
Cellular requirements for lymphokine secretion by rainbow trout Salmo gairdneri leucocytes
10
Developmental and ComparativeImmunology, Vol. 14, pp. 59-68, 1990 0145-305X/90 $3.00 + .00 Printed in the USA. AlL rights reserved. Copyright © 1990 Pergamon Press plc CELLULAR REQUIREMENTS FOR LYMPHOKINE SECRETION BY RAINBOW TROUT SALMO GAIRDNERI LEUCOCYTES Susan Graham and Christopher J. Secombes Department of Zoology, University of Aberdeen, TiLlydrone Ave., Aberdeen,AB9 2TN, UK (Submitted March 1989; Accepted May 1989) [~Abstract--The ability of different popula- tions of rainbow trout blood leucocytesto pro- duce MAF following stimulation with Con A/PMA was assessed by the amount of NBT reduction in target macrophages. The effect of varying lymphocyteor macrophagenumber on MAF production in the presence of a constant number of macrophages or iymphocytes re- spectively, showed that in both cases MAF ac- tivity initially increased with increasing cell number and then plateaued. Macrophages alone did not produce MAF whereas some MAF activity was produced by macrophage- depleted lymphocytes, although significantly lower than in the presence of macrophages. Separation of leucocytes into sIg- and sIg + cells by panning showed that only sIg- lym- phocytes could produce MAF and that macro- phages were necessaryas accessorycells. These results support the contention that fish lym- phocytes can be divided into sIg- T cells and sIg+ B cells. []Keywords--Lymphokine secretion; Cellular requirements; Rainbow trout; Panning; slg + and sIg- cells; MAF; Macrophage-depletion; Lymphocyte-depletion. Introduction It is generally agreed that the earliest manifestations of the specific immune system occur in fish. Their ability to produce antibody and reject allografts demonstrates the possession of both hu- moral and cell-mediated immunity. However, formal proof for the existence of T and B lymphocytes in fish is Address correspondence to Dr. Christopher J. Secombes, Department of Zoology, Uni- versity of Aberdeen, Tillydrone Avenue, Aberdeen, U.K. lacking, due to difficulties encountered in thymectomizing young fish, the lack of inbred strains, and the lack of conve- nient surface markers. That two lym- phocyte populations exist in fish has been suggested by studies looking at mi- togen responses and the hapten-carrier effect (1-4), and indeed in these reac- tions two functionally distinct popula- tions have been separated by adherence to nylon wool columns (5) or on the basis of nonimmune rosetting with heter- ologous erythrocytes (6). More conclu- sive evidence that the populations pre- sent are similar to T and B cells of higher vertebrates have been forthcoming over the last few years from studies using an- tibodies to lymphocyte surface deter- minants to enrich or deplete for these putative lymphocyte populations. Monoclonal antibodies (MoAbs) to serum IgM of a range of teleost fish have been raised that react with only a subpopulation (30-40%) of blood lym- phocytes (7-9) suggesting that surface Ig (slg) can be used as a marker for B- like cells. By selectively enriching for slg+ or slg- lymphocytes by "pan- ning" with these MoAbs the different populations have been separated for functional studies in vitro. Such studies have shown that slg+ lymphocytes re- spond to LPS whereas slg- lympho- cytes respond to Con A, although the re- sponse to LPS is not completely re- moved (8,10). In the mixed leucocyte response (MLR) only slg- cells were found to undergo proliferation, but both slg + and slg- lymphocytes could act as stimulator cells (11). In addition, slg+ lymphocytes were found to be 59
-
Upload
susan-graham -
Category
Documents
-
view
212 -
download
0
Transcript of Cellular requirements for lymphokine secretion by rainbow trout Salmo gairdneri leucocytes

Developmental and Comparative Immunology, Vol. 14, pp. 59-68, 1990 0145-305X/90 $3.00 + .00 Printed in the USA. AlL rights reserved. Copyright © 1990 Pergamon Press plc
CELLULAR REQUIREMENTS FOR LYMPHOKINE SECRETION BY RAINBOW TROUT SALMO GAIRDNERI LEUCOCYTES
Susan Graham and Chr is topher J. Secombes
Department of Zoology, University of Aberdeen, TiLlydrone Ave., Aberdeen, AB9 2TN, UK (Submitted March 1989; Accepted May 1989)
[~Abstract--The ability of different popula- tions of rainbow trout blood leucocytes to pro- duce MAF following stimulation with Con A/PMA was assessed by the amount of NBT reduction in target macrophages. The effect of varying lymphocyte or macrophage number on MAF production in the presence of a constant number of macrophages or iymphocytes re- spectively, showed that in both cases MAF ac- tivity initially increased with increasing cell number and then plateaued. Macrophages alone did not produce MAF whereas some MAF activity was produced by macrophage- depleted lymphocytes, although significantly lower than in the presence of macrophages. Separation of leucocytes into sIg- and sIg + cells by panning showed that only sIg- lym- phocytes could produce MAF and that macro- phages were necessary as accessory cells. These results support the contention that fish lym- phocytes can be divided into sIg- T cells and sIg+ B cells.
