
Buffalo (Bubalus bubalis) ES Cell–Like Cells are Capable of In Vitro Skeletal Myogenic...
8
Buffalo (Bubalus bubalis) ES Cell–Like Cells are Capable of In Vitro Skeletal Myogenic Differentiation MK Singh, KP Singh, D Kumar, RA Shah, T Anand, MS Chauhan, RS Manik, SK Singla and P Palta Embryo Biotechnology Laboratory, Animal Biotechnology Centre, National Dairy Research Institute, Karnal, India Contents When buffalo embryonic stem (ES) cell–like cells that expressed surface markers SSEA-4, TRA-1-60, TRA-1-81, CD9 and CD90 and intracellular markers OCT4, SOX2 and FOXD3, as shown by immunofluorescence, and that expressed REX-1 and NUCLEOSTEMIN as confirmed by RT-PCR, were subjected to suspension culture in hanging drops in absence of LIF and buffalo foetal fibroblast feeder layer support, they differentiated to form three-dimensional embry- oid bodies (EBs). Of 231 EBs examined on Day 3 of suspension culture, 141 (61.3 ± 3.09%) were of compact type, whereas 90 (38.4 ± 3.12%) were of cystic type. The cells obtained from EBs were found to express NF-68 and NESTIN (ectodermal lineage), BMP-4 and a-skeletal actin (mesodermal lineage), and a-fetoprotein, GATA-4 and HNF-4 (endodermal lineage). When these EBs were cultured on gelatin-coated dishes, they spontaneously differentiated to several cell types such as epithelial- and neuron-like cells. When EBs were cultured in the presence of 1 or 2% DMSO or 10 )8 M or 10 )7 M retinoic acid for 25 days, ES cells could be directed to form muscle cell–like cells, the identity of which was confirmed by expression of a-actinin by immunofluorescence and of MYF-5, MYOD and MYOGENIN genes by RT-PCR. MYOD was first detected on Day 10 in both treatment groups and on Day 15 in controls, whereas MYOGENIN was first detected on Day 10, Day 15 and Day 25 in the presence of retinoic acid, in the presence of DMSO and in controls, respectively. The present study demonstrates the ability of buffalo ES cell–like cells to undergo directed differentiation to cells of skeletal myogenic lineage. Introduction Embryonic stem (ES) cells are a special kind of cells derived from the inner cell mass (ICM) of blastocyst stage embryos. These cells possess unique characteristics of being capable of undergoing unlimited number of symmetrical mitotic cell divisions without differentia- tion, maintaining a stable diploid karyotype, and being capable of either entering a path leading to a fully differentiated cell or remaining as stem cells. Following first reports on establishment of ES cells from preim- plantation murine embryos (Evans and Kaufman 1981; Martin 1981), these have been established in a number of mammalian species including many farm animal species such as cattle, sheep goat, pig and horse (For reviews, see Munoz et al. 2009; Nowak-Imialek et al. 2011). Unlimited self-renewal, ability to differentiate to all the cell types found in embryonic and adult organism, refractoriness to senescence and high versatility to genetic manipulation make ES cells very useful as basic research tools for understanding processes such as differentiation, organogenesis and early mammalian development besides being useful in regenerative ther- apy in humans. However, no proven ES cell lines colonizing the germ line have been established for vertebrate species other than mouse and chicken thus far. One of the major limitations for utilizing the thera- peutic potential of human ES cells in muscular dystro- phy has been the very little success at directing these cells into skeletal muscle lineage. In the mouse model, a primary reason for this is the paucity of paraxial mesoderm formation during embryoid body (EB) in vitro differentiation (Darabi et al. 2008). There is a need to develop alternate models that could supplement the human and mouse models for generating informa- tion required for developing sufficient populations of skeletal myocytes for potential cell-based therapies. Buffalo is a suitable candidate for supplementing the mouse model as it does not suffer from some of the limitations of mouse model such as short lifespan, physiology and anatomy very different from humans. We have earlier reported derivation and characteriza- tion of ES cell–like cells from in vitro produced buffalo embryos (Verma et al. 2007; Anand et al. 2009) and the development of a culture system suitable for maintain- ing buffalo ES cells over long periods of time (Sharma et al. 2011). No information is available on the ability of buffalo ES cell–like cells to be directed to differentiate to any particular cell type. The aim of the present study was to study the ability of these cells to undergo directed differentiation to skeletal myogenic lineage. Materials and Methods Reagents and media All the chemicals and media were purchased from Sigma Chemical Co., St. Louis, MO, USA, unless otherwise indicated. Knock-out DMEM and Knock-out serum replacer were purchased from Gibco, NY, USA. Foetal bovine serum (FBS; Hyclone, Logan, UT, US, cat no. CH30160.02) of same batch was used for in vitro embryo production and for feeder layer cell culture. The plasticware were purchased either from Becton, Dickin- son and Co., Lincoln Park, NJ, USA, or from Nunc, Rosklide, Denmark. Syringes of assorted sizes were purchased from Henke Saas Wolf GmBH, Tuttingen, Germany, whereas the 0.22-lm filters were from Millipore Corp., Bedford, MA, USA. The primary antibodies used for the detection of pluripotency-related markers by immunofluorescence staining and the secondary antibody goat anti-rabbit IgG-FITC conju- gate were purchased from Chemicon International, Reprod Dom Anim doi: 10.1111/j.1439-0531.2012.02146.x ISSN 0936-6768 ȑ 2012 Blackwell Verlag GmbH
Transcript of Buffalo (Bubalus bubalis) ES Cell–Like Cells are Capable of In Vitro Skeletal Myogenic...

