
Can ascospore ultrastructure differentiate between the genera Linocarpon and Neolinocarpon and...
9
Can ascospore ultrastructure differentiate between the genera Linocarpon and Neolinocarpon and species therein ? YANNA, Wai Hong HO and Kevin D. HYDE* Department of Ecology & Biodiversity, Centre for Research in Fungal Diversity, The University of Hong Kong, Pokfulam Road, Hong Kong SAR, People’s Republic of China. E-mail : [email protected] Received 24 April 2002; accepted 12 August 2003. Ascospore appendages of two species of Linocarpon and three species of Neolinocarpon are illustrated using electron microscopy. The structure of these appendages can be used to distinguish between species. The ascospore appendages of species in both genera, however, were found to have similar structure. Thus, species of Linocarpon and Neolinocarpon could not be delineated at the generic level based on ascospore appendage structure. INTRODUCTION Linocarpon (Hyponectriaceae) was introduced based on L. pandani (syn. Linospora pandani) (Sydow & Sydow 1917). Neolinocarpon was introduced to accommodate Linocarpon-like species with deeply immersed ascomata forming below a slightly raised or flattened clypeus, and with a refractive globose body below the apical ascus ring (Hyde 1992a). Linocarpon was restricted to species with superficial or slightly immersed ascomata, and the lack of a refractive globose body (Hyde 1992b, Hyde, Taylor & Fro¨hlich 1998). Species of Linocarpon and Neolinocarpon are very similar in producing 8-spored, cylindrical asci with a non-amyloid cylindri- cal apical ring, and filiform, hyaline, usually unicellular, curved ascospores with refringent septum-like bands (Hyde 1988, 1992a, b, 1997, Dulymamode, Cannon & Peerally 1998, Hyde, Taylor & Fro¨hlich 1998). Linocarpon presently comprises 35 species, while Neo- linocarpon comprises eight species (Hyde 1988, 1992a, 1997, Dulymamode et al. 1998, Hyde & Alias 1999, Thongkantha et al. 2003). Both genera are currently placed in the Hyponectriaceae, although Wang & Hyde (1999) suggest their inclusion is unlikely. Species of Linocarpon and Neolinocarpon are ident- ified based on the size of the asci and ascospores, the shape of ascospores, and the presence of ascospore appendages. Ascospores of these two genera are very thin, ranging from 2 to 6 mm. It is therefore difficult to exa- mine the ascospore poles under the light microscope. Poonyth et al. (2000) examined Linocarpon appendiculatum and L. nipae at the ultrastructural level and showed that the morphology of the appendages was a useful character in the delineation of species in Linocarpon. Therefore, to establish further criteria to delineate species of Linocarpon and Neolinocarpon, ascospore appendages of selected species were exam- ined at the electron microscope level. The presence of a refractive globose body below the ascus apical ring in Neolinocarpon appears to be in- consistent, as in species of Lasiosphaeria (Hyde, Taylor & Fro¨hlich 2000). Refractive globose bodies were re- corded in the asci of Neolinocarpon globosicarpon (type species), N. australiense, N. calamae, N. eutypoides, and N. nonappendiculatus (Hyde 1992a, Hyde et al. 1998), but were absent in N. enshiensis, N. inconspicuus and N. nypicola (Hyde 1992a, 1997, Hyde et al. 1998, Hyde & Alias 1999). Therefore, delineation of Linocarpon and Neolinocarpon based on the presence and absence of the refractive globose bodies is unreliable, although no species of Linocarpon has asci with subapical re- fractive globose bodies. Linocarpon and Neolinocarpon species are therefore solely delineated based on the position of the ascomata, i.e. deeply immersed vs superficial. Fro¨hlich & Hyde (2000) have discussed the need to establish fur- ther criteria to delineate species of Linocarpon and Neolinocarpon at the generic level. Ascospore ap- pendage structure of selected species is, therefore, examined at the ultrastructural level. The terminology of Kirk (1966) is used to define the wall layers of * Corresponding author. Mycol. Res. 107 (11): 1305–1313 (November 2003). f The British Mycological Society 1305 DOI: 10.1017/S095375620300861X Printed in the United Kingdom.
Transcript of Can ascospore ultrastructure differentiate between the genera Linocarpon and Neolinocarpon and...

Can ascospore ultrastructure differentiate between the genera
Linocarpon and Neolinocarpon and species therein?
YANNA, Wai Hong HO and Kevin D. HYDE*
Department of Ecology & Biodiversity, Centre for Research in Fungal Diversity, The University of Hong Kong,Pokfulam Road, Hong Kong SAR, People’s Republic of China.E-mail : [email protected]
Received 24 April 2002; accepted 12 August 2003.
Ascospore appendages of two species of Linocarpon and three species of Neolinocarpon are illustrated using electron
microscopy. The structure of these appendages can be used to distinguish between species. The ascospore appendages ofspecies in both genera, however, were found to have similar structure. Thus, species of Linocarpon and Neolinocarponcould not be delineated at the generic level based on ascospore appendage structure.
