
CHARACTERIZATION AND USE OF …plaza.ufl.edu/bbalogh/balogh_b_dissertation.pdf · Effect of a phage...
112
1 CHARACTERIZATION AND USE OF BACTERIOPHAGES ASSOCIATED WITH CITRUS BACTERIAL PATHOGENS FOR DISEASE CONTROL By BOTOND BALOGH A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 2006
Transcript of CHARACTERIZATION AND USE OF …plaza.ufl.edu/bbalogh/balogh_b_dissertation.pdf · Effect of a phage...

1
CHARACTERIZATION AND USE OF BACTERIOPHAGES ASSOCIATED WITH CITRUS BACTERIAL PATHOGENS FOR DISEASE CONTROL
By
BOTOND BALOGH
A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
UNIVERSITY OF FLORIDA
2006

2
Copyright 2006
by
Botond Balogh

3
Unokatestvérem, Dobó Gábor emlékére

4
ACKNOWLEDGMENTS
I would like to thank my committee members, Drs. Jeffrey B. Jones, Robert E. Stall, Timur
M. Momol, Donna H. Duckworth and Paul A. Gulig, for their support, constructive criticism and
guidance through the entire research project and preparation of this manuscript. I especially
appreciate Dr. Jones’ goodwill and loyalty that he showed in the time of need.
I would like to thank all those who helped me in this project: Aaron Hert, Jason Hong,
Frank Figueiredo, Mizuri Marutani, Fanny Iriarte, Ellen Dickstein, Jerry Minsavage, Nelly
Canteros, Alberto Gochez, Debra Jones, Xiaoan Sun, Amber Totten, Tanya Stevens, Mark
Gooch, Jake, Hans-W Ackermann, Donna Williams, Jim Dilley, Henry Yonce, Lee E. Jackson
and the employees of the OmnyLitics Inc., Justyna Kowara, Scott Taylor and the stuff of the
Citra Plant Science Unit, Terry Davoli, Ulla Benny, Kris Beckham, Gary Marlow, Patricia
Rayside, Mark Ross, Eldon Philman, Vanessa Ivanovski, Chandrika Ramadugu and the ones I
forgot to mention.
I would like to thank all those who, while not contributing directly to the project, helped
me during the time I was working on it: Abby Guerra, Gail Harris, Jim Barrel and Aleksa
Obradovic.

5
TABLE OF CONTENTS page
ACKNOWLEDGMENTS ...............................................................................................................4
LIST OF TABLES...........................................................................................................................8
LIST OF FIGURES .........................................................................................................................9
ABSTRACT...................................................................................................................................11
1 CITRUS CANKER.................................................................................................................13
The Citrus Industry .................................................................................................................13 Symptoms, Etiology and Epidemiology .................................................................................14 Disease Impact........................................................................................................................16 Disease Management ..............................................................................................................17 Genetic Variation of the Pathogen..........................................................................................17 Citrus Canker in Florida .........................................................................................................18 Citrus Canker Eradication Program........................................................................................20 Bacteriophages Associated with Citrus Canker......................................................................21 Citrus Bacterial Spot...............................................................................................................22 Project Goal and Objectives ...................................................................................................23
2 THE USE OF BACTERIOPHAGES FOR CONTROLLING PLANT DISEASES..............31
Early History...........................................................................................................................31 Other Uses of Phages in Plant Pathology ...............................................................................32 Return of Phage-Based Disease Control.................................................................................32 Considerations About Phage Therapy ....................................................................................33 Factors Influencing Efficacy of Phages as Biological Control Agents ..................................34 Current Research ....................................................................................................................37 Outlook for the Future ............................................................................................................38
3 CHARACTERIZATION OF BACTERIOPHAGES ASSOCIATED WITH CITRUS CANKER IN FLORIDA AND ARGENTINA ......................................................................40
Introduction.............................................................................................................................40 Materials and Methods ...........................................................................................................41
Bacterial Strains and Bacteriophages ..............................................................................41 Standard Bacteriophage Techniques ...............................................................................42 Phage Isolation from Diseased Plant Tissue ...................................................................44 Phage Typing of 81 Xanthomonas Strains ......................................................................45 Electron Microscopy .......................................................................................................45 Molecular Techniques .....................................................................................................45

6
Results.....................................................................................................................................47 Bacteriophage Isolations .................................................................................................47 Classification of Isolated Phages Based on Chloroform Sensitivity, Plaque
Morphology, Host Range and Virion Morphology......................................................47 Comparison of Florida Group A Phages and CP2 Based on Genome Size and RFLP
Profile...........................................................................................................................48 Phage Typing of 81 Xanthomonas Strains ......................................................................48
Discussion...............................................................................................................................49
4 CONTROL OF CITRUS CANKER AND CITRUS BACTERIAL SPOT WITH BACTERIOPHAGES.............................................................................................................63
Introduction.............................................................................................................................63 Materials and Methods ...........................................................................................................65
Bacterial Strains and Bacteriophages ..............................................................................65 Plant Material and Cultural Conditions in the Greenhouse.............................................65 Standard Bacteriophage Techniques ...............................................................................66 Disease Assessment and Data Analysis ..........................................................................66 Greenhouse Citrus Canker Control Trials .......................................................................67 Citrus Canker Nursery Trials in Argentina .....................................................................68 Citrus Bacterial Spot Disease Control Trials at Dilley and Son Nursery in Avon
Park, Florida.................................................................................................................69 Citrus Bacterial Spot Disease Control Trials at the Plant Science Unit of the
University of Florida in Citra, Florida. ........................................................................69 Results.....................................................................................................................................71
Effect of Bacteriophage Application and Use of Protective Skim Milk Formulation on Citrus Canker Disease Development in the Greenhouse ........................................71
Effect of Phage Application on Disease Severity Incited by a Phage-Sensitive Xac Strain and Its Phage-Resistant Mutant.........................................................................71
Effect of Phage and Copper-Mancozeb Applications on Citrus Canker Disease Development in a Citrus Nursery ................................................................................72
Effect of Bacteriophage Treatment on Citrus Bacterial Spot Disease Development in a Commercial Citrus Nursery ..................................................................................72
Effect of Bacteriophage and Copper-Mancozeb Treatments on Citrus Bacterial Spot Disease Development in an Experimental Nursery .....................................................73
Discussion...............................................................................................................................73
5 INTERACTION OF BACTERIOPAHGES AND THE HOST BACTERIA ON THE PHYLLOPLANE....................................................................................................................79
Introduction.............................................................................................................................79 Material and Methods .............................................................................................................80
Standard Bacteriophage Techniques ...............................................................................80 Changes in Xanthomonas axonopodis pv. citrumelo Phage Populations on the Field....81 Interaction of Xanthomonas perforans and Its Bacteriophage on the Tomato
Foliage in the Greenhouse ...........................................................................................82

7
Interaction of Xanthomonas axonopodis pv. citri and Its Bacteriophages on Grapefruit Foliage in the Greenhouse..........................................................................83
Results.....................................................................................................................................85 Persistence of Bacteriophages on Citrus Leaf Surface Under Field Conditions .............85 Ability of Three Phages of Xanthomonas axonopodis pv. citri to Multiply on
Grapefruit Foliage in the Presence of Their Bacterial Host, and Their Effect on Citrus Canker Disease Development ...........................................................................86
Ability of a Xanthomonas perforans Phage to Multiply on Tomato Foliage in the Presence of Its Bacterial Host, and Its Effect on Tomato Bacterial Spot Disease Development ................................................................................................................87
Discussion...............................................................................................................................88 Overall Summary and Conclusions ........................................................................................89
APPENDIX
A CALCULATIONS..................................................................................................................98
Conversion of Horfall-Barratt Values to Mean Percentages ..................................................98 Calculation of Area Under the Disease Progress Curve.........................................................98
LIST OF REFERENCES.............................................................................................................101
BIOGRAPHICAL SKETCH .......................................................................................................112

8
LIST OF TABLES
Table page 3-1. Bacterial strains used in this study ........................................................................................51
3-2. Bacteriophages used in the study...........................................................................................53
3-3. Chloroform sensitivity, plaque morphology and host range of bacteriophages originating in Florida, Argentina and Japan .........................................................................54
3-4. Summary of morphological characteristics of phage virions. ...............................................57
3-5. Grouping of Florida bacteriophages based on BamH I RLFP profile. ..................................62
4-1. Effect of bacteriophage treatment and use of skim milk formulation on citrus canker disease development incited by Xanthomonas axonopodis pv. citri strain Xac65 on grapefruit plants in the greenhouse .......................................................................................77
4-2. Effect of bacteriophage treatment on citrus canker disease development incited by phage sensitive Xanthomonas axonopodis pv. citri strain Xac65 and its phage-resistant mutant, RF2 on grapefruit plants in the greenhouse .............................................................77
4-3. Effect of phage and copper-mancozeb application on citrus canker disease development, incited by Xanthomonas axonopodis pv. citri strain Xc05-2592 in nursery trial in Bella Vista, Corrientes, Argentina ...............................................................77
4-4. Effect of bacteriophage treatment on citrus bacterial spot disease development incited by naturally occurring Xanthomonas axonopodis pv. citrumelo strains at Dilley and Son Nursery in Avon Park, Florida in 2004, as measured by area under the disease progress curve (AUDPC)......................................................................................................78
4-5. Effect of bacteriophage and copper-mancozeb treatment on citrus bacterial spot disease development incited by Xanthomonas axonopodis pv. citrumelo strain S4m at the UF Plant Science Unit in Citra, Florida in 2006, as measured by area under the disease progress curve (AUDPC)......................................................................................................78
5-1. Effect of a phage treatments on citrus canker disease development, incited by Xanthomonas axonopodis pv. citri strain Xac65, as measured by disease severity..............94
5-2. Effect of a curative application of bacteriophage ΦXv3-3-13h on tomato bacterial spot disease development, incited by Xanthomonas perforans strain ΔopgH:ME-B, as measured by area under the disease progress curve (AUDPC). ...........................................97
A-1. Horsfall-Barratt scale, the corresponding disease intervals and midpoint values. ...............99

9
LIST OF FIGURES
Figure page 1-1. Young citrus canker lesions on grapefruit. ............................................................................24
1-2. Severe citrus canker infection on Key lime...........................................................................25
1-3. Citrus canker lesions on lemon fruit......................................................................................26
1-4. Asian leafminer (Phyllocnistis citrella) tunnels on Swingle citrumelo foliage.....................26
1-5. Citrus canker infection in Asian leafminer tunnels. ..............................................................27
1-6. Covered citrus nursery in Argentina......................................................................................27
1-7. Windbreaks outline an orange grove in Argentina................................................................28
1-8. Citrus bacterial spot lesions on grapefruit leaves. .................................................................29
1-9. Citrus bacterial spot lesions in Asian leafminer tunnels........................................................30
3-1. Transmission electron micrographs of representative phages. A) CP2, B) ΦXaacA1, C) ΦXaacF1, D) ΦXaacF2, E) ΦXaacF3, F) ΦXaacF5, G) ΦXaacF8. .....................................56
3-2. Dendogram and phage sensitivity matrix showing relationship amongst Xanthomonas strains causing citrus canker and citrus bacterial spot based on similarity of sensitivity profile against a battery of 12 phages.. .................................................................................59
3-3. Undigested bacteriophage DNA............................................................................................60
3-4. Bacteriophage DNA digested with BamH I ..........................................................................61
3-5. Representatives of the 7 RFLP groups based on BamH I. digestion profile .........................62
4-1. Experimental citrus nursery, the location of the 2006 citrus canker trials, at the INTA research station in Bella Vista, Corrientes, Argentina..........................................................75
4-2. Disease control plots in Dilley and Son Nursery, Avon Park, Florida..................................75
4-3. Field plots at the Plant Science Research and Education Unit of the University of Florida in Citra, Florida ........................................................................................................76
5-1. Bacteriophage populations and sunlight irradiation in Citra, FL on July 18 and 19, 2006...91
5-2. Changes in bacteriophage populations on grapefruit foliage in the presence or absence the host, Xanthomonas axonopodis pv. citri strain Xac65....................................................92

10
5-3. Changes in bacteriophage populations on grapefruit foliage in the presence or absence the host, Xanthomonas axonopodis pv. citri strain Xac65....................................................93
5-4. Symptoms of tomato bacterial spot, incited by Xanthomonas perforans..............................95
5-4. Populations of bacteriophage ΦXv3-3-13h on tomato foliage in the presence or absence of its host Xanthomonas perforans strain ΔopgH:ME-B......................................................96
A-1. Visualization of the disese progress curve and of the area under the disease progress curve. A) Disease progress curve is prepared by plotting the diseases percentage values (y-axis) against the time of disease assessment (x-axis). B) Area under the disease progress curve (AUDPC) is one value describing the overall disease progress throughout the season. ........................................................................................................100

11
Abstract of Dissertation Presented to the Graduate School of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy
CHARACTERIZATION AND USE OF BACTERIOPHAGES ASSOCIATED WITH CITRUS BACTERIAL PATHOGENS FOR DISEASE CONTROL
By
Botond Balogh
December 2006
Chair: Jeffrey B. Jones Major Department: Plant Pathology
Citrus canker, incited by Xanthomonas axonopodis pv. citri and X. axonopodis pv.
aurantifolii, is one of the most damaging citrus diseases in the world. Citrus canker was
reintroduced to Florida in the 1990s and threatens the state’s $9 billion citrus industry. This work
focused on a biological control approach to use bacteriophages for reducing bacterial pathogen
populations and disease severity on citrus. Bacteriophages isolated from citrus canker lesions in
Florida and Argentina were evaluated based on plaque morphology, chloroform sensitivity, host
range, genome size, DNA restriction profile and virion morphology. The phage isolates showed a
lack of diversity, as 61 of 67 bacteriophages were nearly identical and the remaining six were
identical to each other. Mixtures of bacteriophages were evaluated for controlling citrus canker
in greenhouse trials in Florida and in nursery trials in Argentina. Bacteriophages reduced citrus
canker disease severity both in greenhouse and field trials. The level of control was inferior to
chemical control with copper bactericides. The combination of bacteriophage and copper
treatments did not result in increased control. Citrus canker field trials in Florida have been
prohibited until recently, as the disease was under eradication. For this reason we evaluated the
efficacy of phage treatment on a similar bacterial citrus disease, citrus bacterial spot, incited by
X. axonopodis pv. citrumelo. Bacteriophages reduced citrus bacterial spot severity. The level of

12
control was equal or inferior to chemical control with copper bactericides. The combination of
bacteriophage and copper treatments did not result in increased control. In experiments
monitoring the fate of bacteriophages on the citrus foliage following bacteriophage application,
phage populations stayed steady on the foliage during nighttime but were drastically reduced
within hours after sunrise. The rate of reduction varied among the phages. The ability of
bacteriophages to multiply on the plant foliage in the presence of their bacterial host was
investigated. Phages varied in their ability to multiply, and the ones that successfully increased in
populations on the bacterial host on the leaf surface also reduced disease severity, whereas the
ones that were unable to multiply in the target environment did not reduce disease severity. In
summary, bacteriophages show significant promise as part of an integrated management strategy
for controlling citrus canker.

13
CHAPTER 1 CITRUS CANKER
The Citrus Industry
Citrus species originate from the southeast Asia-India region. They were introduced to the
Americas by Portuguese and Spanish explorers in the 16th century (21). Today citrus is produced
in 140 countries (122), mainly between the North and South 40º latitudes (121), and citrus fruits
are first among fruit crops in the international trade based on value (122). Citrus production has
been on the rise throughout the second half of the 20th century, and the total citrus production of
the world is around 105 million tons per year (122). Orange (Citrus sinensis) accounts for almost
two thirds of the total citrus production (65%), followed by tangerine (C. reticulata) (21%),
lemon (C. limon) (6%) and grapefruit (C. paradisi) (5.5%) (124). Other significant commercially
grown species are lime (C. aurantifolia), pummelo (C. grandis) and citron (C. medica). The
largest citrus producers are Brazil (20%), United States (14%), China (12%), Mexico (6%) and
the countries of the Mediterranean Basin (15%) (122). Most citrus fruits are produced for fresh
market consumption and only around 30% is processed (122). Most of the processing is orange
juice production that is carried out almost exclusively in São Paulo state of Brazil and in Florida
(122).
Citrus is a perennial evergreen with around 50 years of commercial production (121).
While originally it was grown on its own root system, in modern commercial production the
producing plants are grafted onto rootstocks (121). Rootstocks influence adaptation to soil types,
tree size, production characteristics and can provide cold hardiness and resistance to diseases,
nematodes and insects (53, 54, 105, 111). Some important rootstock species are sour orange (C.
aurantium L.), trifoliate orange (Poncirus trifoliata) and citrumelo (C. paradisi ×P. trifoliata)
(111).

14
The total citrus production of the United States was 11.6 million tons ($2.68 billion value)
in the 2005-06 growing season (125). Florida is the biggest citrus producer in the country
providing 68% of the total production, followed by California (28%), Texas (4%) and Arizona
(4%).
Florida’s $9 billion citrus industry (49) is in a rather precarious situation presently. It was
heavily hit in 2004 and 2005 by hurricanes; as a result the total citrus production fell 40%
compared to the pre-hurricane 2003-04 season and the producing area shrunk to the lowest since
1994 with 576,400 bearing acres (125). Another effect of the two consecutive extreme hurricane
seasons was the large scale dissemination of the citrus canker bacterium throughout the state,
which eliminated hopes of eradicating the disease (34). The permanent presence of citrus canker
is expected not only to reduce production volume, increase prices and cause market losses (139),
but also to completely eliminate grapefruit production from the state (49). Moreover, citrus
greening, another devastating citrus disease, was detected in Florida in 2005 (60) and is known
to be present in 12 counties in south Florida (36). Citrus greening is caused by a phloem limited
bacterium, Liberibacter asiaticus, and unlike citrus canker it kills the infected trees (60). Due to
its epidemiology and the presence of its vector, the Asian citrus psyllid (Diaphorina citri) in
Florida, its eradication is not feasible (35).
Symptoms, Etiology and Epidemiology
Citrus canker is one of the most devastating diseases affecting citrus production worldwide
(49). Its center of origin is the southeast Asia-India region (21), similar to its host plants. By the
20th century it was present in most citrus growing areas around the globe: Asia, South and
Central Africa, North and South America, Australia, New Zealand and the Pacific islands (71).
Citrus canker causes erumpent lesions on fruit, foliage and young stems (Figures 1-1, 1-2,
1-3) (21). The earliest symptoms on leaves are minuscule, slightly raised blister-like lesions that

15
appear around 7 days after inoculation under optimum conditions (Figure 1-1). Optimum
temperature for disease development is 20-30°C (71). The lesions expand over time and their
color turns light tan and then tan-to-brown (49). Subsequently, water-soaked margins appear
around the lesions frequently surrounded by a chlorotic halo (Figure 1-2) (49). The center of the
lesion becomes raised and corky and the touch of the lesions feels like sand paper (106).
Eventually the lesions form a crater-like appearance (106) and in some hosts they fall out leaving
a shothole appearance (106). Severe disease can cause defoliation (47), dieback and fruit drop
(49, 71).
A single, plasmid-encoded bacterial protein, PthA, is responsible for inciting the symptoms
(30). It is delivered into the cytoplasm of plant cell by the type 3 secretion system of the
pathogen, from where it is transported into the nucleus (136). This protein is believed to be
important in inciting cell division, cell enlargement and cell death (20, 30).
The bacterium multiplies to high populations in the lesions and in the presence of free
moisture the cells will ooze out to the plant surface. Rain water collected from diseased leaves
contains high bacterial concentrations ranging from 104 to 105 cfu/mL (108). The inoculum is
then dispersed by wind driven rain to new growth on the same plant or to other plants (19,49).
Wind blown inoculum was disseminated up to 32 meters in Argentina (107); however, in Florida
rainstorms move the pathogen much further with estimates of up to 7 miles (50). Long distance
spread of inoculum occurs mostly as a result of human involvement either by moving diseased
plant material or on contaminated equipment (50). Extreme weather conditions, such as
hurricanes and tornadoes have also been shown to facilitate long distance disease spread (51,57).
Bacteria enter the tissues through stomatal openings (48,49) or wounds. Wind speeds
above 18 mph aid in penetration of bacterial cells through the stomatal pores (48). Wounding

16
often occurs by thorns, blowing sand or insect damage (49). Asian leafminer (Phyllocnistis
citrella), an insect that was introduced to Florida in 1993, creates wounds that expose the
mesophyll tissues allowing entry of bacterial inoculum (Figure 1-4, 1-5) (51). Leafminer itself is
not a vector of the disease (17), but its actions can lead to significant field infection even on
highly resistant cultivars and species (49).
Bacteria multiply in the expanding lesions (49) and survive only in the margins of old
lesions. The pathogen persists in lesions in the leaves or fruits until the tissues decompose;
however, long term survival, up to a few years, occurs in lesions in woody tissues (49). Outside
of plant tissues the bacterium is quickly eliminated by desiccation or sunlight irradiation (49),
and it cannot survive longer than a few days in the soil, probably due to competition with
saprophytes (43).
The two main determinants of host susceptibility are the stage of leaf expansion and the
resistance of mesophyll tissue (48,58). All above ground citrus tissues of susceptible genotypes
are sensitive when they are young, especially at the second half of the expansion phase of growth
(108).
Disease Impact
Citrus canker impacts the citrus industry on several levels. It reduces both fruit yield and
quality. While yield reduction has an effect in all spheres of citrus production, the quality
problems affect mostly the fresh fruits as the blemishes caused by the disease (Figure 1-3) render
the fruit aesthetically undesirable and thus unmarketable (50). Additional economic damages
result from market losses due to the disease exclusion policies in canker-free citrus growing
regions. Those regions and countries where citrus canker is not present ban importation of fruits
from canker inflicted areas because of the fear of introduction of the pathogen (50). Furthermore,
if citrus canker becomes endemic, commercial production of the most susceptible citrus species,

