
Characterisation of the Bax-nucleophosmin interaction: the importance of the Bax C-terminus
10
ORIGINAL PAPER Characterisation of the Bax-nucleophosmin interaction: the importance of the Bax C-terminus Jane Thompson Keith Finlayson Eliane Salvo-Chirnside David MacDonald James McCulloch Lorraine Kerr John Sharkey Published online: 24 January 2008 Ó Springer Science+Business Media, LLC 2008 Abstract The molecular chaperone nucleophosmin has been identified as a novel Bax binding protein with this interaction proposed to be a key event in the activation and translocation of Bax in mitochondrial dysfunction and apoptotic cell death. Using a proximity assay, we have quantitatively defined the high affinity and saturable inter- action between Bax and nucleophosmin indicative of a competitive and specific mechanism. Binding of full length Bax to nucleophosmin was only observed after conforma- tional change was induced using non-ionic detergents (e.g., NP-40). The Bax-nucleophosmin interaction was inhibited by a Bax C-terminal antibody (IC 50 = 1 nM) but minimally affected by antibodies directed against either the N-terminus or a-helices 4 and 5. Bcl-2 and p53 inhibited the interaction between full length activated Bax and nucleophosmin. The proximity assay based on the Bax-nucleophosmin interac- tion was robust and reproducible (Z 0 = 0.50) facilitating its use for screening a small chemical library. A low molecular weight non-peptide compound, 2-(5-methyl-2-phenyl-1, 3-thiazol-4-yl)ethanohydrazide, partially inhibited the Bax- nucleophosmin interaction (IC 50 = 100 nM) and also attenuated UV-induced cell death of HEK293 cells. The present investigations demonstrate the importance of exposure of the C-terminus of Bax for its interaction with nucleophosmin. These protein–protein interaction assays provide a technical approach both for the study of Bax- interacting proteins and for the discovery of novel anti- apoptotic agents. Keywords Bcl-2 family Apoptosis Cell death Proximity assay Introduction Bax, a pro-apoptotic member of the Bcl-2 family, is pri- marily found in the cytosol of healthy cells in a soluble inactive state while anti-apoptotic members such as Bcl-2 and Bcl-x L act to protect the outer mitochondrial mem- brane (OMM) integrity [1, 2]. Bcl-2 and Bcl-x L both contain domains BH1-4 and protect the OMM from dis- ruption by Bax [3, 4]. Evidence has shown that insertion of the BH3 domain of Bax into a hydrophobic cleft on the surface of Bcl-2 and Bcl-x L appear necessary for their hetero-dimerization with Bax for the suppression of apoptosis [5]. Following an apoptotic stimulus, BH3 only proteins can activate Bax [6], leading to a conformational change and exposure of its N- and C-terminal domains. The C-terminus of Bax is then targeted to the OMM enabling membrane insertion and pore formation [7]. The resultant loss of cytochrome c and apoptogenic factors through mitochondrial pores leads to the formation of apoptosomes, activation of effector caspses and cell death ensues [8]. L. Kerr and J. Sharkey have contributed equally to this work. J. Thompson (&) K. Finlayson E. Salvo-Chirnside D. MacDonald J. McCulloch L. Kerr J. Sharkey Astellas CNS Research in Edinburgh, The University of Edinburgh, The Chancellors Building, 49 Little France Crescent, Edinburgh EH16 4SB, UK e-mail: [email protected] Present Address: J. Thompson Cyclacel Pharmaceutical Inc, Dundee Technopole, James Lindsay Place, Dundee DD1 5JJ, UK Present Address: E. Salvo-Chirnside Institute of Stem Cell Research, The University of Edinburgh, Kings Buildings, West Mains Road, Edinburgh EH9 3JQ, UK 123 Apoptosis (2008) 13:394–403 DOI 10.1007/s10495-007-0177-2
-
Upload
jane-thompson -
Category
Documents
-
view
215 -
download
1
Transcript of Characterisation of the Bax-nucleophosmin interaction: the importance of the Bax C-terminus

ORIGINAL PAPER
Characterisation of the Bax-nucleophosmin interaction:the importance of the Bax C-terminus
Jane Thompson Æ Keith Finlayson Æ Eliane Salvo-Chirnside Æ David MacDonald ÆJames McCulloch Æ Lorraine Kerr Æ John Sharkey
Published online: 24 January 2008
� Springer Science+Business Media, LLC 2008
Abstract The molecular chaperone nucleophosmin has
been identified as a novel Bax binding protein with this
interaction proposed to be a key event in the activation and
translocation of Bax in mitochondrial dysfunction and
apoptotic cell death. Using a proximity assay, we have
quantitatively defined the high affinity and saturable inter-
action between Bax and nucleophosmin indicative of a
competitive and specific mechanism. Binding of full length
Bax to nucleophosmin was only observed after conforma-
tional change was induced using non-ionic detergents (e.g.,
NP-40). The Bax-nucleophosmin interaction was inhibited
by a Bax C-terminal antibody (IC50 = 1 nM) but minimally
affected by antibodies directed against either the N-terminus
or a-helices 4 and 5. Bcl-2 and p53 inhibited the interaction
between full length activated Bax and nucleophosmin. The
proximity assay based on the Bax-nucleophosmin interac-
tion was robust and reproducible (Z0 = 0.50) facilitating its
use for screening a small chemical library. A low molecular
weight non-peptide compound, 2-(5-methyl-2-phenyl-1,
3-thiazol-4-yl)ethanohydrazide, partially inhibited the Bax-
nucleophosmin interaction (IC50 = 100 nM) and also
attenuated UV-induced cell death of HEK293 cells. The
present investigations demonstrate the importance of
exposure of the C-terminus of Bax for its interaction with
nucleophosmin. These protein–protein interaction assays
provide a technical approach both for the study of Bax-
interacting proteins and for the discovery of novel anti-
apoptotic agents.
Keywords Bcl-2 family � Apoptosis � Cell death �Proximity assay
Introduction
Bax, a pro-apoptotic member of the Bcl-2 family, is pri-
marily found in the cytosol of healthy cells in a soluble
inactive state while anti-apoptotic members such as Bcl-2
and Bcl-xL act to protect the outer mitochondrial mem-
brane (OMM) integrity [1, 2]. Bcl-2 and Bcl-xL both
contain domains BH1-4 and protect the OMM from dis-
ruption by Bax [3, 4]. Evidence has shown that insertion of
the BH3 domain of Bax into a hydrophobic cleft on the
surface of Bcl-2 and Bcl-xL appear necessary for their
hetero-dimerization with Bax for the suppression of
apoptosis [5]. Following an apoptotic stimulus, BH3 only
proteins can activate Bax [6], leading to a conformational
change and exposure of its N- and C-terminal domains. The
C-terminus of Bax is then targeted to the OMM enabling
membrane insertion and pore formation [7]. The resultant
loss of cytochrome c and apoptogenic factors through
mitochondrial pores leads to the formation of apoptosomes,
activation of effector caspses and cell death ensues [8].
