
Cell–Cell Interaction with APC, not IL-23, Is Required for
8
The Journal of Immunology Cell–Cell Interaction with APC, not IL-23, Is Required for Naive CD4 Cells To Acquire Pathogenicity during Th17 Lineage Commitment Guangpu Shi,* Jenna D. Lovaas,* Cuiyan Tan,* Barbara P. Vistica,* Eric F. Wawrousek, ‡ Mehak K. Aziz,* Rachael C. Rigden,* Rachel R. Caspi,* and Igal Gery* Subpopulations of pathogenic or nonpathogenic Th17 cells were reported to develop when presensitized CD4 cells were activated with their target Ag during polarization by either IL-23 or IL-6 and TGF- b, respectively. In this study, we generated two Th17 subpopulations by using a system in which naive CD4 cells from TCR transgenic mice specific to hen egg lysozyme (HEL) are polarized with IL-6/TGF-b and, concurrently, are activated either with HEL presented by APCs, or with anti-CD3/CD28 Abs. Only the former cells were pathogenic, inducing inflammation in eyes expressing HEL. Naive CD4 cells activated by the anti- CD3/CD28 Abs acquired pathogenicity, however, when cocultured with HEL/APC. Importantly, the naive CD4 cells did not acquire pathogenicity when cocultured with APCs stimulated with LPS or when separated from the HEL-presenting cells by a semipermeable membrane. Unlike with presensitized Th17, soluble IL-23 does not participate in pathogenicity acquisition by naive CD4 cells; no pathogenicity was induced by adding IL-23 to cultures activated with anti-CD3/CD28 Abs. Furthermore, Abs against IL-23 or IL-23R did not inhibit acquisition of pathogenicity in cultures of naive CD4 cells activated by HEL/APC. Our data thus show that, unlike presensitized CD4 cells, naive CD4 cells polarized toward Th17 phenotype acquire pathogenicity only by dir ect int eraction with APCs pre senting the Ag, with no appar ent invol vement of soluble IL- 23. We sugges t tha t the Th17 lymphocytes derived from naive CD4 cells participate in pathogenic and other immune processes, along with the IL-23– dependent Th17 cells. The Journal of Immunology, 2012, 189: 000–000. N umero us studies in recent years have estab lish ed the importance of the Th17 population in the immune sys- tem. Th1 7 cells are cru cial in the protec tio n aga ins t bacteria, parasites, and fungi, in particular in the gut and lungs, and they have a critical role in the majority of immune-mediated in- flammatory conditions in humans and experimental animals (1–5). The process of Th17 generation had been controversial in early studies, in particular with regard to the role of IL-23 in the process (2, 6). More recent studies have clarified the issue, however, by providing evidence to show that Th17 cells are normally generated by the pol arizin g act ivity of two cyt oki nes, IL-6 and TGF-b, whereas IL-23 is crucial for the maturity and pathogenic effec- tiveness of these cells (4, 7). In addition, IL-1 and IL-21 contribute to a vigorous generation of Th17 cells (8, 9). Unlike the phenotype stability of Th1, Th17 cells were found to exhibit a high level of plasticity, readily acquiring non-Th17 pheno types when exposed to othe r cyto kine environ ment s (10– 12). Thus, cultu ring Th17 cells in medi a conta inin g polar izin g cytokines specific for Th1 or Treg resulted in the majority of Th17 expressing the corresponding new phenotype within 2–3 d. Im- portantly, the acquisition of new phenotypes by Th17 cells was also observed in vivo (10, 13). Another unique feature of the Th17 population is its hetero- geneity; Th17 lines generated by different procedures were found to exhibit different immunological features (14). Ghoreschi et al. (15) reported on a Th17 subpopulation generated by polarization of naive precursors by IL-6, IL-23, and IL-1b, but without TGF-b. These Th17 cells were pathogenic and capable of inducing EAE. More recently, Kim et al. (16) discovered the existence of a sub- population of “natural Th17,” which are generated in the mouse thymus, before Ag exposure. Of particular interest to the current study is the early observation made by McGeachy et al. (17) of two Th17 subsets, one highly pathogenic and the other completely lacking this capa city. The two subsets ori ginated from pre- sensitized T cells, reactivated in culture with the Ag in the pres- ence of either IL-23, or IL-6 and TGF- b. Only the previous subset dev elope d path ogeni city , moni tored by their capac ity to induc e immune-medi ated inflammati on. We hav e prev ious ly repor ted on the activit ies of Th17 cells generated from naive CD4 cells with transgenic (Tg) TCR specific to hen egg lysozyme (HEL). When activated with HEL, presented by APCs, in the presence of IL-6 and TGF-b, these Th17 cells induced ocular inflammation in recipient mice expressing HEL in their lens (10, 18, 19). In this study, we analyzed the phenotype of these pathogenic cells and compared it with that of another Th17 subp opula tion , whic h is not pathogenic, that we gener ated by activating the naive CD4 cells with anti-CD3/CD28 Abs, along with the same polarizing cytokines, IL-6 and TGF-b. Our data indicate that the major factor determining pathogenicity of these Th17 cells, or lack thereof, is the availability of cell-cell contact with APCs presenting HEL, rather than soluble IL-23. *Laboratory of Immunology, National Eye Institute, National Institutes of Health, Bethesda, MD 20892; and ‡ Labora tory of Molec ular and Devel opmen tal Biolog y, Nation al Eye Institute, National Institutes of Health , Bethe sda, MD 20892 Received for publication October 21, 2011. Accepted for publication May 25, 2012. This work was supported by the Intramural Research Program of the National Eye Institute, National Institutes of Health. Address correspondence and reprint requests to Dr. Igal Gery, Laboratory of Immu- nology , Nation al Eye Institute, National Institute s of Health , Buildi ng 10, Room 10N208, Bethesda, MD 20892-1857. E-mail address: [email protected] The online versio n of this article contains supplementa l materi al. Abbreviations used in this article: DC, dendritic cell; HEL, hen egg lysozyme; HEL/ APC, HEL presented by APC; PbAb, plate-bound anti-CD3/CD28 Ab; qPCR, quan- titative PCR; Tg, transgenic. www.jimmunol.org/cgi/doi/10.4049/jimmunol.1103033 Published June 27, 2012, doi:10.4049/jimmunol.1103 033
-
Upload
neto-pacifico-neto -
Category
Documents
-
view
216 -
download
0
Transcript of Cell–Cell Interaction with APC, not IL-23, Is Required for

7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 1/8
The Journal of Immunology
Cell–Cell Interaction with APC, not IL-23, Is Required for
Naive CD4 Cells To Acquire Pathogenicity during Th17
Lineage Commitment
Guangpu Shi,* Jenna D. Lovaas,* Cuiyan Tan,* Barbara P. Vistica,* Eric F. Wawrousek,‡
Mehak K. Aziz,* Rachael C. Rigden,* Rachel R. Caspi,* and Igal Gery*
Subpopulations of pathogenic or nonpathogenic Th17 cells were reported to develop when presensitized CD4 cells were activated
with their target Ag during polarization by either IL-23 or IL-6 and TGF-b, respectively. In this study, we generated two Th17
subpopulations by using a system in which naive CD4 cells from TCR transgenic mice specific to hen egg lysozyme (HEL) are
polarized with IL-6/TGF-b and, concurrently, are activated either with HEL presented by APCs, or with anti-CD3/CD28 Abs.
