
B.Sc Micro II Microbial physiology Unit 1 Bacterial Photosynthesis
68
Microbial physiology Bacterial Photosynthesis Unit 1
-
Upload
rai-university -
Category
Science
-
view
198 -
download
6
Transcript of B.Sc Micro II Microbial physiology Unit 1 Bacterial Photosynthesis

Microbial physiologyBacterial Photosynthesis Unit 1

Introduction
• In all of the metabolic pathways just discussed, organismsobtain energy for cellular work by oxidizing organiccompounds.
• But where do organisms obtain these organic compounds?Some, including animals and many microbes, feed onmatter produced by other organisms. For example, bacteriamay catabolize compounds from dead plants and animals,or may obtain nourishment from a living host.
• Other organisms synthesize complex organic compoundsfrom simple inorganic substances. The major mechanismfor such synthesis is a process called photosynthesis. whichis used by plants and many microbes.

• Essentially, photosynthesis is the conversion oflight energy from the sun into chemicalenergy.
• The chemical energy is then used to convertCO2 from the atmosphere to more reducedcarbon compounds, primarily sugars.
• This synthesis of sugars by using carbon atomsfrom CO2 gas is also called carbon fixation.


• Photosynthesis, in bacteria, is defined as “thesynthesis of carbohydrates by the chlorophyllin the presence of sunlight, CO2 andreductants taken from air and oxygen do notevolve as by product, except in cynobacteria.

• The most important biological process onEarth is photosynthesis, the conversion of lightenergy to chemical energy.
• Organisms that carry out photosynthesis arecalled phototrophs.
1

• Most phototrophic organisms are alsoautotrophs, capable of growing with CO2 asthe sole carbon source.
• Energy from light is used in the reduction ofCO2 to organic compounds (photoautotrophy).However, some phototrophs use organiccarbon as their carbon source; this lifestyle iscalled photoheterotrophy.

• Photoautotrophy requires that two distinct sets of reactionsoperate in parallel: (1) ATP production and (2) CO2 reduction tocell material.
• For autotrophic growth, energy is supplied from ATP, andelectrons for the reduction of CO2 come from NADH (or NADPH).The latter are produced by the reduction of NAD+ (or NADP+) byelectrons originating from various electron donors.
• Some phototrophic bacteria obtain reducing power fromelectron donors in their environment, such asreduced sulfursources, for example hydrogen sulfide (H2S), or from hydrogen(H2).
• By contrast, green plants, algae, and cyanobacteria use electronsfrom water (H2O) as reducing power.

• The oxidation of H2O produces molecularoxygen (O2) as a by-product. Because O2 isproduced, photosynthesis in these organismsis called oxygenic photosynthesis.
• However, in many phototrophic bacteria H2Ois not oxidized and O2 is not produced, andthus the process is called anoxygenicphotosynthesis.

Photosynthetic Microorganisms
• All life can be divided into three domains, Archaea, Bacteria andEucarya, which originated from a common ancestor.
• Historically, the term photosynthesis has been applied to organismsthat depend on chlorophyll (or bacteriochlorophyll) for theconversion of light energy into chemical free energy.
• These include organisms in the domains Bacteria (photosyntheticbacteria) and Eucarya (algae and higher plants).
• The most primitive domain, Archaea, includes organisms known ashalobacteria, that convert light energy into chemical free energy.
• However, the mechanism by which halobacteria convert light isfundamentally different from that of higher organisms becausethere is no oxidation/reduction chemistry and halobacteria cannotuse CO2 as their carbon source. Consequently some biologists donot consider halobacteria as photosynthetic.
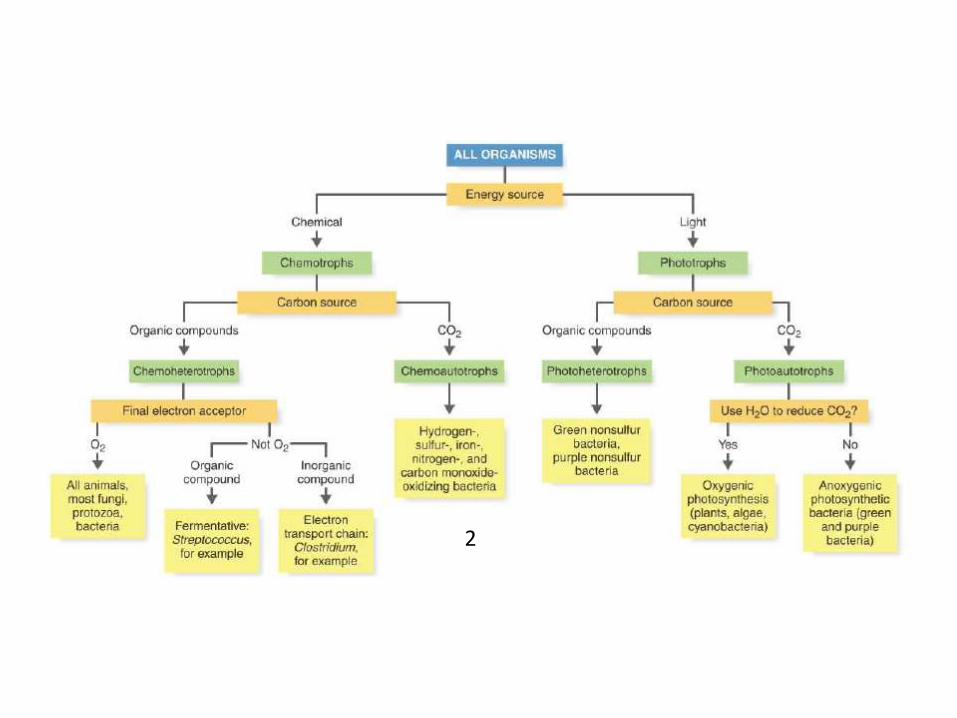
2

