
Efficacy of postoperative radiotherapy combined with taxol ...
description

e Journal of Biological Sciences Volume: 1, Issue 1 (December 2009)
ISSN: 2076-9946, EISSN: 2076-9954 ©All rights reserved.
46
BIOTECHNOLOGICAL APPROACHES FOR THE PRODUCTION
OF POTENTIAL ANTICANCER LEADS PODOPHYLLOTOXIN
AND PACLITAXEL: AN OVERVIEW
Anrini Majumder, Sumita Jha*
Centre of Advanced Study in Cell and Chromosome Research, Department of Botany,
University of Calcutta, 35 Ballygunge Circular Road, Kolkata 700019, India
*E-mail: [email protected]; [email protected]
Telephone no.: 91-33-2461-5445
Fax: 91-33-2461-4849
ABSTRACT
A number of plant-derived compounds with diverse chemical structures have played significant roles in the
development of several potent anticancer drugs. Podophyllotoxin, a lignan and paclitaxel, a diterpenoid are two such
anticancer lead molecules. Compounds like etoposide, teniposide, taxotere etc. have emerged as successful
antineoplastic agents upon modification of these two natural leads. The original sources of podophyllotoxin and
paclitaxel have become scarce in occurrence due to overexploitation and their own biological characteristics and
synthetic production of these compounds is still commercially unacceptable. Biotechnological approaches for the
production of these two anticancer leads, involving plant cell and organ cultures, have been considered to be
attractive alternatives. To meet the growing demand by pharmaceutical industries these compounds have been
isolated from sources other than the original plants and different strategies have been adapted to improve their yield
in in vitro cultures. Although large scale production of paclitaxel is reported, production of podophyllotoxin on a
commercial scale is yet to be achieved. This review focuses on these two anticancer lead molecules and summarizes
the different biotechnological approaches applied to improve their productivity.
Key words: Diterpenoids, lignan, Podophyllum, Taxus
Abbreviations: Amsl: above mean sea level; B5: Gamborg‟s basal medium (1968); BA: 6-benzyladenine; cDNA:
complementary DNA; GA3: gibberellic acid; IAA: indole-3-acetic acid; IBA: indole-3-butyric acid; m.p.: melting
point; mol. wt.: molecular weight; MS: Murashige and Skoog‟s basal medium (1962); NAA: α-Napthaleneacetic
acid; NADP: nicotinamide adenine dinucleotide phosphate, NADPH: reduced nicotinamide adenine dinucleotide
phosphate 2,4-D: 2,4-Dichlorophenoxyacetic acid
1. INTRODUCTION
Cancer is one of the deadliest diseases known through ages and is projected to become the leading cause of
death worldwide in 2010 [according to a new edition of the World Cancer Report from the International Agency for
Research on Cancer (Medscape Medical News © 2008)]. At present, cancer is the second most common cause of
death in the US, exceeded only by heart disease. In the US, cancer accounts for 1 of every 4 deaths; the estimated
number of deaths in the US alone in 2008 was 565,650 (American Cancer Society, Cancer Facts & Figures 2008,
and Atlanta: American Cancer Society; 2008). Cancer cells divide at a fast pace and thus compounds inhibiting cell
division or cytotoxic compounds destroying cells directly are the two most commonly applied modes of
chemotherapy [1]. The search for natural products as potential anticancer agents dates back to the Ebers papyrus in
1550 BC, but the scientific period of this search is much more recent, beginning in 1950s with the discovery and
development of the vinca alkaloids, vinblastine and vincristine, and the isolation of the cytotoxic podophyllotoxins
[2, 3]. Some of the world‟s best known lead antineoplastic agents are still derived from plants as the complex
structures of these compounds prohibit their synthesis on an industrial scale [1]. These include vinblastine,

e Journal of Biological Sciences www.ejarr.com
47
vincristine from Catharanthus roseus, paclitaxel (Taxol®) from Taxus sp., camptothecin from Camptotheca
acuminata and podophyllotoxin from Podophyllum sp. [1]. Extensive research by pharmaceutical companies have
led to the development of several potential anticancer drugs like vinorelbine, vindesine, topotecan (Hycamtin®),
irinotecan (CPT-11, Camptostar®), docetaxel (Taxotere®), etoposide, teniposide, etc. upon modification of the
natural leads and many more are yet to come. Several other plant-derived compounds are currently in clinical and
preclinical trials.
Production of a plant-based pharmaceutical is always not upto the desired level. It may be produced only at a
specific developmental stage or under specific environmental condition, stress or nutrient availability [4]; the plants
may be very slow growing taking several years to reach a size suitable for product extraction. Indiscriminate
collection of medicinal plants from the wild for extraction of products of interest has led to the extinction of certain
number of species. Thus biotechnological approaches involving plant cell and organ cultures appear to be attractive
alternatives. Such cultures provide a continuous and reliable supply of plant natural products regardless of the state
of the plant in the wild or geographical, political, seasonal or environmental influences [5, 6].
Different strategies have been used with the aim of increasing the production of bioactive secondary metabolites
in plant cell cultures. These include screening and selection of high producing cell lines, optimization of nutrient
media for growth and production, organ culture, culture of immobilized cells etc. [4]. Another promising tool to
improve product yield in cell culture is the use of biotic and abiotic elicitors [7]; feeding of biosynthetic precursors
is yet another effective technique. A novel strategy developed is the in vitro cross-species coculture, through which
metabolites produced by one species can be taken up by another species for biochemical conversion [8, 9, and 10].
Large scale culture of plant cells has been shown to be feasible for industrial production and is an attractive
alternative to the traditional method of cultivation. It ensures controlled supply of phytochemicals and provides a
well defined production system which results in improved yields and consistent quality of the product [11]. Since
the first report on cultivation of plant cells in a large volume [12] considerable progress has been made in the field of
scale up of cell cultures and plant cells have been successfully cultivated in bioreactors of upto 75,000 litres capacity
[13]. But in spite of the progress that has been made, production on an industrial level is limited to only a few
phytochemicals- shikonin [14], ginseng saponins [15], berberine [16] and paclitaxel [17]. Efforts are being
continuously made to optimize conditions for large scale plant cell cultures.
This review focuses on two important plant-derived anticancer leads – podophyllotoxin and paclitaxel and
summarises the different biotechnological approaches that have been applied to improve their yield.
2. PODOPHYLLOTOXIN
Podophyllotoxin (Figure 1a) is a pharmaceutically active natural product belonging to the chemical group of
lignans. It is used as a precursor for the synthesis of important antitumour drugs like etoposide (VP-16-213) (Figure
1b) and teniposide (VM-26) (Figure 1d) which are used in the treatment of lung cancer, testicular cancer, a variety
of leukemias and other solid tumours [18, 19, 20]. NK611, TOP 53, GL 331 and azatoxin are some other promising
antitumour derivatives. Podophyllotoxin, endowed with potent cytotoxicity, acts by inhibiting the assembly of
microtubules. It binds to α/β-tubulin dimer giving podophyllotoxin-tubulin complex; this stops the polymerisation of
microtubules at one end but does not stop the disassembly at the other end leading to the degradation of
microtubules; cells are arrested in the metaphase stage of mitosis and thus cell growth stops [21, 22].
Podophyllotoxin itself has got severe gastrointestinal side effects and thus too toxic for therapeutic purposes. So
with an aim to develop more potent and less toxic anticancer agents several glycosides and aglycon derivatives were
prepared by the chemists in the pharmaceutical research department of Sandoz (Switzerland) and this ultimately led
to the discovery of etoposide in 1966, which received FDA (Food and Drug Administration) approval in 1983. In
order to overcome the limitations associated with poor water solubility of etoposide, etopophos or etoposide
phosphate (Figure 1c) was introduced by Bristol-Myers Squibb Co., USA which was approved by the FDA in 1996.
Teniposide is another derivative [20]. These derivatives of podophyllotoxin are topoisomerase II inhibitors [22, 23]
arresting cells in the late S or early G2 phase of the cell cycle [24, 25]. Podophyllotoxin is also the precursor of other
derivatives such as CPH82 (Reumacon®) that has entered phase III of clinical trials against rheumatoid arthritis in
Europe [26]. It has also been applied topically for the treatment of veneral warts (condyloma acuminata) caused by
papilloma virus [27] and was found to inhibit the replication of measles and herpes simplex type I viruses [28].
Because of its tubulin binding property it disrupts the cellular cytoskeleton and thereby interferes with viral
replication [29].

e Journal of Biological Sciences www.ejarr.com
48
2.1 BIOSYNTHESIS OF PODOPHYLLOTOXIN
Podophyllotoxin (C22H22O8, mol. wt. 414.41, m.p.183.3-184°C) is a member of the group of lignans which are
dimers of phenylpropanoid pathway intermediates linked by the central carbons of their side chain [30, 31]. Total
biosynthetic pathway of podophyllotoxin has not yet been elucidated, a hypothetical scheme is proposed. The
pathway, like most other plant secondary metabolite pathways, is branched at several places. The early steps of
lignan biosynthesis until matairesinol were elucidated by Lewis and Davin and their coworkers in Forsythia species
[32]. The biosynthesis of lignans originates from phenylalanine and tyrosine [33] which are converted via a series of
intermediates to coniferyl alcohol, a key precursor in the biosynthetic pathway of podophyllotoxin [34]. Two
molecules of coniferyl alcohol are stereospecifically coupled to (+)-pinoresinol, the first lignan of the pathway by a
protein system pinoresinol synthase, consisting of a radical-forming oxidase and a 78 kDa dirigent protein [35]. (+)-
pinoresinol is reduced by NADPH dependant pinoresinol/lariciresinol reductase to (–)-secoisolariciresinol via (+)-
lariciresinol [36, 37] which is oxidized to matairesinol by NADP dependant secoisolariciresinol dehydrogenase [32,
38, 39]. The first cDNAs encoding dirigent proteins and pinoresinol/lariciresinol reductase were isolated from
Forsythia intermedia [37, 40]. cDNAs encoding secoisolariciresinol dehydrogenase were isolated not only from
Forsythia intermedia but also from Podophyllum peltatum [39]. Biosynthetic steps from matairesinol to
deoxypodophyllotoxin are not yet known; the steps include the formation of methylenedioxy bridge leading to ring
A, hydroxylation and methylation in ring E and closure of ring C [21]. Yatein may be an intermediate [41, 42].
Deoxypodophyllotoxin is hydroxylated in position 4 (ring C) by deoxypodophyllotoxin 4-hydroxylase to
podophyllotoxin [43].

e Journal of Biological Sciences www.ejarr.com
49
TABLE 1. PHYSICOCHEMICAL PROPERTIES OF PODOPHYLLOTOXIN [219, 220]
IUPAC systematic name (7α,7'α,8α,8'β)-7-hydroxy-3',4',5'-trimethoxy-4,5-
methylenedioxy-2,7'-cyclolignano-9',9-lactone
Molecular formula C22H22O8
Molecular weight 414.41
Melting point 183.3-184°C
Solubility in water 120 mg l-1
at RT
Optical rotation[α]² 0
D 132,5 (C 0,2 CHCl3)
2.2 PODOPHYLLOTOXIN FROM PLANT SOURCES
Podophyllotoxin has been reported to occur in both gymnosperms and angiosperms. In gymnosperms its
presence is noted only in some of the members of the family Cupressaceae while in angiosperms few members
belonging to families Berberidaceae, Polygalaceae, Lamiaceae and Linaceae contain this compound [22] (Table 2).
TABLE 2. PLANT SOURCES OF PODOPHYLLOTOXIN [22]
Family Plant species
Gymnosperms Cupressaceae Callitris drummondii
Juniperus sabina
J. sabina var. tamariscifolia
J. virginiana
J. chinensis
J. lucayana
J. scopulorum
Angiosperms Berberidaceae Dyphylleia cymosa
D. grayi
Podophyllum hexandrum
P. peltatum
P. pleianthum
P. versipellis
Polygalaceae Polygala polygama
Lamiaceae Hyptis verticillata
Linaceae Linum album
L. capitatum
L. flavum
L. arboretum
L. campanulatum
L. elegans
L. pamphylicum
Commercially exploitable plant sources of podopyllotoxin are few and this compound is currently extracted for
drug use from the roots and rhizomes of two species of Podophyllum – P. hexandrum Royle or the Indian
Podophyllum and P. peltatum L. or the American Podophyllum of family Berberidaceae. P. peltatum as a source of
podophyllotoxin is commercially inferior to P. hexandrum [44] as the levels of podophyllotoxin in P. peltatum are
lower than P. hexandrum and purification procedures are complicated by the presence of similar levels of the related
lignans – α and β peltatin [45].
P. hexandrum is a perennial rhizomatous herb growing in the subalpine forests of the Himalayas [46, 47]. The
rhizome is the chief source of a resin known as podophyllin [42, 48] which is a mixture of several lignans with
antitumour properties, podophyllotoxin being the most active cytotoxic compound [49]. Podophyllotoxin content of
rhizomes ranges between 0.36-1.08% (on dry weight basis) [50].