[]Keywords--Lymphokine secretion; Cellular requirements; Rainbow trout; Panning; slg + and sIg- cells; MAF; Macrophage-depletion; Lymphocyte-depletion.
Introduction
It is generally agreed that the earliest manifestations of the specific immune system occur in fish. Their ability to produce antibody and reject allografts demonstrates the possession of both hu- moral and cell-mediated immunity. However, formal proof for the existence of T and B lymphocy te s in fish is
Address correspondence to Dr. Christopher J. Secombes, Department of Zoology, Uni- versity of Aberdeen, Tillydrone Avenue, Aberdeen, U.K.
lacking, due to difficulties encountered in thymectomizing young fish, the lack of inbred strains, and the lack of conve- nient surface markers. That two lym- phocyte populations exist in fish has been suggested by studies looking at mi- togen responses and the hapten-carrier effect (1-4), and indeed in these reac- tions two functionally distinct popula- tions have been separated by adherence to nylon wool columns (5) or on the basis of nonimmune rosetting with heter- ologous erythrocytes (6). More conclu- sive evidence that the populations pre- sent are similar to T and B cells of higher vertebrates have been forthcoming over the last few years from studies using an- tibodies to lymphocyte surface deter- minants to enrich or deplete for these putative lymphocyte populations.
Monoclonal antibodies (MoAbs) to serum IgM of a range of teleost fish have been raised that react with only a subpopulation (30-40%) of blood lym- phocytes (7-9) suggesting that surface Ig (slg) can be used as a marker for B- like cells. By selectively enriching for s lg+ or s l g - lymphocytes by "pan- ning" with these MoAbs the different populations have been separated for functional studies in vitro. Such studies have shown that slg+ lymphocytes re- spond to LPS whereas s l g - lympho- cytes respond to Con A, although the re- sponse to LPS is not completely re- moved (8,10). In the mixed leucocyte response (MLR) only s l g - cells were found to undergo proliferation, but both slg + and s l g - lymphocytes could act as stimulator cells (11). In addition, s lg+ lymphocytes were found to be
59

60 S. Graham and C. J. Secombes
neces sa ry for an t i -hap ten plaque- forming cell (PFC) responses to thymus- independent antigens in contrast to PFC responses to thymus-dependent antigens which required both sIg+ and s I g - lymphocytes (12). By combining pan- ning with anti-lg MoAbs and specific ad- herence to hapten or carrier molecules it has been possible to demonstrate that both hapten-responsive slg + and carrier r e spons ive s l g - l y m p h o c y t e s are needed for the anti-hapten response to a thymus-dependent antigen (12).
The requirement for macrophages or monocytes in these responses has also been investigated since they can be re- moved by adherence to baby hamster kidney cell microexudate-coated plates or by passing cells through a Sephadex G-10 column. Mitogen and MLR re- sponses by s l g - cells were dependent upon the presence of macrophages (10,11), as was the anti-hapten PFC re- sponse to thymus-dependent and -inde- pendent antigens (12). Only the LPS re- sponse by slg + cells has been shown to occur in the absence of macrophages to date (10).
Antibodies to determinants on T-like cells have also been used to investigate lymphocyte heterogeneity in fish. For instance, antibrain serum, thought to react with Thy-l-like antigens present on T cells and nervous tissue, can selec- tively kill PHA responsive cells but not LPS responsive cells (6). More recently a MoAb has been produced that specifi- cally reacts with s I g - channel catfish lymphocytes and has been used to iso- late those cells that provided helper ac- tivity for antibody synthesis to a thymus- dependent antigen (13).
Although the thymus-dependency of these populations has still to be ascer- tained, such data provide compelling ev- idence for the existence of T and B cells in fish. Further proof for functions such as the production of lymphokine and the generation of cytotoxic cells will strengthen this conclusion. To this end
we have recently optimized the produc- tion and detection of a macrophage acti- vating factor (MAF) secreted from rainbow trout (Salmo gairdneri) leuco- cytes (14). In this article we report on the secretion of MAF by slg + and s l g - lymphocytes separated using an anti- trout MoAb (I/14), and on the accessory cell requirements.
Materials and Methods
The Production and Detection of MAF- Containing Supernatants
MAF containing supernatants were produced as described by Graham & Se- combes (14). Briefly, leucocyte suspen- sions were adjusted to 5 x 106 cells per ml in L-15 medium (L-15) containing I% Penicillin/Streptomycin (P/S), 5 x 10 -5 M 2-mercaptoethanol (2-ME), and l ml seeded into 24 well microtitre plates (MTP, Nunc). They were then pulsed with 10 ug/ml Concanavalin A (Con A, Difco) and 5 ng/ml phorbol myristate ac- etate (PMA, Sigma), previously shown to be optimal for MAF production (14), for 3h at 18°C. After this time the cells were gently washed five times with sterile phosphate buffered saline (PBS) to remove any residual Con A and PMA, and then cultured in L-15 containing 1% P/S, 10% foetal calf serum (FCS) and 5 x 10 -5 M 2-ME for 48h. The superna- rants were then harvested and stored at -20°C until use.
