Cap2-HAPComplexIsaCriticalTranscriptionalRegulator ... ·...
18
Cap2-HAP Complex Is a Critical Transcriptional Regulator That Has Dual but Contrasting Roles in Regulation of Iron Homeostasis in Candida albicans * □ S Received for publication, February 24, 2011, and in revised form, May 5, 2011 Published, JBC Papers in Press, May 18, 2011, DOI 10.1074/jbc.M111.233569 Rana Pratap Singh 1 , Himanshu K. Prasad 1 , Ishani Sinha 1 , Neha Agarwal, and Krishnamurthy Natarajan 2 From the School of Life Sciences, Jawaharlal Nehru University, New Delhi 110067, India Iron homeostasis is highly regulated in organisms across evo- lutionary time scale as iron is essential for various cellular pro- cesses. In a computational screen, we identified the Yap/bZIP domain family in Candida clade genomes. Cap2/Hap43 is essen- tial for C. albicans growth under iron-deprivation conditions and for virulence in mouse. Cap2 has an amino-terminal bipar- tite domain comprising a fungal-specific Hap4-like domain and a bZIP domain. Our mutational analyses showed that both the bZIP and Hap4-like domains perform critical and independent functions for growth under iron-deprivation conditions. Tran- scriptome analysis conducted under iron-deprivation condi- tions identified about 16% of the C. albicans ORFs that were differentially regulated in a Cap2-dependent manner. Microar- ray data also suggested that Cap2 is required to mobilize iron through multiple mechanisms; chiefly by activation of genes in three iron uptake pathways and repression of iron utilizing and iron storage genes. The expression of HAP2, HAP32, and HAP5, core components of the HAP regulatory complex was induced in a Cap2-dependent manner indicating a feed-forward loop. In a feed-back loop, Cap2 repressed the expression of Sfu1, a nega- tive regulator of iron uptake genes. Cap2 was coimmunoprecipi- tated with Hap5 from cell extracts prepared from iron-depriva- tion conditions indicating an in vivo association. ChIP assays demonstrated Hap32-dependent recruitment of Hap5 to the promoters of FRP1 (Cap2-induced) and ACO1 (Cap2-re- pressed). Together our data indicates that the Cap2-HAP com- plex functions both as a positive and a negative regulator to maintain iron homeostasis in C. albicans. The generality and evolutionary conservation of transcrip- tional regulatory networks is a vital issue that can now be addressed with the availability of a multitude of genome sequences. The yeast Saccharomyces cerevisiae has been a workhorse for understanding major transcriptional regulatory paradigms including cellular responses to various stress. Sev- eral recent examples, however, showed a divergence of tran- scriptional regulatory networks between S. cerevisiae and other fungal genomes (1, 2). Thus the lessons from the S. cerevisiae transcriptional programs would have to be re-evaluated as new genomes are sequenced and annotated. Iron homeostasis is a highly regulated process as iron is essential for various cellular processes, but becomes toxic at high levels (3, 4). The regulation of iron homeostasis in the yeast S. cerevisiae occurs both at transcriptional and post-transcrip- tional levels (5, 6). The S. cerevisiae transcriptional activator Aft1 and its paralog Aft2 regulate expression of genes encoding reductive iron assimilatory system FRE1–FRE6, FET3, FTR1, iron-siderophore importers ARN1–ARN4, and the low-affinity transporter encoded by FET4 during iron starvation (7, 8). Aft1 expression is constitutive and its nuclear localization occurs upon iron depletion (9). Because iron-dependent deactivation of Aft1/Aft2 is abrogated in cells defective for mitochondrial iron-sulfur (Fe-S) cluster biogenesis, it was concluded that iron sensing by Aft1/Aft2 is dependent on mitochondrial Fe-S export (10). During iron deprivation, Aft1/Aft2 also induces the expression of CTH2, encoding a zinc-finger domain protein, which in turn binds to the AU-rich elements in the mRNAs of iron-utilizing genes and targets them for degradation (11). Although a functional homolog of Aft1/Aft2 has not been found in fungi other than Saccharomyces (12), the AFT2-like gene in Candida albicans, although not required for ferric reductase activity under iron deprivation conditions, was required for optimal ferric reductase activity at high and low pH conditions (13). A GATA family transcription factor Sfu1 in C. albicans (14), or its orthologs in other organisms, function as a negative regulator of iron uptake genes in high iron medium (reviewed in Refs. 15 and 16). In addition, the regulation of iron homeostasis in fungal genomes, including C. albicans, is dependent on the HAP regulatory complex (17–21). In S. cerevisiae, the HAP complex components Hap2, Hap3, and Hap5, are homologous to mammalian NF-YA, NF-YB, and NF-YC subunits, respectively, of the trimeric CCAAT box binding regulator NF-Y (22). Although the trimeric complex is sufficient to regulate gene expression in mammalian cells, the Hap4 regulatory subunit associates with Hap2/Hap3/Hap5 in S. cerevisiae to form the HAP complex, a transcriptional activator of respiratory pathway genes (23, 24). In S. cerevisiae, however, the HAP complex does not function in iron homeostasis (25). Barring Saccharomyces genomes, other fungal genomes studied (17, 21, 26, 27) lacked a full-length Hap4 protein. Instead, HapX from A. nidulans (17) and Php4 from Schizosac- charomyces pombe (18) contained the short 17-amino acid HAP4-like (HAP4L) domain. These proteins have been shown * This work was supported in part by a research grant from the Department of Biotechnology, Ministry of Science and Technology, Government of India, and the UPOE and DST-PURSE grants from Jawaharlal Nehru University. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental “Results,” Fig. S1, and Tables S1–S5. 1 Supported by Junior and Senior Research Fellowships from Council of Sci- entific and Industrial Research, New Delhi. 2 To whom correspondence should be addressed. Fax: 91-11-2674-2558; E-mail: [email protected]. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 28, pp. 25154 –25170, July 15, 2011 © 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 25154 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011 by guest on November 26, 2020 http://www.jbc.org/ Downloaded from
Transcript of Cap2-HAPComplexIsaCriticalTranscriptionalRegulator ... ·...

Cap2-HAP Complex Is a Critical Transcriptional RegulatorThat Has Dual but Contrasting Roles in Regulation of IronHomeostasis in Candida albicans*□S
Received for publication, February 24, 2011, and in revised form, May 5, 2011 Published, JBC Papers in Press, May 18, 2011, DOI 10.1074/jbc.M111.233569
Rana Pratap Singh1, Himanshu K. Prasad1, Ishani Sinha1, Neha Agarwal, and Krishnamurthy Natarajan2
From the School of Life Sciences, Jawaharlal Nehru University, New Delhi 110067, India
Iron homeostasis is highly regulated in organisms across evo-lutionary time scale as iron is essential for various cellular pro-cesses. In a computational screen, we identified the Yap/bZIPdomain family inCandida clade genomes.Cap2/Hap43 is essen-tial for C. albicans growth under iron-deprivation conditionsand for virulence in mouse. Cap2 has an amino-terminal bipar-tite domain comprising a fungal-specific Hap4-like domain anda bZIP domain. Our mutational analyses showed that both thebZIP and Hap4-like domains perform critical and independentfunctions for growth under iron-deprivation conditions. Tran-scriptome analysis conducted under iron-deprivation condi-tions identified about 16% of the C. albicans ORFs that weredifferentially regulated in a Cap2-dependent manner. Microar-ray data also suggested that Cap2 is required to mobilize ironthrough multiple mechanisms; chiefly by activation of genes inthree iron uptake pathways and repression of iron utilizing andiron storage genes. The expression ofHAP2,HAP32, andHAP5,core components of theHAP regulatory complexwas induced ina Cap2-dependent manner indicating a feed-forward loop. In afeed-back loop, Cap2 repressed the expression of Sfu1, a nega-tive regulator of iron uptake genes. Cap2was coimmunoprecipi-tated with Hap5 from cell extracts prepared from iron-depriva-tion conditions indicating an in vivo association. ChIP assaysdemonstrated Hap32-dependent recruitment of Hap5 to thepromoters of FRP1 (Cap2-induced) and ACO1 (Cap2-re-pressed). Together our data indicates that the Cap2-HAP com-plex functions both as a positive and a negative regulator tomaintain iron homeostasis in C. albicans.
The generality and evolutionary conservation of transcrip-tional regulatory networks is a vital issue that can now beaddressed with the availability of a multitude of genomesequences. The yeast Saccharomyces cerevisiae has been aworkhorse for understanding major transcriptional regulatoryparadigms including cellular responses to various stress. Sev-eral recent examples, however, showed a divergence of tran-scriptional regulatory networks between S. cerevisiae and other
fungal genomes (1, 2). Thus the lessons from the S. cerevisiaetranscriptional programs would have to be re-evaluated as newgenomes are sequenced and annotated.Iron homeostasis is a highly regulated process as iron is
essential for various cellular processes, but becomes toxic athigh levels (3, 4). The regulation of iron homeostasis in the yeastS. cerevisiae occurs both at transcriptional and post-transcrip-tional levels (5, 6). The S. cerevisiae transcriptional activatorAft1 and its paralog Aft2 regulate expression of genes encodingreductive iron assimilatory system FRE1–FRE6, FET3, FTR1,iron-siderophore importers ARN1–ARN4, and the low-affinitytransporter encoded by FET4 during iron starvation (7, 8). Aft1expression is constitutive and its nuclear localization occursupon iron depletion (9). Because iron-dependent deactivationof Aft1/Aft2 is abrogated in cells defective for mitochondrialiron-sulfur (Fe-S) cluster biogenesis, it was concluded that ironsensing by Aft1/Aft2 is dependent on mitochondrial Fe-Sexport (10). During iron deprivation, Aft1/Aft2 also induces theexpression of CTH2, encoding a zinc-finger domain protein,which in turn binds to the AU-rich elements in the mRNAs ofiron-utilizing genes and targets them for degradation (11).Although a functional homolog of Aft1/Aft2 has not been
found in fungi other than Saccharomyces (12), the AFT2-likegene in Candida albicans, although not required for ferricreductase activity under iron deprivation conditions, wasrequired for optimal ferric reductase activity at high and lowpHconditions (13). A GATA family transcription factor Sfu1 in C.albicans (14), or its orthologs in other organisms, function as anegative regulator of iron uptake genes in high iron medium(reviewed in Refs. 15 and 16). In addition, the regulation of ironhomeostasis in fungal genomes, including C. albicans, isdependent on the HAP regulatory complex (17–21). In S.cerevisiae, the HAP complex components Hap2, Hap3, andHap5, are homologous to mammalian NF-YA, NF-YB, andNF-YC subunits, respectively, of the trimeric CCAAT boxbinding regulator NF-Y (22). Although the trimeric complex issufficient to regulate gene expression in mammalian cells, theHap4 regulatory subunit associateswithHap2/Hap3/Hap5 in S.cerevisiae to form the HAP complex, a transcriptional activatorof respiratory pathway genes (23, 24). In S. cerevisiae, however,the HAP complex does not function in iron homeostasis (25).Barring Saccharomyces genomes, other fungal genomes
studied (17, 21, 26, 27) lacked a full-length Hap4 protein.Instead, HapX fromA. nidulans (17) and Php4 from Schizosac-charomyces pombe (18) contained the short 17-amino acidHAP4-like (HAP4L) domain. These proteins have been shown
* This work was supported in part by a research grant from the Department ofBiotechnology, Ministry of Science and Technology, Government of India,and the UPOE and DST-PURSE grants from Jawaharlal Nehru University.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental “Results,” Fig. S1, and Tables S1–S5.
1 Supported by Junior and Senior Research Fellowships from Council of Sci-entific and Industrial Research, New Delhi.
2 To whom correspondence should be addressed. Fax: 91-11-2674-2558;E-mail: [email protected].
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 28, pp. 25154 –25170, July 15, 2011© 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
25154 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