L. Kerr and J. Sharkey have contributed equally to this work.
J. Thompson (&) � K. Finlayson � E. Salvo-Chirnside �D. MacDonald � J. McCulloch � L. Kerr � J. Sharkey
Astellas CNS Research in Edinburgh, The University of
Edinburgh, The Chancellors Building, 49 Little France Crescent,
Edinburgh EH16 4SB, UK
e-mail: [email protected]
Present Address:J. Thompson
Cyclacel Pharmaceutical Inc, Dundee Technopole,
James Lindsay Place, Dundee DD1 5JJ, UK
Present Address:E. Salvo-Chirnside
Institute of Stem Cell Research, The University of Edinburgh,
Kings Buildings, West Mains Road, Edinburgh EH9 3JQ, UK
123
Apoptosis (2008) 13:394–403
DOI 10.1007/s10495-007-0177-2

Several proteins in the cytosol have been identified as
molecular chaperones or retention factors which regulate
monomeric Bax, preventing the conformational changes
associated with apoptosis and protecting against activation
by BH3 only proteins. Nucleophosmin has recently been
identified as a Bax interacting protein [9]. It is a nucleolar
phosphoprotein and molecular chaperone which is abun-
dantly expressed in the nucleolus and continuously shuttles
between the nucleus and cytoplasm to prevent protein
aggregation, promote histone assembly and acetylation
dependent transcription [10–12]. Nucleophosmin has been
shown to be localized in the cytosol prior to Bax mito-
chondrial translocation following staurospaurine induced
apoptosis in SHSY5Y cells in vitro and focal cerebral
ischemia in vivo [9].
We have developed an in vitro based binding assay
(amplified luminescent proximity homogenous assay) to
study this novel protein–protein interaction at the molecular
level. We established conditions to examine the binding of a
C-terminal peptide and full length Bax protein to nucleo-
phosmin. Various antibodies or peptides known to interact
with Bax or nucleophosmin were used in the assay to allow
a systematic characterisation of the interaction between
nucleophosmin and full length Bax or a C-terminal Bax
peptide. We have demonstrated that Bax binds to nucleo-
phosmin by its C-terminal tail and that small molecule
inhibitors could be identified. Recent studies indicating that
nucleophosmin may be a chaperone involved in transloca-
ting activated Bax to the mitochondria at the critical point of
apoptotic cell death, highlights the therapeutic potential of
modulating the Bax-nucleophosmin interaction which
should aide the identification of drugs for the treatment of
certain neurodegenerative disorders or neoplasias.
Materials and methods
All chemicals were obtained from Sigma (Dorset, UK)
unless otherwise indicated.
Expression and purification of recombinant Bax
and nucleophosmin
The cDNAs encoding Bax and nucleophosmin (Invitrogen,
Paisley, UK) were subcloned from Gateway� pENTRTM221
UltimateTM human ORF clones. Using forward and reverse
primers respectively for Bax (50-ATG GAC GGG TCC GGG
GAG CA-30 and 50-CTA GCC CAT CTT CTT CCA GAT
GG-30) and nucleophosmin (50-ATG GAA GAT TCG ATG
GAG ATG G-30 and 50-GCT GGG TTC TAA AGA GAC
TTC C-30), PCR was used to obtain the complete coding
regions from the pENTRTM221 clones. Amplified products
for Bax and nucleophosmin were TOPO-TA cloned into the
N-terminal His-tag fusion vector pCRT7-NT-TOPO (Invit-
rogen) and sequenced on both strands by automated
sequencing (MWG, London, UK). The nucleophosmin
construct was transformed into BL21 cells (Invitrogen) and
protein expression induced by 1 mM IPTG for 4 h. Cells
were lysed (Fast-break lysis buffer, 1/10 dilution; Promega,
Southampton, UK) and His-tagged nucleophosmin captured
with MagneHisTM beads (for 2 min, Promega). Beads were
captured using a magnetic stand and washed 4 times in
MagneHisTM binding/wash buffer. Bound protein was eluted
in 500 mM imidazole and dialysed against Tris buffer
(50 mM Tris-HCl pH7.4, 150 mM NaCl, 1 mM EDTA,
5 mM DTT, 0.5% NP-40, 0.5% Triton-X100, 10% glycerol)
using Slide-A-Lyzer cassettes (10 KDa MWCO; Pierce,
Northumberland, UK). Full length His-tagged Bax was
produced using ExpresswayTM in vitro expression system
(Invitrogen) according to the manufacturer’s instructions
using 1 lg pCRT7-TOPO-Bax DNA. Recombinant Bax was
purified using MagneHisTM purification kit (Promega) as
described for nucleophosmin except final dialysis was
against 150 mM NaCl, 10 mM HEPES-KOH pH7.4.
Immunoprecipitation and Western blotting
Immunoprecipitation was performed using an antibody
against active Bax (Bax 6A7, 5/200 mg protein; BD Bio-
sciences, Oxford, UK). Complexes were isolated with protein
G-conjugated paramagnetic dynabeads (Dynal Biotech,
Bromborough, UK) and analysed by Western blotting using
BaxNT antibody (1/2000; Upstate, Chandlers Ford, UK).
Biotinylation of Bax protein
To enable conjugation to streptavidin-coated donor beads
(PerkinElmer, Beaconsfield, UK) full length Bax was
biotinylated using the ProtOnTM Biotin labelling Kit
(Vector Laboratories, Peterborough, UK) as described by
the manufacturer. Briefly, 1 mg of recombinant Bax in
100 ll of buffer (150 mM NaCl, 10 mM HEPES-KOH
pH7.4) was biotinylated by adding 2 ll of labelling
reagent. After 45 min incubation at room temperature, the
reaction was terminated by the addition of 2 ll stop solu-
tion. To remove excess biotin from the labelled Bax,
dialysis was carried out against 2 l of buffer (150 mM
NaCl, 10 mM HEPES-KOH pH7.4) overnight at 4�C.
Proximity assay
Protein–protein interactions were analysed using an
amplified luminescence proximity homogeneous assay
Apoptosis (2008) 13:394–403 395
123

(AlphaScreenTM technology; PerkinElmer) in 384-well
white polystyrene proxiplates (PerkinElmer). Assays com-
prised nucleophosmin monoclonal antibody-conjugated
acceptor beads and streptavidin-coated donor beads (20 lg/
ml of each), Bax (biotinylated peptide or full length protein)
and nucleophosmin, with putative inhibitors as indicated, in
a total assay volume of 25 ll unless otherwise stated.
A monoclonal antibody directed against the C-terminus
of nucleophosmin (Sigma) was conjugated with Alpha-
ScreenTM acceptor beads (PerkinElmer) using a standard
amination procedure. The nucleophosmin antibody
(0.01 mg) was added to 0.3 mg of unconjugated acceptor
beads in 5% sodium cyanoborohydride, 0.02 M 2-[N-
morpholino] ethanesulfonic acid (MES), pH6.0 and incu-
bated for 48 h at 37�C in the dark. A block was performed
with 0.1 M carboxymethoxylamine-hemihydrochloride
(CMO; 1 h) and the beads washed 3 times in 0.1 M Tris
buffer pH8.0. Conjugated beads were finally prepared as a
5 mg/ml solution in 25 mM HEPES, 0.1 M NaCl, pH 7.4
(PerkinElmer).
