyme with a molecular weight of 37,000 Daltons (Gilli- kin and Graham, 1991). Since the seed coat of the soybean is a byproduct of the soybean food industry, soybean peroxidase (SBP) has the potential of being a cost eective alternative to HRP for wastewater treat- ment. Therefore, the objectives of this study were to: (1) characterize SBP in terms of its catalytic activity and susceptibility to inactivation; and (2) compare the Bioresource Technology 70 (1999) 69–79 * Corresponding author. E-mail: [email protected]. 0960-8524/99/$ – see front matter Ó 1999 Elsevier Science Ltd. All rights reserved. PII: S 0 9 6 0 - 8 5 2 4 ( 9 9 ) 0 0 0 0 7 - 3
-
Upload
harold-wright -
Category
Documents
-
view
213 -
download
0
Transcript of Characterization of soybean peroxidase for the treatment of aqueous phenols

Characterization of soybean peroxidase for the treatment of aqueousphenols
Harold Wright, James A. Nicell*
Department of Civil Engineering & Applied Mechanics, McGill University, 817 Sherbrooke Street West, Montreal PQ,Canada H3A 2K6
Received 24 July 1998; revised 12 January 1999; accepted 18 January 1999
Abstract
The application of soybean peroxidase (SBP) to catalyze the polymerization and precipitation of aqueous phenols by hydrogen
peroxide is potentially promising because this peroxidase is less expensive than horseradish peroxidase (HRP), which has been the
focus of most wastewater research. SBP can act on a broad range of compounds and retains its catalytic ability over wide ranges of
temperature and pH. Activity was optimal at pH 6.4, with signi®cant activity observed between pH 3 and 9. SBP was very stable at
25°C at neutral and alkaline conditions but experienced rapid inactivation below pH 3. SBP underwent biphasic inactivation by
hydrogen peroxide in the absence of a reductant substrate. SBP was most e�ective when used to treat phenolic solutions between pH
6 and 9. In comparison with HRP, the activity of SBP was only slightly more sensitive to pH, was more stable at elevated tem-
peratures, and was less susceptible to permanent inactivation by hydrogen peroxide. However, SBP was catalytically slower than
HRP and a larger molar quantity of SBP was usually required to remove a given quantity of phenolic substrate. Ó 1999 Elsevier
Science Ltd. All rights reserved.
Keywords: Soybean peroxidase; Wastewater treatment; Inactivation; Stability; Temperature; pH
1. Introduction
The catalytic removal of phenolic and other aromaticcompounds from wastewaters using peroxidase enzymeand hydrogen peroxide has been the focus of extensiveresearch since the initial work of Klibanov et al. (1980).Treatment involves the catalytic oxidation of the aro-matic contaminants by hydrogen peroxide to form ar-omatic free radicals that subsequently combine to createhigh molecular weight products. These products pre-cipitate from solution due to their low solubility. Themethod presents a possible alternative for the treatmentof industrial wastewaters when conventional methodssuch as biological treatment, activated carbon and ad-vanced oxidation may be ine�ective due to the nature ofthe wastewater stream (Aitken, 1993; Nicell et al., 1993).
While a number of peroxidase enzymes obtainedfrom plant, animal and microbial sources have beeninvestigated for their ability to catalyze the removal ofaromatic compounds from wastewaters, the majority ofstudies have focused on using horseradish peroxidase
(HRP). HRP is commercially available in puri®ed formand has a proven ability to remove a variety of phenoliccontaminants from aqueous solution over relativelywide ranges of pH and temperature (Nicell et al., 1993).However, one of the major challenges associated withperoxidase catalyzed phenol removal is the prohibitivecost of the enzyme (Cooper and Nicell, 1996). Enzymecosts are signi®cant because the enzyme is susceptible toinactivation by various side reactions of the treatmentprocess (Buchanan and Nicell, 1997).
While recent work has shown that enzyme inactiva-tion can be reduced using chemical additives (Nakamotoand Machida, 1991), the problem of enzyme cost couldbe circumvented by using a less expensive source ofenzyme. In 1991, the seed coat of the soybean wasidenti®ed as a rich source of a single peroxidase isoen-zyme with a molecular weight of 37,000 Daltons (Gilli-kin and Graham, 1991). Since the seed coat of thesoybean is a byproduct of the soybean food industry,soybean peroxidase (SBP) has the potential of being acost e�ective alternative to HRP for wastewater treat-ment. Therefore, the objectives of this study were to:(1) characterize SBP in terms of its catalytic activity andsusceptibility to inactivation; and (2) compare the
Bioresource Technology 70 (1999) 69±79
* Corresponding author. E-mail: [email protected].
0960-8524/99/$ ± see front matter Ó 1999 Elsevier Science Ltd. All rights reserved.
PII: S 0 9 6 0 - 8 5 2 4 ( 9 9 ) 0 0 0 0 7 - 3

performance of SBP with HRP to assess its potentialapplication to the treatment of industrial wastewaterscontaining phenolic contaminants.
2. Methods
Medium purity soybean peroxidase, with a nominalactivity of 50 PPU/mg based on a purpurogallin assay,was purchased as a dry powder from Enzymol Inter-national, Columbus, OH. Phenolic compounds were atleast 98% pure and were purchased from either Fluka,Ronkonkoma, NY or Aldrich, Milwaukee, WI. ACSgrade 30% H2O2 w/v was purchased from BDH Chem-icals, Toronto, Canada. ACS grade potassium ferricya-nide and 98% pure 4-aminoantipyrine (4-AAP) werepurchased from Fisher Scienti®c, Fair Lawn, NJ andAldrich, respectively. ACS grade conjugate acids andbases were obtained from BDH Chemicals and wereused to prepare bu�ers in accordance with the methodsof Gomori (1955). Except for SBP solutions, all aqueoussolutions were prepared using deionized water obtainedfrom a D4741 Nanopure Ultrapure Water System(Barstead/Thermolyne, USA). Aqueous solutions ofSBP were prepared using either deionized water (ad-justed to an alkaline pH using concentrated NaOH) orpH bu�er. SBP powder and all aqueous solutions werestored at 4°C until the time of use.
All spectrophotometric measurements were perform-ed in a 1 cm cuvette using a Beckman DU-65 Spectro-photometer. Reaction temperatures were maintainedusing a constant temperature room and a Haake A81Temperature Bath. Phenolic polymers generated duringphenol removal reactions were separated from the re-action mixtures by centrifugation at 3000 g in a Centra-8Centrifuge (International Equipment, USA). ReactionpH and temperature were measured using a Radiome-ter-Copenhagen Ion 83 Meter.
SBP activity was measured by a colorimetric assay inwhich the 1 ml assay volume was composed of 500 ll of20 mM phenol, 250 ll of 9.6 mM AAP, 100 ll of 2 mMhydrogen peroxide (H2O2), 50±150 ll of sample, and 0±100 ll of 0.1 M monobasic±dibasic phosphate bu�er(pH 7.4) prepared according to the method of Gomori(1955). All assay reactants were prepared in the phos-phate bu�er. Prior to signi®cant substrate depletion, therate of colour development at 510 nm is proportional tothe concentration of active SBP. The rate of colour de-velopment was converted to enzyme activity in the cu-vette using a conversion factor of 7100 Mÿ1 cmÿ1 basedon hydrogen peroxide. One unit of activity (U) is de®nedas the number of micromoles of peroxide consumed inone min at pH 7.4 and 25°C.