Only the former cells were pathogenic, inducing inflammation in eyes expressing HEL. Naive CD4 cells activated by the anti-
CD3/CD28 Abs acquired pathogenicity, however, when cocultured with HEL/APC. Importantly, the naive CD4 cells did not
acquire pathogenicity when cocultured with APCs stimulated with LPS or when separated from the HEL-presenting cells by
a semipermeable membrane. Unlike with presensitized Th17, soluble IL-23 does not participate in pathogenicity acquisition by
naive CD4 cells; no pathogenicity was induced by adding IL-23 to cultures activated with anti-CD3/CD28 Abs. Furthermore, Abs
against IL-23 or IL-23R did not inhibit acquisition of pathogenicity in cultures of naive CD4 cells activated by HEL/APC. Our
data thus show that, unlike presensitized CD4 cells, naive CD4 cells polarized toward Th17 phenotype acquire pathogenicity only
by direct interaction with APCs presenting the Ag, with no apparent involvement of soluble IL-23. We suggest that the
Th17 lymphocytes derived from naive CD4 cells participate in pathogenic and other immune processes, along with the IL-23–
dependent Th17 cells. The Journal of Immunology, 2012, 189: 000–000.
Numerous studies in recent years have established the
importance of the Th17 population in the immune sys-
tem. Th17 cells are crucial in the protection against
bacteria, parasites, and fungi, in particular in the gut and lungs, and
they have a critical role in the majority of immune-mediated in-
flammatory conditions in humans and experimental animals (1–5).The process of Th17 generation had been controversial in early
studies, in particular with regard to the role of IL-23 in the process
(2, 6). More recent studies have clarified the issue, however, by
providing evidence to show that Th17 cells are normally generated
by the polarizing activity of two cytokines, IL-6 and TGF-b,
whereas IL-23 is crucial for the maturity and pathogenic effec-
tiveness of these cells (4, 7). In addition, IL-1 and IL-21 contribute
to a vigorous generation of Th17 cells (8, 9).
Unlike the phenotype stability of Th1, Th17 cells were found
to exhibit a high level of plasticity, readily acquiring non-Th17
phenotypes when exposed to other cytokine environments (10–
12). Thus, culturing Th17 cells in media containing polarizing
cytokines specific for Th1 or Treg resulted in the majority of Th17
expressing the corresponding new phenotype within 2–3 d. Im-
portantly, the acquisition of new phenotypes by Th17 cells was
also observed in vivo (10, 13).
Another unique feature of the Th17 population is its hetero-
geneity; Th17 lines generated by different procedures were found
to exhibit different immunological features (14). Ghoreschi et al.(15) reported on a Th17 subpopulation generated by polarization
of naive precursors by IL-6, IL-23, and IL-1b, but without TGF-b.
These Th17 cells were pathogenic and capable of inducing EAE.
More recently, Kim et al. (16) discovered the existence of a sub-
population of “natural Th17,” which are generated in the mouse
thymus, before Ag exposure. Of particular interest to the current
study is the early observation made by McGeachy et al. (17) of
two Th17 subsets, one highly pathogenic and the other completely
lacking this capacity. The two subsets originated from pre-
sensitized T cells, reactivated in culture with the Ag in the pres-
ence of either IL-23, or IL-6 and TGF-b. Only the previous subset
developed pathogenicity, monitored by their capacity to induce
immune-mediated inflammation.We have previously reported on the activities of Th17 cells
generated from naive CD4 cells with transgenic (Tg) TCR specific
to hen egg lysozyme (HEL). When activated with HEL, presented
by APCs, in the presence of IL-6 and TGF-b, these Th17 cells
induced ocular inflammation in recipient mice expressing HEL in
their lens (10, 18, 19). In this study, we analyzed the phenotype of
these pathogenic cells and compared it with that of another Th17
subpopulation, which is not pathogenic, that we generated by
activating the naive CD4 cells with anti-CD3/CD28 Abs, along
with the same polarizing cytokines, IL-6 and TGF-b. Our data
indicate that the major factor determining pathogenicity of these
Th17 cells, or lack thereof, is the availability of cell-cell contact
with APCs presenting HEL, rather than soluble IL-23.
*Laboratory of Immunology, National Eye Institute, National Institutes of Health,Bethesda, MD 20892; and ‡Laboratory of Molecular and Developmental Biology,National Eye Institute, National Institutes of Health, Bethesda, MD 20892
Received for publication October 21, 2011. Accepted for publication May 25, 2012.
This work was supported by the Intramural Research Program of the National EyeInstitute, National Institutes of Health.
Address correspondence and reprint requests to Dr. Igal Gery, Laboratory of Immu-nology, National Eye Institute, National Institutes of Health, Building 10, Room10N208, Bethesda, MD 20892-1857. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: DC, dendritic cell; HEL, hen egg lysozyme; HEL/ APC, HEL presented by APC; PbAb, plate-bound anti-CD3/CD28 Ab; qPCR, quan-titative PCR; Tg, transgenic.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1103033
blished June 27, 2012, doi:10.4049/jimmunol.1103033
7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 2/8
Materials and Methods Mice
All studies were performed with (B10.BR3 FVB/N) F1 mice, generated asdetailed elsewhere (10, 18). The lines used in this study included Tg miceexpressing HEL in their lens (HEL-Tg), HEL-specific TCR on the majorityof their T lymphocytes (designated 3A9) (10, 18), or syngeneic wild typemice. HEL is expressed exclusively in eyes of the HEL-Tg mice; this neo-self Ag cannot be detected in the circulation or any other tissue of thesemice (20). All manipulations were performed in compliance with the
National Institutes of Health Resolution on the Use of Animals in Re-search.
Generation of Th17 lines
Naive T cells were enriched from splenocytes and lymph node cells of 3A9mice, using T cell columns (R&D Systems). CD4 cells expressing the TgTCR were then sorted by FACSAria II (BD Biosciences), using the clo-notypic mAb 1G12 (10, 18). The naive CD4 cells were incubated for 4 d in12-well plates, at 0.5 3 106 in 2 ml of Th17 polarizing medium (10, 18).The polarizing cytokines IL-6 and TGF-b were added at 100 and 10 ng/ml,respectively, similar to the study by McGeachy et al. (17). Activation of thenaive CD4 cells, concurrently with the polarization, was performed byHEL (at 2 mg/ml, presented by irradiated wild type splenocytes as indi-cated by “HEL/APC”), or by spleen dendritic cells (as indicated by “HEL/ DC”), or by anti-CD3/CD28 Abs bound to the plate walls (PbAb) by in-
cubation for 1 h (2 mg/well) (21).In certain experiments, additional agents and Abs were added to theculture medium, alone or in combination, as indicated. These included: IL-23(eBioscience), 20 ng/ml onday 1 or2 and at10 ng/ml onday 3 ofculture;LPS (100 ng/ml; Difco); anti–IL-23–p40 Ab (50 mg/ml; a gift from GiorgioTrinchieri, National Cancer Institute, National Institutes of Health); oranti–IL-23R Ab (10 mg/ml; R&D Systems).
