
CellWallGrowth Bacillus Cytoplasmic Radioactivity After ... · DAP on unwashed Whatman no. 4 paper...
11
JOURNAL OF BACTERIOLOGY, Sept. 1975, p. 1197-1207 Copyright © 1975 American Society for Microbiology Vol. 123, No. 3 Printed in U.S.A. Cell Wall Growth of Bacillus megaterium: Cytoplasmic Radioactivity After Pulse-Labeling with Tritiated Diaminopimelic Acid CHANTAL DE CHASTELLIER, CLAUDE FREHEL, AND ANTOINETTE RYTER* Unite de Microscopie Electronique, Departement de Biologie Moleculaire, Institut Pasteur, 75015 Paris, France Received for publication 3 April 1975 Study of the cell wall growth in Bacillus megaterium by pulse-labeling a DAP- Lys- mutant with tritiated diaminopimelic acid (DAP) had revealed the presence of intracytoplasmic radioactivity. The nature of this radioactivity was studied on one hand by autoradiographic analysis of bacteria treated in different ways and on the other hand by chromatography of the radioactive compounds extracted with boiling water. It is shown that cytoplasmic radioactivity corre- sponds neither to free DAP nor to DAP metabolized into lysine, but to murein precursors. Autoradiographic analysis of bacteria in which all murein precursors were removed gives exactly the same cell wall growth pattern as the one previously obtained for untreated bacteria. It can be concluded that, in B. megaterium, cell wall elongation occurs by diffuse intercalation of newly synthesized murein along the cylindrical part of the cell wall and that only cross wall formation occurs in a precise growth zone. In a previous study (4), the cell wall growth pattern was determined in Bacillus megaterium by high-resolution autoradiography of bacteria pulse-labeled with tritiated diaminopimelic acid (DAP). It was concluded that cell wall elongation occurs by diffuse intercalation of newly synthesized murein into the cylindrical part of the cell wall, the cross wall alone representing a precise growth zone. Autoradio- graphs of thin sections revealed however that about 50% of the total radioactivity was located in the cytoplasm. This cytoplasmic radioactiv- ity raised some technical difficulties in the analysis of the silver grain distribution along the cell wall because it can interfere with that of the cell wall. But these problems could be nearly completely overcome by determining the resolution of our autoradiographs (i.e., distance from the cell wall at which silver grains could still be considered as representing DAP incorpo- rated into the cell wall) on thin sections of isolated cell walls. However the nature of this cytoplasmic radioactivity remained unknown. It could be attributed either to free DAP mole- cules trapped in the cell during fixation or, eventually, to DAP metabolized into lysine. Although this seemed improbable since the strain used is a DAP- Lys- double mutant, it could not be completely excluded. Finally, this radioactivity could be attributed to different precursors of the murein biosynthetic pathway. Studies made on this biochemical pathway (1, 5-7) have shown that the first step corresponds to the formation of uridine 5'-diphosphate-Mur- NAc-pentapeptide and uridine 5'-diphosphate- GlNAc in the cytoplasm. These two precursors are then transferred to, a undecaprenol phos- phate carrier of the cytoplasmic membrane thereby forming the dissaccharide pentapeptide GlNAc-MurNAc-pentapeptide. Finally, the complete disaccharide pentapeptide, linked to the lipid intermediate, is integrated into the expanding cell wall with release of the terminal amino acid, D-alanine. The nature and the distribution of the cyto- plasmic radioactivity was therefore studied in different conditions. Autoradiographs of pulse- labeled bacteria were analyzed after washing of cells, after a chase, or after extraction of radio- active compounds not incorporated into the cell wall. The nature of the latter was also deter- mined by chromatography after extraction with boiling water. MATERIALS AND METHODS Growth and labeling conditions. The strain and the experimental conditions are identical to those described in our previous paper (4). The DAP- Lys- mutant of B. megaterium MB was grown at 30 C in Salton synthetic medium (2) supplemented with 1% yeast extract, 0.2% glucose, and 50 ,ug of DAP per ml. When the culture counted 108 cells/ml, it was spun down and the pellet was resuspended in the same 1197 on April 20, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript of CellWallGrowth Bacillus Cytoplasmic Radioactivity After ... · DAP on unwashed Whatman no. 4 paper...

JOURNAL OF BACTERIOLOGY, Sept. 1975, p. 1197-1207Copyright © 1975 American Society for Microbiology
Vol. 123, No. 3Printed in U.S.A.