e Journal of Biological Sciences www.ejarr.com
50
The demand for podophyllotoxin is ever increasing. But the occurrence of P. hexandrum is scarce due to its
long juvenile phase and poor fruit setting ability [51]. Overexploitation and lack of organised cultivation have made
the plant „critically endangered‟ [52, 53]. Owing to its immense clinical importance new routes for total synthesis of
podophyllotoxin have been discovered [54, 55]. But these are low yielding processes and not economically feasible.
Thus isolation from plant sources continues to be the only viable option [29]. A lot of effort has been put in the past
several years to improve its production from different podophyllotoxin producing plant species.
2.3 PROPAGATION OF Podophyllum SPECIES BY CONVENTIONAL AND IN VITRO TECHNIQUES
There are several problems associated with the isolation of compounds for production of pharmaceuticals from
biomass collected from wild populations of plants. Destruction of plant populations due to over exploitation or
natural calamities affects drug supply and the content of bioactive secondary metabolite in the plant. Moreover, wild
populations may be represented by various genotypes growing under different environmental conditions which may
affect drug profile leading to problems in the purity of the final product. Thus cultivation of suitable clones would
ensure a reliable supply of the material with consistent quality. Seeds of P. hexandrum remain dormant for about 10
months under natural conditions [56]. Strategies towards conventional and in vitro propagation of Podophyllum sp.
have been developed. [56] reported 44% germination upon sowing P. hexandrum seeds with fruit pulp immediately
after collection. Cultivation trials using rhizome as the planting material at higher altitudes (3600m amsl) and seeds
at a relatively lower altitude (1150m amsl) have been reported [57, 58]. Nadeem et al. [48] reported 5 fold
improvement in seed germination of P. hexandrum following pretreatment with sodium hypochlorite and also
improved vegetative multiplication using rhizome segments treated with IBA. The authors also reported in vitro
multiplication of the Indian Podophyllum via multiple shoot formation from zygotic embryos and subsequent
rooting. Plant regeneration via somatic embryogenesis has been reported [59]. Callus derived from zygotic embryos
differentiated globular somatic embryos which developed into plantlets. In our laboratory, we observed occasional
shoot organogenesis in mature root derived callus cultures maintained on MS basal medium supplemented with BA
and NAA for 12 weeks. Nodular structures developed on such calli after 12 weeks which developed into bud
primordia like structures within another 3 weeks of culture. Further development of bud primordia into adventitious
shoot buds was found to occur after 2 weeks of culture on the same medium. Shoot buds developed upto 1-2cms
with leaf differentiation when transferred onto fresh medium under 16/8 hrs (light/dark) photoperiod but did not
grow further even after trials with other combination and concentration of different cytokinins with or without auxin
[60]. In vitro propagation of P. peltatum by using rhizome tips as explants has been reported by Moraes-Cerdeira et
al. [61]. Different types of buds were induced from a terminal bud derived from rhizome tips of P. peltatum which
developed into plantlets.
2.4 PRODUCTION OF PODOPHYLLOTOXIN BY PLANT CELL AND ORGAN CULTURES
Since total chemical synthesis of podophyllotoxin is complicated and expensive biotechnological approaches
particularly plant cell and tissue cultures appear to be attractive alternatives for the production of this
pharmaceutically important lignan. Induction of callus culture from P. peltatum and detection of podophyllotoxin
from such cultures was first reported by Kadkade [62, 63]. Van Uden et al. [49] initiated podophyllotoxin producing
callus cultures from in vitro plantlets of the Indian Podophyllum; dark-grown cultures accumulated upto 0.3%
podophyllotoxin (dry weight basis). After several tedious trials we initiated and established callus cultures of the
Indian Podophyllum from in vitro grown axenic seedling explants and roots and rhizomes isolated from 1 year old
mature plants on B5 and MS media supplemented with growth regulators [64]. Podophyllotoxin could be detected
from all the callus lines that survived after 1 year of initiation, induced from different juvenile and mature explants
[64]. Callus cultures producing podophyllotoxin have also been initiated from needles of Callitris drummondii [65],
leaves of Juniperus chinensis [66].
Suspension cultures have been proposed to be a viable alternative for the production of economically important
phytochemicals. Such cultures have a relatively fast growth rate and are easy to manipulate. Suspension cultures
producing podophyllotoxin were initiated from different plant species which are tabulated below (Table 3).

e Journal of Biological Sciences www.ejarr.com
51
TABLE 3. PODOPHYLLOTOXIN PRODUCTION IN CELL SUSPENSION CULTURES OF DIFFERENT
PLANT SPECIES
Species References
Callitris drummondii [65]
Linum album [221]
L. nodiflorum [222]
L. mucronatum spp. armenum [223]
Podophyllum hexandrum [49, 60, 224]
P. peltatum [225, 226]
In a recent study, Anbazhagan et al. [67] established embryogenic cell and adventitious root culture systems in
P. peltatum and high performance liquid chromatography analysis revealed that adventitious roots contained higher
podophyllotoxin than embryogenic cell clumps.
2.5 STRATEGIES FOR IMPROVING THE IN VITRO PRODUCTION OF PODOPHYLLOTOXIN
Several studies have been carried out for improving the accumulation of podophyllotoxin in in vitro cultures by
optimizing culture conditions and nutrient levels, addition of elicitors and precursors, immobilization etc. with
limited success.
Culture conditions had a direct effect on biomass yield and in vitro accumulation of podophyllotoxin. Light
inhibited podophyllotoxin accumulation in callus cultures of P. hexandrum [49]. Cultures grown in light contained
3-4 times less podophyllotoxin than dark grown cultures. Similarly, dark conditions favoured cell growth and
podophyllotoxin accumulation in suspension cultures of P. hexandrum [68]. The accumulation of podophyllotoxin
in callus cultures of P. peltatum was also strongly affected by the quality and intensity of light [63]; total
podophyllotoxin level was promoted by red light at an intensity of 750µW/cm2. The pH of the culture medium
proved to be another critical factor; a medium pH of 6.0 was conducive for high biomass production and
podophyllotoxin accumulation in cell cultures of P. hexandrum [69]. Manipulation of the hydrodynamic stress is yet
another important parameter for enhancing secondary metabolite production in in vitro cultures. Plant cells are
sensitive to shear forces which may be due to large plant cell sizes, presence of a cell wall and vacuoles and
aggregation patterns [70]. P. hexandrum cells were also found to be sensitive to hydrodynamic stress [68]. Higher
rotational speed (>150 rpm) reduced cell viability and thereby had an inhibitory effect on podophyllotoxin
production.
Optimization of the culture medium is an important parameter to improve productivity. To optimize
podophyllotoxin production the effects of major medium components (carbon source, nitrogen and phosphate) and
different types of basal media in P. hexandrum cell suspension culture have been studied [68, 71]. It was found that
podophyllotoxin content could be correlated to the amount and type of sugar used and to the amount of nitrogen and
phosphate present in the culture medium. Glucose was a better carbon source than sucrose for both cell growth and
podophyllotoxin production, the optimum concentration being 60g l-1
. The optimum level of nitrogen was 60mM
with ammonium salts and nitrate in the ratio of 1:2 and that of phosphate was 1.25mM. Also from a wide range of
basal media tested MS medium was found to be most suitable for growth and production of podophyllotoxin [71].
Contrary to these observations, among different basal media tested, we found B5 basal medium to be most suitable
for establishing cell suspension cultures of the Indian Podophyllum [64]. Kadkade [63] showed that the growth of
callus cultures of P. peltatum as well as accumulation of podophyllotoxin in such cultures were also dependant on
the type and concentration of plant growth regulators and other complex supplements (yeast extract, casamino acid,
corn steep liquor and peptone) used. A combination of IAA and/or NAA in the presence of kinetin and casamino
acid promoted callus growth while 2,4-D yielded maximum podophyllotoxin accumulation.
Individual cells of a heterogenous cell suspension culture often differ in their ability to synthesize secondary
metabolites [72]. Cell cloning provides a promising way of screening and selecting high producing cell lines [5].
This approach has been found to be successful for the production of quite a few important plant derived compounds
like shikonin from Lithospermum erythrorhizon [14], berberine from Coptis japonica [73], 10-deacetylbaccatin III
from Taxus baccata [74], forskolin from Coleus forskohlii [75]. Cell plating is a simple technique to select cell lines
with improved growth rates and productivity. Selection of the best performing cell line, its maintenance and
stabilization are necessary prerequisites for the production of compounds in bioreactors and subsequent scale up of

e Journal of Biological Sciences www.ejarr.com
52
the cultivation process to the industrial level. Extensive screening programmes were employed in our laboratory
with an aim of obtaining fast growing cell lines of P. hexandrum with high podophyllotoxin producing capacity.
Indeed, cell lines differed distinctly in growth rates and podophyllotoxin content and selection of fast growing cell
lines and recloning of such lines led to establishment of cell lines capable of optimum growth accumulating
optimum levels of podophyllotoxin [64].
Exogenous supply of a biosynthetic precursor is another interesting alternative to increase the level of a desired
product. Since podophyllotoxin is a product of the phenylpropanoid pathway compounds from the pathway
(phenylalanine, tyrosine, cinnamic acid, caffeic acid, coumaric acid, ferulic acid and coniferin) and one related
compound 3,4-methylenedioxycinnamic acid were added to suspension cultures of P. hexandrum [76]. Of these
precursors only coniferin significantly increased the level of podophyllotoxin – a 12.8 fold increase in content was
observed. We also studied the effect of different concentrations of direct (phenylalanine, tyrosine, trans-cinnamic
acid, para-coumaric acid) and one indirect precursor (tryptophan) of podophyllotoxin on cell suspension cultures of
the Indian Podophyllum [60]. In contrast to observations made by van Uden et al. [76], maximum accumulation of
podophyllotoxin was noted after the addition of para-coumaric acid which was 4.5 fold more than that accumulated
by untreated control cultures. Significant enhancement (2 to 4 fold over control) in podophyllotoxin content was
achieved after adding tyrosine, tryptophan and phenylalanine to cell suspension cultures [60]. Addition of coniferyl
alcohol, the poorly water soluble precursor, complexed with β-cyclodextrin resulted in enhanced podophyllotoxin
accumulation in P. hexandrum cell suspension cultures [77]. Podophyllotoxin content increased 11 fold when
phenylalanine was added to callus cultures of J. chinensis induced from leaves of young trees on Schenk and
Hildebrandt medium [66].
Coniferin is a precursor of podophyllotoxin, but it is not available commercially. Reports indicate that cross
species coculture system using Linum flavum hairy roots (a natural source of coniferin) and P. hexandrum cell
suspensions was an effective strategy for improving podophyllotoxin accumulation. Coniferin generated by L.
flavum hairy roots was effectively used by P. hexandrum cells for in vitro production of podophyllotoxin [10].
Coculture of cell suspensions of P. hexandrum with Beta vulgaris hairy roots (another natural source of coniferin)
revealed that podophyllotoxin was exuded in the culture medium [60]. The study demonstrated the feasibility of
cross species coculture of B. vulgaris hairy roots and cells of the Indian Podophyllum for release of podophyllotoxin
into the culture medium, thereby recycling cells for future use.
Among the manipulative techniques applied to promote the accumulation of secondary metabolites, elicitation
results in significant increases in product yield [78, 79, 80, 81, 82]. In nature plants produce secondary metabolites
as a defense response to pathogen attack or as a wound response and the use of elicitors is usually intended to mimic
these responses in plants. Elicitors are signal molecules triggering the formation of secondary metabolites by
activating novel genes encoding enzymes in different biosynthetic pathways [5, 83, 84]. Addition of elicitors with an
aim of improving the accumulation of bioactive secondary metabolites is well documented [78, 79, 80, 85, 86, 87,
88, 89, 90, 91, 92, 93, 94, 95]. However, there are very few reports on enhancement of podophyllotoxin
accumulation following elicitation. A 15 fold increase in podophyllotoxin accumulation by callus cultures of J.
chinensis was noted after the addition of chito-oligosaccharides [66]. In contrast addition of laminaran enzyme
hydrolysates to the same culture resulted in only a 3.5 fold increase in podophyllotoxin content over the control.
Significant enhancement (10 fold) in podophyllotoxin accumulation was noted when methyl jasmonate was added to
suspension cultures of Linum album line X4SF [96]. We analysed the effect of two well known elicitors (salicylic
acid and methyl jasmonate) on cell suspension cultures of the Indian Podophyllum and a very significant increase in
podophyllotoxin accumulation was noted after the addition of salicylic acid [60].
Another approach of improving productivity by plant cell cultures is the immobilization of plant cells in which
improved cell to cell interaction may lead to improved productivity [4]. Reports also indicate that addition of
precursors to immobilized plant cells resulted in improved accumulation of secondary metabolites than freely
suspended cells under the same condition [97, 98]. However, entrapment of P. hexandrum cells in calcium alginate
as such or addition of precursors to immobilized cells did not improve podophyllotoxin accumulation [76].
Bioreactor studies represent the final step leading to commercial production of economically important
phytochemicals from plant cell cultures. Although a number of economically important compounds have been
produced by plant cell and tissue culture techniques, production of compounds on an industrial scale is still
restricted. Major constraints of industrial or commercial production of compounds of interest have been slow growth
of plant cells, susceptibility to microbial contamination, oxygen needs and susceptibility to shearing stress due to
large cell size [99]. Plant cell cultures often undergo spontaneous genetic variation in terms of secondary metabolite
accumulation resulting in a heterogenous population. For large scale production of any compound selection of a high
yielding and genetically stable cell line is desirable. Also, a multitude of other parameters like growth and
production medium, physicochemical conditions, type of reactor, inoculum density can affect scale up of plant cell