The left and right arms of the proximity assay were
verified to determine if the assay was functional. To
ascertain if biotinylated Bax C-terminal peptide bound to
the streptavidin-coated donor beads, biotinylated acceptor
beads (20 ug/ml) were incubated for 30 min at 30�C with
biotinylated Bax C-terminal peptide (biotin-TVTIF-
VAGVLTASLTIWKKMG; CSS-Albachem, Gladsmuir,
UK) prepared in 150 mM NaCl, 10 mM HEPES-KOH
pH7.4, 0.1% Tween 20) at a concentration range (0.03–
3,000 nM). Streptavidin-coated donor beads were added
and incubation performed for a further 60 min prior to
analysis using a multi-label plate reader (Envision Exci-
teTM; PerkinElmer). Similarly to determine successful
conjugation of nucleophosmin monoclonal antibody to
acceptor beads, the putatively conjugated acceptor beads
were incubated for 30 min with biotinylated secondary
antibody at a concentration range (0.003–3 nM). Strepta-
vidin-coated donor beads were added and incubation
performed for a further 60 min prior to analysis.
The optimal concentrations of Bax C-terminal peptide
and nucleophosmin required to produce a maximal signal
in the assay were established by incubating constant con-
centrations of nucleophosmin (5, 15, 150 and 350 nM) with
Bax C-terminal peptide at a concentration range (0.3–
1,000 nM). The binding of the two molecules at the opti-
mal concentrations (100 nM Bax C-terminal peptide and
15 nM nucleophosmin) was also measured over time (0–
4 h). An optimal incubation for the proteins following
donor bead addition was established at 90 min and was
used thereafter.
To investigate the interaction between full length Bax
and nucleophosmin, serial dilutions of each protein were
performed in assay buffer (150 mM NaCl, 10 mM HEPES-
KOH pH7.4, 0.01% BSA) containing either 0.1% CHAPS,
0.1% Nonidet P-40 (NP-40) or 0.1% Tween 20. Due to the
instability of the full length Bax protein in long-term
storage, production was limited to small quantities. The
concentration of full length Bax protein used in subsequent
competition assays (38 nM) was determined from dilution
of stock preparation producing the maximal signal in a
serial dilution study.
Competition studies were performed using Bax biotin-
ylated C-terminal peptide (100 nM) or full length protein
(38 nM), nucleophosmin (15 nM) and putative inhibitors (if
present) at concentrations ranging from (0.03–1,000 nM)
preincubated at room temperature for 45 min. Nucleo-
phosmin antibody-conjugated acceptor beads were added
and a further incubation of 30 min at 30�C performed. After
the addition of streptavidin-coated donor beads, the plate
was incubated at 30�C for a further 90 min prior to analysis.
Putative peptide inhibitors used were Bcl-2 (His-tagged,
C-terminal truncated; Merck, Nottingham, UK), p53 pep-
tide (Abcam, Cambridge, UK), antibodies raised against
the Bax N-terminus (Bax-NT; Upstate), a-helices 4 and 5
(Ab4; Merck) and C-terminus (1D3; Abcam). C-terminal
Bax antibody at concentration range (0.03–1,000 nM) was
used as a positive inhibition control of the Bax-nucleo-
phosmin interaction while buffer and the individual
proteins alone served as negative controls.
Identification of non-peptide inhibitors
of the Bax-nucleophosmin interaction
A small library of 800 chemically diverse compounds
(HitfinderTM, Maybridge, Cornwall, UK) was assessed for
the ability to block the Bax-nucleophosmin interaction.
Compounds, supplied as a dry film (1 lmole) in 96 well
plates, were diluted to a 50 mM master stock in 100%
DMSO. The final concentration of compounds used in the
assay was 1 lM in 0.02% DMSO. C-terminal Bax peptide
and nucleophosmin were prepared in assay buffer contain-
ing 0.1% Tween 20 to a final assay concentration of 100 nM
and 15 nM respectively and the assay performed as
described above. C-terminal Bax antibody (3 nM final
concentration) was used as a positive inhibition control of
the Bax peptide-nucleophosmin interaction while buffer and
the individual proteins alone served as negative controls.
Cell culture and UV-induced apoptosis
HEK293 cells were seeded into 4 well plates at a density of
0.5 9 105 cells/well and maintained in Dulbecco’s Modi-
fied Eagles Medium (DMEM) supplemented with 2 mM
Glutamine and 1% non essential amino acids at 37�C in a
396 Apoptosis (2008) 13:394–403
123

humidified 5% CO2 atmosphere. Following a 24 h incu-
bation period media was removed from wells and treated
with UV-C (30 J/m2, 254 nm) for 5 s using a GS gene
linker UV chamber (BioRad, Hemel Hempstead, UK) with
the lids removed. After irradiation, the cells were supplied
with complete medium containing either vehicle alone
(0.01% DMSO) or vehicle plus 10 lM inhibitor (2-(5-
methyl-2-phenyl-1,3-thiazol-4-yl) ethanohydrazide). Con-
trol cells which did not receive UV irradiation were
replaced with the same medium as experimental plates.
Cell viability was then measured by MTS assay (Promega)
following a further incubation period of 24 h.
Results
Development of a Bax-nucleophosmin proximity assay
Nucleophosmin was identified as a Bax-binding protein by
affinity chromatography [9]. We have subsequently devel-
oped a proximity assay to characterise this novel protein–
protein interaction in detail. The principle behind the
amplified luminescent proximity homogenous assay (Al-
phaScreenTM) is that upon excitation, a chemiluminescent
signal is produced when donor and acceptor beads are sep-
arated by a distance of less than 200 nm. Thus when two
proteins of interest (separately conjugated to donor and
acceptor beads) interact, the beads are brought into close
proximity and a luminescent signal is produced [13]. We
have used this technology to characterise the interaction
between Bax and nucleophosmin and a schematic diagram of
the assay design is depicted in Fig. 1a. A biotinylated peptide
corresponding to the last 21 C-terminal amino acids of Bax
was attached to streptavidin-coated donor beads. Attachment
was verified by the ability of this peptide to disrupt the
interaction between streptavidin-coated donor beads and
biotin-coated acceptor beads (Fig. 1b, c). A monoclonal
antibody raised against nucleophosmin was directly conju-
gated to acceptor beads and to confirm this, a biotinylated
secondary antibody was used to link streptavidin-coated
donor beads thus generating a signal (Fig. 1d, e). Having
verified both halves of the proximity assay (biotinylated Bax
peptide attachment to streptavidin-coated donor beads and
acceptor beads conjugated with nucleophosmin monoclonal
antibody) optimal assay conditions were determined.