Cell Wall Growth of Bacillus megaterium: CytoplasmicRadioactivity After Pulse-Labeling with Tritiated
Diaminopimelic AcidCHANTAL DE CHASTELLIER, CLAUDE FREHEL, AND ANTOINETTE RYTER*
Unite de Microscopie Electronique, Departement de Biologie Moleculaire, Institut Pasteur, 75015 Paris, France
Received for publication 3 April 1975
Study of the cell wall growth in Bacillus megaterium by pulse-labeling a DAP-Lys- mutant with tritiated diaminopimelic acid (DAP) had revealed thepresence of intracytoplasmic radioactivity. The nature of this radioactivity was
studied on one hand by autoradiographic analysis of bacteria treated in differentways and on the other hand by chromatography of the radioactive compoundsextracted with boiling water. It is shown that cytoplasmic radioactivity corre-sponds neither to free DAP nor to DAP metabolized into lysine, but to mureinprecursors. Autoradiographic analysis of bacteria in which all murein precursorswere removed gives exactly the same cell wall growth pattern as the onepreviously obtained for untreated bacteria. It can be concluded that, in B.megaterium, cell wall elongation occurs by diffuse intercalation of newlysynthesized murein along the cylindrical part of the cell wall and that only cross
wall formation occurs in a precise growth zone.
In a previous study (4), the cell wall growthpattern was determined in Bacillus megateriumby high-resolution autoradiography of bacteriapulse-labeled with tritiated diaminopimelicacid (DAP). It was concluded that cell wallelongation occurs by diffuse intercalation ofnewly synthesized murein into the cylindricalpart of the cell wall, the cross wall alonerepresenting a precise growth zone. Autoradio-graphs of thin sections revealed however thatabout 50% of the total radioactivity was locatedin the cytoplasm. This cytoplasmic radioactiv-ity raised some technical difficulties in theanalysis of the silver grain distribution alongthe cell wall because it can interfere with that ofthe cell wall. But these problems could benearly completely overcome by determining theresolution of our autoradiographs (i.e., distancefrom the cell wall at which silver grains couldstill be considered as representing DAP incorpo-rated into the cell wall) on thin sections ofisolated cell walls. However the nature of thiscytoplasmic radioactivity remained unknown.It could be attributed either to free DAP mole-cules trapped in the cell during fixation or,eventually, to DAP metabolized into lysine.Although this seemed improbable since thestrain used is a DAP- Lys- double mutant, itcould not be completely excluded. Finally, thisradioactivity could be attributed to differentprecursors of the murein biosynthetic pathway.
Studies made on this biochemical pathway (1,5-7) have shown that the first step correspondsto the formation of uridine 5'-diphosphate-Mur-NAc-pentapeptide and uridine 5'-diphosphate-GlNAc in the cytoplasm. These two precursorsare then transferred to, a undecaprenol phos-phate carrier of the cytoplasmic membranethereby forming the dissaccharide pentapeptideGlNAc-MurNAc-pentapeptide. Finally, thecomplete disaccharide pentapeptide, linked tothe lipid intermediate, is integrated into theexpanding cell wall with release of the terminalamino acid, D-alanine.The nature and the distribution of the cyto-
plasmic radioactivity was therefore studied indifferent conditions. Autoradiographs of pulse-labeled bacteria were analyzed after washing ofcells, after a chase, or after extraction of radio-active compounds not incorporated into the cellwall. The nature of the latter was also deter-mined by chromatography after extraction withboiling water.
MATERIALS AND METHODSGrowth and labeling conditions. The strain and
the experimental conditions are identical to thosedescribed in our previous paper (4). The DAP- Lys-mutant of B. megaterium MB was grown at 30 C inSalton synthetic medium (2) supplemented with 1%yeast extract, 0.2% glucose, and 50 ,ug of DAP per ml.When the culture counted 108 cells/ml, it was spundown and the pellet was resuspended in the same
1197
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

1198 DE CHASTELLIER, FREHEL, AND RYTER
volume of fresh medium devoid of DAP. Five milli-liters of this suspension were labeled for 2 min with0.12 mCi of 3H-labeled DAP per ml (specific activity25 Ci/mM, Service des Molecules Marquees, C.E.A.,France). The concentration of radioactive DAP wastherefore 0.9 ug/ml. The reaction was stopped with250 Asg of unlabeled DAP per ml. Bacteria wereimmediately prefixed with osmium tetroxide (0.2%)and prepared for electron microscopy with or withoutwashing.Washing of bacteria after prefixation. The pulse-
labeled bacteria were prefixed with osmium tetroxide(0.2%) and immediately centrifuged. One half of thecells were prepared for high-resolution autoradiogra-phy; the other half was centrifuged and washed twicein complex DAP-free culture medium before beingfixed, dehydrated, and embedded.
Pulse-chase experiments. Bacteria were labeled asusual. Then 500 jg of cold DAP per ml was added tothe 5-ml aliquots of labeled bacteria. After 3 and 9min, respectively, of incubation at 30 C, cells wereprefixed as usual and prepared for electron micros-copy.
Extraction procedures. (i) Extraction of bacteriawith boiling water. Bacteria were labeled, prefixed,and fixed as usual. They were partially dehydratedwith acetone (up to 90% acetone) and treated with 2ml of boiling water for 1 min to extract radioactivecompounds not incorporated in the cell wall. Thesuspension was cooled in ice and centrifuged. Thepellet containing bacteria and cellular debris wasdehydrated again and embedded. The supernatantcontaining the extracted radioactive compounds wasused for chromatography.