MAF activity was detected using pu- rified head kidney (HK) macrophages as targets. These cells were isolated using Percoll density gradients followed by ad- herence to 96 well MTP as previously described (15,16). Following a 48h incu- bation with the MAF containing super- natants the respiratory burst activity of triplicate wells of macrophages was as- sessed by their ability to reduce ni- troblue tetrazolium (NBT). The macro- phage monolayers were covered with

Lymphokine secretion by trout 61
100 ~1 of 1 mg/ml NBT containing 1 ixg/ml PMA, to trigger the respiratory burst, for 30 min before being fixed in methanol. After washing in 70% meth- anol the reduced formazan within the macrophages was solubilized with KOH/ DMSO (17) and the optical density de- termined at 620 nm in a multiscan spec- trophotometer. All results were ex- pressed per l0 s macrophages af ter counting nuclei released with 0.1M citric acid, Tween 20 and 0.05% crystal violet in control wells. In some cases the re- sults were expressed as a stimulation index by dividing the mean optical den- sity reading from macrophages incu- bated with active supernatants with that from macrophages incubated with con- trol supernatants obtained from leuco- cytes not stimulated with Con A/PMA.
Effect of Lymphocyte Number on MAF Production
The effect of varying lymphocyte numbers on the production of MAF was studied by isolating a constant number of macrophages and adding to them varying numbers of leucocytes exten- sively depleted of macrophages (as as- sessed by a lack of adherent cells), prior to addition of Con A/PMA. Blood leuco- cytes (I ml) obtained after centrifugation over a 51% Percoll density gradient, were seeded into 24 well MTP at 1 × 107 cells per ml L-15 containing 0.1% FCS and 1% P/S. After 2h at 18°C for adher- ence, the nonadherent cells were re- moved and the adherent cells were cov- ered with 1 ml L-15, 5% FCS, 1% P/S. The nonadherent cells were then centri- fuged at 400g for 10 min and the pellet of cells resuspended to 2 × 107 cells per ml in L-15, 10% FCS, 1% P/S. The lympho- cytes were then extensively depleted of macrophages by passage through a Sephadex G-10 column prepared as de- scribed by Mishell and Mishell (18). Ap-
proximately 1 ml of the cell suspension was put onto the column containing 6 ml sterile Sephadex G-10. The cells were allowed to penetrate the column before addition of 10 ml of medium to elute the nonadherent cells. After centrifugation the cells were resuspended in fresh me- dium and cultured overnight in a 25 cm z flask to remove any residual macro- phages that did not adhere to the column. The nonadherent lymphocytes were then adjusted to 1.6 x 105 to 107 cells per ml, and were added back to the constant number of macrophages pre- viously set up. MAF-containing super- natants were then produced as described above.
Effect of Macrophage Number on MAF Production
The effect of varying macrophage number on the production of MAF was determined by isolating different con- centrations of macrophages and adding to them a constant number of leucocytes which had been extensively depleted of macrophages, prior to the addition of Con A/PMA. Blood leucocytes were seeded into 24 well MTP in two-fold di- lutions from 1.96 x 105 to 5 x 107 cells per ml L-15, 0.1% FCS, 1% P/S to ob- tain varying macrophage numbers. Du- plicate wells of each cell concentration were used to enable macrophage nuclei counts to be performed. The cells were allowed to adhere for 2h at 18°C after which the nonadherent cells were col- lected and the adherent macrophages were supplemented with L-15, 5% FCS, 1% P/S and placed at 18°C until use. The nonadherent ceils were then depleted extensively of macrophages as above and 5 x 10 6 viable iymphocytes were added back to each well of macrophages previously set up. MAF-containing su- pernatants were then produced as de- scribed earlier.

62 S. Graham and C. J. Secombes
Separation of slg + and slg - Lymphocytes by Panning
Petri dishes (I0 cm, Nunc) were coated overnight at 4°C with 8 ml of con- trol ascitic fluid, obtained using a clone against an unrelated antigen, or mono- clonal anti-trout Ig (1/14) ascitic fluid di- luted 1:200 in PBS. The plates were then washed twice with PBS and 8 ml L-15, 10% FCS was added for lh at 18°C be- fore addition of blood leucocytes. The leucocytes were initially panned for lh on control ascitic fluid coated plates at 2 × 107 cells per dish, to remove adherent s l g - leucocytes, and subsequently were panned twice on 1-14 coated plates. The nonadherent cells obtained after the second 1-14 panning were termed s l g - lymphocytes. After removal of the non- adherent cells from the 1-14 coated plates an intermediate wash was carried out before removal of the adherent slg + cells by pipetting medium onto the plate. The two batches of slg+ lymphocytes were pooled before use and both the slg+ and the s l g - lymphocytes were then cultured overnight in 25 cm 2 flasks. Finally the nonadherent cells from these flasks were added to isolated macro- phages from the same fish (see below) and used to generate MAF-containing supernatants as above. In some experi- ments s l g - cells were stimulated with Con A/PMA in the absence of macro- phages. While the panning was occur- ring blood leucocytes from the same fish were added to a 24 well MTP at 5 × 106 cells per well, to serve as a total leuco- cyte control and as a source of mac- rophages for " a d d - b a c k " and mac- rophage-only controls. For this latter purpose the nonadherent cells were re- moved after lh at 18°C and the adherent cells supplemented with L-15, 10% FCS until use.