17
such as grapefruit, will cease as it becomes impossible to grow them profitably (50). More
resistant species such as tangerines will likely take their place (71,103).
Disease Management
Countries that are free of citrus canker apply quarantine measures to stop the introduction
of the pathogen. As a disease exclusion measure, they prohibit importation of fruits and plant
material from canker inflicted areas (50). Introduction of the disease is generally met by
eradication programs in which all infected and exposed citrus is destroyed (50). The disease was
subject to eradication with varying degrees of success; it was successfully eradicated from
Mozambique, New Zealand, Australia, South Africa, the Fiji Islands and twice from the US,
while eradication efforts failed in Argentina, Uruguay, Paraguay, and more recently in Florida
(98,103,106). The disease is currently under active eradication in Australia and São Paulo state
in Brazil (39,103). In endemic situations the emphasis has shifted to implementing integrated
management programs (76). The disease is under management in all Asian countries, Argentina,
Uruguay, Paraguay, several states of Brazil and Florida (49,76,109). These programs rely on
• planting resistant citrus cultivars (76), • production of disease-free nursery stock by locating nurseries outside of citrus canker areas
and/or indoors (Figure 1-6) (76), and • restricting disease spread by establishing windbreaks (Figure 1-7) and fences around
groves, using preventative copper bactericides (76,109) and by controlling Asian leafminer.
Additionally, in order to ensure that fresh fruit destined for internal and export markets is
disease free, producing groves are regularly inspected for the presence of citrus canker and
sanitation protocols are established in the packing houses (75).
Genetic Variation of the Pathogen
Citrus canker is not a disease caused by a single pathogen but rather a of group similar
diseases caused by closely related Xanthomonas species (49). The Asiatic type (Canker A) is

18
caused by X. axonopodis pv. citri (Xac) strains that originated from Asia. This is the most
geographically widespread pathogen, which also has the widest host range and by far the biggest
impact. Almost all commercially grown citrus varieties are susceptible to this bacterium to a
certain level. Grapefruit, Key lime and lemon are most susceptible. Sweet oranges range from
highly susceptible (Hamlins and Navels) to moderately susceptible (Valencias). Tangerines and
mandarins are moderately resistant while only calamondin (C. mitus) and kumquats (Fortunella
spp.) are highly resistant. The B type of citrus canker (cancrosis B or false canker) was present in
Argentina, Uruguay and Paraguay (103) from the 1920s to 1980s, and eventually was eliminated
by strains of the A pathotype. The causal agent, X. axonopodis pv. aurantifolii (Xaa) , has
limited host range compared to canker A: it is only pathogenic on lemon, Key lime, sour orange,
and pummelo. It was not pathogenic on grapefruit. The C type of citrus canker (cancrosis C) is
also caused by X. axonopodis pv. aurantifolii. It has only been found in São Paulo state in Brazil
and its only two known hosts are Key lime and sour orange. A fourth group of strains (A*
pathotype) was isolated in southwest Asia in the 1990s (126). These strains constitute a subgroup
of the A pathotype, X. axonopodis pv. citri, but their host range is limited to Key lime (5). The
different pathotypes can be distinguished by host range, cultural and physiological characteristics
(49), bacteriophage sensitivity (21), serology (6), plasmid fingerprints (94), DNA-DNA
homology (32), and by various RFLP and PCR analyses (26).
Citrus Canker in Florida
Citrus canker was first introduced into Florida in 1912 on rootstocks imported from Japan
(29). It spread to all Gulf states between Texas and South Carolina. A strict eradication program
was implemented and the disease was eliminated from Florida by 1933 and from the entire
United States by 1947 (29). The second eradication program began in 1984, when Xanthomonas
was isolated from lesions from nursery stock (102,103). Research later showed that this pathogen

19
differed from xanthomonads causing citrus canker. The bacterium was determined to be
endemic, exclusive to Florida, and considerably less aggressive than Xac (103,106). The disease
was named citrus bacterial spot (CBS), the pathogen was named X. axonopodis pv. citrumelo,
and the eradication efforts for CBS were cancelled in 1990 (103). By that time 20 million citrus
plants were destroyed and $94 million was spent (103). In 1986 genuine citrus canker was
discovered in the Tampa Bay area. An eradication effort was begun and the pathogen was
declared eradicated in 1994 (103). Another introduction was discovered in urban Miami in 1995.
The pathogen, called Miami genotype, is determined to be genetically related to Xac strains from
several geographical areas from Southeast Asia and South America (50). The Miami genotype
later spread throughout the state despite the eradication program (26,103) and was responsible
for the majority of post 1997 outbreaks (50). In 1997 there was a new outbreak in the Tampa Bay
area. The pathogen, called Manatee genotype, was identical to Xac strains from China and
Malaysia, based on rep-PCR analysis (26). Also, it was indistinguishable from Xac strains that
caused the outbreak in the 1980s in the Tampa Bay region (26,103), implying that the earlier
eradication effort was not successful. Interestingly, the Manatee genotype disappeared again in
1999 and then reappeared in 2005 (113). In 2000, a new genotype of Xac was identified and was
designated the Wellington genotype. It was discovered in the Palm Beach area. Its host range
was limited to Key Lime (114). Wellington genotype is closely related to the A* strains and thus
likely originates from South-west Asia (26). It was later determined that the inability of this
genotype to infect grapefruit was due to the presence of an avirulence gene, avrGf1, in its
genome (98). The latest introduction of an exotic strain was in 2003 in Orange County. These
strains were found in a residential area on an Etrog citron tree (Citrus medica) that was probably

20
brought to Florida from Pakistan illegally (Etrog genotype) (X. Sun, D. Jones, R.E. Stall,
personal communication).
Citrus Canker Eradication Program
The Citrus Canker Eradication Program (CCEP) was established in 1995 in response to the
Miami outbreak by the Florida Department of Agriculture and Consumer Services (FDACS),
Division of Plant Industry (DPI) and the USDA, Animal and Plant Health Inspection Service
(APHIS) (49). The original quarantine area was 14 square miles in the urban Miami area (103).
After the discovery of citrus canker in the Tampa Bay area in 1997, the quarantine area was
extended to that region as well (49). Citrus trees infected by the disease or located within the
exposure area were uprooted and burned in the commercial groves, or cut down and chipped in
urban areas (49). The exposure area was originally 125 feet radius of a diseased tree, based on
data collected from Argentina (108). Later research showed that the inoculum spreads further
than 125 feet under the Florida conditions (50,51), and the exposure radius was extended to 1900
feet in January 2000 based on epidemiological data collected in the Miami area (50,51). The
eradication program was hindered by strong public resistance; the ensuing legal battles often
delayed the surveying and tree removal (103). The legal challenges were overcome in spring
2004, but then the hurricanes of 2004 and 2005 spread the pathogen from areas awaiting
eradication widely throughout the citrus growing area. On January 10, 2006, APHIS
discontinued the CCEP (24,123), because its continuation in the new situation was judged
infeasible (i) due to financial constraints associated with the tree removal and reimbursement
programs, and (ii) because the citrus industry could not survive the loss of such a large
production area (24,123). During the program 16.5 million trees were destroyed, including 11.3
million trees from commercial groves, equaling 15% of the total bearing acreage (34). The total
costs of CCEP exceeded $600 million (138). In March 2006 APHIS released a Citrus Health

21
Response Plan, which outlined procedures for managing citrus production in the permanent
presence of the disease (34).
The impact of living with the endemic citrus canker situation in the Florida is estimated at
$254.2 million annually (139) and may result in elimination of grapefruit production (49). The
acceptance of citrus canker as an endemic disease will also result in losses in interstate and
international commerce of the state’s fresh citrus fruit, which currently represents 20% of the
state’s $9 billion dollar citrus industry (88).
Bacteriophages Associated with Citrus Canker
There are several reports on bacteriophages found in association with citrus canker. Phages
CP1 and CP2 have been isolated in Japan (45). Goto found that both CP1 and CP2 had wide host
ranges and more than 97% of Xac strains present in Japan were sensitive to one of these two
phages (44). In fact, the Japanese Xac strains comprised two groups: the strains in the first one
originated mostly from Unshu orange (Citrus unshu) and were sensitive to CP2 only, whereas
the members of the second group had a variety of hosts and were sensitive only to CP1. These
phages have been used for detection of the pathogen (45). Bacteriophage CP3, which was also
isolated in Japan, had a tadpole shape with a spherical head and long tail (46). Goto et al. (46)
found that strains of the B pathotype (X. axonopodis pv. aurantifolii) could be distinguished from
type A strains (Xac) based on sensitivity to phage CP3, with all B strains being sensitive to CP3
and all A strains being resistant. Canker C strains were differentiated from canker A strains by
their resistance to both CP1 and CP2 (106).
Filamentous phage Cf was isolated in Taiwan (27). It has a very narrow host range (44),
contains single stranded DNA that is approximately 1 kb long, produces small and clear plaques
and is a temperate phage (27). The product of pilA gene, a type 4 prepilin of the host bacterium is
required for infection of Cf (27). Phages CP115 and CP122, isolated from citrus canker lesions

22
in Taiwan, tested for their ability to lyse Taiwanese strains of Xac, lysed 97.8% of them when
used in combination (135). These phages, however, did not lyse Xanthomonas strains that did not
cause citrus canker, or any other bacteria tested. The authors concluded that these phages could
be used for specific detection of Xac strains in Taiwan. Temperate phage PXC7 that was isolated
from Japanese Xac strain XCJ18 produces small turbid plaques with irregular borders and is
sensitive to chloroform (134). When Xac strain XCJ19 was lysogenized with the phage, its
colony morphology changed from smooth to dwarf and became resistant to phage CP2 (134).
Bacteriophages were also found in citrus canker lesions in Argentina in 1979 (R.E. Stall,
personal communication).
Citrus Bacterial Spot
Citrus bacterial spot (CBS) was discovered in 1984 as a new Xanthomonas disease of
citrus nursery stock that causes canker-like symptoms (102). Citrus bacterial spot differs from
citrus canker in that it causes flat or sunken lesions (Figure 1-8) instead of the corky, raised ones
and is generally less aggressive than citrus canker (106). The pathogen, Xanthomonas
axonopodis pv. citrumelo (Xacm) only exists in Florida (103) and is genetically different than
Xanthomonas strains causing citrus canker (26). Xacm strains are genetically diverse (26) and
comprise three groups based on aggressiveness as measured by rate of lesion expansion and the
ability to multiply in citrus leaves (56). Only the most aggressive strains are able to maintain
high populations in the lesions. Graham et al. (56) questioned if the less aggressive strains
should be considered a citrus pathogen at all. The most aggressive strains are disseminated by
wind driven rain, whereas strains of intermediate and low aggressiveness are spread mainly by
mechanical means (56). Wounding caused by thorns and citrus leafminer (Figure 1-9) facilitates
pathogen entry and increases CBS disease incidence and severity. The disease causes a reduction
in photosynthetically active leaf area and in severe cases can induce leaf drop. Xacm infection

23
also causes bud failure in Swingle citrumelo plants (55). The CBS bacteria are most aggressive
on trifoliate orange, Swingle citrumelo and grapefruit (103,106).
Project Goal and Objectives
The goal of this project was to develop a bacteriophage-based disease control strategy that
could be used as part of an integrated management program against citrus canker in Florida. The
objectives were (i) establishment of a bacteriophage collection against Xac strains present in
Florida, (ii) collection and characterization of bacteriophage associated with citrus canker in
Florida and Argentina; (iii) determination of bacteriophage sensitivity of Xac strains present in
Florida, and (iv) evaluation of bacteriophages for suppression of citrus canker in greenhouse
experiments in Florida and in field experiments in Argentina, and for suppression of citrus
bacterial spot, as a model disease system, in field experiments in Florida.

24
Figure 1-1. Young citrus canker lesions on grapefruit.

25
Figure 1-2. Severe citrus canker infection on Key lime.

26
Figure 1-3. Citrus canker lesions on lemon fruit.
Figure 1-4. Asian leafminer (Phyllocnistis citrella) tunnels on Swingle citrumelo foliage.

27
Figure 1-5. Citrus canker infection in Asian leafminer tunnels.
Figure 1-6. Covered citrus nursery in Argentina.

28
Figure 1-7. Windbreaks outline an orange grove in Argentina.

29
Figure 1-8. Citrus bacterial spot lesions on grapefruit leaves.

30
Figure 1-9. Citrus bacterial spot lesions in Asian leafminer tunnels.

31
CHAPTER 2 THE USE OF BACTERIOPHAGES FOR CONTROLLING PLANT DISEASES
Early History
Bacteriophages were discovered in the beginning of the 20th century independently by
Twort in 1915 and by d’Herelle in 1917 (112). There were differences in interpretation about the
nature and origin of this “lytic principle”. Twort proposed that a bacterial enzyme caused the
lysis, while d’Herelle speculated that a virus was responsible for the phenomenon. A direct
consequence of d’Herelle’s concept was the idea of using phages for controlling bacterial
diseases. Soon after the first medical (112) and veterinary (28) applications, phages were
evaluated for control of plant diseases.
In 1924 Mallman and Hemstreet (79) isolated the “cabbage-rot organism”, Xanthomonas
campestris pv. campestris, from rotting cabbage and demonstrated that the filtrate of the liquid
collected from the decomposed cabbage inhibited in vitro growth of the pathogen. The following
year Kotila and Coons (73) isolated bacteriophages from soil samples that were active against the
causal agent of blackleg disease of potato, Erwinia carotovora subsp. atroseptica. They
demonstrated in growth chamber experiments that co-inoculation of E. carotovora subsp.
atroseptica with phage successfully inhibited the pathogen and prevented rotting of tubers (73).
These workers also isolated phages against Erwinia carotovora subsp. carotovora and
Agrobacterium tumefaciens from various sources such as soil, rotting carrots and river water
(25). Thomas (120) treated corn seeds that were infected with Pantoea stewartii, the causal agent
of Stewart’s wilt of corn, with bacteriophage isolated from diseased plant material. The seed
treatment reduced disease incidence from 18% to 1.4%. Despite the promising early work, phage
therapy did not prove to be a reliable and effective means of controlling phytobacteria. Several
workers questioned if positive results were possible. In 1963, Okabe stated, “in general, the

32
phage seems to be ineffective for [controlling] the disease development” (91). Three decades
later, Goto concluded, “practical use of phages for control of bacterial plant disease in the field
has not been successful.” (44). Chemical control with antibiotics and copper compounds became
the standard for controlling bacterial plant diseases (31,81).
Other Uses of Phages in Plant Pathology
Bacteriophages still remained in use in plant pathology and have been used as tools for
detection, identification, classification, and enumeration of pathogenic bacteria and were also
used for disease forecasting. Phage typing, as a method of differentiating different races or
pathovars of the same bacterial species became a standard method in plant epidemiological
studies (70). Phages CP1 and CP2 of Xanthomonas axonopodis pv. citri, the causal agent of
citrus canker, were used for species-specific identification and classification of strains of the
pathogen in Japan (44). These two phages in combination with phage CP3 were used for
differentiating worldwide strains causing citrus canker (46). Wu et al. (135) used phages CP115
and CP122 for identification of X. axonopodis pv. citri strains in Taiwan. Phages were used for
detection of the host bacterium from crude samples and seed lots (68) and directly from lesions
on the plant foliage (91) by monitoring increases in homologous phage concentration. Okabe and
Goto (91) demonstrated that phages could be used for quantifying bacterial cells based on the
number of newly produced phages and average burst size. They developed a method for
indirectly forecasting bacterial leaf blight by monitoring phage titers in rice fields (91).
Reliability of this latter method was questioned later by Civerolo (22).
Return of Phage-Based Disease Control
Several factors have contributed to the re-evaluation of phage therapy for plant disease
control. The use of antibiotics has been largely discontinued in agriculture due to the emergence
of antibiotic-resistant bacteria in the field (80,84,119) and because of concerns of possible

33
transfer of antibiotic resistance from plant pathogens to human pathogens. The feasibility of
reliance on copper compounds is questioned, because of the emergence of copper-tolerant strains
among phytobacteria (81,129); phytotoxicity caused by ionic copper (85,109) and soil
contamination from extended heavy use (72). Additionally, concerns about food safety and
environmental protection and the goal of achieving sustainable agriculture necessitated
development of safer, more specific and environment-friendly pesticides (96). These factors,
together with the expanding knowledge base about phage application in medicine (13,31,74), led
to renewed interest in bacteriophage-based disease control in modern agriculture.
Considerations About Phage Therapy
Greer (59) and Kutter (74) identified the several advantages of using phages for disease
control.
1. Phages are self-replicating and self-limiting; they replicate only as long as the host bacterium is present in the environment, but are quickly degraded in its absence (74).
2. Bacteriophages are natural components of the biosphere; they can readily be isolated from everywhere bacteria are present, including soil, water, plants, animals (2,52,133) and the human body (93).
3. Phages could be targeted against bacterial receptors that are essential for pathogenesis, so resistant mutants would be attenuated in virulence (74).
4. Bacteriophages are non-toxic to the eukaryotic cell (59). Thus, they can be used in situations where chemical control is not allowed due to legal regulations, such as for treatment of peach fruit before harvest (137) or for control of human pathogens in fresh-cut produce (77,78).
5. Phages are specific or highly discriminatory, eliminating only target bacteria without damaging other, possibly beneficial members of the indigenous flora. Thus their use can also be coupled with the application of antagonistic bacteria for increased pressure on the pathogen (117); or they can be used to promote a desired strain against other members of the indigenous flora (14).
6. Phage preparations are fairly easy and inexpensive to produce and can be stored at 4 ºC in complete darkness for months without significant reduction in titer (59). Application can be carried out with standard farm equipment and since phages are not inhibited by the majority of agrochemicals (9, 137), they can be tank-mixed with them without significant loss in titer. Copper-containing bactericides have been shown to inactivate phages (9,66); however, inhibition was eliminated if phages were applied at least three days after copper (66).
A number of disadvantages and concerns have been raised in relation to phage therapy
(59,74,127); moreover, additional problems specific to agricultural applications have surfaced.

34
1. Limited host range can be a disadvantage, as often there is diversity in phage types of the target bacterium (59). Several approaches have been tried for addressing this problem: using broad host range phages (99,115), using host range mutant phages (37,38,89), applying phages in mixtures (38) or even to breed them (62).
2. The requirement of threshold numbers of bacteria (104-106 cfu/mL) may limit the impact of phages (131).
3. Emergence of phage resistant mutants can render phage treatment ineffective. However, using mixtures of phages that utilize distinct cell receptors can suppress the emergence of resistance (118). Also, phage resistance often comes at some metabolic cost to the bacteria. Loss of virulence was observed with phage-resistant mutants of Ralstonia solanacearum (61), Xanthomonas campestris pv. pruni (95), and Pantoea stewartii (120).
4. Environmental effects, such as temperature, pH and physiology of bacterium can hinder control. Civerolo (22) observed that Xanthomonas phaseoli phages attacked Xanthomonas and Pseudomonas species only at temperatures above 20ºC. Vidaver (128) suggested that P. syringae and P. phaseolicola, causal agents of halo blight and brown spot of bean, may be more prevalent below 22ºC because of phage resistance. Leverentz (78) noted that phage treatment caused a significant population reduction of the Listeria monocytogenes on melons but not on apples, because phages were unstable on apple slices, possibly due to low pH (4.37 in apple vs. 5.77 in melon) (78).
5. Unavailability of target organism can hinder control. Plant pathogenic bacteria often occur in non-homogenous masses surrounded by extracellular polysaccharides that protect them from phage attachment (44,91), or reside in protected spaces on the surface, or inside the plant and unavailable for the control agents (22).
6. There is a concern that phages have the potential of transducing undesirable characteristics, such are virulence factors, between bacteria (127).
7. Lysogenic conversion, alteration of phenotypic characteristics of lysogenized bacteria by their prophages, have been found to have undesirable consequences, such as resistance to bacteriophages, toxin production or even increased virulence. When Xanthomonas axonopodis pv. citri strain XCJ19 was lysogenized with temperate phage PXC7, it became resistant to phage CP2 (134). Phage-associated toxin production has not been documented amongst phytobacteria, but such cases are known amongst human pathogens (130) and in bacteria of plant associated nematodes (92). Goto (44) reported that Xanthomonas campestris pv. oryzae strains lysogenized by phages Xf or Xf-2 became more virulent on rice.
8. Consumer perception of adding viruses to food products also could become an issue (59). 9. Vidaver (127) raised the concern that “transducing phages can introduce active prokaryotic
genes into plant and animal cells”. 10. Despite the generally narrow host ranges of phages, negative side effects due to inhibition of
beneficial bacteria are possible. Examples for negative phage impact in agriculture include studies in which the phage-incited reduction in symbiotic nitrogen fixing bacteria reduced growth and nitrogen content of cowpea (4) and in which biocontrol ability of Pseudomonas flourescens was abolished by a lytic bacteriophage (69).
Factors Influencing Efficacy of Phages as Biological Control Agents
Goodridge stated that the efficacy of phage therapy depends on the ability of a phage to
find its host before it is destroyed (42). According to Johnson (67) the success of a particular

35
biocontrol treatment is influenced by agent and target densities. A component of Johnson’s
model is the possibility that the target resides in spatial refuges where the biocontrol agent
cannot penetrate. Gill and Abedon (40) proposed several additional factors specifically in
relation to phage therapy: the location in which the target pathogen resides; the presence of
adequate water as a medium for virus diffusion; rates of virion decay; timing of phage
application; phage in situ multiplication ability; and relative fitness of phage-resistant bacterial
mutants.
Gill and Abedon (40) looked at the factors that could contribute to success or failure of
phage therapy in the rhizosphere and in the phyllosphere. They suggested that phage therapy
might meet with more success in the rhizosphere because phages are readily isolated from there
and can survive longer in the soil than on the leaf surface. However, they identified several
factors that can hinder success of disease control in the rhizosphere. The rate of diffusion through
the heterogeneous soil matrix is low and changes as a function of available free water. Phages
can become trapped in biofilms (110) and reversibly adsorb to particles of the soil, such as clay
(132). Low soil pH can also inactivate them (116). Physical refuges can protect bacteria from
coming into contact with phages, and due to the low rates of phage diffusion and high rates of
phage inactivation only a low number of viable phages is available to lyse target bacteria (40).
An additional problem is the need for high population of both phage and bacterium in order to
start the “chain reaction” of bacterial lysis (40).
The phyllosphere is a harsh environment, because of high UV and visible light irradiation
and desiccation (40). It has been noted that phages were harder to isolate from aerial plant tissue
than from soil even for pathogens of aerial tissues (37,41,91). Phages applied to aerial tissues
degrade extremely rapidly during the day (8,10,23,66,83). Additionally, the lack of moisture on