Based on standard activity measurements of nineaqueous solutions of SBP, the solid enzyme preparationused in this study had a speci®c activity of 12.3 � 1.1 U/
mg. A Reinheitzahl (RZ) number of 0.49 � 0.06 wascalculated based on a ratio of absorbance measured at280 nm and 403 nm. Assuming a Soret extinction coef-®cient (Dunford and Stillman, 1976) of e403 nm� 105
Mÿ1cmÿ1 is valid for ferric SBP and assuming all Soretabsorbance is attributable to active enzyme, 1 U/ml ofperoxidase activity corresponds to 0.37 � 0.02 lM ofSBP.
The e�ect of assay pH and H2O2 concentration on thecatalytic activity of SBP was investigated by varying thepH and H2O2 concentration in the activity assay andmeasuring the resulting colour generation rate at 510 nm.After colour generation, adjustment of the pH to 7.4 didnot change the peak wavelength nor absorbance signif-icantly, therefore the absorbance of the coloured prod-uct was not a signi®cant function of pH over the rangeexamined.
The stability of SBP was evaluated by incubating theenzyme at 25°C in various pH bu�ers, at elevated tem-peratures in various pH bu�ers, and at 25°C and pH 7.4with various concentrations of H2O2. SBP activity in theincubation mixture was measured over time usingthe standard activity assay and aliquots obtained fromthe incubation mixture. As the incubation progressed, 1ml aliquots were removed from the incubation mixtureand quickly cooled using an ice bath to halt the inacti-vation process. Once cooled, aliquots were removed andallowed to warm to 25°C. Repeated activity measure-ments on the aliquot gave reproducible measurements.When addition of the aliquot to the assay a�ected theassay H2O2 concentration and pH, the measured activitywas adjusted to the expected activity under standardconditions using the experimentally determined depen-dence of peroxidase activity on assay H2O2 concentra-tion and pH. The method is described in detail in Nicelland Wright (1997).
Three-hour stirred batch reactions involving 1 mMphenolic compound, 2 mM H2O2 and various doses ofSBP reacting at 25°C in various pH bu�ers were used toevaluate the ability of SBP to catalyze phenol removalfrom wastewaters. In all batch reactions, concentratedaqueous SBP and phenol were added to 10 ml of pHbu�er. The reaction was initiated by the addition ofconcentrated H2O2. After 3 h, the precipitated productswere removed by centrifugation at 3000 g for 30 min andthe residual phenol in the supernatant was measured. Areaction time of 3 h was su�cient to ensure that all re-actions had gone to completion over the range of pHsstudied. Reaction blanks were used at each reaction pHto con®rm that phenolic compound removal was solelya consequence of SBP activity.
Phenolic compound concentrations were measuredusing a colorimetric assay consisting of 100 ll of phenolsample, 50 ll of 41.6 mM 4-AAP, 800 ll of 0.15 Msodium borate, and 50 ll of 166.8 mM potassiumferricyanide. When the extent of the assay reaction is
70 H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79
limited only by the concentration of the phenolic com-pound, the quantity of colour formed under alkalineconditions at 510 nm is proportional to the initial con-centration of phenolic compound in the assay. The so-dium borate in the assay maintained the mixture at a®xed alkaline pH such that molar absorptivities mea-sured for various phenolic compounds were relativelyinsensitive to sample pH.
Since phenolic compounds have absorbance peaks inthe UV range, the phenolic concentrations in a batchreactions were also estimated by measuring the UVabsorbance peak of the solution and then calculatingconcentration using an appropriate extinction coe�-cient. Molar extinction coe�cients were evaluated foreach phenolic compound and for each pH.
An end-point assay, based on the AAP/phenol systemused in the activity assay, was used to determine thehydrogen peroxide concentration. The concentration ofperoxidase in the microcuvette was approximately 100mg/l, while those of the other reactants (except peroxide)were as cited above for the peroxidase assay. The extentof color development at 510 nm was proportional tohydrogen peroxide concentration, provided this con-centration did not exceed 0.05 mM in the cuvette. Aftera 10-minute reaction time, the absorbance at 510 nmwas converted to peroxide concentration using a cali-bration curve.
3. Results and discussion
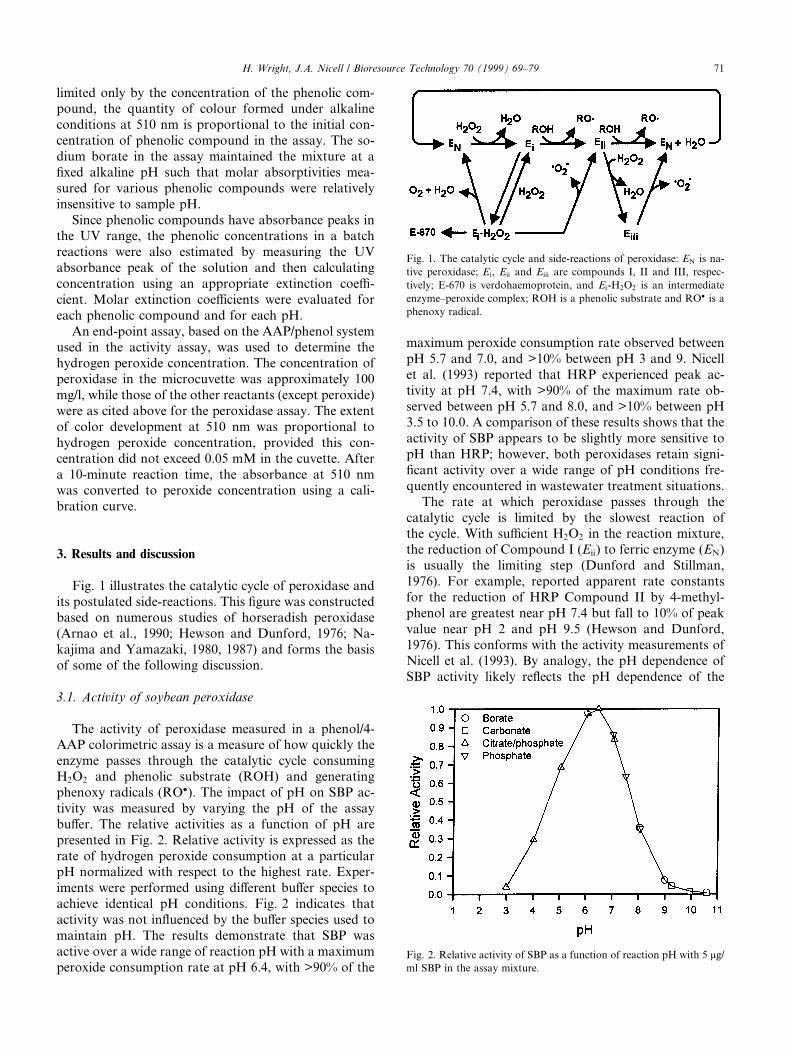
Fig. 1 illustrates the catalytic cycle of peroxidase andits postulated side-reactions. This ®gure was constructedbased on numerous studies of horseradish peroxidase(Arnao et al., 1990; Hewson and Dunford, 1976; Na-kajima and Yamazaki, 1980, 1987) and forms the basisof some of the following discussion.