(ii) Extraction of unfixed bacteria with trichlo-roacetic acid. A 5-ml aliquot of bacteria labeled inthe usual conditions was added to 5 ml of 10%trichloroacetic acid before prefixation and kept at 0 Cfor 10 min to extract acid-soluble radioactive com-pounds. Bacteria were then centrifuged and washedthree times with 5 ml of 5% trichloroacetic acid. Thebacterial pellet was fixed and embedded.Chromatography. The supernatant of bacteria
treated with boiling water was concentrated to aboutone-fourth its volume under a stream of nitrogen. Theextracted radioactive compounds were identified byradloautography. For this purpose, ascending one-dimensional chromatography was done with aliquotsof the extract in parallel to aliquots of 3H-labeledDAP on unwashed Whatman no. 4 paper or oncellulose thin-layered chromatography plates 0.1 mmthick, using phenol-water or butanol-propionic acid-water as solvent (3). 8H-Labeled DAP (10 MA) or ex-tract were deposited on cellulose thin-layer chro-matography plates (4 x 101 counts/min in a Packardmodel 3003 Tri-Carb liquid scintillation spectrometerwhich represents 0.4 MCi) and 100 Ml on paper (4 x 10'counts/min which represents 4 ,uCi). Autoradiogramswere prepared by exposing the air-dried chromato-grams to Kodak nonscreen X-ray films for 3 weeks.Identity of the radioactive compounds was based onninhydrin spray tests after co-chromatography withknown compounds. In this case, 2 Mg of nonradioac-
tive DAP or 2.5 Mg of nonradioactive lysine wereadded as tracer to two 40-ul aliquots (1.6 x 106counts/min which represents 1.6 MCi) of the concen-trated extract. The aliquots were concentrated undera stream of nitrogen until complete evaporation. Eachresidue was taken up separately in a drop of 20%alcohol and used for two-dimensional ascending chro-matography on thin-layer chromatography plates,first in phenol-water and then in butanol-propionicacid-water (3). Autoradiograms were prepared byexposing the two air-dried chromatograms to Kodaknonscreen X-ray films for 2 weeks. The chromato-grams were then sprayed with ninhydrin and devel-oped in an air oven at 100 C for 2 to 3 min.
Electron microscopy and autoradiography. Fixa-tion, dehydration, embedding, and autoradiographytechniques were described in detail in our previouspaper (4).
Analysis of autoradiographs. Silver grain distri-butions were determined on electron micrographsenlarged 30,000 times and are represented in histo-grams.. The distance between the center of silvergrains and the chosen reference marker (median lineof the cell wall or cross wall) was measured with aruler. All histograms concerning the analysis of cyto-plasmic radioactivity were prepared in distance stepsof 33.3 nm. They were normalized to 100, the columncontaining the highest count of silver grains beingconsidered as reference. To determine the distributionof cell wall radioactivity after extraction procedures,bacteria were divided according to their stage into thefour classes previously described (4). Histograms wereprepared in distance steps of 83.3 nm and the numberof silver grains was expressed as percentages of thetotal number of cell wall silver grains.
RESULTSAutoradiography of thin sections of pulse-
labeled bacteria. As already described in ourprevious paper (4), about 50% of the silvergrains are located in the cytoplasm. Theirdistribution was determined by measuring thedistance between all silver grains located insideand outside bacteria and the median line of thecell wall (the median line was at an equaldistance from the inner and outer edges of thecell wall). Silver grains concentrated along thecross wall of dividing bacteria or very close to itwere neglected because they are a source ofconfusion. A normalized histogram was plottedin distance steps of 33.3 nm as described and isrepresented in Fig. 1. The silver grain distribu-tion is not symmetrical on each side of the cellwall as opposed to the one obtained with thinsections of isolated cell walls (4; Fig. 2). Acomparison of these two figures shows that theextracellular silver grain distributions wereidentical and can be perfectly superposed. Thismeans that all the extracellular silver grainsfound on thin sections of whole bacteria were
J. BACTERIOL.
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

CELL WALL GROWTH IN B. MEGATERIUM 1199
100
Ia
9.752I
500 333 166 0 166 333 500distance (in nm)
FIG. 1. Normalized histograms prepared in dis-tance steps of 33.3 nm, representing the distributionof silver grains inside and outside bacteria withrespect to the median line of the cell wall. Longitudi-nal thin sections of exponentially growing bacterialabeled for 2 min with 3H-labeled DAP were analyzed.(Solid line) Unwashed bacteria after 32 days ofexposition. 1,054 silver grains were considered. Theobserved asymmetry indicates that a large amount ofthe radioactivity is located elsewhere than in the cellwall. (Dashed line) Bacteria were washed twice afterprefixation with DAP-free culture medium (42 days ofexposition). 796 silver grains were considered. Theasymmetry of the histogram indicates that intracellu-lar radioactivity is still present.