e Journal of Biological Sciences www.ejarr.com
53
cultures. Batch cultivation of P. hexandrum for production of podophyllotoxin was conducted in a 3 litre stirred tank
bioreactor using optimized medium which resulted in biomass yield of 21.4g l-1
in 24 days and podophyllotoxin
concentrations of 13.8mg l-1
in 26 days [100]. In another study, upto 0.2% podophyllotoxin (on dry weight basis)
was obtained when L. album cell suspensions were cultured in a 20 litre bioreactor [42]. Commercial production of
podophyllotoxin using plant cell cultures is yet to be achieved.
2.6 PODOPHYLLOTOXIN PRODUCTION IN TRANSGENIC CULTURES
The formation of secondary metabolites is correlated with the degree of organization of cell structures and is
often low and unstable in undifferentiated callus and suspension cultures [101]; for example, several alkaloids which
are scarcely synthesized in undifferentiated cultures are produced at higher levels in cultured roots thereby
correlating alkaloid production with root organogenesis [102]. But the major constraint associated with in vitro root
cultures is their slow growth rate. On the other hand, hairy roots transformed with Agrobacterium rhizogenes have a
fast growth rate, are genetically stable and produce secondary metabolites at levels comparable to or greater than
that of intact plants [103, 104, 105]. There is a single report on transformation of P. hexandrum by different strains
of A. rhizogenes (A4, 15834 and K599) in which transformed calli obtained from embryo were reported to contain 3
fold more podophyllotoxin compared to controls [106]. However, till date establishment of hairy roots following
transformation using A. rhizogenes has not been reported.
2.7 ALTERNATIVE SOURCES OF PODOPHYLLOTOXIN
Apart from plant sources some endophytic fungi have also been reported to produce podophyllotoxin. Eyberger
et al. [107] reported that two endophytic fungi, both strains of Phialocephala fortinii isolated from the rhizomes of
P. peltatum produced podophyllotoxin at measurable amounts in the culture broth. In another study Kour et al. [108]
reported that an endophytic fungus Fusarium oxysporum isolated from Juniperus recurva produced
podophyllotoxin. Although optimization studies to increase the production by the cultured fungal endophytes are in
progress, these organisms can be promising candidates for large scale production of podophyllotoxin.
3. PACLITAXEL
Paclitaxel (Taxol®: is the registered trade name of a formulated drug based on the chemical paclitaxel and
marketed by Bristol-Myers Squibb Co., USA) is a novel complex diterpenoid originally isolated from the stem bark
of Pacific yew (Taxus brevifolia Nutt.) (Taxaceae) [109]. It is of immense importance as an antineoplastic agent,
being widely used for the treatment of advanced, progressive and drug refractory ovarian cancer [110, 111, 112] and
breast cancer [113]. Since 1992 it was approved by the FDA for the treatment of ovarian cancer; approval for
treatment of breast cancer followed in 1994 [114]. Clinical use of paclitaxel has increased steadily since then and
now it is also used for the treatment of lung cancer [115], head and neck cancer [116], renal, prostrate, colon, cervix,
gastric and pancreatic cancers [111, 117, 118, 119]. Apart from being an excellent antineoplastic agent it is also
effective against noncancerous conditions like polycystic kidney diseases [120] and has shown promising results in
multiple sclerosis [121] and AIDS related Karposi‟s sarcoma [122].
Paclitaxel has a unique mode of action. Unlike other antimicrotubule agents like podophyllotoxin,
colchicine, vinca alkaloids, combretastatin which inhibit microtubule assembly, paclitaxel stabilises microtubules
against depolymerisation; it promotes the polymerization of microtubules but inhibits depolymerisation [123, 124].
This unusual stability blocks the cell‟s ability to disassemble the mitotic spindle during cell division; cells are
blocked in the G2/M phase of the cell cycle [124] and this finally leads to cell death. Paclitaxel thus represents a new
class of antineoplastic agents [125].
The original source, T. brevifolia Nutt., is also the only FDA approved source of paclitaxel [126]. However, the
supply of paclitaxel from the bark is limited because the plant is not abundantly found in nature [127], it grows
slowly taking several decades to increase a few inches in diameter [128] and contains trace amounts of paclitaxel
(0.01% of dry weight of the bark) [129]. 1kg paclitaxel is produced after extraction from 10,000kg bark [130]. Also,
the removal of the bark results in the death of the tree [131]. Total synthesis of paclitaxel has been achieved [132,
133], but the process is complicated and not economically feasible. Thus, pharmaceutical companies still rely
heavily on plant sources. High demand, combined with such low yields from such slow-growing trees has prompted
researchers to explore alternative sources of paclitaxel.

e Journal of Biological Sciences www.ejarr.com
54
TABLE 4. PHYSICOCHEMICAL PROPERTIES OF PACLITAXEL [130]
IUPAC systematic name [2aR-(2a,4,4a,6,9,11,12,12a,12b)]
-Benzoylamino-hydroxy benzenepropionic acid 6,
12b-bis(acetyloxy)-12-(benzoyloxy)-2a,3,4,4a,5,6,
9,10,11,12,12a,12b-dodecahydro-4,11-dihydroxy-
4a,8,13,13-tetramethyl-5-oxo-7,11-methano-1H-
cyclodeca[3,4]benz[1]ester
Molecular formula C47H51NO14
Molecular weight 853.92g mol-1
Melting point 213-216°C
Solubility in water 4mg l-1
at RT
% Composition C(66.11%), H(6.02%), N(1.64%), O(26.23%)
Specific rotation -49º in methanol
UV max 227, 273nm in methanol
Mass spec M+H calculated for C47H51NO14=854.3388
3.1 OTHER NATURAL SOURCES OF PACLITAXEL
Paclitaxel has been isolated from various parts of several other species of Taxus like T. baccata, T. cuspidata, T.
canadensis, T. chinensis, T. x media, T. floridana, T. yunannensis, T. mairei, T. sumatrana and T. wallichiana [79,
131, 134, 135, 136, 137, 138, 139, 140, 141, 142]. However significant variation in taxane content exists among and
within population and species [135]. The authors also demonstrated that paclitaxel levels in shoot tissues of T.
brevifolia may exceed or equal that in the bark tissue of the same plant. Studies by Witherup et al. [143] and ElSohly
et al. [144] also revealed that the needles of Taxus species represent a rich and renewable source for paclitaxel and
other related taxanes. The needles of T. wallichiana (the Himalayan yew) are particularly good sources of 10-
deacetylbaccatin III which can be used as a precursor for the semisynthesis of paclitaxel and docetaxel (taxotere®)
[145], an analogue of paclitaxel [146]. Kwak et al. [131] detected paclitaxel from seeds of T. baccata, T. brevifolia,
T. cuspidata and T. cuspidata var. latifolia, the content varied according to different stages of seed maturation. It has
also been isolated from needles and bark of Austrotaxus spicata and Torreya grandis [147].
3.2 ALTERNATIVE SOURCES OF PACLITAXEL
The increase in the use of paclitaxel for cancer chemotherapy and basic research warrants efforts to improve the
existing production processes. Semisynthesis is the major route for the production of paclitaxel and related taxoids
[148, 149, 150, 151]. It involves a limited number of steps to convert paclitaxel pathway intermediates (viz. baccatin
III isolated from Taxus needles) to the target compound. But purification of semisynthetic precursors from plant
tissues is required to separate the desired intermediates from phenolics, lipids and other contaminants occurring in
planta.
Few organisms producing paclitaxel include a group of endophytic fungi of Taxus species viz. Taxomyces
andraenae, Fusarium sp. Alternaria sp. [152, 153], an unknown fungus from Taxus yunannensis [154],
Pestalotiopsis microspora and Phoma sp. isolated from T. wallichiana [155, 156]. In a recent study, genes coding
for 10-deacetylbaccatin III-10-O-acetyl transferase and C-13 phenylpropanoid side chain-CoA acyltransferase were
used as molecular markers for screening of taxol-producing endophytic fungi. Using PCR, three out of ninety
endophytic fungi, isolated from T x media and T. yunnanensis, gave positive results. These 3 strains, when grown in
300 ml potato/dextrose liquid medium at 25°C for 10 days, contained 100-160 mg taxol g-1
dry wt of mycelium
[157]. A paclitaxel producing bacterium Erwinia sp. was reported from T. canadensis [158].
3.3 BIOSYNTHESIS OF PACLITAXEL
Paclitaxel (C47H51NO14, m.p.213~216°C, mol. wt. 853.92g mol-1
) (Figure 2) belongs to the class of taxane
diterpenoids which currently has over 300 known members [159]. The structure of paclitaxel differs from the other
members in having a complex β-phenylisoserine side chain esterifying the C-13 position and has the unusual

e Journal of Biological Sciences www.ejarr.com
55
oxetane ring [114]. The biosynthetic pathway is complex involving several enzymatic steps and is not totally
elucidated. The taxane skeleton is synthesized from geranylgeranyl diphosphate. The committed step in the
biosynthesis of paclitaxel and related taxoids is the cyclization of the universal diterpenoid precursor geranylgeranyl
diphosphate to taxa-4(5),11(12)-diene by taxadiene synthase. A cDNA encoding taxadiene synthase (tasy) from T.
brevifolia was isolated by [160]. Hydroxylation of taxa-4(5),11(12)-diene at C-5 to taxa-4(20),11(12)-dien-5α-ol
catalysed by cytochrome P450 taxadiene 5α-hydroxylase is probably the next step in the biosynthetic pathway. The
following steps include acetylation of taxa-4(20),11(12)-dien-5α-ol to taxa-4(20),11(12)-dien-5α-yl acetate by taxa-
4(20),11(12)-dien-5α-ol-O-acetyltransferase and hydroxylation of taxa-4(20),11(12)-dien-5α-yl acetate to taxadien-
5α,10β-diol monoacetate by cytochrome P450 taxane 10β-hydroxylase. Biosynthetic steps from taxadien-5α,10β-
diol monoacetate to 2-debenzoyltaxane are not yet known. Bezoylation of 2-debenzoyltaxane to 10-
deacetylbaccation III by a taxane 2α-O-benzoyltransferase and acetylation of 10-deacetylbaccatin III to baccatin III
by a 10-deacetylbaccatin III-10-O-acetyltransferase are the next steps. Phenylisoserine derived from phenylalanine
by phenylalanine aminomutase, is condensed with baccatin III by baccatin III-10-O-phenylpropanoyltransferase to
3'-N-debenzoyl-2-deoxy-taxol. N-debenzoyl-deoxytaxol N-benzoyltransferase adds a benzoyl-CoA moiety to the 2-
deoxytaxol and a subsequent benzamidation produces paclitaxel. Of the several genes involved in the biosynthesis
of paclitaxel, those encoding taxadiene synthase, taxadien-5α-ol-O-acetyltransferase, cytochrome P450 taxanedienyl
acetate 10β-hydroxylase, 10-deacetylbaccatin III-10β-O-acetyltransferase and taxane 2α-O-benzoyltransferase have
been isolated, expressed and characterized [161, 162, 163, 164]. A thorough understanding of the pathway and its
rate limiting steps is required to improve the biological production of paclitaxel and related taxoids. With the
corresponding enzymes identified and assays developed, the role of each enzymatic step in the control of pathway
flux can be assessed by in vitro and in vivo studies and suitable strategies can be developed to isolate and
overexpress the corresponding genes in Taxus cell cultures for improving the production of paclitaxel.
3.4 PROPAGATION OF Taxus SPECIES BY CONVENTIONAL AND IN VITRO TECHNIQUES
Species of Taxus are propagated by seeds, cuttings and grafting [165]. Propagation by cuttings is well
documented for T. brevifolia, T x media, T. cuspidata [166] and T. baccata [167]. Cutting and grafting techniques
have been employed in the propagation of Himalayan yew T. wallichiana [168, 169]. Of the different treatments
applied to vegetatively propagate the Himalayan yew using branch cuttings, IBA dip treatment proved to be the best
method for induction of roots in stem cuttings [170].
Propagation of Taxus sp. by in vitro techniques is also reported. Taxus species are found to be recalcitrant
towards direct regeneration from axillary buds, nodal, internodal and leaf explants [171]. Taxus seeds have a lengthy
seed dormancy period of nearly 1.5-2.0 years [172] which can be overcome by embryo culture techniques [173].
Propagation by embryo culture technique has been reported for T. brevifolia, T. x media, T. cuspidata, T. baccata, T.
baccata stricta, T. mairei, T. wallichiana [128, 172, 174, 175, 176, 177].
Plant regeneration via direct organogenesis from zygotic embryos has been reported in T. brevifolia[178] and by
indirect organogenesis in T. chinensis [179]. Chee in 1996 [180] reported regeneration of T. brevifolia from