3.1. Activity of soybean peroxidase
The activity of peroxidase measured in a phenol/4-AAP colorimetric assay is a measure of how quickly theenzyme passes through the catalytic cycle consumingH2O2 and phenolic substrate (ROH) and generatingphenoxy radicals (RO�). The impact of pH on SBP ac-tivity was measured by varying the pH of the assaybu�er. The relative activities as a function of pH arepresented in Fig. 2. Relative activity is expressed as therate of hydrogen peroxide consumption at a particularpH normalized with respect to the highest rate. Exper-iments were performed using di�erent bu�er species toachieve identical pH conditions. Fig. 2 indicates thatactivity was not in¯uenced by the bu�er species used tomaintain pH. The results demonstrate that SBP wasactive over a wide range of reaction pH with a maximumperoxide consumption rate at pH 6.4, with >90% of the
maximum peroxide consumption rate observed betweenpH 5.7 and 7.0, and >10% between pH 3 and 9. Nicellet al. (1993) reported that HRP experienced peak ac-tivity at pH 7.4, with >90% of the maximum rate ob-served between pH 5.7 and 8.0, and >10% between pH3.5 to 10.0. A comparison of these results shows that theactivity of SBP appears to be slightly more sensitive topH than HRP; however, both peroxidases retain signi-®cant activity over a wide range of pH conditions fre-quently encountered in wastewater treatment situations.
The rate at which peroxidase passes through thecatalytic cycle is limited by the slowest reaction ofthe cycle. With su�cient H2O2 in the reaction mixture,the reduction of Compound I (Eii) to ferric enzyme (EN)is usually the limiting step (Dunford and Stillman,1976). For example, reported apparent rate constantsfor the reduction of HRP Compound II by 4-methyl-phenol are greatest near pH 7.4 but fall to 10% of peakvalue near pH 2 and pH 9.5 (Hewson and Dunford,1976). This conforms with the activity measurements ofNicell et al. (1993). By analogy, the pH dependence ofSBP activity likely re¯ects the pH dependence of the
Fig. 1. The catalytic cycle and side-reactions of peroxidase: EN is na-
tive peroxidase; Ei, Eii and Eiii are compounds I, II and III, respec-
tively; E-670 is verdohaemoprotein, and Ei-H2O2 is an intermediate
enzyme±peroxide complex; ROH is a phenolic substrate and RO� is a
phenoxy radical.
Fig. 2. Relative activity of SBP as a function of reaction pH with 5 lg/
ml SBP in the assay mixture.
H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79 71

apparent rate constants describing the reactions of SBPCompound II.
The rate of colour generation at pH 7.4 and 25°C wasinvestigated as a function of the assay H2O2 concentra-tion. The resulting reaction rates, expressed as peroxideconsumption rates, are presented in Fig. 3 as a functionof initial H2O2 concentration for three concentrations ofSBP. The maximum activity was consistently observed at0.33 mM H2O2. At low H2O2 concentrations, the per-oxidase cycle is limited by the rate at which ferric per-oxidase is oxidized by H2O2 to Compound I. At higherH2O2 concentrations, the peroxidase cycle is limited bythe formation of inactive Compound III (Eiii). This is nota permanently inactivated form of peroxidase since it canrevert to the ferric state and reenter the catalytic cycle, asshown in Fig. 1; however, the return is very slow and anyaccumulation in this state represents a loss in catalytice�ciency. Nicell and Wright (1997) developed a kineticmodel of this assay and extracted kinetic constants todescribe the rate at which peroxidase passes through itscatalytic cycle and its susceptibility to Compound IIIformation. A comparison of the rate constants for HRPand SBP revealed that SBP is more than an order ofmagnitude slower than HRP during the oxidation ofphenol and SBP is far more susceptible to inactivationthrough Compound III formation than HRP (Nicell andWright, 1997).
3.2. Stability of soybean peroxidase
3.2.1. E�ect of pHThe stability of SBP incubated at 25°C as a function
of pH is presented in Fig. 4. Since activity was measuredusing the standard activity recipe, the addition of analiquot of incubation mixture to the assay caused anotable pH shift that in¯uenced the rate of colour for-mation. Accordingly, the pH of the assay mixture was
measured and the activity corrected to an equivalentvalue at pH 7.4 using the results presented in Fig. 2.
In acidic bu�er, SBP undergoes time dependent in-activation whose rate and extent depend on the pH ofthe incubation mixture. At pH 2 and 3, ®rst order in-activation led to essentially zero activity within two andfour days, respectively. Inactivation at pH 5 was bi-phasic and did not result in complete inactivation. Incomparison, SBP incubated in either neutral or alkalinebu�er was essentially stable for over 20 days of incu-bation.
3.2.2. Thermal inactivationThe stability of SBP when it was incubated at ap-
proximately 80°C in various pH bu�ers is presented inFig. 5. The thermal inactivation depended on incuba-
Fig. 3. Rate of H2O2 consumption in the activity assay as a function of
H2O2 and SBP concentration. Smooth curves are based on a ®t of the
model of Nicell and Wright (1997).
Fig. 4. Stability of SBP incubated at 25°C in various pH bu�ers (pH
2.0 HCl/KCl bu�er, pH 3.0 citrate/phosphate bu�er, pH 5.0 citrate/
phosphate bu�er, pH 7.0 phosphate bu�er, pH 9.0 borate bu�er, pH
10.7 carbonate bu�er).
Fig. 5. Stability of SBP incubated at 80.3 � 0.4°C in various bu�ers
(pH 5.0 citrate/phosphate bu�er, pH 6.0 phosphate bu�er, pH 7.0
phosphate bu�er, pH 8.0 phosphate bu�er, pH 9.0 borate bu�er, pH
10.3 carbonate bu�er).
72 H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79

tion pH and could be modelled using the ®rst orderequation:
A�t� � A0eÿkt � A010ÿtD; �1�
where A(t) is the activity at time t, A0 is the initial ac-tivity, k is the inactivation decay constant, and D is theinactivation decimal reduction value (D-value). Fig. 6presents D-values for SBP thermal inactivation as afunction of incubation temperature and pH. The D-value at 70°C and pH 6.0 could not be measured due tothe SBP's extreme thermal stability under these condi-tions. Consistently, SBP was most resistant to thermalinactivation at approximately pH 6.
Inactivation decay constants may be related to incu-bation temperature using the Arrhenius Law and anactivation energy (De Cordt et al., 1992). D-values overa limited temperature range may be related to temper-ature using:
D � DREF10�TREFÿT �
z �2�where DREF is the decimal reduction value at tempera-ture TREF and Z is the temperature change required toobtain a ten-fold increase or decrease in the decimalreduction value (De Cordt et al., 1992). Unlike inacti-
vation decay constants and D-values, the activationenergies and Z-values obtained from the data in Fig. 6tend to be independent of pH with average values of234 � 13 kJ/mol and 10.2 � 0.6°C, respectively.