C-terminal Bax peptide (0.3–100 nM) produced a con-
centration-dependent increase in binding signal when
incubated with constant concentrations of nucleophosmin.
The maximal signal was observed when the nucleophosmin
concentration was 15 nM (Fig. 2a). Using this concentra-
tion of nucleophosmin and varying the Bax concentration,
a maximal signal was observed with 100 nM C-terminal
Bax peptide (Fig. 2b). Using these optimal concentrations,
the luminescent signal increased with incubation time
reaching equilibrium at 90 min (Fig. 2c). Therefore all
subsequent experiments utilizing Bax peptide were per-
formed with 100 nM C-terminal Bax peptide and 15 nM
nucleophosmin with incubation for at least 90 min.
Nucleophosmin only binds to active Bax
To investigate whether nucleophosmin interacts with full
length Bax, recombinant protein was expressed in vitro,
biotinylated and attached to streptavidin-coated donor
beads. However, no binding between nucleophosmin and
full length Bax was observed unless a conformational
change was induced by the non-ionic detergents Nonidet
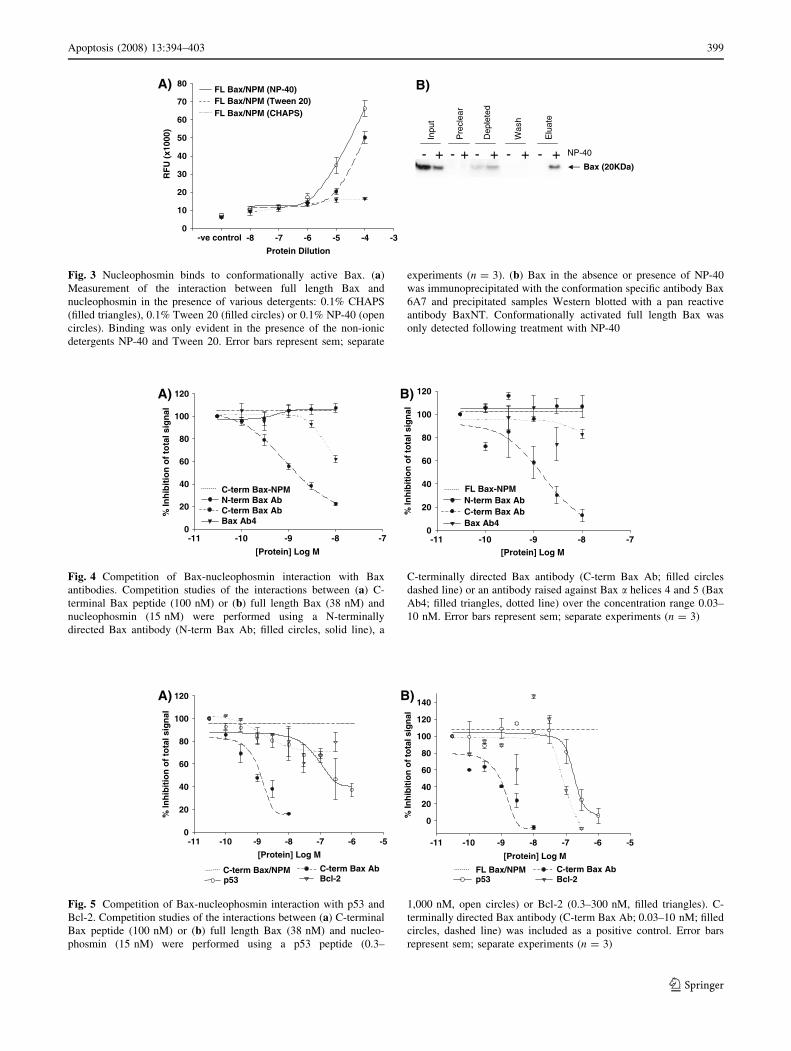
P-40 (NP-40) and Tween 20 (Fig. 3a). This is consistent
with the view that the C-terminus of inactive Bax is
sequestered within a hydrophobic pocket of the BH3
domain and therefore not available to interact with other
proteins unless conformationally activated [14]. Our initial
experiments were performed in 0.1% CHAPS-containing
buffer which does not alter the conformation of Bax, unlike
non-ionic detergents [15]. Verification that NP-40 induced
a Bax conformational change was obtained by immuno-
precipitation using the conformation-sensitive Bax 6A7
antibody (Fig. 3b). Moreover, in the presence of both non-
ionic detergents nucleophosmin-Bax interactions were
detected (Fig. 3a), the greatest being observed with NP-40.
In all further experiments using full length Bax, NP-40 was
included in the reaction buffer.
Molecular analysis of Bax-nucleophosmin interaction
To elucidate further which regions of Bax are involved in
binding nucleophosmin, competition studies were per-
formed with antibodies raised against specific domains of
Bax. An antibody targeted to the C-terminus of Bax (C-
term Ab) inhibited the Bax peptide-nucleophosmin inter-
action with an apparent IC50 of 1.66 ± 0.54 nM
(mean ± SEM; Fig. 4a). While no inhibition was observed
using an N-terminally directed antibody (N-term Ab), an
antibody raised against a-helices 4 and 5 (Ab4) partially
inhibited binding (*40%), although only at the highest
concentration tested (10 nM; Fig. 4a). Similarly the inter-
action between full length Bax and nucleophosmin was
inhibited by the C-terminal antibody (IC50 = 1.43 ± 0.65
nM) but not by either of the other two antibodies (Fig. 4b).
Data obtained from the competition studies gave Hillslope
values = 1 for Bax and nucleophosmin suggesting a
stoichiometric binding of 1:1.
Competition studies were performed using reagents
known to interact with nucleophosmin and Bax. p53, which
Apoptosis (2008) 13:394–403 397
123

binds to nucleophosmin [16], partially inhibited (by 60%)
the Bax peptide-nucleophosmin interaction (IC50 = 65 ±
24 nM; Fig. 5a). In contrast Bcl-2, a known Bax inter-
acting protein [3], had little effect on Bax peptide-
nucleophosmin binding (Fig. 5a). Similar competition
studies were performed using full length Bax protein. In
contrast to the data obtained with the C-terminal Bax
peptide, the interaction between the full-length proteins
was disrupted by both reagents, generating IC50 values of
100 nM and 200 ± 70 nM for Bcl-2 and p53 respectively
(Fig. 5b).