issued from the disintegration of 3H-labeledDAP located in the cell wall. In contrast,intracellular silver grain distributions were verydifferent. There were indeed many more silvergrains in the cytoplasm of whole bacteria,indicating that in this case an importantamount of silver grains represents radioactivecompounds located elsewhere than in the cellwall.As it appears in Fig. 1, the distribution of
these intracellular silver grains decreased regu-larly from the cell wall to the center of the cell(longitudinal axis of symmetry). The presenceof such a gradient suggests that the intracyto-plasmic radioactivity was not randomly distrib-uted in the bacterium but was much moreconcentrated at its periphery. This was con-firmed by the silver grain distribution deter-mined with respect to the cross wall (Fig. 3). Inthis case, the histogram was perfectly symmet-
c10
7 Outside baceri inide bacteria
50-
_1
25
0500 333 166 0 166 333 500
distance (in nm)
FIG. 2. Normalized histogram of the distributionof silver grains with respect to the median line of thecell wall obtained on thin sections of isolated cellwalls (distance step 33.3 nm). 1,450 silver grains werecounted.
100 ,
75
25
0500 333 166 0 166 333 500
distance (in nm)
FIG. 3. Normalized histogram representing thedistribution of silver grains with respect to the crosswall of labeled dividing bacteria not subject to anyother treatment. 740 silver grains were considered.The histogram is quite symmetrical with respect tothe reference line.
VOL. 123, 1975
0
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

1200 DE CHASTELLIER, FREHEL, AND RYTER
rical as expected. Silver grain frequency de-creased very quickly during the first 100 nm andthen reached a plateau at about 200 nm whichwas due to random distribution of radioactivecompounds lengthwise in the cell.
Effect of washing on the intracytoplasmicradioactivity. Pulse-labeled bacteria werewashed twice at room temperature with 3H-labeled DAP-free culture medium after theirprefixation with osmium tetroxide. A randomobservation of autoradiographs with the elec-tron microscope did not reveal any visiblechange in the distribution of silver grains (Fig.4b, c, and d) as compared to untreated bacteria(Fig. 4a). The normalized histogram represent-ing a quantitative analysis of their distribution(Fig. 1) still presented a strong asymmetry withrespect to the cell wall. However there was aslight decrease of the intracytoplasmic radioac-tivity.Autoradiography of pulse-chased bacteria.
Bacteria labeled for 2 min with tritiated DAPreceived a large amount of cold DAP and werefixed 3 or 9 min later. Figure 5 shows that after a3-min chase, silver grains were already muchmore precisely located along the cell wall. Thestatistical analysis of these autoradiographsconfirms this impression (Fig. 6). Indeed thesilver grain distribution became perfectly sym-metrical on each side of the cell wall withrespect to its median line, as was the case forthin sections of isolated cell walls (Fig. 2).
Effect of extraction on cytoplasmic radio-activity. (i) Bacteria treated with boilingwater. The purpose of this treatment was toextract radioactive compounds not incorporatedinto the cell wall and identify their nature bychromatography. To be in conditions as identi-cal as possible to previous morphological experi-ments, labeled bacteria were fixed and nearlycompletely dehydrated with acetone beforebeing treated with boiling water for 1 min.Indeed, during fixation and dehydration ofbacteria for electron microscopy, a certainamount of radioactive compounds were lost.Controls were done to check how many countswere extracted during these processes. It seemsthat prefixation and fixation extract 30% anddehydration 9% of the total number of countsfound before prefixation.
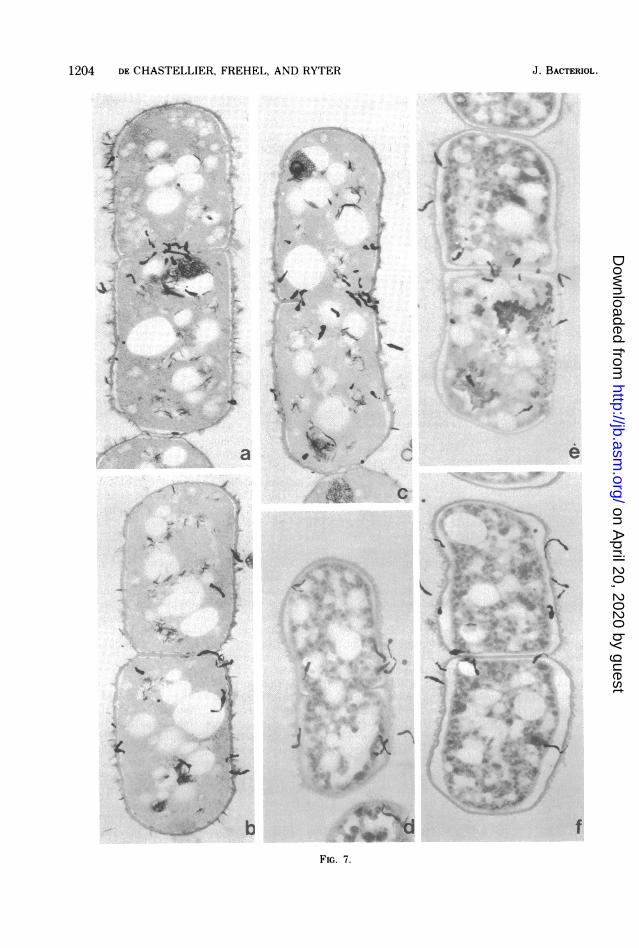
Figure 7 (a, b, and c) shows that the extrac-tion with boiling water somewhat altered thecells morphologically, especially the capsularmaterial exterior to the cell wall and the meso-somes. Small gaps could be seen between thecell wall and the ctyoplasmic membrane, butthe cell wall itself did not appear altered and
seemed to be labeled as untreated cells. As forthe number of silver grains in the cytoplasm, itclearly decreased after the extraction as com-pared to untreated bacteria.The statistical distribution of silver grains on
thin sections of these bacteria (Fig. 8) confirmsa clear decrease of the cytoplasmic radioactivityas compared to untreated bacteria (Fig. 1).However, the histogram was not as perfectlysymmetrical on each side of the cell wall (half-distance inside = 100 nm; half-distance outside= 75 nm) as the one obtained with thin sectionsof isolated cell walls (Fig. 2) or in the pulse-chase experiment (Fig. 6). The relative amountof silver grains was a little higher inside the cellthan outside. The decrease of intracytoplasmicsilver grain frequency was very sharp in the first133 nm. It then became weak but constant until500 nm without reaching a plateau.
