A comparison of the effects of calponin on smooth and skeletal ...

1
Calponin 3 regulates actin cytoskeleton rearrangement in trophoblastic cell fusion 1 2 Yukinao Shibukawa1, Natsuko Yamazaki1, Keiichi Kumasawa3, Etsuko Daimon1, Michiko 3 Tajiri1, 2, Yuka Okada3, Masahito Ikawa3, and Yoshinao Wada1† 4 5
1Department of Molecular Medicine, Osaka Medical Center and Research Institute for 6 Maternal and Child Health, 840 Murodo-cho, Izumi, Osaka 594-1101, Japan 7
2CREST, Japan Science and Technology Agency, 4-1-8 Honcho Kawaguchi, Saitama 8 332-0012, Japan 9
3Research Institute for Microbial Diseases and Graduate School of Pharmaceutical Sciences, 10 Osaka University, 3-1 Yamada-oka, Suita, Osaka 565-0871, Japan 11 12 Running title: Calponin3 regulates cell-cell fusion (less than 40 charactors) 13 14 † To whom all correspondence should be addressed. 15 Department of Molecular Medicine 16 Osaka Medical Center and Research Institute for Maternal and Child Health 17 840 Murodo-cho, Izumi, Osaka 594-1101, Japan 18 Tel : 81-725-56-1220 19 Fax : 81-725-57-3021 20 E-mail: [email protected] 21
22
http://www.molbiolcell.org/content/suppl/2010/09/21/E10-03-0261.DC1Supplemental Material can be found at:

2
1The abbreviations used are: PMSF, phenylmethylsulfonyl fluoride; DTT, dithiothreitol; 1 DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum; HS, horse serum; PBS, 2 phosphate-buffered saline; CNN3, calponin 3; CHD, calponin homology domain; ABS, actin 3 binding site; PM, plasma membrane; CAPMP, cytoplasmically attached peripheral membrane 4 protein; DsRed, red fluorescent protein; ECFP, enhanced cyan fluorescent protein; GFP, green 5 fluorescent protein; EYFP, enhanced yellow fluorescent protein; shRNA, short hairpin RNA; 6 CK, casein kinase 7
8

3
ABSTRACT 1 Cell-cell fusion is an intriguing differentiation process, essential for placental development and 2 maturation. A proteomic approach identified a cytoplasmic protein, calponin 3 (CNN3), 3 related to the fusion of BeWo choriocarcinoma cells. CNN3 was expressed in cytotrophoblasts 4 in human placenta. CNN3 gene knockdown promoted actin cytoskeletal rearrangement and 5 syncytium formation in BeWo cells, suggesting CNN3 to be a negative regulator of trophoblast 6 fusion. Indeed, CNN3 depletion promoted BeWo cell fusion. CNN3 at the cytoplasmic face 7 of cytoskeleton was dislocated from F-actin with forskolin treatment and diffused into the 8 cytoplasm in a phosphorylation dependent manner. Phosphorylation sites were located at 9 Ser293/296 in the C-terminal region and deletion of this region or site-specific disruption of 10 Ser293/296 suppressed syncytium formation. These CNN3 mutants were co-localized with 11 F-actin and remained there after forskolin treatment, suggesting that dissociation of CNN3 from 12 F-actin is modulated by the phosphorylation status of the C-terminal region unique to CNN3 in 13 the CNN family proteins. The mutant missing these phosphorylation sites displayed a 14 dominant negative effect on cell fusion, while replacement of Ser293/296 with aspartic acid 15 enhanced syncytium formation. These results indicated that CNN3 regulates actin 16 cytoskeleton rearrangement which is required for the plasma membranes of trophoblasts to 17 become fusion competent. 18 19
20

4
Introduction 1 Cellular fusion is a dramatic biological event observed in a wide variety of organisms. The 2
fusion process has been studied independently in different species and cells: yeast, C. elegans 3 epidermal cells, myoblasts, macrophages and trophoblasts, as well as during both physiological 4 and pathological events such as fertilization, tumorigenesis and tissue regeneration (Chen and 5 Olson, 2005). Furthermore, virus- or chemical-induced cell-cell fusion is currently an 6 indispensable tool for studying gene expression, chromosomal mapping, antibody production 7 and cancer immunotherapy. Although the mechanisms underlying cellular fusion are not fully 8 understood, some fusogens and transcription factors participating in cell type-specific processes 9 have been identified; e.g., a fusogenic membrane protein called syncytin and transcription factor 10 GCMa (glial cell missing) are known to be required for placental development (Mi et al., 2000; 11 Schreiber et al., 2000; Yu et al., 2002). In addition to Eff-1 and FUS-1 in C. elegans epithelial 12 cell fusion, Duf, Rst and other immunoglobulin (Ig) domain-containing transmembrane proteins 13 are essential for muscle cell fusion and development (Ruiz-Gomez et al., 2000; Strunkelnberg et 14 al., 2001; Mohler et al., 2002; Kontani et al., 2005; Sohn et al., 2009). While most of these 15 molecules are specific to certain systems, a few such as the guanine-nucleotide exchange factors, 16 e.g. Dock180 and Brag2, are involved in myoblast as well as macrophage fusion (Pajcini et al., 17 2008). However, the dynamics of cell-cell adhesion, alignment and membrane mixing are 18 apparently quite similar among the fusion processes of these cell types, and it is therefore 19 conceivable that cytoskeletal rearrangement is finely regulated during the course of cell fusion. 20
Calponin was originally identified as a molecule binding to F-actin, calmodulin and 21 tropomyosin, and to be involved in regulating the contraction/relaxation cycle in smooth muscle 22 cells (Takahashi et al., 1988; Takahashi and Nadal-Ginard, 1991). Three CNN family proteins 23 are characterized by the N-terminal calponin homology domain (CHD) and a middle region 24 containing actin binding site 1 (ABS1) and ABS2, and are distinguished from each other by 25 their unique C-terminal tails (Takahashi and Nadal-Ginard, 1991; Strasser et al., 1993; 26 Applegate et al., 1994; Morgan and Gangopadhyay, 2001; Rozenblum and Gimona, 2007): 27 basic, neutral and acidic CNNs, or CNN1, CNN2 and CNN3, respectively. 28
In this study, we carried out a proteomic search for fusion-related molecules in BeWo 29 choriocarcinoma cells and found calponin3 (CNN3) to be required for cellular fusion. This 30 novel function of CNN3 was associated with actin cytoskeletal rearrangement regulated by 31 phosphorylation of the CNN3-specific C-terminal region. 32
33