Buffalo (Bubalus bubalis) ES Cell–Like Cells are Capable of In Vitro Skeletal
Myogenic Differentiation
MK Singh, KP Singh, D Kumar, RA Shah, T Anand, MS Chauhan, RS Manik, SK Singla and P Palta
Embryo Biotechnology Laboratory, Animal Biotechnology Centre, National Dairy Research Institute, Karnal, India
Contents
When buffalo embryonic stem (ES) cell–like cells thatexpressed surface markers SSEA-4, TRA-1-60, TRA-1-81,CD9 and CD90 and intracellular markers OCT4, SOX2 andFOXD3, as shown by immunofluorescence, and that expressedREX-1 and NUCLEOSTEMIN as confirmed by RT-PCR,were subjected to suspension culture in hanging drops inabsence of LIF and buffalo foetal fibroblast feeder layersupport, they differentiated to form three-dimensional embry-oid bodies (EBs). Of 231 EBs examined on Day 3 ofsuspension culture, 141 (61.3 ± 3.09%) were of compact type,whereas 90 (38.4 ± 3.12%) were of cystic type. The cellsobtained from EBs were found to express NF-68 and NESTIN(ectodermal lineage), BMP-4 and a-skeletal actin (mesodermallineage), and a-fetoprotein, GATA-4 and HNF-4 (endodermallineage). When these EBs were cultured on gelatin-coateddishes, they spontaneously differentiated to several cell typessuch as epithelial- and neuron-like cells. When EBs werecultured in the presence of 1 or 2% DMSO or 10)8
M or10)7
M retinoic acid for 25 days, ES cells could be directed toform muscle cell–like cells, the identity of which was confirmedby expression of a-actinin by immunofluorescence and ofMYF-5, MYOD and MYOGENIN genes by RT-PCR. MYODwas first detected on Day 10 in both treatment groups and onDay 15 in controls, whereasMYOGENIN was first detected onDay 10, Day 15 and Day 25 in the presence of retinoic acid, inthe presence of DMSO and in controls, respectively. Thepresent study demonstrates the ability of buffalo ES cell–likecells to undergo directed differentiation to cells of skeletalmyogenic lineage.
Introduction
Embryonic stem (ES) cells are a special kind of cellsderived from the inner cell mass (ICM) of blastocyststage embryos. These cells possess unique characteristicsof being capable of undergoing unlimited number ofsymmetrical mitotic cell divisions without differentia-tion, maintaining a stable diploid karyotype, and beingcapable of either entering a path leading to a fullydifferentiated cell or remaining as stem cells. Followingfirst reports on establishment of ES cells from preim-plantation murine embryos (Evans and Kaufman 1981;Martin 1981), these have been established in a numberof mammalian species including many farm animalspecies such as cattle, sheep goat, pig and horse (Forreviews, see Munoz et al. 2009; Nowak-Imialek et al.2011).
Unlimited self-renewal, ability to differentiate to allthe cell types found in embryonic and adult organism,refractoriness to senescence and high versatility togenetic manipulation make ES cells very useful as basicresearch tools for understanding processes such asdifferentiation, organogenesis and early mammalian
development besides being useful in regenerative ther-apy in humans. However, no proven ES cell linescolonizing the germ line have been established forvertebrate species other than mouse and chicken thusfar.
One of the major limitations for utilizing the thera-peutic potential of human ES cells in muscular dystro-phy has been the very little success at directing these cellsinto skeletal muscle lineage. In the mouse model, aprimary reason for this is the paucity of paraxialmesoderm formation during embryoid body (EB)in vitro differentiation (Darabi et al. 2008). There is aneed to develop alternate models that could supplementthe human and mouse models for generating informa-tion required for developing sufficient populations ofskeletal myocytes for potential cell-based therapies.Buffalo is a suitable candidate for supplementing themouse model as it does not suffer from some of thelimitations of mouse model such as short lifespan,physiology and anatomy very different from humans.We have earlier reported derivation and characteriza-tion of ES cell–like cells from in vitro produced buffaloembryos (Verma et al. 2007; Anand et al. 2009) and thedevelopment of a culture system suitable for maintain-ing buffalo ES cells over long periods of time (Sharmaet al. 2011). No information is available on the ability ofbuffalo ES cell–like cells to be directed to differentiate toany particular cell type. The aim of the present studywas to study the ability of these cells to undergo directeddifferentiation to skeletal myogenic lineage.
Materials and Methods
Reagents and media
All the chemicals and media were purchased from SigmaChemical Co., St. Louis, MO, USA, unless otherwiseindicated. Knock-out DMEM and Knock-out serumreplacer were purchased from Gibco, NY, USA. Foetalbovine serum (FBS; Hyclone, Logan, UT, US, cat no.CH30160.02) of same batch was used for in vitro embryoproduction and for feeder layer cell culture. Theplasticware were purchased either from Becton, Dickin-son and Co., Lincoln Park, NJ, USA, or from Nunc,Rosklide, Denmark. Syringes of assorted sizes werepurchased from Henke Saas Wolf GmBH, Tuttingen,Germany, whereas the 0.22-lm filters were fromMillipore Corp., Bedford, MA, USA. The primaryantibodies used for the detection of pluripotency-relatedmarkers by immunofluorescence staining and thesecondary antibody goat anti-rabbit IgG-FITC conju-gate were purchased from Chemicon International,
Reprod Dom Anim doi: 10.1111/j.1439-0531.2012.02146.x
ISSN 0936-6768
� 2012 Blackwell Verlag GmbH

Temecula, CA, USA, whereas goat anti-rat IgM-FITCconjugate was purchased from Pierce BiotechnologyInc., Rockford, IL, USA.