3

Classification of photosynthetic bacteria
• Two broad groups:
1) Anoxygenic photosynthetic bacteria
2) Oxygenic photosynthetic bacteria

Anoxygenic photosynthetic bacteria
• Some photosynthetic bacteria can use lightenergy to extract electrons from molecules otherthan water.
• These organisms are of ancient origin, presumedto have evolved before oxygenic photosyntheticorganisms.
• Anoxygenic photosynthetic organisms occur inthe domain Bacteria and have representatives infour phyla – Purple-Sulphur Bacteria, Purple non-Sulphur Bacteria, Green-Sulfur Bacteria, Greennon-Sulfur Bacteria.

• Anoxygenic photosynthesis depends on electrondonors such as reduced sulphur compounds,molecular hydrogen or organic compounds.
• They are found in fresh water, brackish water,marine and hypersaline water.
• Anoxygenic photosynthetic bacteria have beendivided into three groups on the basis ofpigmentation: purple bacteria, green bacteria andheliobacteria.

Purple Bacteria
• The anoxygenic phototrophs grow underanaerobic conditions in the presence of lightand do not use water as electron donor ashigher plants.
• They grow autotrophically with CO2 andhydrogen or reduced sulphur compounds actas electron donor.
• The pigment synthesis is repressed by O2.

• Purple bacteria contain Bchl a and b as photosyntheticpigment.
• The colour of purple bacteria shows brown, pink brown-red, purple-violet based on carotenoid contents.
• The photosynthetic pigments are innfluenced by lightintensity. At high intensity, photo-apparatus is inhibited.
• Carotenoids give rise to purple colour; mutants lackcarotenoids are blue green reflecting the actual colour ofBChl a.
• Purple Bacteria are of two types: purple-suphur bacteriaand purple non-sulphur bacteria.

Purple-sulphur bacteria
• Family: Chromatiaceae
• They are gram negative bacteria which containBChl a and b and grow chemolithotrophicallyin dark with thiosulphate as electron donor.
• They are also chemoorganotrophs, utilizeacetate, pyruvate and few other compounds.
• The mole % of G+C varies from 46-70.

• The cells of purple-sulphur bacteria are largerthan green bacteria and packed withintracellular sulfide deposition.
• They are found in anoxic zone of lakes andsulphur springs.
• They are photolithotrophs and motile innature e.g. Ectothiorhodospira, Chromatiium,Thiocapsa, Thiospirillum, Thiodictyon,Thiopedia etc.

Purple non-sulphur Bacteria
• Family: Ectothiorhodospiraceae
• They also contain BChl a and b and use lowconcentration of sulphide.
• The concn of sulphide utilized by purple sulphurbacteria proved toxic to this category of bacteria.
• Earlier, scientists thought that these bacteria areunable to use sulphide as ele donor for reductionof CO2 to cell material, thus named them non-sulphur.