INTRODUCTION
Linocarpon (Hyponectriaceae) was introduced based onL. pandani (syn. Linospora pandani) (Sydow & Sydow1917). Neolinocarpon was introduced to accommodateLinocarpon-like species with deeply immersed ascomataforming below a slightly raised or flattened clypeus,and with a refractive globose body below the apicalascus ring (Hyde 1992a). Linocarpon was restricted tospecies with superficial or slightly immersed ascomata,and the lack of a refractive globose body (Hyde 1992b,Hyde, Taylor & Frohlich 1998). Species of Linocarponand Neolinocarpon are very similar in producing8-spored, cylindrical asci with a non-amyloid cylindri-cal apical ring, and filiform, hyaline, usually unicellular,curved ascospores with refringent septum-like bands(Hyde 1988, 1992a, b, 1997, Dulymamode, Cannon& Peerally 1998, Hyde, Taylor & Frohlich 1998).Linocarpon presently comprises 35 species, while Neo-linocarpon comprises eight species (Hyde 1988, 1992a,1997, Dulymamode et al. 1998, Hyde & Alias 1999,Thongkantha et al. 2003). Both genera are currentlyplaced in the Hyponectriaceae, although Wang & Hyde(1999) suggest their inclusion is unlikely.
Species of Linocarpon and Neolinocarpon are ident-ified based on the size of the asci and ascospores, theshape of ascospores, and the presence of ascosporeappendages. Ascospores of these two genera are verythin, ranging from 2 to 6 mm. It is therefore difficult toexa-
mine the ascospore poles under the light microscope.Poonyth et al. (2000) examined Linocarponappendiculatum and L. nipae at the ultrastructural leveland showed that the morphology of the appendageswas a useful character in the delineation of species inLinocarpon. Therefore, to establish further criteria todelineate species of Linocarpon and Neolinocarpon,ascospore appendages of selected species were exam-ined at the electron microscope level.
The presence of a refractive globose body below theascus apical ring in Neolinocarpon appears to be in-consistent, as in species of Lasiosphaeria (Hyde, Taylor& Frohlich 2000). Refractive globose bodies were re-corded in the asci of Neolinocarpon globosicarpon (typespecies), N. australiense,N. calamae, N. eutypoides, andN. nonappendiculatus (Hyde 1992a, Hyde et al. 1998),but were absent in N. enshiensis, N. inconspicuus andN. nypicola (Hyde 1992a, 1997, Hyde et al. 1998, Hyde& Alias 1999). Therefore, delineation of Linocarponand Neolinocarpon based on the presence and absenceof the refractive globose bodies is unreliable, althoughno species of Linocarpon has asci with subapical re-fractive globose bodies.
Linocarpon and Neolinocarpon species are thereforesolely delineated based on the position of the ascomata,i.e. deeply immersed vs superficial. Frohlich & Hyde(2000) have discussed the need to establish fur-ther criteria to delineate species of Linocarpon andNeolinocarpon at the generic level. Ascospore ap-pendage structure of selected species is, therefore,examined at the ultrastructural level. The terminologyof Kirk (1966) is used to define the wall layers of* Corresponding author.
Mycol. Res. 107 (11): 1305–1313 (November 2003). f The British Mycological Society 1305
DOI: 10.1017/S095375620300861X Printed in the United Kingdom.
ascospores : mesosporium (the innermost layer) ; epi-sporium (the middle layer, except where the exosporiumis absent and generally the first formed layer) ; andexosporium (the outer layer). The term appendageis used to describe structures at the ends of theascospores. Ultrastructural work was carried out with
the following objectives : (1) to determine and com-pare the ultrastructure of ascospore appendages ofselected taxa in the two genera; and (2) to determineif species in both genera can be delineated at the gen-eric level by the ultrastructure of ascospore append-ages.
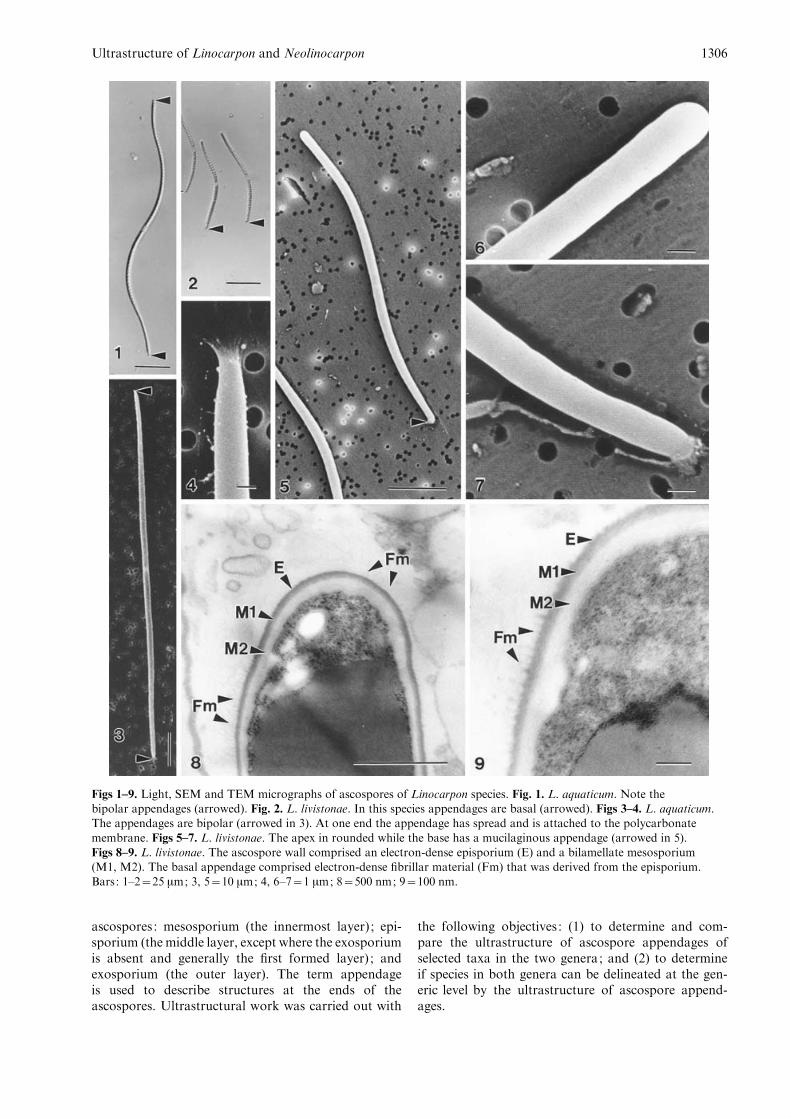
Figs 1–9. Light, SEM and TEM micrographs of ascospores of Linocarpon species. Fig. 1. L. aquaticum. Note thebipolar appendages (arrowed). Fig. 2. L. livistonae. In this species appendages are basal (arrowed). Figs 3–4. L. aquaticum.The appendages are bipolar (arrowed in 3). At one end the appendage has spread and is attached to the polycarbonate
membrane. Figs 5–7. L. livistonae. The apex in rounded while the base has a mucilaginous appendage (arrowed in 5).Figs 8–9. L. livistonae. The ascospore wall comprised an electron-dense episporium (E) and a bilamellate mesosporium(M1, M2). The basal appendage comprised electron-dense fibrillar material (Fm) that was derived from the episporium.