Production and maintenance of ES cell–like cells
Buffalo foetal fibroblast (BFF) feeder layers wereprepared as described earlier (Verma et al. 2007).Briefly, skin biopsies taken from buffalo foetuses(approximately 60 days old) were washed several timeswith Dulbecco’s phosphate buffered saline (DPBS), andthe tissue pieces were transferred to 60-mm cell culturedishes and cultured in Dulbecco’s modified Eagle’smedium (DMEM) supplemented with 20% FBS and50 lg ⁄ml gentamicin sulphate in a CO2 incubator (5%CO2 in air) at 37�C. The explants were removed afterproliferation and establishment of fibroblasts. Mono-layer fibroblasts were allowed to grow till confluence.Subsequent cell layers used as feeder layers wereobtained by disaggregating the cells with 0.25% trypsin–EDTA solution. For the preparation of feeder layer,buffalo embryonic fibroblasts were inactivated by treat-ment with 10 lg ⁄ml mitomycin-C for 3 h, after whichthe monolayer was trypsinized and the cells were washedseveral times with DMEM + 10% FBS + 50 lg ⁄mlgentamicin sulphate. The feeder layer was prepared byseeding these cells in 4-well plates or in tissue culturedishes 1 day prior to culture of ES cell–like cells.
In vitro production of blastocysts and derivation ofbuffalo ES cell–like cells was carried out as describedearlier (Sharma et al. 2011) with some modifications.Briefly, ICM cells isolated mechanically from in vitroproduced day 8 hatched blastocysts using Micro-bladesTM (MicroBlades, MTB-05; MicromanipulatorMicroscope Company, Inc. Washington DC, USA)under a zoom stereomicroscope were cultured on BFFfeeder layers in ES cell culture medium consisting ofKnockout-DMEM + 15% Knockout Serum Repla-cer + 2 mM L-glutamine + 50 lg ⁄ml gentamicin sul-phate + 1% MEM non-essential amino acids +0.1 mM b-mercaptoethanol + 4 ng ⁄ml bFGF +1000 I.U ⁄ml mLIF. For subculture, ES cell colonieswere split mechanically with the help of MicrobladesTM
every 5–6 days in a split ratio of 1 : 2–1 : 4 dependingupon the colony size.
Characterization of ES cell–like cells
ES cell–like cells were regularly checked for theirpluripotency by alkaline phosphatase staining per-formed using a kit (Catalog No. 86C; Sigma ChemicalCo) according to the manufacturer’s protocol and bystudying the expression of intracellular markers such asOCT4, FOXD3 and SOX2, and surface markers such asSSEA-1, SSEA-3, SSEA-4, TRA-1-60, TRA-1-81, CD9and CD90 by immunofluorescence staining as describedearlier (Anand et al. 2009). Briefly, the ES cell colonieswere fixed in 4% paraformaldehyde in DPBS for30 min, washed 3 times with DPBS and then permea-bilized by 0.1% Triton X-100 in DPBS for 30 min. Afterthorough washing with DPBS, the colonies were incu-bated with the blocking solution (4% normal goatserum) for 30 min and then with the primary antibody
at a dilution of 1 : 10–1 : 20 for 1 h. The addition ofprimary antibody was omitted in the respective controls.After washing 3 times with DPBS, the ES cell colonieswere incubated with the appropriate FITC-labelledsecondary antibody (goat anti-rat IgM or goat anti-rabbit IgG or anti-mouse IgG or IgM, diluted 1 : 100–1 : 200) for 2 h. The colonies were then examined undera fluorescence microscope (Diaphot, Nikon, Tokyo,Japan). For examining the chromosomal integrity,colonies of ES cell–like cells were incubated with0.1 lg ⁄ml colchicine for 2–3 h at 37�C, washed twicewith Ca2 + - and Mg2 + -free DPBS, treated with0.25% trypsin-EDTA for disaggregation of cells andthen suspended in 75 mM KCl. The cells were fixedin ice-cold fixative (methanol:acetic acid 3 : 1) andspread on clean microscopic slides. The slides were airdried, and the cells were examined under a microscopeafter staining with 2% Giemsa stain. The occurrence ofa stable karyotype, as indicated by normal diploid setof chromosomes, was examined at passage-3 andpassage-6.
The expression of different genes (Table 1) wasexamined by RT-PCR, for which total RNA wasisolated from the test material using the ‘Cells-to-cDNAkit-II’ (Ambion, Austin, TX, USA) as per the manu-facturer’s instructions. Briefly, the cells were washedwith ice-cold PBS, after which 20–50 ll of cold cell lysisbuffer was added and the mixture was incubated at 75�Cfor 10 min in a thermal cycler. The genomic DNA wasdegraded by incubating the cell lysate in DNase-I at37�C for 30 min, and the activity of DNase-I wasinactivated by heating at 75�C for 5 min. cDNA wasprepared by taking 10 ll of the cell lysates using randomprimers (Operon, Nattermannallee 1, Cologne, Ger-many). The PCR cycle included denaturation (94�C for2 min) followed by repeated cycles of denaturation(94�C for 30 s), annealing (30 s at the temperaturesgiven in Table 1) and extension (72�C for 45 s) followedby a final extension (72�C for 10 min). A negative RTreaction (RT reaction without MMLV enzyme) and anegative PCR reaction without template were also setup. 18S rRNA was amplified at each stage as the house-keeping gene. Feeder cells and ES cell–like cells wereused as negative controls for ES cell–like cells anddifferentiated cells, respectively.