• Some non-sulphur bacteria grow anaerobically in the darkusing fermentative metabolism, while others can growanaerobically in the dark by respiration in which ele donormay be an organic/inorganic compound as H2.
• This group is most versatile energetically due to broadrequirements and are photoorganotrophs i.e. use organicacids, amino acids, benzoate and ethanol.
• They also grow as chemoorganotrophs and requirevitamins.
• The DNA base composition is 61-73 mole % (G+C) and thesulphur granules are formed outside the cell.
• Eg, Rhodomicrobium, Rhodopseudomonmas,Rhodospirillum, Rhodocyclus, etc.

Green Bacteria
• Instead of green in colour, these are brown dueto the presence of carotenoids components.
• They are gram-negative.
• They contain BChl c, d and e plus small amount ofBchl a.
• The photosyntheic apparatus is chlorosomes.
• They do not require vitamins for their growth.
• Green bacteria are of two types: green sulphurbacteria and green non-sulphur bacteria.

Green Sulphur Bacteria
• Family: Chlorobiaceae
• They are non-motile, rods, spiral and cocci.
• Chlorosomes are present in the cell.
• They are strictly anaerobic and obligate phototroph.
• Deposit sulphur extracellularly.
• Mol % G+C is 45-58.
• Eg; Chlorobium, Prostheochloris, Pelodictyon, Chloroherpeton

Green non-Sulphur Bacteria
• The Green non-Sulphur Bacteria are filamentous,gliding bacteria and thermophilic in nature.
• The pigments are Bchl a, Bchl c, and carotenes.• Chlorosomes are present when grown
anaerobically.• They are photoheterotrophic and
photoautotrophic and show gliding movement.• They do not deposit sulphur.• The mol % G+C vary 53-55.• Eg, Chloroflexus

Heliobacteria
• Based on 16S rRNA sequencing and othermorphological and biochemical characters,helicobacter are quite different with otheranoxygenic photosynthetic bacteria.
• They are gram-positive, rod shaped, motileeither by gliding or by means of flagella.
• The mol % G+C is between 50-55.
• Eg, Heliobacterium, Helophilum, Heliobacillus

• Most of them produce endospores and growup to 42°C.
• The heliobacteria are green in colour.
• Most of the heliobacteria are found in tropicalsoils of paddy fields.
• They contain BChl g.

Oxygenic Photosynthetic Bacteria
• The Oxygenic Photosynthetic Bacteria areunicellular or multicellular and possessbacteriochlorophyll a and carry out oxygenicphotosynthesis.
• They are mostly represented by gram-negativecynobacteria.
• Carboxysomes and gas vesicles are present andalso show gliding movement.
• Photosynthesis is oxygenic and autotrophic.• Photosynthates get accumulated in the form of
glycogen.

Photosynthetic Pigments
• Because of presence of carotenoids in allphotosynthetic tissues their role is anticipatedin photosynthesis.
• The cells rich in carotenoids devoid ofclorophyll do not photosynthesize.
• Light energy absorbed by carotenoids appearsto be transferred to chlorophyll a or bacteriochlorophyll a and utilized in photosynthesis.

Chlorophylls andBacteriochlorophylls
• Phototrophic organisms contain some form ofchlorophyll (oxygenic phototrophs) orbacteriochlorophyll (anoxygenic hototrophs).
• Chlorophyll a is green in color because it absorbsred and blue light preferentially and transmitsgreen light.
• The absorption spectrum of cells containingchlorophyll a shows strong absorption of redlight(maximum absorption at a wavelength of 680nm) and blue light (maximum at 430 nm).

• There are a number of different chlorophylls andbacteriochlorophylls, and each is distinguished byits unique absorption spectrum.
• Chlorophyll b, for instance, absorbs maximally at660 nm rather than the 680-nm absorbancemaximum of chlorophyll a.
• All plants contain chlorophylls a and b. Someprokaryotes contain chlorophyll d, whilechlorophyll c is found only in certain eukaryoticphototrophs.

• Among prokaryotes, cyanobacteria producechlorophyll a and prochlorophytes producechlorophylls a and b.
• Anoxygenic phototrophs, such as thephototrophic purple and green bacteria,produce one or more bacteriochlorophylls.
• Bacteriochlorophyll a, present in most purplebacteria absorbs maximally between 800 and925 nm, depending on the species.

• Why do different phototrophs have different forms ofchlorophyll or bacteriochlorophyll that absorb light ofdifferent wavelengths?
• This allows phototrophs to make better use of the availableenergy in the electromagnetic spectrum.
• Only light energy that is absorbed is useful for energyconservation. By having different pigments with differentabsorption properties, different phototrophs can coexist inthe same habitat, each organism using wavelengths of lightthat others are not using.
• Thus, pigment diversity has ecological significance for the successful coexistence of different phototrophs in the same habitat.