Identification of non-peptide inhibitors of the
Bax-nucleophosmin interaction
In addition to investigating protein–protein interactions at
the molecular level, the proximity assay can also be used as
D BAmret-C
xaBMPNAS
D
B mret-CxaB
AS B A D B AAS
A)
B)
C) E)
D)
MgoL]editpepxaBlanimret-C[
5-6-7-8-9-01-11-
% In
hib
itio
n o
f to
tal s
ign
al
0
02
04
06
08
001
021
041
xaBdetalynitoiB
MgoL]nietorP[
8-9-01-11-21-
RF
U (
x100
0)
0
01
02
03
04
lortnocev-
GgIesuomdetalynitoiB
Fig. 1 Validation of proximity assay to analyse Bax-nucleophosmin
interaction. (a) Diagrammatic representation of the proximity assay
design to analyse the Bax-nucleophosmin interaction. Streptavidin-
coated donor beads (D) attached to a biotinylated Bax C-terminal
peptide and acceptor beads (A) conjugated with a nucleophosmin
monoclonal antibody attached to nucleophosmin. (b) and (c) Com-
petition of the interaction between streptavidin-coated donor beads
and biotin-coated acceptor beads with increasing concentration of
biotinylated Bax C-terminal peptide (0.03–3,000 nM) verified attach-
ment of the peptide to the donor beads. (d) and (e) In the presence of
streptavidin-coated donor beads and acceptor beads conjugated with a
nucleophosmin monoclonal antibody, a concentration dependent
increase in signal was observed with a biotinylated secondary
antibody (0.003–3 nM) verifying successful conjugation of acceptor
beads to the nucleophosmin monoclonal antibody
)nim( emiT042081090654035101521
RF
U (
x100
0)
0
5
01
51
02
52
03
53A) B) C) MPN/xaB mret-C
M goL ]xaB[
6-7-8-9-01-
RF
U (
x100
0)
0
02
04
06
08
001 Mn053 MPN
Mn51 MPN Mn5 MPN
Mn051 MPN
M goL ] xaB[
5-6-7-8-9-01-
RF
U (
x100
0)
0
02
04
06
08
001 Mn51 MPN
Fig. 2 Interaction of Bax C-terminal peptide with nucleophosmin.
(a) Binding of Bax C-terminal peptide (0.3–100 nM) and constant
concentrations of nucleophosmin (5 nM, open triangles; 15 nM, filled
triangles; 150 nM, open circles and 350 nM, filled circles) demon-
strated a concentration dependent increase in signal with a maximal
response using 15 nM nucleophosmin. (b) Bax C-terminal peptide
and nucleophosmin (15 nM) interaction demonstrated a maximal
signal at 100 nM Bax. At higher concentrations of protein the
characteristic hook effect was seen due to quenching of the signal. (c)
Time course of C-terminal Bax peptide (100 nM) and nucleophosmin
(15 nM) binding measured over 4 h demonstrated equilibrium
conditions had been obtained by 90 min. Negative control reactions
contained donor and acceptor beads plus either protein alone. Error
bars represent sem; separate experiments (n = 3)
398 Apoptosis (2008) 13:394–403
123
B)A)
04-PN+-+-+-+-+-
Inpu
t
Pre
clea
r
Dep
lete
d
Was
h
Elu
ate
)aDK02(xaB
noituliD nietorP
3-4-5-6-7-8-
RF
U (
x100
0)
0
01
02
03
04
05
06
07
08
lortnoc ev-
)04-PN( MPN/xaB LF)02 neewT( MPN/xaB LF
)SPAHC( MPN/xaB LF
Fig. 3 Nucleophosmin binds to conformationally active Bax. (a)
Measurement of the interaction between full length Bax and
nucleophosmin in the presence of various detergents: 0.1% CHAPS
(filled triangles), 0.1% Tween 20 (filled circles) or 0.1% NP-40 (open
circles). Binding was only evident in the presence of the non-ionic
detergents NP-40 and Tween 20. Error bars represent sem; separate
experiments (n = 3). (b) Bax in the absence or presence of NP-40
was immunoprecipitated with the conformation specific antibody Bax
6A7 and precipitated samples Western blotted with a pan reactive
antibody BaxNT. Conformationally activated full length Bax was
only detected following treatment with NP-40
B)A)
M goL ]nietorP[
7-8-9-01-11-
% In
hib
itio
n o
f to
tal s
ign
al
0
02
04
06
08
001
021
7-M goL ]nietorP[
8-9-01-11-
% In
hib
itio
n o
f to
tal s
ign
al
0
02
04
06
08
001
021
bA xaB mret-NbA xaB mret-C
4bA xaB
MPN-xaB LFbA xaB mret-NbA xaB mret-C
4bA xaB
MPN-xaB mret-C
Fig. 4 Competition of Bax-nucleophosmin interaction with Bax
antibodies. Competition studies of the interactions between (a) C-
terminal Bax peptide (100 nM) or (b) full length Bax (38 nM) and
nucleophosmin (15 nM) were performed using a N-terminally
directed Bax antibody (N-term Bax Ab; filled circles, solid line), a
C-terminally directed Bax antibody (C-term Bax Ab; filled circles
dashed line) or an antibody raised against Bax a helices 4 and 5 (Bax
Ab4; filled triangles, dotted line) over the concentration range 0.03–
10 nM. Error bars represent sem; separate experiments (n = 3)
A) B)
M goL ]nietorP[
5-6-7-8-9-01-11-
% In
hib
itio
n o
f to
tal s
ign
al
0
02
04
06
08
001
021
041
35pbA xaB mret-C
2-lcBMPN/xaB LF
M goL ]nietorP[
5-6-7-8-9-01-11-
% In
hib
itio
n o
f to
tal s
ign
al
0
02
04
06
08
001
021
35pbA xaB mret-C
2-lcBMPN/xaB mret-C
Fig. 5 Competition of Bax-nucleophosmin interaction with p53 and
Bcl-2. Competition studies of the interactions between (a) C-terminal
Bax peptide (100 nM) or (b) full length Bax (38 nM) and nucleo-
phosmin (15 nM) were performed using a p53 peptide (0.3–
1,000 nM, open circles) or Bcl-2 (0.3–300 nM, filled triangles). C-
terminally directed Bax antibody (C-term Bax Ab; 0.03–10 nM; filled
circles, dashed line) was included as a positive control. Error bars
represent sem; separate experiments (n = 3)
Apoptosis (2008) 13:394–403 399
123

a robust pharmacological screen with a signal to noise ratio
of 135 and a Z0 value of 0.50 ± 0.01 over 3 separate
experiments (Fig. 6a).
To identify modulators of the Bax-nucleophosmin
interaction, we evaluated a small, chemically diverse
library of 800 compounds. This screen yielded 17 com-
pounds that were either potential inhibitors or enhancers of
the Bax peptide-nucleophosmin interaction (defined as
inhibiting or enhancing the signal by [50%). The results
from a representative experiment are shown in Fig. 6b.
Putative positive compounds were further analysed over a
concentration range of 1 pM to 10 lM. One compound,
2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)ethanohydrazide with
a chemical composition of C12H13N3OS and a molecular
weight of 247.3 Daltons (Da), showed inhibition similar to
that observed in the first round of screening with an IC50 of
around 100 nM although the maximal inhibition pro-
duced by this agent was approximately 50% (Fig. 6c).
Specific inhibition of the Bax-nucleophosmin interaction by
2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)ethanohydrazide was
confirmed by titration of the compound over the same
concentration range 1 pM-10 uM in a streptavidin-biotin
detection assay. Inhibition was not observed in this assay
indicating there was no quench of the chemiluminescent
signal and action of the compound was specific to the Bax-
nucleophosmin interction (Fig. 6d).