to regulate iron homeostasis by a mechanism involving tran-scriptional repression of iron utilizing genes (reviewed in Refs.16 and 28). The S. cerevisiaeHap5 protein contains a conservedHap4 interaction domain, required for interaction with Hap4(29). The interaction of Hansenula polymorpha Hap4a andHap4b proteins with Hap5 has also been demonstrated usingthe yeast two-hybrid assay (21). Furthermore,HapX interactionhas also been uncovered with the HAP2-HAP3-HAP5 complexusing bimolecular fluorescence complementation assay (17).Together these reports showed that HAP4L family proteinsinteract with the entire HAP complex, in a manner dependenton the HAP4L domain in Hap4, and the Hap4 interactiondomain in Hap5.C. albicans is an opportunistic human fungal pathogen that
causes Candidiasis. Because iron is sequestered by ferritin,transferrin, and lactoferrin, the mammalian host is an iron-poor environment. Moreover, C. albicans also competes withthe hostmicrobiome in biofilms and the gut for the limited ironsupply (30). Thus, the ability of C. albicans to acquire ironbecomes critical for its survival as well as for pathogenesis (31).As in S. cerevisiae, there are multiple pathways for iron uptakein C. albicans as well, including high-affinity uptake, sidero-phore uptake, and heme uptake (32–35). Previous reportsshowed that FTR1 is required forC. albicans pathogenesis (33).The C. albicans genome contains genes encoding sidero-
phore transporter, four iron permeases, five multicopper oxi-dases, a family of 17 putative ferric reductases, as well as hemeuptake components indicating the multiplicity of iron acquisi-tion pathways (28, 32). C. albicans can also acquire iron fromferritin in a manner dependent on the Als3 protein (36), but,iron acquisition throughAls3/ferritin does not exist in S. cerevi-siae. Thus a complete understanding of the regulatory networkgoverning iron homeostasis is required to assess its full impactonC. albicans pathogenesis. In this study, we demonstrate thatthe Cap2-HAP complex is a critical transcriptional regulatorthat has dual but contrasting functions to maintain iron home-ostasis in C. albicans.
EXPERIMENTAL PROCEDURES
Media andGrowth Conditions—C. albicans strains were cul-tured in synthetic complete limited iron medium (SC-LIM)3containing 1.7 g/liter of yeast nitrogen base without iron andcopper (MP Biomedicals LLC), 5 g/liter of ammonium sulfate,20 g/liter of dextrose, complete amino acid supplements, and2.5 �M CuSO4. Iron-depleted medium contained SC-LIM plusBPS (Sigma) and iron-replete medium contained SC-LIM plusFeCl3 (100 �M) alone or FeCl3 and BPS together.Strains and Plasmids—Plasmids andC. albicans strains used
in this study are listed under supplemental Tables S1 and S2.The C. albicans strain SN152 (37), a kind gift from SuzanneNoble and A. D. Johnson, was used as the parental strain for allstrains derived in this study. The list of oligonucleotides is pro-vided under supplemental Table S3. Details of strain and plas-mid constructions are available on request.
The cap2� mutant strains RPC51 and RPC75 were con-structed inC. albicans strain SN152 background using the PCRfusion strategy (37) by electroporation (38). Correct integrationwas confirmed by PCR, and the absence of the CAP2 ORF instrain RPC75 was confirmed by PCR.Plasmid pRC20was constructed by replacing theCaURA3 in
CIp10 (39) with the C.d.ARG4 gene (37), and contained theCAP2ORF flanked by 631 bp upstream and 323 bp downstreamregions. The 2859-bp insert was sequenced completely byprimer walking with oligonucleotides ONC213, ONC10,ONC56, ONC57, ONC58, and ONC81. The sequencing datarevealed that the clone contained the CAP2 allele orf19.8289sequence, and showed eight nucleotide differences comparedwith theCAP2/orf19.681 sequence in CGD (Assembly 19). Thecloned gene contained additional changesA774G,C1279T, andT1848C; of the three, the first and third resulted in silent muta-tions, whereas C1279T led to the Pro to Ser change. Thesenucleotide differences in pRC20 did not cause a measurablereduction in CAP2 function, as judged by growth of strainRPC192 in BPS-containing medium. Isogenic wild-type strainsRPC132 (His� Leu�) and RPC206 (His� Arg�) were con-structed by transforming C.d.ARG4 (37) and C.d.HIS1 (40)DNA fragments.The cap2mutant plasmids bearing amino acid substitutions
in the bZIP domain (cap2–1; pRC62) or a deletion of theHAP4L domain (cap2–2; pRC53) were constructed by site-di-rected mutagenesis using primers ONC230/ONC231 (cap2–1allele) or ONC214/ONC215 (cap2-2 allele) and pRC20 as tem-plate. The coding region in the resultant plasmids weresequenced completely and all mutagenic changes were con-firmed. Plasmids pRC62 and pRC53 were linearized with Stu1and integrated at the RPS1 locus in strain RPC75 to createstrains RPC309 (cap2–1) and RPC286 (cap2–2). Because thebZIP domain mutation could not be discerned by the PCRproduct size alone, we used primers ONC101/ONC315 orONC101/ONC316 that specifically amplify the wild-type andbZIP mutant sequences, respectively.Plasmids pNA7, pNA17, and pNA13 bearing the CAP2,
cap2–1, and cap2–2 alleles, respectively, contained the caSAT1gene (38), in lieu of the C.d.ARG4 marker gene in plasmidspRC20, pRC62, and pRC53. Strains RPC506 (cap2–2 CAP2)and RPC517 (cap2–2 cap2–1) were created by integratingDraIII-cut pNA7 and pNA17, respectively, into strain RPC286.Strains RPC511 (cap2–1 CAP2) and RPC521 (cap2–1 cap2–2)were similarly created by integrating DraIII-cut pNA7 andpNA13 into strain RPC326. The control strain RPC510 wasconstructed by integrating pNA7 into RPC278, which was gen-erated by integrating CIp10-C.d.ARG4 into strain RPC75. Thepresence of the variousCAP2 alleles was confirmed by diagnos-tic PCR using primers ONC101/ONC315 or ONC101/ONC316 (data not shown).C. albicans hap32� mutant strain RPC 431 was constructed
using the split-marker strategy (41). Strains RPC445 (hap31�/�)andRPC434 (sfu1�/�) were produced in SN152 by sequentiallyintroducing PCR amplicons generated using locus-specific longoligonucleotides and pFA-CdHIS1 and pFA-CmLEU2 (42) astemplates. The strain RPC449 (hap31�/� hap32�/�) was con-structed in the RPC445 background. C. albicans hap2�/� and
3 The abbreviations used are: SC-LIM, synthetic complete limited iron medium;BPS, bathophenanthroline-disulfonic acid; HAP4L, HAP4-like; CGD, CandidaGenome Database; GEO, Gene Expression Omnibus; IP, immunoprecipitation;SOD, superoxide dismutase; qRT, quantitative RT.
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25155
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

hap5�/� mutant strains (20) were obtained from FungalGenetics Stock Center (43).The cap2�/� mutation was constructed in strain RPC434
(sfu1�/�) using pRC11, which contained the cap2�::SAT1 flip-per cassette flanked by CAP2 upstream (�432 to �32) anddownstream (�1872 to �2233) sequences. The cap2�::SAT1flipper cassette in pRC11 was excised by KpnI-SacI digestionand transformed by electroporation. The various strainsdescribed above were confirmed using locus-specific andORF-specific PCR primers.TAP tag plasmids Ip21 and Ip22 contained C.m.LEU2 and
C.d.HIS1marker genes, respectively (37), and the ACT1 termi-nator region (44) and the BamHI-EcoRV TAP tag fragmentfrom pPK335 (45). The SFU1-TAP tagging cassettes were gen-erated by PCR using primers ONC257/ONC258 and Ip21 andIp22 as template. The targeted integration of the tagging cas-settes was confirmed by diagnostic PCR and the expression ofSfu1-TAP was confirmed by Western blot using rabbit poly-clonal anti-TAP antibody (Open Biosystems). The HAP5-TAPstrains RPC453 (wild-type) and RPC562 (hap32�) were con-structed and verified using the same strategies as mentionedabove for the SFU1-TAP.The HIS6-FLAG3-tagged HAP5 was constructed from plas-
mid pSH26-5 (caSAT1-FLIP cassette) and split-marker strat-egy. The HAP5 sequence was PCR amplified as the up-split(�2640 bp) and down-split (�3100 bp) fragments using prim-ers ONC460/ONC140 and ONC141/ONC461. The up-splitand down-split amplicons, bearing �1 kb overlap, contained60-bp homology to either side of the HAP5 stop codon. Theamplicons were transformed into SN152 and correct integra-tion of the tagging cassette in RPC552 was confirmed by diag-nostic PCR and expression of tagged Hap5 was confirmed byWestern blot using affinity purified mouse monoclonal anti-FLAGM2 antibody (Sigma).Production of Anti-Cap2 Polyclonal Antibody—Plasmid Ip9
contained truncated CAP2 (�1 to �542 with respect to ATG)in pET28b� vector (Novagen). The recombinant Cap2-His6protein was expressed in Escherichia coli, purified, and poly-clonal antibodies were raised in rabbit. The anti-Cap2 antibodywas affinity purified using recombinant Cap2.Virulence Assay—C. albicans strains RPC206, RPC278, and
RPC192 were grown overnight in YEPD, inoculated into freshYEPD medium to a starting A600 of 0.5, and grown for 4 h at30 °C. The cells were pelleted, washed, and resuspended in ster-ile normal saline, cells were counted, and �2 � 106 cells in 100�l were injected into the lateral tail vein of 8–10-week-oldBALB/c mice with 5–7 mice per group. The mice were fedwater and food ad libitum and monitored daily for vital signsand activity.Whenmice appearedmoribund (hunched postureand minimal motor activity), they were euthanized, organswere dissected, and the fungal burden in kidney and brain wasexamined. The mice were handled humanely as per protocolsfor animal experiments approved by the Jawaharlal Nehru Uni-versity Institutional Animal Ethics Committee.Immunoblotting—C. albicans strains were grown overnight
in SC-LIM medium and inoculated to a starting A600 of 0.25(CAP2) or 0.5 (cap2�) into fresh SC-LIM medium with BPSalone or with BPS plus FeCl3 or FeCl3 alone. Cultures were
grown at 30 °C for different time periods and harvested, pelletswere resuspended in 300 �l of extraction buffer (46) and cellswere lysed by vortexing. The lysate was spun at 18,500 � g at4 °C for 15 min, the supernatant was centrifuged again and thecleared lysate collected. Protein concentration was determinedby the Bradford assay (Bio-Rad) using BSA as standard. About60 �g of whole cell extract was resolved by 6–12% SDS-PAGEand transferred to HybondTM-ECLTM nitrocellulose mem-brane (GEHealthcare). The blot was probedwith primary anti-body, followed by a secondary HRP-conjugated antibody anddetected using ECL Plus Western blotting detection system(GE Healthcare).RNA Extraction and RT-PCR Analysis—C. albicans strains
were grown as described under “Immunoblotting.” Cells equiv-alent toA600 � 10 were harvested rapidly by filtration and snapfrozen in liquid nitrogen vapors. Total RNA was isolated usinghot phenol method (47) and the concentration was determinedusing Nanodrop spectrophotometer.For all RT-PCR experiments, total RNA was treated with
RNase-free DNase I (Invitrogen) to remove any residual DNA.About 150 ng of DNase I-treated RNA was used for single-stranded cDNA synthesis in 7.5 �l of reaction mixture using aHigh-Capacity cDNA Reverse Transcription kit (Applied Bio-systems). For semi-quantitative RT-PCR analysis, 1.0 �l of10-fold dilution of cDNA reaction mixture was used as a tem-plate using Taq DNA polymerase. SCR1 RNA, an RNA poly-merase III transcript (48), was used as endogenous control.Real-time qRT-PCR was carried out in Applied Biosystems7500 Real-time PCR system, where each 10 �l of reaction mix-ture contained 4 �l of 500-fold diluted cDNA, 0.1 �M gene-specific primers and SYBRGreenPCRmastermixture (AppliedBiosystems). The comparativeCTmethod (2���CT) was used todetermine the relative gene expression (49). Control reactionswithout reverse transcriptase were carried out for each cDNApreparation and ascertained that no amplificationwas obtainedas judged by high CT (�39) values and gel analysis.Microarray Analysis—Total RNA was isolated by the hot
phenol method from control SN152 (CAP2) and RPC75(cap2�) strains grown in SC-LIMmedium plus 100 �M BPS for5 h.C. albicansmicroarrays (50) were obtained from theWash-ington University Microarray Core facility. All steps inmicroarray analysis including image quantitation and data nor-malization steps are detailed in GEO entry GSE 28429. Themicroarray hybridization was repeated with two independentsets of RNA preparations. The data were filtered for good spotsin GenePix Pro 6.0 as described before (51), data were normal-ized, and the log2 ratio (CAP2/cap2�) for each spot obtained.The data for triplicate spots (with log2 ratio �1 S.D.) for eachgene in each array was merged and used for further statisticalanalysis. Significance analysis of microarrays (52) was per-formed as described previously (53), with the exception of 1000permutations to get differentially expressed genes with lowfalse discovery rate. GeneOntology analysis of the differentiallyexpressed genes was carried out using Bingo 2.42 (54) in Cyto-scape version 2.7 visualization and analysis software (55) withCGD Candida GO annotation (downloaded August 10, 2010)and overrepresented biological processes with high statisticalsignificance (p value � 0.05) obtained using a hypergeometric
Central Regulator of Iron Homeostasis
25156 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