• Prokaryotes do not contain chloroplasts. Theirphotosynthetic pigments are integrated intointernal membrane systems. These systems arise(1) from invagination of the cytoplasmicmembrane (purple bacteria); (2) from thecytoplasmic membrane itself (heliobacteria); (3)in both the cytoplasmic membrane andspecialized structures enclosed in a nonunitmembrane, called chlorosomes (green bacteria)or (4) in thylakoid membranes (cyanobacteria)

The chlorosome of green sulfur andgreen nonsulfurbacteria. (a) Transmission electronmicrograph of a cross-section ofa cell of the green sulfur bacteriumChlorobaculum tepidum. Note thechlorosomes (arrows). (b) Model ofchlorosome structure. The chlorosome(green) lies appressed to the insidesurface of the cytoplasmic membrane.Antenna bacteriochlorophyll (Bchl)molecules are arranged in tubelikearrays inside the chlorosome, andenergy is transferred from these toreaction center (RC) Bchl a in thecytoplasmic membrane (blue) througha protein called FMO. Base plate (BP)proteins function as connectorsbetween the chlorosome and thecytoplasmic membrane.4

Carotenoids and Phycobilins
• Although chlorophyll or bacteriochlorophyll isrequired for photosynthesis, phototrophicorganisms contain an assortment of accessorypigments as well.
• These include, in particular, the carotenoids andphycobilins. Carotenoids primarily play aphotoprotective role in both anoxygenic andoxygenic phototrophs, while phycobilins functionin energy metabolism as the major light-harvesting pigments in cyanobacteria.

Carotenoids
• The most widespread accessory pigments inphototrophs are the carotenoids.
• Carotenoids are hydrophobic light-sensitive pigmentsthat are firmly embedded in the photosyntheticmembrane.
• Carotenoids are typically yellow, red, brown, or greenin color and absorb light in the blue region of thespectrum.
• Carotenoids are closely associated withbacteriochlorophyll in photosynthetic complexes, andenergy absorbed by carotenoids can be transferred tothe reaction center.

• Nevertheless, carotenoids primarily functionin phototrophic organisms as photoprotectiveagents.
• Bright light can be harmful to cells;Carotenoids quench toxic oxygen species byabsorbing much of this harmful light andprevent these dangerous photooxidations.

Phycobiliproteins and Phycobilisomes
• Cyanobacteria and the chloroplasts of red algae containphycobiliproteins, which are the main light-harvestingsystems of these phototrophs.
• The red phycobiliprotein, called phycoerythrin, absorbsmost strongly at wavelengths around 550 nm, whereas theblue phycobiliprotein, phycocyanin, absorbs most stronglyat 620 nm.
• Phycobiliproteins assemble into aggregates calledphycobilisomes.
• In a fashion similar to how light-harvesting systemsfunction in anoxygenic phototrophs, phycobilisomesfacilitate energy transfer to allow cyanobacteria to grow atfairly low light intensities.

Photosynthetic Electron Transport System
• In the photosynthetic light reactions, electrons travelthrough a series of electron carriers arranged in aphotosynthetic membrane in order of their increasinglymore electropositive reduction potential (E0).
• This generates a proton motive force that drives ATPsynthesis.
• Anoxygenic photosynthesis occurs in at least five phylaof Bacteria: the proteobacteria (purple bacteria); greensulfur bacteria; green nonsulfur bacteria; the gram-positive bacteria (heliobacteria); and the acidobacteria.

Photosynthetic Reaction Centers
• The photosynthetic apparatus of purplebacteria has been best studied.
• Membrane vesicles, sometimes calledchromatophores, or membrane stacks calledlamellae are common membranemorphologies.

Membranes in anoxygenicphototrophs. (a) Chromatophores.Section through a cell of the purplebacterium Rhodobactershowing vesicular photosyntheticmembranes. The vesicles arecontinuouswith and arise by invagination of thecytoplasmic membrane. A cell isabout 1 micro m wide. (b) Lamellarmembranes in the purple bacteriumEctothiorhodospira. A cell is about 1.5micro m wide. These membranes arealso continuous with and arise frominvagination of the cytoplasmicmembrane, but instead of formingvesicles, they form membrane stacks.
5

Photosynthetic Electron Flow in Purple Bacteria
• Pohotosynthetic reaction centers are surrounded by antennapigments that function to funnel light energy to the reactioncenter.
• The energy of light is transferred from the antenna to thereaction center in packets called excitons.
• The light reactions begin when exciton energy strikes the specialpair of bacteriochlorophyll a molecules.
• The absorption of energy excites the special pair, converting itfrom a relatively weak to a very strong electron donor.
• Once this strong donor has been produced, the remaining stepsin electron flow simply conserve the energy released whenelectrons flow through a membrane from carriers of low E0 tothose of high E0, generating a proton motive force.