e Journal of Biological Sciences www.ejarr.com
56
immature zygotic embryos via somatic embryogenesis. Regeneration of T. wallichiana plants via shoot
organogenesis from callus cultures derived from zygotic embryos is also reported [181]. In a recent study, Datta and
Jha [182] reported regeneration of complete plantlets of the Himalayan yew via somatic embryogenesis from zygotic
embryo derived callus cultures.
3.5 IN VITRO PRODUCTION OF PACLITAXEL AND RELATED TAXANES
Cell cultures of Taxus represent a potential alternative source of paclitaxel and related taxanes. Rohr [183] first
established callus cultures from different explants of T. baccata, including microspores while production of
paclitaxel using cell cultures of Taxus was first reported by Christen et al. [184]. Since then, cell cultures have been
reported from different explant sources of different Taxus species with detectable amount of taxanes. Almost every
Taxus species has been studied in terms of media optimization to yield maximum biomass [185]. Fett-Neto et al.
[134] reported induction of callus cultures from different types of explants of T. cuspidata and T. canadensis and
paclitaxel in amounts comparable to T. brevifolia bark was detected from callus cultures of T. cuspidata. Paclitaxel
was also produced by immobilized cell cultures of T. cuspidata. Improved growth and paclitaxel yield in calli of T.
cuspidata were obtained by several sequential modifications of the culture medium viz. addition and/or variation in
the concentration of sucrose, B5 organic supplements, GA3, combinations of 2,4-D/kinetin ratios, media salts and
organic supplements, phenylalanine, casein hydrolysate and variation of medium pH [186]. Wickremesinhe and
Arteca [187] reported induction of callus cultures from stems and needles of T. brevifolia, T. cuspidata and cultivars
of T. baccata and T. x media using different concentrations of 2,4-D, IBA or NAA in combination with kinetin;
paclitaxel could be detected from the callus lines. The authors also reported establishment of cell suspension cultures
from T. x media cv. Hicksii callus cultures after optimizing the carbon source in the medium and detected paclitaxel
from such cultures [188]. Jha and Jha [174] reported establishment of fast growing cell lines of the Himalayan yew
by selection and cloning; though the level of paclitaxel was low, high levels of 10-deacetylbaccatin III were detected
in suspension cultures and culture media, thereby representing a potential source of the precursor of paclitaxel. In a
similar study, screening of cultures derived from different types of explants collected from a number of T.
wallichiana trees growing naturally in different regions of Darjeeling led to the establishment of high paclitaxel
yielding cell lines [189]. Recently, taxane analysis in embryogenic and non embryogenic calli derived from zygotic
embryos of the Himalayan yew revealed that paclitaxel accumulation was higher in embryogenic calli than in non
embryogenic calli [182].
For efficient production of paclitaxel, the effects of cultivation environment and various precursors and
elicitors were studied. Mirjalili and Linden [190, 228] and their co-workers investigated the effects of different
concentrations and combinations of oxygen, carbon dioxide and ethylene on cell growth and paclitaxel production in
cell suspension cultures of T. cuspidata. They found that a low headspace oxygen concentration promoted early
production of paclitaxel while a high carbon dioxide concentration was inhibitory. The effect of temperature shift on
cell growth and paclitaxel production was studied in suspension cultures of T. chinensis [191]. Cell growth was
optimum at 24°C while paclitaxel production reached its maximum at 29°C. The effect of osmotic pressure on
paclitaxel production was investigated in cell suspension cultures of T. chinensis [192]. High osmotic pressure
conditions generated by mannitol, sorbitol and polyethylene glycol enhanced paclitaxel production.
The accumulation of paclitaxel and related taxanes in Taxus is thought to be a biological response to specific
external stimuli [78]. Supplementation of biotic and abiotic elicitors in cell cultures of different Taxus species has
shown to affect both cell growth and paclitaxel accumulation. The use of fungal elicitors and other selected
compounds for the production of paclitaxel by suspension cultures of T. brevifolia was first demonstrated by
Christen et al. [193]. Ciddi et al. [194] demonstrated improved taxoid production in cell suspension cultures of
Taxus sp. after the addition of cell extracts and culture filtrates of Penicillium minioluteum, Botrytis cinerea,
Verticillium dahliae and Gilocladium deliqucescens. Aspergillus niger, an endophytic fungus isolated from the inner
bark of T. chinensis, resulted in a two fold increase in paclitaxel yield when added as an elicitor in the late
exponential growth phase of T. chinensis cell suspension cultures [195]. Reports indicate methyl jasmonate to be an
effective elicitor for the in vitro production of paclitaxel and related taxanes [78, 79, 80, 88]. Probably jasmonates
act as signal compounds in the elicitation process leading to de novo transcription and translation of genes and
ultimately to the biosynthesis of secondary metabolites in plant cell cultures [79]. Yukimune et al. [78] suggested
that methyl jasmonate contributes to paclitaxel production by activating the biosynthetic steps from geranylgeranyl
diphosphate to baccatin III and subsequent steps leading to the formation of paclitaxel from baccatin III. Mirjalili
and Linden [190] claimed that a higher level of paclitaxel was obtained in cell suspension cultures of T. cuspidata
when ethylene was added in combination with methyl jasmonate compared to treatment with methyl jasmonate

e Journal of Biological Sciences www.ejarr.com
57
alone. Paclitaxel accumulation in cell suspension cultures of T. media was improved by adding methyl jasmonate
together with mevalonate and N-benzoylglycine [196]. Another study showed that the application of ethylene
inhibitors to Taxus cell cultures before elicitor treatment was an effective strategy to enhance elicitor-induced
paclitaxel production suggesting that the control of ethylene production is important for the elicitation of paclitaxel
biosynthesis in cultured cells [197]. Yu et al. [198] reported that a T. chinensis culture treated with a fungal elicitor
in combination with salicylic acid achieved higher biomass and paclitaxel production than when treated with the
fungal elicitor alone. The effects of several other elicitors like arachidonic acid [199], silver ion [200], chitosan [200,
201] and La 3+
ion [202] on paclitaxel production have also been studied.
Brassinolide, a member of brassinosteroid group of steroidal lactones, increased paclitaxel content in cell
suspension culture of T. chinensis [203]. Clinton et al. [204] reported that a strong correlation exists between
paclitaxel and abscisic acid recovered from yew and suggested that an increase in abscisic acid concentration might
lead to an increase in paclitaxel production. Indeed, Luo et al. [205] demonstrated that abscisic acid could stimulate
paclitaxel accumulation in cell suspension cultures of T. chinensis and the stimulating effect depended on the dosage
of abscisic acid as well as the stage of cell growth.
Paclitaxel yields in both callus and cell suspension cultures of T. cuspidata were improved after the addition of
phenylalanine and other potential paclitaxel side chain precursors at different concentrations [206]. The promotion
of paclitaxel accumulation in in vitro cultures by phenylalanine might be due to its involvement as a precursor for
the C13 N-benzoylphenylisoserine side chain of paclitaxel. Significant enhancement in the level of paclitaxel was
noted in a cell line of T. wallichiana by supplementation of the basal medium with IAA conjugates like IAA-
phenylalanine and IAA-glycine [207]. Using three different cell lines of T. wallichiana the authors demonstrated
that 2,4-D and IAA-phenylalanine when present alone favoured cell growth and paclitaxel production but when
combined enhanced biomass to a maximum without increasing paclitaxel accumulation, suggesting that a two stage
culture might be needed to optimize paclitaxel accumulation in cell cultures of T. wallichiana. Use of a two stage
culture method for optimum growth and taxane production was also suggested by Furmanowa et al. [79]. In
another study, callus cultures established from young leaf explants of the Himalayan yew showed the presence of
taxanes viz. paclitaxel, 10-DAB and baccatin III [208]. Different precursors and growth retardants like
phenylalanine, sodium benzoate, hippuric acid, leucine, ancymidol, 2-chloroethyl phosphonic acid and
chlorocholine chloride improved the accumulation of taxanes in these cultures.
Expósito et al. [209] conducted an interesting study using cell cultures of T. baccata. A T. baccata cell line
growing for 20 days in a selected growth medium was treated at the beginning of the experiment with different
concentrations of taxol (25, 50, 100 and 200 mg l−1
). Compared with an untreated control, these exogenous taxol
concentrations stimulated cell-associated taxol content (up to 32.7 times in the presence of 200 mg l−1
exogenous
taxol), although higher concentrations significantly depressed cell viability. On the basis of RT-PCR expression
analysis of genes encoding taxadiene synthase (ts) and 1-deoxy-d-xylulose-5-phosphate synthase (dxs) from treated
and nontreated T. baccata cell line cultures, it was observed that exogenous taxol clearly induced the mRNA levels
of both taxane-related enzymes. Exogenous taxol also caused a considerable increase in taxadiene synthase activity.
Cross species coculture proved to be another effective technique for successful production of paclitaxel. In a
recent study, cell suspensions of Taxus chinensis var. mairei were cocultured with its endophytic fungi, Fusarium
mairei, in a 20 litre co-bioreactor, resulting in a production of 25.63 mg l-1
paclitaxel within 15 days; it was
equivalent to a productivity of 1.71 mg l-1
per day and 38-fold higher than that by uncoupled culture [210].
3.6 GENETIC TRANSFORMATION OF Taxus FOR PACLITAXEL PRODUCTION
Genetic transformation of T. baccata and T. brevifolia by Agrobacterium tumefaciens strains Bo542 and C58 is
reported [211]. Strain Bo542 was more efficient in inducing galls on shoot segments of mature yew trees than strain
C58. Although tumours were induced on both the species, T. baccata was more susceptible to transformation than T.
brevifolia. In contrast to untransformed callus cultures, the gall cell lines proliferated on phytohormone free
medium. Paclitaxel and related taxanes were detected in the transgenic cell lines.
In another study, A. rhizogenes strain LBA9402 was used to develop hairy root cultures of T. x media var.
Hicksii [212]. These cultures accumulated twice the amount of paclitaxel than that in the bark of T. brevifolia after
treatment with methyl jasmonate.
e Journal of Biological Sciences www.ejarr.com
58
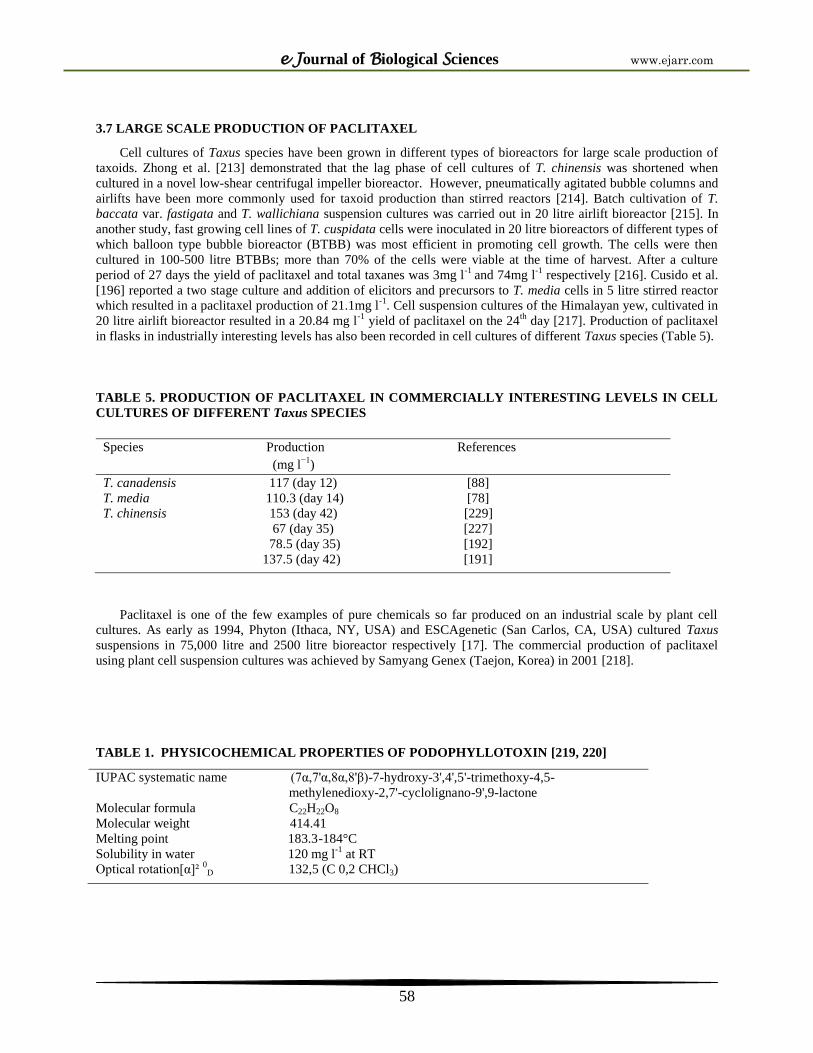
3.7 LARGE SCALE PRODUCTION OF PACLITAXEL
Cell cultures of Taxus species have been grown in different types of bioreactors for large scale production of
taxoids. Zhong et al. [213] demonstrated that the lag phase of cell cultures of T. chinensis was shortened when
cultured in a novel low-shear centrifugal impeller bioreactor. However, pneumatically agitated bubble columns and
airlifts have been more commonly used for taxoid production than stirred reactors [214]. Batch cultivation of T.
baccata var. fastigata and T. wallichiana suspension cultures was carried out in 20 litre airlift bioreactor [215]. In
another study, fast growing cell lines of T. cuspidata cells were inoculated in 20 litre bioreactors of different types of
which balloon type bubble bioreactor (BTBB) was most efficient in promoting cell growth. The cells were then
cultured in 100-500 litre BTBBs; more than 70% of the cells were viable at the time of harvest. After a culture
period of 27 days the yield of paclitaxel and total taxanes was 3mg l-1
and 74mg l-1
respectively [216]. Cusido et al.
[196] reported a two stage culture and addition of elicitors and precursors to T. media cells in 5 litre stirred reactor
which resulted in a paclitaxel production of 21.1mg l-1
. Cell suspension cultures of the Himalayan yew, cultivated in
20 litre airlift bioreactor resulted in a 20.84 mg l-1
yield of paclitaxel on the 24th
day [217]. Production of paclitaxel
in flasks in industrially interesting levels has also been recorded in cell cultures of different Taxus species (Table 5).
TABLE 5. PRODUCTION OF PACLITAXEL IN COMMERCIALLY INTERESTING LEVELS IN CELL
CULTURES OF DIFFERENT Taxus SPECIES
Species Production References
(mg l−1
)
T. canadensis 117 (day 12) [88]
T. media 110.3 (day 14) [78]
T. chinensis 153 (day 42) [229]
67 (day 35) [227]
78.5 (day 35) [192]
137.5 (day 42) [191]
Paclitaxel is one of the few examples of pure chemicals so far produced on an industrial scale by plant cell
cultures. As early as 1994, Phyton (Ithaca, NY, USA) and ESCAgenetic (San Carlos, CA, USA) cultured Taxus
suspensions in 75,000 litre and 2500 litre bioreactor respectively [17]. The commercial production of paclitaxel
using plant cell suspension cultures was achieved by Samyang Genex (Taejon, Korea) in 2001 [218].
TABLE 1. PHYSICOCHEMICAL PROPERTIES OF PODOPHYLLOTOXIN [219, 220]
IUPAC systematic name (7α,7'α,8α,8'β)-7-hydroxy-3',4',5'-trimethoxy-4,5-
methylenedioxy-2,7'-cyclolignano-9',9-lactone
Molecular formula C22H22O8
Molecular weight 414.41
Melting point 183.3-184°C
Solubility in water 120 mg l-1
at RT
Optical rotation[α]² 0
D 132,5 (C 0,2 CHCl3)