Flow Cytofluorometric Analysis
To confirm that the separation of lym- phocytes into sIg+ and s Ig - cells had
been successful on each occasion that this experiment was performed, a sub- sample of the cells was analyzed by flow cytofluorometry. 5 × 105 to 10 6 cells per ml were incubated with 1/14 supernatant containing 0.1% sodium azide, 1% bo- vine serum albumin (BSA). After lh at 4°C the cells were washed three times by centrifugation and incubated for a fur- ther lh with sheep anti-mouse IgG/FITC diluted 1:40 in PBS, 0.1% sodium azide, 1% BSA. Finally the cells were washed three additional times in PBS, 0.1% so- dium azide and were then analyzed in an EPICS-C flow cytometer (Coulter Elec- tronics) with the argon laser tuned to 488 nm and PMT voltage at 1600 V. Graphs of log fluorescence intensity versus cell number were plotted for 10,000 cells.
Results were analyzed using Student's two-tailed t-test.
Results
Effect of Lymphocyte Number on MAF Production
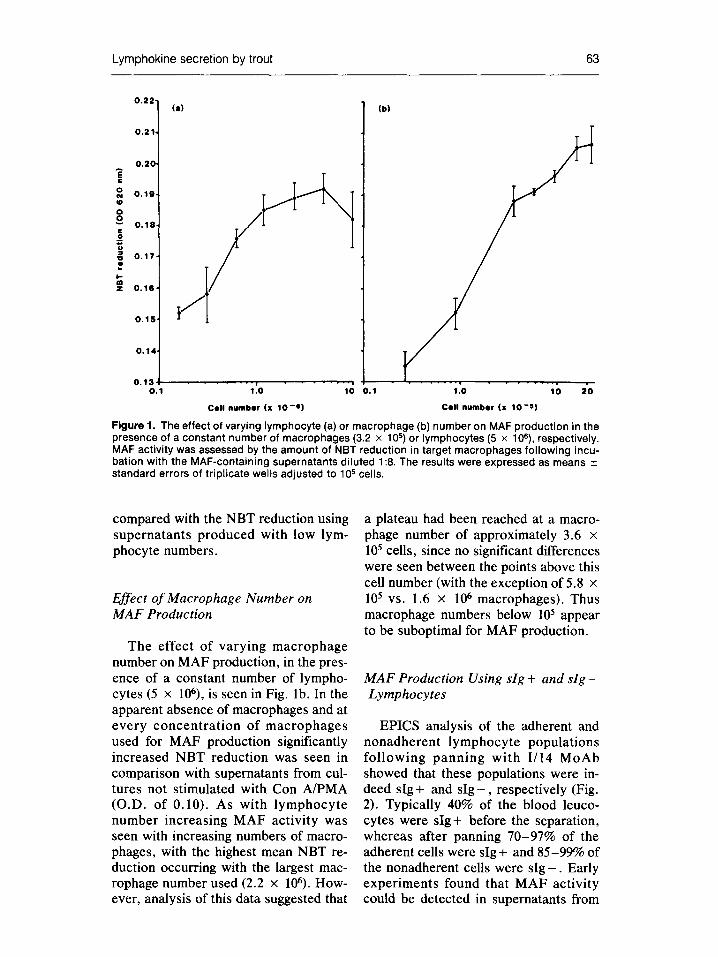
The effect of varying lymphocyte number on MAF production, in the pres- ence of a constant number of macro- phages (3.2 × 105), is seen in Fig. la. In the absence of lymphocytes no MAF was produced by macrophages, as deter- mined by the amount of NBT reduced by macrophages incubated with these supernatants in comparison with super- natants from cultures not stimulated with Con A/PMA (O.D. of 0.15). Low numbers of lymphocytes, 1.6-3.1 × 105 , also did not produce significant levels of MAE With larger numbers of lymphocytes significant MAF activity was seen, and inc reased with cell number up to a maximum at 5 × 10 6
cells. The MAF activity appeared to pla- teau using more than 106 lymphocytes in that there were no significant differences in the amount of NBT reduction be- tween these cell concentrations, but it was significantly higher in each case
Lymphokine secretion by trout 63
0 .22
0.21
0.20
S ~ 0 . 1 9
Q o ~ o . 1 8 .
,D 0 . 1 7 - e
i,- m z 0 . 1 6 -
0.15.
0.14"
0.13 0.1
(e)
Cell number (x 1 0 - e )
(b)
1.0 10 2"0
Cell number (x 10 - s )
Figure 1. The effect of varying lymphocyte (a) or macrophage (b) number on MAF production in the presence of a constant number of macrophages (3.2 × 10 s) or lymphocytes (5 × 106), respectively. MAF activity was assessed by the amount of NBT reduction in target macrophages following incu- bation with the MAF-containing supernatants diluted 1:8. The results were expressed as means _+ standard errors of triplicate wells adjusted to 10 s cells.
compared with the NBT reduction using supernatants produced with low lym- phocyte numbers.