• Before excitation, the purple bacterial reactioncenter, which is called P870, has an E0 of about+0.5 V; after excitation, it has a potential of about-1.0 V
• The excited electron within P870 proceeds toreduce a molecule of bacteriochlorophyll a withinthe reaction center.
• This transition takes place incredibly fast, takingonly about three-trillionths (3X10-12) of a second.

• Once reduced, bacteriochlorophyll a reducesbacteriopheophytin a and the latter reducesquinone molecules within the membrane.
• These transitions are also very fast, taking lessthan one-billionth of a second.
• Relative to what has happened in the reactioncenter, further electron transport reactionsproceed rather slowly, on the order ofmicroseconds to milliseconds.

• From the quinone, electrons are transported inthe membrane through a series of iron–sulfurproteins and cytochromes, eventually returningto the reaction center.
• Key electron transport proteins includecytochrome bc1 and cytochrome c2.
• Cytochrome c2 is a periplasmic cytochrome thatfunctions as an electron shuttle between themembrane-bound bc1 complex and the reactioncenter.
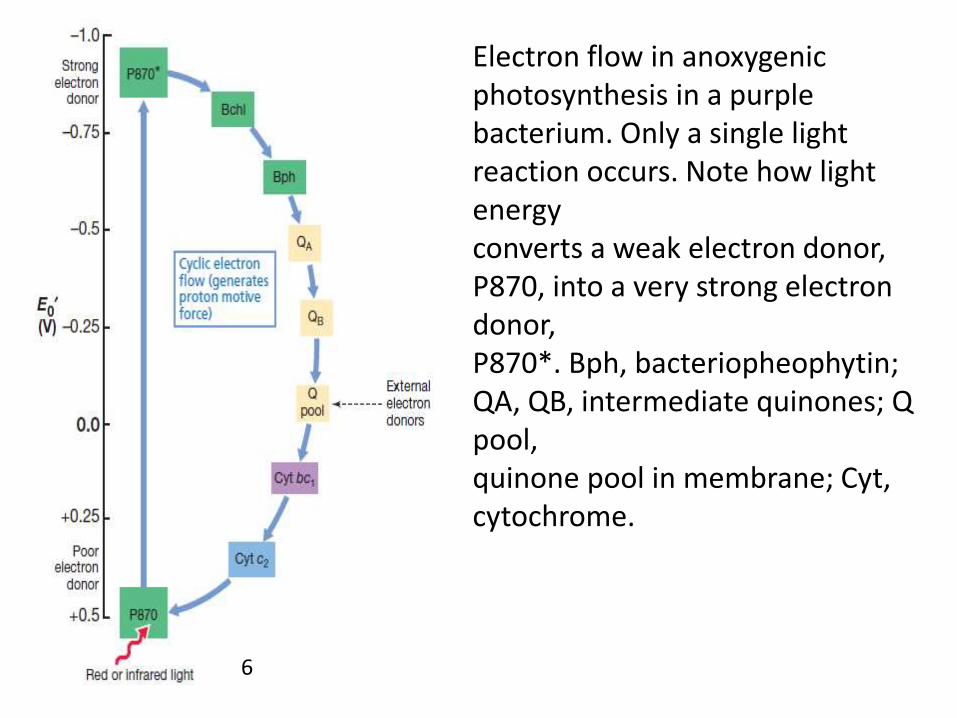
Electron flow in anoxygenicphotosynthesis in a purplebacterium. Only a single light reaction occurs. Note how light energyconverts a weak electron donor, P870, into a very strong electron donor,P870*. Bph, bacteriopheophytin; QA, QB, intermediate quinones; Q pool,quinone pool in membrane; Cyt, cytochrome.
6

Arrangement of protein complexes in the purplebacterium reaction center. The light-generated proton gradient isused in the synthesis of ATP by the ATP synthase (ATPase). LH, lightharvestingbacteriochlorophyll complexes; RC, reaction center; Bph,bacteriopheophytin; Q, quinone; FeS, iron–sulfur protein; bc1, cytochromebc1 complex; c2, cytochrome c2.
7

Photophosphorylation
• ATP is synthesized during photosynthetic electron flowfrom the activity of ATPase that couples the proton motiveforce to ATP formation.
• Electron flow is completed when cytochrome c2 donates anelectron to the special pair.
• This returns these bacteriochlorophyll molecules to theiroriginal ground-state potential (E0= +0.5 V).
• The reaction center is then capable of absorbing newenergy and repeating the process.
• This mechanism of ATP synthesis is calledphotophosphorylation, specifically cyclicphotophosphorylation, because electrons move within aclosed loop.