To generate Th1 lines, naive CD4 cells were incubated for 4 d in 12-wellplates, at 0.5 3 106 in 2 ml of Th1 polarizing medium, as detailed else-where (10, 18).
DC isolation
DCs were isolated from naive syngeneic wild type mouse spleens, using themethod described by Tang et al. (22). Spleens were minced into smallfragments and digested with collagenase D (Roche) and DNase I (Sigma-Aldrich) for 45 min at 37˚C. Cells were collected after passing through
a 70-mM nylon cell strainer (BD Falcon) and treated with EDTA for 5 min.Following red cell lysis and Fc receptor blocking, cells were incubatedwith anti–CD11c-conjugated magnetic beads (Miltenyi Biotec) at 4˚C for15 min. The positive population was purified using autoMACS (MiltenyiBiotec). The isolated preparation consisted of ∼90% cells positive forCD11c.
HEL/APC and PbAb Th17 cells generated in transwell cultures
For HEL/APC and PbAb Th17 generated in transwell cultures, the sortedCD4 cells (0.5 3 106) were cultured with APCs at the top compartment ina volume of 0.5 ml containing HEL and Th17 polarizing mixture (10, 18,19), while the same number of sorted CD4 cells in a volume of 1.5 mlcontaining HEL and the polarizing mixture, but with no APCs, werecultured in the bottom compartments that were coated with anti-CD3/ CD28 Abs, as described above. Th1 cells were similarly cultured for
comparison. The system is also demonstrated in Fig. 6 A.
Quantitative PCR analysis
Total RNA was extracted from cultured cells with TRIzol (Invitrogen-LifeTechnologies). RNA (5 mg), SuperScript III Reverse Transcriptase (Invi-trogen Life Technologies), and oligo(dT)12–16 were used for first-strandcDNA synthesis. Primer-probe sets for quantitative PCR (qPCR) quanti-fying the expression of mouse IL-23R, CD40L, ICOS, AHR, CCL2, CCL5,CCL20, CCL22, CXCL2, CXCL10, CCR2, CCR6 , CXCR3 and GAPDH orb-actin (internal control) were purchased from Applied Biosystems. Pri-mers used for ROR-gt and ROR-a were as follows: ROR-gt, 59-CC-GCTGAGAGGGCTTCAC-39 and 59-TGCAGGAGTAGGCCACATTACA-39; and ROR-a, 59-CGTGTCCATGGCAGAACTAGAA-39 and 59-GCAAGTACTGGCAGGTTTCCA-39. Fluorescence-labeled probes usedare: ROR-gt, 59-AAGGGCTTCTTCCGCCGCAGCCAGCAG-BHQ-1–39;
and ROR-a, 59-CCTTGCCCAGAACATATCCAAATCCCA-BHQ-1–39.PCR parameters were as recommended for the TaqMan Universal PCRMaster Mix kit (Applied Biosystems).
Measurement of cytokines released into Th17 cell culture
supernatant
Cell culture supernatants were sampled on days 2, 3, or 4 of culture. Thesupernatants were tested for the levels of IL-10, IL-17, IL-22, and GM-CSF,using ELISA kits from R&D System, and for the levels of IL-21 and IL-17F, using ELISA kits from BioLegend, according to the manufacturers’instructions.
Flow cytometric analysis of surface, nuclear, and intracellular
moleculesConventional methods were used for analysis of surface molecule ex-pression.
For Foxp3 staining, Th17 cells were fixed and permeabilized with thefixation or permeabilization buffer for 1 h at 4˚C before intracellularstaining with allophycocyanin-conjugated anti-Foxp3, following the pro-cedure recommended by the manufacturer (eBioscience).
For intracellular cytokine staining, polarized Th17 cell cultures werestimulated with 20 ng/ml PMA and 1 mM ionomycin (Sigma-Aldrich), plusGolgi-Stop (BD Biosciences), for 5 h. The stimulated cells were thenstained with the corresponding Abs for surface and intracellular molecules.Data were acquired using a FACSCalibur (BD Biosciences) and analyzedusing FlowJo (Tree Star).
Adoptive transfer of Th17 line cells
After 4 d of incubation, PbAb or HEL/APC Th17 cells were injected (at 43106) via the tail vein, into groups of naive HEL-Tg mice. Recipient eyeswere collected on day 5 after cell transfer and processed for histologic
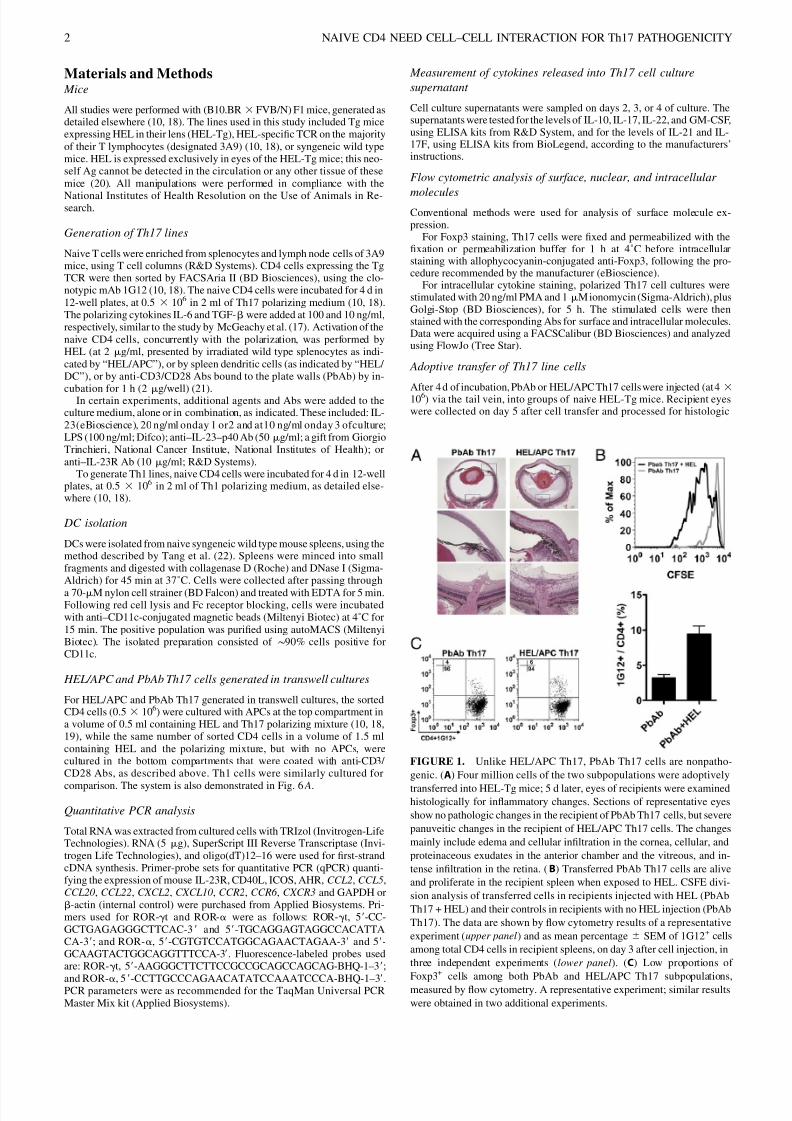
FIGURE 1. Unlike HEL/APC Th17, PbAb Th17 cells are nonpatho-
genic. (A) Four million cells of the two subpopulations were adoptively
transferred into HEL-Tg mice; 5 d later, eyes of recipients were examinedhistologically for inflammatory changes. Sections of representative eyes
show no pathologic changes in the recipient of PbAb Th17 cells, but severe
panuveitic changes in the recipient of HEL/APC Th17 cells. The changes
mainly include edema and cellular infiltration in the cornea, cellular, and
proteinaceous exudates in the anterior chamber and the vitreous, and in-
tense infiltration in the retina. (B) Transferred PbAb Th17 cells are alive
and proliferate in the recipient spleen when exposed to HEL. CSFE divi-
sion analysis of transferred cells in recipients injected with HEL (PbAb
Th17 + HEL) and their controls in recipients with no HEL injection (PbAb
Th17). The data are shown by flow cytometry results of a representative
experiment (upper panel) and as mean percentage 6 SEM of 1G12+ cells
among total CD4 cells in recipient spleens, on day 3 after cell injection, in
three independent experiments (lower panel). (C) Low proportions of
Foxp3+ cells among both PbAb and HEL/APC Th17 subpopulations,
measured by flow cytometry. A representative experiment; similar results
were obtained in two additional experiments.
