
C Primate Antibody Response Immunotoxin: Serological Computer
11
INFECTION AND IMMUNITY, Nov. 1994, p. 5055-5065 Vol. 62, No. 11 0019-9567/94/$04.00+0 Copyright C 1994, American Society for Microbiology Primate Antibody Response to Immunotoxin: Serological and Computer-Aided Analysis of Epitopes on a Truncated Form of Pseudomonas Exotoxin DONNA M. ROSCOE, SUN-HEE JUNG, ITAI BENHAR, LEE PAI, B. K. LEE, AND IRA PASTAN* Laboratory of Molecular Biology, National Cancer Institute, Bethesda, Maryland Received 2 June 1994/Returned for modification 2 August 1994/Accepted 26 August 1994 NLysPE38 is a 38-kDa derivative of Pseudomonas exotoxin (PE) in which domain Ia (amino acids 1 to 252) and part of domain lb (365 to 380) are deleted and an 11-amino-acid N-terminal peptide is added. LMB-1 is an immunotoxin in which the monoclonal antibody B3 is coupled to NLysPE38 near its N terminus. LMB-7 is a single-chain immunotoxin in which the Fv fragment of B3 is fused to PE38. To identify the antigenic regions of PE38, 12 polyclonal serum samples from monkeys immunized with the immunotoxins LMB-1 (six monkeys) and LMB-7 (six monkeys) were tested for their reactivity to a panel of 120 synthetic, overlapping peptides representing the amino acid sequence of NLysPE38. The antibody responses to peptides were similar among the 12 serum specimens, identifying several major immunodominant B-cell epitopes. Predominant reactivity was seen in six locations: amino acids 272 to 287, 341 to 359, 504 to 516, 540 to 564, and 573 to 591 and the C-terminal amino acids 591 to 613. The sera did not react with approximately 75% of the peptides. Furthermore, a computer-aided analysis was done to predict the immunologically relevant areas and revealed the same antigenic regions defined by serum reactivity to peptides. Competition enzyme-linked immunosorbent assays and neutralization assays were performed with domain 11, m, or m plus lb of PE38 and confirmed the immunodominance of domain HI. To analyze the role of specific amino acids in antibody binding, individual amino acids of PE38 with large accessible surface areas were altered by site-directed mutagenesis. These results also show that the predicted areas of immunogenicity agree with the reactivity of the anti-PE38 antibodies to peptides and to the mutants of PE. Pseudomonas aeruginosa exotoxin (PE) is a bacterial toxin that kills eukaryotic cells by inhibiting protein synthesis (9, 10). PE is a single-chain polypeptide consisting of three major structural domains (1). Domain Ia (amino acids 1 to 252) mediates binding to the cellular receptor, domain II (amino acids 253 to 364) translocates the toxin into the cytosol, and domain III (amino acids 400 to 613) inhibits protein synthesis by ADP-ribosylating elongation factor 2. Domain Tb (amino acids 365 to 399) has no identified function. Toxins can be coupled to monoclonal antibodies to generate immunotoxins which are capable of targeting and killing cells expressing the antigen recognized by the antibody (23, 24, 30). In the case of PE, the cell-binding domain (domain Ta) has been removed and the resulting molecule has been coupled to monoclonal antibody B3 to generate a cytotoxic molecule with specificity for cells bearing the B3 antigen (25). In particular, deletion of domain Ia and part of domain Tb (amino acids 365 to 380) and addition of an 11-amino-acid N-terminal linker to enhance chemical coupling to the antibody result in a mutant toxin referred to as NLysPE38 (see Fig. 1A) (6, 12). Conjuga- tion of NLysPE38 to the murine monoclonal antibody B3 forms the immunotoxin LMB-1, a potent therapeutic reagent designed to destroy cancer cells expressing the B3 antigen (20, 21, 23). Similarly, genetic fusion of the Fv portion of B3 to PE38 results in a smaller yet more cytotoxic recombinant immunotoxin referred to as LMB-7 (3, 20). Immunotoxins are being evaluated as novel therapeutic agents for the treatment of cancer and have shown some * Corresponding author. Mailing address: Laboratory of Molecular Biology, DCBDC, NCI, National Institutes of Health, Building 37, Room 4E16, 9000 Rockville Pike, Bethesda, MD 20892. Phone: (301) 496-4797. Fax: (301) 402-1344. efficacy in early clinical trials. However, a major drawback to the success of immunotoxins is their immunogenicity (22). Antibodies to the immunotoxin typically appear shortly after the first course of treatment, limiting the ability to give repeated courses and preventing the therapeutic benefits of continued treatment. Humanization of the antibody portion can decrease the host response to the antibody (11). However, the antigenicity of the toxin is still significant. Identifying the major immunodominant epitopes of the toxin would be of great value to the design of immunotoxins so that antigenic sites could be removed or altered in order to decrease the overall immune response to the immunotoxin. As part of the preclinical evaluation of LMB-1 and LMB-7, it was necessary to perform toxicity studies with monkeys, using the protocol proposed for human trials. Sera from the monkeys were therefore available for immunological analyses. In the present study, synthetic overlapping peptides corresponding to the complete NLysPE38 sequence and mutant PE molecules were used to map the immunodominant epitopes recognized by polyclonal antibodies from monkeys immunized with two PE38-containing immunotoxins, LMB-1 and LMB-7. The NLysPE38 toxin is shown to contain several immunodominant continuous sequences. MATERIALS AND METHODS Peptide synthesis. Peptides were synthesized by Cambridge Research Biochemicals (Cambridge, United Kingdom) by pre- viously described methods (7). Peptides corresponding to the sequence NLysPE38 were 10 amino acids long and overlapped by 7 amino acids. Each peptide contained an additional 4-amino-acid spacer SGSG at the N terminus to which biotin was attached. 5055 Downloaded from https://journals.asm.org/journal/iai on 23 October 2021 by 84.95.205.19.
Transcript of C Primate Antibody Response Immunotoxin: Serological Computer

INFECTION AND IMMUNITY, Nov. 1994, p. 5055-5065 Vol. 62, No. 110019-9567/94/$04.00+0Copyright C 1994, American Society for Microbiology
Primate Antibody Response to Immunotoxin: Serological andComputer-Aided Analysis of Epitopes on a Truncated
Form of Pseudomonas ExotoxinDONNA M. ROSCOE, SUN-HEE JUNG, ITAI BENHAR, LEE PAI, B. K. LEE, AND IRA PASTAN*
Laboratory of Molecular Biology, National Cancer Institute, Bethesda, Maryland
Received 2 June 1994/Returned for modification 2 August 1994/Accepted 26 August 1994
NLysPE38 is a 38-kDa derivative of Pseudomonas exotoxin (PE) in which domain Ia (amino acids 1 to 252)and part of domain lb (365 to 380) are deleted and an 11-amino-acid N-terminal peptide is added. LMB-1 isan immunotoxin in which the monoclonal antibody B3 is coupled to NLysPE38 near its N terminus. LMB-7 isa single-chain immunotoxin in which the Fv fragment of B3 is fused to PE38. To identify the antigenic regionsof PE38, 12 polyclonal serum samples from monkeys immunized with the immunotoxins LMB-1 (six monkeys)and LMB-7 (six monkeys) were tested for their reactivity to a panel of 120 synthetic, overlapping peptidesrepresenting the amino acid sequence of NLysPE38. The antibody responses to peptides were similar amongthe 12 serum specimens, identifying several major immunodominant B-cell epitopes. Predominant reactivitywas seen in six locations: amino acids 272 to 287, 341 to 359, 504 to 516, 540 to 564, and 573 to 591 and theC-terminal amino acids 591 to 613. The sera did not react with approximately 75% of the peptides.Furthermore, a computer-aided analysis was done to predict the immunologically relevant areas and revealedthe same antigenic regions defined by serum reactivity to peptides. Competition enzyme-linked immunosorbentassays and neutralization assays were performed with domain 11, m, orm plus lb of PE38 and confirmed theimmunodominance of domain HI. To analyze the role of specific amino acids in antibody binding, individualamino acids of PE38 with large accessible surface areas were altered by site-directed mutagenesis. Theseresults also show that the predicted areas of immunogenicity agree with the reactivity of the anti-PE38antibodies to peptides and to the mutants of PE.