36
the leaf surfaces does not allow phage dispersion except for temporary leaf wetness periods after
application, during rain events or when dew is present on the leaves at night and early morning.
There were several approaches to increase efficacy of control in the phyllosphere environment,
including applying treatments in the evening or early morning (10,38), using protective
formulations that increase phage longevity on the foliage (8,10,66,89) and using carrier bacteria
for phage propagation in the target environment (115). Using phages isolated from aerial tissues
might be advantageous, as they might be better adapted for surviving and multiplying on the
plant surfaces. A phyllosphere phage investigated by Iriarte et al. (66) turned out to be resistant
to desiccation.
Timing of bacteriophage applications relative to the arrival of the pathogen influenced
efficacy of disease control in several instances. Civerolo and Keil (23) achieved a marked
reduction of peach bacterial spot only if phage treatment was applied one hour or one day before
inoculation with the pathogen. There was a slight disease reduction when phage was applied one
hour after inoculation and no effect if applied one day later. Civerolo (22) suggested that bacteria
were inaccessible to phage in the intercellular spaces, or there were not enough phages reaching
the pathogen. Schnabel et al. (101) achieved a significant reduction of fire blight on apple
blossoms when the phage mixture was applied at the same time as the pathogen, Erwinia
amylovora. In contrast, disease reduction was not significant when phages were applied a day
before inoculation. Berghamin Filho (18) investigated the effect of timing on the efficacy of
phage treatment in greenhouse trials with two pathosystems: black rot of cabbage, caused by
Xanthomonas campestris pv. campestris and bacterial spot of pepper, caused by Xanthomonas
campestris pv. vesicatoria. Phage treatment was applied once varying from 7 days before to 4
days after pathogen inoculation. On cabbage significant disease reduction was achieved if the

37
phage treatment was applied 3 days before to 1 day after inoculation, whereas on pepper it was
achieved when applied from 3 days before to the day of inoculation. The greatest disease
reduction occurred with application of phages the same day as inoculation in both pathosystems.
The effect of phage concentration on disease control efficacy has also been investigated.
Balogh (8) treated tomato plants with phage mixtures of 104, 106 and 108 PFU/mL before
inoculating with Xanthomonas perforans, causal agent of tomato bacterial spot. The two higher
concentrations significantly reduced disease severity, whereas the lowest concentration did not.
Current Research
There has been considerable amount of work on the use of phages for control of bacterial
spot of peach, caused by Xanthomonas campestris pv. pruni. Civerolo and Keil (23) reduced
bacterial spot severity on peach leaves under greenhouse conditions with a single application of a
single-phage suspension. Zaccardelli et al. (137) isolated eight phages active against the
pathogen, screened them for host range and lytic ability, and selected a lytic phage with the
broadest host range for disease control trials. Biweekly spray-applications of the phage
suspension in producing orchards significantly reduced bacterial spot incidence on fruits (99).
Tanaka et al. (117) treated tobacco bacterial wilt, caused by Ralstonia solanacearum, by
co-application of an antagonistic avirulent R. solanacearum strain and a bacteriophage that was
active against both the pathogen and the antagonist. The avirulent strain alone reduced the ratio
of wilted plants from 95.8% to 39.5%, whereas the co-application of the avirulent strain and the
phage resulted in 17.6% wilted plants.
Control of Erwinia amylovora, the fire blight pathogen of apple, pear and raspberry, with
bacteriophages is currently under investigation in Canada and the USA. Schnabel et al. (101)
used a mixture of three phages for controlling fire blight on apple blossoms and achieved
significant (37%) disease reduction. Gill et al. (41) isolated 47 phages capable of lysing E.

38
amylovora and categorized them based on plaque morphology and host range. Later the phages
were evaluated for disease control ability in pear blossom bioassays, and the ones with broad
host ranges and best disease control ability were selected for subsequent orchard trials (115).
Pantoea agglomerans, a bacterial antagonist that was also sensitive to the phages, was used to
deliver and propagate them on the leaf surface. Disease control comparable to streptomycin was
achieved (115).
There has been extensive research on suppressing tomato bacterial spot with phage.
Flaherty et al. (38) effectively controlled the disease in greenhouse and field experiments with a
mixture of four host-range mutant phages active against the two predominant races of the
pathogen, X. campestris pv. vesicatoria. Balogh et al. (10) enhanced the efficacy of phage
treatment with protective formulations that increased phage persistence on tomato foliage.
Obradovic et al. (89,90) used formulated phages in combination with other biological control
agents and systemic acquired resistance inducers, as a part of integrated an disease management
approach. Phage-based integrated management of tomato bacterial spot is now officially
recommended to tomato growers in Florida (85), and bacteriophage mixtures against the
pathogen are commercially available (Agriphage from OmniLytics Inc. Salt Lake City, UT, EPA
Registration # 67986-1).
Other important work includes the reduction of incidence of bacterial blight of geranium
with foliar applications of a mixture of host-range mutant phages (37), disinfection of
Streptomyces scabies-infected seed potatoes using a wide host range phage (82), and a reduction
in the loss of cultivated mushrooms caused by bacterial blotch with phage applications (86,87).
Outlook for the Future
Use of bacteriophages for controlling plant diseases is an emerging field with great
potential. The concern about environment-friendly sustainable agriculture and the rise of organic

39
production necessitates improvements in biological disease control methods, including the use of
bacteriophages against bacterial plant pathogens. On the other hand, the lack of knowledge about
the biology of phage-bacterium-plant interaction and influencing factors hinders progress in the
field. Much research in these areas is needed before phages can become effective and reliable
agents of plant disease management.

40
CHAPTER 3 CHARACTERIZATION OF BACTERIOPHAGES ASSOCIATED WITH CITRUS CANKER
IN FLORIDA AND ARGENTINA
Introduction
In order to develop a phage based disease control strategy it is necessary to (i) establish a
collection of phages that could be used for control; (ii) evaluate diversity of the target organism,
and, (iii) determine if there are phages associated with the pathogen in nature and if so, assess
their impact.
Xanthomonas strains causing citrus canker have been classified based on phage sensitivity
in the past. Goto (44) categorized Xanthomonas axonopodis pv. citri (Xac) strains present in
Japan, causing A type of citrus canker, into two groups based on their sensitivity to phages CP1
and CP2. More than 97% of Xac strains were sensitive to one of these two phages, but none to
both of them. The strains that were sensitive to CP2 originated mostly from Unshu orange
(Citrus unshu), whereas the ones sensitive only to CP1 had a variety of hosts. Goto et al. (46)
found that strains of the B pathotype of citrus canker (X. axonopodis pv. aurantifolii (Xaa))
could be distinguished from type A strains (Xac) based on sensitivity to phage CP3, with all B
strains sensitive to CP3 and all A strains resistant. Strains of the C pathotype (Xaa) were also
differentiated from A strains by their resistance to both CP1 and CP2 (106).
There is known diversity amongst Xac strains in Florida. Four distinct genotypes have
caused outbreaks since 1993 (49): the Miami, the Manatee, the Wellington and the Etrog
genotypes. Cubero et al. (26) used rep-PCR protocol for distinguishing these genotypes and to
determine their geographic origin. They determined that (i) the Miami genotype was related to
Xac strains from several geographical areas in southeast Asia and South America; (ii) the
Manatee genotype was identical to Xac strains from China and Malaysia and also to Xac that was
present in Florida in the 1980s and was supposedly eradicated; and (iii) the Wellington genotype

41
was related to A* strains (Xac) from southwest Asia. There has not been any genetic analysis
published in relation to the Etrog strains, but it is suspected that they were brought into Florida
on plant material from Pakistan.
The objective of this project was to assemble a phage collection active against citrus
canker, partly from academic and commercial sources, and partly by isolating them from plant
tissue showing citrus canker symptoms in Florida, where the disease was under eradication, and
in Argentina, where the disease is endemic. Once a collection was established and the phages
were grouped based on host range, they were used to type the prevalent Xac strains in Florida.
The typing results provided information about the diversity of the pathogen and determined
which phages could be used for disease control. Changes in phage sensitivity of Xac strains
isolated in different years may provide information on what impact naturally occurring phages
have on the pathogen.
Materials and Methods
Bacterial Strains and Bacteriophages
Bacterial strains were grown on nutrient agar (NA) medium (0.8% (wt/V) nutrient broth
(NB) (BBL, Becton Dickinson and Co., Cockeysville, MD) and 1.5% (wt/V) Bacto Agar (Difco,
Becton Dickinson and Co., Sparks, MD)) at 28°C. For bacteriophage detection and propagation
either semisolid nutrient agar yeast extract medium (NYA), (0.8% Nutrient Broth, 0.6% Bacto
Agar and 0.2% Yeast Extract (Difco, Becton Dickinson and Co., Sparks, MD)) or liquid nutrient
broth medium was used. Sterilized tap water or SM buffer (0.05 M Tris-HCl (pH 7.5), 0.1 M
NaCl, 10 mM MgSO4 and 1% (w/V) gelatin) was used for preparing phage suspensions.
Bacterial strains used in this study (Table 3-1) were stored at -80ºC in NB supplemented with
30% glycerol. Bacteriophages (Table 3-2) were stored at 4ºC and protected from light.

42
Standard Bacteriophage Techniques
Purification and storage. Phages were purified by three subsequent single plaque
isolations. Single plaque isolations were carried out by transferring phages from isolated plaques
to a fresh lawn of the host bacterium using sterile toothpicks and then quadrant streaking them
with sterile plastic transfer loops. Following purification the phages were propagated by mass
streaking on fresh lawns of the host. After a 24-h incubation at 28ºC, the phages were eluted by
pouring 5 mL sterilized tap water into the 100 mm×15 mm Petri dishes (Fischer Scientific Co.
LLC, Suwannee, GA) and gently shaking the plates (~20 rpm) for 30 min. The eluate was
centrifuged (10,000 g, 10 min), treated with chloroform or filter-sterilized, depending on the
phage, then quantified as described below, and stored in 2-mL plastic vials at 4°C in complete
darkness. The concentrations of these suspensions were approximately 109 plaque forming units
(PFU) per mL.
Determination of titer. Phage concentrations were determined by dilution-plating-plaque-
count assay on NYA plates without bottom agar as previously described (97). One hundred
microliter aliquots of dilutions of phage suspensions were mixed with 100 µL of concentrated
bacterial suspension in empty Petri dishes and then 12 mL warm (48°C) NYA medium was
poured in each dish. The dishes were gently swirled to evenly distribute the bacteria and the
phages within the medium. After the medium solidified, the plates were transferred to 28°C
incubators and the plaques were counted on the appropriate dilutions after 24 or 48 hours. The
phage concentration was calculated from the plaque number and specific dilution and was
expressed as PFU/mL.
Phage propagation. Phages were recovered from storage, purified by single plaque
isolations and then mass streaked on the freshly prepared lawn of the propagating host. The next
day phages were eluted from the plate, sterilized and enumerated, as described above. The eluate

43
was used for infecting 500 mL actively growing culture of the propagating strain (108 cfu/mL)
grown in NB liquid medium in 1 liter flasks, at 0.1 multiplicity of infection (MOI), (i.e., the
phage concentration at the beginning of the incubation was 107 PFU/mL). After addition of the
phage and 5-min incubation on the bench top, the culture was shaken at 150 rpm at 28ºC for 18
h. The resulting culture was sterilized; phages were enumerated and stored at 4 ºC in the dark
until use. This method yielded phage titers of approximately 1010 PFU/mL.
Phage concentration by high speed centrifugation. High titer phage lysates (~1010
PFU/mL) were concentrated and purified according to methods described by Hans-W.
Ackermann (personal communication). One hundred milliliters of the sterilized lysate was
centrifuged at 10,000 g for 10 min to sediment the bacterial debris. Forty milliliters of the
supernatant was transferred to a new centrifuge tube and centrifuged at 25,000 g for 60 min to
sediment the phage particles. The supernatant was discarded and replaced with 0.1 M ammonium
acetate solution (pH 7.0). Following an additional centrifugation (60 min, 25,000 g) the
supernatant was discarded and the pellet was resuspended in 1.5 mL SM buffer. The final phage
concentration was approximately 1012 PFU/mL.
Evaluation of bacterial sensitivity. Sensitivity of a bacterial strain to phages was
determined based on the ability of the phage to produce plaques on the bacterial lawn, and the
level of sensitivity was evaluated based on efficiency of plating (EOP) on the test strain in
comparison with the propagating host strain of the phage as follows. A phage suspension of
known concentration was plated simultaneously on the test and the host strains and EOP was
calculated as the number of plaques on the test strain divided by the number of plaques on the
host strain. For example, if the phage produced 55 plaques on the host and 36 plaques on the test
strain, then EOP = 36/55 = 0.65. The higher the EOP, the more efficient the phage is in initiating

44
disease of the bacterium. Consequently, the more sensitive the bacterial strain is to the phage. If
the EOP was higher than or equal to 0.1, the test strain was considered sensitive; if the EOP was
less than 0.1, but more than or equal to 0.01 the strain was considered moderately sensitive. If
EOP was less than 0.01, the strain was considered resistant.
Phage Isolation from Diseased Plant Tissue
Bacteriophages were isolated from leaves and fruits with characteristic citrus canker
lesions in Florida and in Argentina. In Florida, phage was isolated from diseased tissue received
from the Florida Department of Agriculture & Consumer Services, Division of Plant Industry,
Gainesville, FL, between May and August 2003, as a part of the Citrus Canker Eradication
Program (53). In Argentina, diseased tissue was collected directly from infected trees located at
the Instituto Nacional de Tecnología Agropecuaria (INTA) research center in Bella Vista,
Corrientes, and from commercial citrus groves in Corrientes province. The tissue samples were
placed in plastic freezer bags or 125 mL flasks and after the addition of 50 mL deionized (DI) or
sterilized tap water were shaken for 20 min. Two milliliters were collected and centrifuged at
10,000 g for 10 min to remove debris. The supernatants were either treated with chloroform or
filter-sterilized and then were checked for the presence of bacteriophages by spotting 20 µL onto
freshly prepared lawns of the indicator bacteria. In Florida three Xac strains were used for
detection: Xac65, a Miami type; Xac60, a Manatee type, and Xac66, a Wellington-type. In
Argentina a battery of 11 Xac strains of diverse origins plus one Xaa strain were used (1622-4,
1528-7-3, 1635, 1660-1, 94-358-1, 1319, 1617, 1604, 1322, 2525, 78-4-3-2-4B and 1311) (Table
3-1). If lysis was observed after 24 h incubation at 28°C, the phage was purified by three
successive single plaque isolations and then propagated and stored, as described above.

45
Phage Typing of 81 Xanthomonas Strains
Twelve phages were used in the phage typing study: α-MME, Φ5536, ΦXacm4-11,
ΦXv3-21, ΦXaacA1, ΦXaacF1, ΦXaacF8, Φcc19-1, Φcc13-2, CP1, CP2 and CP3 (Table 3-2).
The bacterium-phage interactions were scored as sensitive = 2, moderately sensitive = 1 and
resistant = 0. Similarity matrix was calculated from the phage typing scores using the Pearson
correlation, and a dendrogram of relatedness was prepared in which the clustering was achieved
by UPGMA (unweighted pair group method using arithmetic averages) with Bionumerics
software package version 3.0 (Applied Math, Kotrijk, Belgium).
Electron Microscopy
Transmission electron microscopy (TEM) was carried out at the Electron Microscopy
Laboratory of the Microbiology and Cell Science Department of the University of Florida,
Gainesville, FL. The phages were visualized using negative staining protocol with 1% aqueous
uranyl acetate, as follows. A drop of the phage suspension was applied to a 300 mesh formvar-
coated copper grid. After 2 min the liquid was blotted away and the grid was rinsed with DI
water. A 1% uranyl acetate solution was applied to the grid and blotted away after 1 min. The
phages were observed and photographed on a Zeiss EM-10CA transmission electron microscope
operating at 100 kV.
Molecular Techniques
Phage DNA extraction. Fifteen hundred microliters of concentrated and purified
bacteriophage suspensions (~1012 PFU/mL, SM buffer) were treated with nuclease (12.3
unit/sample DNase I (Qiagen Inc., Valencia, CA) and 6.3 units/sample RNase A (Qiagen), 30
min, 37ºC) to digest any contaminating bacterial nucleic acids. Subsequently, samples were
divided into 500 µL subsamples, placed in 1.5 mL microcentrifuge tubes, and 375 µL of a
phenol-chloroform-isoamyl alcohol mixture (25:24:1) was added. The samples were vortexed

46
and centrifuged for 5 min at 10,000 g. The top, aqueous layer was transferred into a new
microfuge tube and the organic layer was discarded. The sample volume was brought up to 500
µL again with the addition of sterile DI water and was subjected to phenol-chloroform-isoamyl
alcohol extraction two more times. Sterilized DI water was added to the sample to bring the
volume up to 500 µL and then the solution was subjected to chloroform-isoamyl alcohol
extraction once by adding 250 µL of a chloroform-isoamyl alcohol mixture (24:1), vortexing and
then centrifuging for 5 min at 10,000 g. The top, aqueous phase was saved and transferred to a
new tube, while the remainder was discarded. The DNA was then precipitated by adding 40 µL
sodium acetate (3 M, pH 5.2) and 800 µL of cold (-4°C) 95% ethanol to the tube, vortexing and
incubating at -80°C for 30 min. The sample was then centrifuged (10,000 g, 20 min) at 4°C and
the supernatant was discarded and replaced with 1 mL of 70% ethanol. After another
centrifugation (10,000 g, 5 min) the supernatant was gently removed by pipetting and the pellet
containing the precipitated DNA was allowed to air dry. The pellet was resuspended in 20 µL
sterile DI water and stored at 4°C.
Digestion of DNA with restriction enzymes. One microliter of the phage DNA was mixed
with 7 µL sterile DI water, 1 µL enzyme buffer and 1 µL restriction enzyme (EcoRI or BamHI)
in a 1.5 mL microcentrifuge tube. The mixture was incubated at 37°C for 90 minutes.
Immediately following the incubation, the entire mixture was subjected to agarose gel
electrophoresis (100) using 1% agarose gel (SeqKem GTG agarose, Cambrex Bio Science
Rockland Inc., Rockland, ME) to separate restriction fragments. EcoRI plus HinDIII digested
Lambda phage DNA (Promega Co., Madison, WI) was used as size marker. The gel was
photographed after 15 min ethidium bromide staining.

47
Results
Bacteriophage Isolations
Bacteriophages were isolated from leaf samples exhibiting citrus canker symptoms in
spring and summer 2003, in Florida. Phages were detected in 37 of the 138 samples (27%).
Bacteriophages were isolated from leaf and fruit tissues with characteristic citrus canker lesions
in December 2003 in Corrientes province, Argentina. Phages were detected in 30 of 56 samples
tested (54%). An attempt was made to isolate phages in Florida in 2006; none were detected in
62 samples.
Classification of Isolated Phages Based on Chloroform Sensitivity, Plaque Morphology, Host Range and Virion Morphology
Chloroform sensitivity, plaque morphology and host range were determined for each
newly isolated phage and for CP1, CP2 and CP3 phages originating in Japan. The phages
isolated in Florida comprised two groups. Thirty one of 37 phages (group A) were resistant to
chloroform, produced clear plaques of 2-3 mm in diameter on strain Xac65 and had identical
host ranges (Table 3-3). Virion morphology of four members of this group was determined: they
were all tailed phages with symmetrical heads and short tails (Figure 3-2, Table 3-4), which is
characteristic of phages in the Podoviridae family. The remaining six phages (group B) were
chloroform sensitive, produced 1 mm turbid plaques on Xac65 and had an identical host range,
which was different than the host range of group A phages. Virion morphology of one phage in
this group, ΦXaacF8, was found to be filamentous, similar to members of Inoviridae (Figure 3-1,
Table 3-4). The 30 phages collected in Argentina were all chloroform resistant, produced 3-4
mm clear plaques on Xac65 and had an identical host range, which only differed from the host
range of group A phages in the reaction on Argentina Xac strain BV42. The Argentina phages
lysed BV42 whereas the Florida group A phages did not (Table 3-3). One Argentina phage,

48
ΦXaacA1, belonged to Podoviridae based on virion morphology (Figure 3-1, Table 3-4). CP1,
CP2 and CP3 were all resistant to chloroform. CP1 did not lyse any strains tested, while CP3
only lysed its propagating strain, XC90. CP2 had the same host range as the Florida phage group
A, produced clear, 3 mm plaques on Xac65 (Table 3-3) and belonged to Podoviridae based on
virion morphology (Figure 3-2, Table 3-4).
Comparison of Florida Group A Phages and CP2 Based on Genome Size and RFLP Profile
Genome sizes of all 31 members of Florida phage group A and CP2 were identical and
were approximately 22 kb (Figure 3-3). However, the group A phages constituted seven groups
based on RFLP profile after BamHI digestion (Figures 3-4, 3-5, Table 3-5). The RFLP profile of
CP2 was different from that of all seven groups (Figure 2-4, panel B). Restriction profiles based
on EcoRI digestion resulted in six groups, five of which matched five BamHI groups; whereas
the sixth one contained two BamHI groups (data not shown). Virion morphology of members of
different BamHI groups, ΦXaacF1 (group 1), ΦXaacF2 (group 7), ΦXaacF3 (group 2) and
ΦXaacF5 (group 3) did not differ considerably (Figure 3-1, Table 3-4).
Phage Typing of 81 Xanthomonas Strains
Sixty nine xanthomonads of worldwide origin causing citrus canker were phage typed on a
battery of 12 phages: 49 Xac strains of pathotype A, three Xac strains of type A*, eight strains of
type B (Xaa) and nine strains of type C (Xaa). Also included were eight strains of X. axonopodis
pv. citrumelo (Xacm), causal agent of citrus bacterial spot, one strain each of X. perforans, X.
vesicatoria and X. euvesicatoria, causal agents of bacterial spot of tomato, and one strain of X.
axonopodis pv. dieffenbachiae, causal agent of anthurium bacterial blight.
In general, citrus canker strains of the same genotype did tend to cluster together (Figure
3-2). On the other hand, Xacm strains showed considerable diversity. Most of Etrog strains were
in one group (G1), clustering together with a Pakistani A strain. Two Etrog strains comprised an