The thermal inactivation of SBP observed in thiswork is compared to observations reported in the liter-ature in Table 1. Using SBP which was puri®ed usingdialysis, fractionation and chromatography, Sessa andAnderson (1981) and Chuang and Chen (1988) observedconsiderably faster thermal inactivation of SBP. Sinceenzyme thermal inactivation is in¯uenced by a multitudeof factors such as ionic strength and bu�er species (DeCordt et al., 1992), calcium ion concentration (EnzymolInternational Inc., 1994), and polyhydric alcohol andsugar concentration (Asther and Meunier, 1990), dif-ferences between the rates of thermal inactivation re-ported in the literature and the rates observed in thiswork may be due to di�erences in the constituents of theincubation mixture which are a result of the puri®cationtechniques used to extract SBP from the original plantmaterial. The Z-value observed in this work compareswell with the value reported by Chuang and Chen(1988). A pH optimum for peroxidase thermal stabilityhas been previously reported with Japanese radish per-oxidase (Tamura and Morita, 1975) and pH indepen-dent Z-values have been observed with non-peroxidaseenzymes (De Cordt et al., 1992). While a 3±4 minD-value has been observed with HRP incubated at 72°Cand pH 7.4 (from the data of Nicell et al., 1993), SBPincubated under similar conditions in this work exhib-ited a D-value of several days thereby indicating a muchgreater stability at elevated temperatures.
3.2.3. Inactivation by hydrogen peroxideThe stability of SBP incubated at 25°C in pH 7.4
bu�er with various concentrations of H2O2 is presentedin Fig. 7. Since the concentration of H2O2 within theassay was increased by the addition of an aliquot fromthe incubation mixture, the measured rate of colourgeneration was corrected to standard conditions usingthe curves of Fig. 3 and the method described by Nicell
Fig. 6. SBP decimal reduction values as a function of temperature and
pH.
Table 1
Thermal inactivation parameters of SBP
Enzyme source pH and bu�er species Temperature (°C) D (min) Z (°C) Reference
Composite of puri®ed
isoenzymes
pH 5.5 (0.1 M citrate/
phosphate bu�er)
70 60 ÿ Sessa and Anderson
(1981)
Puri®ed 37 000 Dalton
isoenzyme
pH 7.0 (0.2 M phosphate
bu�er)
70 103 8 Chuang and Chen
(1988)80 9.1
90 0.38
Enzymol medium purity
SBP
pH 5.0 (0.1 M citrate/
phosphate bu�er)
70.2 1450 10.2 � 0.6 This work
pH 7.0 (0.1 M phosphate
bu�er)
70.2 4000
80.3 300
90.8 30
H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79 73

and Wright (1997). This correction was applied assum-ing that the catalytic consumption of H2O2 in the in-cubation mixture was negligible. However, it should benoted that after 80 minutes of incubation with initialH2O2 concentrations of 0.02, 0.2, 2.0, and 20 mM, theconcentration of H2O2 in the incubation mixtures fell by62, 50, 30 and 20% of its initial value, respectively. Thisreduction in H2O2 may result in a worst-case activityoverestimation of 6.6% and 1.4% after 80 min of incu-bation with initial H2O2 concentrations of 20 mM and2.0 mM, respectively.
When peroxidase and H2O2 are incubated togetherwithout a phenolic reducing agent present, H2O2 acts toreduce the +5 oxidation state of the enzyme in either oneor two electron steps (Nakajima and Yamazaki, 1987).Arnao et al. (1990) reported that H2O2 also reacts withHRP resulting in an inactive form referred to as ve-rdohemoprotein (see E-670 in Fig. 1). While the exactmechanism of verdohemoprotein formation is uncertainand may involve a sequence of reversible and irreversiblereactions (Nakajima and Yamazaki, 1980), Arnao et al.(1990) proposed the scheme shown in Fig. 1 for itsformation.
SBP inactivation over time presented in Fig. 7 in-creased with H2O2 concentration and was biphasic. Thebiphasic inactivation observed with SBP can be ex-
plained based on the relative magnitudes of kineticconstants published for HRP reacting with H2O2 (Ar-nao et al., 1990). Since the reaction of ferric peroxidasewith H2O2 occurs quickly (Yamazaki and Nakajima,1986), ferric SBP is expected to oxidize to Compound Ivirtually immediately following the addition of H2O2 tothe incubation mixture. Since the reduction of Com-pound I by H2O2 occurs relatively slowly (Arnao et al.,1990), the rate of verdohemoprotein formation at thispoint can be expected to occur at its fastest rate as-suming its formation originates at the Compound Istate. As the incubation proceeds, the majority of SBP istransformed to Compound II and Compound III at theexpense of Compound I and the rate of verdohemo-protein formation slows. However, spontaneous de-composition of Compound III to ferric SBP ensuresthat a residual level of Compound I is maintained. Thus,the ®rst phase of the biphasic inactivation re¯ects thetransformation of active enzyme to Compound III at theexpense of Compound I thereby slowing verdohemo-protein formation. The second phase re¯ects the loss ofactive enzyme to verdohemoprotein due to the presenceof small quantities of Compound I. In addition, thegradual catalytic consumption of H2O2 simultaneouslyacts to slow compound III and verdohemoprotein for-mation.
Arnao et al. (1990) modeled the biphasic inactivationof HRP incubated with H2O2 by deriving two eigen-values from a ®rst-order linear di�erential equationapproximation of the reacting system. The two eigen-values, k1 and k2, were estimated from the inactivationdata by ®tting the equation:
A�t� � A1eÿk1t � A2eÿk2t �3�Arnao et al. (1990) used these eigenvalues to estimatekinetic constants for verdohemoprotein formation andCompound I reduction by H2O2.
The curves drawn through the SBP inactivation datapresented in Fig. 7 were obtained by ®tting Eq. (3) usingregression and the method of Arnao et al. (1990). Theparameters of the curve ®t of SBP data for comparisonwith similar data published for HRP are presented inTable 2. While Eq. (3) provided for an excellent ®t tothe data in Fig. 7, the consumption of H2O2 mentionedabove is so signi®cant as to invalidate treating the re-acting system as a steady-state system that can be
Fig. 7. Stability of 0.37 U/ml SBP at 25°C as a function of time and
initial concentration of hydrogen peroxide in pH 7.4 phosphate bu�er.
Smooth curlves are based on Eq. (3) and parameters summarized in
Table 2.
Table 2
Summary of parameters describing the time-dependant inactivation of SBP and HRP by H2O2 without the presence of a phenolic substrate
Initial [H2O2] (mM) SBP ± This work HRP±Arnao et al. (1990)
A1 (U/mL) k1 (minÿ1) A2 (U/mL) k2 (minÿ1) k1 (minÿ1) k2 (minÿ1)
0.02 ÿ ÿ 0.361 0.00084 ÿ ÿ0.2 0.160 0.0324 0.235 0.00082 0.159 0.0104
2 0.176 0.0353 0.198 0.00133 0.357 0.0314
20 0.136 0.0512 0.173 0.00632 0.695 0.0379
74 H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79
modelled using ®rst-order linear di�erential equations,as was done by Arnao et al. (1990). Therefore, the termsk1 and k2 cannot be treated as eigenvalues and kineticconstants cannot be evaluated.