test and Benjamini andHochberg FDR correction. Heat map ofthe differentially expressed genes was constructed using the RBioconductor package running under Linux. The microarraydata has been deposited under GEO entry GSE 28429.Coimmunoprecipitation Assay—C. albicans strains SN152
and RPC552 were cultivated in BPS containing medium at30 °C for 5 h, andwhole cell extractswere prepared as describedunder “Immunoblotting.” About 1.2 mg each of the whole cellextracts were mixed with 10 �l of anti-FLAG M2 affinity gel(Sigma, A-2220), and incubated on a rotating platform at 4 °Cfor 4 h. After IP, beadswere collected, washed, and resuspendedin 25 �l of 1� SDS-PAGE sample buffer. The entire immuno-precipitated sample and 100 �g of input protein extracts wereseparated in a gradient 6–12% SDS-polyacrylamide gel, trans-ferred to Hybond-ECLTM nitrocellulose membrane (GEHealthcare), and immunoblotting was performed. The follow-ing antibodies were used to probe the immunoblots: affinitypurified monoclonal anti-FLAG M2 antibody (1:1000; Sigma),affinity purified polyclonal anti-Cap2 antibody (1:2500), and ascontrol anti-yeast glucose-6-phosphate dehydrogenase anti-body (1:500; Sigma).Chromatin Immunoprecipitation (ChIP) Assay—HAP5-TAP
strains RPC453 (HAP32/HAP32) and RPC562 (hap32�/hap32�) were grown in SC-LIM containing 0.1 mM BPS or 0.1mM BPS plus iron for 5 h at 30 °C to an A600 of 0.8–1.0. Form-aldehyde (1% v/v) was added to the cultures and cross-linkedfor 20 min at 30 °C and cells were spheroplasted in a buffercontaining 1 M sorbitol, 50 mM Tris-Cl (pH 7.4), and 10 mM
�-mercaptoethanol, and 0.03 mg/ml of zymolyase (ICN).Spheroplasts were resuspended in 50 mM HEPES-KOH (pH7.5), 140mMNaCl, 1mMEDTA, 1%TritonX-100, 0.1% sodiumdeoxycholate, 0.1% SDS plus protease inhibitors (1 mM PMSFand 1 mg/ml each of leupeptin, aprotinin, and pepstatin) andsheared by sonication using Bioruptor (model UCD 300, Diag-enode), at high power for 30 cycles (30 s on and 30 s off). Theaverage chromatin size obtained was �0.25 kb. About 25 �l ofpre-blocked IgG-Sepharose (GE Healthcare) was used forimmunoprecipitation from the sheared chromatin extractequivalent to�20A600 cells and the ChIP assay was carried outessentially as described previously (56). Whole cell extracts(Input) and the IP eluatewere treated for reversal of cross-links,DNA purified by phenol-chloroform extraction, and analyzedby qPCR. ChIP-qPCR data analysis was done as described pre-viously (49).
RESULTS
C. albicans Cap2 Is a HAP4L-bZIP Bipartite Domain Protein—We identified the complete repertoire of bZIP domain contain-ing sequences from S. cerevisiae, Candida glabrata, and 10Candida clade species (Fig. 1A, and supplemental Fig. S1). The124 bZIP domain sequences identified were used to constructmultiple sequence alignment (supplemental Fig. S1) of a mini-mal region of 38 amino acids and a phylogenetic tree was con-structed (Fig. 1A). The phylogenetic tree revealed clustering ofthe bZIP domains into distinct clusters occupied by the S.cerevisiae Yap1–Yap8, Hac1, Sko1/Aca1–2, Gcn4, and Met4–Met28-proteins, respectively (Fig. 1A). Analysis of the treerevealed that the Candida clade genomes had four Yap/bZIP
proteins named CAP1 through CAP4, compared with the eightin S. cerevisiae andC. glabrata (Fig. 1B). The Yap/bZIP domainof each of the four C. albicans Cap proteins contained the fourYap-specific residues described previously (57), and are indi-cated in Fig. 1B. CAP1 has been previously characterized as afunctional ortholog of the S. cerevisiae YAP1 gene (58), andCAP3/FCR3 upon overexpression conferred fluconazole resist-ance to the S. cerevisiae pdr1�pdr3�mutant (59, 60).However,thus far no function has been attributed to the CAP4 gene.
Ourmutational analyses showed that theCAP2/HAP43 genewas required for growth of C. albicans under iron deprivationconditions (Fig. 2A). Furthermore, the cap2� mutant showeddelayed virulence in a mouse model of C. albicans virulence(Fig. 2B). Analysis of the fungal burden from infected miceshowed at least 50% recovery of cap2� cells as compared withwild-type from the kidney and brain as determined by colonyforming units (Fig. 2C).Sequence analysis showed that the Cap2 contains the 17-
amino acid HAP4L domain (Pfam accession PF10297) at itsamino-terminal region. Because the bZIP and HAP4L domainsare located next to each other, we have named them the bipar-tite domain. Cap2 was of great interest to us because Cap2orthologs found in multiple fungal genomes have been shownto be required for growth under iron-deprivation conditionsand for regulation of iron homeostasis (17, 19–21, 61–63).Although a point mutation in the HAP4L domain of Hap4aimpaired the growth of Histoplasma capsulatum in glycerolmedium (26), the requirement of the HAP4L domain or thebZIP domain for Cap2 function was not understood.Both bZIP and HAP4-like Domains Are Required for CAP2
Function—To examine the requirement of bZIP domain forCap2 function in vivo, we carried out site-directedmutagenesis.Previous high resolution crystal structure of the S. pombe Pap1Yap/bZIP domain had revealed that the basic region motifNXXAQXXFR makes direct contacts with the 5�-TTACG-TAA-3� DNA element (64). Moreover, mutational analysis hadalso established the requirement of basic region residues in theGcn4 bZIP domain for DNA interaction (57, 65). Therefore weconstructed strains bearing the cap2–1 allele containing substi-tutions in four of the five critical amino acid residues, Asn-93,Gln-97, Phe-100, and Arg-101 (Fig. 3A). Western blot analysisshowed that expression of Cap2–1 protein was comparablewith the wild-type (Fig. 3B). The cap2–1 allele was unable tofully complement the BPS-sensitive growth defect of the cap2�strain (Fig. 3C). These data indicated that the bZIP domain iscritical for Cap2 function under iron limiting conditions.To investigate the requirement of the HAP4L domain for
Cap2 function, we constructed the cap2–2 allele, lacking theHAP4L domain (residues 63 to 79) (Fig. 3D). Although theCap2–2 expressionwas not impaired (Fig. 3E, lane 3), the cap2-2 strain was unable to grow in BPS-containing medium (Fig.3F). Thus the HAP4L domain is critically required for Cap2function under iron limiting conditions. We carried out a sys-tematic identification and phylogeny analyses of hapX andCap2-like protein sequences bearing the bipartite domain andidentified 47 sequences, the bulk of them not previously stud-ied, from 46 of the 62 fungal genomes analyzed (supplementalTable S4B), indicating that this protein family is highly perva-
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25157
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