Bars: 1–2=25 mm; 3, 5=10 mm; 4, 6–7=1 mm; 8=500 nm; 9=100 nm.
Ultrastructure of Linocarpon and Neolinocarpon 1306

MATERIAL AND METHODS
Light microscopy (LM)
Freshly collected decaying palm samples were in-cubated separately in plastic bags lined with moistpaper towels and examined within one month. Fungalfruit bodies were mounted on slides in water or lacto-phenol for microscopic examination. The collectiondetails of material examined are listed in Table 1.
Scanning electron microscopy (SEM)
Fresh samples were fixed with 2% (w/v) osmiumtetroxide at 4 xC overnight. Samples were then washedwith distilled water, dehydrated through a gradedalcohol series to absolute alcohol, followed by criticalpoint drying, sputter-coating with gold/palladium for40 s, and examined using a Leica Cambridge Stereo-scan 440 scanning electron microscope operating at20 kV.
Transmission electron microscopy (TEM)
Fresh samples were embedded in 2% (w/v) ion agar,fixed with 4% (v/v) glutaraldehyde at room tempera-ture for 4 h, and post-fixed with 2% (w/v) osmiumtetroxide at 4 x overnight. Fixed samples were washedwith distilled water and dehydrated through a gradedalcohol series and acetone series. Dehydrated sampleswere embedded in Mollenhauer’s resin (Mollenhauer1964) through a series : 25, 50, 75 and 90% resin inacetone for 36 h for each concentration. Samples werethen transferred to 100% resin and infiltrated in avacuum chamber at 20 cmHg for 4 h, followed bypolymerization at 60 x for 3 d. Ultra-thin sections werestained with lead citrate for 15 min and post-stained
Table 1. Collection details of material examined.
Fungus Site Substratum Date
Herbarium
assession no.
Linocarpon aquaticum Australia, North Queensland,
Atherton Tablelands
Decaying rachis of
Oraniopsis appendiculata
May 1998 YAN M6
(HKU(M) 3358)
L. livistonae Brunei Darussalam, Temburong,
Batu Apoi Forest Reserve, The
University of Brunei Darussalam
Kuala Belalong Field Studies
Centre (KBFSC), Baki Tributary
Decaying rachis of
Oncosperma horridum
Feb. 1999 YAN 92 On
(HKU(M) 13181)
Neolinocarpon australiense Brunei Darussalam, Temburong, Batu
Apoi Forest Reserve, The University
of Brunei Darussalam Kuala Belalong
Field Studies Centre (KBFSC),
Baki Tributary
Decaying rachis of
Oncosperma horridum
Feb. 1999 YAN 93 On
(HKU(M) 13210)
N. globosicarpon Brunei Darussalam, Temburong,
mangrove
Decaying rachis of
Nypa fruticans
April 1998 YAN M3
(HKU(M) 3359)
N. nonappendiculatus Brunei Darussalam, Temburong, Batu
Apoi Forest Reserve, The University
of Brunei Darussalam Kuala Belalong
Field Studies Centre (KBFSC),
Baki Tributary
Decaying rachis of
Oncosperma horridum
Dec. 1997 YAN 108 On
(HKU(M) 10153)
Figs 10–16. Light and SEM micrographs of ascospores ofNeolinocarpon australiense. Figs 10–11. Note the roundedapex and basal truncate appendage (arrowed in 11). Fig. 12.
Rounded apex. Figs 13–16. The truncate base of each asco-spore is surrounded by a membranous collar with enclosedgranular material. Bars: 10=25 mm; 11=10 mm; 12–16=1 mm.
Yanna, W. H. Ho and K. D. Hyde 1307
with uranyl acetate for 40 min and examined using aJEOL 100S transmission electron microscope operat-ing at 80 kV.
RESULTS
Linocarpon
Linocarpon aquaticum
The ascospores comprise round appendages of uniformwidth at both ends (Figs 1, 3). Usually one of the asco-spore appendages was attached to the polycarbonatemembrane, while the other was unattached (Fig. 3).Unattached ends were thick, pad-shaped, with smoothirregular edges, which may have folded. Attached ap-pendages had slightly spread out forming thin layers ofmucilage which adhered to the polycarbonate mem-brane (Fig. 4).