(ii) Bacteria treated with 5% trichloroace-tic acid. Thin sections of bacteria treated with5% trichloroacetic acid before their fixation areshown in Fig. 7 (d, e, f). As expected, theextraction procedure dramatically affected themorphology of bacteria. The rod shape wassomewhat altered; cells appeared rounder andshorter and the wall was wavy, which probablymeans that some modifications occurred in thecell wall. Also the capsular material on theexterior side of the cell wall was altered as wellas inner structures of the cell. Finally, impor-tant gaps could be seen between the cell walland the cytoplasmic membrane, as if the cyto-plasm had "shrunk" during the extraction pro-cedure. In spite of these alterations it was clearthat silver grains are evenly distributed alongthe cell wall, with a concentration on the crosswall during its formation (Fig. 7d, e, and f) andthat the cytoplasm is barely labeled. The histo-gram (Fig. 8) shows that the silver grain distri-bution was quite symmetrical (half-distanceinside = 83 nm; half-distance outside = 75 nm),and in any case more symmetrical than that ofbacteria treated with boiling water. Small dif-ferences in distribution patterns of extracellularsilver grains were probably due to the wavyaspect of the cell wall of trichloroacetic acid-treated bacteria which slightly affected autora-diographic resolution. This symmetry meansthat most of the radioactivity was located in thecell wall.Chromatography of compounds extracted
with boiling water. Chromatography was doneto identify the nature of the radioactive com-pounds not incorporated into the cell wall andextracted with boiling water.By one-dimensional chromatography, it was
J. BACTERIOL.
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

S0 4
i
.o.-
/If .ef0 ",.f1`
i ._ "I';
- U
*
_at
4.~~~~~~~~* 2_ ( \~~~*_. |w~~2
¾
Vt4..
-^
b9'
iI
N
b
.¼
ti
4
1
_
i1.3*_
"I
*$ s .*4 .I"..
4ie4.
'tF
?,
.1
4,
I
IV*,
dFIG. 4. Autoradiographs of longitudinal thin sections of exponentially growing bacteria labeled for 2 min
with 3H-labeled DAP. (a) Untreated bacteria (32 days of exposition); (b, c, d) bacteria washed twice with
DAP-free complex medium after prefixation (42 days of exposition). x25,000.1201
./
C4.
..d'........I. tA
E
* ie."
I
t la Air
* %a%*;d a
p
r1S0rC -
a
I ..-, -.o
I._
-il
..
..-fl
J
i1%
I
-4.I
ii
Jr
c
IF
.1
. f ...
.9 I
4.rt. 1
\06,16 4 A&.
. I.
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

N~~~~~~~~~~~F-4 so
St!
a....,,,,..C
b -
e
fFIG. 5. Autoradiographs of longitudinal thin sections of exponentially growing bacteria labeled for 2 min
with 'H-labeled DAP which was chased with cold DAP added in excess, for 3 (a, b, c) or 9 min (d, e, f) (28 daysof exposition). x 18,000.