5
Results 1 Identification of calponin 3 as a fusion-related protein 2
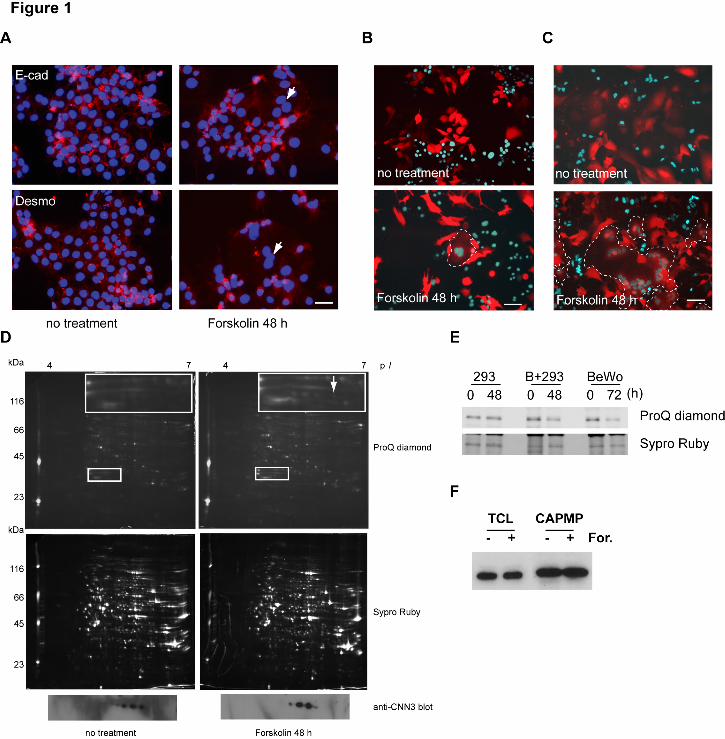
BeWo cells differentiate to form syncytiotrophoblasts via a cyclic AMP-dependent pathway 3 following forskolin treatment (Wice et al., 1990; Keryer et al., 1998). Syncytium formation is 4 usually observed by immuno-staining for E-cadherin or desmoplakin localized at intercellular 5 boundaries (Keryer et al., 1998), but discriminating between multi- and mononuclear cells is 6 often quite difficult (Fig. 1A). To overcome the problem, we developed a differential coloring 7 system by employing co-culture, e.g. red fluorescent protein in the cytoplasm (DsRed cells) and 8 enhanced-cyan fluorescent protein with a nuclear localization signal (ECFP-Nuc cells), to 9 visualize fusion by detecting DsRed/ECFP-Nuc-double positive cells (Fig. 1B). 10 DsRed/ECFP-Nuc-double positive syncytia were detected after 2-day forskolin treatment, but 11 fusion efficiency was less than 5% for homotypic BeWo/BeWo cell fusion (Fig. 1B). In 12 contrast, heterotypic fusion of BeWo with HEK293 cells, which formed a huge hybrid 13 syncytium, was found to be more efficient (Fig. 1C), allowing robust proteomic analysis. 14
In order to identify fusion-related molecules in the plasma membrane (PM), "cytoplasmically 15 attached peripheral membrane proteins" (CAPMPs) were isolated from the PM fraction using 16 cationic colloidal silica (Chaney and Jacobson, 1983; Ghitescu et al., 1997). Two-dimensional 17 electrophoresis (2-DE) followed by ProQ Diamond staining and peptide mass fingerprinting 18 identified several cytoskeletal, transmembrane protein-associated molecules showing altered 19 expressions, isoelectric points and/or phosphorylation levels in response to forskolin treatment 20 (Supplemental Table I). Among them, we focused on CNN3, or acidic CNN (Supplemental 21 Table II), because this molecule was believed to be involved in cytoskeletal rearrangement and 22 thereby to be related to membrane mixing and fusion. CNN3 was identified in at least five 23 spots by 2-DE followed by Western blotting (Fig. 1D, bottom panels). After forskolin 24 treatment, the intensity of acidic spots was decreased, and ProQ Diamond staining suggested 25 this change to be due to dephosphorylation (Fig. 1D). To confirm the change in CNN3 26 phosphorylation levels with fusion induction, endogenous CNN3 molecules were 27 immunoprecipitated and phospho-CNN3 was detected by ProQ-Diamond staining. The overall 28 phosphorylation level of CNN3 was decreased in both homotypic BeWo/BeWo and heterotypic 29 BeWo/HEK293 fusion, while CNN3 protein levels were unchanged (Figs. 1E and 1F). In the 30 absence of BeWo cells, HEK293 cells did not fuse with each other (data not shown) and there 31 were no other associated changes in CNN3 phosphorylation even with forskolin treatment (Fig. 32 1E). These results suggest CNN3 to have multiple phosphorylation sites and that CNN3 33 undergoes dephosphorylation during BeWo cell fusion. RT-PCR analysis showed no 34 expression of other members of the CNN family, i.e. CNN1 and CNN2, in BeWo or HEK293 35 cells (Supplemental Fig. S1A). 36

6
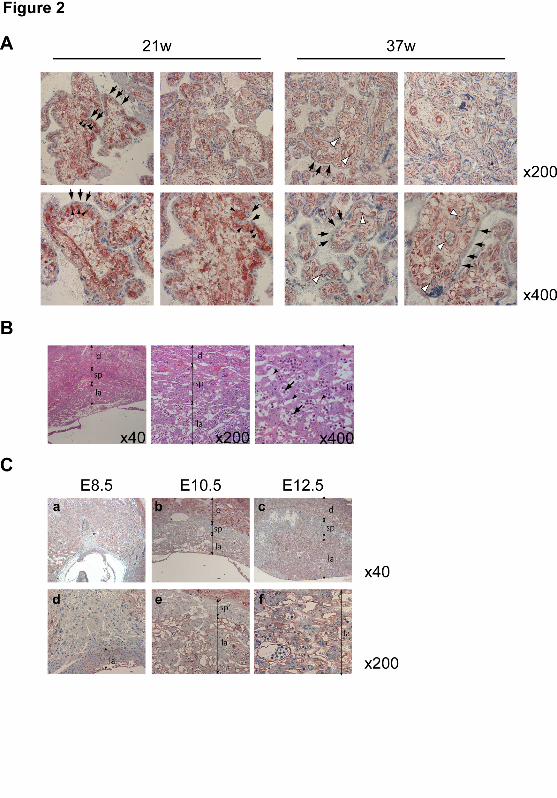
1 In vivo CNN3 expression in the placental tissues of humans and mice 2 BeWo is a human choriocarcinoma cell line. In the human placenta, cytotrophoblasts fuse 3 together to form syncytiotrophoblasts or fuse with syncytiotrophoblasts, replenishing the 4 syncytium that covers the surfaces of chorionic villi. To investigate CNN3 expression in vivo, 5 chorionic villi in human placenta were analyzed by immunohistchemistry. As shown in 6 Figure 2A, CNN3 was expressed in cytotrophoblasts beneath the syncytiotrophoblast layer but 7 not in this layer itself. In term placenta, the population of cytotrophoblasts is quite small, and 8 CNN3 expression was observed only in endothelial cells in chorionic villi. This histological 9 distribution indicated that CNN3 expression disappears during the course of tropoblastic cell 10 differentiation. 11
In mice, the labyrinth is a tissue in which trophoblasts differentiate and fuse to form a 12 syncytiotrophoblast layer (for review see (Rossant and Cross, 2001)). As shown in Figures 2B 13 and 2C, CNN3 was detected in both labyrinth and maternal deciduas at 10.5 dpc but not in the 14 spongiotrophoblast layer at any time during placental development. CNN3 was expressed by 15 fetal endothelial cells as well (Fig. 2C, panel f). These observations suggested CNN3 to play a 16 role in placental development in both human and mouse. 17
18 CNN3 depletion by RNAi enhances BeWo cell fusion 19
To study the role of CNN3 in trophoblastic cell fusion, lentivirus vector-based 20 RNA-interference (RNAi) constructs, which allowed simultaneous expressions of a short 21 hairpin RNA (shRNA) and a marker protein, either enhanced yellow-green fluorescent protein 22 (EYFP) or ECFP-Nuc, were introduced into BeWo cells. The resulting depletion with either 23 CNN3 RNAi sequence was at least 90%, as demonstrated by Western blotting (Supplemental 24 Fig. S1B). Phalloidin staining revealed that the CNN3 gene knockdown induced 25 morphological change in BeWo cells. In contrast to the cubic and island-like morphology of 26 control cells, the CNN3-depleted cells had a broad, flat cytoplasm (Fig. 3A). This observation 27 of morphological change was supported by Phalloidin staining for F-actin. As shown in Figure 28 3A, actin bundles disappeared from CNN3-depleted cells even in the absence of forskolin. 29 Furthermore, CNN3 depletion promoted syncytium formation even in the absence of forskolin 30 and was further augmented by forskolin treatment (Figs. 3A and 3B), as were the morphological 31 features described above. These results indicated that CNN3 prevents BeWo cells from 32 undergoing cell-cell fusion. 33
Syncytin is a fusogenic protein playing a central role in trophoblast fusion in both humans 34 and mice, and the expression is regulated by GCMa, a cAMP-responsible transcription factor 35 (Mi et al., 2000; Frendo et al., 2003; Dupressoir et al., 2005; Dupressoir et al., 2009). In this 36

7
study, syncytin-1 expression levels were decreased by forskolin treatment for BeWo cell fusion 1 as reported by Vargas et al. (Vargas et al., 2009), and this syncytin-1 down-regulation was 2 suppressed by CNN3 knockdown (Supplemental Fig. S1C). These findings suggested that 3 CNN3 might regulate syncytin-1 expression or affect stabilization of this membrane protein. 4 Only trophoblastic cell lines are able to induce GCMa and form syncytium in response to cAMP 5 signaling (Yu et al., 2002), and thus neither morphological change nor syncytium formation was 6 induced by CNN3 depletion in HEK293 cells (data not shown). 7 8 CNN3 phosphorylation sites involved in cell fusion 9
The actin-binding property of CNN1 is regulated by kinase- and phosphatase-dependent 10 phosphorylation/dephosphorylation of serine, threonine or tyrosine residues. Two 11 phosphorylation sites at Ser175 and Thr184 of CNN1 are conserved in this family (Fig. 4A), 12 and phosphorylation of these residues dissociates CNN1 from F-actin in vitro (Nakamura et al., 13 1993; Tang et al., 1996; Kaneko et al., 2000; Abouzaglou et al., 2004). 14
To determine CNN3 phosphorylation sites, Flag-CNN3 was recovered from HEK293 cells 15 and subjected to tryptic digestion. Mass spectrometry (MS) of the phosphopeptides, enriched 16 by TiO2 (Sugiyama et al., 2007), identified at least three, two major and one minor, 17 phosphorylation sites in the peptide corresponding to residues 272-318 (Supplemental Figs. 18 S2A-S2E) and two phosphorylation sites in a smaller peptide corresponding to residues 272-298 19 (Supplemental Figs. S2F and S2G). These peptides are located in the acidic tail which is 20 unique to CNN3 in the CNN family, suggesting this specific region to be involved in the 21 regulation of fusion. Indeed, the mutant (ΔC) lacking the C-terminal sequence from residue 22 274 was negative for ProQ Diamond phospho-specific staining (Figs. 4B and 4D), indicating the 23 phosphorylation sites to be located exclusively within the C-terminal region. Sypro Ruby 24 staining revealed similar amounts of Flag-tagged WT or ΔC CNN3 to be recovered by 25 immunoprecipitation using anti-Flag-agarose beads. In Figure 4B, a 40kDa protein 26 co-precipitated with Flag-tagged WT or ΔC CNN3 was identified as actin by peptide mass 27 fingerprinting (PMF) analysis. The ΔC mutant retained its actin-binding property, because the 28 ABS1 and ABS2 remained intact (Danninger and Gimona, 2000). Indeed, actin amounts in the 29 immunoprecipitates were similar in cells transfected with wild type or ΔC CNN3 before 30 forskolin treatment. 31
Subsequently, to identify the phosphorylation sites of CNN3 (Supplemental Figs. S2G-S2H 32 for sequence), various Flag-CNN3 mutants were transiently expressed in HEK293 cells, and 33 Flag-CNN3 was recovered by immunoprecipitation using anti-Flag agarose beads. The 34 phosphorylation levels of each CNN3 mutant were evaluated by ProQ-Diamond staining. 35 Among the putative phosphorylation sites within the 272-298 region, the substitution of Ser293 36