Due to the similarity between these experiments andthose of Arnao et al. (1990), it is very likely that H2O2
consumption was also signi®cant in their experiments.For this reason, a direct comparison of kinetic coe�-cients of SBP and HRP cannot be made. However, acomparison of the parameters presented in Table 2suggests that with the same initial H2O2 concentration inthe incubation mixture, inactivation observed over timewill be greater with HRP than with SBP. The resultsimply that either the formation of verdohemoproteinoccurs at a faster rate with HRP, or SBP tends to par-tition more as Compound II and/or Compound IIIthereby protecting the enzyme from permanent inacti-vation. The suggestion that SBP tends to partition moreas Compound III is supported by the kinetic model ofNicell and Wright (1997) which quanti®es the inhibitionof peroxidase by hydrogen peroxide.
3.3. Phenolic compound removal
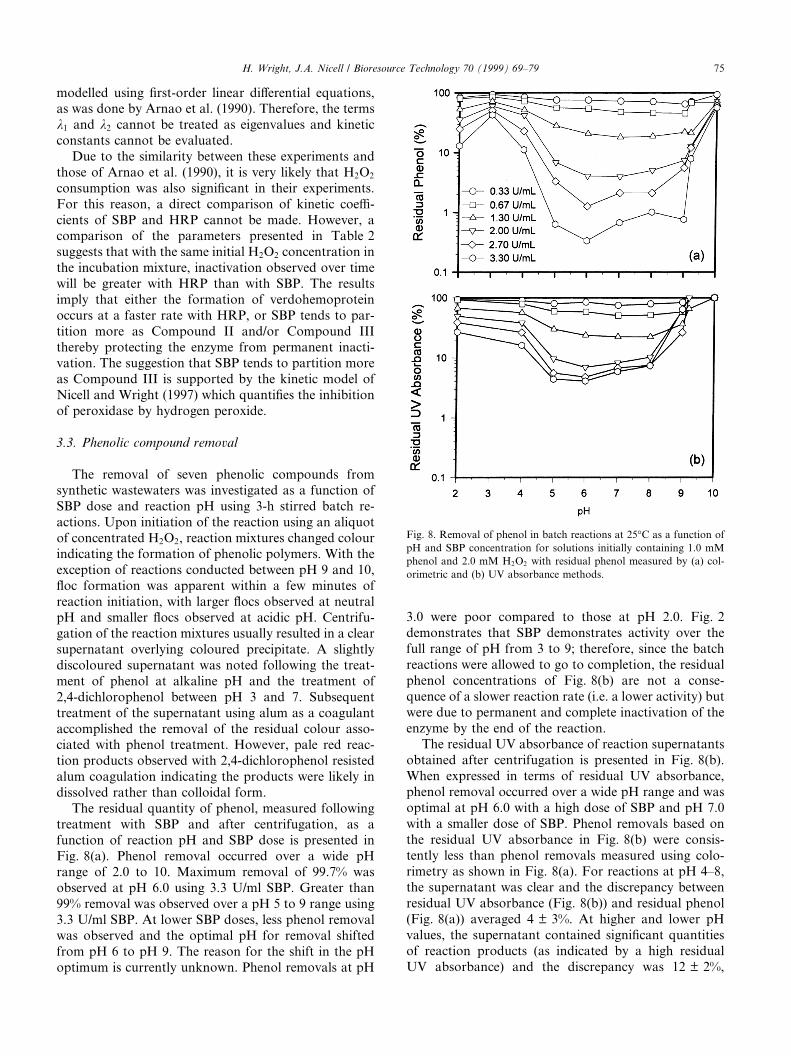
The removal of seven phenolic compounds fromsynthetic wastewaters was investigated as a function ofSBP dose and reaction pH using 3-h stirred batch re-actions. Upon initiation of the reaction using an aliquotof concentrated H2O2, reaction mixtures changed colourindicating the formation of phenolic polymers. With theexception of reactions conducted between pH 9 and 10,¯oc formation was apparent within a few minutes ofreaction initiation, with larger ¯ocs observed at neutralpH and smaller ¯ocs observed at acidic pH. Centrifu-gation of the reaction mixtures usually resulted in a clearsupernatant overlying coloured precipitate. A slightlydiscoloured supernatant was noted following the treat-ment of phenol at alkaline pH and the treatment of2,4-dichlorophenol between pH 3 and 7. Subsequenttreatment of the supernatant using alum as a coagulantaccomplished the removal of the residual colour asso-ciated with phenol treatment. However, pale red reac-tion products observed with 2,4-dichlorophenol resistedalum coagulation indicating the products were likely indissolved rather than colloidal form.
The residual quantity of phenol, measured followingtreatment with SBP and after centrifugation, as afunction of reaction pH and SBP dose is presented inFig. 8(a). Phenol removal occurred over a wide pHrange of 2.0 to 10. Maximum removal of 99.7% wasobserved at pH 6.0 using 3.3 U/ml SBP. Greater than99% removal was observed over a pH 5 to 9 range using3.3 U/ml SBP. At lower SBP doses, less phenol removalwas observed and the optimal pH for removal shiftedfrom pH 6 to pH 9. The reason for the shift in the pHoptimum is currently unknown. Phenol removals at pH
3.0 were poor compared to those at pH 2.0. Fig. 2demonstrates that SBP demonstrates activity over thefull range of pH from 3 to 9; therefore, since the batchreactions were allowed to go to completion, the residualphenol concentrations of Fig. 8(b) are not a conse-quence of a slower reaction rate (i.e. a lower activity) butwere due to permanent and complete inactivation of theenzyme by the end of the reaction.
The residual UV absorbance of reaction supernatantsobtained after centrifugation is presented in Fig. 8(b).When expressed in terms of residual UV absorbance,phenol removal occurred over a wide pH range and wasoptimal at pH 6.0 with a high dose of SBP and pH 7.0with a smaller dose of SBP. Phenol removals based onthe residual UV absorbance in Fig. 8(b) were consis-tently less than phenol removals measured using colo-rimetry as shown in Fig. 8(a). For reactions at pH 4±8,the supernatant was clear and the discrepancy betweenresidual UV absorbance (Fig. 8(b)) and residual phenol(Fig. 8(a)) averaged 4 � 3%. At higher and lower pHvalues, the supernatant contained signi®cant quantitiesof reaction products (as indicated by a high residualUV absorbance) and the discrepancy was 12 � 2%,
Fig. 8. Removal of phenol in batch reactions at 25°C as a function of
pH and SBP concentration for solutions initially containing 1.0 mM
phenol and 2.0 mM H2O2 with residual phenol measured by (a) col-
orimetric and (b) UV absorbance methods.
H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79 75
35 � 23%, and 93 � 77% at pH 2.0, 9.0 and 9.2,respectively. It is concluded that for pH <4 and pH >8there is a tendency to form signi®cantly greater quanti-ties of soluble reaction products.