Pseudomonas aeruginosa exotoxin (PE) is a bacterial toxinthat kills eukaryotic cells by inhibiting protein synthesis (9, 10).PE is a single-chain polypeptide consisting of three majorstructural domains (1). Domain Ia (amino acids 1 to 252)mediates binding to the cellular receptor, domain II (aminoacids 253 to 364) translocates the toxin into the cytosol, anddomain III (amino acids 400 to 613) inhibits protein synthesisby ADP-ribosylating elongation factor 2. Domain Tb (aminoacids 365 to 399) has no identified function.
Toxins can be coupled to monoclonal antibodies to generateimmunotoxins which are capable of targeting and killing cellsexpressing the antigen recognized by the antibody (23, 24, 30).In the case of PE, the cell-binding domain (domain Ta) hasbeen removed and the resulting molecule has been coupled tomonoclonal antibody B3 to generate a cytotoxic molecule withspecificity for cells bearing the B3 antigen (25). In particular,deletion of domain Ia and part of domain Tb (amino acids 365to 380) and addition of an 11-amino-acid N-terminal linker toenhance chemical coupling to the antibody result in a mutanttoxin referred to as NLysPE38 (see Fig. 1A) (6, 12). Conjuga-tion of NLysPE38 to the murine monoclonal antibody B3forms the immunotoxin LMB-1, a potent therapeutic reagentdesigned to destroy cancer cells expressing the B3 antigen (20,21, 23). Similarly, genetic fusion of the Fv portion of B3 toPE38 results in a smaller yet more cytotoxic recombinantimmunotoxin referred to as LMB-7 (3, 20).Immunotoxins are being evaluated as novel therapeutic
agents for the treatment of cancer and have shown some
* Corresponding author. Mailing address: Laboratory of MolecularBiology, DCBDC, NCI, National Institutes of Health, Building 37,Room 4E16, 9000 Rockville Pike, Bethesda, MD 20892. Phone: (301)496-4797. Fax: (301) 402-1344.
efficacy in early clinical trials. However, a major drawback tothe success of immunotoxins is their immunogenicity (22).Antibodies to the immunotoxin typically appear shortly afterthe first course of treatment, limiting the ability to giverepeated courses and preventing the therapeutic benefits ofcontinued treatment. Humanization of the antibody portioncan decrease the host response to the antibody (11). However,the antigenicity of the toxin is still significant. Identifying themajor immunodominant epitopes of the toxin would be ofgreat value to the design of immunotoxins so that antigenicsites could be removed or altered in order to decrease theoverall immune response to the immunotoxin.As part of the preclinical evaluation of LMB-1 and LMB-7,
it was necessary to perform toxicity studies with monkeys, usingthe protocol proposed for human trials. Sera from the monkeyswere therefore available for immunological analyses. In thepresent study, synthetic overlapping peptides corresponding tothe complete NLysPE38 sequence and mutant PE moleculeswere used to map the immunodominant epitopes recognizedby polyclonal antibodies from monkeys immunized with twoPE38-containing immunotoxins, LMB-1 and LMB-7. TheNLysPE38 toxin is shown to contain several immunodominantcontinuous sequences.
MATERIALS AND METHODS
Peptide synthesis. Peptides were synthesized by CambridgeResearch Biochemicals (Cambridge, United Kingdom) by pre-viously described methods (7). Peptides corresponding to thesequence NLysPE38 were 10 amino acids long and overlappedby 7 amino acids. Each peptide contained an additional4-amino-acid spacer SGSG at the N terminus to which biotinwas attached.
5055
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

5056 ROSCOE ET AL.
Polyclonal sera. Twelve cynomolgus monkeys, male andfemale, received either LMB-1 (monkeys 67, 107, 1201, 1202,4201, and 4202) or LMB-7 (monkeys 58, 61, 122, 222, 151, and242) intravenously on days 1, 3, and 5 as part of a preclinicaltoxicity protocol (performed at Batelle Laboratories, Colum-bus, Ohio) (animal use protocol approved by the NationalInstitutes of Health animal use committee). Each dose was asfollows (in milligrams per kilogram of body weight): monkey107, 3; monkeys 67, 4201, and 4202, 4; monkeys 1201, 1202, 58,and 242, 1; monkeys 122 and 222, 0.25; and monkeys 61 and151, 0.75 and 1.25, respectively. Monkeys were bled on day 19to measure antibody responses to the immunotoxin. Titerswere determined by direct enzyme-linked immunosorbentassay (ELISA). Preimmune sera were screened and did notcontain any anti-PE antibodies. Immunotoxins were preparedas described previously (23).PE mutants and derivatives. Plasmids expressing amino
acids 381 to 613 and 400 to 613 have been described elsewhere(17). A plasmid expressing domain II amino acids 253 to 264was made by PCR using plasmid pMS38f+t (6), which containsthe sequence for NLysPE38. Construction of PE mutants andprotein expression and purification have been described else-where (2).ELISA. The ELISA with peptides was as described else-
where (7, 27). Briefly, biotinylated peptide stocks were sup-plied as a lyophilized powder and reconstituted in 200 ,ul ofdimethyl sulfoxide (final concentration, approximately 8 mg/ml). Prior to the assay, peptides were further diluted 1:1,000 inphosphate-buffered saline (PBS) plus 0.1% bovine serumalbumin (BSA) (ICN Biomedicals Inc.) and 0.1% sodium azide(Sigma). Polystyrene plates (96 well, Immulon 4; DynatechLaboratories) were coated for 1 h at 37°C with 5 ,ug of avidin(Pierce) per ml in bicarbonate buffer (0.2 M NaHCO3 and 0.2M Na2CO3; pH 9.4). The plates were washed three times withTPBS (0.5% Tween 20 in PBS), and peptides (100 ,ul) wereadded in duplicate to the plates, which were then incubated at37°C for 1 h. The plates were washed again with TPBS andblocked with dilution buffer (TPBS plus 1% BSA and 1%ovalbumin [Sigma]) for 1 h at 37°C. After three washings, theplates were incubated overnight with the polyclonal antiseradiluted 1:500 in dilution buffer. The plates were washed fourtimes with TPBS and incubated with peroxidase-conjugatedgoat anti-monkey immunoglobulin G (Cappel) diluted to 1pug/ml in dilution buffer. The plates were washed four timeswith TPBS and then four times with PBS before 100 pul ofsubstrate (1 mg of ABTS [Boehringer Mannheim], 1 pul of 30%H202 per ml in 0.1 M citric acid buffer; pH 4) was added perwell. The reaction was stopped by the addition of 100 ,ul of10% sodium dodecyl sulfate. The A405 was measured with amicroplate reader. For the competitive ELISA, 50 ,ul ofpolyclonal sera was diluted with an equal volume of variouscompetitors in an NLysPE38-coated (1 pug/ml) microtiter plate.Competitors were diluted in PBS plus 0.2% BSA so that theirmolar ratios to the plate-bound NLysPE38 would be 50, 5, 0.5and 0.05 to 1. The plates were incubated for 1 h and thenprocessed as described above. Inhibition of binding to the solidphase was plotted as relative absorbance as a function of themolar ratio of the competitor to the NLysPE38, as follows:relative absorbance (%) = [(A - Ab)/(Am,x - Ab)] x 100,whereA is the absorbance of each well with competitor, Ab isthe absorbance of a well to which no serum is added, andAmaxis the absorbance when no competitor is added.