8
or Ser296 with Ala decreased phosphorylation levels of CNN3 and these mutants also retained 1 actin binding properties (Figs. 4D and 4E). S293/296A double mutation drastically reduced 2 CNN3 phosphorylation (Fig. 4D and Supplemental Fig. S2J). S293/296 phosphorylation was 3 also identified in BeWo cells (data not shown). None of the point mutations at other putative 4 S/T/Y phosphorylation sites altered phosphorylation status (Fig. 4E), indicating Ser293 and 5 Ser296 to be the major phosphorylation sites in the C-terminal region of CNN3. 6
To address whether CNN3 phosphorylation is involved in cell fusion and syncytium 7 formation, ΔC, S293/296A, or S293/296D was introduced into BeWo cells expressing DsRed or 8 CFP-Nuc, and the ratio of the numbers of DsRed/CFP-Nuc double positive cells to total DsRed 9 positive cells was calculated. As shown in Fig. 4F, ΔC or S293/296A mutation significantly 10 suppressed forskolin-induced cell fusion via a dominant negative effect, while the cells 11 expressing S293/296D showed retention of fusion efficiency. The number of nuclei in the 12 syncytium, or fusion index, decreased by 30-40% in cells expressing ΔC or S293/296A and 13 increased by 20% in those expressing S293/296D (Fig. 4G). The effects of S293A and S296A 14 mutations were additive (data not shown), and ΔC exerted the most prominent suppression on 15 cell fusion. 16
Several lines of evidence indicate that CNN1 is phosphorylated by multiple kinases including 17 Rho-associated kinase (ROCK), Erk, CaMK and PKC (Winder and Walsh, 1990; Kaneko et al., 18 2000). To identify CNN3-specific kinases, we performed a search using NetPosK 19 (http://www.cbs.dtu.dk/services/), and found casein kinase (CK) 1/2 to be a candidate kinase 20 for Ser296 phosphorylation. We then focused on ROCK as well as CK, and carried out an in 21 vitro phosphorylation assay using a rCNN3 substrate. ProQ Diamond phospho-specific 22 staining revealed that the active forms of both ROCK2 and CK1 were capable of 23 phosphorylating CNN3. Ser293 was preferentially phosphorylated by ROCK rather than 24 CK1, while the overall phosphorylation activity of CK1 was higher than that of ROCK2 25 (Supplemental Fig. S1G). However, pretreatment with specific inhibitors against these 26 kinases did not change the constitutive CNN3 phosphorylation in unstimulated BeWo cells 27 (data not shown). 28
29 Phosphorylation status and subcellular distribution of endogenous CNN3 in cell fusion 30
We prepared antibodies separately recognizing phosphoserine at either Ser293 or Ser296 (See 31 supplemental Fig. S3 for properties of these antibodies). The anti-pSer293 antibody was much 32 less reactive to Ser296A mutant than Ser296D mutant or wild-type CNN3 (Supplemental Fig. 33 3E), suggesting that Ser296 phosphorylation preceded Ser293 phosphorylation. Next, we 34 analyzed site-specific phosphorylation of endogenous CNN3 from different subcellular fractions. 35 With forskolin treatment, cytosolic CNN3 was dephosphorylated at these residues (Fig. 5A), 36

9
and phospho-S293 spots recovered from CAPMP fraction were sifted to the basic side 1 (Supplemental Fig. 1H). These results were consistent with the reduction of total CNN3 2 phosphorylation levels shown in Figure 1E. 3
In Figure 5B, the amount of actin bound to endogenous CNN3 was markedly decreased in 4 both homotypic (BeWo/BeWo) and heterotypic (BeWo/HEK293) cell fusion. This result was 5 consistent with that of Flag-tagged CNN3 shown in Figure 4. Indeed, endogenous CNN3 was 6 well co-localized with F-actin before forskolin treatment and diffused into the cytoplasm in the 7 syncytium after stimulation, as demonstrated by microscopic observations (Fig. 5C). 8 9 Binding of CNN3 to actin cytoskeleton is regulated by CNN3 C-terminal region 10
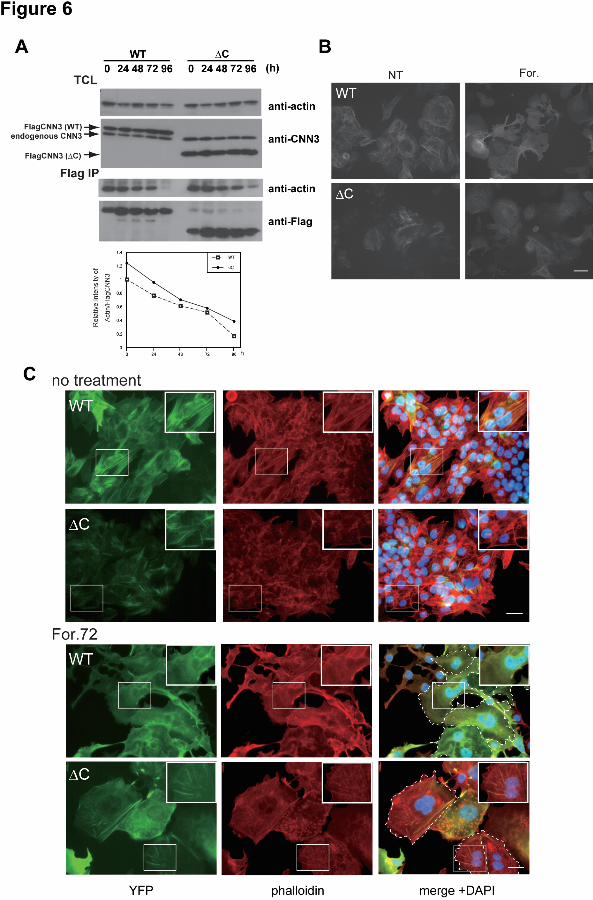
Implications of S293/296 phosphorylation in cytoskeletal rearrangement and cell fusion were 11 then investigated using BeWo cells expressing ΔC mutant. Compared with the wild-type 12 CNN3 transfectant, a significant amount of actin remained bound to ΔC CNN3 at 96h after 13 forskolin treatment, though dissociated from actin in a time dependent manner (Fig. 6A). The 14 levels of actin and ΔC CNN3 in the total cell lysate were unchanged (Fig. 6A, upper panels). 15 This result was consistent with the previous reports showing the tail region of CNNs to 16 negatively regulate actin binding capacity (Danninger and Gimona, 2000; Burgstaller et al., 17 2002). Microscopic observations of living cells revealed the YFP-tagged ΔC mutant as well 18 as the wild-type CNN3 to be localized at stress fiber-like bundles and the cytoplasmic face of 19 the PM before forskolin treatment (Fig. 6B left panels). After stimulation, wild-type CNN3 20 diffused into the cytoplasm, but the ΔC mutant remained at the cytoskeletal bundles (Fig. 6B 21 right panels). Phalloidin staining demonstrated co-localization of wild or ΔC CNN3 with 22 F-actin before forskolin treatment (Fig. 6C). The wild-type CNN3 was not co-localized with 23 F-actin in multinucleated cells after stimulation, indicating dissociation from the actin 24 cytoskeleton. The S293/296D CNN3 underwent a similar dislocation (Supplemental Fig. S4). 25 In contrast, ΔC CNN3 remained at the actin cytoskeleton even in the multinucleated cells, which 26 were found only a few among the ΔC CNN3-expressing cells and retained F-actin bundles (Fig. 27 6C). The Ser293/296A CNN3 behaved in a similar fashion to the ΔC mutant, and the F-actin 28 cytoskeleton remained after forskolin treatment (Supplemental Fig. S4). These results 29 indicated that the phosphorylated Ser293/296 are required for dissociation of CNN3 from actin 30 cytoskeleton and thus for efficient trophoblastic cell fusion. 31
32