Autotrophy in Purple Bacteria: Electron Donors and Reverse Electron
Flow• For a purple bacterium to grow as a photoautotroph,
the formation of ATP is not enough.• Reducing power (NADH or NADPH) is also necessary so
that CO2 can be reduced to the redox level of cellmaterial.
• As previously mentioned, reducing power for purplesulfur bacteria comes from hydrogen sulfide (H2S),although sulfur (S0), thiosulfate (S2O3
2-), ferrous iron(Fe2+), nitrite (NO2-), and arsenite (AsO3
2-) can also beused by one or another species.
• When H2S is the electron donor in purple sulfurbacteria, globules of S0 are stored inside the cells.

Phototrophic purple and green sulfur bacteria. (a) Purplebacterium, Chromatium okenii. Notice the sulfur granules depositedinside the cell (arrows). (b) Green bacterium, Chlorobium limicola. Therefractile bodies are sulfur granules deposited outside the cell (arrows). Inboth cases the sulfur granules arise from the oxidation of H2S to obtainreducing power. Cells of C. okenii are about 5 m in diameter, and cellsof C. limicola are about 0.9 m in diameter. Both micrographs are brightfieldimages.
8

• Reduced substances used as photosynthetic electron donors areoxidized and electrons eventually end up in the “quinone pool”of the photosynthetic membrane.
• However, the E0 of quinone (about 0 volts) is insufficientlyelectronegative to reduce NAD+ (- 0.32 V) directly.
• Instead, electrons from the quinone pool travel backwardsagainst the thermodynamic gradient to eventually reduceNAD(P)+ to NAD(P)H.
• This energy-requiring process, called reverse electron transport,is driven by the energy of the proton motive force
• If NADPH is needed as a reductant instead of NADH, it can beproduced from NADH by enzymes called transhydrogenases.

Photosynthesis in OtherAnoxygenic Phototrophs
• Our discussion of photosynthetic electron flow hasthus far focused on the process as it occurs in purplebacteria.
• Although similar membrane reactions drivephotophosphorylation in other anoxygenicphototrophs, there are differences in certain details.
• The reaction centers of green nonsulfur bacteria andpurple bacteria are structurally quite similar; however,the reaction centers of green sulfur bacteria andheliobacteria differ significantly from those of purpleand green nonsulfur bacteria.

A comparison of electron flow in purple bacteria, green sulfur bacteria, and heliobacteria.Note that reverse electron flow inpurple bacteria is necessary to produce NADH because the primary acceptor (quinone, Q) ismore positive in potential than the NAD+/NADH couple. In green and heliobacteria, ferredoxin(Fd), whose E0 is more negative than that of NADH, is produced by light-driven reactions forreducing power needs. Bchl, Bacteriochlorophyll; BPh, bacteriopheophytin. P870 and P840 arereaction centers of purple and green bacteria, respectively, and consist of Bchl a. The reactioncenter of heliobacteria (P798) contains Bchl g, and the reaction center of Chloroflexus is of thepurple bacterial type. Note that forms of chlorophyll a are in the reaction centers of greenbacteria and heliobacteria.
9

• In green bacteria and heliobacteria the excited state of thereaction center bacteriochlorophylls resides at a significantlymore electronegative E0 than in purple bacteria and that actualchlorophyll a (green bacteria) or a structurally modified form ofchlorophyll a, called hydroxychlorophyll a (heliobacteria), ispresent in the reaction center.
• Thus, unlike in purple bacteria, where the first stable acceptormolecule (quinone) has an E0 of about 0 V, the acceptors ingreen bacteria and heliobacteria iron–sulfur (FeS) proteins havea much more electronegative E0 than does NADH.
• In green bacteria a protein called ferredoxin (reduced by theFeS protein) is the direct electron donor for CO2 fixation.
• This has a major effect on reducing power synthesis in theseorganisms, as reverse electron flow, necessary in purplebacteria, is not required in green sulfur bacteria or heliobacteria.