Neutralization assay. A 25-pl volume of serum was incu-bated with or without 1 jig of various PE derivatives for 2 h at37°C, and immunotoxin LMB-1 or LMB-7 was added at a finalconcentration of 10 or 1 ng/ml, respectively, along with whole
murine immunoglobulin at 100 ng/ml to preadsorb neutralizingantibodies directed at the B3 portion of the immunotoxin. Theability to neutralize was measured as a function of cytotoxicitywith A431 (human epidermal carcinoma) cells as describedpreviously (21).
Computer-aided analyses of PE amino acids. The X-raycrystal structure of PE is available (1) but is missing fourcontiguous sequences, amino acids 485 to 492, 547 to 550, 575to 580, and 601 to 613. The structures of the missing sequenceswere replaced by using the program QUANTA and availablecrystal structure information for human proteins selected fromthe Protein Data Bank files which have similar main-chaingeometry to three residues on either side of the missing aminoacids. The C-terminal residues were constructed with in-housegraphics software GEMM. The surface accessibility profile wasobtained by rolling a 10-A (1-nm) probe over the surface of therevised crystal structure as described previously (14-16). No-votny et al. (16) calculated the contact surface area of ly-sozyme, using a probe with a radius of 10 A (1 nm) rather thanthe solvent-accessible area used here. However, comparison ofthe different strategies for determining surface area versusresidue number for lysozyme are very similar. Data are notsmoothed and are plotted against the residue number.
Elution of sera from peptides. Microtiter plates (96 well)were coated with peptides and incubated with sera as describedabove. The plates were washed four times with TPBS and thenfour times with PBS. A 100-pAl volume of elution buffer (0.1 MHCl [pH 2.4], 1 mg of BSA per ml, and 1 mg of glycine per ml)was added per well, and the plates were incubated for 10 min.Buffer was collected and immediately neutralized with 2 M Trisbase. Eluted antibodies were concentrated on Centricon 30filters (Amicon) which were prewashed with 2 ml of PBS-0.2%BSA. Concentrated antibody was then used in a neutralizationassay as described above.
RESULTS
Serum reactivity to peptides. Polyclonal serum specimensfrom 12 monkeys immunized with either the immunotoxinLMB-1 (monkeys 67, 107, 1201, 1202, 4201, and 4202) orLMB-7 (monkeys 58, 61, 122, 222, 151, and 242) were testedfor reactivity to a panel of 120 synthetic peptides representingthe sequence of NLysPE38, the toxin portion of LMB-1 andLMB-7. The amino acid sequence for NLysPE38 and corre-sponding peptide numbers are shown in Fig. lB. Each peptidecontained 10 amino acids of the NLysPE38 sequence, and thepeptides overlap each other by 7 amino acids. The reactivitiesof the polyclonal monkey sera to the peptides identified severalcontinuous epitopes, as shown in Fig. 2. Peptides to which seradid react are summarized in Table 1. The polyclonal sera didnot react with 75% of the peptides. Comparisons of the profilesof reactivity to the peptides by the monkey sera reveal fourmajor and two minor areas of reactivity. Four of the 12 serumspecimens (from monkeys 1202, 4202, 61, and 122) recognizedamino acids 341 to 359 (peptides 36 to 39). Four of the 12serum specimens (67, 4202, 58, and 122) recognized aminoacids 504 to 516 (peptides 85 and 86). Four of the 12 serumsamples (67, 61, 122, and 242) recognized amino acids 573 to591 (peptides 110 to 113), and 9 of the 12 serum specimensrecognized C-terminal amino acids 591 to 613 (peptides 116 to121). Additional minor antigenic regions are located in pep-tides 13 to 15 (amino acids 272 to 287), showing reactivity in 4of the 12 serum specimens (67, 58, 122, and 222), and 7 of the12 serum samples (67, 1201, 1202, 4201, 122, 222, and 151)recognized peptides 99 to 104 (amino acids 540 to 564). Strongreactivity to peptides 78 and 79 (amino acids 483 to 495) was
INFECr. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

EPITOPE MAPPING OF PE38 5057
PE [
NLysPE38
253
la 1I365 400
l b| III
253 364 381
MLQGTKLMAEE E
253M L Q G T K L MAE E G GS L AA L TA H Q A C3 4 5 6 7 8 9 10
274
R H R Q P R G W E Q L E Q C G Y P V Q R L V A L14 15 16 17 18 19 20 21
306N Q V D Q VI R NA LA S PG S G G D L GE AI
25 26 27 28 29 30 31 32
338L A35
LT LAAA ESE R F VR Q G T G ND EA GAA36 37 38 39 40 41 42 43
273
H L P L E T F T11 12 13
305Y L A A R L S W
22 23 24
337
R E Q P E Q A R33 34
385N G P A D S
44 45
386G D A L L E R N Y P
46 47 48
400T G A E F L49 50
G D G G D V S F S T51 52 53 54
417
R G T Q N W55 56
418TV E RL L QA H R Q LEE R GYVFVGY H G T FL
57 s5 59 60 61 62 63 64 65
450
I V F G G V R A R S Q D L D A I67 68 69 70 71 72
482
A Q D Q E P D78 79
W R G F Y I A G D P A L A73 74 75 76
AR GR I RNGAL LRVYVP RS S L P G80 81 82 83 84 8s 86 87
481
Y G Y77
513F Y R
88
514T B L T L A A
89 90
P A A G V E R L I91 92 93 94
546
545
G H P L P L R L D A I T G P95 96 99 100
577
EEE G GR LET IL GW P LAE RTVV I P S AI PT DP RN101 102 103 104 105 106 107 108 109 110 1ll
578 609
VG G D L DPS S I P DXE QA IS ALP D Y A S Q P G R P P R112 113 114 115 116 117 118 119 120/121
610 613
D L RFIG. 1. (A) Schematic map of PE and NLysPE38. (B) Single-letter code for the amino acid sequence for NLysPE38. Numbers below the amino
acids are the peptide numbers and represent the first amino acid of each 10-amino-acid peptide. Peptides 1, 2, 97, and 98 were synthesized ascontrol peptides and are not related to NLysPE38 and therefore are not depicted. Numbers above the sequence are the amino acid numbers asdefined by the original sequence of PE.
displayed by sera 4201 and 122. Preimmune sera did not reactto any peptides (data not shown).Immunodominant epitopes are located in domain III. To
determine if the continuous epitopes recognized by the poly-clonal sera are representative of the whole polyclonal responseto NLysPE38, competitive ELISAs using individual domains ascompetitors for antibody binding to ELISA plates coated withNLysPE38 were performed. For the assay, 96-well microtiterplates were coated with NLysPE38 and then various competi-tors were added at molar ratios of 0.05, 0.5, 5, and 50 to 1. Sera
were added at the time of addition of competitors at a dilutionof 1:1,000. Figure 3 demonstrates that the immunodominantepitopes recognized by the anti-PE38 serum are located pri-marily in domain III because in 11 of the 12 serum samples (allbut 1202) domain II (amino acids 253 to 364) failed to competefor binding, while domain III, amino acids 400 to 613, com-peted as well or almost as well as whole NLysPE38 in 10 of the12 serum samples (all but 1202 and 61). In four of the 12 serumsamples, denatured NLysPE38 competed better than nativeNLysPE38 (1202 and 61) or as well as native NLysPE38 or
A
B
613
66 kDa
61338 kDa
449E A A Q 8
66
VOL. 62, 1994
1
I
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

5058 ROSCOE ET AL.
1-t'l
3* 3
0
too
ion0a
ll.