1202
tk *4
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

CELL WALL GROWTH IN B. MEGATERIUM 1203
100
c outside bacteria insidi bacteria
750
25
0
500 333 166 0 166 333 500
distance (in nm)
FIG. 6. Normalized histograms representing the
distribution of silver grains inside and outside pulse-
chased bacteria with respect to the median line of the
cell wall. (Solid line) 3-min chase; 859 silver grainsconsidered; (Dashed line) 9-min chase; 863 silvergrains considered. The symmetry of these histogramsindicates that the whole radioactivity is located in thecell wall.
difficult to determine the nature of the radioac-tive compounds, in particular to detect thepresence of free 3H-labeled DAP molecules. Wetherefore tried to better separate the radioactivecompounds by two dimensional chromatogra-phy done as described. After 2 weeks of exposi-tion, spots on radioautograms were compared tothose of DAP and lysine revealed by ninhydrinon the chromatograms. As shown in Fig. 9, spotsdid not correspond. Therefore there were no free3H-labeled DAP or [3H ]lysine molecules in theextract.Control of cell wall growth pattern in
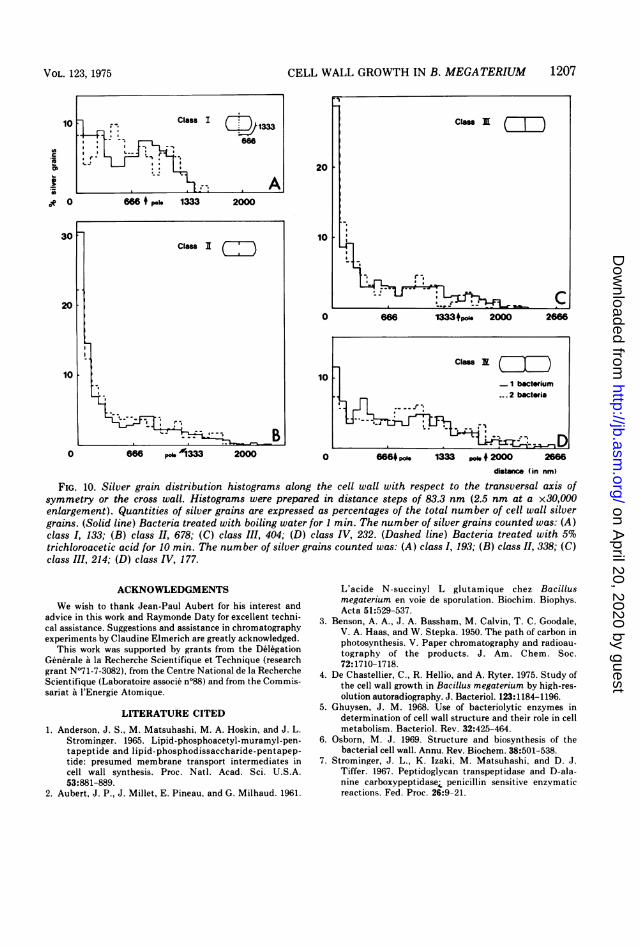
pulse-labeled bacteria treated with boilingwater or trichloroacetic acid. Silver graindistributions with respect to the median line ofthe cell wall, obtained on bacteria treated withboiling water or trichloroacetic acid, showedthat most of the radioactivity still present in thebacteria corresponded to DAP incorporated intothe cell wall. In these conditions the correctnessof the growth pattern deduced from autoradio-graphs of unwashed bacteria can be verified.For this purpose, the distribution of silvergrains along the cell wall was established withrespect to the transversal axis of symmetry of
the cells, or the cross wall when it exists, as inour previous experiments with untreated bacte-ria (4). Histograms obtained from bacteriatreated with boiling water or trichloroaceticacid (Fig. 10) were quite similar to those ob-tained previously with unwashed bacteria (4).Incorporation of DAP stayed homogeneousalong the cylindrical part of the cell wall duringthe whole division cycle, poles were poorlylabeled, whereas the cross wall was a very activesite of DAP incorporation during its formation.No small peaks of radioactivity appeared any-where along the cell wall after removal of themurein precursors.
DISCUSSIONThis study shows that the intracytoplasmic
radioactivity found in untreated bacteria iscompletely removed by a 3-min chase since thesilver grain distribution pattern becomes sym-metrical with respect to the cell wall. Thismeans that all the radioactivity located in thecytoplasm before the chase is now incorporatedinto the murein. If the exact nature of theseradioactive compounds stays unknown (free3H-labeled DAP molecules or murein precur-sors), the possibility of it being DAP metabo-lized into lysine and incorporated into proteinscan be eliminated. The chromatographic analy-sis of the extracted radioactive compoundsconfirms that there are no detectable free[3H]lysine molecules in the bacteria. This wasexpected since the strain used is a DAP- Lys-mutant of B. megaterium. Moreover, no free3H-labeled DAP molecules were found in theextract. Since all the cytoplasmic radioactivityis incorporated into the cell wall after a chase, itnecessarily represents murein precursors.A statistical analysis of autoradiographs of
untreated bacteria has shown that the distribu-tion of these precursors is not random butdecreases regularly from the cell wall to thecenter of the bacterium. It is tempting toassume that most of the silver grains locatednear the cytoplasmic membrane correspond toimmediate precursors, that is to say to dissac-charide pentapeptides linked to the glycosyllipid carrier of the cytoplasmic membrane, andthat silver grains located towards the center ofthe cell represent first-step precursors of themurein biosynthetic pathway. The observeddistribution of radioactivity is rather surprisingand it seems unlikely that it is an artefactoccurring during the first minutes of prefixa-tion. It is more probable that all enzymesinvolved in murein synthesis are located at the
VOL. 123, 1975
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
1204 DE CHASTELLIER, FREHEL, AND RYTER
i; s"N .V14
JO'1/
4 .. w--. I s
S
j
,.tK X
9q,..wi
'~~~~~~~~~~~~~1
N
, ½;- I
iw
a 14F
:Vk
6f
-I
c
.P
.f rN,A..t
6 .. /
4-C
.t' I
P.