Central Regulator of Iron Homeostasis
25158 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

sive in the fungal kingdom (Fig. 3G). Multiple sequence align-ment of the 47 bipartite domain sequences revealed the Yap-like amino acid residues in the bZIP domain of all sequences.Next, we wished to test if the bZIP and HAP4L domains
perform independent functions in Cap2. Because Yap proteinsform homodimers (64), we carried out trans-complementation
assays by co-expressing the two cap2 alleles, cap2–1 (bZIPmutant) and cap2–2 (HAP4L deletion mutant). Results of thetrans-complementation assay showed that the strains express-ing both cap2–1 and cap2–2 did not confer BPS resistance,indicating that the two defective copies did not complementeach other (rows 6 and 9, Table 1). Together these data demon-strated that both bZIP and HAP4L domains are required forCap2 function, and the lack of trans-complementation by thetwo mutant proteins suggests that the two domains performindependent functions in vivo.Transcriptome Profiling and Gene Ontology Analysis of Cap2
Regulon—Because the CAP2 gene is required for growth underiron-deprivation conditions, we examined if the CAP2 mRNAand protein levels were regulated by iron availability. RT-PCRanalysis showed that iron deprivation led to up-regulation ofCAP2 mRNA in a time-dependent manner, with a maximuminduction of 5.7-fold after 3 h of BPS addition (Fig. 4A). Addi-tion of iron to BPS-containing medium led to repression ofCAP2mRNA levels (Fig. 4A, lanes 7 and 8). Western blot anal-ysis also showed that theCap2 proteinwas highly induced uponBPS treatment and was undetectable in cell extracts from iron-containing medium (Fig. 4B).To identify Cap2-regulated genes, we carried out transcrip-
tome profiling (66, 67) using the C. albicans whole genomemicroarray (50).We compared themRNA profiles of wild-typeand cap2� cells cultured in BPS-containing medium and iden-tified 420 up-regulated and 509 down-regulated genes (total15.8% of the ORFs examined) that showed statistically signifi-cant differential expression (supplemental Table S5). To iden-tify Cap2-regulated biological processes, we carried out GeneOntology analysis of the differentially expressed genes usingBingo 2.42 software.We obtained several overrepresented, sta-tistically significant GO functional processes. Ten genes werefound to be associated with the transition metal ion transportincluding several genes in the iron uptake pathway and will bediscussed later.Moreover, seven up-regulated genes were asso-ciated with the pyruvatemetabolic process. Interestingly, pyru-vate production and increased glycolysis has also been observedduring iron deprivation in S. cerevisiae (68). Our microarraydata also revealed several transcriptional regulator genes thatshowed Cap2 dependence, of which 25 genes were up-regu-lated, and 15 were down-regulated. The differential regulationof these transcriptional regulator geneswould likely amplify thesignal perceived by CAP2 during the iron deprivation responseand account for the large Cap2 transcriptome, thus indicatingthat Cap2 is a master regulator of iron homeostasis in C.albicans.GO analysis of the down-regulated genes also identified sev-
eral biological processes including generation of precursormetabolites and energy, aerobic respiration, respiratory chain
FIGURE 1. Bioinformatics of Candida clade bZIP domain sequences. A, comparative phylogeny of bZIP domains from Candida and Saccharomyces cladegenomes. Neighbor-joining tree (1000 bootstrap replicates) was constructed based on the multiple sequence alignment of Candida clade, S. cerevisiae, and C.glabrata bZIP domains using MEGA4 software. The known bZIP subfamily proteins from S. cerevisiae (blue box) are indicated by their gene names as given in theSaccharomyces Genome Database. The putative orthologs from C. albicans (red circle) and other organisms are indicated. The unique identifiers of proteins/fungal species are provided under supplemental Table S4A. B, structural features of C. albicans Cap proteins. Top, schematic diagram of the predicted domainsin the CAP proteins: HAP4L domain, bZIP domain, cysteine-rich domains (CRD-1, -2, and -3) and the low complexity regions (mostly serine-rich) identified usingSMART and Motif Scan software. Bottom, multiple sequence alignment of the bZIP domain sequence from Cap1– 4, Yap1– 8, S. pombe Pap1, and S. cerevisiaeGcn4. Characteristic Yap-specific residues found in all Yap-like proteins but not in Gcn4 are indicated (*).
FIGURE 2. Effect of cap2� mutation on C. albicans growth and virulence.A, the growth phenotype of cap2�/� mutant in iron-deplete medium. C. albi-cans strains SN152 (CAP2�/�), RPC51 (cap2�/�), RPC75 (cap2�/�), andRPC192 (cap2�/�/�) were grown overnight in SC-LIM liquid medium at30 °C. About 5 �l each from 0.001, 0.01, and 0.1 A600 equivalent cell suspen-sions were spotted onto SC-LIM, SC-LIM � 50 �M BPS, or SC-LIM � 50 �M BPSplus 100 �M FeCl3 plates and incubated at 30 °C. B, the cap2�/� mutantshowed delayed virulence in mouse. Strains RPY206 (CAP2�/�), RPC278(cap2�/�), and RPC192 (cap2�/�/�) were injected into BALB/c mice (n 5–7) and the percentage of surviving mice was plotted for each group.Kaplan-Meier test yielded a p value 0.004 between wild-type and cap2�/�mutant. C, the fungal burden in the brain and kidneys of the dead or mori-bund mice were determined by plating aliquots of appropriate dilutions oftissue homogenates on YPD containing ampicillin (50 �g/ml) and gentamy-cin (0.8 mg/ml). The recovered colonies were tested for their auxotrophicmarkers to confirm the identity of the strains.
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25159
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

complex assembly, mitochondrial ATP synthesis-coupled elec-tron transport, heme and other cofactor metabolism, de novoNADbiosynthesis, 2-oxoglutaratemetabolism, andprotein tar-
geting to peroxisomes (data not shown). The bulk of these pro-cesses require iron, and their down-regulation can be viewed asamechanism geared to reduce iron utilization.Of the six super-
Central Regulator of Iron Homeostasis
25160 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

oxide dismutase (SOD) genes, SOD1–SOD6 in C. albicansgenome (69), the manganese superoxide dismutase genes,SOD2 and SOD3, are repressed in a Cap2-dependent manner.There are previous reports that deletion of CuZn and/or man-ganese SOD genes leads to accumulation of free iron in S. cerevi-siae cells (70, 71). Thus it appears that Cap2-mediated repres-sion of SOD gene expression could additionally mobilize ironin vivo.Cap2-dependent Differential Expression of Genes Regulated
by IronAvailability—Toobtain insights intoCap2 regulation ofiron homeostasis, we analyzed our microarray data in the con-text of genes involved in iron homeostasis.We retrieved the lowiron_versus_high iron (L_H_Fe) microarray data of Lan et al.(14), and a list of all 133 genes annotated in CGD (downloadedApril 21, 2010) bearing the keyword “iron,” plus five additionalgenes CSA1, CSA2, PGA7, FRE9, and orf19.867, related to ironhomeostasis (32). Comparison of the three datasets showedthat 33 genes were up-regulated in L_H_Fe data and showedCap2 dependence. Twelve of these genes were annotated asiron-related and included genes involved in three iron uptakepathways. Of the 48 down-regulated genes in both the experi-mental data, 25 were annotated in CGD as iron-related.To examine the behavior of the Cap2-dependent iron-re-
lated genes, we generated a heatmap for the above discussed 12up-regulated and 25 down-regulated genes (Fig. 4C). TheL_H_Fe data (14) for these genes was also included in the heatmap for comparison. We also included an additional 15 iron-related genes that showed Cap2-dependent up-regulation (sixgenes) or down-regulation (nine genes), for which the L_H_Fe
data were not available. Interestingly, eight of the 18 up-regu-lated genes in the CAP2/cap2� data have been shown experi-mentally to be involved in iron uptake pathways, viz., SIT1encoding siderophore transporter (72), FET3, FTR1, FRP1,CFL5, and FRE9 encoding reductive iron uptake (73, 74), andRBT5 and HMX1 being part of heme uptake (35, 75, 76) path-ways (Fig. 4C). Our microarray data also identified Cap2-de-pendent up-regulation of CSA2, PGA7 (predicted GPIanchored cell wall protein),PHR2, FET3, and FRE9not found inthe L_H_Fe data (Fig. 4C). We also found that HAP2 andHAP32, two components of the trimeric DNA-binding HAPcomplex, were induced in a CAP2-dependent manner.Iron-sulfur containing genes BIO2, SDH2, ACO1, CYC1,
ISA1, and heme containing genes HEM1, HEM3, HEM4,HEM13, and HEM14 were down-regulated in a CAP2-depen-dent manner. There were indications that Cap2 also facilitatediron mobilization by down-regulating SMF3 and YFH1, shownpreviously to be involved in iron storage in S. cerevisiae (77, 78).The heat map also showed that several respiratory pathwaygenes including ACO1, CYC1, SDH2, ISA1, RIP1, and YAH1were down-regulated in a Cap2-dependent manner upon irondeprivation. The behavior of these genes can be expectedbecause iron is an essential cofactor for their activity.Cap2-dependent Activation of Three Iron-acquisition Path-
ways and Repression of Iron-requiring Enzymes—Expression ofkey genes involved in iron homeostasis, identified in ourmicroarray data, was validated by qRT-PCR. The CAP2mRNAwas highly induced in iron-depleted versus iron-repletedmedia(Fig. 4D) confirming the RT-PCR data (Fig. 4A). The data alsoshowed that the expression of genes encoding three differentiron uptake pathways, viz., siderophore, reductive, and hemeuptake, were highly induced upon iron deprivation in the wild-type strain, but substantially impaired in the cap2� strain (irondependence, Fig. 4D). Iron deprivation induced the expressionof FRP1 and SIT1 to �124- and 44-fold, respectively, in thewild-type strain; their induction in the cap2� strain, however,was substantially reduced but not completely lost. Among theinduced genes analyzed, only the induction of HMX1 showedabsolute Cap2 dependence. Whereas the activation of othergenes showedunmistakableCap2dependence, the activation ofFRP1 and SIT1 was still retained in the cap2� strain indicatingan additional means of regulation of expression.The two iron utilization genesACO1 andCOX5 also showed
CAP2-dependent strong repression in iron-depleted medium
FIGURE 3. Both bZIP and the HAP4L domains are required for Cap2 function. A, schematic diagram of Cap2 protein showing amino acid residues within thebasic region of the bZIP domain. The wild-type (Cap2) residues from 93 to 101 are shown above and the corresponding quadruple mutant (Cap2–1) residues,generated by site-directed mutagenesis, are shown below. B, Western blot analysis of the Cap2–1 protein. Whole cell extracts prepared from strains grown iniron-limiting conditions (SC-LIM plus 0.1 mM BPS for 5 h) were blotted and probed with affinity-purified rabbit anti-Cap2 polyclonal antibody (1:2500). Theposition of the �95-kDa Cap2 band is indicated; a nonspecific band (*) below the 72-kDa marker was consistently observed in all the extracts and served as aloading control. The cap2� extract only contained the 72-kDa nonspecific band, establishing the identity of the 95-kDa Cap2 protein band. C, the cap2–1mutant strain is growth impaired under iron-limiting conditions (50 �M BPS). D, schematic diagram showing the evolutionarily conserved HAP4L domain inCap2 and the S. cerevisiae Hap4 (ScHap4) proteins. The amino acid residues deleted in Cap2–2 protein are depicted by dotted lines. E, Western blot analysis ofCap2–2 protein expression. F, deletion of the HAP4L domain abrogated growth under iron-depleted conditions. Strains used were CAP2�/� (SN152), cap2�/�(RPC75), RPC309 (cap2�/�/cap2–1), and RPC286 (cap2�/�/cap2–2). G, the bipartite HAP4L-bZIP domain was identified in 46 fungal genomes. The multiplesequence alignment of the HAP4L-bZIP bipartite domain, created using MUSCLE 3.0 software, was edited by removing gaps in the alignment between theHAP4L and bZIP domains. A core 60-amino acid region was obtained comprising a highly conserved 20-amino acid HAP4L domain, an insert of 7 amino acids,and 33 amino acids of the bZIP domain. A phylogenetic tree was constructed using the neighbor-joining method with 1000 bootstrap replicates using MEGA4software. The clustering represented Pezizomycotina sequences including A. nidulans HapX (I), Saccharomycotina sequences including Cap2 (II), Agaricomy-cotina and Ustilaginomycotina sequences (III), and a second subset of the Saccharomycotina (IV) branch occupied by Yap7. The scale bar shows the number ofamino acid differences per site. The details of the unique identifiers of proteins and that of the fungal species are provided under supplemental Table S4B.
TABLE 1Trans-complementation assay to examine the requirement of HAP4L-bZIP bipartite domainThe caSAT1-marked plasmids bearing CAP2, cap2–2, and cap2–1 were integratedinto RPC278 (cap2�), RPC286 (cap2–2), or RPC326 (cap2–1) to generate strainsexpressing wild-type and/or mutant cap2 in all combinations shown. The strainsbearing the constructs were verified by PCR and spot assay carried out in SC-LIMorSC-LIM � 50 �M BPS plates. Plates were incubated at 30 °C and relative growthscored by visual examination. � indicates no growth.
S. No. Strain CAP2 allele 1 CAP2 allele 2 Growth on BPS plate
1 SN152 CAP2 CAP2 2.0�2 RPC278 cap2� cap2� �3 RPC510 cap2� CAP2 1.5�4 RPC286 cap2� cap2–2 �5 RPC506 cap2–2 CAP2 1.5�6 RPC517 cap2–2 cap2–1 �7 RPC326 cap2� cap2–1 �8 RPC511 cap2–1 CAP2 2.0�9 RPC521 cap2–1 cap2–2 �
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25161
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