49
independent group (G6), however. Most of the B strains clustered together (G2) and were
sensitive to phage CP3, however strains 6B, 7B (G7) and 8B differed from the rest and were not
sensitive to CP3. Wellington strains clustered together with the A* strains in groups G3 or G5.
The majority of Miami strains and the majority of A strains of various geographical origins
clustered together and were sensitive to at least 8 of 12 phages (G4). However, three Miami
strains and one Brazilian A strain constituted another group (G8) and were resistant to all but two
phages, including ΦXaacF1 and ΦXaacF8 that are naturally present in Florida. All C strains
clustered in group G9 and were resistant to CP1 and CP2. None of the Manatee strains were
sensitive to any phages tested (G11). On the other hand, strain ATCC 49118 (G10), which was
isolated from the 1980s Tampa Bay outbreak was sensitive to 4 phages. Two A strains, isolated
in Florida in 2005 (XC2005-00344-1) and in 2006 (XI2006-00204), have not been classified into
any of the Florida genotypes by genetic methods. One of them had an identical profile to most
Miami types (G4), whereas the other one was resistant to 11 of 12 phages tested. Host ranges of
phages CP2 and ΦXaacA1 were identical and were almost identical to that of ΦXaacF1: the only
difference was ΦXaacF1 was not able to lyse strain XC427 from Thailand.
Discussion
In this study bacteriophages were frequently associated with citrus canker lesions in
Argentina, where the disease is endemic. Also phages were widespread in Florida in 2003, where
citrus canker was under eradication and was located in relatively concise location, mainly in the
Miami-Palm Beach area. It seems that there are near identical population of phages around the
world, as all phages isolated from Argentina and the majority of the ones isolated in Florida were
almost identical to phage CP2 originating in Japan. It is possible that CP2, or a progenitor of CP2
spread around the world with the Xac strains. Interestingly, attempts to isolate phages in Florida
in 2006 were unsuccessful. Since 2003 citrus canker was spread by hurricanes widely within

50
Florida, and it may be that the bacteria managed to escape its pathogens in the process. Also, this
study showed the presence of strains resistant to the phages isolated in Florida and it is possible
that their prevalence resulted in decline of phage.
Results of this study also showed that phages can be used for distinguishing Xac genotypes
present in Florida. Our results generally supported the findings of Cubero et al. (26); however
based on phage typing results, the representative strain associated with the 1980s outbreak and
strains from the 1997 Manatee outbreak were different. This finding contradicts the theory that
the 1997 citrus canker outbreak in Manatee County was caused by strains of the 1980s outbreak
that survived the eradication program. Strain XC2005-00344-1, which is an A strain isolated in
Polk County in 2005 and is unclassified genetically, was resistant to 11 of 12 phages, which
indicates the possibility of a population shift to phage resistance. That in turn could mean that the
naturally occurring phages pose a selection pressure on the pathogen populations, that is, they
have a significant impact on the pathogen’s life. Thus, artificial introduction of exotic
bacteriophages, phage therapy, may well be effective for reducing pathogen populations and
suppressing disease.
The slight differences in host ranges of the Argentina phages, Florida group A phages and
CP2 could be caused by minor differences in phage DNA sequences: the presence or absence of
DNA sequence patterns in the phage genome that are recognized by the host bacterial
restriction-modification system.

51
Table 3-1. Bacterial strains used in this study Strain Origin Genotypea Provided by X. axonopodis pv. citri 1311 Pakistan Canteros, B.I.b
1319 India Canteros, B.I. 1322 Brazil Canteros, B.I. 1604 Argentina Canteros, B.I. 1617 Argentina Canteros, B.I. 1635 Argentina Canteros, B.I. 2525 Argentina Canteros, B.I. 1528-7-3 Argentina Canteros, B.I. 1622-4 Argentina Canteros, B.I. 1660-1 Paraguay Canteros, B.I. 306 Brazil Hartung, J.S.c
94-358-1 Argentina Canteros, B.I. BV38 Argentina Canteros, B.I. BV42 Argentina Canteros, B.I. RF2f Florida Miami This study Xac5 Florida Miami Sun, X.d
Xac6 Florida Miami Sun, X. Xac7 Florida Miami Sun, X. Xac8 Florida Manatee Sun, X. Xac15 Florida Wellington Sun, X. Xac25 Florida Miami Sun, X. Xac30 Florida Miami Sun, X. Xac31 Florida Miami Sun, X. Xac41 Florida Manatee Sun, X. Xac51 Florida Miami Sun, X. Xac65 Florida Miami Sun, X. Xac66 Florida Wellington Sun, X. Xac71 Florida Wellington Sun, X. Xc05-2592 Argentina Canteros, B.I. XC62 Japan Hartung, J.S. XC63 Japan Hartung, J.S. XQ05-1-2 Florida Miami Sun, X. X. axonopodis pv. aurantifolii 78-4-3-2-4B Argentina Canteros, B.I. XC 90 Mexico Hartung, J.S. X. axonopodis pv. citrumelo Xacm 36 Florida This study Xacm 45 Florida This study Xacm 47 Florida This study S4 Florida This study

52
Table 3-1. Continued Strain Origin Genotypea Provided by S4mg Florida This study X. vesicatoria MME Jones, J.B.e
X. perforans ΔopgH:ME-B Jones, J.B.
a Genotype of X. axonopodis pv. citri strains isolated in Florida. b Instituto Nacional de Tecnología Agropecuaria, Bella Vista, Corrientes, Argentina. c United States Department of Agriculture, Agricultural Research Service, Fruit Laboratory, Beltsville, MD. d Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL. e University of Florida, Plant Pathology Department, Gainesville, FL. f RF2 is a phage-resistant mutant of Xac65. g Chloramphenicol and Rifamycin-resistant mutant of S4.

53
Table 3-2. Bacteriophages used in the study Phage Host Source Origin Provided by CP1 XC62 Japan Hartung, J.S.a
CP2 XC63 Japan Hartung, J.S. CP3 XC90 soil Japan Hartung, J.S. CP31 Xac65 Hartung, J.S. ΦXaacF1 to 37 Xac65 citrus, aerial Florida This study ΦXaacA1 to 30 BV42 citrus, aerial Argentina This study ccΦ5 Xac65 Jackson, L.E.b
ccΦ7 Xac65 Jackson, L.E. ccΦ13-2 Xac65 Jackson, L.E. ccΦ19-1 Xac65 Jackson, L.E. ccΦ24 Xac65 Jackson, L.E. ccΦ25 Xac65 Jackson, L.E. ccΦ28 Xac65 Jackson, L.E. ccΦ35 Xac65 Jackson, L.E. ccΦ36 Xac65 Jackson, L.E. ΦXV3-21 Xac65 Jackson, L.E. α-MME MME lake water Florida Minsavage, G.V.c
Φ5536 Xacm 47 Jackson, L.E. ΦXv3-3-13h ΔopgH:ME-B Jackson, L.E. ΦX44 Xacm47 Jackson, L.E. ΦXacm4-4 Xacm36 citrus, aerial Florida This study ΦXacm4-16 Xacm36 citrus, aerial Florida This study ΦXacm4-11 Xacm36 citrus, aerial Florida This study
a United States Department of Agriculture, Agricultural Research Service, Fruit Laboratory, Beltsville, MD. b OmniLytics, Inc., Salt Lake City, UT. c University of Florida, Plant Pathology Department, Gainesville, FL.

54
Table 3-3. Chloroform sensitivity, plaque morphology and host range of bacteriophages originating in Florida, Argentina and Japan
Chloroform Plaque typeb, Host ranged
sensitivity diameter (mm) BV38 XC90 Xac41 Xac65 Xac15 ΦXaacF1 Ra clear, 2-3 -c - ++ ++ - ΦXaacF2 R clear, 2-3 - - ++ ++ - ΦXaacF3 R clear, 2-3 - - ++ ++ - ΦXaacF4 R clear, 2-3 - - ++ ++ - ΦXaacF5 R clear, 2-3 - - ++ ++ - ΦXaacF6 R clear, 2-3 - - ++ ++ - ΦXaacF7 S turbid, 1 ++ - - ++ ++ ΦXaacF8 S turbid, 1 ++ - - ++ ++ ΦXaacF9 S turbid, 1 ++ - - ++ ++ ΦXaacF10 R clear, 2-3 - - ++ ++ - ΦXaacF11 S turbid, 1 ++ - - ++ ++ ΦXaacF12 R clear, 2-3 - - ++ ++ - ΦXaacF13 S turbid, 1 ++ - - ++ ++ ΦXaacF14 R clear, 2-3 - - ++ ++ - ΦXaacF15 R clear, 2-3 - - ++ ++ - ΦXaacF16 R clear, 2-3 - - ++ ++ - ΦXaacF17 R clear, 2-3 - - ++ ++ - ΦXaacF18 R clear, 2-3 - - ++ ++ - ΦXaacF19 R clear, 2-3 - - ++ ++ - ΦXaacF20 R clear, 2-3 - - ++ ++ - ΦXaacF21 R clear, 2-3 - - ++ ++ - ΦXaacF22 R clear, 2-3 - - ++ ++ - ΦXaacF23 R clear, 2-3 - - + ++ - ΦXaacF24 R clear, 2-3 - - ++ ++ - ΦXaacF25 S turbid, 1 ++ - - ++ ++ ΦXaacF26 R clear, 2-3 - - ++ ++ - ΦXaacF27 R clear, 2-3 - - ++ ++ - ΦXaacF28 R clear, 2-3 - - ++ ++ - ΦXaacF29 R clear, 2-3 - - ++ ++ - ΦXaacF30 R clear, 2-3 - - ++ ++ - ΦXaacF31 R clear, 2-3 - - ++ ++ - ΦXaacF32 R clear, 2-3 - - ++ ++ - ΦXaacF33 R clear, 2-3 - - ++ ++ - ΦXaacF34 R clear, 2-3 - - ++ ++ - ΦXaacF35 R clear, 2-3 - - ++ ++ - ΦXaacF36 R clear, 2-3 - - ++ ++ - ΦXaacF37 R clear, 2-3 - - ++ ++ - ΦXaacA1 R clear, 3-4 ++ - ++ ++ - ΦXaacA2 R clear, 3-4 ++ - ++ ++ - ΦXaacA3 R clear, 3-4 ++ - ++ ++ - ΦXaacA4 R clear, 3-4 ++ - ++ ++ - ΦXaacA5 R clear, 3-4 ++ - ++ ++ - ΦXaacA6 R clear, 3-4 ++ - ++ ++ - ΦXaacA7 R clear, 3-4 ++ - ++ ++ - ΦXaacA8 R clear, 3-4 ++ - ++ ++ - ΦXaacA9 R clear, 3-4 ++ - ++ ++ -

55
Table 3-3. Continued Chloroform Plaque typeb, Host ranged sensitivity diameter (mm) BV38 XC90 Xac41 Xac65 Xac15 ΦXaacA10 R clear, 3-4 ++ - ++ ++ - ΦXaacA11 R clear, 3-4 ++ - ++ ++ - ΦXaacA12 R clear, 3-4 ++ - ++ ++ - ΦXaacA13 R clear, 3-4 ++ - ++ ++ - ΦXaacA14 R clear, 3-4 ++ - ++ ++ - ΦXaacA15 R clear, 3-4 ++ - ++ ++ - ΦXaacA16 R clear, 3-4 ++ - ++ ++ - ΦXaacA17 R clear, 3-4 ++ - ++ ++ - ΦXaacA18 R clear, 3-4 ++ - ++ ++ - ΦXaacA19 R clear, 3-4 ++ - ++ ++ - ΦXaacA20 R clear, 3-4 ++ - ++ ++ - ΦXaacA21 R clear, 3-4 ++ - ++ ++ - ΦXaacA22 R clear, 3-4 ++ - ++ ++ - ΦXaacA23 R clear, 3-4 ++ - ++ ++ - ΦXaacA24 R clear, 3-4 ++ - ++ ++ - ΦXaacA25 R clear, 3-4 ++ - ++ ++ - ΦXaacA26 R clear, 3-4 ++ - ++ ++ - ΦXaacA27 R clear, 3-4 ++ - ++ ++ - ΦXaacA28 R clear, 3-4 ++ - ++ ++ - ΦXaacA29 R clear, 3-4 ++ - ++ ++ - ΦXaacA30 R clear, 3-4 ++ - ++ ++ - CP1 R clear, 4-5 - - - - - CP2 R clear, 3 - - ++ ++ - CP3 R clear, 1 - ++ - - -
Summary 32 phages R clear - - ++ ++ - 30 phages R clear, 3-4 ++ - ++ ++ - 6 phages S turbid, 1 ++ - - ++ ++ 1 phage R clear, 1 - ++ - - - 1 phage R clear, 4-5 - - - - - a S: sensitive, R: resistant. b Plaque morphology was evaluated on the propagating host (Table 3-2) after 36 h. c ++: sensitive, -: resistant. d Strains Xacm36, Xacm45, Xacm47 and 306 were resistant to all phages. Strain BV42 had identical profile to Xac65.

56
A B
C D
E F
G
Figure 3-1. Transmission electron micrographs of representative phages. A) CP2, B) ΦXaacA1, C) ΦXaacF1, D) ΦXaacF2, E) ΦXaacF3, F) ΦXaacF5, G) ΦXaacF8.

57
Table 3-4. Summary of morphological characteristics of phage virions.
Phage Head shape Head
(Lengtha, nm) Tail shape Classificationb
CP2 symmetrical 50.8 short Podoviridae ΦXaacA1 symmetrical 53.3 short Podoviridae ΦXaacF1 symmetrical 58.3 short Podoviridae ΦXaacF2 symmetrical 51.2 short Podoviridae ΦXaacF3 symmetrical 56.6 short Podoviridae ΦXaacF5 symmetrical 62.5 short Podoviridae ΦXaacF8 Inoviridae
a Length measurement represents the average value obtained by measuring three virions per phage. b Classification based on virion morphology as described by Ackermann Fig. 4.1 (1).

58
100
806040200 Phi
Xac
m4-
11
Phi
Xv3
-21
ccP
hi19
-1
Phi
5536
Phi
Xaa
cF8
CP2
Phi
Xaa
cA1
Phi
Xaa
cF1
alph
a-M
ME
CP3
ccP
hi13
-2
CP1
XN2003-00012-7XN2004-00225-2
XS2004-00159
XC2005-00344-1
XN2003-00012-3XN2003-00013-2
X2001-00042XN2003-00011-1
XN2003-00011-2
XN2003-00011-8
XC159LMG 7484
XS1999-00061
JH96E
XC64
JH96-2XCC88B
JH96CXC90
S4
XC2005-00110XC2000-00042
X2000-12884
X2001-00006
X2003-01008
XC165X2001-00032
XC261
XN2003-01036
X2000-12862X2000-00071
X2003-03516SNU6B
X1999-12813
XC392
XCC03-1635XI2000-00075
XI2000-00080
XI2000-00120
XI2006-00204
XS1999-00038XC214
XC100X2003-02912
XC2002-00010XI2000 00194
Xac A EtrogXacm
Xacm
Xac A Miami
Xac A EtrogXac A Etrog
XacmXac A Etrog
Xac A Etrog
Xac A Etrog
Xac AXa diffenbachiae
Xacm
Xaa B
Xaa B
Xaa BXaa B
Xaa BXaa D
Xacm
XacmXacm
Xac A Wellington
Xac A Wellington
Xac A Wellington
Xac A*Xac A Wellington
Xac A*
Xac A Wellington
XacmXac A Miami
Xac A MiamiXac A
Xac A Miami
Xac A
Xac AXac A Miami
Xac A Miami
Xac A Miami
Xac A Miami
Xac A MiamiXac A
Xac AXac A Miami
Xac A MiamiX A Mi i
FloridaFlorida
Florida
Florida
FloridaFlorida
FloridaFlorida
Florida
Florida
Pakistan
Florida
Argentina
Japan
ArgentinaArgentina
ArgentinaMexico
Florida
FloridaFlorida
Florida
Florida
Florida
IndiaFlorida
Saudi Arabia
Florida
FloridaFlorida
FloridaKorea
Florida
Indonesia
ArgentinaFlorida
Florida
Florida
Florida
FloridaTaiwan
YemenFlorida
Florida
Fl id Figure 3-2. Dendogram and phage sensitivity matrix showing relationship amongst
Xanthomonas strains causing citrus canker and citrus bacterial spot based on similarity of sensitivity profile against a battery of 12 phages. Typing phages: 1:ΦXacm4-11, 2:ΦXV3-21, 3:ccΦ19-1, 4:Φ5536, 5:ΦXaacF8, 6:ΦCP2, 7:ΦXaacA1, 8:ΦXaacF1, 9:α-MME, 10:CP3, 11:ccΦ13-2, 12:CP1. Percent similarity values were calculated using the Pearson correlation, and clustering was achieved by UPGMA using Bionumerics software package version 3.0. Black rectangle = sensitive; grey rectangle = moderately sensitive; no rectangle = resistant. Xac=Xanthomonas axonopodis pv. citri; Xaa: Xanthomonas axonopodis pv. aurantifolii; Xacm= Xanthomonas axonopodis pv. citrumelo.
Strain Class Origin
G1
G2
G4
G3
Typing Phages_____ 1 2 3 4 5 6 7 8 9 10 11 12
kjkj
Percent Similarity

59
100
806040200 PhiX
acm
4-11
PhiX
v3-2
1
ccPh
i19-
1
Phi5
536
PhiX
aacF
8
CP
2
PhiX
aacA
1
PhiX
aacF
1
alph
a-M
ME
CP
3
ccPh
i13-
2
CP
1
X2003-02912
XC2002-00010
XI2000-00194
X2003-00008
XC63
XC273
XN2003-01035
XN2003-00012-1XN2003-00012-6
6B
7B
306
X2000-00067XI1999-00112
XI2001-00098
10537C
17C
1C
2C
3C
4C
JH70C
MME
10535C
IAPAR9691/90
91-106
91-118
XC205
8B
XC427
ATCC 49118
XC362
XC62
XS1995-00051
XS1997-00006
XS1997-00018
XS1997-00082
XS2003-00004
Xac A Miami
Xac A Miami
Xac A Miami
Xac A Miami
Xac A
Xac A*
Xac A Wellington
Xac A EtrogXac A Etrog
Xaa B
Xaa B
Xac A
Xac A MiamiXac A Miami
Xac A Miami
Xaa C
Xaa C
Xaa C
Xaa C
Xaa C
Xaa C
Xaa C
X. vesicatoria
Xaa C
Xaa C
X. euvesicatoria
X. perforans
Xac A
Xaa B
Xac A
Xac A
Xac A
Xac A
Xac A Manatee
Xac A Manatee
Xac A Manatee
Xac A Manatee
Xac A Manatee
Florida
Florida
Florida
Florida
Japan
Saudi Arabia
Florida
FloridaFlorida
Argentina
Argentina
Brazil
FloridaFlorida
Florida
Brazil
Brazil
Brazil
Brazil
Brazil
Brazil
Brazil
Brazil
Brazil
Taiwan
Argentina
Thailand
Florida
Australia
Japan
Florida
Florida
Florida
Florida
Florida Figure 3-2. Continued
Strain Class Origin
G4
G5
G6
G7
G8
G9
G11
G10
Typing Phages_____ 1 2 3 4 5 6 7 8 9 10 11 12Percent Similarity
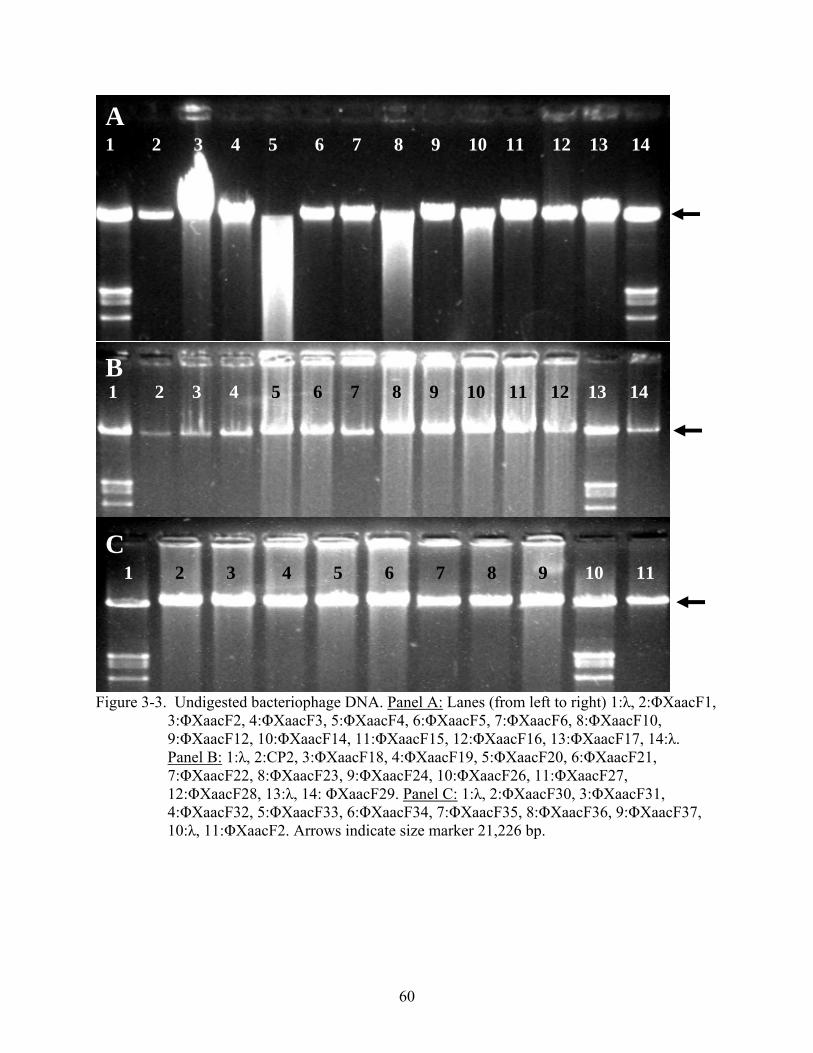
60
Figure 3-3. Undigested bacteriophage DNA. Panel A: Lanes (from left to right) 1:λ, 2:ΦXaacF1,
3:ΦXaacF2, 4:ΦXaacF3, 5:ΦXaacF4, 6:ΦXaacF5, 7:ΦXaacF6, 8:ΦXaacF10, 9:ΦXaacF12, 10:ΦXaacF14, 11:ΦXaacF15, 12:ΦXaacF16, 13:ΦXaacF17, 14:λ. Panel B: 1:λ, 2:CP2, 3:ΦXaacF18, 4:ΦXaacF19, 5:ΦXaacF20, 6:ΦXaacF21, 7:ΦXaacF22, 8:ΦXaacF23, 9:ΦXaacF24, 10:ΦXaacF26, 11:ΦXaacF27, 12:ΦXaacF28, 13:λ, 14: ΦXaacF29. Panel C: 1:λ, 2:ΦXaacF30, 3:ΦXaacF31, 4:ΦXaacF32, 5:ΦXaacF33, 6:ΦXaacF34, 7:ΦXaacF35, 8:ΦXaacF36, 9:ΦXaacF37, 10:λ, 11:ΦXaacF2. Arrows indicate size marker 21,226 bp.
A
C
B
1 2 3 4 5 6 7 8 9 10 11 12 13 14
1 2 3 4 5 6 7 8 9 10 11
1 2 3 4 5 6 7 8 9 10 11 12 13 14