Spontaneous and induced differentiation of ES cell–likecells
ES cell–like cells were subjected to spontaneous differ-entiation in suspension culture in the absence of mLIFand without feeder layer support. Briefly, passage-3 andpassage-6 ES cell–like cell colonies were disaggregatedinto small clumps and were cultured in hanging drops ofES cell culture medium without mLIF. The medium waschanged every 48 h by tilting the plate and allowing thecells to settle. The EBs formed were seeded in 4-wellgelatin-coated (0.1%) tissue culture dishes for sponta-neous differentiation. The EBs differentiated to severaltypes of cells, depending on the duration of culture.These were characterized by the lineage-specific geneexpression by RT-PCR (Table 1) on days 5, 10, 15 and20 of culture.
2 MK Singh, KP Singh, D Kumar, RA Shah, T Anand, MS Chauhan, RS Manik, SK Singla and P Palta
� 2012 Blackwell Verlag GmbH

For directed differentiation, the EBs produced by thehanging drop method were placed in suspension for 3additional days and were then transferred to tissueculture dishes or 4-well plates in medium containingeither 1 or 2% dimethylsulfoxide (DMSO) or 10)8
M or10)7
M retinoic acid for directing the differentiation tomuscle cell–like cells. The differentiated cells formedwere characterized by skeletal muscle cell–specific geneexpressions by RT-PCR (Table 1) on days 5, 10, 15, 20and 25 of culture.
Results
Expression of pluripotency-related markers
The ES cell–like cells produced in the present studyexpressed surface markers SSEA-4, TRA-1-60, TRA-1-81, CD9 and CD90, and intracellular markers OCT4,SOX2 and FOXD3 (Fig. 1a–h) as shown by immuno-fluorescence. They were also found to express REX-1and NUCLEOSTEMIN (Fig. 1i–k) by RT-PCR. Thesecells also showed a high expression of alkaline phos-phatase (Fig. 1l) and had a normal diploid set ofchromosomes at passage-3 and passage-6 (Fig. 1m).
Formation and characterization of embryoid bodies
When ES cell–like cells were subjected to suspensionculture in the absence of mLIF and buffalo foetalfibroblast feeder layer, they differentiated to form three-
dimensional aggregates called EBs. When the EBs(n = 231) were harvested from the hanging drops onDay 3 of suspension culture, 141 (61.3 ± 3.09%) ofthem were in the form of compact mass (Fig. 2a),whereas 90 (38.4 ± 3.12%) were of cystic type (Fig. 2b). When EBs were cultured on gelatin-coated dishes,they spontaneously differentiated to several types of cellssuch as neuron- and epithelial-like cells (Fig. 2c–d).
The EBs were harvested on Days 5, 10, 15 and 20 ofculture, and the cell types formed were characterized byexamining lineage-specific gene expression by RT-PCR.The cells were found to express NESTIN and NF-68 ofectodermal lineage, bone morphogenetic protein-4(BMP-4) and a-skeletal actin (ASA) of mesodermallineage and alpha fetoprotein (AFP), GATA-4 andhepatocyte nuclear factor-4 (HNF-4) of endodermallineage (Fig. 3).
Induced differentiation of ES cell–like cells to muscle cell–like cells
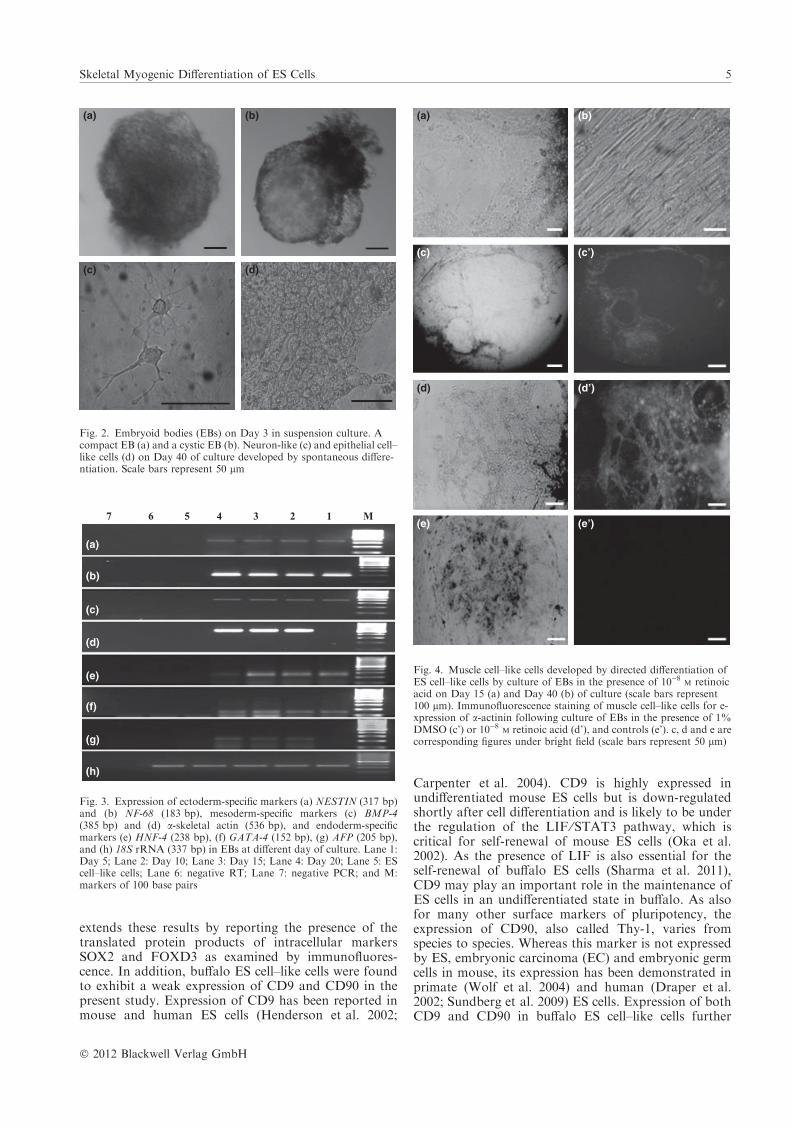
The outgrowths at the periphery of the EBs increased insize and assumed muscle cell–like morphology followingculture in the presence of retinoic acid or DMSO(Fig 4a, b). The muscle cell–like cells were found toshow an expression for a-actinin by immunofluorescencestaining (Fig. 4c–e). When the cells were examined forthe expression of skeletal muscle cell–specific genes(Figs 5 and 6), the earliest expressed gene was MYF-5.