10
Discussion 1 Three CNN family proteins have an N-terminal CHD followed by a region containing actin 2
binding sites, and the C-terminal region specifies each isoform. CNN1 is involved in 3 regulating the contraction/relaxation cycle in smooth muscle cells. Binding of CNN1 to the 4 actin cytoskeleton inhibits actomyosin ATPase activity (el-Mezgueldi et al., 1996), and is 5 regulated by phosphorylation at the conserved residues in the ABS region by protein kinase C or 6 Ca2+/calmodulin dependent protein kinase II (Winder and Walsh, 1990; Kaneko et al., 2000). 7 Gene targeting of CNN1 results in ectopic bone formation, suggesting a negative role of CNN1 8 in osteogenesis (Yoshikawa et al., 1998). CNN2 is involved in macrophage differentiation, 9 and other CNN isoforms do not compensate for loss of this CNN2 function (Huang et al., 2008). 10 These reports indicate the CNN family to be involved in differentiation as well as contractile 11 mechanics and cellular motility. In the present study, CNN3 was shown to participate in the 12 cytoskeletal reorganization necessary for trophoblastic fusion, playing a role similar to that of 13 CNN2 in cytoskeletal rearrangement and migration of endothelial cells during vascular 14 development (Fukui et al., 1997; Tang et al., 2006). Depletion of CNN3 by RNAi enhanced 15 cell fusion of BeWo cells, suggesting that CNN3 impedes the cytoskeletal rearrangement 16 necessary for cell fusion by binding F-actin. 17
CNN binding to the actin cytoskeleton is conferred by ABS 1 and ABS2 and the C-terminal 18 region negatively regulates this basal property of CNN as demonstrated for CNN1 and CNN2 19 (Danninger and Gimona, 2000; Burgstaller et al., 2002). We examined the C-terminal region 20 of CNN3 and found the phosphorylation status at S293/296 to regulate CNN3 binding to the 21 actin cytoskeleton and thereby cytoskeletal remodeling, which renders the PM flexible and cells 22 competent to fuse. In fact, cells expressing ΔC and S293/296A mutants which lack this 23 regulatory system displayed decreased fusion efficiency. The phenotype of cells expressing 24 S293/296A CNN3 was mild compared with those expressing ΔC CNN3, probably due to minor 25 phosphorylation sites remaining in the C-terminal region. On the other hand, cells expressing 26 the S293/296D mutant displayed a fusion efficiency similar to that of those expressing 27 wild-type CNN3. The S293/296D CNN3 was localized at F-actin bundle and actin 28 cytoskeleton. After forskolin treatment, the cells expressing S293/296D CNN3 underwent 29 normal processes including F-actin bundle disappearance, and the actin cytoskeleton 30 rearrangement. In unstimulated BeWo cells, the role of Ser293/296 phosphorylation would be 31 to keep actin cytoskeleton flexible for de novo actin polymerization and/or treadmilling. 32 Phosphorylated CNN3 is co-localized with F-actin and undergoes release, re-cycling and re-binding, 33 while lack of C-terminal phosphorylation sites, or the regulatory tail, enhances and stabilizes the 34 CNN3 binding to actin and reduces actin dynamics and membrane flexibility. Upon stimulation 35 by forskolin, factor (or signal) “X” recognizes or binds to the phosphorylated C-terminal region, 36

11
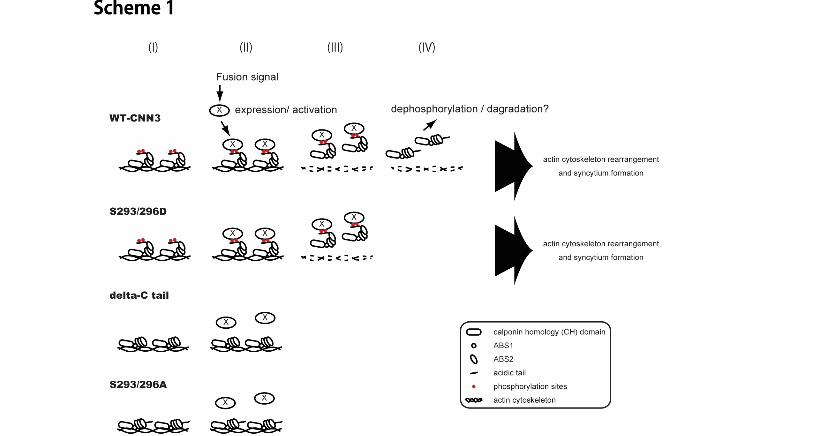
which regulates the affinity of the actin binding region to actin, and thereby CNN3 is released from 1 F-actin. The overall process involving S293/296 phosphorylation is illustrated in Scheme 1; (i) In 2 unstimulated cells, CNN3 is constitutively phosphorylated at Ser293 and Ser296, and is 3 associated with F-actin cytoskeleton via actin-binding sites. (ii, iii) After fusion induction, 4 factor (or signal) “X” is induced/activated and binds to phosphorylated CNN3, which is then 5 released from actin cytoskeleton. (iv) Dephosphorylation of S293/296 or degradation of 6 CNN3 occurs after dissociation from actin. Upregulation of phosphatases might be involved 7 in the last phase. The dephosphorylated CNN3 is not re-cycled, because F-actin bundle, or the 8 binding target, rapidly disappears in the forskolin-treated, fused or unfused, BeWo cells. In 9 summary, CNN3 phosphorylation probably plays distinct roles of regulating actin binding in a 10 steady-state condition and of receiving factor “X” after fusion induction. 11
In human placenta, CNN3 expression was markedly decreased in the course of trophoblastic 12 cell differentiation (Fig. 2A), indicating CNN3 undergoes down-regulation by decreased gene 13 expression or protein degradation after cell fusion in vivo. It has been reported that CNN2 14 synthesis and degradation were regulated by mechanical tension (Hossain et al. , 2006) and 15 that CNNs 1 and 3 were cleaved by μ-calpain (Tsunekawa et al. , 1989; Yoshimoto et al. , 16 2000). In BeWo cells, CNN3 might be cleaved and down-regulated in mature syncytium, 17 although we did not study the fate of CNN3 after cell fusion in vitro. 18
In the present study, forskolin promoted dissociation of CNN3 from actin in BeWo cells, but 19 had the opposite effect in HEK293 cells (Fig. 4A). This discrepancy is probably due to a 20 difference in motility between these cells. The cellular motility and membrane ruffling of 21 HEK293 cells in culture are inherently much faster than those in BeWo cells, and forskolin 22 accelerates the movement of HEK293 cells while decelerating that of BeWo cells (our 23 unpublished observations with time-lapse imaging). 24
ROCK, protein kinase C, CaM kinase and Src family kinases have been reported to 25 phosphorylate Ser, Thr or Tyr residues of CNN1 (Nakamura et al., 1993; Tang et al., 1996; 26 Kaneko et al., 2000; Abouzaglou et al., 2004). In addition, a line of studies demonstrated the 27 involvement of MAP kinase family or ROCK in labyrinth formation (for review see (Rossant 28 and Cross, 2001; Hatano et al., 2003; Thumkeo et al., 2003)). In our preliminary study, 29 ROCK1/2 and ERK1/2 were capable of phosphorylating S293 of CNN3 in vitro, while casein 30 kinase phosphorylated other residues (Supplemental Fig. S1H). On the other hand, it is likely 31 that specific phosphatases are also involved in this mechanism, as protein phosphatases 1 and 32 2B are responsible for CNN1 dephosphorylation (Ichikawa et al., 1993; Fraser and Walsh, 33 1995). Indeed, Vargas et al. reported acceleration of trophoblast fusion with inhibition of 34 tyrosine phosphatase (Vargas et al., 2008), and we identified altered expressions of protein 35 phosphatase(s) alpha/beta according to BeWo cell fusion (Supplemental Table I). 36

12
Membrane flexibility is necessary for membrane fusion, though it is not a sufficient condition. 1 In the present study, a novel function of CNN3 in trophoblastic cell fusion was involved in 2 cytoskeletal dynamics which impacts on plasma membrane flexibility and mixing, consequently 3 on cell fusion. This novel function of CNN3 was identified in myoblasts as well as 4 trophoblasts and works in myotube formation (our unpublished observations), suggesting that 5 regulation of actin cytoskeletal rearrangement by CNN3 is required for these cells to become 6 fusion competent. This regulatory mechanism probably underlies a wide range of cellular 7 events and manipulations including virus-cell fusion, hybridoma production and stem 8 cell-mediated tissue regeneration. 9
10