41i0
4
41aI
411111IsaMiaa
A:~~~~~q na te q1
o ~~~i
- "~~~
fit ti aI
.1
I
IjA
I
AII
3* ag
=
U
a
=
U
aUU
IIUU
0 UN
IU.? .3* .@
N o;~~~~~~CY ~~~I
I
-ji
i
aa
10
0CY
III0
-A - t - N Iso ro
I
I
N-N f
_* U*
|-~~"~* aI
E
I
I_ S~~~
_~~
5 .S. *I
N~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~IN~~~~~~~~
N~~~~~~~~~~~~~~
3* a
aIIa
Ia
N0
N
I I 4t aNW I
d
I
a 3
iia
imaiII
I
INFEcr. IMMUN.
an
I
a
iaIaI
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

EPITOPE MAPPING OF PE38 5059
FIG. 2. Binding of polyclonal anti-LMB-1 and anti-LMB-7 antibodies to synthetic overlapping peptides covering the sequence of NLysPE38.Twelve monkey serum samples were tested for their reactivities to decapeptides covering the whole NLysPE38 sequence by ELISA as describedin Materials and Methods. Sera were diluted 1:200 or 1:500, depending on the titer to NLysPE38. Peptides were biotinylated and overlapped by7 amino acids. Monkeys 67, 107, 1201, 1202, 4201, and 4202 were immunized with LMB-1; the others were immunized with LMB-7. Each valuerepresents the average of samples done in duplicate. Each experiment was performed at least three times. Standard error was less than 10%. O.D.405, optical density at 405 nm.
domain III (58 and 242). Serum binding to individual domainswas measured by direct ELISA, which demonstrated thatbinding to plate-bound domain II was detectable but was lessthan binding to plate-bound domain III plus Ib, amino acids381 to 613, in 10 of the 12 serum samples (all except 1202 and61; data not shown). Since soluble and therefore nativeNLysPE38 competed for binding to plate-bound NLysPE38almost fully in most of the sera, the major immunodominantepitopes recognized by the sera are still accessible underconditions in which NLysPE38 is bound to the plate.
Neutralizing epitopes are located in domain Ill. To deter-mine the location of neutralizing epitopes, sera were preincu-bated with either domain II (amino acids 253 to 364), domainIII plus part of Ib (amino acids 381 to 613), or domain III alone(amino acids 400 to 613) and tested for their ability toneutralize the immunotoxin LMB-1. If neutralizing antibodiesare directed at a particular domain, then preadsorption withthat domain would make those antibodies unavailable forneutralization of the immunotoxin. Sera were first tested for
their ability to neutralize immunotoxin (Fig. 4). Sera 1201,1202, 61, 222, and 242 did not efficiently neutralize immuno-toxin. This lack of neutralizing antibodies correlated with lowtiters. Sera 67, 107, 4201, 4202, 58, 122, and 151 had highertiters and contained neutralizing antibodies to immunotoxin.The results shown in Fig. 4 demonstrate that NLysPE38preadsorbs all neutralizing antibodies from the sera havingsignificant ability to neutralize the immunotoxin (67, 107, 4201,4202, 58, 122, and 151). Preadsorption with domain II, domainIII plus Ib, or domain III alone by the sera followed byincubation with immunotoxin shows that the neutralizing an-
tibodies are directed primarily at domain III, with the excep-tion of one (4201). In addition, the antibodies which neutralizeLMB-1 in serum 4202 were preadsorbed by heat-denaturedNLysPE38 as well as by domain III.Comparison of predicted regions of immunogenicity and
continuous epitopes recognized by the polyclonal monkey sera.Previous studies have shown that amino acid segments withmore exposed surface area than average are more likely than
TABLE 1. Summary of polyclonal serum reactivities to various peptidesa
ReactivityPeptide no. No.ofAnti-LMB-7sera re(amino acids) Anti-LMB-1 sera Anti_LMB_7_sera_ reactota
67 107 1201 1202 4201 4202 58 61 122 222 151 242
13 (272-281) - - - - - ++ - + - - 2/1214 (275-284) ++ - - - - - - - - + - - 2/1215 (278-287) + - - - - - - - + + - - 3/1222 (299-308) - - - - - - - - - - + - 1/1232 (329-338) - - - - - - - - ++ - - - 1/1233 (332-341) - - - - - - - - + - - - 1/1236 (341-350) - - - + - - - +++ + - - - 3/1237(344353) - - - + - - - 1/1238 (347-356) - - - - - - - - - - - 1/1239 (350-359) - - - - + - - - - - - 2/1240 (353-362) - - - - - + - - - - - - 1/1246 (387-396) - - - - - - + - - - - - 1/1278 (483-492) - - - - + - - - ++ - - - 2/1279 (486-495) - - - - - - - + - - - 2/1285 (504-513) +++ - - - - - 3/1286 (507-516) - - - - - - ++ - - - - 1/1296 (537-546) - - - - + - - - - - - - 1/1299(540-549) - - - + - - - 2/12102 (549-558) - - - - - - - - - - - + 1/12103 (552-561) - - - - + - - - - + - - 2/12104 (555-564) ++ - + - - - - - - - + - 3/12105 (558-567) - - - - - - - - - - + - 1/12109 (570-579) +++ - - - - - - - - - - - 1/12110 (573-582) +++ - - - - - - - - - - - 1/12111 (576-585) - - - - - - - - - - - + 1/12113 (582-591) - - - - - - - ++ +++ - - - 2/12115 (588-597) - - - - - - - ++ - - - 2/12116 (591-600) - ++ ++ - - - ++ - - - +++ ++ 4/12117 (594-603) - + ++ - - - ++ - - - ++ - 4/12119 (600-609) - - - ++ - - - ++ ++ +++ ++ 6/12120 (603-612) - - - + +++- - +++ + ++ + 7/12121 (604-613) - - - - + - +++- - - - 3/12
a Scoring was based on the A405 as follows: +, A405 75 to 100% above background but <1.0; + +, A405> 1.0; + + +, A405 > 2.0.
VOL. 62, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

5060 ROSCOE ET AL.
IN 11avlq
a2
V-4
Aa4#tA
a
a
X
X
C4
(%) Wuvqosqy OARfVt[(h) Wutq3osqV OAfltjOf
INFECT. IMMUN.
Q0
3
0.1:
a0
. 0_
,
0
.
-0
Ia.8
0~o2_U
00
o_ I0
*04,0Z
0
0
.8_0
0I0
800
(%6) -I-UE'qO" OAjjTj0'f
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

EPITOPE MAPPING OF PE38 5061
FIG. 3. Reactivities of polyclonal sera to NLysPE38 in the presence of various competitors. NLysPE38 was immobilized on a 96-well microtiterplate and incubated with various competitors at molar ratios of 50, 5, 0.5, or 0.05 to 1. Sera were added in equal volumes at a dilution of 1:1,000.Competitors were NLysPE38 (0), NLysPE38 denatured by being heated to 100°C for 20 min (0), domain II (amino acids 253 to 364) (A), domainIII plus lb (amino acids 381 to 613) (E), or domain III alone (amino acids 400 to 613) (-). Results are plotted as percent A405 of the control(without competitor). Each value represents the average of three determinations. Experiments were performed at least three times. Standard errorwas less than 12%.
segments with less surface area to be antigenic sites recognizedby antibodies (8, 15, 16). It has been suggested (16) that amongthe residues in an epitope, only a few residues favorablycontribute to antigen-antibody binding, and the energeticallymost important residues belong to the most protruding part ofthe protein surface. Knowledge of the crystal structure of PEmade it possible to use a computer-aided analysis of theexposed surface area of each amino acid and predict itscontribution toward defining immunogically distinct epitopes.The accessible surface area of PE was calculated by using a10-A (1-nm) probe as described previously (14-16). Figure 5compares the average absorbances of all 12 serum specimens
A130
10-
5-430
10
61 107 4 15
B130S
I;11090-
O70-
50-
30-
Z 10-
-10- abcdef abcdef abcdef58 122 151
SERAFIG. 4. Neutralization of immunotoxin LMB-1 (A) or LMB-7 (B)
preadsorbed with various domains. A 25-,ul sample of each serum waspreincubated with either nothing (a), NLysPE38 (b), denaturedNLysPE38 (c), domain II (d), domain III plus Ib (e), or domain III (f)for 2 h at 37°C. The final concentration of all competitors wasapproximately 1 ,uM. LMB-1 (10 ng/ml with whole immunoglobulin at100 ng/ml; final concentration, 50 pM) or LMB-7 (1 ng/ml; finalconcentration, 15 pM) was added to each sample and incubated withA431 cells overnight. The ability to neutralize LMB-1 was measured ina cytotoxicity assay.