4, A-s
I* F}~
4!
I.W
1 . .404,_. IN. DS~~~
i
w,rFI..
FIG. 7.
_
4\_ \ -V_I'9
f
J. BACTERIOL.
:a
ly- 14.
% 10,
I
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
CELL WALL GROWTH IN B. MEGATERIUM 1205
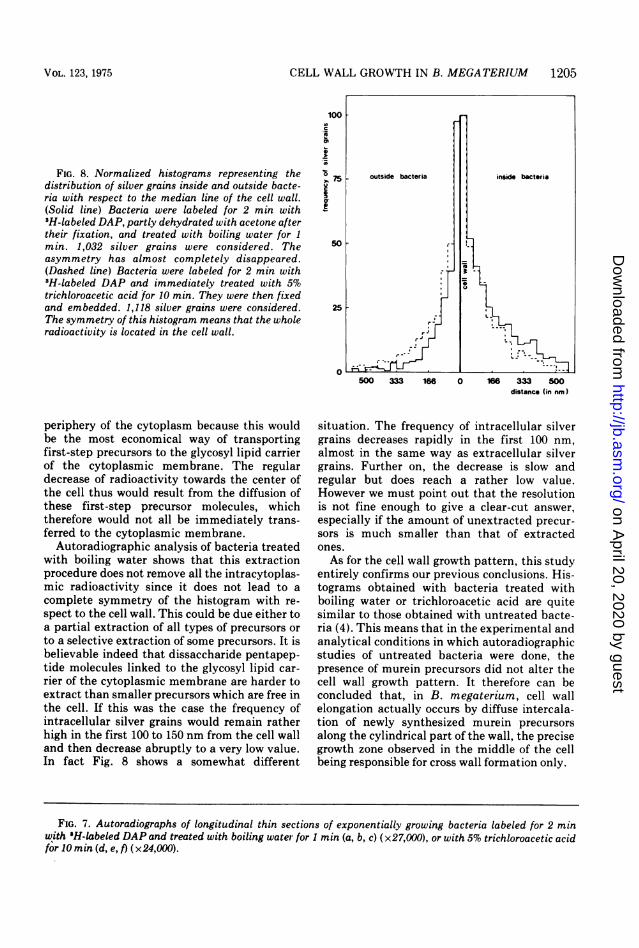
FIG. 8. Normalized histograms representing thedistribution of silver grains inside and outside bacte-ria with respect to the median line of the cell wall.(Solid line) Bacteria were labeled for 2 min with3H-labeled DAP, partly dehydrated with acetone aftertheir fixation, and treated with boiling water for 1min. 1,032 silver grains were considered. Theasymmetry has almost completely disappeared.(Dashed line) Bacteria were labeled for 2 min with3H-labeled DAP and immediately treated with 5%trichloroacetic acid for 10 min. They were then fixedand embedded. 1,118 silver grains were considered.The symmetry of this histogram means that the wholeradioactivity is located in the cell wall.
100
.m
.a
Nl 75
500 333 166 0 166 333 500distance (in nm)
periphery of the cytoplasm because this wouldbe the most economical way of transportingfirst-step precursors to the glycosyl lipid carrierof the cytoplasmic membrane. The regulardecrease of radioactivity towards the center ofthe cell thus would result from the diffusion ofthese first-step precursor molecules, whichtherefore would not all be immediately trans-ferred to the cytoplasmic membrane.
Autoradiographic analysis of bacteria treatedwith boiling water shows that this extractionprocedure does not remove all the intracytoplas-mic radioactivity since it does not lead to acomplete symmetry of the histogram with re-spect to the cell wall. This could be due either toa partial extraction of all types of precursors orto a selective extraction of some precursors. It isbelievable indeed that dissaccharide pentapep-tide molecules linked to the glycosyl lipid car-rier of the cytoplasmic membrane are harder toextract than smaller precursors which are free inthe cell. If this was the case the frequency ofintracellular silver grains would remain ratherhigh in the first 100 to 150 nm from the cell walland then decrease abruptly to a very low value.In fact Fig. 8 shows a somewhat different
situation. The frequency of intracellular silvergrains decreases rapidly in the first 100 nm,almost in the same way as extracellular silvergrains. Further on, the decrease is slow andregular but does reach a rather low value.However we must point out that the resolutionis not fine enough to give a clear-cut answer,especially if the amount of unextracted precur-sors is much smaller than that of extractedones.As for the cell wall growth pattern, this study
entirely confirms our previous conclusions. His-tograms obtained with bacteria treated withboiling water or trichloroacetic acid are quitesimilar to those obtained with untreated bacte-ria (4). This means that in the experimental andanalytical conditions in which autoradiographicstudies of untreated bacteria were done, thepresence of murein precursors did not alter thecell wall growth pattern. It therefore can beconcluded that, in B. megaterium, cell wallelongation actually occurs by diffuse intercala-tion of newly synthesized murein precursorsalong the cylindrical part of the wall, the precisegrowth zone observed in the middle of the cellbeing responsible for cross wall formation only.