13
METHODS 1 Materials and Methods 2
The following reagents and antibodies were used: mouse monoclonal anti-E-cadherin IgG 3 from BD Bioscience (Franklin Lakes, NJ); mouse monoclonal anti-desmoplakin 1/2 IgG from 4 Progen (Heidelberg, Germany); mouse polyclonal anti-human CNN3 IgG from Abnova (Taipei, 5 Taiwan), mouse monoclonal anti-FLAG (M2) IgG, and mouse monoclonal anti-actin IgG from 6 Sigma (St. Louis, MO); rabbit polyclonal anti-CNN3 IgG and anti-syncytin-1 from Santa Cruz 7 (Santa Cruz, CA); rabbit polyclonal anti-FLAG IgG from Cell Signaling (Danvers, MA); 8 ProQ-Diamond Phosphoprotein Gel Stain and SYPRO Ruby from Molecular Probes (Eugene, 9 OR); lysylendopeptidase (Achromobacter lyticus protease I) from Wako (Osaka, Japan); trypsin 10 (Sequence Grade Modified Trypsin, from porcine pancreas) from Promega (Madison, WI). 11 12 Phospho-specific CNN3 antibodies 13 Anti-CNN3 pS293 and pS296 rabbit antibodies were raised against phosphorylated peptides: 14 N’-CQGTGTNG(phos)SEI; and N’-EISD(phos)SDYQAEC (MBL, Nagoya, Japan). 15 Antibodies were affinity-purified from serum by using the corresponding phosphorylated 16 peptide-coupled agarose beads. The phospho-specific antibodies were then affinity-purified by 17 immunoadsorption with non-phosphorylated peptides. The specificities of the resulting 18 antibodies were verified by ELISA. 19 20 Cloning and site-directed mutagenesis of human CNN3 21
Human CNN3 cDNA was amplified from the random-primed in-house cDNA library of 22 BeWo cells (American Type Culture Collection, Manassas, VA), and inserted into a XhoI/EcoRI 23 site of pENTR/flag to generate N-terminal Flag-tagged CNN3, or a XhoI/BamHI site of 24 EYFP-C1 (Clontech, Mountain View, CA) to generate EYFP-CNN3. C-terminal deletion (ΔC) 25 or site-directed mutagenesis was performed using a KOD-Plus Mutagenesis kit (TOYOBO, 26 Osaka, Japan) according to the manufacturer’s protocol. For the ΔC mutant, a stop codon 27 followed by an EcoRI site was introduced by PCR. 28 29 Cell culture, treatment, transfection and transduction of lentivirus vectors 30
BeWo cells constitutively expressing fluorescent protein (CFP-Nuc or DsRed) were 31 maintained in an undifferentiated state in F12 Ham medium (Wako) supplemented with 10% 32 fetal bovine serum (FBS). Differentiation was induced by treatment with 50 μM forskolin 33 (Wako), for up to 96 h (Wice et al., 1990; Lyden et al., 1993; Keryer et al., 1998) with daily 34 exchange of the forskolin-containing medium. Recombinant lentiviruses kindly provided by 35 Dr. Hiroyuki Miyoshi (RIKEN BRC, Ibaragi, Japan), were produced by transient transfection of 36

14
HEK293T cells according to reported protocols (Zufferey et al., 1997; Miyoshi, 2004). Briefly, 1 sub-confluent HEK293T cells were co-transfected with 20 µg of a self-inactivating (SIN) vector, 2 10 µg of a pCAG-HIVgp and 10 µg of a pCMV-VSV-G-RSV-Rev by calcium phosphate 3 precipitation. The medium was changed 18h later, and the recombinant lentivirus vectors were 4 harvested after an additional 48h incubation. 5 6 Detection of CNN3 in placenta 7
Human placentas at 21 and 37 weeks gestation were obtained, under informed consent, after 8 elective abortion and Caesarian section, respectively, and transferred to our laboratory after 9 anonymization according to the Ethical Guidelines for Clinical Research in Osaka Medical 10 Center and Research Institute for Maternal and Child Health. The placentas were cut into 11 small pieces, which were then fixed with formaldehyde and paraffin-embedded. Mouse 12 placentas were isolated at 8.5 to 12.5 dpc and subjected to the same fixation and embedding 13 procedures as the human placentas. For immunohisotochemistry, rabbit polyclonal IgG against 14 human CNN3 was used for these human and mouse tissues, and peroxidase-based color 15 development was performed using ABC and AEC (3-amino-9-ethylcarbazole) kits (Vector, 16 Burlingame, CA). 17 18 Detection of cell fusion in BeWo cells 19
After fixation with 4% paraformaldehyde, cells were immunostained for E-cadherin and 20 desmoplakin with monoclonal antibodies, and cell nuclei were counterstained with DAPI. 21 Photographs were taken with an Olympus microscope IX51 (Tokyo, Japan) equipped with a 22 cooled charge-coupled device camera, and the images were prepared for presentation using 23 Q-Capture Pro. For BeWo or BeWo/HEK293 cell fusion, the cells constitutively expressing 24 DsRed and CFP-Nuc were harvested by trypsinization, mixed at a ratio of 1:1 for BeWo or 2:1 25 for BeWo/HEK293 and then seeded in microplate wells (22 mm diameter) at a density of 2x104 26 cells/mL. After cell adhesion, the cells were fed daily with forskolin-containing media. The 27 fusion frequency was counted as fusion induction, represented by the number of nuclei in 28 DsRed and CFP-Nuc double positive syncytia against the total number of nuclei in the DsRed 29 positive cells, as determined by microscopic observation (x100 magnification). The degree of 30 maturation was calculated as the fusion index, which was represented by the average number of 31 nuclei per DsRed and CFP-Nuc double positive cells, as determined by microscopic observation 32 (x100 magnification). 33 34 shRNA-induced degradation of CNN3 35 The shRNA-encoding lentiviral vectors were created by inserting the annealed complementary 36

15
oligonucleotides into the BamHI and HindIII site located at the 3’ end of a human U6 1 promoter-containing the pENTER4 vector. The shRNAs in pENTR4 were then subcloned into 2 a CSII-RfA-CMV-GFP or a CSII-RfA-CMV-CFP-Nuc vector by an L-R clonase reaction 3 (Invitrogen, Carlsbad, CA) according to the manufacturer’s protocol. Purified lentiviruses 4 allow simultaneous expression of both shRNA and marker genes. The sequences for the 5 coding strand of shRNA were (5’- 3’) 6 GATCCGTCGACTGTGGATTGGCATCTGTGAAGCCACAGATGGGATGCCAATCCACAG7 TCGACTTTTTTA and (5’- 3’) 8 AGCTTAAAAAAGTCGACTGTGGATTGGCATCCCATCTGTGGCTTCACAGATGCCAAT9 CCACAGTCGACG for controls, (5’- 3’) 10 GATCCGCACATGCTCGGAAGGAAACTGTGAAGCCACAGATGGGTTTCCTTCCGAGC11 ATGTGCTTTTTTA and (5’- 3’) 12 AGCTTAAAAAAGCACATGCTCGGAAGGAAACCCATCTGTGGCTTCACAGTTTCCTTC13 CGAGCATGTGCG for hCNN3 shRNA1, (5’- 3’) 14 GATCCGGCTCAACACCTTGCTCATCTGTGAAGCCACAGATGGGATGAGCAAGGTGTT15 GAGCCTTTTTTA and (5’- 3’) 16 AGCTTAAAAAAGGCTCAACACCTTGCTCATCCCATCTGTGGCTTCACAGATGAGCAA17 GGTGTTGAGCCG for hCNN3 shRNA2. 18 19 Immunoblot analysis 20
Cells were washed twice with ice-cold PBS, scraped and lysed at 4ºC in a buffer containing 21 20 mM Tris-HCl pH 7.2, 150 mM NaCl, 0.1% NP-40, 0.5% Triton-X 100, 5 mM EDTA, 1 mM 22 Na3VO4, 1 mM NaF, 1 mM phenylmethylsulfonyl fluoride (PMSF), 2 μg/ml aprotinin and 1 23 mM dithiothreitol (DTT) and then centrifuged at 15,000 x g for 15 min. The supernatants 24 were collected and the protein concentrations were determined by the Bradford method (BioRad, 25 Hercules, CA). Equal amounts of proteins were loaded on a 10% SDS-PAGE gel, and then 26 transferred to PVDF membranes (Schleicher & Schuell, Dassel, Germany). The membrane 27 was incubated with primary and secondary antibodies for 1h each and detection was performed 28 using an ECL kit (GE Healthcare, Piscataway, NJ) according to the manufacturer's instructions. 29
30 Purification of CAPMPs from the apical-PM protein fraction 31
PMs from BeWo cells were isolated using a cationic colloidal silica method (Chaney and 32 Jacobson, 1983; Ghitescu et al., 1997; Stolz et al., 1999; Ghitescu et al., 2001; Rahbar and 33 Fenselau, 2004). The cells were washed twice with ice-cold PBS (-), then with PM coating 34 (PMC) buffer (0.5 mM CaCl2, 1 mM MgCl2, 20 mM MES, 135 mM NaCl, pH 5.3). The cells 35 were next incubated in a 5% suspension of cationic colloidal silica (Sigma-Aldrich, St. Louis, 36