While the UV absorbance of residual SBP may ex-plain a fraction of the discrepancies observed betweenthe UV absorbance and colorimetric phenol measure-ments, it is not the most signi®cant factor. Rather, thehigher absorbance values obtained using the UV ab-sorbance method can most likely be attributed to solubleand colloidal reaction products that resist removal bycentrifugation. Zou and Taylor (1994) reported that 0.40lM o,o0-biphenol, 0.083 lM p-phenoxyphenol and 0.90lM p,p0-biphenol remained as soluble byproducts aftertreatment of 1.0 mM phenol at pH 7.4 using excessHRP. Yu et al. (1994) also observed o,o0-biphenol, p-phenoxyphenol and p,p0-biphenol in the precipitatephase after phenol treatment with HRP at neutral pH.Thus it appears that a percentage of the original phenolmay be transformed to soluble reaction products inequilibrium with a solid precipitate phase. Furthermore,while these soluble reaction products may undergo fur-ther enzyme catalyzed oxidation (Yu et al., 1994) andcombine to form less soluble reaction products, theequilibrium with the solid phase will ensure that someresidual levels of these products are maintained. Solublearomatic reaction products at the concentrations ob-served by Zou and Taylor (1994) would only require UVmolar extinction coe�cients an order of magnitudelarger than the coe�cients for phenol in order to explainthe discrepancies observed between the UV and colori-metric phenol measuring methods.
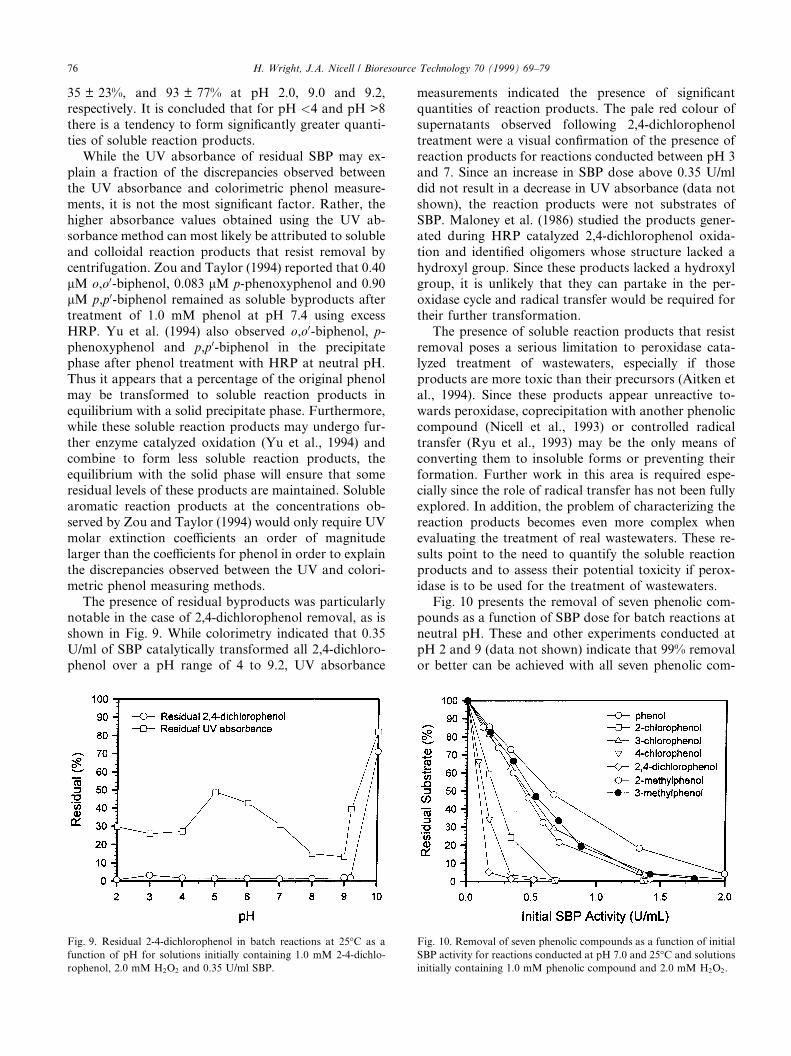
The presence of residual byproducts was particularlynotable in the case of 2,4-dichlorophenol removal, as isshown in Fig. 9. While colorimetry indicated that 0.35U/ml of SBP catalytically transformed all 2,4-dichloro-phenol over a pH range of 4 to 9.2, UV absorbance
measurements indicated the presence of signi®cantquantities of reaction products. The pale red colour ofsupernatants observed following 2,4-dichlorophenoltreatment were a visual con®rmation of the presence ofreaction products for reactions conducted between pH 3and 7. Since an increase in SBP dose above 0.35 U/mldid not result in a decrease in UV absorbance (data notshown), the reaction products were not substrates ofSBP. Maloney et al. (1986) studied the products gener-ated during HRP catalyzed 2,4-dichlorophenol oxida-tion and identi®ed oligomers whose structure lacked ahydroxyl group. Since these products lacked a hydroxylgroup, it is unlikely that they can partake in the per-oxidase cycle and radical transfer would be required fortheir further transformation.
The presence of soluble reaction products that resistremoval poses a serious limitation to peroxidase cata-lyzed treatment of wastewaters, especially if thoseproducts are more toxic than their precursors (Aitken etal., 1994). Since these products appear unreactive to-wards peroxidase, coprecipitation with another phenoliccompound (Nicell et al., 1993) or controlled radicaltransfer (Ryu et al., 1993) may be the only means ofconverting them to insoluble forms or preventing theirformation. Further work in this area is required espe-cially since the role of radical transfer has not been fullyexplored. In addition, the problem of characterizing thereaction products becomes even more complex whenevaluating the treatment of real wastewaters. These re-sults point to the need to quantify the soluble reactionproducts and to assess their potential toxicity if perox-idase is to be used for the treatment of wastewaters.
Fig. 10 presents the removal of seven phenolic com-pounds as a function of SBP dose for batch reactions atneutral pH. These and other experiments conducted atpH 2 and 9 (data not shown) indicate that 99% removalor better can be achieved with all seven phenolic com-
Fig. 9. Residual 2-4-dichlorophenol in batch reactions at 25°C as a
function of pH for solutions initially containing 1.0 mM 2-4-dichlo-
rophenol, 2.0 mM H2O2 and 0.35 U/ml SBP.
Fig. 10. Removal of seven phenolic compounds as a function of initial
SBP activity for reactions conducted at pH 7.0 and 25°C and solutions
initially containing 1.0 mM phenolic compound and 2.0 mM H2O2.
76 H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79
pounds over a wide range of reaction pH given a su�-cient dose of SBP.