61
Figure 3-4. Bacteriophage DNA digested with BamH I. Panel A: Lanes (from left to right) 1:λ,
2:ΦXaacF1, 3:ΦXaacF2, 4:ΦXaacF3, 5:ΦXaacF4, 6:ΦXaacF5, 7:ΦXaacF6, 8:ΦXaacF10, 9:ΦXaacF12, 10:ΦXaacF14, 11:ΦXaacF15, 12:ΦXaacF16, 13:ΦXaacF17, 14:λ. Panel B: 1:λ, 2:ΦXaacF18, 3:ΦXaacF19, 4:ΦXaacF20, 5:ΦXaacF21, 6:ΦXaacF22, 7:ΦXaacF23, 8:ΦXaacF24, 9:ΦXaacF26, 10:CP2, 11:ΦXaacF27, 12:ΦXaacF28, 13: ΦXaacF29, 14: ΦXaacF30, 15: ΦXaacF31, 16:λ. Panel C: 1:ΦXaacF32, 2:ΦXaacF33, 3:ΦXaacF34, 4:ΦXaacF35, 5:λ, 6:ΦXaacF36, 7:ΦXaacF37, 8:ΦXaacF2. Arrows indicate size marker 21,226 bp.
A
C
B
1 2 3 4 5 6 7 8 9 10 11 12 13 14
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
1 2 3 4 5 6 7 8

62
Table 3-5. Grouping of Florida bacteriophages based on BamH I RLFP profile. Group 1 Group 2 Group 3 Group 4 Group 5 Group 6 Group 7 ΦXaacF1 ΦXaacF3 ΦXaacF5 ΦXaacF27 ΦXaacF31 ΦXaacF34 ΦXaacF2ΦXaacF6 ΦXaacF4 ΦXaacF32 ΦXaacF36 ΦXaacF12 ΦXaacF10 ΦXaacF33 ΦXaacF37 ΦXaacF15 ΦXaacF14 ΦXaacF16 ΦXaacF17 ΦXaacF18 ΦXaacF30 ΦXaacF19 ΦXaacF20 ΦXaacF21 ΦXaacF22 ΦXaacF23 ΦXaacF24 ΦXaacF26 ΦXaacF28
Figure 3-5. Representatives of the 7 RFLP groups based on BamH I. digestion profile. Lanes
(from left to right) 1:λ, 2:ΦXaacF24 (group 1), 3:ΦXaacF30 (group 2), 4:ΦXaacF5 (group 3), 5:ΦXaacF27 (group 4), 6:ΦXaacF31 (group 5), 7:ΦXaacF34 (group 6), 8:ΦXaacF2 (group 7), 9:λ. Arrow indicates size marker 21,226 bp.
1 2 3 4 5 6 7 8 9

63
CHAPTER 4 CONTROL OF CITRUS CANKER AND CITRUS BACTERIAL SPOT WITH
BACTERIOPHAGES
Introduction
The traditional control strategies for managing bacterial plant diseases have utilized
chemical control with reliance on antibiotics and copper bactericides (3,81,119). However, the
use of antibiotics has been largely discontinued in agriculture due to the emergence of antibiotic-
resistant bacteria in the field (119) and due to concerns that phytobacteria can harbor and transfer
antibiotic resistance to human pathogens. The extended heavy use of copper compounds led to
soil contamination (72) and the emergence of copper-tolerant strains amongst phytobacteria (81)
led to reduced efficacy of copper treatments. High doses of ionic copper also can cause
phytotoxicity (85,109). These factors have prompted a search for identifying novel, effective
means for managing bacterial diseases of plants. Additionally, concerns about food safety and
environmental protection, and the growing organic production necessitate development of safer,
more specific and environment-friendly pesticides (96).
Bacteriophages have been used effectively for controlling several Xanthomonas diseases.
Severity of bacterial spot of peach, caused by X. campestris pv. pruni, has been reduced both on
foliage (23) and on fruits (99) with phage treatment. Phage applications decreased geranium
bacterial blight severity, caused by X. campestris pv. pelargonii (37). Tomato bacterial spot
disease severity was reduced and fruit yield was increased by phage applications (10,38,89,90).
Phage-based integrated management of tomato bacterial spot is recommended to tomato growers
in Florida (85), and bacteriophage mixtures against the pathogen are commercially available
(Agriphage from OmniLytics Inc., Salt Lake City, UT, EPA Registration # 67986-1). However,
phage therapy has not been evaluated for the control of citrus diseases.

64
Our objective was to evaluate phage therapy for treatment of citrus canker. Additionally,
we wished to investigate the effect of protective formulations on efficacy of bacteriophage
treatment and also to use phages in combination with chemical bactericides as a part of an
integrated approach. Skim milk-based protective formulations have been used to increase phage
persistence on plant foliage and contributed to enhanced control of tomato bacterial spot (10).
Ionic copper is toxic to all cells, since it interacts with sulfhydryl (-SH) groups of amino acids
and causes denaturation of proteins ((3), p.345). Ionic copper reduces bacteriophage populations
(11, 66); however, it only persists on plant foliage a short time and does not affect bacteriophage
survival if applied three or more days in advance of phage application (66).
Citrus canker until recently has been under eradication in Florida, and field trials in groves
could not be carried out in the state. Therefore, we conducted disease control trials (i) in the
greenhouse of the citrus canker quarantine facility of the Florida Department of Agriculture &
Consumer Services, Division of Plant Industry (DPI) and (ii) in Argentina, where disease canker
is endemic in cooperation with researchers of the Instituto Nacional de Tecnología Agropecuaria
(INTA). Additionally, we carried out field trials in Florida for suppressing citrus bacterial spot
(CBS), another Xanthomonas disease of citrus (102). CBS, incited by X. axonopodis pv.
citrumelo (Xacm), is similar to citrus canker, as both diseases cause lesions on citrus leaves and
stems, infect young tissue, spread by wind driven rain and their severity is greatly increased by
Asian leafminer (Phyllocnistis citrella) tunneling activity. CBS differs from citrus canker in that
it does not affect full grown plants, causes flat or sunken lesions (Figure 1-8) instead of the
corky, raised ones of canker (Figure 1-1) and is generally less aggressive than citrus canker
(106). Additionally, CBS is not regulated allowing the implementation of field trials.

65
Materials and Methods
Bacterial Strains and Bacteriophages
Bacterial strains (Table 3-2) were grown on nutrient agar (NA) medium (0.8% (wt/V)
Nutrient Broth (NB) (BBL, Becton Dickinson and Co., Cockeysville, MD) and 1.5% (wt/V)
Bacto Agar (Difco, Becton Dickinson and Co., Sparks, MD)) at 28°C. For preparation of
bacterial suspensions 24 h cultures were suspended in sterile tap water, their concentration was
adjusted to 5×108 cfu/mL (A600=0.3), and then were diluted appropriately. Bacterial inoculations
were carried out by misting the bacterial suspension on the plant foliage with a hand-held
sprayer. For detection and propagation of bacteriophages (Table 3-1) either semisolid, soft
nutrient agar yeast extract medium (NYA), (0.8% Nutrient Broth, 0.6% Bacto Agar and 0.2%
Yeast Extract (Difco, Becton Dickinson and Co., Sparks, MD)), or liquid nutrient broth medium
was used. Sterilized tap water or SM buffer (0.05 M Tris-HCl (pH 7.5), 0.1 M NaCl, 10 mM
MgSO4 and 1% (w/V) gelatin) was used for preparing phage suspensions. Bacterial strains used
in this study (Table 3-2) were stored at -80ºC in NB supplemented with 30% glycerol.
Bacteriophages were stored at 4ºC and protected from light.
Plant Material and Cultural Conditions in the Greenhouse
Duncan grapefruit plants were grown from seed in 15 cm plastic pots in Terra-Lite®
agricultural mix (Scott Sierra Horticultural Products Co., Marysville, OH) and fertilized as
needed using Osmocote® Outdoor & Indoor Smart Release® Plant Food (Scott Sierra
Horticultural Products Co.) controlled release fertilizer (NPK 19-6-12). The plants were kept in
the glasshouse of the citrus canker quarantine facility of the Division of Plant Industry in
Gainesville, Florida at temperature ranging from 25-30ºC.

66
Standard Bacteriophage Techniques
Determination of titer. Phage concentrations were determined by the dilution-plating-
plaque count assay on NYA plates without bottom agar as previously described (97).
Propagation. Phages were recovered from storage, purified by single plaque isolations and
then mass streaked on the freshly prepared lawn of the propagating host. The following day the
phages were eluted from the plate, sterilized and enumerated. The eluate was used for infecting
500 mL of actively growing culture of the propagating strain (108 cfu/mL) grown in NB liquid
medium in 1 liter flasks, at 0.1 multiplicity of infection (MOI). After addition of the phage and 5
min incubation on the bench top, the culture was shaken at 150 rpm at 28ºC for 18 h. Then the
culture was sterilized, enumerated and stored at 4 ºC in the dark until use. This method yielded
phage titers of approximately 1010 PFU/mL.
Phage concentration by high speed centrifugation. High titer phage lysates (~1010
PFU/mL) were concentrated and purified according to methods described by Hans-W.
Ackermann (personal communication). One hundred milliliters of the sterilized lysate was
centrifuged at 10,000 g for 10 min to sediment the bacterial debris. Forty milliliters of the
supernatant was transferred to a new centrifuge tube and centrifuged at 25,000 g for 60 min to
sediment the phage particles. The supernatant was discarded and replaced with 0.1M ammonium
acetate solution (pH 7.0). Following an additional centrifugation (60 min, 25,000 g) the
supernatant was discarded and the pellet was resuspended in 1.5 mL of SM buffer. The final
phage concentration was approximately 1012 PFU/mL.
Disease Assessment and Data Analysis
The ratio of diseased leaf surface area was estimated using the Horsfall-Barratt (HB) scale
(12): 1=0%, 2=0-3%, 3=3-6%, 4=6-12%, 5=12-25%, 6=25-50%, 7=50-75%, 8=75-87%, 9=87-
94%, 10=94-97%, 11=97-100%, 12=100%. The HB values were converted to estimated mean

67
percentages by using the Elanco Conversion Tables for Barratt-Horsfall Rating Numbers
(ELANCO Products Co., Indianapolis, IN), as described in Appendix A. If disease was assessed
more than once, the area under the disease progress curve (AUDPC) was computed by
trapezoidal integration of disease percentage values taken at different timepoints using the
formula proposed by Shaner and Finnley (104), as described in Appendix A.
Analysis of variance (ANOVA) and subsequent separation of sample means by the Waller-
Duncan K-ratio t Test (for balanced data) or by least square means method (for unbalanced data)
was carried out using SAS System for Windows program release 8.02 (Cary, NC).
Greenhouse Citrus Canker Control Trials
Control with phage mixture applied with or without skim milk formulation. Duncan
grapefruit plants were heavily pruned and fertilized to induce a new flush of growth that is highly
susceptible to citrus canker infection. Approximately three weeks later the emerging-uniform-
new-foliage was treated with a equal mixture of phages CP2, CP31, ccΦ7 and ccΦ13-2 (test 1 -
5×109 PFU/mL, test 2 - 1×109 PFU/mL, test 3 - 1×108 PFU/mL of each phage) in the evening
and the plants were covered with white plastic bags. Phages were applied with or without skim
milk formulation (0.75% (wt/V) non-fat dry milk powder). The goal of placing in plastic bags
was to maintain high relative humidity, simulate dew conditions and also to contribute to the
opening of the stomata that increases the penetration of bacteria into the leaf tissues. On the next
morning the bags were removed from the plants, and the plants were inoculated with Xac65 (test
1 - 1×108 cfu/mL, tests 2 and 3 - 5×106 cfu/mL). After inoculation the plants were placed inside
the bags for an additional 24 h of high moisture. After removal from the bags and after the
foliage was allowed to dry the plants were arranged in a completely randomized pattern on a
greenhouse bench. The disease was assessed 3-4 weeks after inoculation. Four plants were used
per treatment.

68
Control of disease caused by phage-sensitive strain and its phage-resistant mutant. A
mutant of Xac65, designated RF2, which was selected for resistance to phage ΦXaacF2, was
found also to be resistant to CP2, CP31, ccΦ7 and ccΦ13-2. In three tests grapefruit plants were
inoculated either with Xac65 or RF2, at 5×106 cfu/mL 12 h after treatment with the mixture of
CP2, CP31, ccΦ7 and ccΦ13-2, each at 1×108 PFU/mL. These tests were carried out similarly to
the previous ones.
Citrus Canker Nursery Trials in Argentina
The trials were conducted at the experimental nursery of the Instituto Nacional de
Tecnología Agropecuaria (INTA) research center in Bella Vista, Corrientes (Figure 4-1) from
January to April 2006 in cooperation with scientists of INTA. The experiment was set up in a
split-plot design: there were two main treatments, phage and no phage, and within them there
were two sub-treatments, copper-mancozeb and no copper-mancozeb. The sub-treatments were
randomly replicated four times within the main treatments. The replications were 5-m long plots.
The main plots were replicated twice. The plants were inoculated with a copper-resistant Xac
strain, Xc05-2592 in January 2006 and then re-inoculated in February. The phage treatment
(ΦXaacA1, ~106 PFU/mL) was mixed with a protective formulation (0.75% (wt/V) non-fat dry
milk powder) and spray-applied twice weekly in. Copper-mancozeb (3 g/L Caurifix WG (BASF)
(a.i. 84% copper oxychloride) plus 2 g/L Dithane M80 (a.i. 80% mancozeb)) was also spray-
applied twice weekly. Disease assessment was carried out once, after 12 weeks of phage and
copper-mancozeb applications. All diseased leaves were collected and the average number of
lesions per leaf was determined. This experiment was carried out at two sites at the nursery
simultaneously.

69
Citrus Bacterial Spot Disease Control Trials at Dilley and Son Nursery in Avon Park, Florida
The experiment was carried out at Roland L. Dilley & Son, Inc., a citrus nursery in Avon
Park, FL (Figure 4-2), which had a long history of CBS, caused by X. axonopodis pv. citrumelo.
The cultural methods and the high plant density in the nursery resulted in active plant growth,
long leaf-wetness periods and easy pathogen spread, all contributing to CBS development. Three
test sites were established in the nursery: two were set in a section containing Valencia orange
scions on Volkamer lemon (Citrus volkameriana) rootstock (Figure 4-2), and the third one was
set in a section of Ray Ruby grapefruit scions on Cleopatra mandarin (C. reticulata Blanco)
rootstocks. In each test site a row was divided into six 6-m-long plots, spaced 3 m apart, with
three receiving phage-treatment (phage), and three not (control). The phage-treated plots were
sprayed twice-weekly at dawn with a mixture of three bacteriophages (ΦXacm2004-4,
ΦXacm2004-16 (both isolated from the nursery), and ΦX44 (OmniLytics Inc., Salt Lake City,
UT), 2.4 × 108 PFU/mL each) blended in a suspension of 0.75% (wt/V) non-fat dry milk powder
using a non-CO2, backpack sprayer. The treatment was applied from August 9 until November
11, 2004. The high winds associated with the hurricanes caused a great deal of physical damage
to the plants opening up entry for pathogen ingress. Disease severity, caused by natural
inoculum, was evaluated five times during the experiment (biweekly between September 16 and
November 11).
Citrus Bacterial Spot Disease Control Trials at the Plant Science Unit of the University of Florida in Citra, Florida.
Two trials were carried out during summer and fall of 2006 at the Plant Science Unit of the
University of Florida in Citra, Florida (Figure 4-3). Swingle citrumelo plants, donated by Dilley
and Son, Inc. (Avon Park, FL) were transplanted in April 2005 and were fertilized monthly with
Osmocote® Outdoor & Indoor Smart Release® Plant Food (Scott Sierra Horticultural Products

70
Co., Marysville, OH) and irrigated as needed using an overhead irrigation system. The
experiment was set up in a split-plot design: there were two main treatments, phage and no
phage, and within them there were two sub-treatments, copper-mancozeb and no treatment. The
sub-treatments were randomly replicated five times within the main treatments. Each replicate
was one plot consisting of nine plants spaced 25 cm apart. The plots were separated by 2 m. The
plants were pruned and heavily fertilized to achieve uniform emerging young foliage before
inoculation with S4m, an antibiotic resistant mutant of a Xacm strain, S4 (Table 3-2). Inoculation
was carried out by spraying plants with the suspension of the pathogen (107 cfu/mL) amended
with 0.025% Silwet L-77 (Loveland Industries, Co., Greely, CO), a wetting agent, for increased
inoculum penetration. In the first trial every plant was inoculated, and pruned back after
developing disease symptoms, and then the emerging foliage rated for disease. This was done to
simulate the situation similar to that of the Dilley’s nursery, where the pathogen is present in the
nursery but is not directly inoculated on the plants. Since the disease pressure was extremely low
using this method, in the second trial the middle plant of every plot was inoculated and then the
disease was assessed on the inoculated foliage.
Phage and copper-mancozeb applications started before inoculations and continued until
the last rating was taken. The phage treatment was applied twice weekly in the evenings. It
consisted of a mixture of three phages, (ccΦ13-2, α-MME and Φ5536, 1×108 PFU/mL each)
applied with skim milk formulation (0.75% (wt/V) non-fat dry milk powder) using a non-CO2,
backpack sprayer. The copper-mancozeb treatment (2.5 g/L Manzate 75DF (Griffin Co.,
Valdosta, GA) (a.i. 75% mancozeb) and 3.6 g/L Kocide 2000 ( E. I. du Pont de Nemours & Co.)
(a.i. 53.8% copper hydroxide)) was applied once a week, 2 days apart from the phage

71
applications. Disease severity was assessed weekly using the Horsfall-Barratt scale, as described
above.
Results
Effect of Bacteriophage Application and Use of Protective Skim Milk Formulation on Citrus Canker Disease Development in the Greenhouse
In order to evaluate whether the presence of high populations of bacteriophages on the
grapefruit foliage suppresses citrus canker disease development, a mixture of four bacteriophages
(CP2, CP31, ccΦ7 and ccΦ13-2) was applied on grapefruit plants 12 h before inoculation with X.
axonopodis pv. citri (Xac) strain, Xac65. The phage mixture was applied with or without a skim
milk formulation. Three tests were carried out and in two of them there were significant
differences between treatments (Table 4-1). In both of those tests the application of phage
mixture without skim milk significantly reduced disease severity (Table 4-1). No phages could
be recovered two days after application from plants if they were applied without skim milk
formulation, whereas if applied with the formulation phage populations ranged from 104 to 107
PFU/mL. However, interestingly, formulated phage treatment did not decrease disease severity
(Table 4-1).
Effect of Phage Application on Disease Severity Incited by a Phage-Sensitive Xac Strain and Its Phage-Resistant Mutant
In order to reduce possible cross-treatment interference caused by phage spread between
plants, the effect of phage treatment on the development of citrus canker incited by a phage
sensitive strain, Xac65, and its phage resistant mutant, RF2 were compared in three experiments.
The disease caused by the two strains was not significantly different (Table 4-2), indicating that
the mutation to phage resistance did not affect pathogenicity of RF2. Phage application
significantly reduced the disease severity incited by Xac65 in all three experiments (Table 4-2).
Interestingly, the phage treatment also decreased disease severity incited by the RF2 in of one of

72
three tests (Table 4-2). It is likely that this resulted from an error during the execution of the
experiment: the plants may have been inoculated with Xac65 instead of RF2, or the phages may
have been contaminated with a phage that was able to lyse RF2.
Effect of Phage and Copper-Mancozeb Applications on Citrus Canker Disease Development in a Citrus Nursery
The effect of bacteriophage treatment, copper-mancozeb treatment, and their combination
was evaluated for reducing citrus canker disease development under field conditions in an
experimental citrus nursery in Argentina (Figure 4-1). Nursery plants were inoculated with a
copper tolerant Xac strain and were treated (i) twice weekly with a suspension of phage
ΦXaacA1; or (ii) twice weekly with a copper-mancozeb solution, or (iii) both with phage and
copper-mancozeb. Phage application alone significantly reduced citrus canker disease severity,
but it was not as effective as the copper-mancozeb treatment (Table 4-3). The combination of
phage and copper-mancozeb treatments did not result in increased control (Table 4-3); to the
contrary it reduced the efficacy of copper treatment in one of two trials.
Effect of Bacteriophage Treatment on Citrus Bacterial Spot Disease Development in a Commercial Citrus Nursery
Disease control trials were set up in the summer and fall of 2004 in a commercial citrus
nursery in Florida to evaluate the effect of preventative bacteriophage applications on CBS
disease development incited by naturally occurring Xacm populations. A mixture of three phages
was applied twice weekly in skim milk formulation at three sites. Two sites were established in
sections of Valencia orange, which is moderately sensitive to CBS, whereas the third site was in
a grapefruit section, which is highly susceptible to the disease. Phage treatment contributed to a
significant disease reduction in both Valencia sites (Table 4-4). Some disease suppression also
occurred at the grapefruit site, but it was not significant (p = 0.1585) (Table 4-4).