Effect of Macrophage Number on MAF Production
The effect of varying macrophage number on MAF production, in the pres- ence of a constant number of lympho- cytes (5 x 106), is seen in Fig. lb. In the apparent absence of macrophages and at every concentrat ion of macrophages used for MAF production significantly increased NBT reduction was seen in comparison with supernatants from cul- tures not stimulated with Con A/PMA (O.D. of 0.10). As with lymphocyte number increasing MAF activity was seen with increasing numbers of macro- phages, with the highest mean NBT re- duction occurring with the largest mac- rophage number used (2.2 x 106). How- ever, analysis of this data suggested that
a plateau had been reached at a macro- phage number of approximately 3.6 × 105 cells, since no significant differences were seen between the points above this cell number (with the exception of 5.8 × 105 vs. 1.6 × 106 macrophages). Thus macrophage numbers below 105 appear to be suboptimal for MAF production.
MAF Production Using slg + and slg - Lymphocytes
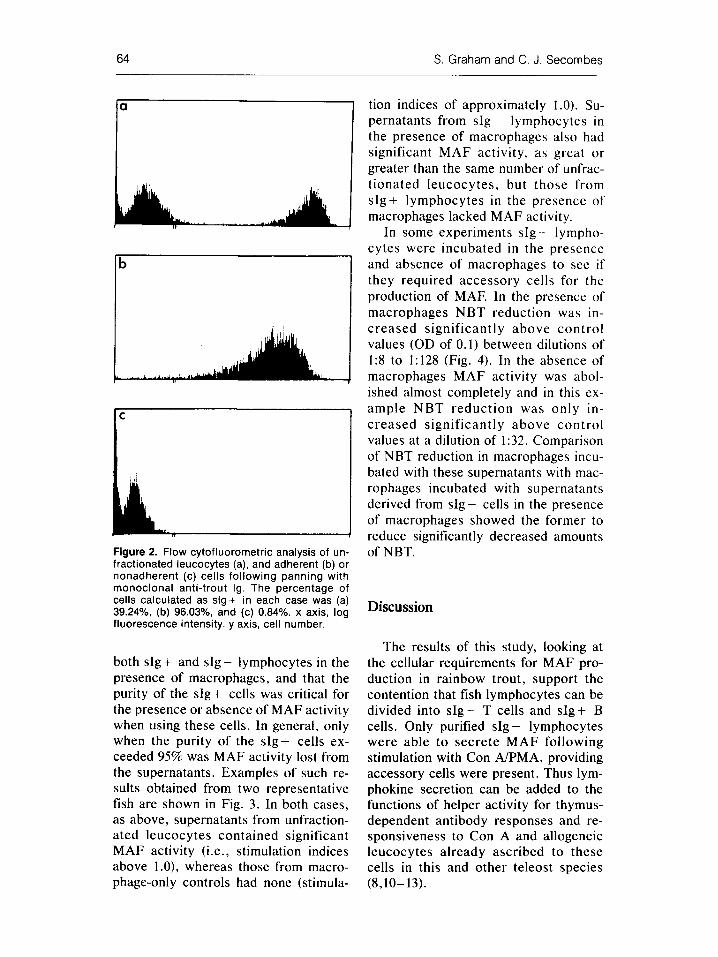
EPICS analysis of the adherent and nonadherent lymphocyte populations fol lowing panning with 1/14 MoAb showed that these populations were in- deed slg + and s l g - , respectively (Fig. 2). Typically 40% of the blood leuco- cytes were slg+ before the separation, whereas after panning 70-97% of the adherent cells were slg + and 85-99% of the nonadherent cells were s l g - . Early experiments found that MAF activity could be detected in supernatants from
64 S. Graham and C. J. Secombes
6
I b
IC .i
Figure 2. Flow cytofluorometric analysis of un- fractionated leucocytes (a), and adherent (b) or nonadherent (c) cells following panning with monoclonal anti-trout Ig. The percentage of cells calculated as slg+ in each case was (a) 39.24%, (b) 96.03%, and (c) 0.84%. x axis, log fluorescence intensity, y axis, cell number.
both slg + and s l g - lymphocytes in the presence of macrophages, and that the purity of the slg+ cells was critical for the presence or absence of MAF activity when using these cells. In general, only when the purity of the slg+ cells ex- ceeded 95% was MAF activity lost from the supernatants. Examples of such re- sults obtained from two representative fish are shown in Fig. 3. In both cases, as above, supernatants from unfraction- ated leucocytes contained significant MAF activity (i.e., stimulation indices above 1.0), whereas those from macro- phage-only controls had none (stimula-
tion indices of approximately 1.0). Su- pernatants from s l g - lymphocytes in the presence of macrophages also had significant MAF activity, as great or greater than the same number of unfrac- tionated leucocytes, but those from slg+ lymphocytes in the presence of macrophages lacked MAF activity.