Linocarpon livistonae
The apex of each ascospore was rounded and lackedappendages, while the base was covered with muci-laginous material (Figs 2, 5–7). The ascospore wallcomprised an electron-dense episporium, 10.5–14 nmthick and a bilamellate mesosporium, 40–60 nm thick(Figs 8–9). The outer mesosporium (11–15 nm thick)was more electron-dense than the inner one (34.5–40 nm thick) (Figs 8–9). The basal appendages com-prised electron-dense fibrillar material that was derivedfrom the episporium (Figs 8–9).
Neolinocarpon
Neolinocarpon australiense
The apex of each ascospore was rounded and lackedappendages (Figs 10–12), while the base was sur-rounded by a membranous collar with enclosed
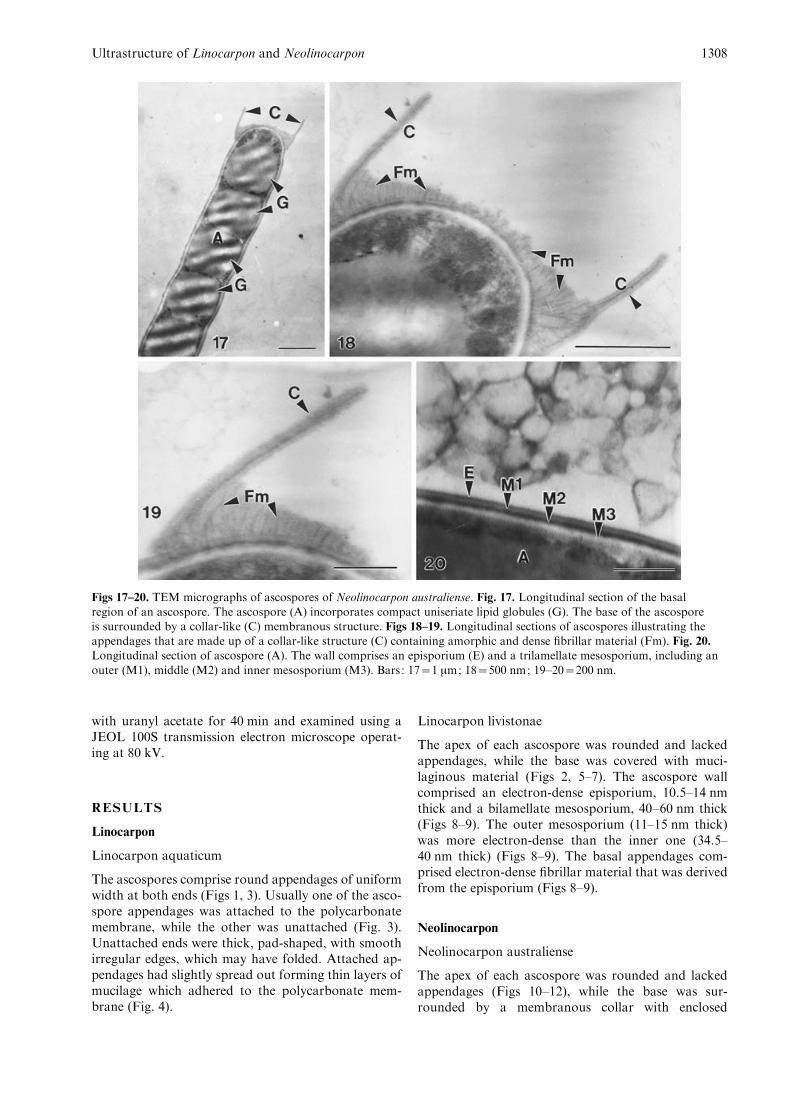
Figs 17–20. TEM micrographs of ascospores of Neolinocarpon australiense. Fig. 17. Longitudinal section of the basal
region of an ascospore. The ascospore (A) incorporates compact uniseriate lipid globules (G). The base of the ascosporeis surrounded by a collar-like (C) membranous structure. Figs 18–19. Longitudinal sections of ascospores illustrating theappendages that are made up of a collar-like structure (C) containing amorphic and dense fibrillar material (Fm). Fig. 20.Longitudinal section of ascospore (A). The wall comprises an episporium (E) and a trilamellate mesosporium, including an
outer (M1), middle (M2) and inner mesosporium (M3). Bars: 17=1 mm; 18=500 nm; 19–20=200 nm.
Ultrastructure of Linocarpon and Neolinocarpon 1308
granular material (Figs 13–16). The edge of the mem-brane collar was irregular and usually spread outwards(Figs 14–16). When attached, the membranous ma-terial of the collar may break open, releasing thegranular material that sticks to the polycarbonatemembrane.
The ascospore wall comprised a 3.6–4 nm thickelectron-dense episporium and a 48–52 nm thick, tri-lamellate mesosporium (outer : 15–16.5 nm thick,middle: 9–10 nm thick, inner: 22–28 nm thick) (Fig.20). The middle layer of the mesosporium was the mostelectron-dense and the inner layer of the mesosporiumwas least electron-dense (Fig. 20). The basal end com-prised a trilamellate collar which was not continuousfrom the ascospore wall (Figs 17–19). The collar was50–58 nm thick, comprising a thin electron-dense layer(7.5–11 nm thick) which was sandwiched between twoless electron-dense layers (21–25 nm thick). The asco-spore wall surrounded by the collar was covered withamorphic material and electron-dense fibrillar materialwhich extended from the middle layer of the meso-sporium (Figs 17–19).