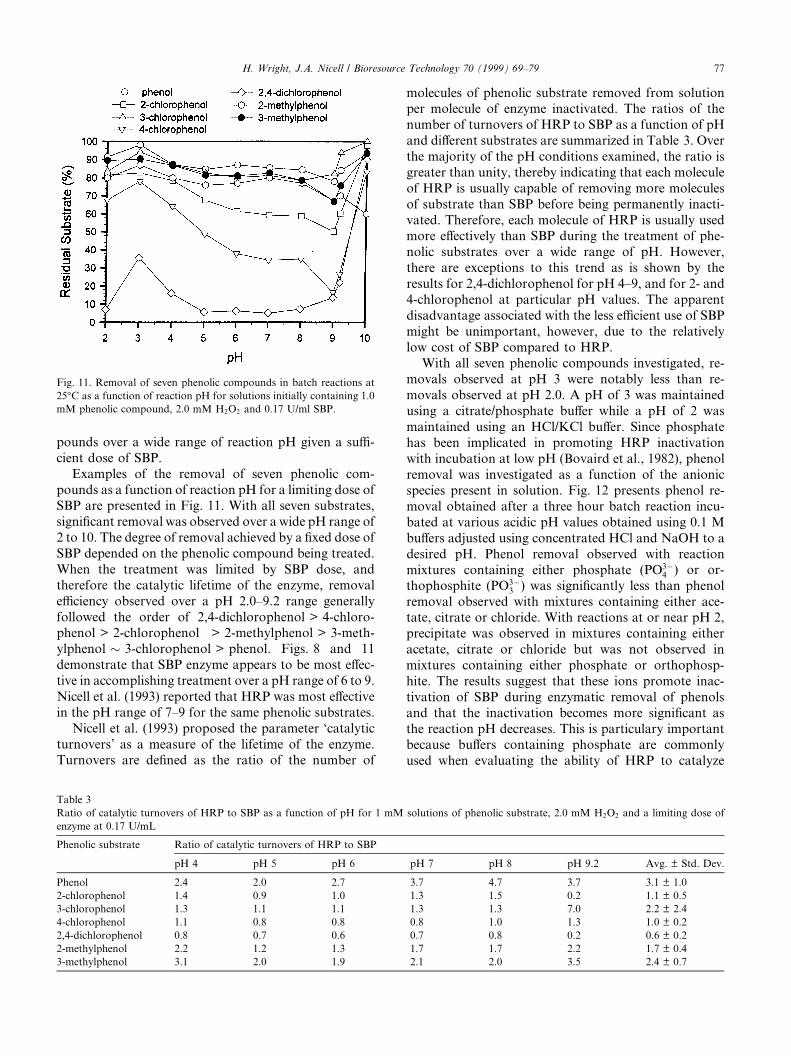
Examples of the removal of seven phenolic com-pounds as a function of reaction pH for a limiting dose ofSBP are presented in Fig. 11. With all seven substrates,signi®cant removal was observed over a wide pH range of2 to 10. The degree of removal achieved by a ®xed dose ofSBP depended on the phenolic compound being treated.When the treatment was limited by SBP dose, andtherefore the catalytic lifetime of the enzyme, removale�ciency observed over a pH 2.0±9.2 range generallyfollowed the order of 2,4-dichlorophenol > 4-chloro-phenol > 2-chlorophenol > 2-methylphenol > 3-meth-ylphenol � 3-chlorophenol > phenol. Figs. 8 and 11demonstrate that SBP enzyme appears to be most e�ec-tive in accomplishing treatment over a pH range of 6 to 9.Nicell et al. (1993) reported that HRP was most e�ectivein the pH range of 7±9 for the same phenolic substrates.
Nicell et al. (1993) proposed the parameter `catalyticturnovers' as a measure of the lifetime of the enzyme.Turnovers are de®ned as the ratio of the number of
molecules of phenolic substrate removed from solutionper molecule of enzyme inactivated. The ratios of thenumber of turnovers of HRP to SBP as a function of pHand di�erent substrates are summarized in Table 3. Overthe majority of the pH conditions examined, the ratio isgreater than unity, thereby indicating that each moleculeof HRP is usually capable of removing more moleculesof substrate than SBP before being permanently inacti-vated. Therefore, each molecule of HRP is usually usedmore e�ectively than SBP during the treatment of phe-nolic substrates over a wide range of pH. However,there are exceptions to this trend as is shown by theresults for 2,4-dichlorophenol for pH 4±9, and for 2- and4-chlorophenol at particular pH values. The apparentdisadvantage associated with the less e�cient use of SBPmight be unimportant, however, due to the relativelylow cost of SBP compared to HRP.
With all seven phenolic compounds investigated, re-movals observed at pH 3 were notably less than re-movals observed at pH 2.0. A pH of 3 was maintainedusing a citrate/phosphate bu�er while a pH of 2 wasmaintained using an HCl/KCl bu�er. Since phosphatehas been implicated in promoting HRP inactivationwith incubation at low pH (Bovaird et al., 1982), phenolremoval was investigated as a function of the anionicspecies present in solution. Fig. 12 presents phenol re-moval obtained after a three hour batch reaction incu-bated at various acidic pH values obtained using 0.1 Mbu�ers adjusted using concentrated HCl and NaOH to adesired pH. Phenol removal observed with reactionmixtures containing either phosphate (PO3ÿ
4 ) or or-thophosphite (PO3ÿ
3 ) was signi®cantly less than phenolremoval observed with mixtures containing either ace-tate, citrate or chloride. With reactions at or near pH 2,precipitate was observed in mixtures containing eitheracetate, citrate or chloride but was not observed inmixtures containing either phosphate or orthophosp-hite. The results suggest that these ions promote inac-tivation of SBP during enzymatic removal of phenolsand that the inactivation becomes more signi®cant asthe reaction pH decreases. This is particulary importantbecause bu�ers containing phosphate are commonlyused when evaluating the ability of HRP to catalyze
Fig. 11. Removal of seven phenolic compounds in batch reactions at
25°C as a function of reaction pH for solutions initially containing 1.0
mM phenolic compound, 2.0 mM H2O2 and 0.17 U/ml SBP.
Table 3
Ratio of catalytic turnovers of HRP to SBP as a function of pH for 1 mM solutions of phenolic substrate, 2.0 mM H2O2 and a limiting dose of
enzyme at 0.17 U/mL
Phenolic substrate Ratio of catalytic turnovers of HRP to SBP
pH 4 pH 5 pH 6 pH 7 pH 8 pH 9.2 Avg. � Std. Dev.
Phenol 2.4 2.0 2.7 3.7 4.7 3.7 3.1 � 1.0
2-chlorophenol 1.4 0.9 1.0 1.3 1.5 0.2 1.1 � 0.5
3-chlorophenol 1.3 1.1 1.1 1.3 1.3 7.0 2.2 � 2.4
4-chlorophenol 1.1 0.8 0.8 0.8 1.0 1.3 1.0 � 0.2
2,4-dichlorophenol 0.8 0.7 0.6 0.7 0.8 0.2 0.6 � 0.2
2-methylphenol 2.2 1.2 1.3 1.7 1.7 2.2 1.7 � 0.4
3-methylphenol 3.1 2.0 1.9 2.1 2.0 3.5 2.4 � 0.7
H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79 77

phenolic compound removal from solution under acidicconditions (Klibanov et al., 1980; Dec and Bollag,1990). In future work, the use of acidic bu�ers (pH < 5)containing phosphate or orthophosphite should beavoided in order to prevent inaccuracies associated withinactivation of the enzyme by these ionic species.
4. Conclusions
SBP has demonstrated its ability to catalyze the po-lymerization and precipitation of phenolic compoundsfrom water. It can act on a broad range of compoundsand retains its catalytic ability over wide ranges oftemperature and pH. Its catalytic activity was optimal atpH 6.4, with more than 90% of its maximum activityobserved between pH 5.7 and 7.0, and more than 10%between pH 3 and 9. SBP activity was very stable at25°C at neutral and alkaline pHs but experienced rapidinactivation below pH 3. It was most thermally stable atapproximately pH 6. SBP underwent biphasic inactiva-tion by hydrogen peroxide in the absence of a reductantsubstrate. SBP was most e�ective when used to treatphenolic solutions between pH 6 and 9.