73
Effect of Bacteriophage and Copper-Mancozeb Treatments on Citrus Bacterial Spot Disease Development in an Experimental Nursery
The effects of bacteriophage treatment, copper-mancozeb treatment and their combination
were evaluated for reducing CBS disease development in an experimental citrus nursery. Phage
treatment contributed to significant disease reduction (Table 4-5); however, it provided less
control than the copper-mancozeb in one of two trials. The combination of phage and copper
treatments did not result in increased disease control, but did not result in reduced control, either.
Discussion
Phage treatments proved to be effective means of disease reduction both under greenhouse
and field conditions, both for citrus canker and citrus bacterial spot. Their efficacy would likely
be further increased if the phage mixtures were carefully designed and consisted of higher
number of phages with distinct cell receptor specificities. In these studies the number of phages
included in the mixture ranged from one to four and their receptor specificity was not evaluated
at all.
The greenhouse trials provided a good example of what could happen if phages with
similar receptor specificity are used. Strain RF2 became resistant to all four phages used in the
greenhouse trials with presumably a single mutation. It indicates that those four phages had the
same receptor specificity. Mutation to resistance by RF2 resulted in abolishment of disease
control by the phage treatment without loss in aggressiveness of the pathogen. This phenomenon
also underscores the need for constant stringent monitoring of pathogen populations in a phage-
based disease management program.
Skim milk formulation, despite its effect of increasing phage longevity on plant foliage,
totally abolished disease control achieved by the phages. It is possible that skim milk, which acts
as a wetting agent and breaks down surface tension, helped in the pathogen ingress. It could have

74
also served as a carbon source for the bacterium. There is clearly a need for further research on
developing protective formulations that give protection without the adverse effects.
The twice weekly spray schedule used in this study might be too frequent for a practical
application in a commercial grove setting. The relation of application frequency and control
efficacy is an economically important factor and should be investigated before considering
commercialization.
The combination of phage treatment and copper-mancozeb did not prove to be
advantageous. It was expected that the presence of ionic copper may reduce phage efficacy as it
generally inactivates proteins. However, phage application also reduced copper efficacy in these
trials. It is possible that phage virions and the proteins in the skim milk formulation tied up a
significant part of free copper ions thus reducing the copper concentration available for
eliminating the bacterial pathogen. The phage sprays also could have washed off a considerable
amont of copper from the plant foliage. If phages are to be used as a part of an integrated pest
management program, they likely will perform better together with other biological control
methods or with chemical methods of different modes of action such as systemic acquired
resistance inducers.

75
Figure 4-1. Experimental citrus nursery, the location of the 2006 citrus canker trials, at the
INTA research station in Bella Vista, Corrientes, Argentina.
Figure 4-2. Disease control plots in Dilley and Son Nursery, Avon Park, Florida.

76
Figure 4-3. Field plots at the Plant Science Research and Education Unit of the University of
Florida in Citra, Florida.

77
Table 4-1. Effect of bacteriophage treatment and use of skim milk formulation on citrus canker disease development incited by Xanthomonas axonopodis pv. citri strain Xac65 on grapefruit plants in the greenhouse
Percent disease severity Treatment Phagey Formulated phage Water pz
Test 1x 53b 75a 72a 0.0116 Test 2 6b 12ab 16a 0.1967 Test 3 13b 23a 24a 0.0211
x Inoculum concentration was 1×108 cfu/mL in test 1 and 5×106 cfu/mL in tests 2 and 3. y Means within the same row followed by the same letter are not significantly different according to the Waller-Duncan K-ratio t Test at p=0.05 level. z p = Probability that there are no differences in treatment means according to analysis of variance. Table 4-2. Effect of bacteriophage treatment on citrus canker disease development incited by
phage sensitive Xanthomonas axonopodis pv. citri strain Xac65 and its phage-resistant mutant, RF2 on grapefruit plants in the greenhouse
Percent disease severity Pathogen Xac65 RF2 Treatment Phagey Water Phage Water pz
Test 1 16b 55a 47a 51a 0.0454 Test 2 17b 39a 8b 48a 0.0052 Test 3 5b 42a 46a 36a 0.0262
y Means within the same row followed by the same letter are not significantly different according to the Waller-Duncan K-ratio t Test at p=0.05 level. z p = Probability that there are no differences in treatment means according to analysis of variance. Table 4-3. Effect of phage and copper-mancozeb application on citrus canker disease
development, incited by Xanthomonas axonopodis pv. citri strain Xc05-2592 in nursery trial in Bella Vista, Corrientes, Argentina
Disease intensityz Treatment UTCy,x Phage Cu-Mzv Phage+Cu-Mz
pw
Test 1 2.6a 2.0b 1.6c 1.8bc <0.0001 Test 2 3.1a 2.1b 1.6c 2.0b <0.0001
v Cu-Mz = copper-mancozeb. w p = Probability that there are no differences in treatment means according to analysis of variance. x UTC = untreated control y Means within the same row followed by the same letter are not significantly different based on least square means differences at p=0.05 level. z Disease intensity = average number of lesions on diseased leaves.

78
Table 4-4. Effect of bacteriophage treatment on citrus bacterial spot disease development incited
by naturally occurring Xanthomonas axonopodis pv. citrumelo strains at Dilley and Son Nursery in Avon Park, Florida in 2004, as measured by area under the disease progress curve (AUDPC)
AUDPC Phage Control pz
Site 1y 173 331 0.0023 Site 2 169 261 0.0357 Site 3 128 154 0.1585
y Sites 1 and 2 consisted of Valencia orange, site 3 of grapefruit. z Probability of equality of sample means according to t test.
Table 4-5. Effect of bacteriophage and copper-mancozeb treatment on citrus bacterial spot disease development incited by Xanthomonas axonopodis pv. citrumelo strain S4m at the UF Plant Science Unit in Citra, Florida in 2006, as measured by area under the disease progress curve (AUDPC)
AUDPC Treatment UTCy,z Phage Cu-Mzw Phage+Cu-Mz
px
Test 1 0.53a 0.20b 0.08b 0.13b 0.0115 Test 2 24.3a 14.9b 4.0c 2.0c <0.0001
w Cu-Mz = copper-mancozeb. x p = Probability that there are no differences in treatment means according to analysis of variance. y UTC = untreated control. z Means within the same row followed by the same letter are not significantly different based on least square means differences at p=0.05 level.

79
CHAPTER 5 INTERACTION OF BACTERIOPAHGES AND THE HOST BACTERIA ON THE
PHYLLOPLANE
Introduction
Bacteriophages, when used as biological control agents for foliar plant diseases interact
with the target organism on the leaf surface, the phylloplane. The phylloplane is a constantly
changing environment; there are changes in temperature, sunlight irradiation, leaf wetness,
relative humidity, osmotic pressure, pH, microbial flora, and in the case of agricultural plants,
also chemical compounds (40). These factors are harmful to bacteriophages to varying extents.
Sunlight irradiation, especially in the UV A and B range, is highly detrimental to
microorganisms in general (4,65) and is mainly responsible for eliminating bacteriophages
within hours of application (66). Different temperatures and levels of relative humidity have
different effects on phage longevity. Water is necessary for disseminating phage virions and it
provides the medium for phage-bacterium interactions. It also contributes to virion stability,
since many phages are sensitive to desiccation. Microbial activity and enzymes degrade phages.
Modern intensive agriculture relies heavily on application of pesticides for control of plant
diseases and insect pests. Most chemicals tested did not affect bacteriophage persistence (9,
137); however, copper compounds are clearly detrimental (9,66).
Materials that increase the longevity of viruses in the field have been identified
(7,8,15,16,63,64,65). The use of such protective materials also increased disease control
achieved with bacteriophages (10). Timing phage treatments to apply in the evenings and early
mornings when sunlight UV irradiation is minimal, also increased efficacy (10). One important
factor that remains unexplored is the phages themselves. Bacteriophages used in previous studies
and also for commercial phage products available for agricultural use were selected strictly based
on in vitro tests (10, 89, 90, 99) without any evaluation in the target environment. It is not known

80
whether the different phages included in the mixture persist similarly on the plant surface,
whether the ratio of active ingredients in the phage treatments changes in the target environment
after application. It is not known if different phages in the phage mixture contribute equally to
disease control. It is not known if there are such characteristics that make a phage more suitable
to disease control on a leaf surface than others. The goal was to try to shed some light in this
darkness. The two objectives were (i) to monitor the fate of bacteriophages, individual
constituents of a phage mixture used for disease control in a real field situation after being
applied to the leaf surface, and (ii) to test if the ability to multiply on the leaf surface influences
phage disease control efficacy and makes a detectable difference on disease control.
Material and Methods
Standard Bacteriophage Techniques
Determination of titer. Phage concentrations were determined by the dilution-plating-
plaque-count assay on nutrient agar yeast extract plates without bottom agar, as previously
described (97).
Phage propagation. Phages were recovered from storage, purified by single plaque
isolations and then mass streaked on the freshly prepared lawn of the propagating host. After
overnight incubation the phages were eluted from the plate, sterilized and enumerated, as
described above. The eluate was used for infecting 500 mL of actively growing culture of the
propagating strain (108 cfu/mL) grown in NB liquid medium in 1 liter flasks at 0.1 multiplicity of
infection (MOI). After addition of the phage and 5-min incubation on the bench top, the culture
was shaken at 150 rpm at 28ºC for 18 h. Then the culture was sterilized, enumerated and stored
at 4ºC in the dark until use. This method yielded phage titers of approximately 1010 PFU/mL.
Phage concentration by high speed centrifugation. High titer phage lysates (~1010
PFU/mL) were concentrated and purified according to methods described by Hans-W.

81
Ackermann (personal communication). One hundred milliliters of the sterilized lysate was
centrifuged at 10,000 g for 10 min to sediment the bacterial debris. Forty milliliters of the
supernatant was transferred to a new centrifuge tube and centrifuged at 25,000 g for 60 min to
sediment the phage particles. The supernatant was discarded and replaced with 0.1 M ammonium
acetate solution (pH 7.0). Following an additional centrifugation step (60 min, 25,000 g) the
supernatant was discarded and the pellet was resuspended in 1.5 mL SM buffer. The final phage
concentration was approximately 1012 PFU/mL.
Changes in Xanthomonas axonopodis pv. citrumelo Phage Populations on the Field
The experiment was conducted at the Plant Science Unit of the University of Florida in
Citra, Florida. A mixture of three bacteriophages was sprayed onto Swingle citrumelo plants at 8
PM on July 18, 2006. The phages used were ccΦ13-2, α-MME and Φ5536, all adjusted to 1×108
PFU/mL as determined on their common propagating bacterium, Xanthomonas axonopodis pv.
citrumelo (Xacm) strain, S4m. The mixture was applied in skim milk formulation (0.75% (wt/V)
non-fat dry milk powder) using a non-CO2, Solo backpack sprayer. The phage populations were
monitored in the evening and the next morning as follows. Leaf samples were taken at 8:15 PM,
7:45 AM, 9:00 AM, 10:00 AM and 11:00 AM. For each sample three trifoliate leaves,
originating from a singe plant, were removed and placed into a plastic freezer bag (Hefty OneZip
quart size, Pactiv Co., Lake Forest, IL). At each timepoint four samples were collected from
plants within the same plot. The samples were placed on ice in a portable plastic cooler and
immediately carried to the laboratory. The bags were then weighed and 50 mL of sterilized tap
water was poured into each bag. The bags were shaken on a wrist action shaker (Burrel Co.,
Oakland, CA) for 20 min and then 1 mL of leaf-wash was removed from each bag and
transferred into 1.5 mL microcentrifuge tubes containing 20 µL of chloroform. The tubes were
stored at 4ºC until all samples were taken and processed and then transported to the Gainesville

82
laboratory for determination of phage titer. Phages were enumerated by dilution plating and
plaque counts on three bacterial strains: MME (X. vesicatoria), Xacm45 (Xanthomonas
axonopodis pv. citri) and S4m (Xacm). MME selectively detected phage α-MME, Xacm45
selectively detected phage Φ5536, and S4m detected all three phages. Titers of phages α-MME
and Φ5536 were calculated from the plaque number, dilution information and leaf weight and
were expressed as PFU/g leaf tissue. Titer of phage ccΦ13-2 was determined indirectly, as
described. The efficacy of plating of phage α-MME on strain MME versus strain S4m was
determined concurrently to phage titer determinations by plating known concentration of phage
α-MME on both strains (5 replication each) and dividing the average number of plaques
produced on MME by the average number of plaques produced on S4m. EOP of phage Φ5536
on Xacm45 versus S4m was determined similarly. The EOP values were multiplied with the
phage concentrations determined on the appropriate selective host to calculate the phage
concentration on S4m for each sample. The concentration of ccΦ13-2 in each sample was
determined by the following equation: ccΦ13-2 concentration = total phage concentration (as
determined on S4m) – α-MME concentration (calculated) - Φ5536 concentrated (calculated).
Finally, the data were converted by the y=log10(x+1) function and were graphed. The sunlight
irradiation data were downloaded from the Florida Automated Weather Network website
(http://fawn.ifas.ufl.edu).
Interaction of Xanthomonas perforans and Its Bacteriophage on the Tomato Foliage in the Greenhouse
Population study. Six young tomato plants, in 3-4 leaf stage, were dipped in a suspension
of X. perforans strain, ΔopgH:ME-B (105 cfu/mL), amended with 0.025% Silwet L-77 (Loveland
Industries, Co., Greely, CO). Five days later three of the plants were sprayed with sterilized tap
water and three with a suspension of bacteriophage ΦXv3-3-13h (108 PFU/mL). The plants were

83
placed in clear polyethylene bags for 1 day after phage application in a growth chamber (28ºC).
Before removing the bags, the plants were transferred to a greenhouse where they remained
throughout the experiment in a completely randomized arrangement. Phage populations were
monitored daily for a week by removing three leaflets and recovering the phages from the
leaflets.
Disease control study. Phage ΦXv3-3-13h was also used for controlling tomato bacterial
spot severity incited by ΔopgH:ME-B. Six young tomato plants were inoculated with strain
ΔopgH:ME-B; and later three of them were sprayed with ΦXv3-3-13h and three were sprayed
with sterilized tap water. Methods of bacterial inoculation and phage application were the same
as for the population studies. The ratio of diseased leaf tissue was estimated on the bottom three
fully expanded leaves of each plant twice, 19 and 22 days after inoculation using the Horsfall-
Barratt (HB) scale (12): 1=0%, 2=0-3%, 3=3-6%, 4=6-12%, 5=12-25%, 6=25-50%, 7=50-75%,
8=75-87%, 9=87-94%, 10=94-97%, 11=97-100%, 12=100%. The HB values were converted to
estimated mean percentages by using the Elanco Conversion Tables for Barratt-Horsfall Rating
Numbers (ELANCO Products Co., Indianapolis, IN), as described in Appendix A. The area
under the disease progress curve (AUDPC) was computed by trapezoidal integration of disease
percentage values taken in different timepoints using the formula proposed by Shaner and
Finnley (104), as described in Appendix A. The sample means of the phage-treated and non-
treated plants were compared by t-test using SAS System for Windows program release 8.02
(Cary, NC).
Interaction of Xanthomonas axonopodis pv. citri and Its Bacteriophages on Grapefruit Foliage in the Greenhouse
Disease control studies. Two experiments were carried out at the greenhouse of the
Department of Agriculture & Consumer Services, Division of Plant Industry, citrus canker

84
quarantine facility. Duncan grapefruit plants were heavily pruned and fertilized to induce the
simultaneous boom of a new flush that is susceptible to citrus canker infection. Three weeks later
the emerging uniform new foliage was treated with on of three different single-phage
suspensions (ΦXV3-21, ΦXaacF1 or ccΦ19-1, 5x109 PFU/mL) or with sterilized tap water. Fifty
milliliters of the phage suspension was spayed on each plant in the evening, and then the plants
were placed in white plastic bags. On the next morning the bags were removed from the plants,
and the plants were inoculated with Xac strain Xac65 (test 1×106 cfu/mL). After inoculation the
plants were placed inside the bags for an additional 24 h of high moisture. After removal from
the bags and after the foliage was allowed to dry the plants were arranged in a completely
randomized pattern on a greenhouse bench. The disease was assessed 3-4 weeks after
inoculation. Four plants were used per treatment. In the first experiment the phage suspensions
were prepared by diluting high titer lysates, but because of concerns that the presence of nutrient
broth in the phage lysate may contribute to increased disease severity, in the second experiment
the phage lysates were concentrated in order to remove nutrient broth. The ratio of diseased leaf
surface area was estimated using the Horsfall-Barratt (HB) scale (12). The HB values were
converted to estimated mean percentages by using the Elanco Conversion Tables for Barratt-
Horsfall Rating Numbers (ELANCO Products Co., Indianapolis, IN), as described in Appendix
A. Analysis of variance (ANOVA) and subsequent separation of sample means by the Waller-
Duncan K-ratio t Test was carried out using SAS System for Windows program release 8.02
(Cary, NC).
Population studies. In order to determine if bacteriophages ΦXV3-21, ΦXaacF1 and
ccΦ19-1 are able to multiply on the grapefruit foliage in the presence of their host, Xac65,
grapefruit plants were sprayed with a mixture of the three phages at low concentration (5×106

85
PFU/mL) and immediately followed by application of the bacterial suspension at a much higher
concentration (1×108 cfu/mL) or with sterilized tap water for control. Phage populations were
monitored by removing three leaflets 0, 3, 6 and 9 h after application, recovering the phages
from the leaflets and determining concentrations. In order to determine populations of the
individual phages, the leaf washes were plated on three Xac strains that specifically detected
each of the tree phages. These strains were only sensitive to one of three phages. Strain Xac41
was used for specific detection of ΦXaacF1; Xac15 for ccΦ19-1 and Xac30 for ΦXv3-21.
Results
Persistence of Bacteriophages on Citrus Leaf Surface Under Field Conditions
The fate of three phages, ccΦ13-2, α-MME and Φ5536, was monitored on citrus foliage.
The mixture of these three phages was used for biological control of citrus bacterial spot, and
was applied at 8 PM. The three phages were applied at equal concentrations, at 1×108 PFU/mL,
in skim milk formulation. The sunlight irradiation at the sites was 0 until 6:30 AM then gradually
increased to approximately 400 W/m2 at 9:45 AM (Figure 5-1). Then the irradiation suddenly
jumped to almost 600 W/m2. The three phages persisted differently on the foliage (Figure 5-1).
Φ5536 populations reduced slightly from 8 PM until 10 AM, 132 and 135 fold in sites 1 and 2,
respectively (Figure 5-1). However, concurrently with the suddenly higher sunlight irradiation,
Φ3356 populations sharply dropped between 10 AM and 11AM, decreasing 2159 and 801 folds,
in sites 1 and 2, respectively, and were practically eliminated by 11 AM. The populations of
phage ccΦ13-2 decreased 32 and 12 fold between 8 PM and 10 AM, in sites 1 and 2, respectively
(Figure 5-1). The rate of population decline increased between 10 AM and 11 AM, but it was
much lower than that of Φ3356, with 66 and 30 fold in sites 1 and 2, respectively (Figure 5-1).
At 11 AM still there were, 632 and 142 PFU per leaf ccΦ13-2 at site 1 and site 2, respectively.
ΦMME populations were lower from the beginning compared to the other two phages (Figure

86
5-1). The nighttime reduction was low, but the populations decreased earlier in the morning than
other two phages. In site 1 there was no population reduction between 8 PM and 7:45 AM, but
between 7:45 and 9 AM there was a 2490 fold decrease and from 9 AM or later the populations
were hardly detectable, 19 PFU/leaf (Figure 5-1). In site 2, ΦMME populations reduced slightly,
6 folds, between 8PM and 7:45AM, and more than 500 fold between 7:45 and 10 AM to 121
PFU/leaf (Figure 5-1).
Ability of Three Phages of Xanthomonas axonopodis pv. citri to Multiply on Grapefruit Foliage in the Presence of Their Bacterial Host, and Their Effect on Citrus Canker Disease Development
In order to elucidate if a phage need to be able to multiply on the leaf surface for effective
disease control, three phages active against Xac were evaluated for their ability to (i) multiply on
grapefruit leaf surface in the presence of high populations of the host bacterium, and to (ii)
reduce citrus canker disease severity under greenhouse conditions. The populations of phage
ΦXV3-21 decreased in the absence of Xac65 to 1.6 and 1.9% of the original populations after
nine hours on the grapefruit leaves in experiments 1 and 2, respectively (Figures 5-2A and 5-
3A). In the presence of Xac65 ΦXV3-21 populations persisted better than in the absence of the
host, although they still decreased over time and after 9 hours were 4.2 and 26.2% of the starting
populations. Populations of phage ΦXaacF1 also decreased in the absence of the Xac65 and were
4.2 and 5.8% at the end of the experiment in experiment 1 and 2, respectively (Figures 5-2B and
5-3B). However, in the presence of the host, ΦXaacF1 increased in population, to 324 and 557%,
after nine hours in experiments 1 and 2, respectively (Figures 5-2B and 5-3B). The third phage,
ccΦ19-1 decreased in populations in the absence of Xac65 to 0.4 and 1.5% by the end of the
experiment (Figures 5-2B and 5-3B). In experiment 1, 6 hours after application ccΦ19-1
populations were higher in the presence of the host than in its absence (35% vs. 9.2%); however

87
at nine hours no ccΦ19-1 was detected in the presence of the host (Figure 5-2C). In the second
experiment the presence of Xac65 ccΦ19-1 populations were at 45% after 9 h (Figure 5-2C).
These three phages were used to treat grapefruit plants before inoculating with the
pathogen, Xac65, in two experiments. In the first experiment none of the phages reduced citrus
canker disease severity (Table 5-1). On the contrary, ΦXV3-21 application resulted in significant
disease increase compared to the water control (Table 5-1). However there were significant
differences between plants treated with different phages: ccΦ19-1-treated plants had significantly
less disease than ΦXV3-21-treated plants and significantly more disease than ΦXaacF1-treated
ones (Table 5-1). It was suspected that the nutrient broth present in phage suspensions
contributed to the disease increase on the ΦXV3-21 treated plants and it could have also caused
the loss of control provided by the phages. Thus, for the second experiment nutrient broth was
removed from phage suspensions before applications. In this experiment ΦXaacF1 treatment
significantly reduced disease severity, while the other two phages did not have any significant
effect.
Ability of a Xanthomonas perforans Phage to Multiply on Tomato Foliage in the Presence of Its Bacterial Host, and Its Effect on Tomato Bacterial Spot Disease Development
To investigate if the correlation between a phage’s ability to multiply on the plant surface
and effectively reduce disease is a general and present in other pathosystems, the interactions of
a X. perforans strain, the causal agent of tomato bacterial spot (Figure 5-4), and its bacteriophage
were studied. The ability of phage, ΦXv3-3-13h, to multiply on tomato foliage in the presence of
its host, ΔopgH:ME-B, was evaluated in greenhouse studies. ΦXv3-3-13h populations
continuously decreased in the absence of the bacterial host and were under the detection limit 5
or 6 days after application, in experiments 1 and 2, respectively (Figure 5-5). In the presence of
ΔopgH:ME-B ΦXv3-3-13h populations increased 11 and 87 fold in the first 24 h, in experiments

88
1 and 2, respectively, and then declined at slower rate than without the host (Figure 5-5). After 7
days, the populations were only 8.5 and 17 times lower than at the time of application.
Phage ΦXv3-3-13h was evaluated for its ability to reduce bacterial spot of tomato disease
development incited by ΔopgH:ME-B. Disease severity was significantly reduced on phage-
treated plants compared to water-treated plants (Table 5-2).
Discussion
Phages constituting a phage mixture were shown to persist differently in the phyllosphere.
The phages in the mixture had different levels of persistence and in the morning soon after
sunrise the phage diversity was reduced from three phages to two and by late morning to one.
Experiments with formulations showed that increasing phage persistence contributes to increased
disease control ability. Thus it seems likely that using phages that can persist longer on the
foliage either by to resisting desiccation and sunlight irradiation better, or being more effective in
multiplying in the presence of the pathogen would be better suited for disease control.
The phage that was effective in multiplying on the leaf surface did control the disease.
Conversely, the phages that were not able to multiply efficiently enough to even maintain their
populations could not reduce the disease. We only showed this correlation with a limited number
of phages and only in two pathosystems, and it would be interesting to continue such evaluations
on a broader scale. If it is true that those phages are more effective, which persist longer and can
multiply efficiently in the phyllosphere than the phages naturally inhabiting the phyllosphere
should be the most effective ones. Thus the phyllosphere may be the best place for finding
phages for disease control purposes. It is discouraging, however, that in this study we found no
variability amongst the phyllosphere phages in host ranges. However, our results also showed
that the phages sensitivity profiles of Xanthomonas species overlap, thus phage diversity may be
increased by isolating phages from different Xanthomonas pathosystems.