Central Regulator of Iron Homeostasis
25162 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

compared with iron-replete medium (Fig. 4D). Interestingly,the expression of ACO1 was higher by �3-fold in the cap2�strain. Analysis of the data also showed Cap2 dependence foractivation and repression of the same genes only in iron limitedbut not in high iron medium. Taken together, the qRT-PCRdata demonstrated Cap2-dependent activation, and repressionof selected iron homeostasis genes under iron-deprivationconditions.Expression of ACO1 and other iron utilizing genes is
repressed by CTH2 in S. cerevisiae. Therefore we examined therequirement of C. albicans CTH2/19.5334 for repression ofACO1 and found that its expression was derepressed �1.5-foldin the cth2� strain (data not shown). Moreover, growth of thecth2� strain was also not compromised in BPS medium. ThusCaCTH2/19.5334 does not seem to have a pronounced effect inrepressing ACO1 expression.Cap2-dependent Repression of the Negative Regulator Sfu1—
Sfu1 was previously shown to be a repressor of iron uptakegenes and of Cap2 under iron-replete conditions (14). More-over, our CAP2/cap2� microarray data (Fig. 4C) revealed thatunder iron-deprivation conditions, SFU1 expression wasrepressed in a CAP2-dependent manner. To confirm this, wecarried out real-time qPCR analysis of SFU1 transcript fromwild-type and cap2� cells. The data showed that SFU1 wasrepressed �5–10-fold by 1 and 5 h of BPS treatment (Fig. 5A).Significantly, this down-regulation of SFU1 mRNA was com-pletely abrogated in cap2� cells (Fig. 5A), indicating that Cap2is required to repress SFU1 in BPS medium. In iron-repletemedium, however, SFU1 expression was derepressed in bothwild-type and cap2� cells (Fig. 5A). In agreement with mRNAanalysis, Sfu1 protein expression was repressed in BPSmediumin the wild-type strain but derepressed in the cap2� strain (Fig.5B, lanes 2 and 4).Together the data presented here showed that Cap2 and Sfu1
regulate expression of each other in a reciprocal manner iniron-deprived and iron-containing media, respectively. More-over, repression of SFU1 mRNA by Cap2 during iron-limitedconditions would further relieve the negative regulatory loopandmaximize iron acquisition. To further probe the regulatoryinteraction between SFU1 and CAP2, we carried out epistasisanalysis. We constructed the cap2� sfu1� double deletionstrain and tested its ability to grow in BPS-containing medium.The phenotype test showed that the cap2� sfu1� doublemutant strain displayed the same phenotype as the cap2�strain, indicating that CAP2 is epistatic to SFU1 under iron-depleted conditions (Fig. 5C).We hypothesized that if SFU1 had a role in repression of
CAP2 in the iron-replete medium, as indicated previously by
microarray analysis (14), the attendant derepression of CAP2upon sfu1 deletion could result in induction of Cap2-targetgenes. Our qRT-PCR analysis showed that indeed the CAP2mRNA level was derepressed �4.0-fold in the sfu1� straincompared with the level in the wild-type strain in high ironmedium (Fig. 5D). However, the mRNA level of Cap2 up-regu-lated genes FRP1, HAP2, HAP32, and HMX1 was not substan-tially altered indicating that expression ofCAP2mRNAwas notsufficient to activate its target genes in high iron medium (Fig.5D). Moreover, expression of ACO1 (Fig. 5D) and CYC1 (datanot shown), down-regulated in a Cap2-dependent manner inlow iron medium, was unchanged in the sfu1� strain in highironmedium.Western blot analysis showed that the Cap2 pro-tein level was only slightly elevated in the sfu1� extract com-pared with the wild-type extract from the high iron medium(data not shown), indicating that regulation ofCAP2 expressionlikely occurs at both mRNA and protein levels.Cap2-dependent Up-regulation of Components of the HAP
Regulatory Complex—There are previous reports that Cap2 orits orthologs likely function in a complex with Hap2-Hap3-Hap5 (19, 21, 29). In a previous study, three HAP4-like genesand two HAP3-like genes were identified in the C. albicansgenome (27). Our bioinformatic analyses further showed thatthe Candida clade genomes had three each of HAP3 (HAP31,HAP32, and orf19.5825 named HAP33), HAP4 (HAP41,HAP42, and HAP43/CAP2), and HAP5 (orf19.1973/HAP5,orf19.3063 namedHFL1, and orf19.2736 namedHFL2) but onlya singleHAP2 sequence. Although the three C. albicansHap5-like sequences each had a histone-fold domain like that of the S.cerevisiae Hap5, only Hap5 is likely to be a functional orthologof Hap5 due to the presence of the Hap4-interaction domain(data not shown).As discussed previously, CAP2/HAP43 was induced in iron-
depleted versus iron-repleted media in the wild-type strain.Therefore we investigated which of the multiple HAP geneswere also induced under iron-deprivation conditions. Theexpression of HAP41 and HAP42 were unaffected (Fig. 6A),whereas of the HAP3-like genes, only HAP32 mRNA expres-sionwas highly induced upon iron deprivation (�93.5-fold) in aCap2-dependent manner (Fig. 6A). HAP31 was repressed iniron-depleted conditions but this repression was not Cap2-de-pendent. The expression ofHAP33was unchanged in the wild-type strain but was down-regulated in the cap2� mutant. ThusonlyHAP32 expression was up-regulated by low iron as well asCap2. TheHAP2mRNAwas induced 4.5-fold in iron-depletedrelative to iron-replete conditions, and this inductionwas com-pletely lost in the cap2� strain (Fig. 6A). Interestingly, althoughHAP5 expression was not induced by iron deprivation in the
FIGURE 4. CAP2-dependent transcriptome profiling. A, time course of CAP2 mRNA induction in iron-limiting medium. Total RNA isolated from C. albicansstrain CAP2�/� (SN152) and cap2�/� (RPC75) were used for first strand cDNA synthesis. 10-fold dilution of the cDNA preparation was used as a template forPCR using CAP2 and endogenous control SCR1 primers and the PCR products were resolved on a native polyacrylamide gel and stained with ethidium bromide.Total rRNA profile obtained in the agarose gel is shown. The relative expression level of CAP2 mRNA, normalized to SCR1 RNA (an RNA polymerase III transcript(48)), was expressed for each time point with reference to the 0 min sample. B, Western blot analysis of Cap2 expression. Whole cell extracts from CAP2�/�(SN152) and cap2�/� (RPC75) strains were used for immunoblot analysis. C, heat map of iron-related genes differentially expressed in a CAP2-dependentmanner. Two-color microarray data expressed as log2 CAP2/cap2� ratio (four replicates 1a, 1b, 2a, and 2b and average of the four replicates, Ave) is plotted asheat map. The L_H_Fe data (14) is also shown for comparison. The color scale at the bottom indicates the log2 ratio (CAP2/cap2�). D, qRT-PCR analysis ofselected genes differentially expressed in CAP2/cap2� microarray data. For relative quantification, �CT values were derived after normalization to SCR1transcript levels (endogenous control), and ��CT values were derived using either BPS � FeCl3 sample as calibrator for iron dependence or cap2� sample ascalibrator for CAP2-dependence. Fold-change (2���CT) was calculated from the average of at least three RNA preparations, and S.D. was determined.
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25163
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

wild-type strain, HAP5 expression was reduced in the cap2�strain (Fig. 5A), indicating that Cap2 was required to maintainHAP5 expression in iron-deprivation conditions.Whereas acti-vation of HAP2 expression was almost entirely dependent onCap2, the activation of HAP32 expression, although reduced�10-fold, was still maintained in the cap2� mutant indicatingother forms of regulation.
We carried out mutational analysis to examine the require-ment of HAP3-like genes and found that the hap32� mutantbut not the hap31� mutant was impaired for growth in BPSmedium (Fig. 6B), but the defective phenotype was not as tightas that of the cap2� mutant (Fig. 6B). The phenotype of thehap32� hap31� double mutant could not be accuratelyassessed in BPSmedium due to the growth defect of the double
FIGURE 5. Reciprocal regulation of Cap2 and Sfu1 expression. A, qRT-PCR analysis of SFU1 mRNA expression. For relative quantification, �CT values werederived after normalization to SCR1 transcript levels, and ��CT values were derived with the 0-h sample as calibrator in CAP2�/� (SN152) and cap2�/� (RPC75)backgrounds. The average SFU1 mRNA expression ratio was derived from two independent RNA preparations and S.D. of the measurements shown as errorbars. B, Western blot analysis of Sfu1 protein expression. Cell extracts from C. albicans SFU1-TAP strains RPC391 (CAP2�/�), RPC422 (cap2�/�), and untaggedcontrol (SN152) cultured in either iron-deplete (SC-LIM plus 100 �M BPS) or in iron-replete (SC-LIM plus 100 �M FeCl3) media were resolved by SDS-PAGE,blotted, and probed with 1:1000 dilution of rabbit anti-TAP polyclonal antibody (Open Biosystems; top panel), or with 1:1000 dilution of monoclonal anti-�-tubulin antibody (Sigma; middle panel). The Ponceau S stain of the blot is also shown (bottom panel). C, the sfu1�/� cap2�/� double mutant showed the samephenotype as cap2�/� mutant in iron-depleted medium. C. albicans strains SN152, RPC75, RPC434 (sfu1�/�), and RPC474 (sfu1�/� cap2�/�) were used for thespot assay. D, qRT-PCR analysis of Cap2 target genes in wild-type (SN152) and sfu1�/� mutant (RPC434) strains grown under iron-replete (SC-LIM plus 100 �M
FeCl3) medium for 5 h.
Central Regulator of Iron Homeostasis
25164 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