e Journal of Biological Sciences www.ejarr.com
59
4. CONCLUSIONS
Podophyllotoxin and paclitaxel are two potent plants derived antitumour compounds. Supply of these
compounds from traditional sources is limited either due to the dwindling condition of plant sources or due to trace
amounts of compounds present in the plants. So several alternative strategies have been adapted to increase the
production of these compounds. Plant tissue culture represents a sustainable and contained production system which
is constantly improving with respect to reliability and capacity. Indeed, recent years have seen great achievements in
the production of these compounds using plant cell culture systems. Although large scale production of paclitaxel is
reported, production of podophyllotoxin on a commercial scale is yet to be achieved; technological developments
are required. Development of transgenics and metabolic engineering are other important tools. Genes encoding
enzymes for the biosynthesis of paclitaxel and podophyllotoxin can be expressed in fast growing microorganisms.
Genes controlling slow steps in the pathways can be overexpressed or synthesis of unwanted metabolites can be
suppressed by various sense and antisense technologies. However, more basic knowledge about the pathways to
paclitaxel and podophyllotoxin and their regulation is required. The challenge remains to elucidate the complete
biosynthetic pathways, thereby exploiting cell cultures more rationally for the production of these compounds so
much in need by the pharmaceutical industry.
Acknowledgement
A. M. thanks Council of Scientific and Industrial Research, New Delhi, for the award of Senior Research
Fellowship. The financial assistance from Department of Biotechnology, Ministry of Science and Technology, India,
is gratefully acknowledged.
REFERENCES:
[1] Wink M, Alfermann AW, Franke R, Wetterauer B, Distl M, Windhövel J, Krohn O, Fuss E, Garden H,
Mohagheghzadeh A, Wildi E, and Ripplinger P, “Sustainable bioproduction of phytochemicals by plant in
vitro cultures: anticancer agents”, Plant Genetic Resources, Vol. 3, 2005, pp. 90–100.
[2] Cragg GM, and Newman DJ, “Plants as a source of anti-cancer agents”, In: Elisabetsky E, Etkin NL (ed)
Ethnopharmacology, in Encyclopedia of Life Support Systems (EOLSS), Developed under the Auspices of
the UNESCO, Eolss Publishers, Oxford ,UK, http://www.eolss.net, 2004
[3] Srivastava V, Negi AS, Kumar KJ, Gupta MM, and Khanuja SPS, “Plant-based anticancer molecules: A
chemical and biological profile of some important leads”, Bioorganic and Medicinal Chemistry, Vol. 13,
2005, pp. 5892–5908.
[4] Verpoorte R, Contin A, and Memelink J, “Biotechnology for the production of plant secondary metabolites”,
Phytochemistry Reviews, 1, 2002, pp. 13–25.
[5] Mulabagal V, and Tsay HS, “Plant cell cultures- an alternative and efficient source for the production of
biologically important secondary metabolites”, International Journal of Applied Science and Engineering,
Vol. 1, 2004, pp. 29–48.
[6] Rischer H, and Oksman-Caldentey KM, “Biotechnological utilization of plant genetic resources for the
production of phytopharmaceuticals”, Plant Genetic Resources, Vol. 3, 2005, pp. 83–89.
[7] DiCosmo F, and Misawa M, “Eliciting secondary metabolism in plant cell cultures”, Trends in Biotechnology,
Vol. 3, 1985, pp. 318–322.
[8] Mahagamasekara MGP, and Doran PM, “Intergeneric coculture of genetically transformed organs for the
production of scopolamine”, Phytochemistry, Vol. 47, 1998, pp. 17–25.
[9] Pereira AMS, Bertoni BW, Câmara FLA, Duarte IB, Queiroz MEC, Leite VGM, Moraes RM, Carvalho D, and
França SC “Co-cultivation of plant cells as a technique for the elicitation of secondary metabolite
production”, Plant Cell, Tissue and Organ Culture,Vol. 60, 2000, pp. 165–169.
[10] Lin H, Kian HK, and Doran PM, “Production of podophyllotoxin using cross-species coculture of Linum
flavum hairy roots and Podophyllum hexandrum cell suspensions”, Biotechnology Progress, Vol. 19, 2003,
pp. 1417–1426.

e Journal of Biological Sciences www.ejarr.com
60
[11] Paek KY, Chakrabarty D, and Hahn EJ, “Application of bioreactor systems for large scale production of
horticultural and medicinal plants”, Plant Cell, Tissue and Organ Culture,Vol. 81, 2005, pp. 287–300.
[12] Tulecke W, and Nickell LG, “Production of large amounts of plant tissue by submerged culture”, Science, Vol.
130, 1959, pp. 863–864.
[13] Ritterhaus E, Ulrich J, and Westphal K, “Large-scale production of plant-cell cultures”, International
Association of Plant Tissue Culture Newsletter,Vol. 61, 1990, pp. 2-10.
[14] Fujita Y, Tabata M, Nishi A, and Yamada Y, “New medium and production of secondary compounds with the
two staged culture method”, In: Fujiwara A (ed.) Plant Tissue Culture 1982, Tokyo, the Japanese
Association for Plant Tissue Culture, Maruzen Co. Ltd., 1982, pp. 399-400.
[15] Hibino K, and Ushiyama K “Commercial production of ginseng by plant tissue culture technology”, In: Fun TJ,
Singh G, Curtis WR (ed) Plant Cell and Tissue Culture for the Production of Food Ingredients, Kluwer
Academic, Plenum Publisher, 1999, pp. 215–224.
[16] Fujita M, “Industrial production of shikonin and berberine”, In: Brock E, Marsh J (ed) Applications of plant cell
and tissue culture, Ciba foundation symposium 137 Wiley, New York, 1988, pp. 228-238.
[17] Smith MAL, “Large scale production of secondary metabolites”, In Terzi M, Cella R and Falavigna A (ed)
Current issues in plant molecular and cellular biology. Kluwer Academic Publishers, Dordrecht, The
Netherlands , 1995, pp. 669-674.
[18] Holthius JJM, “Etoposide and teniposide: Bioanalysis, metabolism and clinical pharmokinetics”,
Pharmaceutisch Weekblad , Vol. 10, 1988, pp. 101–116.
[19] Stahelin HF, and Wartburg AV von, “The chemical and biological route from podophyllotoxin glucoside to
etoposide”, Cancer Research,Vol. 51, 1991, pp. 5–15.
[20] Imbert TF, “Discovery of podophyllotoxins”, Biochimie Vol. 80, 1998, pp. 207–222.
[21] Petersen M, and Alfermann AW “The production of cytotoxic lignans by plant cell cultures”, Applied
Microbiology and Biotechnology, Vol. 55, 2001, pp. 135–142.
[22] Koulman A “Podophyllotoxin: a study of the biosynthesis, evolution, function and use of podophyllotoxin and
related lignans”, http://irs.ub.rug.nl/ppn/252657888, 2003.
[23] Desbène S, and Giorgi-Renault S, “Drugs that Inhibit Tubulin Polymerization: The Particular Case of
Podophyllotoxin and Analogues”, Current Medicinal Chemistry, Vol. 2, 2002, pp. 71–90.
[24] Hainsworth JD, and Greco FA, “Etoposide: twenty years later”, Annals of Oncology, Vol. 6, 1995, pp. 325–
341.
[25] Hande KR, “Etoposide: four decades of development of a topoisomerase II inhibitor”, European Journal of
Cancer, Vol. 34, 1998, pp. 1514-1521.
[26] Carlstrom K, Hedin PJ, Jansson C, Lerndal T, Lien J, Weitaft TT, and Axelson T, “Endocrine effects of
podophyllotoxin derivatives drug CPH 82 (Reumacon) in patients with rheumatoid arthritis”, Scandinavian
Journal of Rheumatology, Vol. 29, 2000, pp. 89–94.
[27] Kaplan IW, “Condyloma acuminata”, New Orleans Med Surg J, Vol. 94, 1942, pp. 388–395.
[28] Bedows E, and Hatfield GM, “An investigation of the antiviral activity of Podophyllum peltatum”, Journal of
Natural Products, Vol. 45, 1982, pp. 725–729.
[29] Canel C, Moraes RM, Dayan FE, and Ferreira D, “Molecules of interest podophyllotoxin”, Phytochemistry Vol.
54, 2000, pp. 115–120.
[30] MacRae WD, and Towers GHN, “Biological activities of lignans”, Phytochemistry Vol. 23, 1984, pp. 1207–
1220.
[31] Pelter A, “Lignans: Some properties and syntheses”, Recent Advances in Phytochemistry, Vol. 20, 1986, pp.
201–241.
[32] Lewis NG, and Davin LB, “Lignans: Biosynthesis and function”, In: Barton DHR, Nakanishi K and Meth-Coon
O (ed) Comprehensive natural products chemistry, vol. 1, Sankawa U (ed) Polyketides and other secondary
metabolites including fatty acids and their derivatives. Elsevier Science, Oxford, 1999, pp. 639-712.
[33] Ward RS, “The synthesis of lignans and neolignans”, Chem Soc Rev, Vol. 11, 1982, pp. 75–125.
[34] Jackson DE, and Dewick PM, “Biosynthesis of Podophyllum lignans-I. Cinnamic acid precursors of
podophyllotoxin in Podophyllum hexandrum”, Phytochemistry, Vol. 23, 1984b, pp. 1029–1035.
[35] Davin LB, Wang HB, Crowell AL, Bedgar DL, Martin DM, Sarkanen S, and Lewis NG, “Stereoselective
biomolecular phenoxy radical coupling by an auxiliary (dirigent) protein without an active centre.” Science,
Vol. 275, 1997, pp. 362–366.