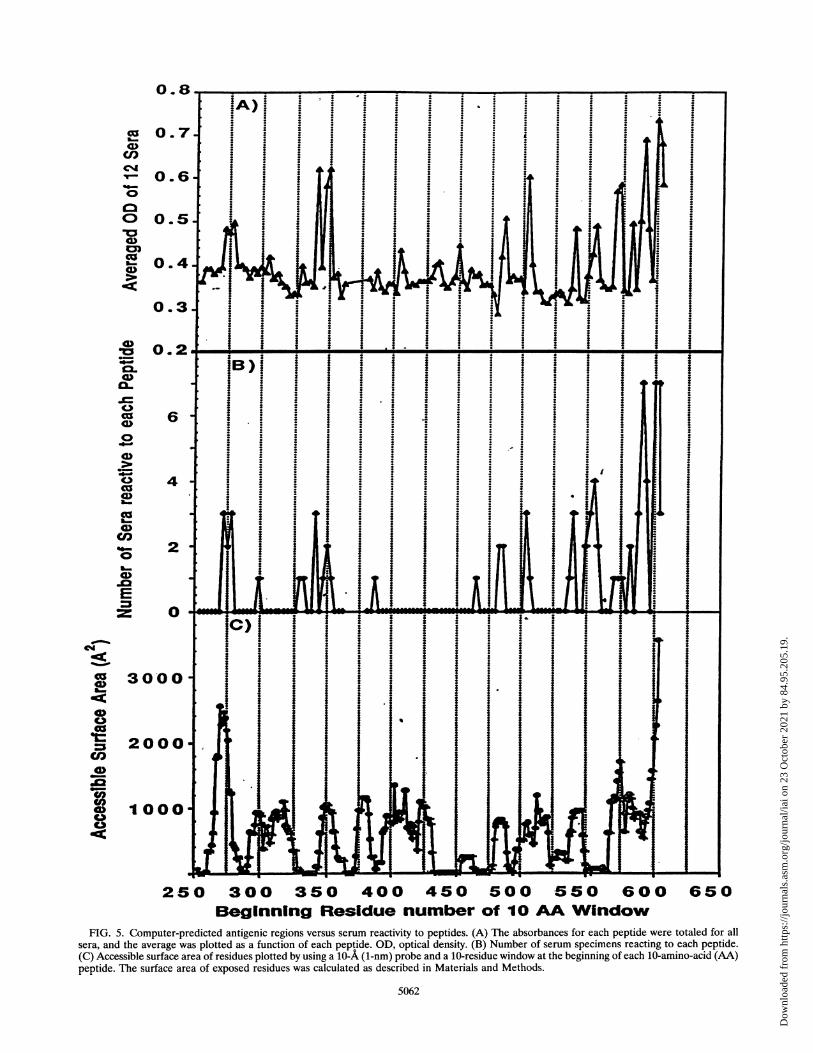
for each peptide (Fig. SA) and the frequency of recognition ofeach peptide (Fig. SB) with the accessible surface area of theresidues in PE (Fig. SC). The comparison of the epitopesidentified by peptide ELISA with the surface areas of thepeptides indicates that all antigenic regions are strongly pre-dicted, although reactivity to an individual peptide is not. Forexample, peptide 103, containing amino acids 552 to 561, is notpredicted to be antigenic, although amino acids 565 to 591 are.In addition, the analysis of the exposed areas predicts severalantigenic sites which do not show any peptide reactivity. Theseare amino acids 300 to 325 and 400 to 425; they could beantigenic sites not detectable with synthetic peptides or couldbe altered in their structure in PE38 because of the deletion ofamino acids 365 to 380.
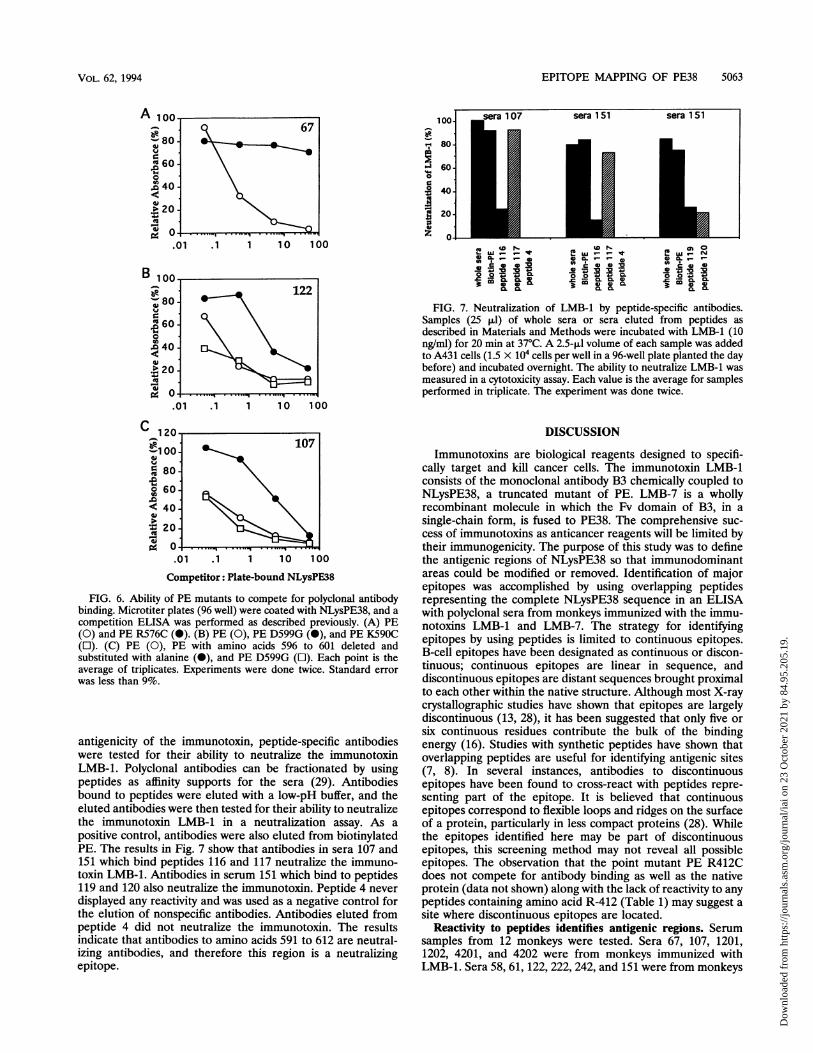
Polyclonal reactivity of sera to PE mutants. The computer-analyzed surface exposure of individual amino acids indicatesthat the amino acids with the greatest surface area are R-276,R-279, R-412, E-522, R-576, and the C-terminal amino acidsQ-603, P-608, and 610 to 613 (EDLK). All of these residuesexcept R-412 and E-522 are located in areas of primary peptidereactivity. To determine the role of individual amino acidspredicted to be residues involved in the antigenicity of thecontinuous epitopes, mutations at specific sites in PE weremade and the mutant proteins were compared with native PEin a competitive ELISA. The mutant proteins produced werePE R576C (R-to-C mutation at position 576), PE K590C, PED599G, and PE A596-601/A (amino acids 596 to 601 deletedand replaced with a single alanine). The results in Fig. 6 showthat these mutants compete less effectively for antibody bind-ing to plate-bound NLysPE38 than native PE. The R576Cmutation results in a protein that fails to compete for bindingto plate-bound NLysPE38 with antibodies in serum 67 com-pared with native PE (Fig. 6A). PE mutants D599G andK59OC were tested for binding by serum 122 (Fig. 6B). Theresults demonstrate that while the K59OC mutant does com-pete as well as PE, the mutant D599G competes much lesseffectively. Similarly, PE A596-601/A was less effective thannative PE at competing for binding of serum 107 (Fig. 6C). Themutant D599G, however, competes as well as native PE. Thisindicates that residue 599 is not critical for binding to peptide116 or 117 (amino acids 591 to 603) or, alternatively, that theantibodies binding to those peptides in serum 107 are notpresent in sufficient quantity to detect any measurable differ-ence in binding. The inability to compete was not universal forall sera recognizing the same peptide (data not shown),indicating differences in the concentration of specific antibod-ies in each serum. Interestingly, R412C was tested and did notcompete as effectively as native PE for antibody binding in 5 ofthe 12 serum specimens, suggesting that amino acid 412 isinvolved in an epitope not identified by the overlappingpeptides (data not shown). The results do correlate, however,with the predicted role of amino acid 412 in immunogenicitybased on the large surface area of the residue.