mutant in BPS � FeCl3 medium (Fig. 6B). It needs to be seen ifHAP33 has any role in growth of C. albicans in BPS conditions,and if that contributed to the residual growth seen in thehap32� mutant. In any case, our data showed that HAP32,but not HAP31, had a predominant role for growth underiron-deprivation conditions. Recent reports showed thatorf19.1973/HAP5 (19, 20) and orf19.1228/HAP2 (20) arerequired for growth of C. albicans in BPS-containing medium.
Thus all three core HAP components HAP2, HAP32, andHAP5, are required for growth of C. albicans under iron-depri-vation conditions.Cap2 Target Gene Regulation Is Dependent on Core HAP
Components—To investigate the requirement of the core HAPcomplex in the regulation of Cap2-target gene expression, wealso measured transcript levels in hap2�, hap5�, and hap32�strains under iron-deprivation conditions. Data showed that in
FIGURE 6. Cap2-dependent differential expression of HAP genes. A, qRT-PCR analysis of HAP gene transcripts. For relative quantification, �CT values werederived after normalization to the SCR1 transcript levels, and ��CT values were derived using the BPS � FeCl3 sample as calibrator. The bar graph shows thefold-change (BPS versus BPS� FeCl3) of HAP gene transcripts in wild-type and cap2�/� background from two RNA preparations. B, HAP32 but not HAP31 isrequired for growth under iron-depleted medium. Strains WT (SN152), cap2�/� (RPC75), hap32�/� (RPC432), hap31�/� (RPC445), and hap32�/� hap31�/�(RPC449) were used. C, qRT-PCR analysis of Cap2 target genes in cap2�/� (RPC75), hap2�/� (087), hap5�/� (093), and hap32�/� (RPC432) and expressed asfold-change. Data are the average values from 2–3 RNA preparations and replicate assays.
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25165
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

both hap2� and hap5� strains, FRP1, SIT1, andHMX1 expres-sionwas impaired, andACO1 expressionwas elevated (Fig. 6C),indicating that both Hap5 and Hap2 subunits, in addition toCap2 are required for the regulation. However, the expressionof thesemRNAswas only slightly affected in the hap32� strain.We also investigated the requirement of the HAP complexcomponents for activation of the expression of HAP2, HAP32,andHAP5 genes. Data showed that only cap2� but not the hapmutants impaired the expression of the mRNA encoding HAPcomponents in iron-deprived medium (Fig. 6C). These datasuggests that Cap2 alone was required for activation of expres-sion of HAP complex core components. Interestingly, regula-tion of SFU1 expression was dependent on both Cap2 and theHAP complex core components.Cap2 Associates with HAP Complex in Whole Cell Extracts—
We hypothesized that the subset of Cap2/HAP43, Hap2,Hap32, and Hap5, by virtue of their functional requirementsunder iron-deprivation conditions, likely interacted in a com-plex to regulate iron homeostasis in C. albicans. To test thispossibility, we chromosomally tagged one allele of HAP5 at itscarboxyl terminus using a dual epitope tag comprising His6-FLAG3 and performed coimmunoprecipitation experiments.Immunoprecipitation of Hap5-His6-FLAG3 with anti-FLAGbeads could efficiently pulldownHap5 from tagged cell extractsbut not fromuntagged control cell extracts (Fig. 7). Upon prob-ing the blot with affinity-purified anti-Cap2 antibody, wefound that a significant fraction of Cap2 was coimmunopre-cipitated specifically from the tagged HAP5 strain but notfrom untagged control strain. Comparison of the band inten-sities in the input and the immunoprecipitated samples indi-cated that Cap2 and Hap5 proteins were not recovered inequivalent proportions in the immunoprecipitated samplesuggesting either a weaker association of Cap2 with Hap5 orthat a fraction of Cap2 exists outside of Hap5 in cell extracts.We also probed the blot with control glucose-6-phosphatedehydrogenase antibody and found that the glucose-6-phos-phate dehydrogenase band was detectable only in the input
but not in immunoprecipitated samples (Fig. 7). These dataled us to conclude that Cap2 associated with Hap5 in vivounder iron-deprivation conditions.Hap5 Is Recruited to Cap2 Target Gene Promoters in a
Hap32-dependent Manner—To assess promoter occupancyof the HAP complex using ChIP assay, we constructed aHAP5-TAP strain in wild-type and hap32� backgrounds. We ascer-tained that introduction of TAP tag did not disruptHAP5 func-tion by examining growth of theHAP5-TAP/HAP5-TAP strainon BPS medium (data not shown). The HAP5-TAP strainswere grown in BPS or BPS plus iron containing medium for5 h and the Hap5-TAP-bound chromatin was immunopre-cipitated. The immunopurified DNA was analyzed for Hap5target promoters by qRT-PCR. The data showed that theFRP1 promoter (�84 to �281 with respect to ATG) wasefficiently precipitated, �9.8-fold, as compared with a con-trol nonspecific region in BPS medium but not in iron-sup-plemented medium (Fig. 8). Interestingly, theHap5-TAP occu-pancy was completely abolished in the hap32� background. The�84 to�281 sequence contained the 5�-CCAAT-3� box at�138,shown previously to be important for activation of the FRP1-lacZreporter (19).Examination of the FRP1 promoter sequence showed two
additional CCAAT box motifs at �241 and �532 in theupstream region (data not shown). Therefore we examinedHap5-TAP occupancy at these locations as well. The datashowed �7.2- and �4.0-fold enrichment at both locations inthe FRP1 promoter (Fig. 8), indicating Hap5 recruitment to
FIGURE 7. Cap2 interacts with Hap5 in cell extracts. Protein extracts fromuntagged (SN152) and HAP5-FLAG (HAP5::His6-FLAG3; RPC552), cultured iniron-limiting condition, were subjected to immunoprecipitation using anti-FLAG M2 affinity gel. The IP and input (100 �g) samples were analyzed byWestern blotting. The bands marked with an asterisk in the IP lanes are non-specific bands and correspond to signals of IgG eluted from the IP beads; notethat the size of this species is also larger than the glyceraldehyde-6-phos-phate dehydrogenase (G6PDH) in the input lanes. FIGURE 8. Hap5 is recruited to FRP1 and ACO1 promoters. Chromatin
immunoprecipitation assay was performed using HAP5-TAP strains RPC453(HAP32/HAP32) and RPC562 (hap32�/hap32�) cultured in BPS or in BPS plusiron medium as described under “Experimental Procedures.” Total input DNAand immunoprecipitated DNA from multiple locations of the two target pro-moters FRP1 and ACO1, and a control non-coding region from chromosome 1(ca21chr1_1573500 –1574000) were analyzed by qRT-PCR. The �CT valueswere derived after normalization to input samples separately for the targetand control regions, and ��CT values were derived using the control regionas a calibrator. The fold-enrichment (calculated as 2���CT) for Hap5-TAP ateach location was expressed relative to that of the control location. ControlChIP reactions using untagged strain chromatin extracts yielded undetect-able signal demonstrating the specificity of the chromatin IP reactions.
Central Regulator of Iron Homeostasis
25166 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

multiple locations in this promoter. Furthermore, higheroccupancy was seen at the two proximal (�138, �241) FRP1promoter regions compared with the distal region examined.We also examined Hap5-TAP occupancy at the ACO1 pro-
moter in the same ChIP assay. The data showed high levels ofHap5 occupancy at twoACO1 upstream regions bearing CCAATmotifs at positions�250 and�658 (Fig. 8). The Hap5 occupancyat this promoter was alsoHap32-dependent (Fig. 8). Interestingly,Hap5occupancy, althoughelevated inBPSmedium,wasnot abro-gated in iron-supplementedmedium.ThusHap5 binding at FRP1and ACO1 promoters responded differently to iron status. Thesedata demonstrated thatHap5 is recruited to bothFRP1 andACO1promoters in vivo in a Hap32-dependent manner, providing amechanistic framework for a direct role for the Cap2-HAP com-plex in activation and repression of transcription of iron homeo-stasis genes.
DISCUSSION
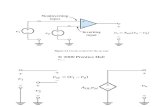
In this study, we demonstrate that the Cap2-HAP complexis the central and a critical regulator of iron homeostasis inC. albicans. First, CAP2 is required for growth of C. albicansin iron-limited medium in vitro and for virulence in themouse model. The requirement of CAP2, also called HAP43,for growth under iron-deprivation conditions was indepen-dently identified in other studies (19, 20, 61). Our mutationalanalyses showed that both bZIP and HAP4L domains arerequired for Cap2 function. Second, CAP2 expression is up-regulated in iron-limited medium and repressed in iron-re-plete medium. Third, transcriptome profiling indicated thatCAP2 regulates iron homeostasis chiefly by transcriptionalactivation of genes in three iron uptake pathways and repres-sion of iron utilizing and iron storage pathway genes. Fourth,several lines of evidence indicated that Cap2 functions in acomplex with the core HAP complex, namely Cap2-depen-dent up-regulation of HAP32, HAP2, and HAP5, require-ment of the HAP components for regulation of Cap2 targetgenes, and association of Cap2 with Hap5 in cell extracts.Finally, iron deprivation elicited Hap5 recruitment to FRP1(Cap2-induced) and ACO1 (Cap2-repressed) promoters in aHap32-dependent manner, indicating a direct role in vivo forthe Cap2-HAP complex for regulation of iron homeostasis.
Transcriptional Regulation of Cap2 Expression
Iron deprivation lead to expression and activation of Cap2,which has a Yap-like bZIP domain and the fungal-specificHAP4-like domain (Fig. 9). The iron-dependent regulation ofCAP2 expression was first noted in a C. albicans microarraystudy (14), and indeed for CAP2 orthologs in other systems aswell (17, 21, 79). In iron-replete media, although the CAP2mRNA level was diminished to a low basal level, the Cap2 pro-tein level was reduced to an undetectable level (Fig. 4B), indi-cating a tighter control of Cap2 protein expression. However,the mechanism(s) controlling the expression of Cap2 in iron-depleted or iron-replete conditions are not understood. Exam-ination of the Cap2 amino acid sequence showedmultiple Cys-rich regions (Fig. 1B); it needs to be determined if one or moreof the Cys-rich regions are required for Cap2 activity in vivo. Inany case, because Cap2 expression is tightly controlled at both
mRNA and protein levels, together our data suggests that reg-ulation of Cap2 expression is the primary step in the iron-de-privation response by C. albicans.
Cap2-dependent Regulation of Iron HomeostasisTranscriptome
Microarray data revealed Cap2-dependent up-regulationof numerous genes including eight iron-uptake genesbelonging to three pathways, viz., siderophore, reductiveiron, and heme uptake. Although most of the Cap2-inducediron uptake genes were also induced upon iron depletion(14), our data identified Cap2-dependent induction of addi-tional genes in the iron uptake pathway. Interestingly, atleast three of the iron uptake genes, FRE1, FRE2, and FTH1,that were induced upon iron deprivation in a wild-type straindid not show Cap2 dependence indicating other modes ofregulation of this subset of genes. Our microarray data alsorevealed that genes encoding iron-utilizing proteins such asFe-S proteins and heme proteins and numerous other genesare repressed in a Cap2-dependent manner (Fig. 9). More-over, iron storage genes are also repressed in a Cap2-depen-dent manner, likely contributing to mobilization of ironfrom endogenous pools.
Cap2-HAP Regulatory Network
Feedback Regulation of the Sfu1Negative Regulator—Howdocells deal with the negative regulator Sfu1 under iron-depriva-tion conditions? First, in a feedback loop, the expression ofSFU1 is repressed in a Cap2-dependent manner (Fig. 9). TheCap2 control seems to be limited to iron-deprivation condi-tions because under high iron conditions SFU1mRNAandpro-tein levels were comparable in wild-type and cap2� strains.Second, it seems likely that any residual Sfu1 protein could alsobe inactive in the absence of iron, as shown previously for itsortholog Sre1 in H. capsulatum (80). Although deletion ofSFU1 led to 4-fold higher expression of CAP2mRNA (Fig. 5D),but not Cap2 protein levels, the CAP2 target genes are notinduced. These data suggest that regulation of Cap2 expressionand function operate atmultiple levels possibly to enforce strin-gent regulation of iron homeostasis. Furthermore, expressionanalysis suggested that Sfu1 seems to exert its repressive activ-ity predominantly at theCap2 promoter, but not at the promot-ers of the core HAP complex components.Feed-forward Regulation and Cap2-HAP Complex Forma-
tion—We obtained several lines of evidence indicating thatCap2 functions in a complex with the Hap2-Hap32-Hap5 com-plex in vivo. Iron deprivation led to Cap2-dependent activationof HAP2 and HAP32 and expression of HAP5 genes (Fig. 9).Although low iron-elicited up-regulation ofHAP32 expressionwas previously found (14), Cap2 dependence was not known.Our data also showed that although HAP5 expression was notregulated by iron availability in wild-type cells, Cap2 proteinwas required to maintain HAP5 mRNA levels during iron-de-privation conditions. Second, mutational analyses showed thatboth bZIP and HAP4L domains are required for Cap2 functionin iron-deprivation response. Remarkably, only Cap2, but notthe HAP core subunits, was required for low iron-elicited acti-vation of coreHAP components revealing an additional facet of
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25167
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