16
MO) in PMC buffer on ice for 1 min. The solution phase was removed, and the cells were 1 washed with PMC buffer to remove excess silica. The cells were then incubated in a 10 mg/ml 2 of polyacrylic acid (Sigma) solution with PMC buffer at pH 6.1 on ice for 1 min. The 3 polyacrylic acid solution was removed, and the cells were washed with PMC buffer. The cells 4 were washed briefly in lysis buffer (2.5 mM imidazole, pH 7.0), and then incubated with lysis 5 buffer containing a protease inhibitor (1 mM PMSF and 50 μg/ml aprotinin) on ice for 30 min. 6 The cells were scraped and the cell suspension was recovered and homogenized by 20 strokes 7 with a Dounce homogenizer. The lysate was mixed with an equal volume of 100% Nycodenz 8 and then placed over a 1.0 ml layer of 70 % Nycodenz. The tube was topped off with lysis 9 buffer and then centrifuged in a swing bucket at 20,000 x g for 30 min. Following removal of 10 the layer containing nuclei, the pellet containing silica-coated PMs was washed 3 times with 11 lysis buffer. CAPMPs were extracted from the silica-coated PMs by incubation in 100 mM 12 Na2CO3, at pH 11.4 on ice for 30 min followed by centrifugation at 12,000 x g for 10 min 13 (Hubbard and Ma, 1983; Ghitescu et al., 1997). Back extraction was performed once with the 14 same procedure except for omission of the 10 min centrifugation. 15
16 Two-dimensional electrophoresis (2-DE) 17
The CAPMPs were precipitated with trichloroacetic acid (TCA) and then washed with 18 ethanol. The pellet was solubilized in 2-DE buffer (7M urea, 2M thiourea and 4% CHAPS) 19 and a 40 μg sample was subjected to 2-DE using an IPG strip (13 cm, pH4-7L) (GE Healthcare) 20 for the first dimension separation. Fluorescent detection was carried out for phosphoproteins 21 using ProQ-Diamond Phosphoprotein Gel Stain according to the manufacturer’s protocol, and 22 the images were acquired with a FluoroPhoreStar 3000 (Anatech, Tokyo, Japan). Subsequently, 23 the gels were washed and stained with SYPRO Ruby to visualize total proteins. 24
25

17
Acknowledgments 1 The authors thank Dr. Katsuhito Takahashi for helpful insights on properties of the CNN family. 2 The authors also thank Dr. Masahiro Nakayama for helpful comments on the histology of the 3 human placenta. This work was supported in part by Grants-in-Aid for Young Scientists (B) 4 (17790273) from Japan Society for the Promotion of Science and from Takeda Science 5 Foundation. 6
7

18
Figure legends 1 Figure 1. Identification of CNN3 as a fusion-related protein. (A) BeWo cell fusion 2 visualized by the conventional staining with anti-E-cadherin or anti-desmoplakin in 3 combination with nuclear staining with DAPI. Cells were treated with or without 50 μM 4 forskolin for 48 h. Arrows indicate a syncytium in which these marker proteins were 5 translocated and diffused into the cytoplasm. Scale bar: 50 μm. (B) Detection of 6 forskolin-induced syncytium between DsRed- and CFP-Nuc-expressing BeWo cells. A 7 syncytium is easily recognized by the cyan-colored multiple nuclei within the homogeneously 8 red-colored cytoplasm, whereas unfused cells have either red cytoplasm or cyan nuclei. Scale 9 bar: 100 μm. (C) Heterologous fusion of CFP-Nuc-expressing HEK293 cells with BeWo cells 10 expressing DsRed. The incidence of BeWo/HEK293 fusion was higher than that of 11 homologous BeWo cell fusion and the hybrid syncytium was much larger than that of 12 BeWo-derived homologous syncytium. Scale bar: 250 µm. The broken lines in B and C 13 show the periphery of the syncytium. (D) Cytoplasmically attached peripheral membrane 14 proteins (CAPMPs) isolated from BeWo/HEK293 co-culture were separated by 2-DE. The gels 15 were stained with ProQ Diamond phospho-specific staining (top panels) or SYPRO Ruby 16 protein staining (middle panels). A spot (arrow) among a series of protein spots in inset was 17 subjected to in-gel digestion followed by peptide mass fingerprinting, and CNN3 was identified. 18 All these spots in line were confirmed to be CNN3 by Western blotting (bottom panels) (E) 19 Endogenous CNN3 was immunopurified using anti-Flag agarose beads from total cell lysate 20 (TCL) of the cells incubated with 50 μM forskolin for the indicated times. The 21 phosphorylation status of CNN3 was analyzed by ProQ Diamond phospho-specific staining, and 22 the same gel was stained by Sypro Ruby to verify each sample equal in the amount of CNN3. 23 "B+293" indicates a BeWo/HEK293 mixed culture. (F) Western blot of CNN3 in the TCL or 24 the CAPMP fraction from BeWo cells in the presence or absence of 50 μM forskolin for 72 h. 25 26 Figure 2. CNN3 expression in human and mouse placental tissues. (A) 27 Immunohistochemistry of chorionic villi of human placentas at 21 and 37 weeks of gestation. 28 CNN3 was detected in the cytotrophoblasts (black arrowheads) and fetal endothelial cells (white 29 arrowheads) but not in the syncytiotrophoblast layer (arrows). (B) Histology of mouse 30 placenta. Transverse sections (4 μm) were stained with hematoxylin/eosin. A few maternal 31 blood sinuses (arrowheads) and fetal blood vessels (arrows) are found within the labyrinth 32 region. Localizations of the labyrinth layer (la), spongiotrophoblast layer (sp), and maternal 33 decidua (d) are indicated. (C) Immunohistochemistry of mouse placental tissues at 8.5 dpc (a, 34 d) 10.5 dpc (b, e) and 12.5 dpc (c, f). CNN3 was detected in the labyrinth layer and maternal 35 deciduas. 36

19
1 Figure 3. CNN3 gene knockdown (A) Control or CNN3-specific shRNA was introduced into 2 BeWo cells expressing EYFP or CFP-Nuc. After washing, different marker expressing cells 3 were trypsinized and mixed in the culture in the absence or presence of 50 μM forskolin for 72 h. 4 After fixation, cells were stained with Alexa Fluor 568-conjugated phalloidin (red). Scale bar: 5 40μm. Arrows indicate the multinucleated cells identified by an EYFP/CFP-Nuc double-color 6 system. (B) Higher magnification images of A. The cell morphology was visualized by Alexa 7 Fluor 568-conjugated phalloidin staining. Broken lines show the periphery of the 8 EYFP/CFP-Nuc double positive syncytium. Note that CNN3-depletion induced syncytium 9 formation without forskolin treatment and increased the number of multinucleated cells after 10 forskolin treatment. Scale bar: 100 μm. 11 12 Figure 4. CNN3 phosphorylation in BeWo cell fusion. (A) Comparison of the amino acid 13 sequences of human CNN family proteins. CNN1 phosphorylation sites are indicated by 14 asterisks. The sequence of ΔC mutant is truncated after Cys274 (arrow). (B) HEK293 cells 15 transiently expressing WT or ΔC CNN3. Flag-CNN3 with a wild orΔC sequence was 16 recovered by immunoprecipitation from cell lysate using anti-Flag-agarose beads. The eluate 17 was subjected to SDS-PAGE, and the phosphorylated CNN3s were visualized by ProQ 18 Diamond phospho-specific staining (upper panel) followed by Sypro Ruby protein staining 19 (lower panel). Note that the ΔC mutant was not phosphorylated but retained the actin-binding 20 property. (C) Flag-tagged WT or ΔC CNN3 identified by Western blotting using anti-FLAG 21 (left panel) or anti-CNN3 (right panel) antibodies. Each twenty μg sample of HEK293 cell 22 lysate was loaded. (D, E) Phosphorylation levels of WT or mutant Flag-CNN3 transiently 23 expressed in HEK293 cells. Flag-CNN3 was immunopurified using anti-Flag-agarose beads, 24 and was subjected to SDS-PAGE followed by ProQ Diamond phospho-specific staining. The 25 ratio of phosphorylated and total CNN3 was calculated and indicated below each lane, showing 26 decreased phosphorylation of S293A and S296A CNN3s. (F, G) BeWo cell fusion at 96 h after 27 forskolin treatment in various mutants. The fusion frequency was counted as “fusion induction” 28 (F), and the degree of maturation was calculated as “fusion index” (G). Data from three 29 independent experiments were averaged, and standard deviations were indicated by error bars. 30 31 Figure 5. Phosphorylation status and subcellular distribution of endogenous CNN3. (A) 32 site-specific phosphorylation of CNN3. Each twenty μg sample of the cytosolic or plasma 33 membrane (PM) fraction from BeWo cells was subjected to SDS-PAGE followed by Western 34 blotting using antibodies against specific phosphorylation site at S293 or S296 (see 35 Supplemental Fig. S3 for their specificity). Enolase (cytosol) and E-cadherin (PM) were used 36