Table 1. PCR primers used for the confirmation of lineage of differentiated cells
Gene Primer
Annealing
temperature ⁄ Cyc Product size (bp) Accession no
Pluripotency-related markers
REX-1 5¢-CAGCTGCCATAGGCTCTACC-3¢ 55 ⁄ 35 239 XM_001255545
5¢-GTCTGGCCTGACAAAGGTGT-3¢NUCLEOSTEMIN 5¢-AGAATCGGATGAGCTGTTGG-3¢ 60 ⁄ 35 400 NM_001034307.1
5¢-ATCAGCATTGCCAGAGAGGT-3¢Lineage-specific markers
Ectoderm
NF-68 5¢-AGGAAGATGCTGAGGAAGCA-3¢ 60 ⁄ 35 183 NM174121
5¢- GTTGACCTGATTTCGGGAGA-3¢NESTIN 5¢-AGTGTGAAGGCAAAGATAGC -3¢ 52 ⁄ 35 317 Sritanaudomchai
et al. (2007)5¢-TCTGTCAGGATTGGGATGGG -3¢Mesoderm
ASA 5¢-GGCATCATCACCAACTGGGA-3¢ 55 ⁄ 35 536 Sritanaudomchai
et al. (2007)5¢-TTGCCGATGGTGATGACCTG-3¢BMP-4 5¢- GGTAACCGAATGCTGATGGT-3¢ 55 ⁄ 35 385 NM001045877.1
5¢-CTGGGATGTTCTCCAGATGT -3¢Endoderm
AFP 5¢-TCTTCCCCATGTTCTTCAGG-3¢ 55 ⁄ 35 205 NM_001034262
5¢-TTTCACGGCAAATTTCTTCC-3¢HNF-4 5¢-ACATCCCAGCCTTCTGTGAG-3¢ 58 ⁄ 35 238 AF250028
5¢-TCGTCAATCTGCAGCTCTTG-3¢GATA-4 5¢-CGGAAAGAGGGGATTCAAAC-3¢ 58 ⁄ 35 152 DQ487031
5¢-ATCTCCTCACTGCTGCTGGT-3¢Skeletal muscle cell–specific markers
MYF-5 5¢-AGACGCCTGAAGAAGGTCAA-3¢ 52 ⁄ 35 220 EF128444
5¢-TGCCATCAGAGCAACTTGAG-3¢MYOD 5¢-CGACGGCATGATGAACTACA -3¢ 52 ⁄ 35 159 AB257293
5¢- GAGATGCGCTCCACGATG-3¢MYOGENIN 5¢-CCAGACACTCTGCCTCCTTC-3¢ 52 ⁄ 35 187 EF636460
5¢-CCCACCCCCTATTTCAGAAT-3¢18S rRNA 5¢-GAGAAACGGCTACCACATCC-3¢ 60 ⁄ 35 337 AF176811
5¢- GGACACTCAGCTAAGAGCATCG-3¢
Skeletal Myogenic Differentiation of ES Cells 3
� 2012 Blackwell Verlag GmbH

However, its expression was detected even in the absenceof DMSO and retinoic acid, and it continued up to thelast day of experiment, that is, Day 25 of culture. Theexpression of MYOD was first detected on Day 10 inboth the treatment groups and on Day 15 in controls.The expression of MYOGENIN was first detected onDay 10 in the presence of retinoic acid and on Day 15 inthe presence of DMSO, whereas in the controls, it wasdetected on Day 25.