e Journal of Biological Sciences www.ejarr.com
61
[36] Chu A, Dinkova A, Davin LB, Bedgar DL, and Lewis NG, “Stereospecificity of (+)-pinoresinol and (+)-
lariciresinol reductase from Forsythia intermedia”, Journal of Biological Chemistry, Vol. 268, 1992, pp.
27026–27033.
[37] Dinkova-Kostova AT, Gang DR, Davin LB, Bedgar DL, Chu A, and Lewis NG, “(+)-Pinoresinol/(+)-
lariciresinol reductase from Forsythia intermedia: protein purification, cDNA cloning, heterologous
expression and comparison to isoflavone reductase”, Journal of Biological Chemistry, Vol. 271, 1996, pp.
29473–29482.
[38] Umezawa T, Davin LB, and Lewis NG, “Formation of lignans (–)-secoisolariciresinol and (–)-matairesinol with
Forsythia intermedia cell-free extracts”, Journal of Biological Chemistry, Vol. 266, 1991, pp. 10210–
10217.
[39] Xia ZQ, Costa MA, Pélissier HC, Davin LB, and Lewis NG, “Secoisolariciresinol dehydrogenase purification,
cloning and functional expression: Implications for human health protection”, Journal of Biological
Chemistry, Vol. 276, 2001, pp. 12614–12623.
[40] Gang DR, Costa MA, Fujita M, Dinkova-Kostova AT, Wang HB, Burlat V, Martin W, Sarkanen S, Davin LB,
and Lewis NG, “Regiochemical control of monolignol radical coupling: a new paradigm for lignin and
lignan biosynthesis”, Chemistry & Biology, Vol. 6, 1999, pp. 143–151.
[41] Kuhlmann S, Kranz K, Lücking B, Alfermann AW, and Petersen M, “Aspects of cytotoxic lignan biosynthesis
in suspension cultures of Linum nodiflorum”, Phytochemistry Reviews, Vol. 1, 2002, pp. 37–43.
[42] Arroo RRJ, Alfermann AW, Medarde M, Petersen M, Pras N, and Woolley JG, “Plant cell factories as a source
of anticancer lignans”, Phytochemistry Reviews Vol. 1, 2002 pp. 27–35.
[43] Henges A, "Biosynthese und Kompartimentierung von Lignanen in Zellkulturen von Linum album", PhD
Thesis, University of Düsseldorf, 1999.
[44] Jackson DE, and Dewick PM, “Aryltetralin lignans from Podophyllum hexandrum and Podophyllum peltatum”,
Phytochemistry, Vol. 23, 1984a, pp. 1147–1152.
[45] Heyenga AG, Lucas JA, and Dewick PM, “Production of tumour inhibitory lignans in callus culltures of
Podophyllum hexandrum”, Plant Cell Reports, Vol. 9, 1990, pp. 382–385.
[46] Bhadula SK, Singh A, Lata H, Kunyal CP, and Purohit AN, “Genetic resources of Podophyllum hexandrum
Royle, an endangered medicinal species from Garhwal Himalaya, India”, International Plant Genetic
Resources Newsletter, Vol. 106, 1996, pp. 26–29.
[47] Nadeem M, Palni LMS, Purohit AN, Pandey H, and Nandi SK, “Propagation and conservation of Podophyllum
hexandrum Royle: an important medicinal herb”, Biological Conservation, Vol. 92, 2000, pp. 121–129.
[48] Greenwald RB, Conover CD, Pendri A, Choe YH, Martinez A, Wu D, Guan S, Yao Z, and Shum KL, “Drug
delivery of anticancer agents: water soluble 4-poly (ethylene glycol) derivatives of the lignan,
azadirachtin”, Journal of Controlled Release, Vol. 61, 1999, pp. 281–294.
[49] Uden W van, Pras N, Visser JF, and Malingre TM, “Detection and identification of podophyllotoxin produced
by cell cultures derived from Podophyllum hexandrum Royle”, Plant Cell Reports, Vol. 8, 1989, pp. 165–
168.
[50] Nadeem M, Palni LMS, Kumar A, and Nandi SK, “Podophyllotoxin content, above- and belowground biomass
in relation to altitude in Podophyllum hexandrum populations from Kumaun region of the Indian Central
Himalaya”, Planta Medica, Vol. 73, 2007, pp. 388–391.
[51] Choudhary DK, Kaul BL, and Khan S, “Cultivation and conservation of Podophyllum hexandrum – an
overview”, Journal of Medicinal and Aromatic Plant Sciences, Vol. 20, 1998, pp. 1071–1073.
[52] Foster S, “Medicinal plant conservation and genetic resources: examples from the temperate northern
hemisphere”, Acta Horticulturae, Vol. 330, 1993, pp. 67–73.
[53] Airi S, Rawal RS, Dhar U, and Purohit AN, “Population studies on Podophyllum hexandrum Royle- a
dwindling medicinal plant of the Himalaya”, Plant Genetic Resources Newsletter, Vol. 110, 1997, pp. 20–
34.
[54] Bush EJ, and Jones DW, “Asymmetric total synthesis of (−)-podophyllotoxin”, Journal of the Chemical Society
− Perkin Transactions, Vol. 1, 1995, pp. 151–155.
[55] Berkowitz D, Choi S, and Maeng J, “Enzyme-assisted asymmetric total synthesis of (−)-podophyllotoxin and
(−)-picropodophyllin”, Journal of Organic Chemistry, Vol. 65, 2000, pp. 847–860.
[56] Badhwar RL, and Sharma BK, “A note on germination of Podophyllum seeds”, Indian Forester, Vol. 89, 1963,
pp. 445–447.
[57] Singh A, “Ecophysiology of Podophyllum hexandrum Royle”, Ph.D thesis, H.N.B. Garhwal University, 1992.

e Journal of Biological Sciences www.ejarr.com
62
[58] Pandey H, Nandi SK, Kumar A, Palni UT, and Palni LMS, “Podophyllotoxin content in Podophyllum
hexandrum Royle plants of known age of seed origin and grown at a lower altitude”, Acta Physiologiae
Plantarum, Vol. 29, 2007, pp. 121–126.
[59] Arumugam N, and Bhojwani SS, “Somatic embryogenesis in tissue cultures of Podophyllum hexandrum”,
Canadian Journal of Botany, Vol. 68, 1990, pp. 487–491.
[60] Majumder A, “Biotechnological approaches for the production of cytotoxic anticancerous compounds”, PhD
thesis, Calcutta University, Calcutta, India, 2008.
[61] Moraes-Cerdeira RM, Burandt CL, Bastos JK, Nanayakkara NPD, and McChesney JD, “In vitro propagation of
Podophyllum peltatum”, Planta Medica, Vol. 64, 1998, pp. 42–45.
[62] Kadkade PG, “Formation of podophyllotoxin by Podophyllum peltatum tissue cultures”, Naturwissenchaften,
Vol. 68, 1981, pp. 481–482.
[63] Kadkade PG, “Growth and podophyllotoxin production in callus tissues of Podophyllum peltatum”, Plant
Science Letters, Vol. 25, 1982, pp. 107–115.
[64] Majumder A, and Jha S, “Growth and podophyllotoxin production in callus and cell suspension cultures of
Podophyllum hexandrum”, Proceeding of National Symposium on Plant Biotechnology: New Frontiers,
CIMAP, Lucknow, 2007, pp. 433–438.
[65] Uden W van, Pras N, and Malingre TM, “The accumulation of podophyllotoxin-β-D-glucoside by cell
suspension cultures derived from the conifer Callitris drumondii”, Plant Cell Reports, Vol. 9, 1990c, pp.
257–260.
[66] Muranaka T, Miyata M, Kazutaka I, and Tachibana S, “Production of podophyllotoxin in Juniperus chinensis
callus cultures treated with oligosaccharides and a biogenetic precursor”, Phytochemistry, Vol. 49, 1998,
pp. 491–496.
[67] Anbazhagan VR, Ahn CH, Harada E, Kim YS, and Choi YE “Podophyllotoxin production via cell and
adventitious root cultures of Podophyllum peltatum”, In Vitro Cellular and Developmental Biology, Vol.
44, 2008, pp. 494–501.
[68] Chattopadhyay S, Srivastava AK, Bhojwani SS, and Bisaria VS, “Production of podophyllotoxin by plant cell
cultures of Podophyllum hexandrum in bioreactor”, Journal of Bioscience and Bioengineering, Vol. 93,
2002b, pp. 215–220.
[69] Chattopadhyay S, Srivastava AK, and Bisaria VS, “Optimisation of culture parameters for production of
podophyllotoxin in suspension culture of Podophyllum hexandrum”, Applied Biochemistry and
Biotechnology, Vol. 102/103, 2002a, pp. 381–393.
[70] Kieran PM, Malone DM, and MacLoughlin PF, “Effects of hydrodynamic and interfacial forces on plant cell
suspension systems” Advanced Biochemical Engineering/Biotechnology,Vol. 139, 2000, p. 177.
[71] Chattopadhyay S, Mehra RS, Srivastava AK, Bhojwani SS, and Bisaria VS, “Effect of major nutrients on
podophyllotoxin production in Podophyllum hexandrum suspension cultures”, Applied Microbiology and
Biotechnology, Vol. 60, 2003c, pp. 541–546.
[72] Rhodes MJC, Hamill J, Parr AJ, Robins RJ, and Walton NJ, “Strain improvement by screening and selection
techniques”, In: Robins RJ and Rhodes MJC (ed.) Manipilating secondary metabolism in culture.
Cambridge University Press, Cambridge, 1988, p. 82
[73] Yamada Y, and Sato F, “Production of berberine in cultured cells of Coptis japonica”, Phytochemistry, Vol. 20,
1981, pp. 545–547.
[74] Jha S, and Jha TB “A fast growing cell line of Taxus baccata L. (Himalayan yew) as a potential source of taxol
precursor” Current Science, Vol. 69, 1995, pp. 971–972.
[75] Mukherjee S, Ghosh B, and Jha S, “Establishment of forskolin yielding transformed cell suspension cultures of
Coleus forskohlii as controlled by different factors”, Journal of Biotechnology, Vol. 76, 2000a, pp. 73–81.
[76] Uden W van, Pras N, and Malingre TM “On the improvement of the podophyllotoxin production by
phenylpropanoid precursor feeding to cell cultures of Podophyllum hexandrum Royle”, Plant Cell, Tissue
and Organ Culture, Vol. 23, 1990a, pp. 217–224.
[77] Woerdenbag HJ, Uden W van, Frijlink HW, Lerk CF, Pras N, and Malingre TM, “Increased podophyllotoxin
production in Podophyllum hexandrum cell suspension cultures after feeding coniferyl alcohol as a β-
cyclodextrin complex”, Plant Cell Reports, Vol. 9, 1990, pp. 97–100.
[78] Yukimune Y, Tabata H, Higashi Y, and Hara H, “Methyl-jasmonate induced overproduction of paclitaxel and
baccatin III in Taxus cell suspension cultures”, Nature Biotechnology ,Vol. 14, 1996, pp. 1129–1132.
[79] Furmanowa M, Glowniak K, Syklowska-Baranek K, Zgórka G, and Józefczyk A, “Effect of picloram and
methyl jasmonate on growth and taxane accumulation in callus culture of Taxus x media var. Hatfieldii”,
Plant Cell, Tissue and Organ Culture, Vol. 45, 1997, pp. 75–79.