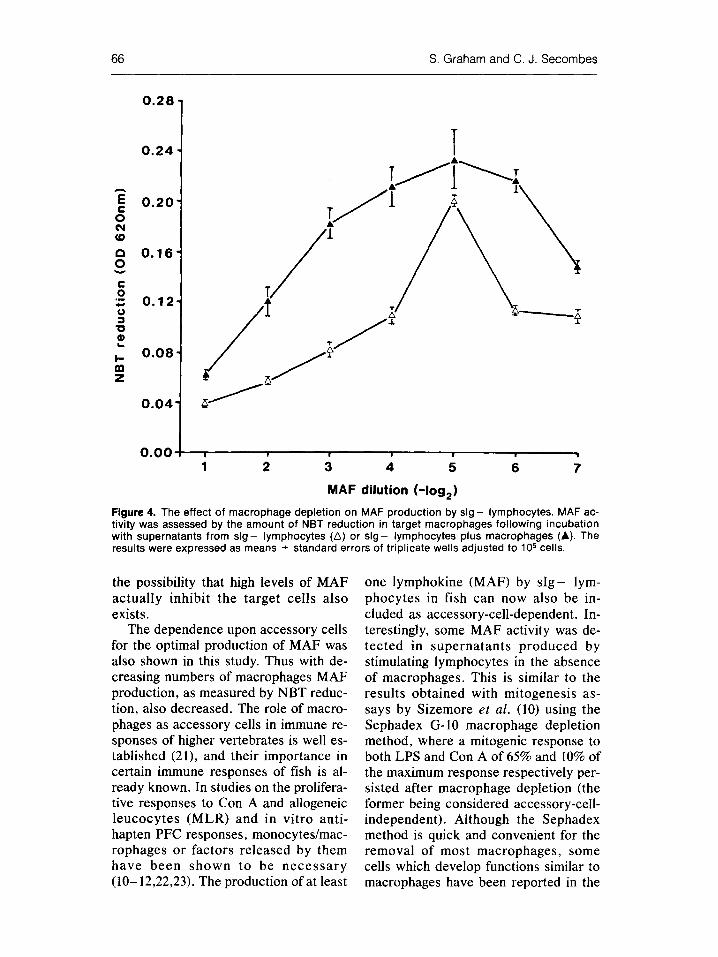
In some experiments s l g - lympho- cytes were incubated in the presence and absence of macrophages to see if they required accessory cells for the production of MAE In the presence of macrophages NBT reduction was in- creased significantly above control values (OD of 0.1) between dilutions of 1:8 to 1:128 (Fig. 4). In the absence of macrophages MAF activity was abol- ished almost completely and in this ex- ample NBT reduction was only in- creased significantly above control values at a dilution of 1:32. Comparison of NBT reduction in macrophages incu- bated with these supernatants with mac- rophages incubated with supernatants derived from s l g - cells in the presence of macrophages showed the former to reduce significantly decreased amounts of NBT.
Discussion
The results of this study, looking at the cellular requirements for MAF pro- duction in rainbow trout, support the contention that fish lymphocytes can be divided into s l g - T cells and slg+ B cells. Only purified s l g - lymphocytes were able to secrete MAF following stimulation with Con A/PMA, providing accessory cells were present. Thus lym- phokine secretion can be added to the functions of helper activity for thymus- dependent antibody responses and re- sponsiveness to Con A and allogeneic leucocytes already ascribed to these cells in this and other teleost species (8,10-13).

Lymphokine secretion by trout 65
2.5"
2.0"
i 1.5" !
-$
W 1.0'
0.5 '
Fish 1 Fish 2
MAF dilution ( - log 2)
Figure 3. MAF production by unfractionated blood leucocytes (0), macrophages (O), s lg - lym- phocytes plus macrophages (A) and slg + lymphocytes plus macrophages (A). MAF activity was assessed by the amount of NBT reduction in target macrophages following incubation with the MAF-containing supernatants The results were expressed as a stimulation index by dividing the mean optical density reading from macrophages incubated with test supernatants with that from macrophages incubated with control supernatants obtained from leucocytes not stimulated with Con A/PMA.
To demonstrate whether this teleost MAF was truly a lymphokine secreted from lymphocytes, experiments were carried out using varying numbers of macrophage-depleted leucocytes added to a constant number of macrophages and vice versa. Since salmonid blood leucocytes normally contain < 10% gran- ulocytes (19), after macrophage-deple- tion predominantly lymphocytes and some thrombocytes will remain, and the major role of the latter is in blood coagu- lation and general hemostasis (20). In both experimental approaches MAF ac- tivity initially increased with increasing numbers of macrophages or lympho- cytes and then appeared to plateau. Perhaps more significantly MAF activity was completely absent from macrophage cultures without lymphocytes, whereas in the presence of optimal lymphocyte
numbers even in the apparent absence of macrophages significant MAF activity was present. In addition the panning ex- periments showed that in the presence of optimal numbers of macrophages MAF activity was only found using s l g - lymphocytes and not s lg+ lym- phocytes. Thus s l g - lymphocytes and not macrophages or s lg+ lymphocytes would appear to be the responding cells.