2 NAIVE CD4 NEED CELL–CELL INTERACTION FOR Th17 PATHOGENICITY

7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 3/8
analysis by conventional H&E staining. Severity of the inflammation wasscored according to histologic changes, as described elsewhere (23).
Measuring cell division rate by CFSE dilution
PbAb or HEL/APC Th17 cells (107 cells/ml) were labeled with CFSE (10mM; Molecular Probes) for adoptive transfer, as described elsewhere (19,24). CFSE-labeled cells were washed, resuspended in RPMI 1640, andinjected i.v. into recipient mice (4 3 106 cells). Recipient mice were eu-thanized on the indicated days after cell transfer, and splenocytes werecollected and stained with anti–CD4-PE. Data were acquired using a
FACSCalibur. CFSE dye dilution was analyzed using FlowJo.
Statistical analysis
Unpaired, two-tail t test was performed for comparison of transcript levelin Th17 or cytokine level in supernatant, as well as for comparison of thefrequency of transferred cells recovered from recipient spleen. A p value #0.05 was defined as statistically significant.
ResultsTh17 cells generated by activation with HEL/APC are
pathogenic, whereas those generated by activation with
anti-CD3/CD28 Abs are not
We generated two subpopulations of Th17 cells by activation of
sorted naive CD4 cells specific to HEL with either the target Ag,
HEL, presented by APCs (HEL/APC), or with plate-bound anti-CD3/CD28 Abs (PbAb), in the presence of the same polarizing
cytokines, IL-6 and TGF-b. To test for the pathogenic capacity of
the two Th17 subpopulations, we adoptively transferred the cells
into syngeneic recipients expressing HEL in their eyes (10, 18,
19). Fig. 1A depicts representative sections of eyes from recipients
injected with cells of the two subpopulations. Severe ocular in-
flammatory changes are seen in the recipient of HEL/APC Th17
cells, whereas no inflammatory changes could be detected in the
eyes of the recipient of PbAb Th17 cells.
The lack of pathogenicity by PbAb-activated Th17 cells could
be attributed to their low viability in vivo. To examine this pos-
sibility, we used the CFSE division method to test the capacity of the transferred cells to proliferate when exposed to their target Ag.
We and others (25, 26) have reported that adoptively transferred
autopathogenic T cells initially migrate to the host spleen, where
they proliferate before invading the target tissue. Fig. 1B shows
a representative experiment. Adoptively transferred CFSE-labeled
PbAb Th17 cells proliferated poorly in the spleen of recipient
mice, but vigorous proliferation was seen in recipient mice that
were also injected with HEL. This observation thus provides ev-
idence that the injected PbAb-activated cells are viable and fully
capable of responding to their target Ag. Importantly, the PbAb-
activated Th17 cells in this system retained their nonpathogen-
icity despite their reactivation by the Ag in vivo (data not shown).
The possibility that the nonpathogenic activity of PbAb-stimulated
Th17 cells was due to high levels of regulatory T cells wasruled out by the finding that the proportion of cells expressing
Foxp3 in PbAb-activated cells was similar to that of the patho-
genic HEL/APC Th17 cells (Fig. 1C).
FIGURE 2. HEL/APC and PbAb Th17 subpopulations differ in their cytokine production profiles. (A) Expression of ROR-a and ROR-gt transcripts by
the two Th17 subpopulations, after 4 d of incubation, determined by qPCR. ( B) Mean levels6 SEM of IL-17, IL-22, and IL-10 in supernatants collected at
different time points of cultures in which the two subpopulations were generated. The data were collected in four independent experiments. ( C) Immu-
nostaining for intracellular expression of IL-17, IL-22, and IL-10 of cells from the two Th17 subpopulations. The cells were collected after 4 d in culture
and stained with the specific mAbs, or isotype controls (IgG), as detailed in Shi et al. (10). Note that HEL/APC Th17, but not PbAb Th17, cells stained for
intracellular IL-22 and that different subsets within this subpopulation stained for either IL-17 or IL-22, with just a minor portion of cells coexpressing both
cytokines. Also of note is the coexpression of IL-17 and IL-10 by cells of the two Th17 subpopulations. A representative experiment; similar observations
were made in another experiment. ★ p , 0.05.
The Journal of Immunology 3

7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 4/8
Molecular analysis of the two Th17 subpopulations
To define the two Th17 subpopulations molecularly, we compared
their capacity to produce several molecules characteristic for the
Th17 lineage. Both subpopulations expressed transcripts of the
two Th17 specific transcription factors, ROR-a and ROR-gt, with
higher levels seen in the PbAb cultures (Fig. 2A).
Next, we analyzed the cytokine production by the two sub-
populations, by measuring in the culture supernatants the levels of
three Th17 products—IL-17, IL-10, and IL-22—at different timepoints (Fig. 2B). PbAb cultures yielding nonpathogenic Th17
produced exceedingly high levels of IL-17 that were notably
higher than those produced by HEL/APC cultures, which generate
the pathogenic Th17. The two types of cultures produced similar
or only moderately different levels of IL-10, but striking differ-
ences were seen in the pattern of IL-22 production between the
cultures activated by PbAb or HEL/APC, with the latter cultures
producing IL-22 levels higher by two orders of magnitude. The
vigorous production of IL-22 by CD4 cells stimulated by HEL/
APC was not affected by the addition of Ab against IL-23R, or
against 12p40, but was moderately inhibited by the addition of
PbAb (Supplemental Fig. 1).