20
as the control for cell fractionation. (B) Co-immunoprecipitation of actin with CNN3. 1 Endogenous CNN3 was immunopurified from BeWo and/or HEK293 cells after forskolin 2 treatment. Immuno-purified CNN3 or total cell lysates (TCL) were analyzed by Western 3 blotting for actin or CNN3. (C) Dissociation of endogenous CNN3 from actin cytoskeleton in 4 BeWo cell fusion. CNN3 was immunostained using rabbit anti-CNN3 IgG, and the cells were 5 visualized by Alexa Fluor 488-conjugated secondary antibody (green). Subsequently, Alexa 6 Fluor 568-conjugated phalloidin (red) and DAPI staining was performed (blue). Endogenous 7 CNN3 in multinucleated BeWo cells was dissociated from actin cytoskeleton after forskolin 8 treatment. Broken line shows the periphery of syncytium. Scale bar: 50 μm. 9 10 Figure 6. Phosphorylation-dependent association of CNN3 with actin cytoskeleton. (A) 11 Association of ΔC CNN3 with actin. Wild type (WT) or ΔC Flag-CNN3 expressing BeWo 12 cells were treated with forskolin for the indicated times, and association of CNN3 and actin was 13 analyzed by co-IP assay using anti-Flag agarose beads. Note that a significant amount of ΔC 14 CNN3 remained in association with actin at 96h after forskolin treatment, compared with WT 15 CNN3. (B) Subcellular localization of EYFP-CNN3. WT or ΔC EYFP-CNN3 was 16 introduced into BeWo cells, and the localization of EYFP-CNN3 was visualized in the presence 17 or absence of forskolin in living cells. (C) Co-localization of CNN3 and F-actin. BeWo cells 18 expressing EYFP-CNN3 were cultivated in the presence (lower panels) or absence (upper 19 panels) of 50 μM forskolin for 72 h. After fixation, the cells were stained by Alexa fluor 20 568-conjugated phalloidin (red) and DAPI (blue). Note that WT EYFP-CNN3 expressing 21 syncytium did not co-localize with F-actin after forskolin treatment, but ΔC EYFP-CNN3 still 22 overlapped with the F-actin even in multinucleated cells. The dotted lines show the periphery 23 of the syncytium. Scale bar: 50 μm. 24 25 Scheme 1. Model of CNN3-dependent trophoblast fusion 26 (I) In un-stimulated cells, CNN3 is constitutively phosphorylated at Ser293 and Ser296 in the 27 regulatory tail, and is associated with F-actin cytoskeleton via actin-binding sites. (II) When 28 cells receive a fusion signal, factor (or signal) “X” is induced/activated and binds to (or 29 recognizes) the phosphorylated regulatory tail, and triggered CNN3 dissociation from actin. 30 The phosphomimetic S293/296D mutant responds to the signal, while ΔC and S293/296A are 31 unresponsive and remain attached to F-actin. (III) After CNN3 release, actin bundles are 32 rearranged and then disappeared. (IV) Released CNN3 is dephosphorylated or degradaded in 33 cytoplasm. 34
35

21
References 1 2 Abouzaglou, J., Benistant, C., Gimona, M., Roustan, C., Kassab, R., and Fattoum, A. (2004). 3 Tyrosine phosphorylation of calponins. Inhibition of the interaction with F-actin. Eur J Biochem 4 271, 2615-2623. 5 Applegate, D., Feng, W., Green, R.S., and Taubman, M.B. (1994). Cloning and expression of a 6 novel acidic calponin isoform from rat aortic vascular smooth muscle. J Biol Chem 269, 7 10683-10690. 8 Burgstaller, G., Kranewitter, W.J., and Gimona, M. (2002). The molecular basis for the 9 autoregulation of calponin by isoform-specific C-terminal tail sequences. Journal of cell science 10 115, 2021-2029. 11 Chaney, L.K., and Jacobson, B.S. (1983). Coating cells with colloidal silica for high yield 12 isolation of plasma membrane sheets and identification of transmembrane proteins. J Biol Chem 13 258, 10062-10072. 14 Chen, E.H., and Olson, E.N. (2005). Unveiling the mechanisms of cell-cell fusion. Science. 308, 15 369-373. 16 Danninger, C., and Gimona, M. (2000). Live dynamics of GFP-calponin: isoform-specific 17 modulation of the actin cytoskeleton and autoregulation by C-terminal sequences. Journal of 18 cell science 113 Pt 21, 3725-3736. 19 Dupressoir, A., Marceau, G., Vernochet, C., Benit, L., Kanellopoulos, C., Sapin, V., and 20 Heidmann, T. (2005). Syncytin-A and syncytin-B, two fusogenic placenta-specific murine 21 envelope genes of retroviral origin conserved in Muridae. Proceedings of the National Academy 22 of Sciences of the United States of America 102, 725-730. 23 Dupressoir, A., Vernochet, C., Bawa, O., Harper, F., Pierron, G., Opolon, P., and Heidmann, T. 24 (2009). Syncytin-A knockout mice demonstrate the critical role in placentation of a fusogenic, 25 endogenous retrovirus-derived, envelope gene. Proceedings of the National Academy of 26 Sciences of the United States of America 106, 12127-12132. 27 el-Mezgueldi, M., Strasser, P., Fattoum, A., and Gimona, M. (1996). Expressing functional 28 domains of mouse calponin: involvement of the region around alanine 145 in the actomyosin 29 ATPase inhibitory activity of calponin. Biochemistry 35, 3654-3661. 30 Fraser, E.D., and Walsh, M.P. (1995). Dephosphorylation of calponin by type 2B protein 31 phosphatase. Biochemistry 34, 9151-9158. 32 Frendo, J.L., Olivier, D., Cheynet, V., Blond, J.L., Bouton, O., Vidaud, M., Rabreau, M., 33 Evain-Brion, D., and Mallet, F. (2003). Direct involvement of HERV-W Env glycoprotein in 34 human trophoblast cell fusion and differentiation. Mol Cell Biol 23, 3566-3574. 35 Fukui, Y., Masuda, H., Takagi, M., Takahashi, K., and Kiyokane, K. (1997). The presence of 36

22
h2-calponin in human keratinocyte. J Dermatol Sci 14, 29-36. 1 Ghitescu, L.D., Crine, P., and Jacobson, B.S. (1997). Antibodies specific to the plasma 2 membrane of rat lung microvascular endothelium. Exp Cell Res 232, 47-55. 3 Ghitescu, L.D., Gugliucci, A., and Dumas, F. (2001). Actin and annexins I and II are among the 4 main endothelial plasmalemma-associated proteins forming early glucose adducts in 5 experimental diabetes. Diabetes 50, 1666-1674. 6 Hatano, N., Mori, Y., Oh-hora, M., Kosugi, A., Fujikawa, T., Nakai, N., Niwa, H., Miyazaki, J., 7 Hamaoka, T., and Ogata, M. (2003). Essential role for ERK2 mitogen-activated protein kinase 8 in placental development. Genes Cells 8, 847-856. 9 Hossain, M.M., Smith, P.G., Wu, K., and Jin, J.P. (2006). Cytoskeletal tension regulates both 10 expression and degradation of h2-calponin in lung alveolar cells. Biochemistry 45, 11 15670-15683. 12 Huang, Q.Q., Hossain, M.M., Wu, K., Parai, K., Pope, R.M., and Jin, J.P. (2008). Role of 13 H2-calponin in regulating macrophage motility and phagocytosis. J Biol Chem 283, 14 25887-25899. 15 Hubbard, A.L., and Ma, A. (1983). Isolation of rat hepatocyte plasma membranes. II. 16 Identification of membrane-associated cytoskeletal proteins. J Cell Biol 96, 230-239. 17 Ichikawa, K., Ito, M., Okubo, S., Konishi, T., Nakano, T., Mino, T., Nakamura, F., Naka, M., 18 and Tanaka, T. (1993). Calponin phosphatase from smooth muscle: a possible role of type 1 19 protein phosphatase in smooth muscle relaxation. Biochem Biophys Res Commun 193, 20 827-833. 21 Kaneko, T., Amano, M., Maeda, A., Goto, H., Takahashi, K., Ito, M., and Kaibuchi, K. (2000). 22 Identification of calponin as a novel substrate of Rho-kinase. Biochem Biophys Res Commun 23 273, 110-116. 24 Keryer, G., Alsat, E., Tasken, K., and Evain-Brion, D. (1998). Cyclic AMP-dependent protein 25 kinases and human trophoblast cell differentiation in vitro. J Cell Sci. 111, 995-1004. 26 Kontani, K., Moskowitz, I.P., and Rothman, J.H. (2005). Repression of cell-cell fusion by 27 components of the C. elegans vacuolar ATPase complex. Dev Cell. 8, 787-794. 28 Lyden, T.W., Ng, A.K., and Rote, N.S. (1993). Modulation of phosphatidylserine epitope 29 expression by BeWo cells during forskolin treatment. Placenta 14, 177-186. 30 Mi, S., Lee, X., Li, X., Veldman, G.M., Finnerty, H., Racie, L., LaVallie, E., Tang, X.Y., 31 Edouard, P., Howes, S., Keith, J.C., Jr., and McCoy, J.M. (2000). Syncytin is a captive retroviral 32 envelope protein involved in human placental morphogenesis. Nature 403, 785-789. 33 Miyoshi, H. (2004). Gene delivery to hematopoietic stem cells using lentiviral vectors. Methods 34 Mol Biol. 246, 429-438. 35 Mohler, W.A., Shemer, G., del Campo, J.J., Valansi, C., Opoku-Serebuoh, E., Scranton, V., 36