Neolinocarpon globosicarpon
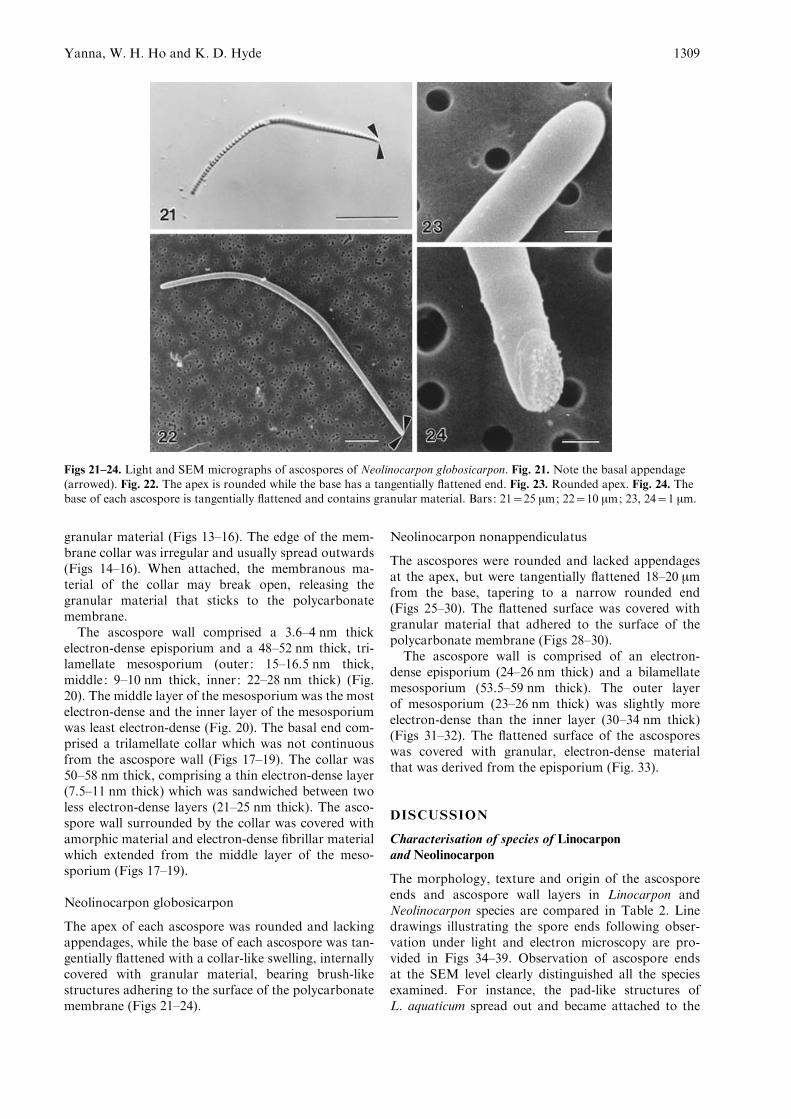
The apex of each ascospore was rounded and lackingappendages, while the base of each ascospore was tan-gentially flattened with a collar-like swelling, internallycovered with granular material, bearing brush-likestructures adhering to the surface of the polycarbonatemembrane (Figs 21–24).
Neolinocarpon nonappendiculatus
The ascospores were rounded and lacked appendagesat the apex, but were tangentially flattened 18–20 mmfrom the base, tapering to a narrow rounded end(Figs 25–30). The flattened surface was covered withgranular material that adhered to the surface of thepolycarbonate membrane (Figs 28–30).
The ascospore wall is comprised of an electron-dense episporium (24–26 nm thick) and a bilamellatemesosporium (53.5–59 nm thick). The outer layerof mesosporium (23–26 nm thick) was slightly moreelectron-dense than the inner layer (30–34 nm thick)(Figs 31–32). The flattened surface of the ascosporeswas covered with granular, electron-dense materialthat was derived from the episporium (Fig. 33).
DISCUSSION
Characterisation of species of Linocarponand Neolinocarpon
The morphology, texture and origin of the ascosporeends and ascospore wall layers in Linocarpon andNeolinocarpon species are compared in Table 2. Linedrawings illustrating the spore ends following obser-vation under light and electron microscopy are pro-vided in Figs 34–39. Observation of ascospore endsat the SEM level clearly distinguished all the speciesexamined. For instance, the pad-like structures ofL. aquaticum spread out and became attached to the
Figs 21–24. Light and SEM micrographs of ascospores of Neolinocarpon globosicarpon. Fig. 21. Note the basal appendage(arrowed). Fig. 22. The apex is rounded while the base has a tangentially flattened end. Fig. 23. Rounded apex. Fig. 24. Thebase of each ascospore is tangentially flattened and contains granular material. Bars: 21=25 mm; 22=10 mm; 23, 24=1 mm.