Production of the three cytokines by Th17 cells was also directlydemonstrated by flow cytometry. The data of a representative ex-
periment are recorded in Fig. 2C and show that the intracellular
staining patterns of the tested cytokines are in accord with our
ELISA data. Of interest are the findings that (1) IL-10 was
expressed by the same subset of CD4 cells that produced IL-17,
which is in line with our finding (Fig. 2B) that pathogenic Th17
cells in our study also secrete IL-10, and (2) in contrast toIL-10, IL-
22 production in cultures activated by HEL/APC was localized to
a large extent in a subset of CD4 cells that did not coexpress IL-17.
The subpopulations of pathogenic and nonpathogenic Th17
also differed in their staining for certain surface Ags. Differences
were particularly clear in the expression of CCR5, CCR6, and
a4b7 (Fig. 3A). More dramatic differences between the two sub-
populations were found in their expression profiles of several
chemokines and chemokine receptors (Fig. 3B). Whereas tran-
scripts of CCL20 and CXCR3 were more elevated in PbAb cul-
tures, those of CXCL10, CCL2, CCL22, and CXCL2 were higher
in HEL/DC cultures.
Th17 cells generated by stimulation with HEL/APC or with
PbAb were also tested for production of other molecules: cells of
the two subpopulations expressed similar levels of CD40L tran-
scripts, but HEL/APC Th17 expressed higher levels of ICOStranscript, whereas moderately higher levels of AHR transcript
were expressed by PbAb Th17 cells (Supplemental Fig. 2A).
Supernatants of these cultures contained similar levels of GM-CSF
and IL-21, but PbAb-stimulated cultures were superior in their
production of IL-17F (Supplemental Fig. 2B), in line with their
higher levels of IL-17A (Fig. 2B).
Th17 generated by stimulation with either PbAb or HEL/APC
proliferated vigorously, but a marked difference was noted in
the kinetics of their response, with PbAb-stimulated Th17 cells
responding earlier (Supplemental Fig. 3). This difference is in
accord with the two mechanisms of stimulation process: the Abs
directly stimulate the naive CD4 cells, whereas stimulation with
HEL/APC requires prior processing of the Ag by the APC.
The two Th17 subpopulations differ in their homing to and
proliferation in the recipient mouse spleen
As mentioned above, adoptively transferred activated T cells mi-
grate to the recipient spleen, where they proliferate before invading
the target organ (25, 26). To compare between the two Th17 sub-
populations for this capacity, we adoptively transferred the tested
cells into HEL-Tg mice and tracked them in the recipient spleen by
the clonotypic Ab, 1G12. Fig. 4A depicts data of a representative
experiment in which 1G12+ cells were identified by flow cytometry
in recipient spleens 4 d after cell transfer; the figure shows that
Th17 generated by activation with HEL/APC were superior to Th17
generated by PbAb activation in their capacity to accumulate and
FIGURE 3. The two Th17 subpopulations differ in their expression of surface molecules and certain chemokine and chemokine receptor transcripts. (A)
Suspensions of PbAb and HEL/APC Th17 cells, harvested after 4 d in culture, were stained for the indicated surface molecules and examined by flow
cytometry. (B) qPCR analysis of Th17 cells collected after 4 d of incubation at the HEL/DC or PbAb conditions. Transcript expression is presented relative
to GAPDH. A representative experiment is shown; similar expression profiles were observed in four other independent experiments.
4 NAIVE CD4 NEED CELL–CELL INTERACTION FOR Th17 PATHOGENICITY

7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 5/8
proliferate in the recipient spleen. Repeated experiments in which
recipient spleens were collected on days 2, 4, and 7 after cell
transfer are summarized in Fig. 4B, showing the superiority of the
HEL/APC Th17 at all tested time points.
Next, we used the CFSE dilution procedure to compare the two
subpopulations of Th17 cells for their proliferation rate in the
recipient spleen. As seen in Fig. 4C, the HEL/APC Th17 prolif-
erated considerably more vigorously than the nonpathogenic
PbAb cells, a finding in line with their higher proportion among
the total spleen CD4 cells (Fig. 4A, 4B).
We have reported previously (10) that Th17 cells generated by
activation with HEL/APC exhibit plasticity and acquire the Th1
phenotype when exposed to other cytokine environments. To
compare the two Th17 subpopulations for their plasticity, we
collected recipient spleen cells on day 4 after cell injection and
analyzed them for intracellular expression of IL-17 and IFN-g. As
shown in Fig. 4D, the proportion of cells expressing IFN-g washigher among the HEL/APC recipients than among the PbAb
recipients.
Naive CD4 cells activated by plate-bound Abs acquire
pathogenicity when cocultured with DC and HEL
Soluble IL-23 was shown to be critical for acquisition of patho-
genicity by CD4 cells preimmunized against the immunopatho-
genic Ag (7, 17). No pathogenicity was acquired, however, in our
system by naive CD4 activated in culture by the plate-bound Abs
when IL-23 was added to the medium during Th17 polarization,
on day 1 (Fig. 5A, upper panel) or even on days 2 and 3 (not
shown), when the expression of IL-23R was elevated (Fig. 5B). In
contrast, PbAb-activated CD4 cells acquired pathogenic capacity
when DCs and HEL were also added to the culture medium; theseTh17 cells induced inflammation levels similar to those achieved
by Th17 cells of cultures stimulated by DCs and HEL (Fig. 5A,
upper panel). Importantly, however, no pathogenicity was ac-
quired in cultures in which DCs were added alone or with LPS,
a potent APC stimulant (27–29) (Fig. 5A, upper panel). All cul-
tures in these experiments were polarized with IL-6 and TGF-b
during the activation process and, as shown in the lower panel of
Fig. 5A, Th17 cells generated in all these cultures released high
levels of IL-17, despite the differences in their immunopathoge-
nicity, but in agreement with data shown in Fig. 2B. It is of in-
terest, however, that the lowest IL-17 levels were released by the
pathogenic cultures activated by only DCs and HEL, again in
agreement with the data recorded in Fig. 2B, underscoring the IL-17–stimulating capacity of the plate-bound Abs.
It is also noteworthy that the addition of Abs against IL-23 or IL-
23R had no effect on acquisition of pathogenicity by CD4 cells
activated by HEL/DC, or by plate-bound Abs and HEL/DC (data
not shown).