Discussion
To our knowledge, this is the first report on the directeddifferentiation of buffalo ES cell–like cells. We havepreviously reported the expression of surface markersSSEA-4, TRA-1-60 and TRA-1-81 by immunofluores-cence (Anand et al. 2009) and of the presence of mRNAfor intracellular markers OCT4, SOX2, FOXD3, REX-1and NUCLEOSTEMIN by RT-PCR (Sharma et al.2011) in buffalo ES cell–like cells. The present study
(a) (a’)
(b) (b’)
(e)
(f)
(e’)
(f’)
(c) (c’) (g) (g’)
(d)
(i)
(j)
(k)
(d’) (h) (h’)
(l) (m)
Fig. 1. Colonies of ES cell–like cells showing expression of SSEA-4 (a’), TRA-1-60 (b’), TRA-1-81 (c’), CD9 (d’), CD90 (e’), OCT4 (f’) SOX2 (g’)and FOXD3 (h’) examined by immunofluorescence. a, b, c, d, e, f, g and h are their respective photographs under bright light. They also showedalkaline phosphatase activity (l) and normal chromosome number (m). Scale bars represent 100 lm. Expression of REX-1 (i), NUCLEOSTEMIN(j) and 18S rRNA (k) examined by RT-PCR in ES cell–like cell colonies. Lane 1: ES cells at passage-2; Lane 2: ES cells at passage-5; Lane 3: EScells at passage-8; Lane 4: buffalo foetal fibroblast feeder layer cells; Lane 5: negative RT; Lane 6: negative PCR; and M: markers of 100 basepairs
4 MK Singh, KP Singh, D Kumar, RA Shah, T Anand, MS Chauhan, RS Manik, SK Singla and P Palta
� 2012 Blackwell Verlag GmbH
extends these results by reporting the presence of thetranslated protein products of intracellular markersSOX2 and FOXD3 as examined by immunofluores-cence. In addition, buffalo ES cell–like cells were foundto exhibit a weak expression of CD9 and CD90 in thepresent study. Expression of CD9 has been reported inmouse and human ES cells (Henderson et al. 2002;
Carpenter et al. 2004). CD9 is highly expressed inundifferentiated mouse ES cells but is down-regulatedshortly after cell differentiation and is likely to be underthe regulation of the LIF ⁄STAT3 pathway, which iscritical for self-renewal of mouse ES cells (Oka et al.2002). As the presence of LIF is also essential for theself-renewal of buffalo ES cells (Sharma et al. 2011),CD9 may play an important role in the maintenance ofES cells in an undifferentiated state in buffalo. As alsofor many other surface markers of pluripotency, theexpression of CD90, also called Thy-1, varies fromspecies to species. Whereas this marker is not expressedby ES, embryonic carcinoma (EC) and embryonic germcells in mouse, its expression has been demonstrated inprimate (Wolf et al. 2004) and human (Draper et al.2002; Sundberg et al. 2009) ES cells. Expression of bothCD9 and CD90 in buffalo ES cell–like cells further
(a) (b)
(c) (d)
Fig. 2. Embryoid bodies (EBs) on Day 3 in suspension culture. Acompact EB (a) and a cystic EB (b). Neuron-like (c) and epithelial cell–like cells (d) on Day 40 of culture developed by spontaneous differe-ntiation. Scale bars represent 50 lm
7 6 5 4 3 2 1 M
(a)
(b)
(c)
(d)
(e)
(f)
(g)
(h)
Fig. 3. Expression of ectoderm-specific markers (a) NESTIN (317 bp)and (b) NF-68 (183 bp), mesoderm-specific markers (c) BMP-4(385 bp) and (d) a-skeletal actin (536 bp), and endoderm-specificmarkers (e) HNF-4 (238 bp), (f) GATA-4 (152 bp), (g) AFP (205 bp),and (h) 18S rRNA (337 bp) in EBs at different day of culture. Lane 1:Day 5; Lane 2: Day 10; Lane 3: Day 15; Lane 4: Day 20; Lane 5: EScell–like cells; Lane 6: negative RT; Lane 7: negative PCR; and M:markers of 100 base pairs
(a) (b)
(c) (c’)
(d) (d’)
(e) (e’)
Fig. 4. Muscle cell–like cells developed by directed differentiation ofES cell–like cells by culture of EBs in the presence of 10)8
M retinoicacid on Day 15 (a) and Day 40 (b) of culture (scale bars represent100 lm). Immunofluorescence staining of muscle cell–like cells for e-xpression of a-actinin following culture of EBs in the presence of 1%DMSO (c’) or 10)8
M retinoic acid (d’), and controls (e’). c, d and e arecorresponding figures under bright field (scale bars represent 50 lm)
Skeletal Myogenic Differentiation of ES Cells 5
� 2012 Blackwell Verlag GmbH

highlights the multiplicity and the species differences inthe surface marker phenotype of ES cells.
The ability of ES cells to form EBs is considered areliable test of their pluripotency as EBs contains cellsfrom all the three germ layers. Buffalo ES cell–like cellsproduced in the present study were able to form EBsthat expressed genes of lineages of ectoderm (NESTINand NF-68), mesoderm (BMP-4 and ASA) and endo-derm (AFP, GATA-4 and HNF-4), confirming theirpluripotent nature. On Day 3 of suspension culture inhanging drops, 61.3% of them were of compact type,whereas the rest 38.4% had multiple cysts. The size andnumber of cysts in cystic EBs increased with the increasein culture period. In mouse, visceral endoderm formsfrom the ICM, thereby giving rise to simple EBs within2–4 days of culture of EBs (Leahy et al. 1999). Onapproximately day 4, differentiation of columnar epi-thelium with a basal lamina and formation of a centralcavity occurs, at which stage, the EBs are called cysticEBs and bear similarity to the egg-cylinder stage of
mouse embryos (Ng et al. 2005). When the EBs werecultured on gelatin-coated dishes, ES cell–like cellsdifferentiated to several types of cells such as epithelial-and neuron-like cells, as also reported by us earlier(Verma et al. 2007; Anand et al. 2009), confirming theirability to undergo spontaneous differentiation.