Bax-nucleophosmin inhibitor blocks UV
irradiation-induced cell death
The utility of 2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)etha-
nohydrazide in modulating cell death was assessed using
an in vitro model in which apoptotic cell death was
induced in HEK293 cells by exposure to UV irradiation
[17]. Twenty-four hours after exposure to 5 s of UV, cell
viability was reduced to 53.85 ± 3.29%. Treatment with
2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)ethanohydrazide
0
20
40
60
80
100
120
140
160
180
TotL
NPML B2 E2 H2 C3 F3 A4 D4 G4 B5 E5 H5 C6 F6 A7 D7 G7 B8 E8 H8 C9 F9A10 D10 G10 B11 E11 H11
Buff R
Compound number
% S
ign
al
B)2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)ethanohydrazide
C12H13N3OS (247.3Da)
Well Number0 10 15 20
RF
U (
x100
0)
0
20
40
60
80
100 C-terminal Bax/NPM signalBackground signal
A)
% In
hib
itio
n o
f to
tal s
ign
al
0
20
40
60
80
100
120
0
20
40
60
80
100
140
120
2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)ethanohydrazidestreptavidin/ biotin
[Protein] Log M
-13 -12 -11 -10 -9 -8 -7 -6 -5 -4
[Protein] Log M
-13 -12 -11 -10 -9 -8 -7 -6 -5 -4
% In
hib
itio
n o
f to
tal s
ign
al
C-terminal Bax /NPM 2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)
D)C)
5
Fig. 6 Identification of a partial inhibitor of the Bax-nucleophosmin
interaction. (a) Assay performance was confirmed by a Z0 value of
0.5 ± 0.01 for the proximity assay and a signal to noise ratio of 135.
Data from a representative plate are shown of experiments performed
in triplicate. (b) Bar chart showing proximity assay results from a
representative plate in the screen of a small chemical library from
which the hit compound was identified. The chemical structure,
composition and molecular weight (247.3 Da) of 2-(5-methyl-2-
phenyl-1,3-thiazol-4-yl)ethanohydrazide is shown. (c) The interaction
between Bax C-terminal peptide (100 nM) and nucleophosmin
(15 nM) was partially inhibited by 2-(5-methyl-2-phenyl-1,3-thia-
zol-4-yl)ethanohydrazide over the concentration range 1 pM-10 lM
(filled triangles, solid line) as compared to vehicle treatment (filled
circles, dashed line). (d) The interaction between streptavidin coated
donor beads and biotin coated acceptor beads was not inhibited by
2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)ethanohydrazide over the con-
centration range 1 pM-10 lM (filled triangles, solid line)
400 Apoptosis (2008) 13:394–403
123

(10 lM) showed a partial but significant (P-value =
0.002, t-test) rescue of cells (by 17.85%) from the UV-
induced reduction in cell viability (Fig. 7).
Discussion
We have recently identified the nuclear chaperone nucle-
ophosmin as a Bax-binding protein and demonstrated that
in animal models of stroke, nucleophosmin moves from the
nucleolus to the cytosol at a time when Bax is conforma-
tionally active, before it translocates to the mitochondria to
induce apoptotic cell death [9]. We have now developed a
proximity assay which has enabled us to characterise this
novel protein–protein interaction at the molecular level.
The proximity assay demonstrated that nucleophosmin
bound to a peptide corresponding to the Bax C-terminal
domain of the native protein. This region is normally
sequestered within the structure of cytosolic Bax and
therefore not available for binding [14]. In addition, no
interaction between full length Bax and nucleophosmin
was observed unless the detergents NP-40 or Tween 20
were included in the reaction buffer. The non-ionic deter-
gent NP-40 has previously been shown to induce a
conformational change in Bax [15] and we confirmed this
by immunoprecipitation with the conformation-sensitive
Bax antibody 6A7. The 6A7 antibody recognises an epi-
tope at the N-terminus of Bax (amino acids 12–24) which
is exposed when the conformation of Bax changes
following treatment with certain detergents or after the
induction of apoptosis [14]. The full length Bax-nucleo-
phosmin interaction was not disrupted by an antibody
raised against the N-terminus of Bax although it was dis-
rupted by an antibody raised against the Bax C-terminus. It
is less clear why very high concentrations of an antibody
recognising a-helices 4 and 5 caused a modest reduction;
steric hindrance is unlikely as antibodies of a similar size
did not produce a similar effect at this concentration.
However recent data have also demonstrated a role for Bax
a-helices 5 and 6 in the interaction of Bax with mito-
chondrial membranes [18]. Only the C-terminal Bax
antibody markedly inhibited the Bax-nucleophosmin
interaction. Taken together our data suggest that nucleo-
phosmin only binds to conformationally altered (activated)
Bax via its C-terminal domain which is normally inacces-
sible in cytosolic Bax.
The importance of helix a9, the C-terminal domain in
the targeting of Bax to the mitochondria following acti-
vation remains controversial. Following an apoptotic
stimulus, Bax undergoes conformational change [19] and it
has been proposed that exposure of the N- and C-terminal
domains of Bax occurs before targeting to the mitochondria
is mediated by the C-terminal domain alone [20]. Several
studies support this theory; GFP-tagged C-terminal trun-
cated Bax expressed in COS cells does not translocate to
mitochondria following apoptosis [19] whereas a Bax
polypeptide devoid of the N-terminal ART domain (Bax-
DART) is constitutively targeted to the mitochondria [21].
Furthermore mutation of Ser184 to Val (S184V), deletion
of the Ser184 (DSer184) altogether in the C-terminal
domain of full length Bax or a 21 amino acid C-terminal
peptide are also sufficient to induce targeting to the mito-
chondria [22]. The interaction of nucleophosmin with
mutated Bax peptide (S184V) in a similar proximity assay
[9] is consistent with the importance of amino acid residues
in the C-terminus region for biological activity.
Recombinant human Bcl-2, a known Bax-binding pro-
tein [23, 24], blocked the interaction between full length
Bax and nucleophosmin but did not affect the binding
between the Bax C-terminal peptide and nucleophosmin.