Yanna, W. H. Ho and K. D. Hyde 1309
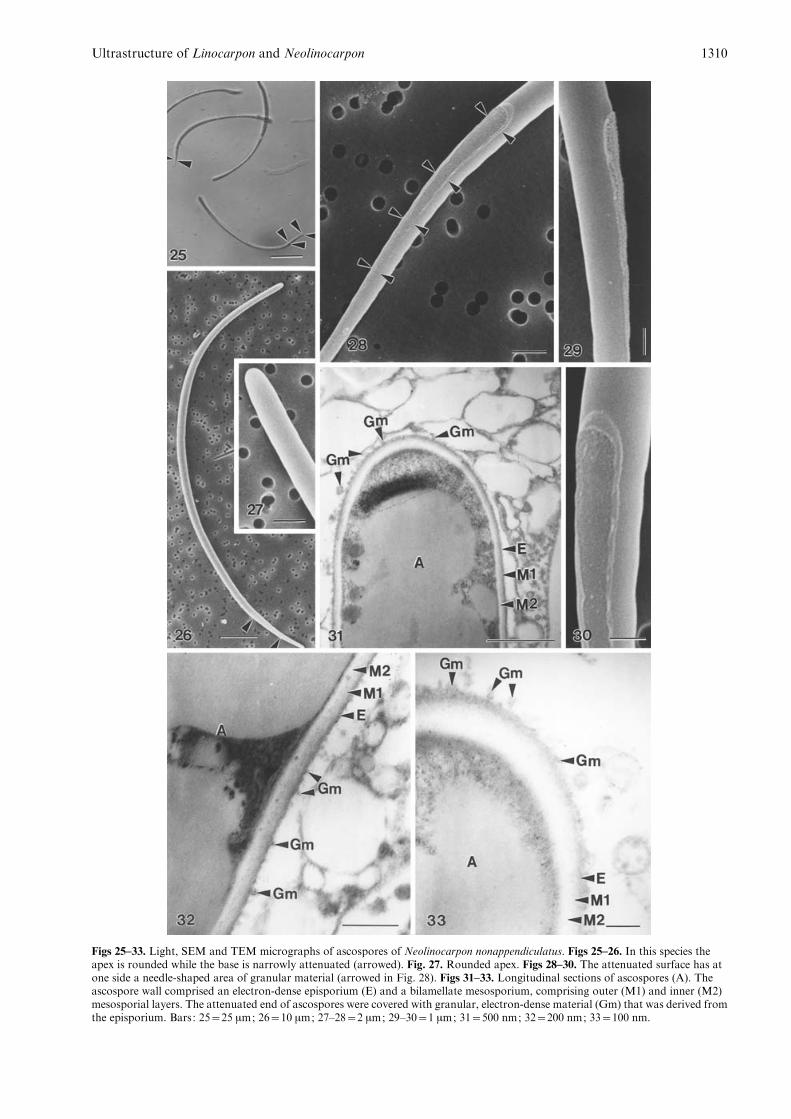
Figs 25–33. Light, SEM and TEM micrographs of ascospores of Neolinocarpon nonappendiculatus. Figs 25–26. In this species theapex is rounded while the base is narrowly attenuated (arrowed). Fig. 27. Rounded apex. Figs 28–30. The attenuated surface has atone side a needle-shaped area of granular material (arrowed in Fig. 28). Figs 31–33. Longitudinal sections of ascospores (A). Theascospore wall comprised an electron-dense episporium (E) and a bilamellate mesosporium, comprising outer (M1) and inner (M2)mesosporial layers. The attenuated end of ascospores were covered with granular, electron-dense material (Gm) that was derived fromthe episporium. Bars: 25=25 mm; 26=10 mm; 27–28=2 mm; 29–30=1 mm; 31=500 nm; 32=200 nm; 33=100 nm.
Ultrastructure of Linocarpon and Neolinocarpon 1310
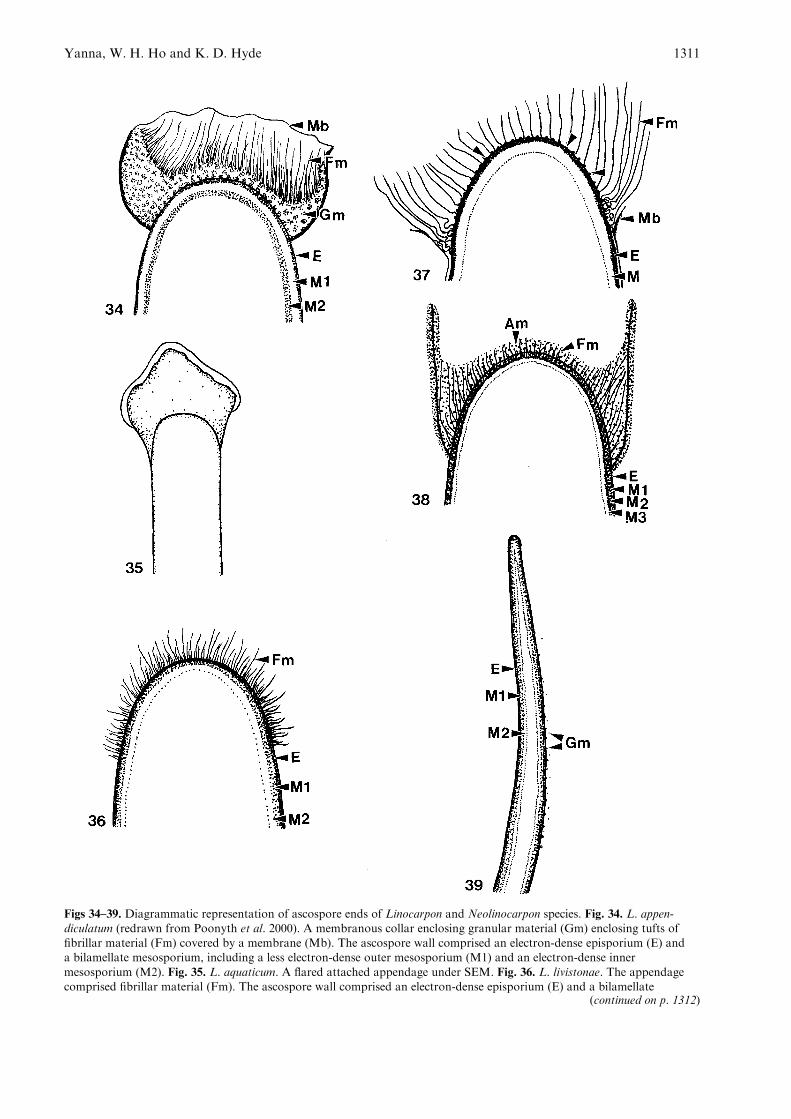
Figs 34–39. Diagrammatic representation of ascospore ends of Linocarpon and Neolinocarpon species. Fig. 34. L. appen-diculatum (redrawn from Poonyth et al. 2000). A membranous collar enclosing granular material (Gm) enclosing tufts offibrillar material (Fm) covered by a membrane (Mb). The ascospore wall comprised an electron-dense episporium (E) anda bilamellate mesosporium, including a less electron-dense outer mesosporium (M1) and an electron-dense inner
mesosporium (M2). Fig. 35. L. aquaticum. A flared attached appendage under SEM. Fig. 36. L. livistonae. The appendagecomprised fibrillar material (Fm). The ascospore wall comprised an electron-dense episporium (E) and a bilamellate
(continued on p. 1312)
Yanna, W. H. Ho and K. D. Hyde 1311

polycarbonate membrane (Fig. 4). L. livistonae asco-spore ends were similar to L. aquaticum in producingmucilaginous structures, but only at the bases (Fig. 5).N. australiense had a distinct collar-like end containinga fibrous structure (Figs 13–16). Neolinocarpon non-appendiculatus produced a layer of mucilage liningthe tapering region towards the basal tip (Fig. 28).Although L. appendiculatum, L. nipae and N. globosi-carpon produced brush-like structures at the SEM level,the structural details of each were distinct. One sporeend of L. appendiculatum comprised a collar which wasfilled with short fibrils (Poonyth et al. 2000). The sporeends of L. nipae were made up of long fibrils (Poonythet al. 2000), whereas those of N. globosicarpon com-prised granular material (Fig. 24).