It is interesting to note that at high concentrations of these lymphokine- containing supernatants a suppressive effect upon NBT reduction was some- times seen, as shown in Figs. 3 and 4. This could be due to a multiplicity of factors being present in the supernatant, some of which were suppressive. How- ever, this effect could still be seen fol- lowing partial purification of the MAF activity by HPLC (unpublished), and so
66 S. Graham and C. J. Secombes
0 . 2 8
0 . 2 4
A
E 0 . 2 0 c O ~N (O
0 . 1 6 O
c O
0 . 1 2 o
"O L .
0 . 0 8
Z
0 .04 '
0 . 0 0
T
! ! i i | •
2 3 4 5 6 7
MAF dilution (-log 2)
Figure 4. The effect of macrophage depletion on MAF production by s l g - lymphocytes. MAF ac- tivity was assessed by the amount of NBT reduction in target macrophages following incubation with supernatants from s l g - lymphocytes (G) or s l g - lymphocytes plus macrophages (A). The results were expressed as means _+ standard errors of triplicate wells adjusted to 10 s cells.
the possibility that high levels of MAF actually inhibit the target cells also exists.
The dependence upon accessory cells for the optimal production of MAF was also shown in this study. Thus with de- creasing numbers of macrophages MAF production, as measured by NBT reduc- tion, also decreased. The role of macro- phages as accessory cells in immune re- sponses of higher vertebrates is well es- tablished (21), and their importance in certain immune responses of fish is al- ready known. In studies on the prolifera- tive responses to Con A and allogeneic leucocytes (MLR) and in vitro anti- hapten PFC responses, monocytes/mac- rophages or factors released by them have been shown to be neces sa ry (10-12,22,23). The production of at least
one lymphokine (MAF) by s l g - lym- phocytes in fish can now also be in- cluded as accessory-cell-dependent. In- terestingly, some MAF activity was de- tected in supernatants produced by stimulating lymphocytes in the absence of macrophages. This is similar to the results obtained with mitogenesis as- says by Sizemore et al. (10) using the Sephadex G-10 macrophage depletion method, where a mitogenic response to both LPS and Con A of 65% and 10% of the maximum response respectively per- sisted after macrophage depletion (the former being considered accessory-cell- independent). Although the Sephadex method is quick and convenient for the removal of most macrophages, some cells which develop functions similar to macrophages have been reported in the

Lymphokine secretion by trout 67
filtered population (18). Thus it is pos- sible that some macrophages were not removed using the depletion technique, and provided a limited accessory cell function. Alternatively, the presence of PMA in the present study may have partly overridden the need for accessory cells since another phorbol ester TPA has been shown to overcome the acces- sory cell requirement for Con A stimula- tion of thymocytes in channel catfish (24). Whatever the explanation, intact macrophages clearly provide most effec- tive accessory signals.
The plateauing of NBT reduction of target macrophages using more than 10 6
lymphocytes is in contrast to the studies of the mitogenic response of channel catfish lymphocytes to Con A and LPS where proliferation increased linearly with increasing lymphocyte number up to 5 x 105 (10) and was used as evidence that lymphocytes were the mitogen-re- sponsive cells in this system. Had the study on mitogen responses used larger numbers of lymphocytes it is possible that a plateauing of the response would also have been seen. Alternatively, since lymphocyte proliferation in response to mitogens is a direct measurement, whereas in this study the level of lym- phokine released is measured indirectly by its effect on target macrophages, the plateauing effect could be due to the
target cells becoming saturated and not being able to respond further.
The panning experiments coupled with flow cytofluorometr ic analysis showed that it was crucial to have highly purified cell types in order to see any differences in MAF production. Con- tamination of the s lg+ lymphocytes with >5% s l g - lymphocytes was often sufficient to detect MAF activity in the supernatants. Assuming s l g - lympho- cytes are the responding cells from the above arguments , then clearly only small numbers are required in order to release enough MAF to stimulate large numbers of target macrophages. If this is the case then the plateauing effect of NBT reduction by target macrophages would not be surpris ing since the number of lymphocytes able to secrete sufficient MAF to stimulate all of the fixed number of target cells would soon be reached.
Further studies are in progress to characterize this teleost lymphokine, to see if it is equivalent to known mamma- lian MAFs such as ~/-interferon.
A c k n o w l e d g e m e n t s - - T h i s w o r k was sup- ported by S E R C grant No. GR/E/15413. The authors gratefully acknowledge the gift of the 1/14 hybr idoma cells f rom Professor G. W. Wart, Univers i ty of South Carolina. Thanks also go to Miss K. E. Henderson and Mr. J. Milton for the EPICS analysis.
References
1. Stolen, J. S.; Makela, O. Cartier preimmuni- zation in the anti-hapten response of a marine fish. Nature. 254:718-719; 1975.
2. Yocum, D.; Cuchens, M.; Clem, L. W. The hapten-carrier effect in teleost fish. J. Im- munol. 114:925-927; 1975.
3. Etlinger, H. M.; Hodgins, H. O.; Chiller, J. M. Evolution of the lymphoid system. I. Evidence for lymphocyte heterogeneity in rainbow trout revealed by the organ distribu- tion of mitogenic responses. J. Immunol. 116:1547-1553; 1976.
4. Chilmonczyk, S. In vitro stimulation by mi- togens of peripheral blood lymphocytes from rainbow trout (Salmo gairdneri). Ann. D'lm- munol. 129C:3-12; 1978.