e Journal of Biological Sciences www.ejarr.com
63
[80] Moon WJ, Yoo BS, Kim D, and Byun SY, “Elicitation kinetics of taxane production in suspension cultures of
Taxus baccata Pendula”, Biotechnology Techniques, Vol. 12, 1998, pp. 79–81.
[81] Zabetakis I, Edwards R, and O'Hagan D, “Elicitation of tropane alkaloid biosynthesis in transformed root
cultures of Datura stramonium”, Phytochemistry Vol. 50, 1999, pp. 53–56.
[82] Kuzovkina IN, Guseva AV, Alterman IE, and Karnachuk RA, “Flavonoid production in transformed Scutellaria
baicalensis roots and ways of its regulation”, Russian Journal of Plant Physiology, Vol. 48, 2001, pp. 448–
452.
[83] Gundlach H, Müller M, Kutchan TM, and Zenk MH, “Jasmonic acid is a signal transducer in elicitor-induced
plant cell cultures”, Proceedings of the National Academy of Sciences, Vol. 89, 1992, pp. 2389–2393
[84] Zhong JJ, “Plant cell culture for production of paclitaxel and other taxanes”, Journal of Bioscience and
Bioengineering, Vol. 94, 2002, pp. 591–599.
[85] Ramakrishna SV, Reddy RG, Curtis WR, and Humphry AE, “Stimulation of solvetivone synthesis in free and
immobilized cells of Hyoscyamus muticus by Rhizoctonia solani fungal components”, Biotechnology
Letters, Vol. 15, 1993, pp. 307-310.
[86] Rajendran L, Suvarnalatha G, Ravishankar GA, and Venkataraman LV, “Enhancement of anthocyanin
production in callus cultures of Daucus carota L. under influence of fungal elicitors”, Applied
Microbiology and Biotechnology 42, 1994, pp. 227-231.
[87] Kim JH, Shin JH, Lee HJ, and Chung IS “Effect of chitosan on indirubin production from suspension culture of
Polygonum tinctorum”, Journal of Fermentation and Bioengineering, Vol. 83, 1997, pp. 206-208.
[88] Ketchum REB, Gibson DM, Croteau RB, and Shuler ML, “The kinetics of taxoid accumulation in cell
suspension cultures of Taxus following elicitation with methyl jasmonate”, Biotechnology and
Bioengineering, Vol. 62, 1999, pp. 97–105.
[89] Pedapudi S, Chin CK, and Pedersen H, “Production and elicitation of benzalacetone and the raspberry ketone in
cell suspension cultures of Rubus idaeus”, Biotechnology Progress, Vol. 16, 2000, pp. 346–349.
[90] Pitta-Alvarez ST, Spollansky TC, and Giulietti AM, “The influence of different biotic and abiotic elicitors on
the production and profile of tropane alkaloids in hairy root cultures of Brugsmansia candida”, Enzyme and
Microbial Technology, Vol. 26, 2000, pp. 252–258.
[91] Bulgakov VP, Tchernoded GK, Mischenko NP, Khodakovskaya MV, Glazunov VP, Radchenko SV, Zvereva
EV, Fedoreyev SA, and Zhuravlev YN, “Effect of salicylic acid, methyl jasmonate, ethephon and
cantharidin on anthraquinone production by Rubia cordifolia callus cultures transformed with the rolB and
rolC genes”, Journal of Biotechnology, Vol. 97, 2002, pp. 213–221.
[92] Kasparova M, and Siatka T, “Effect of salicylic acid on production of anthracene derivatives in a culture of
Rheum palmatum L. in vitro”, Ceská a Slovenská Farmacia, Vol. 51, 2002, pp. 177–181.
[93] Kittipongpatana N, Davis DL, and Porter JR “Methyl jasmonate increases the production of valepotriates by
transformed root cultures of Valerianella locusta”, Plant Cell, Tissue and Organ Culture, Vol. 71, 2002,
pp. 65–75.
[94] Yu KW, Gao WH, Hahn EJ, and Paek KY, “Jasmonic acid improves ginsenoside accumulation in adventitious
root culture of Panax ginseng C.A. Meyer”, Biochemical Engineering Journal, Vol. 11, 2002, pp. 211–215.
[95] Lee-Parsons CWT, Ertürk S, and Tengtrakool J, “Enhancement of ajmalicine production in Catharanthus
roseus cell cultures with methyl jasmonate is dependent on timing and dosage of elicitation”,
Biotechnology Letters, Vol. 0, 2004, pp. 1595-1599
[96] Fürden B van, Humburg A, and Fuss E, “Influence of methyl jasmonate on podophyllotoxin and 6-
methoxypodophyllotoxin accumulation in Linum album cell suspension cultures”, Plant Cell Reports, Vol.
24, 2005, pp. 312–317.
[97] Brodelius P, and Mosbach K, “Immobilised plant cells. General aspects”, Journal of Chemical Technology and
Biotechnology, Vol. 32, 1982, pp. 330–337.
[98] Pras N, Wichers HJ, Bruins AP, and Malingré TM, “Bioconversion of para-substituted monophenolic
compounds into corresponding catechols by alginate entrapped cells of Mucuna puriens”, Plant Cell,
Tissue and Organ Culture, Vol. 13, 1988, pp. 15–26.
[99] Sasson A, “Production of useful biochemicals by higher-plant cell cultures: biotechnological and economic
aspects”, Options Méditerranéennes- Série Seminaires, Vol. 14, 1991, pp. 59–74.
[100] Chattopadhyay S, Bisaria VS, and Srivastava AK, “Enhanced production of podophyllotoxin by Podophyllum
hexandrum using in situ cell retention bioreactor”, Biotechnology Progress, Vol. 19, 2003a, pp. 1026–
1028.
[101] Sevón N, and Oksman-Caldentey KM “Agrobacterium rhizogenes mediated transformation: root cultures as a
source of alkaloids”, Planta Medica, Vol. 68, 2002, pp. 859-868.

e Journal of Biological Sciences www.ejarr.com
64
[102] Sevón N, Hiltunen R, and Oksman-Caldentey KM “Somaclonal variation in Agrobacterium transformed roots
and protoplast-derived hairy root clones of Hyoscyamus muticus”, Planta Medica, Vol. 64, 1998, pp. 37-41.
[103] Oksman-Caldentey KM, Hiltunen R “Transgenic crops for improved pharmaceutical products”, Field Crops
Research, Vol.45, 1996, pp. 57-69
[104] Sevón N, “Tropane alkaloids in hairy roots and regenerated plants of Hyoscyamus muticus”, PhD Thesis,
University of Helsinki, Helsinki, Finland, 1997.
[105] Giri A, and Narasu ML, “Transgenic hairy roots: recent trends and applications”, Biotechnology Advances,
Vol. 18, 2000, pp. 1-22.
[106] Giri A, Giri CC, Dhingra V, and Narasu ML, “Enhanced podophyllotoxin production from Agrobacterium
rhizogenes transformed cultures of Podophyllum hexandrum”, Natural Product Letters, Vol. 15, 2001a ,
pp. 229–235.
[107] Eyberger AL, Dondapati R, and Porter JR, “Endophyte fungal isolates from Podophyllum peltatum produce
podophyllotoxin”, Journal of Natural Products, Vol. 69, 2006, pp. 1121–1124.
[108] Kour A, Shawl AS, Rehman S, Sultan P, Qazi PH, Suden P, Khajuria RK, and Verma V, “Isolation and
identification of an endophytic strain of Fusarium oxysporum producing podophyllotoxin from Juniperus
recurva”, World Journal of Microbiology and Biotechnology, Vol. 24, 2008. pp. 1115–1121.
[109] Wani MC, Taylor HL, Wall ME, Coggon P, McPhail AT, “Plant antitumour agents. VI. The isolation and
structure of Taxol, a novel antileukemic and antitumour agent from Taxus brevifolia”, Journal of the
American Chemical Society, Vol. 93, 1971, pp. 2325–2327.
[110] McGuire WP, Rowinsky EK, Rosenshein NB, Grumbine FC, Ettinger DS, Armstrong DK, and Donehower
RC, “Taxol: A unique antineoplastic agent with significant activity in advanced ovarian epithelial
neoplasms”, Annals of Internal Medicine, Vol. 11, 1989, pp. 273–279.
[111] Einzig AI, Wiernik PH, and Schwartz EL, “Taxol: a new agent active in melanoma and ovarian cancer”,
Cancer Treatment and Research, Vol. 58, 1991, pp. 89–100.
[112] Markman M, “Taxol: an important new drug in the management of epithelial ovarian cancer”, Yale Journal of
Biology and Medicine, Vol. 64, 1991, pp. 583–590.
[113] Holmes FA, Walters RS, Theriault RL, Forman AD, Newton LK, Raber MN, Buzdar AU, Frye DK, and
Hortabagyi GN, “Phase II trial of taxol, an active drug in the treatment of metastatic breast cancer”,
Journal of the National Cancer Institute, Vol. 83, 1991, pp. 1797–1805.
[114] Kingston DGI, “Recent advances in the chemistry of taxol”, Journal of Natural Products, Vol. 63, 2000, pp.
726–734.
[115] Ettinger DS, “Taxol in the treatment of lung cancer”, In: Abstracts of Second National Cancer Institute
Workshop on Taxol and Taxus, Alexandria, Virginia, 23-24 September 1992.
[116] Forastiere AA, Neuberg D, Taylor SG, DeConti R, and Adams G, “Phase II evaluation of taxol in advanced
head and neck cancer: an Eastern Cooperative Oncology Group Trial”, Journal of the National Cancer
Institute, Monographs,Vol. 15, 1993, pp. 181-184.
[117] Arbuck SG, Christian MC, Fisherman JS, Cazenave LA, Sarosy G, Suffness M, Adams J, Canetta R, Cole KE,
and Friedman MA, “Clinical development of Taxol”, Journal of the National Cancer Institute,
Monographs, Vol. 15, 1993, pp. 11–24.
[118] Roth BJ, Yep BY, Wilding G, Kasimes B, McLeod D, and Loehrer PJ,“Taxol in advanced hormone refractory
carcinoma of the prostrate: a phase II trial of the Eastern Cooperative Oncology Group”, Cancer, Vol. 72,
1993, pp. 2457.
[119] Brown T, Tangen C, Flemming T, and Macdonald J , “A phase II trial of taxol and granulocyte colony
stimulating factor (G-CSF) in patients with adenocarcinoma of pancreas”, Proceedings of American
Society of Clinical Oncology 12 (abstracts), 1993, p. 200.
[120] Woo DDl, Miao SYP, Pelayo JC, and Woolf AS “Taxol inhibits progression of congenital polycystic kidney
disease”, Nature, Vol. 368, 1994, pp. 750–753.
[121] Theodoridis G, Laskaris G, and Verpoorte R, “HPLC analysis of taxoids in plants and plant cell tissue
culture”, American Biotechnology Laboratory, Vol. 17, 1999, pp. 40–44.
[122] Skeel RT, “Handbook of Chemotherapy” (5th
ed.) Lippincott, Williams and Wilkins, Baltimore, 1999.
[123] Schiff PB, Fant J, Auster LA, and Horowitz SB, “Effects of taxol on cell growth and in vitro microtubule
assembly”, J Supramol Struct, Vol. 8, 1978, p. 328.
[124] Horowitz SB, Lothsteia L, Manfredi JJ, Mellado W, Parness J, Roy SN, Schiff PB, Sorbara L, and Zeheb R,
“Taxol: Mechanism of action and resistance”, Annals of the New York Academy of Sciences, Vol. 466,
1986, pp. 733-743.