Oxygenic Photosynthesis
• In contrast to electron flow in anoxygenic phototrophs,electron flow in oxygenic phototrophs proceeds throughtwo distinct but interconnected series of light reactions.
• The two light systems are called photosystem I andphotosystem II, each photosystem having a spectrallydistinct form of reaction center chlorophyll a.
• Photosystem I (PSI) chlorophyll, called P700, absorbs lightat long wavelengths (far red light), whereas PSII chlorophyll,called P680, absorbs light at shorter wavelengths (near redlight).
• Oxygenic phototrophs use light to generate both ATP andNADPH, the electrons for the latter arising from thesplitting of water into oxygen and electrons.

Electron Flow in Oxygenic Photosynthesis
• The path of electron flow in oxygenicphototrophs resembles the letter Z turned on itsside, and Figure outlines this so-called “Zscheme” of photosynthesis.
• The reduction potential of the P680 chlorophyll amolecule in PSII is very electropositive, evenmore positive than that of the O2/H2O couple.
• This facilitates the first step in oxygenic electronflow, the splitting of water into oxygen andelectrons.

• Light energy converts P680 into a strong reductantwhich reduces pheophytin a (chlorophyll a minus itsmagnesium atom), a molecule with an E0 of about -0.5V.
• An electron from water is then donated to the oxidizedP680 molecule to return it to its ground-state reductionpotential.
• From the pheophytin the electron travels throughseveral membrane carriers of increasingly morepositive E0 that include quinones, cytochromes, and acopper-containing protein called plastocyanin; thelatter donates the electron to PSI.

• The electron is accepted by the reactioncenter chlorophyll of PSI, P700, which haspreviously absorbed light energy and donatedan electron that will eventually lead to thereduction of NADP+.
• Electrons transferred through severalintermediates terminating with the reductionof NADP+ to NADPH.

ATP Synthesis in Oxygenic Photosynthesis
• Besides the net synthesis of reducing power (thatis, NADPH), other important events take placewhile electrons flow in the photosyntheticmembrane from PSII to PSI. Electron transportgenerates a proton motive force from which ATPcan be produced by ATPase. This mechanism forATP synthesis is called noncyclicphotophosphorylation because electrons do notcycle back to reduce the oxidized P680, butinstead are used in the reduction of NADP+.
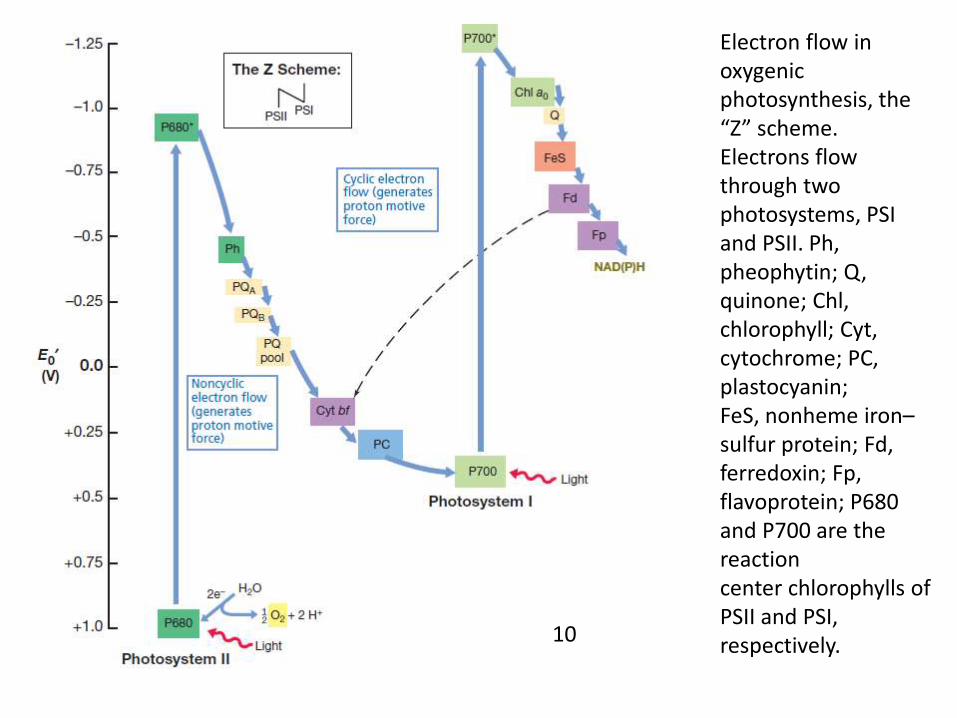
Electron flow in oxygenic photosynthesis, the “Z” scheme. Electrons flow through twophotosystems, PSI and PSII. Ph, pheophytin; Q, quinone; Chl, chlorophyll; Cyt, cytochrome; PC, plastocyanin;FeS, nonheme iron–sulfur protein; Fd, ferredoxin; Fp, flavoprotein; P680 and P700 are the reactioncenter chlorophylls of PSII and PSI, respectively.10