Cell-cell interaction between naive CD4 and APCs is essential
for acquisition of pathogenicity by Th17 cells
APCs affect T cells by the release of cytokines and by cell-cell
interaction (30, 31). To analyze the mechanism whereby APCs
induce pathogenicity in Th17 in our experimental system, we
separated by semipermeable membranes the HEL-presenting cells
and the naive CD4 cells activated with anti-CD3/CD28 Abs (Fig.6A) and examined the development of pathogenicity by the latter
cells. Naive CD4 cells were also added to the top compartment,
along with the APCs. Th17 polarizing cytokines (IL-6 and TGF-b)
and HEL were added to both compartments. Data collected in
repeated experiments are summarized in Fig. 6B, whereas Fig. 6D
shows representative eye sections of recipient mice injected with
Th17 cells collected from the two compartments. As expected,
the CD4 lymphocytes from the top compartment, cultured with
HEL/APC, developed pathogenic capacity. In contrast, no patho-
genicity was demonstrated by the CD4 cells from the bottom
compartment, which were activated by anti-CD3/CD28 Abs.
These observations thus provide evidence showing that acquisi-
tion of pathogenicity by naive CD4 cells during polarization to-
ward Th17 phenotype requires cell-cell interaction with APCs,
rather than soluble cytokines released by APCs.
FIGURE 4. HEL/APC Th17 are superior to PbAb Th17 cells in their
capacity to migrate to and proliferate in the recipient spleen. Four million
Th17 cells of the two subpopulations were adoptively transferred into
HEL-Tg mice, and spleens of the recipients were collected at the indicated
time points. (A) Flow cytometric analysis on day 4, identifying donor cells,
positive for the clonotypic Ab, 1G12, among the CD4 population. The
proportion of donor cells is higher in the recipients injected with HEL/
APC Th17 than in that of the PbAb Th17 recipients. A representative
experiment is shown; similar data were obtained in two other experiments.
(B) A summary of three experiments, depicting the mean percentage 6
SEM of PbAb or HEL/APC Th17 cells (1G12+) among CD4 cells in the
recipient spleens, on days 2, 4, or 7 after cell injection. (C) Proliferation
rate of the two subpopulations determined by the CFSE dilution method, at
the indicated time points. Th17 cells of the HEL/APC subpopulation
proliferated at a faster rate. (D) Th17 generated by HEL/APC activation
switch to the Th1 phenotype in the recipient spleen more readily than
PbAb-generated Th17. Intracellular flow cytometric analysis shows a
higher proportion of the former subpopulation expressing IFN-g on day 4
after cell injection. ★ p , 0.05.
The Journal of Immunology 5

7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 6/8
To further examine the observations we made with Th17 sub-
populations using the transwell system, we performed similar
experiments in which Th1 cell subpopulations were generated and
tested for pathogenicity (Fig. 6C, 6E). Th1 cells were generated bypolarization with IL-12 (10, 18). Unlike with Th17 cells, Th1 cells
generated by activation with PbAb during polarization acquired
pathogenicity. As shown in Fig. 6C and 6E, Th1 collected from
the bottom compartment (activated by PbAb) induced inflamma-
tion in recipient eyes, similar to that induce by Th1 cells collected
from the top compartment (activated by HEL/APC). These
observations thus underscore the profound difference between
Th17 and Th1 populations in their acquisition of pathogenicity.
Activation of naive CD4 cells with PbAb during polarization
elicits pathogenic capacity in Th1 but not in Th17 cells.
Discussion
As cited above, recent publications revealed the heterogenicity of the Th17 population, with different subpopulations being generated
by different procedures (14, 15, 17). Data collected in the current
study further extend the information concerning the variety of
Th17 subpopulations. These data show that activating naive CD4cells with either the Ag presented by APCs, or with Abs against
CD3/CD28, during polarization with IL-6 and TGF-b, generate
two Th17 subpopulations that differ by their immunopathogenic
capacity: the former subpopulation is pathogenic, whereas the
latter is not. It is of note that the pathogenic and nonpathogenic
subpopulations in our study differed from the analogous two Th17
subpopulations described by McGeachy et al. (17) in their cell
source (i.e., naive versus preimmunized cells) as well as the ac-
tivation and polarization procedures used for their generation. In
addition, the pathogenic Th17 lines of the two studies also differed
in their expression of IL-10; pathogenic cells in the study by
McGeachy et al. (17) did not produce this cytokine, whereas the
pathogenic cells in our study did, as shown by both their releaseand intracellular production of this cytokine (Fig. 2B, 2C).
FIGURE 5. Naive CD4 cells activated by PbAb acquire pathogenicity
when DCs and HEL are also added to the culture medium. (A) Top frame,
severity levels of ocular inflammation induced in recipient mice expressingHEL in their eyes, following adoptive transfer of 4 million Th17 cells,
generated as indicated. The dots represent individual eyes, scored as de-
tailed in Kim et al. (23). Bottom panel, Levels of IL-17 secreted by the
different Th17 cultures, generated as indicated, following incubation for
4 d. (B) Expression levels of IL-23R transcripts by activated naive CD4
cells, at different time points. PbAb-stimulated naive CD4 cells, under
Th17 polarizing conditions, were collected at the indicated time points and
assayed by qPCR. Freshly isolated naive CD4 cells were used as control
for the assay.
FIGURE 6. Cell-cell interaction is required for acquisition of pathoge-
nicity by Th17 cells, but not by Th1 cells. Transwell cultures (A) were
established with HEL-specific naive CD4 cells activated with either APCs
in the top compartment or anti-CD3/CD28 Abs in the bottom compart-
ment, along with HEL and polarizing cytokines for either Th17 or Th1 inboth compartments. Following 4 d of incubation, the cells collected from
each compartment were injected into HEL-Tg recipients. (B and C)
Summaries of three repeated experiments, recording the severity levels of
individual eyes of recipient mice injected with Th17 or Th1 cells, collected
from the top or bottom compartments and scored as detailed in Kim et al.
(23). (D and E) Sections of the anterior and posterior eye segments of
representative recipient eyes collected 5 d after cell transfer. Severe
changes were induced by Th1 collected from both culture compartments
(E), but only from the top compartment of Th17 cultures (D). C, Cornea; I,
iris; L, lens; O, optic nerve; R, retina; V, vitreous.
6 NAIVE CD4 NEED CELL–CELL INTERACTION FOR Th17 PATHOGENICITY

7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 7/8
The two subpopulations in our study also differed in their
production of IL-22, a characteristic product of Th17 cells (32):
only cells of the pathogenic Th17 subpopulation produced this
cytokine. Interestingly, flow cytometric analysis revealed that the
majority of cells that express IL-22 do not coproduce IL-17 (Fig.
2C). More studies are needed to further analyze the biological
function of the subset of IL-22–producing Th17 cells and their
possible relationship to the Th22 subpopulation recently identified
in humans (33, 34). The capacity to produce IL-22 is not related topathogenicity of Th17 cells, because treatment of recipients of
pathogenic Th17 with anti–IL-22 Ab had no effect on the devel-
opment of ocular inflammation in these mice (data not shown).
The two subpopulations in our study differed in their profiles of
cell surface Ags, chemokines, and chemokine receptors, and it is
conceivable that these differences determined their pathogenic
capacity to a large extent. This notion is also supported by the
finding that the profiles of chemokines and chemokine receptors of
the pathogenic and nonpathogenic Th17 subpopulations generated
in our study (Fig. 3B) resembled those of the analogous sub-
populations generated by McGeachy et al. (17), despite the re-
markable differences in their cell origin, mode of generation, and
lymphokine profiles.