Generic delineation of Linocarpon and Neolinocarpon
Ascospore appendage ontogeny at the ultrastructurallevel has been widely used in the delineation of fungalgenera. For instance, Moss (1990) and Jones (1995)have demonstrated delineation in genera of Halo-sphaeriaceae based on ascospore appendage ontogeny.Annulatascus, Cataractispora, Diluvicola, Fluminicola,Pseudoproboscispora and Rivulicola described in theAnnulatascaceae were also delineated based on theirdifference in ascospore appendage ontogeny (Ho, Hyde& Hodgkiss 1999, Wong, Hyde & Jones 1999, Ho &Hyde 2000).
Ascospore end structures of all examined Linocarponand Neolinocarpon species originated as a direct out-growth of the episporium (L. livistonae, L. nipae, and
N. nonappendiculatus), mesosporium (N. australiense),and episporium and mesosporium (L. appendiculatum).In a review of generic delineation in Halosphaeriaceae,Jones (1995) recognised ten ascospore appendageontogeny types, including four which do not have anexosporium: (1) appendage formed as a direct out-growth from one or more of the spore wall layers ; (2)appendage exuded through a pore or pores in the as-cospore wall ; (3) ascospores with end chambers formedas outgrowths of the mesosporium and episporium, thechamber containing mucilage ; and (4) ascospores withpolar mucilaginous appendages. In Linocarpon andNeolinocarpon species, the ascospore ends are of similarontogeny, as a direct outgrowth of one or more of thespore wall layers. It is therefore not possible to delin-eate species of Linocarpon and Neolinocarpon at thegeneric level based on ascospore appendage ontogeny.The distinction between species based on ascospore endstructure is also confused.
The position of ascomata therefore remains the maincriterion to delineate Linocarpon and Neolinocarponspecies. However, using the position of ascomata todelineate the two genera is not straightforward. Theascomata of Linocarpon should be superficial and thoseof Neolinocarpon should be deeply immersed (Hyde1992a, Hyde et al. 1998). Unfortunately, L. aquaticumand L. palmetto produce immersed ascomata (Hyde1992b). It is therefore arguable, that these last twospecies should be placed in Neolinocarpon. With theconfusion in delineation of Linocarpon and Neolino-carpon species now demonstrated, the genera need to becompared using molecular techniques.
Table 2. Comparison of ultrastructural morphologies of some Linocarpon and Neolinocarpon species.