The Calvin-Benson cycle• The light-independent reactions of the Calvin cycle can
be organized into three basic stages: fixation, reduction,and regeneration.
• The Calvin cycle has three stages. In stage 1, the enzyme RuBisCO incorporates carbon dioxide into an organic molecule, 3-PGA. In stage 2, the organic molecule is reduced using electrons supplied by NADPH. In stage 3, RuBP, the molecule that starts the cycle, is regenerated so that the cycle can continue. Only one carbon dioxide molecule is incorporated at a time, so the cycle must be completed three times to produce a single three-carbon GA3P molecule, and six times to produce a six-carbon glucose molecule.

11

• Stage 1: Fixation• in addition to CO2,two other components are present to initiate the
light-independent reactions: an enzyme called ribulosebisphosphate carboxylase (RuBisCO) and three molecules ofribulose bisphosphate (RuBP). RuBP has five atoms of carbon,flanked by two phosphates. RuBisCO catalyzes a reaction betweenCO2 and RuBP. For each CO2 molecule that reacts with one RuBP,two molecules of 3-phosphoglyceric acid (3-PGA) form. 3-PGA hasthree carbons and one phosphate. Each turn of the cycle involvesonly one RuBP and one carbon dioxide and forms two molecules of3-PGA. The number of carbon atoms remains the same, as theatoms move to form new bonds during the reactions (3 atoms from3CO2 + 15 atoms from 3RuBP = 18 atoms in 3 molecule of 3-PGA).This process is called carbon fixation because CO2 is "fixed" from aninorganic form into organic molecules.

• Stage 2: Reduction• ATP and NADPH are used to convert the six molecules
of 3-PGA into six molecules of a chemical calledglyceraldehyde 3-phosphate (G3P). This is a reductionreaction because it involves the gain of electrons by 3-PGA. Recall that a reduction is the gain of an electronby an atom or molecule. Six molecules of both ATP andNADPH are used. For ATP, energy is released with theloss of the terminal phosphate atom, converting it toADP; for NADPH, both energy and a hydrogen atom arelost, converting it into NADP+. Both of these moleculesreturn to the nearby light-dependent reactions to bereused and reenergized.

• Stage 3: Regeneration• At this point, only one of the G3P molecules leaves the
Calvin cycle and is sent to the cytoplasm to contributeto the formation of other compounds needed by theplant. Because the G3P exported from the chloroplasthas three carbon atoms, it takes three "turns" of theCalvin cycle to fix enough net carbon to export oneG3P. But each turn makes two G3Ps, thus three turnsmake six G3Ps. One is exported while the remainingfive G3P molecules remain in the cycle and are used toregenerate RuBP, which enables the system to preparefor more CO2 to be fixed. Three more molecules of ATPare used in these regeneration reactions.

Conclusion
• The Calvin cycle refers to the light-independent reactions inphotosynthesis that take place in three key steps.
• Although the Calvin Cycle is not directly dependent on light, it is indirectlydependent on light since the necessary energy carriers (ATP and NADH)are products of light-dependent reactions.
• In fixation, the first stage of the Calvin cycle, light-independent reactionsare initiated; CO2 is fixed from an inorganic to an organic molecule.
• In the second stage, ATP and NADPH are used to reduce 3-PGA into G3P;then ATP and NADH are converted to ADP and NADP+, respectively.
• In the last stage of the Calvin Cycle, RuBP is regenerated, which enablesthe system to prepare for more CO2to be fixed.
• For each molecule of carbon dioxide that is fixed, two molecules of NADPHand three molecules of ATP from the light reactions are required. Theoverall reaction can be represented as follows:

12

References
• Reading
• Brock biology ofmicroorgamism (13th
edition) by Madigan,Martinko, Stahl, Clark
• Microbiology (10th
edition) by Tortora,Funke and Case
• Microbiology (5th
edition) by Prescott
• Images
• 1-12: Brock biology of microorgamism (13th
edition) by Madigan, Martinko, Stahl, Clark


