Our study shows that pathogenic Th17 cells are superior to thenonpathogenic cells in their capacity to migrate into and proliferate
in the recipient spleen. The migration was monitored by tracking
the injected cells with the clonotypic Ab 1G12, and their prolif-
eration rate was measured by the CFSE dilution method (Fig. 4A–
C). It is conceivable that these cellular capacities are essential for
the pathogenic Th17 cells in their induction of inflammation in the
target organ, the eye in the current study. Our study thus provides
a new parameter for the capacity of Th populations to migrate into
and proliferate in the recipient organs.
In accord with our previous study (10), a portion of Th17 cells
generated by HEL/APC activation acquire the Th1 phenotype in
the recipient spleen (Fig. 4D). Lower proportions of Th17 cells
generated by PbAb activation also exhibited phenotype switching,but it seems unlikely this lower activity of the PbAb Th17 cells
contributed significantly to their lack of pathogenicity.
Importantly, data collected in the current study show that, un-
like presensitized CD4 lymphocytes (17), naive CD4 cells are not
affected by soluble IL-23 during their polarization toward the
pathogenic Th17 phenotype. Addition of soluble IL-23 to cultures
stimulated with anti-CD3/CD28 Abs had no effect on the gener-
ation of nonpathogenic phenotype, whereas addition of APCs and
HEL to these cultures yielded pathogenic Th17 (Fig. 5A). Fur-
thermore, the generation of pathogenic Th17 from naive cells
could not be blocked in culture by Abs against IL-23 or IL-23R
(data not shown).
It is noteworthy that the APC effect of generating pathogenicityin naive CD4 cells in our experimental system was achieved only
when the APC presented the target Ag—HEL. No effect was seen
when the APCs were added with no addition of HEL, or even
when LPS, a potent APC stimulator (27–29), was added to the
culture system (Fig. 5A). These data suggest that naive CD4 cells
acquire pathogenicity only by cell-cell interaction with HEL-
presenting APCs, a notion that was further examined by the
transwell method. This method allowed us to test in the same
culture system activation of naive CD4 cells by either cell-cell
interaction with APC-presenting HEL, or by cytokines released
from these APCs. Pathogenicity was acquired only by the former
mode of activation (Fig. 6B).
In conclusion, the data in this study provide evidence to show
that naive CD4 cells polarized into the Th17 phenotype acquire
pathogenic capacity only by directly interacting with APCs pre-
senting the specific Ag. Our data also show that IL-23, which was
found to be crucial for development of pathogenicity by pre-
immunized CD4 cells (4, 7, 12, 17), is not essential for the process
in naive CD4 cells (Fig. 5A). These data thus show for the first
time, to our knowledge, that the process that generates Th17 from
naive CD4 is remarkably different from the process that generates
Th17 from mature preimmunized Th cells. More investigation is
needed to analyze the functions of Th17 originated from naive
cells in the total Th17-induced immune response against microbialinvasion, or the initiation of pathogenic inflammation. We suggest
that the Th17 lymphocytes derived from naive CD4 cells partic-
ipate in these immune processes along with, or ahead of, the IL-
23–dependent Th17 cells.
AcknowledgmentsWe thank Dr. Ronald H. Schwartz for helpful suggestions, Lindsey Nugent
for expert assistance, R. Steven Lee for tail DNA analysis, the National Eye
Institute FlowCytometry Corefor technical support, and the Histology Core
for tissue section preparations.
DisclosuresThe authors have no financial conflicts of interest.
References1. Bettelli, E., M. Oukka, and V. K. Kuchroo. 2007. T(H)-17 cells in the circle of
immunity and autoimmunity. Nat. Immunol. 8: 345–350.2. Weaver, C. T., R. D. Hatton, P. R. Mangan, and L. E. Harrington. 2007. IL-17
family cytokines and the expanding diversity of effector T cell lineages. Annu. Rev. Immunol. 25: 821–852.
3. Park, H., Z. Li, X. O. Yang, S. H. Chang, R. Nurieva, Y. H. Wang, Y. Wang,L. Hood, Z. Zhu, Q. Tian, and C. Dong. 2005. A distinct lineage of CD4 T cellsregulates tissue inflammation by producing interleukin 17. Nat. Immunol. 6:1133–1141.
4. Morrison, P. J., S. J. Ballantyne, and M. C. Kullberg. 2011. Interleukin-23 and Thelper 17-type responses in intestinal inflammation: from cytokines to T-cellplasticity. Immunology 133: 397–408.
5. Shainheit, M. G., K. W. Lasocki, E. Finger, B. M. Larkin, P. M. Smith,A. H. Sharpe, C. A. Dinarello, L. I. Rutitzky, and M. J. Stadecker. 2011. Thepathogenic Th17 cell response to major schistosome egg antigen is sequentiallydependent on IL-23 and IL-1b. J. Immunol. 187: 5328–5335.
6. McKenzie, B. S., R. A. Kastelein, and D. J. Cua. 2006. Understanding the IL-23-IL-17 immune pathway. Trends Immunol. 27: 17–23.
7. McGeachy, M. J., Y. Chen, C. M. Tato, A. Laurence, B. Joyce-Shaikh,W. M. Blumenschein, T. K. McClanahan, J. J. O’Shea, and D. J. Cua. 2009. Theinterleukin 23 receptor is essential for the terminal differentiation of interleukin17-producing effector T helper cells in vivo. Nat. Immunol. 10: 314–324.
8. Geri, G., B. Terrier, M. Rosenzwajg, B. Wechsler, M. Touzot, D. Seilhean,T. A. Tran, B. Bodaghi, L. Musset, V. Soumelis, et al. 2011. Critical role of IL-21in modulating TH17 and regulatory T cells i n Behcet disease. J. Allergy Clin.
Immunol. 128: 655–664.9. Sutton, C., C. Brereton, B. Keogh, K. H. Mills, and E. C. Lavelle. 2006. A
crucial role for interleukin (IL)-1 in the induction of IL-17-producing T cells thatmediate autoimmune encephalomyelitis. J. Exp. Med. 203: 1685–1691.
10. Shi, G., C. A. Cox, B. P. Vistica, C. Tan, E. F. Wawrousek, and I. Gery. 2008.Phenotype switching by inflammation-inducing polarized Th17 cells, but not byTh1 cells. J. Immunol. 181: 7205–7213.
11. Lee, Y. K., H. Turner, C. L. Maynard, J. R. Oliver, D. Chen, C. O. Elson, and
C. T. Weaver. 2009. Late developmental plasticity in the T helper 17 lineage. Immunity 30: 92–107.
12. Hirota, K., J. H. Duarte, M. Veldhoen, E. Hornsby, Y. Li, D. J. Cua, H. Ahlfors,C. Wilhelm, M. Tolaini, U. Menzel, et al. 2011. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat. Immunol. 12: 255–263.