Cap2 function. Third, we showed that Cap2 could be efficientlycoimmunoprecipitated with Hap5 from cell extracts preparedfrom cultures grown under iron-deprivation conditions.Although previous studies indicated an interaction of Hap4-like proteins with Hap2-Hap3-Hap5 proteins from C. albicans(19, 27), our study is the first biochemical demonstration ofCap2-HAP interaction in cell extracts under iron-deprivationconditions. A recent yeast two-hybrid study also showed inter-action betweenH. polymorphaHap4b and Hap5 (21). Thus theHAP4Ldomain alone could be sufficient tomediateCap2 inter-action with Hap5. In this respect, the C. albicans Hap5/orf19.1973 also contains the Hap4-interaction domain previ-ously identified in S. cerevisiae (29).Genetic studies showed a requirement ofHAP32,HAP2, and
HAP5 for the growth of C. albicans in iron-limited medium(Fig. 4B) (19, 20). Moreover, the core HAP complex compo-
nents were also required for the expression of Cap2 targetgenes, thereby providing functional support for HAP com-ponents in iron homeostasis regulation (Fig. 9). Structuralstudies using mammalian NF-YB and NF-YC heterodimers,the orthologs of Hap3 and Hap5 proteins, respectively,showed that their interaction occurred through histoneH2A/H2B-like histone-fold domains (81). Moreover, theHap32 and Hap5 proteins also could heterodimerizethrough their histone-fold domains (as reviewed in Ref. 82),and in association with Hap2 could constitute the core HAPregulatory complex. Such a mechanism for complex forma-tion has previously been reported for the S. cerevisiae Hap2-Hap3-Hap5 complex (29). Although previous studiesshowed that interaction between S. cerevisiae Hap4 andHap5-Hap3-Hap2 required DNA (24), data from the presentstudy and from H. polymorpha (21) indicated that DNA may
FIGURE 9. Summary of the Cap2-HAP-mediated regulation of iron homeostasis in C. albicans. Iron deprivation strongly elicited the induction of Cap2mRNA and protein expression (thick broken arrows); in contrast, in iron containing medium the Cap2 expression was repressed. Both HAP4L and bZIP domainsare required for Cap2 function. Mutational analyses showed that HAP32, HAP2, and HAP5 are required for C. albicans growth in iron-depleted medium.Expression analysis showed Cap2-dependent activation of HAP2 and HAP32 expression; additionally, HAP5 expression was also dependent on Cap2. Irondeprivation also elicited partial induction (thin broken arrows) of HAP32 expression in the cap2� mutant strain indicating a role for other transcriptionalregulator(s). Cap2, but not Hap2, Hap5, and Hap32, was required for induction of HAP2, HAP32, and HAP5 expression in low iron medium. Cap2 functionsin association with the HAP complex as Cap2 was coimmunoprecipitated with Hap5 from cell extracts. Furthermore, SFU1 mRNA expression wasrepressed in a Cap2-HAP complex dependent manner. Because sfu1� mutation led to derepression of CAP2 mRNA expression but did not induce theCap2 target genes, Sfu1 seems to primarily mediate Cap2 repression, and in its wake leading to the repression of iron uptake gene expression in highiron medium. Transcriptome profiling showed Cap2-dependent up-regulation of 10 iron uptake genes and about 410 genes in several functionalcategories. Additionally, the transcriptome data also identified Cap2-dependent down-regulation of 16 genes involved in iron utilization or that requireiron for its activity, and 493 other genes representing several functional categories. Chromatin IP assays showed Hap5 occupancy to the promoters ofboth Cap2-induced (FRP1) and Cap2-repressed (ACO1) genes, thus indicating that Cap2-HAP complex functions by direct promoter binding to up- ordown-regulate gene expression to maintain iron homeostasis.
Central Regulator of Iron Homeostasis
25168 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

not be an absolute requirement at least for the HAP4Ldomain protein interaction with Hap5.
Dual Regulation by Cap2-HAP Complex Involves DirectPromoter Binding
Previous studies on Cap2-HAP orthologs indicated amechanism of repression of genes encoding iron-utilizingproteins under iron-deprivation conditions such asAspergil-lus nidulansHapX (17) and S. pombe Php4 (79). Therefore tounderstand the mechanism of Cap2-HAP function, we car-ried out a in vivo ChIP assay and demonstrated promoteroccupancy of Hap5 to the CCAAT box containing regions ofthe ACO1 gene promoter in vivo under both iron-depletedand iron-replete media. Thus although HAP complex bind-ing to the ACO1 promoter is not regulated by iron availabil-ity, the expression of this promoter is differentially regu-lated. In this respect Cap2 association with the HAP complexin iron-depleted medium is the key regulatory switch torepress ACO1 expression. This conclusion is also strength-ened by our data that CaCTH2/19.5334 does not have a sub-stantial role in ACO1 repression.A previous study showed that the CCAATbox is required for
activated expression of FRP1-lacZ reporter in C. albicans (19).Moreover, EMSA studies have shown that one or more Hap5-containing complexes can interact with CCAAT box contain-ing promoters (19, 27). Indeed using ChIP analysis, we havedemonstrated Hap32-dependent occupancy of Hap5 in vivo toFRP1 promoter regions bearing CCAAT box in iron-deprivedmedium. Interestingly, the Hap5 occupancy at FRP1 was notfound in iron-replete medium. Thus, in contrast to the ACO1promoter, Hap5 occupancy at the FRP1 promoter is differen-tially regulated by iron availability. Herein Hap32 was impor-tant for Hap5 promoter occupancy. At odds with the Hap32requirement for Hap5 occupancy, the Cap2-stimulated expres-sion of FRP1was weakly dependent on Hap32. It is conceivablethat absence of Hap32 weakened Hap5 occupancy, but theHap5 associationwas sufficient to drive FRP1 expression, albeitat reduced levels.A simplistic model from our data is that under iron-depri-
vation conditions, the Cap2-HAP complex binds to and acti-vates the FRP1 promoter by direct recruitment of the transcrip-tional machinery. An alternative model is that Cap2-HAPfunctions as an anti-repressor by promoter binding, whichcould lead to destabilization of a putative repressor under low-iron conditions and consequent assembly of the transcriptionalmachinery. In any case, Cap2-dependent expression of the coreHAP complex components HAP5, HAP32, and HAP2, andrepression of the negative regulator Sfu1, and the large scope ofthe Cap2 regulon together indicate that Cap2 functions as amaster regulator of iron homeostasis. Thus regulation of Cap2expression by iron availability appears to be a pivotal step in theCap2/HAP regulatory cascade to mediate iron homeostasis inC. albicans. Moreover, the dual but contrasting regulation ofiron acquisition and iron storage pathways by the Cap2-HAPregulatory complex suggests a robust coordination of the cellu-lar machinery to maintain iron homeostasis during iron depri-vation in this important human fungal pathogen.
Acknowledgments—We thank Drs. S. Noble and A. D. Johnson, Dr. J.Morschhauser, Dr. J.Wendland,Dr. A. J. Brown, andDr. P. Koetter forplasmids and strains; Dr. S. Noble for advice on fusion PCR protocol;Dr. K. Ganesan for reagents and advice; Dr. M. Heinz and Chris Saw-yer, Washington University, for supply of the C. albicans microarrayslides and advice; Ankhee Dutta for assistance with the microarrayheat map. We thank Dr. C. C. Philpott for advice and helpful com-ments, Dr. R. S. Gokhale and Dr. A. Bhattacharya for stimulatingdiscussions and suggestions on the manuscript, Dr. A. Datta forencouragement and support, and all members of the lab for discus-sions and reading of the manuscript, and the three anonymousreviewers for constructive suggestions.
REFERENCES1. Martchenko, M., Levitin, A., Hogues, H., Nantel, A., and Whiteway, M.
(2007) Curr. Biol. 17, 1007–10132. Tsong, A. E., Tuch, B. B., Li, H., and Johnson, A. D. (2006) Nature 443,
415–4203. Wessling-Resnick, M. (1999) Crit. Rev. Biochem. Mol. Biol. 34, 285–3144. Moye-Rowley, W. S. (2003) Eukaryot. Cell 2, 381–3895. Masse, E., and Arguin, M. (2005) Trends Biochem. Sci. 30, 462–4686. Philpott, C. C. (2006) Biochim. Biophys. Acta 1763, 636–6457. Rutherford, J. C., Jaron, S., and Winge, D. R. (2003) J. Biol. Chem. 278,
27636–276438. Philpott, C. C., and Protchenko, O. (2008) Eukaryot. Cell 7, 20–279. Yamaguchi-Iwai, Y., Ueta, R., Fukunaka, A., and Sasaki, R. (2002) J. Biol.
Chem. 277, 18914–1891810. Rutherford, J. C., Ojeda, L., Balk, J., Muhlenhoff, U., Lill, R., and Winge,
D. R. (2005) J. Biol. Chem. 280, 10135–1014011. Puig, S., Askeland, E., and Thiele, D. J. (2005) Cell 120, 99–11012. Haas, H., Eisendle, M., and Turgeon, B. G. (2008)Annu. Rev. Phytopathol.
46, 149–18713. Liang, Y., Wei, D., Wang, H., Xu, N., Zhang, B., Xing, L., and Li, M. (2010)
Microbiology 156, 2912–291914. Lan, C. Y., Rodarte, G., Murillo, L. A., Jones, T., Davis, R. W., Dungan, J.,
Newport, G., and Agabian, N. (2004)Mol. Microbiol. 53, 1451–146915. Rutherford, J. C., and Bird, A. J. (2004) Eukaryot. Cell 3, 1–1316. Labbe, S., Pelletier, B., and Mercier, A. (2007) Biometals 20, 523–53717. Hortschansky, P., Eisendle, M., Al-Abdallah, Q., Schmidt, A. D., Berg-
mann, S., Thon,M., Kniemeyer, O., Abt, B., Seeber, B.,Werner, E. R., Kato,M., Brakhage, A. A., and Haas, H. (2007) EMBO J. 26, 3157–3168
18. Mercier, A., Watt, S., Bahler, J., and Labbe, S. (2008) Eukaryot. Cell 7,493–508
19. Baek, Y. U., Li, M., and Davis, D. A. (2008) Eukaryot. Cell 7, 1168–117920. Homann, O. R., Dea, J., Noble, S. M., and Johnson, A. D. (2009) PLoS
Genet. 5, e100078321. Sybirna, K., Petryk, N., Zhou, Y. F., Sibirny, A., and Bolotin-Fukuhara, M.
(2010) Yeast 27, 941–95422. Mantovani, R. (1999) Gene 239, 15–2723. Forsburg, S. L., and Guarente, L. (1989) Genes Dev. 3, 1166–117824. McNabb, D. S., Xing, Y., and Guarente, L. (1995) Genes Dev. 9, 47–5825. Kaplan, J.,McVeyWard,D., Crisp, R. J., and Philpott, C. C. (2006)Biochim.
Biophys. Acta 1763, 646–65126. Sybirna, K., Guiard, B., Li, Y. F., Bao, W. G., Bolotin-Fukuhara, M., and
Delahodde, A. (2005) Curr. Genet. 47, 172–18127. Johnson, D. C., Cano, K. E., Kroger, E. C., and McNabb, D. S. (2005)
Eukaryot. Cell 4, 1662–167628. Kornitzer, D. (2009) Curr. Opin. Microbiol. 12, 377–38329. McNabb, D. S., and Pinto, I. (2005) Eukaryot. Cell 4, 1829–183930. Schaible, U. E., and Kaufmann, S. H. (2004) Nat. Rev. Microbiol. 2,
946–95331. Sutak, R., Lesuisse, E., Tachezy, J., and Richardson, D. R. (2008) Trends
Microbiol. 16, 261–26832. Almeida, R. S., Wilson, D., and Hube, B. (2009) FEMS Yeast Res. 9,
1000–1012
Central Regulator of Iron Homeostasis
JULY 15, 2011 • VOLUME 286 • NUMBER 28 JOURNAL OF BIOLOGICAL CHEMISTRY 25169
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