Elution of peptide-specific antibodies. The polyclonal mon-key sera reacted most strongly and frequently to the C-terminal amino acids 591 to 612, represented by peptides 116,117, 119, and 120. To better assess the role of this region in the
VOL. 62, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.
II I
250 300 350 400 450Beginning Residue number
W.Ii
I
S00 550 600 650of 10 AA Window
FIG. 5. Computer-predicted antigenic regions versus serum reactivity to peptides. (A) The absorbances for each peptide were totaled for allsera, and the average was plotted as a function of each peptide. OD, optical density. (B) Number of serum specimens reacting to each peptide.(C) Accessible surface area of residues plotted by using a 10-A (1-nm) probe and a 10-residue window at the beginning of each 10-amino-acid (AA)peptide. The surface area of exposed residues was calculated as described in Materials and Methods.
5062
0
0.7
0.6
0.5
0
0.3
0.2
cn
*-
0
aCl40o
0
ci
0-
4a
a-
0
0
0
1006-
Colcc%
C0
0)S:
%t00.-f!
0C)0
co
C)
CD7.3
:<
3000
2000
1000
I
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.
EPITOPE MAPPING OF PE38 5063
A 100-
,60-
'400, 40-
.20-
.a
B 100-
80.v
0 60-
40-
. 20-
120-81i00.g 80
qo, 60-
< 40.
*! 20.(A:O
00
T!
A
3
01 .1 1 10 100
)1 . .1 .
Dl .1 1 10 10(
I I~~~~~~~~~~~~~~~~
.01 .1 1 10 100
Competitor: Plate-bound NLysPE38
FIG. 6. Ability of PE mutants to compete for polyclonal antibodybinding. Microtiter plates (96 well) were coated with NLysPE38, and a
competition ELISA was performed as described previously. (A) PE(0) and PE R576C (0). (B) PE (0), PE D599G (0), and PE K590C(l). (C) PE (0), PE with amino acids 596 to 601 deleted andsubstituted with alanine (0), and PE D599G (O). Each point is theaverage of triplicates. Experiments were done twice. Standard errorwas less than 9%.
antigenicity of the immunotoxin, peptide-specific antibodieswere tested for their ability to neutralize the immunotoxinLMB-1. Polyclonal antibodies can be fractionated by usingpeptides as affinity supports for the sera (29). Antibodiesbound to peptides were eluted with a low-pH buffer, and theeluted antibodies were then tested for their ability to neutralizethe immunotoxin LMB-1 in a neutralization assay. As apositive control, antibodies were also eluted from biotinylatedPE. The results in Fig. 7 show that antibodies in sera 107 and151 which bind peptides 116 and 117 neutralize the immuno-toxin LMB-1. Antibodies in serum 151 which bind to peptides119 and 120 also neutralize the immunotoxin. Peptide 4 neverdisplayed any reactivity and was used as a negative control forthe elution of nonspecific antibodies. Antibodies eluted frompeptide 4 did not neutralize the immunotoxin. The resultsindicate that antibodies to amino acids 591 to 612 are neutral-izing antibodies, and therefore this region is a neutralizingepitope.
100-
80-
60-
40-
20-
04
sera 1 07 sera 1 51 sera 1 51icmE;t LO ................¢°etem0U) *9.41 s
COCa, *O e
0 n.' . a n .
FIG. 7. Neutralization of LMB-1 by peptide-specific antibodies.Samples (25 p.l) of whole sera or sera eluted from peptides asdescribed in Materials and Methods were incubated with LMB-1 (10ng/ml) for 20 min at 37°C. A 2.5-ilJ volume of each sample was addedto A431 cells (1.5 x 104 cells per well in a 96-well plate planted the daybefore) and incubated overnight. The ability to neutralize LMB-1 wasmeasured in a cytotoxicity assay. Each value is the average for samplesperformed in triplicate. The experiment was done twice.
DISCUSSION
Immunotoxins are biological reagents designed to specifi-cally target and kill cancer cells. The immunotoxin LMB-1consists of the monoclonal antibody B3 chemically coupled toNLysPE38, a truncated mutant of PE. LMB-7 is a whollyrecombinant molecule in which the Fv domain of B3, in asingle-chain form, is fused to PE38. The comprehensive suc-cess of immunotoxins as anticancer reagents will be limited bytheir immunogenicity. The purpose of this study was to definethe antigenic regions of NLysPE38 so that immunodominantareas could be modified or removed. Identification of majorepitopes was accomplished by using overlapping peptidesrepresenting the complete NLysPE38 sequence in an ELISAwith polyclonal sera from monkeys immunized with the immu-notoxins LMB-1 and LMB-7. The strategy for identifyingepitopes by using peptides is limited to continuous epitopes.B-cell epitopes have been designated as continuous or discon-tinuous; continuous epitopes are linear in sequence, anddiscontinuous epitopes are distant sequences brought proximalto each other within the native structure. Although most X-raycrystallographic studies have shown that epitopes are largelydiscontinuous (13, 28), it has been suggested that only five orsix continuous residues contribute the bulk of the bindingenergy (16). Studies with synthetic peptides have shown thatoverlapping peptides are useful for identifying antigenic sites(7, 8). In several instances, antibodies to discontinuousepitopes have been found to cross-react with peptides repre-senting part of the epitope. It is believed that continuousepitopes correspond to flexible loops and ridges on the surfaceof a protein, particularly in less compact proteins (28). Whilethe epitopes identified here may be part of discontinuousepitopes, this screening method may not reveal all possibleepitopes. The observation that the point mutant PE R412Cdoes not compete for antibody binding as well as the nativeprotein (data not shown) along with the lack of reactivity to anypeptides containing amino acid R-412 (Table 1) may suggest asite where discontinuous epitopes are located.