13. Martin-Orozco, N., Y. Chung, S. H. Chang, Y. H. Wang, and C. Dong. 2009. Th17cells promote pancreatic inflammation but only induce diabetes efficiently inlymphopenic hosts after conversion into Th1 cells. Eur. J. Immunol. 39: 216–224.
14. Peters, A., Y. Lee, and V. K. Kuchroo. 2011. The many faces of Th17 cells. Curr.Opin. Immunol. 23: 702–706.
15. Ghoreschi, K., A. Laurence, X. P. Yang, C. M. Tato, M. J. McGeachy,J. E. Konkel, H. L. Ramos, L. Wei, T. S. Davidson, N. Bouladoux, et al. 2010.Generation of pathogenic T(H)17 cells in the absence of TGF-b signalling.
Nature 467: 967–971.16. Kim, J. S., J. E. Smith-Garvin, G. A. Koretzky, and M. S. Jordan. 2011. The
requirements for natural Th17 cell development are distinct from those of conventional Th17 cells. J. Exp. Med. 208: 2201–2207.
17. McGeachy, M. J., K. S. Bak-Jensen, Y. Chen, C. M. Tato, W. Blumenschein,
T. McClanahan, and D. J. Cua. 2007. TGF-b and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain T(H)-17 cell-mediated pathology. Nat.
Immunol. 8: 1390–1397.
The Journal of Immunology 7

7/31/2019 Cell–Cell Interaction with APC, not IL-23, Is Required for
http://slidepdf.com/reader/full/cellcell-interaction-with-apc-not-il-23-is-required-for 8/8
18. Cox, C. A., G. Shi, H. Yin, B. P. Vistica, E. F. Wawrousek, C. C. Chan, andI. Gery. 2008. Both Th1 and Th17 are immunopathogenic but differ in other keybiological activities. J. Immunol. 180: 7414–7422.
19. Shi, G., M. Ramaswamy, B. P. Vistica, C. A. Cox, C. Tan, E. F. Wawrousek,R. M. Siegel, and I. Gery. 2009. Unlike Th1, Th17 cells mediate sustained au-toimmune inflammation and are highly resistant to restimulation-induced celldeath. J. Immunol. 183: 7547–7556.
20. Zhang, M., M. S. Vacchio, B. P. Vistica, S. Lesage, C. E. Egwuagu, C. R. Yu,M. P. Gelderman, M. C. Kennedy, E. F. Wawrousek, and I. Gery. 2003. T celltolerance to a neo-self antigen expressed by thymic epithelial cells: the solubleform is more effective than the membrane-bound form. J. Immunol. 170: 3954–3962.
21. Huter, E. N., G. H. Stummvoll, R. J. DiPaolo, D. D. Glass, and E. M. Shevach.2009. Pre-differentiated Th1 and Th17 effector T cells in autoimmune gastritis:Ag-specific regulatory T cells are more potent suppressors than polyclonalregulatory T cells. Int. Immunopharmacol. 9: 540–545.
22. Tang, J., W. Zhu, P. B. Silver, S. B. Su, C. C. Chan, and R. R. Caspi. 2007.Autoimmune uveitis elicited with antigen-pulsed dendritic cells has a distinctclinical signature and is driven by unique effector mechanisms: initial encounterwith autoantigen defines disease phenotype. J. Immunol. 178: 5578–5587.
23. Kim, S. J., M. Zhang, B. P. Vistica, C. C. Chan, D. F. Shen, E. F. Wawrousek, andI. Gery. 2002. Induction of ocular inflammation by T-helper lymphocytes type 2.
Invest. Ophthalmol. Vis. Sci. 43: 758–765.24. Fujimoto, C., C. R. Yu, G. Shi, B. P. Vistica, E. F. Wawrousek, D. M. Klinman,
C. C. Chan, C. E. Egwuagu, and I. Gery. 2006. Pertussis toxin is superior to TLRligands in enhancing pathogenic autoimmunity, targeted at a neo-self antigen, bytriggering robust expansion of Th1 cells and their cytokine production. J.
Immunol. 177: 6896–6903.25. Chen, J., B. P. Vistica, H. Takase, D. I. Ham, R. N. Fariss, E. F. Wawrousek,
C. C. Chan, J. A. DeMartino, J. M. Farber, and I. Gery. 2004. A unique pattern of
up- and down-regulation of chemokine receptor CXCR3 on inflammation-inducing Th1 cells. Eur. J. Immunol. 34: 2885–2894.
26. Flugel, A., T. Berkowicz, T. Ritter, M. Labeur, D. E. Jenne, Z. Li, J. W. Ellwart,M. Willem, H. Lassmann, and H. Wekerle. 2001. Migratory activity and func-tional changes of green fluorescent effector cells before and during experimentalautoimmune encephalomyelitis. Immunity 14: 547–560.
27. Gery, I., and B. H. Waksman. 1972. Potentiation of the T-lymphocyte response tomitogens. II. The cellular source of potentiating mediator(s). J. Exp. Med. 136:143–155.
28. Piani, A., J. P. Hossle, T. Birchler, C. A. Siegrist, D. Heumann, G. Davies,S. Loeliger,R. Seger,and R. P. Lauener. 2000. Expressionof MHCclassII moleculescontributes to lipopolysaccharide responsiveness. Eur. J. Immunol. 30: 3140–3146.
29. Ulevitch, R. J., and P. S. Tobias. 1995. Receptor-dependent mechanisms of cellstimulation by bacterial endotoxin. Annu. Rev. Immunol. 13: 437–457.
30. Gutcher, I., and B. Becher. 2007. APC-derived cytokines and T cell polarizationin autoimmune inflammation. J. Clin. Invest. 117: 1119–1127.
31. Yokosuka, T., and T. Saito. 2010. The immunological synapse, TCR micro-clusters, and T cell activation. Curr. Top. Microbiol. Immunol. 340: 81–107.
32. Liang, S. C., X. Y. Tan, D. P. Luxenberg, R. Karim, K. Dunussi-Joannopoulos,M. Collins, and L. A. Fouser. 2006. Interleukin (IL)-22 and IL-17 are coex-pressed by Th17 cells and cooperatively enhance expression of antimicrobialpeptides. J. Exp. Med. 203: 2271–2279.
33. Eyerich, S., K. Eyerich, D. Pennino, T. Carbone, F. Nasorri, S. Pallotta,F. Cianfarani, T. Odorisio, C. Traidl-Hoffmann, H. Behrendt, et al. 2009. Th22cells represent a distinct human T cell subset involved in epidermal immunityand remodeling. J. Clin. Invest. 119: 3573–3585.
34. Wan, Q., L. Kozhaya, A. ElHed, R. Ramesh, T. J. Carlson, I. M. Djuretic,M. S. Sundrud, and D. Unutmaz. 2011. Cytokine signals through PI-3 kinasepathway modulate Th17 cytokine production by CCR6+ human memory T cells.
J. Exp. Med. 208: 1875–1887.
8 NAIVE CD4 NEED CELL–CELL INTERACTION FOR Th17 PATHOGENICITY


