Texture Morphology Origin of appendages Ascospore wall layer
Linocarpon appendiculatum
(Poonyth et al. 2000)
Collar with fibrils
(brush-like)
Fibrillar Episporium and
mesosporium
1 episporium
2 mesosporium
L. aquaticum Pad-like and
mucilaginous
– – –
L. livistonae Mucilaginous Fibrillar Episporium 1 episporium
2 mesosporium
L. nipae (Poonyth et al. 2000) Brush-like Fibrillar Episporium 1 episporium
1 mesosporium
Neolinocarpon australiense Collar Fibrillar Mesosporium 2 1 episporium
3 mesosporium
N. globosicarpon Brush-like – – –
N. nonappendiculatus Mucilaginous Granular Episporium 1 episporium
2 mesosporium
mesosporium, including an electron-dense outer mesosporium (M1) and an less electron-dense inner mesosporium (M2).Fig. 37. L. nipae. The appendage comprised fibrillar material (Fm). Also note the remnants of the membrane (Mb) whichpartially covers the fibrillar material and the ascospore wall adjacent to the appendage. The ascospore wall comprised
an electron-dense episporium (E) and a less electron-dense mesosporium (M). Fig. 38. N. australiense. A membranous collarcomprising amorphous material (Am) and fibrillar material (Fm) under TEM. The appendage comprised fibrillar material(Fm). The ascospore wall comprised an electron-dense episporium (E) and a trilamellate mesosporium, including an outer
mesosporium (M1), middle (M2) and inner mesosporium (M3). Fig. 39. N. nonappendiculatus. The ascospore wall comprisedan electron-dense episporium (E) and a bilamellate mesosporium (M1, M2). Note the granular material (Gm) which isderived from the episporium. This is a needle-shaped area of granular material as seen under SEM.
(continued )
Ultrastructure of Linocarpon and Neolinocarpon 1312

REFERENCES
Dulymamode, R., Cannon, P. F. & Peerally, A. (1998) Fungi from
Mauritius: Linocarpon species on Pandanus. Mycological Research
102 : 1331–1337.
Frohlich, J. & Hyde, K. D. (2000) Palm Microfungi. [Fungal
Diversity Research Series Vol. 3.] Fungal Diversity Press,
Hong Kong.
Ho, W. H. & Hyde, K. D. (2000) A new family of freshwater asco-
mycetes. Fungal Diversity 4 : 21–36.
Ho, W. H., Hyde, K. D. & Hodgkiss, I. J. (1999) Ultrastructure
of Annulatascus aquaticus sp. nov., a freshwater ascomycete
on submerged wood from Hong Kong. Fungal Diversity 2 :
119–128.
Hyde, K. D. (1988) The genus Linocarpon from the mangrove palm
Nypa fruticans. Transactions of the Mycological Society of Japan
29 : 339–350.
Hyde, K. D. (1992a) Fungi from decaying intertidal fronds of Nypa
fruticans, including three new genera and four species. Botanical
Journal of the Linnean Society 110 : 95–110.
Hyde, K. D. (1992b) Fungi from palms. I. The genus Linocarpon, a
revision. Sydowia 44 : 32–54.
Hyde, K. D. (1997) Additions to the genus Linocarpon (Ascomycetes :
Hyponectriaceae). Botanical Journal of the Linnean Society 123 :
109–131.
Hyde, K. D. & Alias, S. A. (1999) Linocarpon angustatum sp. nov.,
and Neolinocarpon nypicola sp. nov. from petioles of Nypa fruti-
cans, and a list of fungi from aerial parts of this host. Mycoscience
40 : 145–149.
Hyde, K. D., Taylor, J. E. & Frohlich, J. (1998) Fungi from palms.
XXXIV. The genus Neolinocarpon with five new species and one
new combinations. Fungal Diversity 1 : 115–131.
Hyde, K. D., Taylor, J. E. & Frohlich, J. (2000) Genera of Asco-
mycetes from Palms. [Fungal Diversity research Series Vol. 2.]
Fungal Diversity Press, Hong Kong.
Jones, E. B. G. (1995) Ultrastructure and taxonomy of the aquatic
ascomycetous order Halosphaeriales. Canadian Journal of Botany
73 (Suppl.) : S790–S801.
Kirk, P. W. (1966) Morphogenesis and microscopic cytochemistry
of the marine pyrenomycete ascospores.Nova Hedwigia Beiheft 22 :
1–128.
Moss, S. T. (1990) The relevance of scanning electron microscopy
in the taxonomy of marine ascomycetes. In Scanning Electron
Microscopy in Taxonomy and FunctionalMorphology (D. Claugher,
ed.) : 149–170. Oxford University Press, Oxford.
Mollenhauer, H. H. (1964) Plastic embedding mixtures for use in
electron microscopy. Stain Technology 39 : 111–114.
Poonyth, A. D., Hyde, K. D., Wong, S. W. & Peerally, A. (2000)
Ultrastructure of asci and ascospore appendages in Linocarpon
appendiculatum and L. nipae. Botanica Marina 43 : 213–221.
Sydow, H. & Sydow, P. (1917) Beitrag zur Kenntnis der Pilzflora der
Philippinen-Inseln. Annales Mycologici 15 : 165–268.
Thongkantha, S., Lumyong, S., Lumyong, P., Whitton, S. R.,
McKenzie, E. H. C. & Hyde, K. D. (2003) Microfungi on the
Pandanaceae : Linocarpon lammiae sp nov., L. siamensis sp nov.
and L. suthepensis sp nov. are described with a key to Linocarpon
species from the Pandanaceae. Mycologia 95 : 360–367.
Wang, Y. Z. & Hyde, K. D. (1999) Hyponectria buxi with notes on
the Hyponectriaceae. Fungal Diversity 3 : 159–172.
Wong, S. W., Hyde, K. D. & Jones, E. B. G. (1999) Ultrastructural
studies on freshwater ascomycetes, Fluminicola bipolaris gen. et sp.
nov. Fungal Diversity 2 : 189–197.
Corresponding Editor: I. B. Heath
Yanna, W. H. Ho and K. D. Hyde 1313


