Both retinoic acid and DMSO were able to direct thedifferentiation of buffalo ES cell–like cells to musclecell–like cells following culture of EBs on gelatin-coateddishes. The skeletal myogenic lineage of these cells, asindicated by their morphological resemblance to skeletalmuscle cells, was confirmed by examining a-actininexpression through immunofluorescence and by demon-strating the mRNA expression of early, intermediateand late myogenic regulatory factors (MRFs) MYF-5,MYOD and MYOGENIN, respectively. The earliestexpressed gene was MYF-5, the expression of which wasdetected as early as Day 5 of culture. The expression ofMYOD was first detected on Day 10 in both thetreatment groups and on Day 15 in controls, whereas
(a)
(b)
(c)
(d)
(e)
(g)
(f)
(h)
Fig. 5. Expression pattern of skeletal muscle cell–specific genes inmuscle cell–like cells produced following culture in the presence of 1%DMSO; (a) MYF-5 (220 bp); (c) MYOD (159 bp); (e) MYOGENIN(187 bp); and (g) 18S rRNA at different days of culture, with b, d, fand h being their respective controls. Lane 1: Day 5; Lane 2: Day 7;Lane 3: Day 10; Lane 4: Day 15; Lane 5: Day 25; Lane 6: ES cell–likecells; Lane 7: buffalo foetal fibroblast cells; Lane 8: negative RT; Lane9: negative PCR; and M: markers of 100 base pairs
(a)
(b)
(c)
(d)
(e)
(g)
(f)
(h)
Fig. 6. Expression pattern of skeletal muscle cell–specific genes inmuscle cell–like cells produced following culture in the presence of10)8
M retinoic acid; (a) MYF-5; (c) MYOD; E: MYOGENIN and (g)18S rRNA at different days of culture, with b, d, f and h being theirrespective controls. Lane 1: Day 5; Lane 2: Day 7; Lane 3: Day 10;Lane 4: Day 15; Lane 5: Day 25; Lane 6: ES cell–like cells; Lane 7:buffalo foetal fibroblast cells; Lane 8: negative RT; Lane 9: negativePCR; and M: markers of 100 base pairs
6 MK Singh, KP Singh, D Kumar, RA Shah, T Anand, MS Chauhan, RS Manik, SK Singla and P Palta
� 2012 Blackwell Verlag GmbH

that of MYOGENIN was first detected on Day 10 in thepresence of retinoic acid and on Day 15 in the presenceof DMSO; however, in the controls, it was detected onDay 25. These results show that both DMSO andretinoic acid are effective in inducing myogenic differ-entiation of buffalo ES cell–like cells, as indicated by thesequential expression of mRNA for MRFs in a mannersimilar to that observed during embryonic myogenesis(Seale et al. 2000). The MRFs have received consider-able attention as key regulatory molecules involved indifferentiation of skeletal muscle cells, which is a highlyordered process. Myf5 and MyoD initiate the transcrip-tion of muscle cell–specific genes and direct the cells intoskeletal muscle lineage, whereas Myogenin regulates thelater stages of development such as the fusion ofmyoblasts to myotubes (Weintraub et al. 1989; Tapscott2005). Among these MRFs, MyoD is perhaps the mostimportant one (Olson 1990) as its ectopic expression hasbeen found to be sufficient to force non-muscle cells tocomplete the myogenic programme (Weintraub et al.1991). However, the pathway by which MyoD convertsstem cells into skeletal muscle is still not known.
Both retinoic acid (Kennedy et al. 2009) and DMSO(Angello et al. 1997; Skerjanc 1999) have been used forinducing differentiation of EC cells to muscle cells. Inearlier studies, retinoic acid has been shown to inhibitcardiomyogenesis and enhance skeletal myogenesis(Wobus et al. 1994) although the stage at which thisoccurs and the interaction with other signalling path-ways are not known. Both retinoic acid and DMSOwere found to be capable of inducing skeletal myogen-esis in the buffalo ES cell–like cells. However, in mouse
EC cells, skeletal myogenesis has been reported to occurin the presence of DMSO, but not in its absence(Skerjanc and McBurney 1994). Earlier appearance ofthe expression of MYOGENIN following treatmentwith retinoic acid than after treatment with DMSOindicates that the former may be more effective inrecruitment of buffalo ES cell–like cells to myogeniclineage. However, the concentration of retinoic acidused is important as high concentrations (>10)7
M)have been reported to enhance neuronal differentiationbut inhibit myogenesis, whereas low concentrations(<10)7
M) have been observed to enhance myogenicconversion in ES and embryonic carcinoma cells(Hescheler et al. 1997).
In conclusion, the present study demonstrates theability of buffalo ES cell–like cells to undergo directeddifferentiation to cells of skeletal myogenic lineage.
Acknowledgement
This work was funded by Dept. of Biotechnology (Govt. of India).
Conflict of interests
None of the authors have any conflict of interest to declare.
Author contributions
MKS carried out the research work and wrote the manuscript. KPS,DK, RAS and TA carried out the research work. MSC designed thework.RSM wrote the manuscript. SKS analysed the data. PP designedthe work and wrote the manuscript.
ReferencesAnand T, Kumar D, Singh MK, Shah RA,
Chauhan MS, Manik RS, Singla SK,Palta P, 2009: Buffalo (Bubalus bubalis)embryonic stem cell-like cells and preim-plantation embryos exhibit comparableexpression of pluripotency-related anti-gens. Reprod Domest Anim 46, 50–58.
Angello JC, Stern HM, Hauschka SD, 1997:P19 embryonal carcinoma cells: a modelsystem for studying neural tube inductionof skeletal myogenesis. Dev Biol 192, 93–98.
Carpenter M, Rosier E, Fisk G, Branden-berger R, Ares X, Muira T, Lucero M,Rao MS, 2004: Properties of four humanES cell lines maintained in a feeder-freeculture system. Dev Dyn 229, 243–258.
Darabi R, Gehlbach K, Bachoo RM,Kamath S, Osawa M, Kamm KE, KybaM, Perlingeiro RC, 2008: Functionalskeletal muscle regeneration from differ-entiating embryonic stem cells. Nat Med14, 134–143.
Draper JS, Pigott C, Thomson JA, AndrewsPW, 2002: Surface antigens of humanembryonic stem cells: changes upon differ-entiation in culture. J Anat 200, 249–258.
Evans MJ, Kaufman M, 1981: Establish-ment in culture of pluripotential cellsfrom mouse embryos. Nature 292, 154–156.
Henderson JK, Draper JS, Baillie HS, FishelS, Thomson JA, Moore H, Andrews PW,2002: Preimplantation human embryos
and embryonic stem cells show compara-ble expression of stage-specific embryonicantigens. Stem Cells 20, 329–337.
Hescheler J, Fleischmann BK, Lentini S,Maltsev VA, Rohwedel J, Wobus AM,Addicks K, 1997: Embryonic stem cells: amodel to study structural and functionalproperties in cardiomyogenesis. Cardio-vasc Res 36, 149–162.