Reactivity to peptides identifies antigenic regions. Serumsamples from 12 monkeys were tested. Sera 67, 107, 1201,1202, 4201, and 4202 were from monkeys immunized withLMB-1. Sera 58, 61, 122, 222, 242, and 151 were from monkeys
107
VOL. 62, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

5064 ROSCOE ET AL.
immunized with LMB-7. The results identify several immuno-dominant regions and indicate that the polyclonal antibodyresponses to peptides among all the sera are very similar. Assummarized in Table 1, the peptide reactivities of the 12 serumsamples can be grouped into minor and major antigenicregions. The regions are located primarily in domain III(amino acids 504 to 516, 540 to 564, 573 to 591, and 591 to613), although two of the antigenic regions are located indomain II (amino acids 272 to 287 and 341 to 359). Thedesignation of antigenic regions was based on the proximity ofthe amino acids to each other. For example, the crystalstructure shows that amino acids 485 to 492 are not close toamino acids 504 to 516, and therefore the antibody reactivity topeptides 78 and 79 were not grouped with peptides 85 and 86as an antigenic site. A low level of reactivity was seen forpeptides 13 to 15 (amino acids 272 to 287) with 4 of the 12serum samples (67, 58, 122, and 222). Three of the four serumsamples were from monkeys immunized with LMB-7, indicat-ing that this region may be more available for antibody bindingon LMB-7 than LMB-1. A likely explanation is that in LMB-1the antibody is coupled to domain II and blocks the bindingsite, whereas in LMB-7 the Fv domain fused at the N terminusis significantly smaller than the whole antibody. Previousexperiments by Ogata et al. demonstrated that during isolationof monoclonal antibodies to PE40 (PE amino acids 253-613missing domain Ta) three of the eight antibodies isolated wereto this region of domain H (17). Of the four antigenic sites indomain III, amino acids 540 to 564 had an overall low level ofreactivity but a very high frequency of reactivity (7 of 12 serumspecimens). One explanation may be that this is a highlyantigenic site inadequately represented by peptides.Domain III is the principal domain for antibody reactivity
and neutralizing antibodies. The peptide reactivity resultssuggest that the bulk of the antibody reactivity in the sera isdirected at domain III, which represents two-thirds of the massof PE38. This was confirmed by competition ELISA using PEderivatives representing the individual domains: domain II(amino acids 253 to 364), domain III plus Ib (amino acids 381to 613), and domain III alone (amino acids 400 to 613). Theresults showed that for ten of the 12 serum samples (67, 107,1201, 4201, 4202, 58, 122, 222, 151, and 242) domain IIIcompeted as well as native NLysPE38. Domain II also failed tocompete for any significant binding in any of the sera. In fourserum specimens (1202, 4201, 61, and 242) denaturedNLysPE38 was a significant competitor, and in two cases (1202and 61) it was the only competitor. This indicates that themajor epitopes recognized in these four serum samples aresequential determinants present on NLysPE38 when it iscomplexed to the monoclonal antibody and not in nativeNLysPE38 alone or that they were uncovered as a result ofdegradation of the immunotoxin either in vitro during handlingor in vivo during immunization of the animal. Interestingly, thetwo serum samples (1202 and 61) were similar in having astrong antibody response to peptides 38 and 39, whose sharedamino acids are 350 to 356 (FVRQGTG). This sequence waspredicted to have significant surface exposure as well (Fig. 5).The neutralizing antibodies in the sera are directed primarily
at epitopes in domain III in PE38-containing immunotoxins.The elution of peptide-specific antibodies which recognizeamino acids 591 to 612 revealed these to be neutralizingantibodies. Previously, three neutralizing determinants in PE(amino acids 1 to 613) were identified; one is in domain Ia, thereceptor-binding domain; another is in domain Ib (amino acids392 to 405) (26); and the third is associated with the C-terminalamino acids 581 to 613 (18). Our results confirm the immu-nodominance of the C terminus for PE38 as well as PE but also
indicate that the immunodominant sites are not restricted tothe last 30 amino acids in domain ITT. The neutralizing capacityof sera 1201, 1202, 61, 222, and 242 was low and was reflectedin the low titer for these sera as well, suggesting a decreasedimmune response to the neutralizing epitopes, probably due tothe deletion of domain I and part of Tb.
Residues with large surface areas are located in antigenicepitopes. Because the crystal structure of PE is known (1), itcould be used to calculate the relative contributions of theindividual residues to antigenicity on the basis of their relativesurface exposure. Elucidation of the crystal structure of PEfailed to identify the positions of residues at four sites in themolecule, presumably because of the overall flexibility of thoseparticular regions. Three of the missing segments are predictedto be loops with a significant degree of surface exposure. Thefourth is at the carboxy terminus of the protein. The missingloop regions are composed of amino acids 485 to 492, 547 to550, and 575 to 580. The predominant reactivity by the sera wasto peptides located in domain III, particularly at the regions ofthese loops. Sera 4201 and 122 recognized peptides 78 and 79(amino acids 483 to 495). Sera 4201, 1202, and 58 stronglyrecognized and sera 151 and 242 to a lesser extent recognizedamino acids 540 to 558 (peptides 99 to 102). Additionalreactivity to amino acids surrounding this loop region (pep-tides 103 and 104, amino acids 552 to 564) by sera 67, 1201,4202, and 151 and at a lower dilution serum 122 was seen.Serum 67 reacted strongly to peptides 109 and 110, which
share the amino acids 573 to 579 (TDPRNVG). By usingmodeling techniques, the missing sequences were filled intothe crystal structure and the amino acids with the largestaccessible surface area were identified as R-276, R-279, R-412,E-522, and R-576 and the C-terminal amino acids. Site-directed mutagenesis of R-576 to a cysteine produces a PEmolecule which cannot compete for binding compared withnative PE, indicating that the major antibody response ofserum 67 is directed at this region. The R576C mutant is fullycytotoxic compared with PE, implying that its inability to bindantibodies is not due to improper folding. Peptides 13, 14, and15 share four amino acids (278 to 281 [PRGW]). The comput-er-aided analysis of the predicted antigenic amino acidsshowed that R-276 and R-279 had a high degree of surfaceaccessibility and were likely candidates for epitopes. Thepeptide data confirm these results. However, mutant PEmolecules with mutations at these residues compete for bind-ing as effectively as native PE (data not shown), furthercorroborating the results showing that the antibodies to thisregion are not a substantial part of the entire antibody pool(Table 1). The crystal structure of PE also did not identify theposition of amino acids 600 to 613, which is the region ofhighest immunogenicity detected by this system; 7 of the 12serum specimens (1202, 4201, 58, 122, 222, 151, and 242) bindto peptides 119, 120, and/or 121 (amino acids 600 to 613),whose minimal shared amino acids are QPGKPPR. Twoadditional serum samples (107 and 1201) showed strong reac-tivity to peptides 116 and 117, as did sera 58, 151, and 242 and,to a lesser extent, 122. Peptides 116 and 117 (amino acids 591to 603) share amino acids 594 to 600, whose sequence isISALPDY. When amino acids 596 to 600 are deleted andreplaced with a single A, the mutant competes less than thenative protein for binding to antibodies in serum 107. A toxinwith a mutation of just residue D-599 to G competes fully,indicating that this is not a critical residue in this epitope.These results were not the same for all sera tested and mayreflect a difference in specific antibody populations within eachserum. In addition, strong reactivity to a peptide does notnecessarily indicate that the peptide-specific antibody is
INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.

EPITOPE MAPPING OF PE38 5065
present in a large quantity within the entire pool of NLysPE38antibodies. For example, when a mutant PE in which aminoacids 485 to 492 were deleted and replaced with a stretch ofglycines and serines was tested for its ability to compete forbinding to antibodies in sera 4201 and 122, which bound topeptides 78 and 79 (shared amino acids 486 to 492), the mutantcompeted as well as native PE (data not shown). The resultsfor point mutants in a competition ELISA are difficult tointerpret with polyclonal sera because of the heterogeneousnature of the antibodies.PE is a very immunogenic molecule. Others have identified
domain Ia as a major site for neutralizing antibodies (17).Removal of domain Ia as in NLysPE38 decreases the immuneresponse to this molecule; however, these results show thatstrong neutralizing antibodies to domain III are still produced.It has been previously shown that mutation of amino acid 553destroys the cytotoxic activity of PE (4, 19). This region islocated in a potential antigenic site (Table 1) and lies in orclose to the area to which the neutralizing antibodies in thesesera are directed. Removal of this antigenic site will rely onaltering surrounding amino acids which do not influencecytotoxicity. Antibodies to PE which neutralize cytotoxicitywithout altering ADP-ribosyl transferase activity have beenidentified (5). Sera from mice have shown a pattern ofreactivity similar to that of primate antisera recognizing pep-tides 35, 79, 109, 116, and 117 (data not shown), indicating thatthe results obtained from murine antibodies can be extrapo-lated to primates. Therefore, it should be possible to generatea panel of mouse monoclonal antibodies to PE38 to moreprecisely map the epitopes.We have identified several sites within domain III of PE that
are highly immunogenic in monkeys. If these sites are alsopresent in human serum, it may be possible to eliminate themby mutation or to mask them by polyethylene glycol conjuga-tion in order to make less immunogenic immunotoxins forcancer therapy. Alternatively, these antigenic sites could beused toward vaccine development, and the antibodies theygenerate could be candidates for therapy against pseudomonasinfections.