23
Assaf, N., White, J.G., and Podbilewicz, B. (2002). The type I membrane protein EFF-1 is 1 essential for developmental cell fusion. Dev Cell. 2, 355-362. 2 Morgan, K.G., and Gangopadhyay, S.S. (2001). Invited review: cross-bridge regulation by thin 3 filament-associated proteins. J Appl Physiol 91, 953-962. 4 Nakamura, F., Mino, T., Yamamoto, J., Naka, M., and Tanaka, T. (1993). Identification of the 5 regulatory site in smooth muscle calponin that is phosphorylated by protein kinase C. J Biol 6 Chem 268, 6194-6201. 7 Pajcini, K.V., Pomerantz, J.H., Alkan, O., Doyonnas, R., and Blau, H.M. (2008). Myoblasts and 8 macrophages share molecular components that contribute to cell-cell fusion. J Cell Biol 180, 9 1005-1019. 10 Rahbar, A.M., and Fenselau, C. (2004). Integration of Jacobson's pellicle method into proteomic 11 strategies for plasma membrane proteins. J Proteome Res 3, 1267-1277. 12 Rossant, J., and Cross, J.C. (2001). Placental development: lessons from mouse mutants. Nat 13 Rev Genet 2, 538-548. 14 Rozenblum, G.T., and Gimona, M. (2007). Calponins: Adaptable modular regulators of the actin 15 cytoskeleton. Int J Biochem Cell Biol 31, 31. 16 Ruiz-Gomez, M., Coutts, N., Price, A., Taylor, M.V., and Bate, M. (2000). Drosophila 17 dumbfounded: a myoblast attractant essential for fusion. Cell. 102, 189-198. 18 Schreiber, J., Riethmacher-Sonnenberg, E., Riethmacher, D., Tuerk, E.E., Enderich, J., Bosl, 19 M.R., and Wegner, M. (2000). Placental failure in mice lacking the mammalian homolog of glial 20 cells missing, GCMa. Mol Cell Biol. 20, 2466-2474. 21 Sohn, R.L., Huang, P., Kawahara, G., Mitchell, M., Guyon, J., Kalluri, R., Kunkel, L.M., and 22 Gussoni, E. (2009). A role for nephrin, a renal protein, in vertebrate skeletal muscle cell fusion. 23 Proceedings of the National Academy of Sciences of the United States of America 106, 24 9274-9279. 25 Stolz, D.B., Ross, M.A., Salem, H.M., Mars, W.M., Michalopoulos, G.K., and Enomoto, K. 26 (1999). Cationic colloidal silica membrane perturbation as a means of examining changes at the 27 sinusoidal surface during liver regeneration. Am J Pathol 155, 1487-1498. 28 Strasser, P., Gimona, M., Moessler, H., Herzog, M., and Small, J.V. (1993). Mammalian 29 calponin. Identification and expression of genetic variants. FEBS Lett 330, 13-18. 30 Strunkelnberg, M., Bonengel, B., Moda, L.M., Hertenstein, A., de Couet, H.G., Ramos, R.G., 31 and Fischbach, K.F. (2001). rst and its paralogue kirre act redundantly during embryonic muscle 32 development in Drosophila. Development. 128, 4229-4239. 33 Sugiyama, N., Masuda, T., Shinoda, K., Nakamura, A., Tomita, M., and Ishihama, Y. (2007). 34 Phosphopeptide enrichment by aliphatic hydroxy acid-modified metal oxide chromatography 35 for nano-LC-MS/MS in proteomics applications. Mol Cell Proteomics 6, 1103-1109. 36

24
Takahashi, K., Hiwada, K., and Kokubu, T. (1988). Vascular smooth muscle calponin. A novel 1 troponin T-like protein. Hypertension 11, 620-626. 2 Takahashi, K., and Nadal-Ginard, B. (1991). Molecular cloning and sequence analysis of 3 smooth muscle calponin. J Biol Chem 266, 13284-13288. 4 Tang, D.C., Kang, H.M., Jin, J.P., Fraser, E.D., and Walsh, M.P. (1996). Structure-function 5 relations of smooth muscle calponin. The critical role of serine 175. J Biol Chem 271, 6 8605-8611. 7 Tang, J., Hu, G., Hanai, J., Yadlapalli, G., Lin, Y., Zhang, B., Galloway, J., Bahary, N., Sinha, S., 8 Thisse, B., Thisse, C., Jin, J.P., Zon, L.I., and Sukhatme, V.P. (2006). A critical role for calponin 9 2 in vascular development. J Biol Chem 281, 6664-6672. 10 Thumkeo, D., Keel, J., Ishizaki, T., Hirose, M., Nonomura, K., Oshima, H., Oshima, M., Taketo, 11 M.M., and Narumiya, S. (2003). Targeted disruption of the mouse rho-associated kinase 2 gene 12 results in intrauterine growth retardation and fetal death. Mol Cell Biol 23, 5043-5055. 13 Tsunekawa, S., Takahashi, K., Abe, M., Hiwada, K., Ozawa, K., and Murachi, T. (1989). 14 Calpain proteolysis of free and bound forms of calponin, a troponin T-like protein in smooth 15 muscle. FEBS Lett 250, 493-496. 16 Vargas, A., Moreau, J., Landry, S., LeBellego, F., Toufaily, C., Rassart, E., Lafond, J., and 17 Barbeau, B. (2009). Syncytin-2 plays an important role in the fusion of human trophoblast cells. 18 Journal of molecular biology 392, 301-318. 19 Vargas, A., Moreau, J., Le Bellego, F., Lafond, J., and Barbeau, B. (2008). Induction of 20 trophoblast cell fusion by a protein tyrosine phosphatase inhibitor. Placenta 29, 170-174. 21 Wice, B., Menton, D., Geuze, H., and Schwartz, A.L. (1990). Modulators of cyclic AMP 22 metabolism induce syncytiotrophoblast formation in vitro. Exp Cell Res 186, 306-316. 23 Winder, S.J., and Walsh, M.P. (1990). Smooth muscle calponin. Inhibition of actomyosin 24 MgATPase and regulation by phosphorylation. J Biol Chem 265, 10148-10155. 25 Yoshikawa, H., Taniguchi, S.I., Yamamura, H., Mori, S., Sugimoto, M., Miyado, K., Nakamura, 26 K., Nakao, K., Katsuki, M., Shibata, N., and Takahashi, K. (1998). Mice lacking smooth muscle 27 calponin display increased bone formation that is associated with enhancement of bone 28 morphogenetic protein responses. Genes Cells 3, 685-695. 29 Yoshimoto, R., Hori, M., Ozaki, H., and Karaki, H. (2000). Proteolysis of acidic calponin by 30 mu-calpain. Journal of biochemistry 128, 1045-1049. 31 Yu, C., Shen, K., Lin, M., Chen, P., Lin, C., Chang, G.D., and Chen, H. (2002). GCMa regulates 32 the syncytin-mediated trophoblastic fusion. J Biol Chem 277, 50062-50068. 33 Zufferey, R., Nagy, D., Mandel, R.J., Naldini, L., and Trono, D. (1997). Multiply attenuated 34 lentiviral vector achieves efficient gene delivery in vivo. Nat Biotechnol. 15, 871-875. 35