Kennedy KA, Porter T, Mehta V, Ryan SD,Price F, Peshdary V, Karamboulas C,Savage J, Drysdale TA, Li SC, BennettSA, Skerjanc IS, 2009: Retinoic acidenhances skeletal muscle progenitor for-mation and bypasses inhibition by bonemorphogenetic protein 4 but not domi-nant negative beta-catenin. BMC Biol 7,67.
Leahy A, Xiong J, Kuhnert F, Stuhlmann H,1999: Use of developmental marker genesto define temporal and spatial patterns ofdifferentiation during embryoid body for-mation. J Exp Zool 284, 67–81.
Martin GR, 1981: Isolation of a pluripotentcell line from early mouse embryos cul-tured in medium conditioned by terato-carcinoma stem cells. Proc Nat Acad SciUSA 78, 7634–7638.
Munoz M, Trigal B, Molina I, Dıez C,Caamano JN, Gomez E, 2009: Con-straints to progress in embryonic stemcells from domestic species. Stem Cell Rev5, 6–9.
Ng ES, Davis RP, Azzola L, Stanley EG,Elefanty AG, 2005: Forced aggregation of
defined numbers of human embryonicstem cells into embryoid bodies fostersrobust, reproducible hematopoietic differ-entiation. Blood 106, 1601–1603.
Nowak-Imialek M, Kues W, Carnwath JW,Niemann H, 2011: Pluripotent stem cellsand reprogrammed cells in farm animals.Microsc Microanal 17, 474–497.
Oka M, Tagoku K, Russell TL, Nakano Y,Hamazaki T, Meyer EM, Yokota T,Terada N, 2002: CD9 is associated withleukemia inhibitory factor mediatedmaintenance of embryonic stem cells.Mol Biol Cell 13, 1274–1281.
Olson EN, 1990: MyoD family: a paradigmfor development? Genes Dev 4, 1454–1461.
Seale P, Sabourin LA, Girgis-Gabardo A,Mansouri A, Gruss P, Rudnicki MA,2000: Pax7 is required for the specificationof myogenic satellite cells. Cell 102, 777–786.
Sharma R, George A, Kamble NM, SinghKP, Chauhan MS, Singla SK, Manik RS,Palta P, 2011: Optimization of cultureconditions to support long-term self-renewal of buffalo (Bubalus bubalis)embryonic stem cell-like cells. Cell Repro-gram 13, 539–549.
Skerjanc IS, 1999: Cardiac and skeletalmuscle development in P19 embryonalcarcinoma cells. Trends Cardiovasc Med9, 139–143.
Skerjanc IS, McBurney MW, 1994: The Ebox is essential for activity of the
Skeletal Myogenic Differentiation of ES Cells 7
� 2012 Blackwell Verlag GmbH

cardiac actin promoter in skeletal butnot in cardiac muscle. Dev Biol 163,
125–132.Sritanaudomchai H, Pavasuthipasit K, Kit-
iyanant Y, Kupradinum P, Mitalipova S,Kusamran T, 2007: Characterization andmultilenease differentiation of embryonicstem cells derived from a buffalo parthe-nogenetic embryo. Mol Reprod Dev 74,
1295–1302.Sundberg M, Jansson L, Ketolainen J,
Pihlajamaki H, Suuronen R, SkottmanH, Inzunza J, Hovatta O, Narkilahti S,2009: CD marker expression profilesof human embryonic stem cells andtheir neural derivatives, determinedusing flow-cytometric analysis, reveal anovel CD marker for exclusion ofpluripotent stem cells. Stem Cell Res 2,
113–124.Tapscott SJ, 2005: The circuitry of a master
switch: Myod and the regulation of skel-
etal muscle gene transcription. Develop-ment 132, 2685–2695.
Verma V, Gautam SK, Singh B, Manik RS,Palta P, Singla SK, Goswami SL,Chauhan MS, 2007: Isolation andcharacterization of embryonic stem cell-like cells from in vitro-produced buffalo(Bubalus bubalis) embryos. Mol ReprodDev 74, 520–529.
Weintraub H, Tapscott SJ, Davis RL,Thayer MJ, Adam MA, Lassar AB,Miller AD, 1989: Activation of muscle-specific genes in pigment, nerve, fat, liver,and fibroblast cell lines by forced expres-sion of MyoD. Proc Natl Acad Sci USA86, 5434–5438.
Weintraub H, Davis R, Tapscott S, TahyerM, Krause M, Benezra R, Blackwell TK,Turner D, Rupp R, Hollenberg S, ZhuangY, Lassar A, 1991: The myoD family:nodal point during specification of themuscle cell lineage. Science 251, 761–766.
Wobus AM, Rohwedel J, Maltsev V,Hescheler J, 1994: in vitro differentiationof embryonic stem cells into cardiomyo-cytes or skeletal muscle cells is specificallymodulated by retinoic acid. Roux’s ArchDev Biol 204, 36–45.
Wolf DP, Kuo HC, Francis Pau KY, LesterL, 2004: Progress with Nonhuman Pri-mate Embryonic Stem Cells. Biol Reprod71, 1766–1771.
Submitted: 3 May 2012; Accepted: 31 May
2012
Author’s address (for correspondence): PPalta, Embryo Biotechnology Laboratory,Animal Biotechnology Centre, NationalDairy Research Institute, Karnal-132001,Haryana, India. E-mail: [email protected]
8 MK Singh, KP Singh, D Kumar, RA Shah, T Anand, MS Chauhan, RS Manik, SK Singla and P Palta
� 2012 Blackwell Verlag GmbH


