FIG. 7. Autoradiographs of longitudinal thin sections of exponentially growing bacteria labeled for 2 minwith 'H-labeled DAP and treated with boiling water for 1 min (a, b, c) (x27,000), or with 5% trichloroacetic acidfor 10 min (d, e, f) (x24,000).
VOL. 123, 1975
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

1206 DE CHASTELLIER, FREHEL, AND RYTER
Lysine
$_DAP0
butanol -propionic acid - waterFIG. 9. Comparison of the autoradiograms of the extract to the chromatograms sprayed with ninhydrin, after
2 weeks of exposition. Bacteria were labeled for 2 min with 3H-labeled DAP. Extraction was done with boilingwater. Aliquots of the extract to which DAP or lysine were added as tracer were deposited on cellulose thin-layerchromatography plates and two-dimensional chromatography was done, using phenol-water in the firstdimension and butanol-propionic acid-water in the second dimension.
J. BACTERIOL.
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
CELL WALL GROWTH IN B. MEGATERIUM 1207
10
0o
*2
0
i.o
0 666 po. 0'l333 2000
20 F
10
0
Class
1 bacterium--~~~~~~~~~.2 bacteria
0 6664 po 1333 .* 2000 2866
distance (in nm)
FIG. 10. Silver grain distribution histograms along the cell wall with respect to the transversal axis ofsymmetry or the cross wall. Histograms were prepared in distance steps of 83.3 nm (2.5 nm at a x30,000enlargement). Quantities of silver grains are expressed as percentages of the total number of cell wall silvergrains. (Solid line) Bacteria treated with boiling water for 1 min. The number of silver grains counted was: (A)class I, 133; (B) class II, 678; (C) class III, 404; (D) class IV, 232. (Dashed line) Bacteria treated with 5%trichloroacetic acid for 10 min. The number of silver grains counted was: (A) class I, 193; (B) class II, 338; (C)class III, 214; (D) class IV, 177.
ACKNOWLEDGMENTS
We wish to thank Jean-Paul Aubert for his interest andadvice in this work and Raymonde Daty for excellent techni-cal assistance. Suggestions and assistance in chromatographyexperiments by Claudine Elmerich are greatly acknowledged.
This work was supported by grants from the DelegationGenerale a la Recherche Scientifique et Technique (researchgrant N°71-7-3082), from the Centre National de la RechercheScientifique (Laboratoire associe n°88) and from the Commis-sariat a l'Energie Atomique.
LITERATURE CITED
1. Anderson, J. S., M. Matsuhashi, M. A. Hoskin, and J. L.Strominger. 1965. Lipid-phosphoacetyl-muramyl-pen-tapept ide and lipid-phosphodissaccharide- pentapep-tide: presumed membrane transport intermediates incell wall synthesis. Proc. Natl. Acad. Sci. U.S.A.53:881-889.
2. Aubert, J. P., J. Millet, E. Pineau, and G. Milhaud. 1961.
L'acide N-succinyl L glutamique chez Bacillusmegaterium en voie de sporulation. Biochim. Biophys.Acta 51:529-537.
3. Benson, A. A., J. A. Bassham, M. Calvin, T. C. Goodale,V. A. Haas, and W. Stepka. 1950. The path of carbon inphotosynthesis. V. Paper chromatography and radioau-tography of the products. J. Am. Chem. Soc.72:1710-1718.
4. De Chastellier, C., R. Hellio, and A. Ryter. 1975. Study ofthe cell wall growth in Bacillus megaterium by high-res-olution autoradiography. J. Bacteriol. 123:1184-1196.
5. Ghuysen, J. M. 1968. Use of bacteriolytic enzymes indetermination of cell wall structure and their role in cellmetabolism. Bacteriol. Rev. 32:425-464.
6. Osborn, M. J. 1969. Structure and biosynthesis of thebacterial cell wall. Annu. Rev. Biochem. 38:501-538.
7. Strominger, J. L., K. Izaki, M. Matsuhashi, and D. J.Tiffer. 1967. Peptidoglycan transpeptidase and D-ala-nine carboxypeptidase, penicillin sensitive enzymaticreactions. Fed. Proc. 26:9-21.
Class I CDp3- - 66
0--.A666
200
Class IE )
I@ , C
666 13334po" 2000 266
VOL. 123, 1975
W6 4 b1 133 2000
I
on April 20, 2020 by guest
http://jb.asm.org/
Dow
nloaded from


