REFERENCES1. Allured, V. A., R J. Collier, S. F. Carroll, and D. B. McKay. 1986.
Structure of exotoxin A of Pseudomonas aeruginosa at 3.0 ang-strom. Proc. Natl. Acad. Sci. USA 83:1320-1324.
2. Benhar, I., Q. C. Wang, D. Fitzgerald, and I. Pastan. 1994.Pseudomonas exotoxin A mutants: replacement of surface-ex-posed residues in domain III with cysteine residues that can bemodified with PEG in a site-specific manner. J. Biol. Chem. 269:13398-13404.
3. Brinkman, U., L. H. Pai, D. J. Fitzgerald, M. Willingham, and I.Pastan. 1991. B3(Fv)-PE38KDEL, a single-chain immunotoxinthat causes complete regression of a human carcinoma in mice.Proc. Natl. Acad. Sci. USA 88:8616-8620.
4. Carroll, S. F., and R J. Collier. 1987. Active site of Pseudomonasaeruginosa exotoxin A. J. Biol. Chem. 262:8707-8711.
5. Chia, J. K. S., M. Pollack, D. Avigan, and S. Steinbach. 1986.Functionally distinct monoclonal antibodies reactive with enzy-matically active and binding domains of Pseudomonas aeruginosatoxin A. Infect. Immun. 52:756-762.
6. Debinski, W., and I. Pastan. 1992. Monovalent immunotoxincontaining truncated form of Pseudomonas exotoxin as potentantitumor agent. Cancer Res. 52:1-7.
7. Geysen, H. M., S. J. Rodda, T. J. Mason, G. Tribbick, and P. G.Schoofs. 1987. Strategies for epitope analysis using peptide syn-thesis. J. Immunol. Methods 102:259-274.
8. Geysen, H. M., J. A. Tainer, S. J. Rodda, T. J. Mason, H.Alexander, E. D. Getzoff, and R D. Lerner. 1987. Chemistry ofantibody binding to a protein. Science 235:1184-1190.
9. Hwang, J., D. J. P. Fitzgerald, S. Adhya, and I. Pastan. 1987.Functional domains of Pseudomonas exotoxin identified by dele-tion analysis of the gene expressed in E. coli. Cell 48:129-136.
10. Iglewski, B. H., and D. Kabat. 1975. NAD-dependent inhibition ofprotein synthesis by Pseudomonas aeruginosa toxin A. Proc. Natl.Acad. Sci. USA 72:2284-2288.
11. Jolliffe, L K. 1993. Humanized antibodies: enhancing therapeuticutility through antibody engineering. Int. Rev. Immunol. 10:241-250.
12. Kondo, T., D. Fitzgerald, K. Chaudhury, S. Adhya, and L. Pastan.1988. Activity of immunotoxins constructed with modified Pseudo-monas exotoxin A lacking the cell recognition domain. J. Biol.Chem. 263:9470-9475.
13. Laver, W. G., G. M. Air,R G. Webster, and S. J. Smith-Gill. 1990.Epitopes on protein antigens: misconceptions and realities. Cell61:553-556.
14. Lee, B. K., and F.M. Richards. 1971. The interpretation of proteinstructures: estimation of static accessibility. J. Mol. Biol. 55:379-400.
15. Novotny, J. 1991. Protein antigenicity: a thermodynamic approach.Mol. Immunol. 28:201-207.
16. Novotny, J., R E. Bruccoleri, and F. A. Saul. 1989. On theattribution of binding energy in antigen-antibody complexesMcPC 603, D1.3 and HyHEL-5. Biochemistry 28:4735-4749.
17. Ogata, M., I. Pastan, and D. Fitzgerald. 1991. Analysis of Pseudo-monas exotoxin activation and conformational changes by usingmonoclonal antibodies as probes. Infect. Immun. 59:407-414.
18. Olson, J. C., A. N. Hamood, T. S. Vincent, E. H. Beachey, and B. H.Iglewski. 1990. Identification of functional epitopes of Pseudomo-nas aeruginosa exotoxin A using synthetic peptides and subcloneproducts. Mol. Immunol. 27:981-993.
19. Oppenheimer, N., and J. W. Bodley. 1981. Diphtheria toxin; siteand configuration of diphthamide in elongation factor 2. J. Biol.Chem. 256:8579-8581.
20. Pai, L., and I. Pastan. 1993. Immunotoxin therapy for cancer.JAMA 269:78-81.
21. Pai, L. H., J. K. Batra, D. J. Fitzgerald, M. C. Willingham, and I.Pastan. 1991. Anti-tumor activities of immunotoxins made ofmonoclonal antibody B3 and various forms of Pseudomonasexotoxin. Proc. Natl. Acad. Sci. USA 88:3358-3362.
22. Pai, L. H., M. A. Bookman, R J. Ozols, et al. 1991. Clinicalevaluation of intraperitoneal Pseudomonas exotoxin immunocon-jugate OVB3-PE in patients with ovarian cancer. J. Clin. Oncol.9:2095-2103.
23. Pai, L. H., and I. Pastan. 1993. The use of immunotoxins forcancer therapy. Eur. J. Cancer 29A:1606-1609.
24. Pastan, I., V. Chaudhury, and D. J. Fitzgerald. 1992. Recombinanttoxins as novel therapeutic agents. Annu. Rev. Biochem. 61:331-354.
25. Pastan, I., E. T. Lovelace, M. C. Gallo, A. V. Rutherford, J. L.Magnani, and M. C. Willingham. 1991. Characterization of mono-clonal antibodies Bi and B3 that react with mucinous adenocar-cinomas. Cancer Res. 51:3781-3787.
26. Rutault, K, M. J. Vacheron, M. Guinand, and G. Michel. 1993.Comparative immunochemistry of two fragments from domains Iband II of Pseudomonas aeruginosa exotoxin A. Infect. Immun. 61:5417-5420.
27. Savoca, R, C. Schwab, and H. R Bosshard. 1991. Epitopemapping employing immobilized synthetic peptides. J. Immunol.Methods 141:245-252.
28. Shwab, C., A. Twardek, T. P. Lo, G. D. Brayer, and H. RBosshard. 1993. Mapping antibody binding sites on cytochrome cwith synthetic peptides: are results representative of the antigenicstructure of proteins? Protein Sci. 2:175-182.
29. Tribbick, G., B. Triantafyllou, R Lauricella, S. J. Rodda, T. J.Mason, and H. M. Geysen. 1991. Systematic fractionation ofserum antibodies using multiple antigen homologous peptides asaffinity ligands. J. Immunol. Methods 139:155-166.
30. Vitetta, E. S., R J. Fulton, R D. May, M. Till, and J. W. Uhr. 1987.Redesigning nature's poisons to create antitumor reagents. Sci-ence 238:1098-1104.
VOL. 62, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 23
Oct
ober
202
1 by
84.
95.2
05.1
9.


