89
Overall Summary and Conclusions
Citrus canker, incited by Xanthomonas axonopodis pv. citri and X. axonopodis pv.
aurantifolii, is one of the most damaging citrus diseases in the world. Citrus canker was
reintroduced to Florida in the 1990s and threatens the state’s $9 billion citrus industry. This work
focused on a biological control approach to use bacteriophages for reducing bacterial pathogen
populations and disease severity on citrus. Bacteriophages isolated from citrus canker lesions in
Florida and Argentina were evaluated based on plaque morphology, chloroform sensitivity, host
range, genome size, DNA restriction profile and virion morphology. There was low diversity
among the isolated phages, as 61 of 67 were nearly identical to each other and phage CP2 of
Japan. Mixtures of bacteriophages were evaluated for controlling citrus canker in greenhouse
trials in Florida and in nursery trials in Argentina. Bacteriophages reduced citrus canker disease
severity both in greenhouse and field trials. The level of control was inferior to chemical control
with copper bactericides. The combination of bacteriophage and copper treatments did not result
in increased control. Citrus canker field trials in Florida have been prohibited until recently, as
the disease was under eradication. For this reason we evaluated the efficacy of phage treatment
on a similar bacterial citrus disease, citrus bacterial spot, incited by X. axonopodis pv. citrumelo.
Bacteriophages reduced citrus bacterial spot severity. The level of control was equal or inferior
to chemical control with copper bactericides. The combination of bacteriophage and copper
treatments did not result in increased control. In experiments monitoring the fate of
bacteriophages on the citrus foliage following bacteriophage application, phage populations
stayed steady on the foliage during nighttime but were drastically reduced within hours after
sunrise. The rate of reduction varied among the phages. The ability of bacteriophages to multiply
on the plant foliage in the presence of their bacterial host was investigated. Phages varied in their
ability to multiply, and the ones that successfully increased in populations on the bacterial host

90
on the leaf surface also reduced disease severity, whereas the ones that were unable to multiply
in the target environment did not reduce disease severity.
However there is a need for research in several areas before commercial phage application
will become a feasible option. First of all, the diversity of the citrus canker phage library has to
be increased, either by further isolations or by changing host ranges and receptor specificities of
available phages. The protective formulation also need improvement: the 0.75% skim milk
powder formulation increases phage longevity on the foliage, but it also provides carbon source
to the microorganisms on the leaf surface, including the pathogen. Additionally, the protein
content of skim milk may tie up a considerable amount of ionic copper thus reducing the efficacy
of copper bactericides. Identification of formulations that provide protection without the adverse
effects could potentially result in increased disease control efficacy. The optimal frequency of
application and phage concentration also remains to be determined. Both of these factors greatly
influence the cost of phage treatment, what in turn determines feasibility. In these field tests we
applied phages twice weekly at ~108 PFU/ml, but it is not known if less frequent applications
and lower phage concentrations would have achieved similar control. Combining phage
treatment with other biological control methods and chemicals with different modes of action
than copper also needs to be evaluated. It may be possible to increase disease control efficacy of
individual bacteriophages by selection for increased persistence on the leaf surfaces and higher
in vivo multiplication ability. Also, rapid screening methods could be developed to predict
disease control ability of phages.

91
0
1
2
3
4
5
6
7
8:15 PM 7:45 AM 9:00 AM 10:00 AM 11:00 AM
Log 1
0 PFU
/g le
af ti
ssue
Φ5536ΦMMEccΦ13-2
A
0
1
2
3
4
5
6
7
8:15 PM 7:45 AM 9:00 AM 10:00 AM 11:00 AM
Log 1
0 PFU
/g le
af ti
ssue
Φ5536ΦMMEccΦ13-2
B
0
100
200
300
400
500
600
700
8:00 P
M
9:00 P
M
10:00
PM
11:00
PM
12:00
AM
1:00 A
M
2:00 A
M
3:00 A
M
4:00 A
M
5:00 A
M
6:00 A
M
7:00 A
M
8:00 A
M
9:00 A
M
10:00
AM
11:00
AM
Sun
light
(W/m
2 )
C
Figure 5-1. Bacteriophage populations and sunlight irradiation in Citra, FL on July 18 and 19,
2006. A) Bacteriophage populations at site 1. B) Bacteriophage populations at site 2. C) Sunlight irradiation. Phage was applied at 8:00PM. Error bars indicate the standard error.

92
0
20
40
60
80
100
120
140
160
0 3 6 9
Hours after inoculation
Per
cent
pop
ulat
ion
ΦXV3-21ΦXV3-21 with host
A
0
100
200
300
400
500
600
0 3 6 9
Hours after inoculation
Per
cent
pop
ulat
ion
ΦXaacF1ΦXaacF1 with host
B
0
20
40
60
80
100
120
140
160
0 3 6 9
Hours after inoculation
Per
cent
pop
ulat
ion
ccΦ19-1ccΦ19-1 with host
C
Figure 5-2. Changes in bacteriophage populations on grapefruit foliage in the presence or
absence of the host, Xanthomonas axonopodis pv. citri strain Xac65. Experiment 1. A) Phage ΦXV3-21. B) Phage ΦXaacF1. C) Phage ccΦ19-1. Percent population = percent of population average at 0 hour. Error bars indicate the standard error.

93
0
20
40
60
80
100
120
140
0 3 6 9
Hours after inoculation
Per
cent
pop
ulat
ion
ΦXV3-21ΦXV3-21 with host
A
0
100
200
300
400
500
600
700
800
900
0 3 6 9
Hours after inoculation
Perc
ent p
opul
atio
n
ΦXaacF1ΦXaacF1 with host
B
020406080
100120140160180200
0 3 6 9
Hours after inoculation
Per
cent
pop
ulat
ion
ccΦ19-1ccΦ19-1 with host
C
Figure 5-3. Changes in bacteriophage populations on grapefruit foliage in the presence or
absence the host, Xanthomonas axonopodis pv. citri strain Xac65. Experiment 2. A) Phage ΦXV3-21. B) Phage ΦXaacF1. C) Phage ccΦ19-1. Percent population = percent of population average at 0 hour. Error bars indicate the standard error.

94
Table 5-1. Effect of treatment with three phages on citrus canker disease development, incited by Xanthomonas axonopodis pv. citri strain Xac65, as measured by disease severity.
Average disease severity Treatment Controly ccΦ19-1 ΦXaacF1 ΦXV3-21 pz
Test 1 22bc 33b 13c 51a 0.0018 Test 2 28ab 24bc 18c 32a 0.0049
y Means within the same row followed by the same letter are not significantly different based on least square means differences at p=0.05 level. z p = Probability that there are no differences in treatment means according to analysis of variance.

95
Figure 5-4. Symptoms of tomato bacterial spot, incited by Xanthomonas perforans.

96
0123456789
10
1 2 3 4 5 6 7 8
Days after application
Log 1
0 PFU
/leaf
let
HostNo Host
A
0123456789
10
1 2 3 4 5 6 7 8
Days after application
Log 1
0 P
FU/le
afle
t
HostNo Host
B
Figure 5-4. Populations of bacteriophage ΦXv3-3-13h on tomato foliage in the presence or
absence of its host Xanthomonas perforans strain ΔopgH:ME-B. A) Experiment 1. B) Experiment 2. Error bars indicate the standard error.

97
Table 5-2. Effect of a curative application of bacteriophage ΦXv3-3-13h on tomato bacterial
spot disease development, incited by Xanthomonas perforans strain ΔopgH:ME-B, as measured by area under the disease progress curve (AUDPC).
Treated Control pz
AUDPC 12.5 27.3 0.0026 z Probability of equality of sample means according to t test.

98
APPENDIX A CALCULATIONS
Conversion of Horfall-Barratt Values to Mean Percentages
The Horsfall-Barratt values (12) were converted to estimated mean percentages by using
the Elanco Conversion Tables for Barratt-Horsfall Rating Numbers (ELANCO Products Co.,
Indianapolis, IN). The estimated mean percentage of certain experimental unit, one plant in the
greenhouse experiments or one plot in the field trials, was determined by taking the arithmetic
mean of the percentages of several ratings taken at one time, where each individual percentage
was taken as the midpoint of the appropriate percentage interval (Table A-1). For example, if the
following four HB ratings were assigned to a grapefruit plant, 10, 7, 5 and 10, then the estimated
mean percentage was calculated as (95.31 + 62.5 + 18.75 + 95.31) / 4 = 78.5.
Calculation of Area Under the Disease Progress Curve
The area under the disease progress curve (AUDPC) (Figure A-1) was computed by the
trapezoidal integration of disease percentage values taken in different timepoints using the
formula proposed by Shaner and Finnley (104). Disease progress curve is graphed by plotting the
diseases percentage values (y-axis) against the time of disease assessment (x-axis) (Figure A-1).
The area under this curve consists of several trapezoids. The area of each trapezoid is determined
as (y2+y1)/2*(x2-x1), where (x1,y1) are the coordinate of the earlier and (x2,y2) are the coordinates
of the later observation. For example if four disease ratings were taken one week apart and the
disease percentages are 5, 10, 20 and 25, then there will be three trapezoids. The area of the first
trapezoid is (10+5)/2*7=52.5, the area of the second is 105, the area of the third one is 157.5, so
the total area of the disease progress curve is 315.

99
Table A-1. Horsfall-Barratt scale, the corresponding disease intervals and midpoint values.
HB value Disease Percentage Midpointz
1 0 0 2 0-3 2.34 3 3-6 4.68 4 6-12 9.37 5 12-25 18.75 6 25-50 37.5 7 50-75 62.5 8 75-88 81.25 9 87-94 90.63 10 94-97 95.31 11 97-100 97.66 12 100 100
z Midpoint values are as published in the Elanco Conversion Tables for Barratt-Horsfall Rating Numbers (ELANCO Products Co., Indianapolis, IN).

100
A
0
5
10
15
20
25
30
0 7 14 21
Days after first rating
Per
cent
dis
ease
Disease Progress Curve
B
0
5
10
15
20
25
30
0 7 14 21
Days after first rating
Per
cent
dis
ease
Area Under the Disease Progress Curve
Figure A-1. Visualization of the disese progress curve and of the area under the disease progress
curve. A) Disease progress curve is prepared by plotting the diseases percentage values (y-axis) against the time of disease assessment (x-axis). B) Area under the disease progress curve (AUDPC) is one value describing the overall disease progress throughout the season.

101
LIST OF REFERENCES
1. Ackermann H. 2005. Bacteriophage classification. Pages 67-89 in: Bacteriophages: biology and applications. in: Kutter E, and Sulakvelidze A, eds. CRC Press, Boca Raton, Florida.
2. Adams MH. 1959. Bacteriophages. Interscience Publishers, New York, New York.
3. Agrios GN. 2004. Plant pathology. 5th ed. Elseiver Academic Press, San Diego, California.
4. Ahamd MH, and Morgan V. 1994. Characterization of a cowpea (Vigna undulata) rhizobiophage and its effects on cowpea nodulation and growth. Biol Fertil Soils 18:297-301.
5. Al-saadi A. 2005. Phenotypic characterization and sequence analysis of pthA homologs from five pathogenic variant groups of Xanthomonas citri [dissertation]. Gainesville, Florida: University of Florida.
6. Alvarez AM, Benedict AA, Mizumoto CY, Pollard LW, and Civerolo EL. 1991. Analysis of Xanthomonas campestris pv. citri and X. c. citrumelo with monoclonal antibodies. Phytopathology 81:857-865.
7. Arthurs SP, Lacey LA, and Behle RW. 2006. Evaluation of spray-dried lignin-based formulations and adjuvants as solar protectants for the granulovirus of the codling moth, Cydia pomonella (L). Journal of invertebrate pathology 93:88-95. Epub: 2006 Jun 13.
8. Balogh B. 2002. Strategies of improving the efficacy of bacteriophages for controlling bacterial spot of tomato [thesis]. Gainesville, Florida: University of Florida.
9. Balogh B, Jones JB, Momol MT, and Olson SM. 2005. Momol MT, Ji P, Jones JB, editors. Persistence of bacteriophages as biocontrol agents in the tomato canopy. Proceedings of the 1st IS on tomato diseases; June 21-24, 2004; Orlando, Florida. ISHS:299-302.
10. Balogh B, Jones JB, Momol MT, Olson SM, Obradovic A, King P, and Jackson LE. 2003. Improved efficacy of newly formulated bacteriophages for management of bacterial spot on tomato. Plant Dis 87:949-954.
11. Balogh B, Jones JB, Stall RE, Canteros BI, and Gochez AM. 2004. Association of bacteriophages with citrus canker and their use for control. Phytopathology 94 6.
12. Barratt RW, and Horsfall JG. 1945. An improved grading system for measuring plant disease. Phytopathology 35:655.
13. Barrow PA. 2001. The use of bacteriophages for treatment and prevention of bacterial disease in animals and animal models of human infection. J Chem Technol Biotechnol 76:677-682.
14. Basit HA, Angle JS, Salem S, and Gewaily EM. 1992. Phage coating of soybean seeds reduces nodulation by indigenous soil bradyrhizobia. Can J Microbiol 38:1264-1269.

102
15. Behle RW, McGuire MR, Gillespie RL, and Shasha BS. 1997. Effects of alkaline gluten on the insecticidal activity of Bacillus thuringiensis. J Econ Entomol 90:354-360.
16. Behle RW, McGuire MR, and Shasha BS. 1996. Extending the residual toxicity of Bacillus thuringiensis with casein-based formulations. J Econ Entomol 89 6:1399-1405.
17. Belasque J,Jr, Parra-Pedrazzoli AL, Neto JR, Yamamoto PT, Chagas MCM, Parra JRP, Vinyard BT, and Hartung JS. 2005. Adult citrus leafminers (Phyllocnistis citrella) are not efficient vectors for Xanthomonas axonopodis pv. citri. Plant Dis 89:590-594.
18. Bergamin Filho A, and Kimati H. 1981. Estudos sobre um bacteriofago isolado de Xanthomonas campestris. II. Seu emprego no controle de X. campestris e X. vesicatoria. Sum Phytopath 7:35-43.
19. Bock CH, Parker PE, and Gottwald TR. 2005. Effect of simulated wind-driven rain on duration and distance of dispersal of Xanthomonas axonopodis pv. citri from canker-infected citrus trees. Plant Dis 89:71-80.
20. Brunings AM, and Gabriel DW. 2003. Xanthomonas citri: breaking the surface. Molecular plant pathology 4:141-157.
21. Civerolo EL. 1984. Bacterial canker disease of citrus. J Rio Grande Valley Hortic Assoc 37:127-146.
22. Civerolo EL. 1972. Maas Geesteranus HP, editor. Interaction between bacteria and bacteriophages on plant surfaces and in plant tissues. Proceedings of the third international conference of plant pathogenic bacteria; April 14-21 1971; Wageningen. Wageningen: Centre for Argricultural Publishing and Documentation 25-37.
23. Civerolo EL, and Keil HL. 1969. Inhibition of bacterial spot of peach foliage by Xanthomonas pruni bacteriophage. Phytopathology 12:1966-1967.
24. Compton L. USDA determines citrus canker eradication not feasible Online: Florida Department of Agriculture and Consumer Services - Division of Plant Industry. 2006 January 11 [cited 2006 October 31]. Available from: http://www.doacs.state.fl.us/press/2006/01112006_2.html.
25. Coons GH, and Kotila JE. 1925. The transmissible lytic principle (bacteriophage) in relation to plant pathogens. Phytopathology 15:357-370.
26. Cubero J, and Graham JH. 2002. Genetic relationship among worldwide strains of Xanthomonas causing canker in citrus species and design of new primers for their identification by PCR. Appl Environ Microbiol 68:1257-1264.
27. Dai H, Chiang KS, and Kuo TT. 1980. Characterization of a new filamentous phage Cf from Xanthomonas citri. J Gen Virol 46:277-289.
28. d'Herelle F. 1921. Le bactériophage: Sone rôle dans limmunité. Masson et Cie, Paris.

103
29. Dopson RN. 1964. The eradication of citrus canker. Pl Dis Rept 48:30-31.
30. Duan YP, Castaneda A, Zhao G, Erdos G, and Gabriel DW. 1999. Expression of a single, host-specific, bacterial pathogenicity gene in plant cells elicits division, enlargement, and cell death. Mol Plant-Microbe Interact 12:556-560.
31. Duckworth DH, and Gulig PA. 2002. Bacteriophages: potential treatment for bacterial infections. BioDrugs 16:57-62.
32. Egel DS, Graham JH, and Stall RE. 1991. Genomic relatedness of Xanthomonas campestris strains causing diseases of citrus. Appl Environ Microbiol 57:2724-2730.
33. Erskine JM. 1973. Characteristics of Erwinia amylovora bacteriophage and its possible role in the epidemiology of fire blight. Can J Microbiol:7.
34. FDACS DPI. Comprehensive Report on Citrus Canker Eradication Program in Florida Through 14 January 2006 Revised Online: Florida Department of Agriculture and Consumer Services Division of Plant Industry. 2006 August 29. Available from: http://www.doacs.state.fl.us/pi/canker/pdf/cankerflorida.pdf.
35. FDACS DPI.. Florida Huanglongbing Science Panel Report Online: Florida Department of Agriculture and Consumer Services, Division of Plant Industry. 2006 January 31. Available from: http://www.doacs.state.fl.us/pi/chrp/greening/hlbsciencereport1-31-06.pdf.
36. FDACS DPI. Huanglongbing statewide survey map Online: Florida Department of Agriculture and Consumer Services, Division of Plant Industry. 2006 October 25. Available from: http://www.doacs.state.fl.us/pi/chrp/greening/maps/cgsit_map.pdf.
37. Flaherty JE, Harbaugh BK, Jones JB, Somodi GC, and Jackson LE. 2001. H-mutant bacteriophages as a potential biocontrol of bacterial blight of geranium. HortScience 36:98-100.
38. Flaherty JE, Jones JB, Harbaugh BK, Somodi GC, and Jackson LE. 2000. Control of bacterial spot on tomato in the greenhouse and field with H-mutant bacteriophages. HortScience 35:882-884.
39. Gambley CF. 2005.The distribution of citrus canker in Emerald, Australia and bacterial survival in citrus trash. Second international citrus canker and huanglongbing research workshop; November 7-11, 2005; Orlando,Florida. 70.
40. Gill JJ, and Abedon ST. 2003. Bacteriophage Ecology and Plants. APSnet November. http://www.apsnet.org/online/feature/phages/.
41. Gill JJ, Svircev AM, Smith R, and Castle AJ. 2003. Bacteriophages of Erwinia amylovora. Appl Environ Microbiol 69:2133-2138.
42. Goodridge LD. 2004. Bacteriophage biocontrol of plant pathogens: fact or fiction? Trends in biotechnology 22:384-385.