5. Ruben, L. N.; Warr, G. W.; Decker, J. M.; Marchalonis, J. J. Phylogenetic origins of im- mune recognition: lymphoid heterogeneity and the hapten/carrier effect in the goldfish, Car- assius auratus. Cell. Immunol. 31:266-283; 1977.
6. Cuchens, M. A.; Clem, L. W. Phylogeny of lymphocyte heterogeneity. II. Differential ef- fects of temperature on fish T-like and B-like cells. Cell. Immunol. 34:219-230; 1977.
7. Lobb, C. J.; Clem, L. W. Fish lymphocytes differ in the expression of surface immuno- globulin. Dev. Comp. Immunol. 6:473-479; 1982.
8. Deluca, D.; Wilson, M.; Wart, G. W. Lympho- cyte heterogeneity in the trout, Salmo gaird-

68 S. Graham and C. J. Secombes
neri, defined with monoclonal antibodies to lgM. Eur. J. Immunol. 13:546-551; 1983.
9. Secombes, C. J.; van Groningen, J. J. M.; Eg- hefts, E. Separation of lymphocyte subpopu- lations in carp Cyprinus carpio L. by mono- clonal ant ibodies : immunohis tochemica l studies. Immunology. 48:165-175; 1983.
10. Sizemore, R. C.; Miller, N. W.; Cuchens, M. A.; Lobb, C. J.; Clem, L. W. Phylogeny of lymphocyte heterogeneity: the cellular re- quirements for in vitro mitogenic responses of channel catfish leukocytes. J. Immunol. 133: 2920-2924; 1984.
11. Miller, N. W.; Deuter, A.; Clem, L. W. Phy- logeny of lymphocyte heterogeneity: the cel- lular requirements for the mixed leucocyte re- action with channel catfish. Immunology. 59: 123-128, 1986.
12. Miller, N. W.; Sizemore, R. C.; Clem, L. W. Phylogeny of lymphocyte heterogeneity: the cellular requirements for in vitro antibody re- sponse of channel catfish leukocytes. J. lm- munol. 134:2884-2888; 1985.
13. Miller, N. W.; Bly, J. E.; van Ginkel, F.; Ell- saesser, C. E; Clem, L. W. Phylogeny of lym- phocyte heterogeneity: identification and sep- aration of functionally distinct subpopulations of channel catfish lymphocytes with mono- clonal antibodies. Dev. Comp. Immunol. 11: 739-747; 1987.
14. Graham, S.; Secombes, C. J. The production of a macrophage-activating factor from rain- bow trout Salmo gairdneri leucocytes. Immu- nology. 65:293-297; 1988.
15. Chung, S.; Secombes, C. J. Activation of rainbow trout macrophages. J. Fish. Biol. 31A:51-56; 1987.
16. Chung, S.; Secombes, C. J. Analysis of events occurring within teleost macrophages during the respiratory burst. Comp. Biochem. Physiol. 89B:539-544; 1988.
17. Rook, G. A.; Steele, J.; Umar, S.; Dockrell, H. M. A simple method for the solubilisation of reduced NBT, and its use as a colorimetric assay for activation of human macrophages by 3,-interferon. J. lmmunol. Methods. 82:161- 167; 1985.
18. Mishell, R. I.; Mishell, B. B. Use of Sephadex G-10 to separate macrophages and lympho- cytes. In: di Sabato, G.; Langone, J. J.; van Vunakis, H., eds. lmmunochemical tech- niques. Part G. Separation and characteriza- tion of lymphoid cells. London: Academic Press; 1984:p. 303-307.
19. Yasutake, W. T.; Wales, J. H. Microscopic anatomy of salmonids: an atlas. Washington, DC, United States Department of the Interior Fish and Wildlife Service Resource Publica- tion 150; 1983.
20. Rowley, A. E; Hunt, T. C.; Page, M.; Main- waring, G. Fish. In: Rowley, A. E; Ratcliffe, N. A., eds. Vertebrate blood cells. Cambridge: Cambridge University Press; 1988:p. 19-127.
21. Unanue, E. R.; Belier, D. I.; Yu, C. Y.; Allen, P. M. Antigen presentation: comments on its regulation and mechanisms. J. Immunol. 132: 1-5; 1984.
22. Smith, P. D.; Braun-Nesje, R. Cell-mediated immunity in the salmon: lymphocyte and mac- rophage stimulation, lymphocyte/macrophage interactions, and the production of lympho- kine-like factors by stimulated lymphocytes. Dev. Comp. lmmunol. Supplement 2:233-238; 1982.
23. Clem, L. W.; Sizemore, R .C . ; EIIsaesser, C. E; Miller, N. W. Monocytes as accessory cells in fish immune responses. Dev. Comp. Immunol. 9:803-809; 1985.
24. Ellsaesser, C. E; Bly, J. E.; Clem, L. W. Phy- logeny of lymphocyte heterogenei ty : the thymus of the channel catfish. Dev. Comp. Immunol. 12:787-799; 1988.


