These data suggest that Bcl-2 binds to a domain which is
absent from the C-terminal Bax peptide but present in the
full length protein such as the BH3 domain or N-terminal
region and are consistent with data in the literature as the
BH3 domain of Bax has been shown to be essential for
binding to Bcl-2 [25]. The role of the Bax BH3 domain (a-
helix 2) in binding to nucleophosmin is still unclear as we
were unable to source a suitable antibody against this
region to perform inhibition studies and it is possible that
the block observed using Bcl-2 was due to steric hindrance
of another binding site. In contrast to the data obtained with
Bcl-2, a peptide fragment of p53, a known nucleophosmin-
rotibihnIelciheVrotibihnIelciheV
Cel
l Su
rviv
al (
%)
0
02
04
06
08
001
021
VUoN VUsceS5
*
* p 200.0=
Fig. 7 Attenuation of UV-induced cell death in HEK293 cells by
2-(5-methyl-2-phenyl-1,3-thiazol-4-yl)ethanohydrazide. HEK293
cells were treated with vehicle or inhibitor (2-(5-methyl-2-phenyl-
1,3-thiazol-4-yl)ethanohydrazide). Cell death was induced using UV
irradiation (30 J/m2, 5 s) and cell viability was measured by MTS
assay 24 h later. Following UV treatment, cell viability was reduced
to 53.85 ± 3.29% in vehicle treated cells. However a 17.85%
increase in survival was observed in those cells treated with inhibitor
compared to vehicle following UV irradiation (P-value = 0.002,
t-test). Survival was expressed as a percentage relative to untreated
cells. Error bars represent sem; separate experiments (n = 3)
Apoptosis (2008) 13:394–403 401
123

binding protein [16], disrupted the interaction between
nucleophosmin and both forms of Bax. The p53 binding
region is located in the C-terminal half of nucleophosmin
[16, 26] and recent data produced using chemical cross-
linking, proteolytic digestion and mass spectrometry-based
peptide mapping have narrowed this down to residues 249–
262 [27]. This p53-binding region of nucleophosmin
overlaps with the immunizing peptide (residues 226–294)
used to raise the monoclonal antibody [28] which we used
to attach nucleophosmin to the acceptor beads in our
proximity assay. Therefore it is likely that the p53 peptide
competed for nucleophosmin binding at the C-terminal
domain attached to the acceptor beads rather than the Bax-
nucleophosmin protein–protein interaction. As a proximity
assay signal was generated with Bax when nucleophosmin
was attached to the acceptor beads by its C-terminal
domain, our data also implicate the N-terminal half of
nucleophosmin as important for Bax binding. This region
contains the signals required for nucleophosmin localiza-
tion to the nucleolus and sequences essential for its
chaperone activity [29, 30] consistent with our identifica-
tion of nucleophosmin as a Bax chaperone which
translocates from the nucleolus to the cytoplasm following
apoptotic stimuli [9, 31].
Recently identified chaperones that regulate the target-
ing of Bax to the mitochondria bind to either the N- or C-
terminus and these include humanin, 14-3-3, Ku70 and
clusterin [32–35]. As with nucleophosmin, clusterin inter-
acts with a conformation-altered Bax, but to prevent
apoptotic activity [35]. Data published from this lab sug-
gests nucleophosmin is a regulator of Bax mediated cell
death both in vitro and in vivo [9]. Following stauro-
sporine treatment of SH-SY5Y cells, nucleophosmin was
localised in the cytosol prior to Bax mitochondrial trans-
location. In vivo, focal cerebral ischaemia led to
nucleophosmin translocation and Bax conformational
change [9]. Nucleophosmin acts as a chaperone to regulate
the stability and transcriptional activation of ARF and p53
in another apoptotic pathway [16, 36–38]. Binding of ARF
by nucleophosmin activates mdm2 causing p53 to be down
regulated resulting in cell proliferation [38]. However in
the absence of nucleophosmin, ARF binds to mdm2 caus-
ing p53 accumulation and cell cycle arrest via downstream
targets such as BH3 only proteins and Bax [39].
The proximity assay technology used to characterise the
molecular interplay between Bax and nucleophosmin, can
also be used as a high-throughput pharmacological screen for
inhibitors/modulators of this interaction. The reliability of this
384-well assay is reflected by a signal to noise ratio of 135, an
inter-plate variance of 14.8% (across days) and a Z0 value of
0.50 which puts this assay at the cusp between marginal and
excellent as a high-throughput screen [40]. A manual screen of
a small chemical library (800 compounds) identified 17
potential compounds that modulated Bax peptide-nucleo-
phosmin binding either by inhibiting or enhancing the signal
by 50%. One hit compound, 2-(5-methyl-2-phenyl-
1,3-thiazol-4-yl)ethanohydrazide, produced a concentration-
dependent partial inhibition of the Bax-nucleophosmin
interaction with an IC50 value of 100 nM. The success rate is
supportive of extending the analysis to the major chemical
libraries of the pharmaceutical industry which contain more
than 250,000 diverse compounds. Moreover, this compound
partially reversed the cell death induced by UV treatment of
HEK293 cells. We previously demonstrated that RNAi
knockdown of nucleophosmin expression protected against
staurosporine-induced apoptosis and that the level of protec-
tion was proportionate to the reduction in expression levels
[9]. Together these data suggest that the Bax-nucleophosmin
interaction may be involved in the response to different
apoptotic stimuli. Thus the Bax-nucleophosmin protein–pro-
tein interaction may represent a novel therapeutic target for the
numerous disease states associated with dysregulated
apoptosis.
In conclusion, the present report provides compelling
new data in three aspects of the interaction between Bax
and nucleophosmin:
(1) The kinetics of the binding of Bax and nucleophos-
min are indicative of a competitive and specific
biological mechanism.
(2) The binding of full length Bax to nucleophosmin
requires the conformational change which Bax under-
goes early in the apoptotic cascade, exposing the
C-terminal region.
(3) The proximity assay described provides a drug
discovery platform for the identification of low
molecular weight modulators of apoptosis.
References
1. Gross A, Jockel J, Wei MC, Korsmeyer SJ (1998) Enforced
dimerization of BAX results in its translocation, mitochondrial
dysfunction and apoptosis. EMBO J 17:3878–3885
2. Hsu YT, Wolter KG, Youle RJ (1997) Cytosol-to-membrane
redistribution of Bax and Bcl-X(L) during apoptosis. Proc Natl
Acad Sci USA 94:3668–3672
3. Oltvai ZN, Milliman CL, Korsmeyer SJ (1993) Bcl-2 heterodi-
merizes in vivo with a conserved homolog, Bax, that accelerates
programmed cell death. Cell 74:609–619
4. Boise LH, Gonzalez-Garcia M, Postema CE et al (1993) Bcl-x, a
Bcl-2-related gene that functions as a dominant regulator of
apoptotic cell death. Cell 74:597–608
5. Sedlak TW, Oltvai ZN, Yang E et al (1995) Multiple Bcl-2
family members demonstrate selective dimerizations with Bax.