104
43. Goto M. 1992. Citrus canker. Pages 250-269 in: Plant Diseases of International Importance. in: Kumar J, Chaube HS, Singh US, and Mukhopadhyay AN, eds., Prentice-Hall, Englewood Cliff, NJ.
44. Goto M. 1992. Fundamentals of bacterial plant pathology. Academic Press, Inc., San Diego, California.
45. Goto M, Serizawa S, and Morita M. 1971. Studies on citrus canker disease. II. Leaf infiltration technique for detection of Xanthomonas citri (Hasse) Dowson, with special reference to comparison with phage method.
46. Goto M, Takahashi T, and Messina MA. 1980. A comparative study of the strains of Xanthomonas campestris pv. citri isolated from citrus canker in Japan and cancrosis B in Argentina. Annals of the Phytopathological Society of Japan 46:329-338.
47. Goto M, and Yaguchi Y. 1979. Relationship between defoliation and disease severity in citrus canker (Xanthomonas citri). Ann Phytopathol Soc Japan 45:689-694.
48. Gottwald TR, and Graham JH. 1992. A device for precise and nondisruptive stomatal inoculation of leaf tissue with bacterial pathogens. Phytopathology 82:930-935.
49. Gottwald TR, Graham JH, and Schubert TS. Citrus canker: The pathogen and its impact. Plant Health Progress. 2002 2002 August 12 doi:10.1094/PHP-2002-0812-01-RV:2006 October 31. http://www.plantmanagementnetwork.org/pub/php/review/citruscanker.
50. Gottwald TR, Hughes G, Graham JH, Sun X, and Riley T. 2001. The citrus canker epidemic in Florida: The scientific basis of regulatory eradication policy for an invasive species. Phytopathology 91:30-34.
51. Gottwald TR, Sun X, Riley T, Graham JH, Ferrandino F, and Taylor EL. 2002. Geo-referenced spatiotemporal analysis of the urban citrus canker epidemic in Florida. Phytopathology 92:361-377.
52. Goyal SM, Gerba CP, and Bitton G, editors. 1987. Phage ecology. Wiley, New York.
53. Graham JH. 1995. Root regeneration and tolerance of citrus rootstocks to root rot caused by Phytophthora nicotianae. Phytopathology 85:111-117.
54. Graham JH, Bright DB, and McCoy CW. 2003. Phytophthora - Diaprepes weevil complex: Phytophthora spp. relationship with citrus rootstocks. Plant Dis 87 1:85-90.
55. Graham JH, Drouillar DL, Bannwart DL, and Dilley JR. 2000. Bud failure on Swingle citrumelo in a Florida citrus nursery caused by Xanthomonas axonopodis pv. citrumelo. Proceedings of the 13th annual meeting of the Florida State Horticultural Society 112:60-68.
56. Graham JH, and Gottwald TR. 1990. Variation in aggressiveness of Xanthomonas campestris pv. citrumelo associated with citrus bacterial spot in Florida citrus nurseries. Phytopathology 80:190-196.

105
57. Graham JH, Gottwald TR, Cubero J, and Achor DS. 2004. Pathogen profile: Xanthomonas axonopodis pv. citri : factors affecting successful eradication of citrus canker. Mol Plant Pathol 5:1-15.
58. Graham JH, Gottwald TR, Riley TD, and Achor D. 1992. Penetration through leaf stomata and strains of Xanthomonas campestris in citrus cultivars varying in susceptibility to bacterial diseases. Phytopathology 82:1319-1325.
59. Greer GG. 2005. Bacteriophage control of foodborne bacteria. J Food Prot 68:1102-1111.
60. Halbert S. 2005.The discovery of Huanglongbing in Florida. 2nd international citrus canker/Huanglongbing research workshop; November 7-11, 2005; Orlando,Florida. 50.
61. Hendrick CA, and Sequeira L. 1984. Lipopolysaccharide-defective mutants of the wilt pathogen Pseudomonas solanacearum. Appl Environ Microbiol 48:94-101.
62. Hibma AM, Jassim SAA, and Griffiths MW. 1997. Infection and removal of L-forms of Listeria monocytogenes with bred bacteriophage. Int J Food Microbiol 34:197-207.
63. Ignoffo CM, and Garcia C. 1996. Simulated sunlight-UV sensitivity of experimental dust formulations of the nuclear polyhedrosis virus of Helicoverpa/Heliothis. J Invertebr Pathol 67:192-194.
64. Ignoffo CM, and Garcia C. 1995. Aromatic/heterocyclic amino acids and the stimulated sunlight-ultraviolet inactivation of the Heliothis/Helicoverpa baculovirus. Environ Entomol 24:480-482.
65. Ignoffo CM, Garcia C, and Saathoff SG. 1997. Sunlight stability and rain-fastness of formulations of Baculovirus heliothis. Environ Entomol 26:1470-1474.
66. Iriarte FB, Balogh B, Momol MT, and Jones JB. In press. Factors affecting survival of bacteriophage on tomato leaf surfaces. Appl Environ Microbiol.
67. Johnson KB. 1994. Dose-response relationships and inundative biological control. Phytopathology 84:780-784.
68. Katznelson H, and Sutton MD. 1951. A rapid phage plaque count method for the detection of bacteria as applied to the demonstration of internally borne bacterial infection of seed. J Bacteriol 61:689-701.
69. Keel C, Ucurum Z, Michaux P, Adrian M, and Haas D. 2002. Deleterious impact of a virulent bacteriophage on survival and biocontrol activity of Pseudomonas fluorescens Strain CHA0 in Natural Soil. Mol Plant-Microbe Interact 15:567-576.
70. Klement Z, Rudolf K, and Sands DC, editors. 1990. Methods in phytobacteriology. Akadémiai Kiadó, Budapest.

106
71. Koizumi M. 1985. Citrus canker: the world situation. Pages 2-7 in: Citrus canker: An international perspective. Timmer LW, ed. Citrus Research & Education Center, University of Florida, Lake Alfred.
72. Koller W. 1998. Chemical approaches to managing plant pathogens. in: Handbook of integrated pest management. Ruberson JR, ed. Dekker.
73. Kotila JE, and Coons GH. 1925. Investigations on the blackleg disease of potato. Michigan Agr Exper Sta Tech Bull 67:3-29.
74. Kutter E. Phage therapy: bacteriohages as antibiotics Olympia, Washington: The Evergreen State College. 1997 Novermber 15. Available from: http://www.evergreen.edu/phage/phagetherapy/phagetherapy.htm.
75. Leite JRP. 2005. Integrated management of citrus canker in Southern Brazil. Second international citrus canker and huanglongbing research workshop; November 7-11, 2005; Orlando,Florida.
76. Leite JRP, and Mohan SK. 1990. Integrated management of the citrus bacterial canker disease caused by Xanthomonas campestris pv. citri in the State of Paraná, Brazil. Crop Protection 9:3-7.
77. Leverentz B, Conway WS, Alavidze Z, Janisiewicz WJ, Fuchs Y, Camp MJ, Chighladze., and Sulakvelidze A. 2001. Examination of bacteriophage as a biocontrol method for Salmonella on fresh-cut fruit: a model study. J Food Prot 64:1116-1121.
78. Leverentz B, Conway WS, Camp MJ, Janisiewicz WJ, Abuladze T, Yang M, Saftner R, and Sulakvelidze A. 2003. Biocontrol of Listeria monocytogenes on fresh-cut produce by treatment with lytic bacteriophages and a bacteriocin. Appl Environ Microbiol 69:4519-4526.
79. Mallmann WL, and Hemstreet CJ. 1924. Isolation of an inhibitory substance from plants. Arg Res 28:599-602.
80. Manulis S, Zutra D, Kleitman F, Dror O, David I, Zilberstaine M, and Shabi E. 1998. Distribution of streptomycin-resistant strains of Erwinia amylovora in Israel and occurrence of blossom blight in the autumn. Phytoparasitica 26:223-230.
81. Marco GM, and Stall RE. 1983. Control of bacterial spot of pepper initiated by strains of Xanthomonas campestris pv. vesicatoria that differ in sensitive to copper. Plant Dis 67:779-781.
82. McKenna F, El-Tarabily KA, Hardy GESTJ, and Dell B. 2001. Novel in vivo use of a polyvalent Streptomyces phage to disinfest Streptomyces scabies-infected seed potatoes. Plant Pathol 50:666-675.
83. McNeil DL, Romero S, Kandula J, Stark C, Stewart A, and Larsen S. 2001. Bacteriophages: A potential biocontrol agent against walnut blight (Xanthomonas campestris pv. juglandis). N Z Plant Prot 54:220-224.

107
84. Minsavage GV, Canteros BI, and Stall RE. 1990. Plasmid-mediated resistance to streptomycin in Xanthomonas campestris pv. vesicatoria. Phytopathology 80:719-723.
85. Momol MT, Jones JB, Olson SM, Obradovic A, Balogh B, King P. Integrated management of bacterial spot on tomato in Florida. EDIS, Institute of Food and Agricultural Sciences, University of Florida, Online 2002 September. Report No.: PP110.
86. Munsch P, and Olivier JM. 1995. Biocontrol of bacterial blotch of the cultivated mushroom with lytic phages: Some practical considerations. Pages 595-602 in: Science and cultivation of edible fungi, Vol II: Proceedings of the 14th international congress. Elliot TJ, ed.
87. Munsch P, Olivier JM, and Houdeau G. 1991. Experimental control of bacterial blotch by bacteriophages. Pages 389-396 in: Science and cultivation of edible fungi. in: Maher MJ, ed. Balkema, Rotterdam, The Netherlands.
88. Muraro RP, Roka F, and Spreen TH. 2000.An overview of Argentina’s citrus canker control program with applicable costs for a similar program in Florida. Proceedings of the international citrus canker research workshop; June 20-22, 2000; Ft. Pierce FL. Online: Division of Plant Industry, Florida Department of Agriculture and Consumer Services.
89. Obradovic A, Jones JB, Momol MT, Balogh B, and Olson SM. 2004. Management of tomato bacterial spot in the field by foliar applications of bacteriophages and SAR inducers. Plant Dis 88:736-740.
90. Obradovic A, Jones JB, Momol MT, Olson SM, Jackson LE, Balogh B, Guven K, and Iriarte FB. 2005. Integration of biological control agents and systemic acquired resistance inducers against bacterial spot on tomato. Plant Dis 89:712-716.
91. Okabe N, and Goto M. 1963. Bacteriophages of plant pathogens. Annu Rev Phytopathol 1:397-418.
92. Ophel KM, Bird AF, and Kerr A. 1993. Association of bacteriophage particles with toxin production by Clavibacter toxicus, the causal agent of annual ryegrass toxicity. Phytopathology 83:676-681.
93. Osawa S, Furuse K, and Watanabe I. 1981. Distribution of ribonucleic acid coliphages in animals. Appl Environ Microbiol 45:164-168.
94. Pruvost O, Hartung JS, Civerolo EL, Dubois C, and Perrier X. 1992. Plasmid DNA fingerprints distinguish pathotypes of Xanthomonas campestris pv. citri , the causal agent of citrus bacterial canker disease. Phytopathology 82:485-490.
95. Randhawa PS, and Civerolo EL. 1986. Interaction of Xanthomonas campestris pv. pruni with pruniphage and epiphytic bacteria on detached peach leaves. Phytopathology 76:549-553.
96. Reichelderfer K, and Barry JW. 1995. Introduction. Pages 28-35 in: Biorational pest control agents: formulation and delivery. in: Hall FR, andBarry JW, eds.American Chemical Society, Washington, DC.

108
97. Rizvi S, and Mora PT. 1963. Bacteriophage plaque-count assay and confluent lysis on plates without bottom agar layer. Nature 200:1324-1325.
98. Rybak MA. 2005. Genetic determinants of host range specificity of the Wellington strain of Xanthomonas axonopodis pv. citri [dissertation]. Gainesville, Florida: University of Florida.
99. Saccardi A, Gambin E, Zaccardelli M, Barone G, and Mazzucchi U. 1993. Xanthomonas campestris pv. pruni control trials with phage treatments on peaches in the orchard. Phytopathol Mediterr 32:206-210.
100. Sambrook J, Fritsch EF, and Maniatis T. 1989. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
101. Schnabel EL, Fernando WGD, Meyer MP, Jones AL, and Jackson LE. 1999. Bacteriophage of Erwinia amylovora and their potential for biocontrol.
102. Schoulties CL, Civerolo EL, Miller JW, Stall RE, Krass CJ, Poe SR, and Ducharme EP. 1987. Citrus canker in Florida. Plant Dis 71:388-395.
103. Schubert TS, Rizvi SA, Sun X, Gottwald TR, Graham JH, and Dixon WN. 2001. Meeting the challenge of eradicating citrus canker in Florida - Again. Plant Dis 85:340-356.
104. Shaner G, and Finnley RE. 1977. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat. Phytopathology 67:1051-1056.
105. Shapiro JP, and Gottwald TR. 1995. Resistance of eight cultivars of citrus rootstock to a larval root weevil, Diaprepes abbreviatus L. (Coleoptera: Curculionidae). J Econ Entomol 88:148-154.
106. Stall RE, and Civerolo EL. 1991. Research relating to the recent outbreak of citrus canker in Florida. Annu Rev Phytopathol:399-420.
107. Stall RE, Marco GM, and Canteros de Echenique BI. 1982. Importance of mesophyll in mature-leaf resistance to cancrosis of citrus. Phytopathology 72:1097-1100.
108. Stall RE, Miller JW, Marco GM, and Canteros de Echenique BI. 1980. Population dynamics of Xanthomonas citri causing cancrosis of citrus in Argentina. Proc Fla Hort Soc 93:10-14.
109. Stein B, Ramallo J, Foguet L, and Morandini M. 2005.Chemical control of citrus canker in lemons (Citrus limon, (L.) Burm. f) in Tucuman, Argentina (Abstr.). Second international citrus canker and huanglongbing research workshop; November 7-11, 2005; 25.
110. Storey MV, and Ashbolt NJ. 2001. Persistence of two model enteric viruses (B40-8 and MS-2 bacteriophages) in water distribution pipe biofilms. Water Science and Technology 43:133-138.

109
111. Stover E, Castle B, and Hebb J. Citrus rootstock usage in the Indian river region Online: Electronic Data Information Source (EDIS) HS-852, Horticultural Science Department, IFAS, University of Florida. 2002 April. Available from: http://edis.ifas.ufl.edu/HS129.
112. Summers WC. 2005. Bacteriophage research: Early History. Pages 5-27 in: Bacteriophages: biology and applications. in: Kutter E, and Sulakvelidze A, eds. CRC Press, Boca Raton, Florida.
113. Sun X, Jones D, and Duan YP. 2005.Detection of Manatee genotypes of citrus canker bacteria in Hillsborough Co, Florida. Second international citrus canker and huanglongbing research workshop; November 7-11, 2005; Orlando,Florida. 32.
114. Sun X, Stall RE, Jones JB, Cubero J, Gottwald TR, Graham JH, Dixon WN, Schubert TS, Chaloux PH, Stromberg VK, Lacy GH, and Sutton BD. 2004. Detection and characterization of a new strain of citrus canker bacteria from Key/Mexican lime and alemow in south Florida. Plant Dis 88:1179-1188.
115. Svircev AM, Lehman SM, Kim W-, Barszcz E, Schneider KE, and Castle AJ. 2006. Zeller W, Ullrich C, editors. Control of the fire blight pathogen with bacteriophages. Proceedings of the 1st international symposium on biological control of bacterial plant diseases; October 23-26, 2005; Seeheim/Darmstadt, Germany. Berlin, Germany: Die Deutsche Bibliothek - CIP-Einheitsaufnahme 259-261.
116. Sykes IK, Lanning S, and Williams ST. 1981. The effect of pH on soil actinophage. J Gen Microbiol 122:271-280.
117. Tanaka H, Negishi H, and Maeda H. 1990. Control of tobacco bacterial wilt by an avirulent strain of Pseudomonas solanacearunm M4S and its bacteriophage. Ann Phytopath Soc Japan 56:243-246.
118. Tanji Y, Shimada T, Yoichi M, Miyanaga K, Hori K, and Unno H. 2004. Toward rational control of Escherichia coli O157:H7 by a phage cocktail. Appl Microbiol Biotechnol 64:270-274.
119. Thayer PL, and Stall RE. 1961. A survey of Xanthomonas vesicatoria resistance to streptomycin. Proc Fla Hort Soc 75:163-165.
120. Thomas RC. 1935. A bacteriophage in relation to Stewart’s disease of corn. Phytopathology 25:371-372.
121. Timmer LW, Zitko SE, Gottwald TR, and Graham JH. 2000. Phytophthora brown rot of citrus: Temperature and moisture effects on infection, sporangium production, and dispersal. Plant Dis 84:157-163.
122. UNCTAD INFO COMM Citrus fruit Online: United Nations Conference on Trade and Development, Market Information in the Commodities Area [cited 2006 October 31]. Available from: http://www.unctad.org/infocomm/anglais/orange/market.htm#prod.

110
123. USDA APHIS. Citrus canker Online: United States Department of Agriculture Animal and Plant Health Inspection Service. 2006 October 5. Available from: http://www.aphis.usda.gov/newsroom/hot_issues/citrus_canker/citrus_canker.shtml.
124. USDA FAS. Situation and outlook 2005/06 Online: United States Department of Agriculture Foreign Agricultural Service. 2006 February 20. Available from: http://www.fas.usda.gov/htp/Hort_Circular/2006/02-06/02-20-06%20Citrus%20Feature.pdf.
125. USDA NASS. Citrus fruits - 2006 Summary Online: United States Department of Agriculture National Agricultural Statistics Service. 2006 September 21. Available from: http://usda.mannlib.cornell.edu/usda/current/CitrFrui/CitrFrui-09-21-2006.pdf.
126. Verniere C, Hartung JS, Pruvost OP, Civerolo EL, Alvarez AM, Maestri P, and Luisetti J. 1998. Characterization of phenotypically distinct strains of Xanthomonas axonopodis pv. citri from southwest Asia. Eur J Plant Pathol 104:477-487.
127. Vidaver AK. 1976. Prospects for control of phytopathogenic bacteria by bacteriophages and bacteriocins. Annu Rev Phytopathol:14.
128. Vidaver AK, and Schuster ML. 1960. Characterization of Xanthomonas phaseoli bacteriophages. J Virol 4:300-308.
129. Voloudakis AE, Reignier TM, and Cooksey DA. 2005. Regulation of resistance to copper in Xanthomonas axonopodis pv. vesicatoria. Appl Environ Microbiol 71:782-789.
130. Wagner PL, and Waldor MK. 2002. Bacteriophage control of bacterial virulence. Infection and immunity 70:3985-3993.
131. Wiggins BA, and Alexander M. 1985. Minimum bacterial density for bacteriophage replication: implications for significance of bacteriophages in natural ecosystems. Appl Environ Microbiol 49:19-23.
132. Williams ST, Mortimer AM, and Manchester L. 1987. Ecology of soil bacteriophages. Pages 157-179 in: Phage ecology. in: Goyal SM, Gerba CP, and Bitton G, eds. John Wiley and Sons, New York.
133. Woods TL, Israel HW, and Sherf AF. 1981. Isolation and Partial Characterization of a bacteriophage of Erwinia stewartii from the corn flea beetle, Chaetocnema pulicaria. Prot.Ecol 3:229-236.
134. Wu WC. 1972. Phage-induced alterations of cell disposition, phage adsorption and sensitivity, and virulence in Xanthomonas citri. Ann Phytopath Soc Japan 38:333-341.
135. Wu WC, Lee ST, Kuo HF, and Wang LY. 1993. Use of phages for indentifying the citrus canker bacterium Xanthomonas campestris pv. citri in Taiwan. Plant Pathol 42:389-395.
136. Yang Y, and Gabriel DW. 1995. Xanthomonas avirulence/pathogenicity gene family encodes functional plant nuclear targeting signals. MPMI 8:627-631.

111
137. Zaccardelli M, Saccardi A, Gambin E, and Mazzucchi U. 1992. Xanthomonas campestris pv. pruni bacteriophages on peach trees and their potential use for biological control. Phytopathol Mediterr 31:133-140.
138. Zansler ML, Spreen TH, Muraro RP. Florida's citrus canker eradication program (CCEP): Benefit-cost analysis. Electronic Data Information Source, IFAS, University of Florida, Online 2005 March. Report No.: FE 531.
139. Zansler ML, Spreen TH, Muraro RP. Florida's citrus canker eradication program (CCEP): Summary of annual costs and benefits. Electronic Data Information Source , IFAS, University of Florida; , Online 2005 March. Report No.: FE 532.

112
BIOGRAPHICAL SKETCH
Botond Balogh was born to Balogh Antal and Gyuris Borbála, in Kalocsa, Hungary, on
March 25, 1976. He graduated from the Ságvári Endre High School in Szeged in 1994 with
mathematics major. He attended the University of Agronomy, College of Agricultural Sciences,
in Gödöllő from 1994 to 1998, majoring in horticulture and plant breeding – genetics. He
participated in a research program for pepper breeding via anther and microspore culture under
the guidance of Judith Mitykó in the Agricultural Biotechnological Center, Gödöllő, from 1996
to 1998. In 1998 he took part in a year-long agricultural exchange program organized by the
Communicatin for Agriculture Exchange Program. In the summer and fall of 1999 he attended
Brevard Community College in Cocoa, FL. He was admitted to the University of Florida,
College of Agricultural and Life Sciences, in January 2000, and graduated with the degree of
Bachelor of Science with the plant science major and plant pathology minor in August 2000.
During his undergraduate studies he participated in a research project characterizing a mobile
genetic element found in the genome of several strains of Xanthomonas campestris pv.
vesicatoria, a plant pathogenic bacterium. From August 2000 he enrolled to the graduate
program of the University of Florida, College of Agricultural and Life Sciences, Department of
Plant Pathology. He graduated with a degree of Master of Scence in May 2002. The title of his
thesis was: “Strategies For Improving The Efficacy Of Bacteriophages For Controlling Bacterial
Spot Of Tomato”. He continued with his doctoral studies at the Plant Pathology department and
conducted a research project on evaluating bacteriophages for controlling citrus canker and citrus
bacterial spot.

![[PPT]Transformations! Translations, Reflections, and …plaza.ufl.edu/mel97/EME_4401_Micro_Micro_Teaching.ppt · Web viewTransformations! Translations, Reflections, and Rotations](https://static.fdocuments.us/doc/165x107/5ab1fe777f8b9a7e1d8d114a/ppttransformations-translations-reflections-and-plazaufledumel97eme4401micromicro.jpg)
