33. Ramanan, N., and Wang, Y. (2000) Science 288, 1062–106434. Hu, C. J., Bai, C., Zheng, X. D., Wang, Y. M., and Wang, Y. (2002) J. Biol.
Chem. 277, 30598–3060535. Weissman, Z., and Kornitzer, D. (2004)Mol. Microbiol. 53, 1209–122036. Almeida, R. S., Brunke, S., Albrecht, A., Thewes, S., Laue, M., Edwards,
J. E., Filler, S. G., and Hube, B. (2008) PLoS Pathog. 4, e100021737. Noble, S. M., and Johnson, A. D. (2005) Eukaryot. Cell 4, 298–30938. Reuss, O., Vik, A., Kolter, R., and Morschhauser, J. (2004) Gene 341,
119–12739. Murad, A. M., Lee, P. R., Broadbent, I. D., Barelle, C. J., and Brown, A. J.
(2000) Yeast 16, 325–32740. Gola, S., Martin, R., Walther, A., Dunkler, A., and Wendland, J. (2003)
Yeast 20, 1339–134741. Fairhead, C., Llorente, B., Denis, F., Soler, M., and Dujon, B. (1996) Yeast
12, 1439–145742. Schaub, Y., Dunkler, A., Walther, A., and Wendland, J. (2006) J. Basic
Microbiol. 46, 416–42943. McCluskey, K., Wiest, A., and Plamann, M. (2010) J. Biosci. 35, 119–12644. Park, Y. N., and Morschhauser, J. (2005) Eukaryot. Cell 4, 1328–134245. Corvey, C., Koetter, P., Beckhaus, T., Hack, J., Hofmann, S., Hampel, M.,
Stein, T., Karas, M., and Entian, K. D. (2005) J. Biol. Chem. 280,25323–25330
46. Wu, P. Y., and Winston, F. (2002)Mol. Cell. Biol. 22, 5367–537947. Kohrer, K., and Domdey, H. (1991) in Methods in Enzymology: Guide to
Yeast Genetics and Molecular Biology (Guthrie, C., and Fink, G. R., eds)Vol. 194, pp. 398–405, Academic Press, Inc., San Diego
48. Marck, C., Kachouri-Lafond, R., Lafontaine, I., Westhof, E., Dujon, B., andGrosjean, H. (2006) Nucleic Acids Res. 34, 1816–1835
49. Schmittgen, T. D., and Livak, K. J. (2008) Nat. Protoc. 3, 1101–110850. Brown, V., Sexton, J. A., and Johnston, M. (2006) Eukaryot. Cell 5,
1726–173751. Yatskou, M., Novikov, E., Vetter, G., Muller, A., Barillot, E., Vallar, L., and
Friederich, E. (2008) BMC Res. Notes 1, 8052. Tusher, V. G., Tibshirani, R., and Chu, G. (2001) Proc. Natl. Acad. Sci.
U.S.A. 98, 5116–512153. Kadosh, D., and Johnson, A. D. (2005)Mol. Biol. Cell 16, 2903–291254. Maere, S., Heymans, K., and Kuiper, M. (2005) Bioinformatics 21,
3448–344955. Cline, M. S., Smoot, M., Cerami, E., Kuchinsky, A., Landys, N., Workman,
C., Christmas, R., Avila-Campilo, I., Creech, M., Gross, B., Hanspers, K.,Isserlin, R., Kelley, R., Killcoyne, S., Lotia, S., Maere, S., Morris, J., Ono, K.,Pavlovic, V., Pico, A. R., Vailaya, A., Wang, P. L., Adler, A., Conklin, B. R.,Hood, L., Kuiper, M., Sander, C., Schmulevich, I., Schwikowski, B., War-ner, G. J., Ideker, T., and Bader, G. D. (2007) Nat. Protoc. 2, 2366–2382
56. Zordan, R. E.,Miller,M. G., Galgoczy, D. J., Tuch, B. B., and Johnson, A. D.(2007) PLoS Biol. 5, e256
57. Fernandes, L., Rodrigues-Pousada, C., and Struhl, K. (1997)Mol. Cell. Biol.17, 6982–6993
58. Alarco, A. M., and Raymond, M. (1999) J. Bacteriol. 181, 700–70859. Alarco, A. M., Balan, I., Talibi, D., Mainville, N., and Raymond, M. (1997)
J. Biol. Chem. 272, 19304–1931360. Yang, X., Talibi, D.,Weber, S., Poisson, G., and Raymond,M. (2001) Yeast
18, 1217–122561. Hsu, P. C., Yang, C. Y., and Lan, C. Y. (2011) Eukaryot. Cell 10, 207–22562. Jung,W.H., Saikia, S., Hu, G.,Wang, J., Fung, C. K., D’Souza, C.,White, R.,
and Kronstad, J. W. (2010) PLoS Pathog. 6, e100120963. Schrettl,M., Beckmann,N., Varga, J., Heinekamp, T., Jacobsen, I. D., Jochl,
C., Moussa, T. A., Wang, S., Gsaller, F., Blatzer, M., Werner, E. R., Nier-mann, W. C., Brakhage, A. A., and Haas, H. (2010) PLoS Pathog. 6,e1001124
64. Fujii, Y., Shimizu, T., Toda, T., Yanagida, M., and Hakoshima, T. (2000)Nat. Struct. Biol. 7, 889–893
65. Pu, W. T., and Struhl, K. (1991)Mol. Cell. Biol. 11, 4918–492666. Natarajan, K., Marton, M. J., and Hinnebusch, A. G. (2007) inMethods for
General and Molecular Microbiology (Reddy, C. A., Beveridge, T. J., Br-eznak, J. A., Snyder, L., Schmidt, T. M., and Marzluf, G. A., eds) pp.978–994, ASM Press, Washington, D. C.
67. Bowtell, D., and Sambrook, J. (2003)DNAMicroarrays: AMolecular Clon-ing Manual, Cold Spring Harbor Press, Cold Spring Harbor, NY
68. Shakoury-Elizeh, M., Protchenko, O., Berger, A., Cox, J., Gable, K., Dunn,T.M., Prinz,W. A., Bard,M., and Philpott, C. C. (2010) J. Biol. Chem. 285,14823–14833
69. Martchenko,M., Alarco, A.M., Harcus, D., andWhiteway,M. (2004)Mol.Biol. Cell 15, 456–467
70. Srinivasan, C., Liba, A., Imlay, J. A., Valentine, J. S., and Gralla, E. B. (2000)J. Biol. Chem. 275, 29187–29192
71. De Freitas, J. M., Liba, A., Meneghini, R., Valentine, J. S., and Gralla, E. B.(2000) J. Biol. Chem. 275, 11645–11649
72. Heymann, P., Gerads, M., Schaller, M., Dromer, F., Winkelmann, G., andErnst, J. F. (2002) Infect. Immun. 70, 5246–5255
73. Knight, S. A., Lesuisse, E., Stearman, R., Klausner, R. D., and Dancis, A.(2002)Microbiology 148, 29–40
74. Knight, S. A., Vilaire, G., Lesuisse, E., and Dancis, A. (2005) Infect. Immun.73, 5482–5492
75. Santos, R., Buisson, N., Knight, S., Dancis, A., Camadro, J. M., and Lesu-isse, E. (2003)Microbiology 149, 579–588
76. Weissman, Z., Shemer, R., Conibear, E., and Kornitzer, D. (2008) Mol.Microbiol. 69, 201–217
77. Singh, A., Kaur, N., and Kosman, D. J. (2007) J. Biol. Chem. 282,28619–28626
78. Santos, R., Buisson, N., Knight, S. A., Dancis, A., Camadro, J. M., andLesuisse, E. (2004)Mol. Microbiol. 54, 507–519
79. Mercier, A., Pelletier, B., and Labbe, S. (2006) Eukaryot. Cell 5, 1866–188180. Chao, L. Y., Marletta, M. A., and Rine, J. (2008) Biochemistry 47,
7274–728381. Romier, C., Cocchiarella, F., Mantovani, R., and Moras, D. (2003) J. Biol.
Chem. 278, 1336–134582. Brakhage, A. A., Andrianopoulos, A., Kato, M., Steidl, S., Davis, M. A.,
Tsukagoshi, N., and Hynes, M. J. (1999) Fungal Genet. Biol. 27, 243–252
Central Regulator of Iron Homeostasis
25170 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 28 • JULY 15, 2011
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from

Krishnamurthy NatarajanRana Pratap Singh, Himanshu K. Prasad, Ishani Sinha, Neha Agarwal and
Candida albicansContrasting Roles in Regulation of Iron Homeostasis in Cap2-HAP Complex Is a Critical Transcriptional Regulator That Has Dual but
doi: 10.1074/jbc.M111.233569 originally published online May 18, 20112011, 286:25154-25170.J. Biol. Chem.
10.1074/jbc.M111.233569Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2011/05/19/M111.233569.DC1
http://www.jbc.org/content/286/28/25154.full.html#ref-list-1
This article cites 80 references, 35 of which can be accessed free at
by guest on Novem
ber 26, 2020http://w
ww
.jbc.org/D
ownloaded from