Proc Natl Acad Sci USA 92:7834–7838
6. Desagher S, Osen-Sand A, Nichols A et al (1999) Bid-induced
conformational change of Bax is responsible for mitochondrial
cytochrome c release during apoptosis. J Cell Biol 144:891–901
402 Apoptosis (2008) 13:394–403
123

7. Eskes R, Desagher S, Antonsson B, Martinou JC (2000) Bid
induces the oligomerization and insertion of Bax into the outer
mitochondrial membrane. Mol Cell Biol 20:929–935
8. Li P, Nijhawan D, Budihardjo I et al (1997) Cytochrome c and
dATP-dependent formation of Apaf-1/caspase-9 complex initi-
ates an apoptotic protease cascade. Cell 91:479–489
9. Kerr LE, Birse-Archbold JL, Short DM et al (2007) Nucleo-
phosmin is a novel Bax chaperone that regulates apoptotic cell
death. Oncogene 26(18):2554–2562
10. Feuerstein N, Spiegel S, Mond JJ (1988) The nuclear matrix
protein, numatrin (B23), is associated with growth factor-induced
mitogenesis in Swiss 3T3 fibroblasts and with T lymphocyte
proliferation stimulated by lectins and anti-T cell antigen receptor
antibody. J Cell Biol 107:1629–1642
11. Okuwaki M, Matsumoto K, Tsujimoto M, Nagata K (2001)
Function of nucleophosmin/B23, a nucleolar acidic protein, as a
histone chaperone. FEBS Lett 506:272–276
12. Swaminathan V, Kishore AH, Febitha KK, Kundu TK (2005)
Human histone chaperone nucleophosmin enhances acetylation-
dependent chromatin transcription. Mol Cell Biol 25:7534–7545
13. Ullman EF, Kirakossian H, Singh S et al (1994) Luminescent
oxygen channeling immunoassay: measurement of particle
binding kinetics by chemiluminescence. Proc Natl Acad Sci USA
91:5426–5430
14. Suzuki M, Youle RJ, Tjandra N (2000) Structure of Bax: core-
gulation of dimer formation and intracellular localization. Cell
103:645–654
15. Hsu YT, Youle RJ (1998) Bax in murine thymus is a soluble
monomeric protein that displays differential detergent-induced
conformations. J Biol Chem 273:10777–10783
16. Colombo E, Marine JC, Danovi D, Falini B, Pelicci PG (2002)
Nucleophosmin regulates the stability and transcriptional activity
of p53. Nat Cell Biol 4:529–533
17. Fan Y, Wu D, Jin L, Yin Z (2005) Human glutamylcysteine
synthetase protects HEK293 cells against UV-induced cell death
through inhibition of c-Jun NH2-terminal kinase. Cell Biol Int 29:
695–702
18. Garcia-Saez AJ, Coraiola M, Serra MD, Mingarro I, Muller P,
Salgado J (2006) Peptides corresponding to helices 5 and 6 of
Bax can independently form large lipid pores. FEBS J 273:971–
981
19. Wolter KG, Hsu YT, Smith CL, Nechushtan A, Xi XG, Youle RJ
(1997) Movement of Bax from the cytosol to mitochondria during
apoptosis. J Cell Biol 139:1281–1292
20. Schinzel A, Kaufmann T, Schuler M, Martinalbo J, Grubb D,
Borner C (2004) Conformational control of Bax localization and
apoptotic activity by Pro168. J Cell Biol 164:1021–1032
21. Goping IS, Gross A, Lavoie JN et al (1998) Regulated targeting
of BAX to mitochondria. J Cell Biol 143:207–215
22. Nechushtan A, Smith CL, Hsu YT, Youle RJ (1999) Conforma-
tion of the Bax C-terminus regulates subcellular location and cell
death. EMBO J 18:2330–2341
23. Antonsson B, Conti F, Ciavatta A et al (1997) Inhibition of Bax
channel-forming activity by Bcl-2. Science 277:370–372
24. Otter I, Conus S, Ravn U et al (1998) The binding properties and
biological activities of Bcl-2 and Bax in cells exposed to apop-
totic stimuli. J Biol Chem 273:6110–6120
25. Zha H, Aime-Sempe C, Sato T, Reed JC (1996) Proapoptotic
protein Bax heterodimerizes with Bcl-2 and homodimerizes with
Bax via a novel domain (BH3) distinct from BH1 and BH2. J Biol
Chem 271:7440–7444
26. Li J, Zhang X, Sejas DP, Pang Q (2005) Negative regulation of
p53 by nucleophosmin antagonizes stress-induced apoptosis in
human normal and malignant hematopoietic cells. Leuk Res 29:
1415–1423
27. Lambert B, Buckle M (2006) Characterisation of the interface
between nucleophosmin (NPM) and p53: Potential role in p53
stabilisation. FEBS Lett 580:345–350
28. Ochs R, Lischwe M, O’Leary P, Busch H (1983) Localization of
nucleolar phosphoproteins B23 and C23 during mitosis. Exp Cell
Res 146:139–149
29. Chan HJ, Weng JJ, Yung BY (2005) Nucleophosmin/B23-bind-
ing peptide inhibits tumor growth and up-regulates transcriptional
activity of p53. Biochem Biophys Res Commun 333:396–403
30. Hingorani K, Szebeni A, Olson MO (2000) Mapping the func-
tional domains of nucleolar protein B23. J Biol Chem 275:
24451–24457
31. Lu YY, Lam CY, Yung BY (1996) Decreased accumulation and
dephosphorylation of the mitosis-specific form of nucleophos-
min/B23 in staurosporine-induced chromosome decondensation.
Biochem J 317(Pt 1):321–327
32. Guo B, Zhai DY, Cabezas E et al (2003) Humanin peptide sup-
presses apoptosis by interfering with Bax activation. Nature 423:
456–461
33. Nomura M, Shimizu S, Sugiyama T et al (2003) 14-3-3 interacts
directly with and negatively regulates pro-apoptotic bax. J Biol
Chem 278:2058–2065
34. Sawada M, Sun WY, Hayes P, Leskov K, Boothman DA, Ma-
tsuyama S (2003) Ku70 suppresses the apoptotic translocation of
Bax to mitochondria. Nat Cell Biol 5:320–329
35. Zhang HL, Kim JK, Edwards CA, Xu ZH, Taichman R, Wang
CY (2005) Clusterin inhibits apoptosis by interacting with acti-
vated Bax. Nat Cell Biol 7:909–915
36. Bertwistle D, Sugimoto M, Sherr CJ (2004) Physical and func-
tional interactions of the Arf tumor suppressor protein with
nucleophosmin/B23. Mol Cell Biol 24:985–996
37. Colombo E, Bonetti P, Denchi EL et al (2005) Nucleophosmin is
required for DNA integrity and p19(Arf) protein stability. Mol
Cell Biol 25:8874–8886
38. Korgaonkar C, Hagen J, Tompkins V et al (2005) Nucleophos-
min (B23) targets ARF to nucleoli and inhibits its function. Mol
Cell Biol 25:1258–1271
39. Vousden KH, Lu X (2002) Live or let die: The cell’s response to
p53. Nat Rev Cancer 2:594–604
40. Zhang JH, Chung TDY, Oldenburg KR (1999) A simple statis-
tical parameter for use in evaluation and validation of high
throughput screening assays. J Biomol Screen 4:67–73
Apoptosis (2008) 13:394–403 403
123

![Synthetic Bax-Anti Bcl2 combination module actuated by ......Bcl 2 levels and elevating Bax levels [14–18]. It indicates that the combination of Bax protein and anti-Bcl 2 molecule](https://static.fdocuments.us/doc/165x107/6113a58ae4fe0d22082a45c6/synthetic-bax-anti-bcl2-combination-module-actuated-by-bcl-2-levels-and.jpg)
