In comparison with HRP, the activity of SBP wasonly slightly more sensitive to pH, was signi®cantlymore stable at elevated temperatures, and was less sus-ceptible to permanent inactivation by hydrogen perox-ide. However, SBP was catalytically slower than HRPand a larger molar quantity of SBP was usually requiredto remove a given quantity of phenolic substrate. BothSBP and HRP were susceptible to increased inactivationin the presence of phosphate or orthophosphite ionsunder acidic conditions.
The formation of residual byproducts is a particularconcern which may limit the application of this waste-
water treatment process. Signi®cant quantities of solubleproducts were observed at extremes of pH and the ten-dency for the formation of such products was dependenton the phenol being treated. Additional research is re-quired to assess the toxicity of these products and toexplore methods to reduce their concentration in thetreated e�uents.
Acknowledgements
This work was funded by the Natural Sciences andEngineering Research Council of Canada and the``Fonds pour la formation de chercheurs et l'aide �a larecherche'' of Quebec. The technical assistance of Caro-line Korn is gratefully acknowledged.
References
Aitken, M.D., 1993. Waste treatment applications of enzymes:
opportunities and obstacles. Chem. Eng. J. 52, B49±B58.
Aitken, M.D., Massey, I.J., Chen, T., Heck, P.E., 1994. Characteriza-
tion of reaction products from the enzyme catalyzed oxidation of
phenolic pollutants. Wat. Res. 28, 1879±1889.
Arnao, M.B., Acosta, M., del Rio, J.A., Garcia-Canovas, F.,
1990. Inactivation of peroxidase by hydrogen peroxide and
its protection by a reductant agent. Biochim. Biophys. Acta 1038,
85±89.
Asther, M., Meunier, J.C., 1990. Increased thermal stability of Bacillus
licheniformis a-amylase in the presence of various additives. Enz.
Microb. Tech. 12, 902±905.
Bovaird, J.H., Ngo, T.T., Lenho�, H.M., 1982. Optimizing the o-
phenylenediamine assay for horseradish peroxidase: E�ects of
phosphate and pH, substrate and enzyme concentrations, and
stopping reagents. Clin. Chem. 28, 2423±2426.
Buchanan, I.D., Nicell, J.A., 1997. Model development for horseradish
peroxidase-catalyzed removal of aqueous phenol. Biotech. Bioeng.
54 (3), 251±261.
Chuang, W., Chen, A.O., 1988. Puri®cation and characterization of
vegetable soybean peroxidase I: Puri®cation and heat stability of
vegetable soybean peroxidase isoenzymes. Shih P'in K'o Hsueh
(Taipei) 15, 12±26 (in Chinese).
Cooper, V.A., Nicell, J.A., 1996. Removal of phenols from a foundry
wastewater using horseradish peroxidase. Wat. Res. 30 (4), 954±
964.
De Cordt, S., Vanhoof, K., Hu, J., Maesmans, G., Hendrickx, M.,
Tobback, P., 1992. Thermostability of soluble and immobilized
a-amylase from Bacillus licheniformis. Biotech. Bioeng. 40, 396±
402.
Dec, J., Bollag, J.M., 1990. Detoxi®cation of substituted phenols by
oxidoreductive enzymes through polymerization reactions. Arch.
Environ. Contam. Toxicol. 19, 543±550.
Dunford, H.B., Stillman, J.S., 1976. On the function and mechanism of
action of peroxidases. Coord. Chem. Rev. 19, 187±251.
Enzymol International Inc. 1994. Personal Communication.
Gillikin, J.W., Graham, J.S., 1991. Puri®cation and developmental
analysis of the major anionic peroxidase from the seed coat of
Glycine max. Plant Physiol. 96, 214±220.
Gomori, G., 1955. Preparation of bu�ers for use in enzyme studies. In:
Colowick, S.P., Kaplan, N.O., (Eds.), Methods in Enzymology,
vol. I, New York, Academic Press, p. 138.
Fig. 12. E�ect of pH bu�er species on phenol removal for reactions at
25°C with initial concentrations of 1.0 mM phenol, 2.0 mM H2O2 and
1.48 U/ml SBP and a bu�er concentration of 0.1 M.
78 H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79

Hewson, W.D., Dunford, H.B., 1976. Stoichiometry of the reaction
between horseradish peroxidase and p-cresol. J. Biol. Chem. 251,
6043±6052.
Klibanov, A.M., Alberti, B.N., Morris, E.D., Felshin, L.M., 1980.
Enzymatic removal of toxic phenols and anilines from wastewaters.
J. Appl. Biochem. 2, 414±421.
Maloney, S.W., Manem, J., Mallevialle, J., Fiessinger, F., 1986.
Transformation of trace organic compounds in drinking water by
enzymatic oxidative coupling. Env. Sci. Tech. 20, 249±253.
Nakajima, R., Yamazaki, I., 1987. The mechanism of oxyperoxidase
formation from ferryl peroxidase and hydrogen peroxide. J. Biol.
Chem. 262, 2576±2581.
Nakajima, R., Yamazaki, I., 1980. The conversion of horseradish
peroxidase C to a verdohemoprotein by a hydroperoxide derived
enzymatically from indole-3-acetic acid and by m-nitro-
peroxybenzoic acid. J. Biol. Chem. 255, 2067±2071.
Nakamoto, S., Machida, N., 1991. Phenol removal from aqueous
solutions by peroxidase-catalyzed reaction using additives. Wat.
Res. 26, 49±54.
Nicell, J.A., Bewtra, J.K., Biswas, N., St. Pierre, C.C., 1993. Enzyme
catalyzed polymerization and precipitation of aromatic compounds
from aqueous solution. Can. J. Civ. Eng. 20, 725±735.
Nicell, J.A., Wright, H., 1997. A model of peroxidase activity with
inhibition by hydrogen peroxide. Enz. Microb. Tech. 21 (4), 302±
310.
Ryu, K., McEldoon, J.P., Pokora, A.R., Cyrus, W., Dordick, J.S.,
1993. Numerical and Monte Carlo simulations of phenolic
polymerizations catalyzed by peroxidase. Biotech. Bioeng. 42,
807±814.
Sessa, D.J., Anderson, R.L., 1981. Soybean peroxidases: puri®cation
and some properties. J. Agric. Food Chem. 29, 960±965.
Tamura, Y., Morita, Y., 1975. Thermal denaturation and regeneration
of japanese-radish peroxidase. J. Biochem. 78, 561±571.
Yamazaki, I., Nakajima, R., 1986. Physico-chemical comparison
between horseradish peroxidases A and C. In: Greppin, H., Penel
C., Gaspar, T., (Eds.), Molecular and Physiological Aspects of
Plant Peroxidases, 3rd ed., University of Geneva, Switzerland, pp.
71±84.
Yu, J., Taylor, K.E., Zou, H., Biswas, N., Bewtra, J.K., 1994. Phenol
conversion and dimeric intermediates in horseradish peroxidase-
catalyzed phenol removal from water. Env. Sci. Tech. 28, 2154±
2160.
Zou, H., Taylor, K.E., 1994. Products of oxidative coupling of phenol
by horseradish peroxidase. Chemosphere 28, 1807±1817.
H. Wright, J.A. Nicell / Bioresource Technology 70 (1999) 69±79 79


