e Journal of Biological Sciences www.ejarr.com
65
[125] Suffness M, and Cordell GA, “Antitumour alkaloids”, In: Brossi A (ed) The Alkaloids, vol XXV. Academic
Press Inc., Orlando, 1985, pp. 6-20.
[126] Gibson DM, Ketchum REB, Vance NC, and Christen AA, “Initiation and growth of cell lines of Taxus
brevifolia (Pacific Yew)”, Plant Cell Reports, Vol. 12, 1993, pp. 479–482.
[127] Cragg GM, Boyd MR, Cardellina II JH, Grever MR, Schepartz S, Snader KM, and Suffness M, “The search
for new pharmaceutical crops” In: Janick J and Simon JE (ed) Drug Discovery and Development at the
National Cancer Institute. New Crops, Wiley, New York, 1993, pp. 61-167.
[128] Flores HE, and Sgrignoli PJ, “In vitro culture and precocious germination of Taxus embryos”, In Vitro
Cellular and Developmental Biology, Vol. 27P, 1991, pp. 139–142.
[129] Banerjee S, Upadhyay N, Kukreja AK, Ahuja PS, Kumar S, Saha GC, Sharma RP, and Chattopadhyay SK,
“Taxanes from in vitro cultures of the Himalayan Yew Taxus wallichiana”, Planta Medica, Vol. 62, 1996,
pp. 333–335.
[130] Sohn H, and Okos MR, “Paclitaxel (Taxol): from Nutt to Drug”, Journal of Microbiology and Biotechnology,
Vol. 8, 1998, pp. 427–440.
[131] Kwak SS, Choi MS, Park YG, Yoo JS, and Liu JR, “Taxol content in the seeds of Taxus spp”,
Phytochemistry Vol. 40, 1995, pp. 29–32.
[132] Holton R, Somoza AC, Kim B, Liang F, Biediger J, Boatman PD, Shindo M, Smith CC, and Kim SC, “First
total synthesis of Taxol. 1. Functionalisation of the B ring”, Journal of American Chemical Society, Vol.
116, 1994, pp. 1597–1600.
[133] Nicolaou KC, Yang Z, Liu JJ, Ueno H, Nantermet PG, Guy RK, Clalborne CF, Renaud J, Couladouros EA,
Paulvannan K, and Sorensen EJ, “Total synthesis of taxol”, Nature, Vol. 367, 1994, pp. 630-634.
[134] Fett-Neto AG, DiCosmo F, Reynolds WF, and Sakata K, “Cell culture of Taxus as a source of the
antineoplastic drug taxol and related taxanes”, Biotechnology, Vol. 10, 1992, pp. 1572–1575.
[135] Wheeler NC, Jech K, and Masters S, “Effects of genetic, epigenetic and environmental factors on taxol
content in Taxus brevifolia and related species”, Journal of Natural Products, Vol. 55, 1992, pp. 432–440.
[136] Chang SH, and Yang JC, “Enhancement of plant formation from embryo cultures of Taxus mairei using
suitable culture medium and PVP”, Botanical Bulletin of Academia Sinica, Vol. 37, 1996, pp. 35–40.
[137] Chen Z, and Chen Y “Chemical constituents of Taxus yunnanensis Chen et L.K.Fu.” Chung Kuo Chung Yao
Tsa Chih, Vol. 21, 1996, pp. 230-232.
[138] Ellis DD, Zeldin EL, Brodhagen M, Russin WA, and McCrown BH, “Taxol production in nodule cultures of
Taxus”, Journal of Natural Products, Vol. 59, 1996, pp. 246-250.
[139] Gibson DM, Ketchum REB, Hirasuna TJ, and Shuler ML, “Potential of plant cell culture for taxane
production”, Taxol Science and Application, 1995, pp. 71-95.
[140] Hirasuna TJ, Pestchanker LJ, Srinivasan V, and Shuler ML, “Taxol production in suspension cultures of Taxus
baccata”, Plant Cell, Tissue and Organ Culture, Vol. 441996, pp. 95–102.
[141] Wang HQ, Zhong JJ, and Yu JT, “Enhanced production of taxol in suspension cultures of Taxus chinensis by
controlling inoculum size”. Biotechnology Letters, Vol. 19, 1997, pp. 353-355.
[142] Zhang X, Burt HM, Mangold G, Dexter D, Hoff von D, Mayer L, and Hunter WL, “Anti-tumour efficacy and
biodistribution of intravenous polymeric micellar Paclitaxel”, Anticancer Drugs , Vol. 8, 1997, pp. 696-
701.
[143] Witherup KM, Look SA, Stasko MW, Ghiorzi TJ, and Muschik GM, “Taxus spp. needles contain amounts of
taxol comparable to the bark of Taxus brevifolia: analysis and isolation”, Journal of Natural Products, Vol.
53, 1990, pp. 1249–1255.
[144] ElSohly HN, Croom EM, Kopycki WJ, Joshi AS, ElSohly MA, and McChesney JD, “Concentrations of taxol
and related taxanes in the needles of different Taxus cultivars”, Phytochemical Analysis, Vol. 6, 1995, pp.
149–156.
[145] Chattopadhyay SK, Saha GC, Sharma RP, Kumar S, and Roy R, “A rearranged taxane from the Himalayan
Yew Taxus wallichiana”, Phytochemistry Vol. 42, 1996, pp. 787–788.
[146] Furmanowa M, and Syklowska-Baranek K, “Hairy root cultures of Taxus x media var. Hicksii Rehd. as a new
source of paclitaxel and 10-deacetylbaccatin III” Biotechnology Letters, Vol. 22, 2000, pp. 683–686.
[147] Luo SD, Ning BM, Ruan DC, and Wang HY, “HPLC analysis of taxol and related compounds from Taxus and
its related plants”, Journal of Plant Resources and Environment, Vol. 3, 1994, pp. 31–33.
[148] Guénard D, Guéritte-Voegelein F, and Potier P, “Taxol and Taxotere: discovery, chemistry and structure-
activity relationships”, Accounts of Chemical Research, Vol. 26, 1993, pp. 160–167.
[149] Kingston DGI, Molinero AA, and Rimoldi JM, “The Taxane Diterpenoids” Springer-Verlag, New York,
1993.

e Journal of Biological Sciences www.ejarr.com
66
[150] George GI, Ali S, Zygmut J, and Jayasinghe LR, “Taxol: a novel antitumour agent”, Expert Opinion on
Therapeutic Patents, Vol. 4, 1994, pp. 109–120.
[151] Commercon A, Bourzat JD, Didier E, and Lavelle F, “Practical semisynthesis and antimitotic activity of
docetaxel and side chain analogs”, ACS Symposium Series, Vol. 583, 1995, pp. 233–246.
[152] Shrestha K, Strobel GA, Shrivastava SP, and Gewali MB, “Evidence for paclitaxel from three new endophytic
fungi of Himalayan yew of Nepal”, Planta Medica, Vol. 67, 2001, pp. 374–376.
[153] Su X, Xingguo M, and Wei G, “Study of paclitaxel production from an antifungal variant of Taxus callus”,
Plant Cell, Tissue and Organ Culture, Vol. 68, 2002, pp. 215–223.
[154] Qiu DY, “Isolation of an enophytic fungus associated with Taxus yunnanensis”, Acta-Mycologica-Sinica, Vol.
13, 1994, pp. 314–316.
[155] Li JY, Sidhu RS, Bollon A, and Gary A, “Stimulation of taxol production in liquid cultures of Pestalotiopsis
microspora”, Mycological Research, Vol. 102, 1998, pp. 461–464.
[156] Yang X, Strobel G, Stierle A, Hess WM, Lee J, and Clardy J, “A fungal endophyte-tree relationship: Phoma
sp. in Taxus wallichiana”, Plant Science, Vol. 102, 1994, pp. 1–9.
[157] Zhang P, Zhou PP, Jiang, Yu H, and Yu LJ “Screening of taxol-producing fungi based on PCR amplification
from Taxus”, Biotechnol Letters, Vol. 30, 2008, pp. 2119–2123.
[158] Page M, and Landry N, “Bacterial mass production of taxanes with Erwinia”, US Patent 5,561,055, 1 Oct
1996
[159] Baloglu E, and Kingston DGI, “The Taxane Diterpenoids”, Journal of Natural Products, Vol. 62, 1999,
pp.1448–1472.
[160] Wildung MR, and Croteau R “A cDNA clone for taxadiene synthase, the diterpene cyclase that catalyses the
committed step of taxol biosynthesis”, Journal of Biological Chemistry, Vol. 271, 1996, pp. 9201–9204.
[161] Hezari M, and Croteau R, “Taxol biosynthesis: an update”, Planta Medica, Vol. 63, 1997, pp. 291–295.
[162] Walker K, and Croteau R, “Taxol biosynthetic genes”, Phytochemistry, Vol. 58, 2001, pp. 1–7.
[163] Walker K, Long R, and Croteau R “The final acetylation step in taxol biosynthesis: cloning of the taxoid-C13-
side chain N-benzoyl transferase from Taxus”, Proceedings of the National Academy of Sciences of the
United States of America, Vol. 99, 2002a, pp. 9966–9971.
[164] Walker K, Fujisaki S, Long R, and Croteau R “Molecular cloning and heterologous expression of the C13
phenylpropanoid side chain-CoA acyltransferase that functions in taxol biosynthesis”, Proceedings of the
National Academy of Sciences of the United States of America, Vol. 99, 2002b, pp. 12715–12720.
[165] Suffness M, “Taxol: Science and Applications”, CRC Press, Boca Raton, Florida, 1995.
[166] Bruce SE, and Rowe DB, “Using chlorophyll fluorescence in storage and vegetative propagation of Taxus”,
HortScience, Vol. 35, 2000, pp. 544–549.
[167] Schenk V, “Studies on the influence of clone on rooting ability and quality in the propagation of cuttings from
40-50 year old yews (T. baccata L.)” Silvae Genetica, Vol. 45, 1997, pp. 246–249.
[168] Mitter H, and Sharma A “Propagation of T baccata Linn. by stem cuttings”, Indian Forester, Vol. 125, 1999,
pp. 159–162.
[169] Saini RP, “Vegetative propagation in Silviculture (Hills) Division, Darjeeling (West Bengal)”, Indian
Forester, Vol. 127, 2001, pp. 389–408.
[170] Dubey KP, “Himalayan Yew (T. baccata) conservation: A vegetative approach”, Indian Forester, Vol. 123,
1997, pp. 1150–1154.
[171] Maheshwari P, Garg S, and Kumar A, “Taxoids: Biosynthesis and in vitro production”, Biotechnology and
Molecular Biology Reviews, Vol. 3, 2008, pp. 71–087.
[172] Chee PP, “In vitro culture of zygotic embryos of Taxus species”, HortScience, Vol. 29, 1994, pp. 695–697.
[173] Ho RH, “Embryo Culture”, In JM Bonga and DJ Durzan (ed) Cell and Tissue Culture in Forestry, vol. 2.
Specific Principles and Methods: Growth and Development. Martinus Nijhoff Publishers, Dordercht, 1987,
pp. 37-167.
[174] Flores T, Wagner LJ, Flores HE “Embryo culture and taxane production in Taxus spp”, In Vitro Cellular and
Developmental Biology, Vol. 29P, 1993, pp. 160–165.
[175] Chang SH, and Yang JC, “Enhancement of plant formation from embryo cultures of Taxus mairei using
suitable culture medium and PVP”, Botanical Bulletin of Academia Sinica, Vol. 37, 1996, pp. 35-40.
[176] Choi MS, “In vitro germination and propagation by embryo culture of Taxus cuspidata for taxol production”,
Journal of Plant Biotechnology, Vol. 2, 2000, pp. 29–33.
[177] Datta MM, and Jha S, “Embryo culture of Taxus wallichiana (Zucc.)”, Journal of Plant Biotechnology, Vol. 6,
2004, pp. 213–220.

e Journal of Biological Sciences www.ejarr.com
67
[178] Chee PP, “Organogenesis in Taxus brevifolia tissue cultures”, Plant Cell Reports, Vol. 14, 1995a, pp. 560–
565.
[179] Xie D, and Guo ZC, “Zygotic embryo culture of Taxus chinensis var. mairei and plant regeneration through
organogenesis”, Israel Journal of Plant Sciences, Vol. 47, 1999, pp. 287–290.
[180] Chee PP, “Plant regeneration from somatic embryos of Taxus brevifolia”, Plant Cell Reports, Vol. 16, 1996,
pp. 184–187.
[181] Datta MM, Majumder A, and Jha S, “Organogenesis and plant regeneration in Taxus wallichiana (Zucc.)”,
Plant Cell Reports, Vol. 25, 2006, pp. 11–18.
[182] Datta MM, and Jha S, “Plant regeneration through somatic embryogenesis in Taxus wallichiana”, Journal of
Plant Biochemistry and Biotechnology Vol. 17, 2008, pp. 37–44.
[183] Rohr R, “Production de cals par les gemetophyte males de Taxus baccata L. Cultivars sur un milieu
artifigicial. Etude en microscopie photonique et electronique.” Caryologia, Vol. 25, 1973, pp. 177–189.
[184] Christen AA, Bland J, and Gibson DM, “Cell culture as a means to produce taxol”, Proceedings of the
American Association for Cancer Research, Vol. 30, 1989, pp. 566.
[185] Mirajalili N, and Linden JC, “Methyl jasmonate induced production of taxol in suspension cultures of Taxus
cuspidata: ethylene interaction and induction models”, Biotechnology Progress, Vol. 12, 1996, pp. 110–
118.
[186] Fett-Neto AG, Melanson SJ, Sakata K, and Dicosmo F, “Improved growth and taxol yield in developing calli
of Taxus cuspidata by medium composition modification”, Biotechnology, Vol. 11, 1993, pp. 731–734.
[187] Wickremesinhe ERM, and Arteca RN, “Taxus callus cultures: Initiation, growth optimization, characterization
and taxol production”, Plant Cell, Tissue and Organ Culture, Vol. 35, 1993, pp. 181–193.
[188] Wickremesinhe ERM, and Arteca RN, “Taxus cell suspension cultures: optimizing growth and production of
taxol”, Journal of Plant Physiology, Vol. 144, 1994, pp. 183–188.
[189] Ghosh B, Mukherjee S, Jha TB, and Jha S, “Taxol production in untransformed cell cultures of Taxus
wallichiana”, In: Nandi SK, Palni LMS, Kumar A (ed) Role of plant tissue culture in biodiversity
conservation and economic development, Gyanodaya Prakashan, Nainital, India, 2002, pp. 287–295.
[190] Mirjalili N, and Linden JC “Gas phase composition effects on suspension cultures of Taxus cuspidata”,
Biotechnology and Bioengineering, Vol. 48, 1995, pp. 123–132.
[191] Choi HK, Kim SI, Son JS, Hong SS, Lee HS, and Lee HJ, “Enhancement of paclitaxel production by
temperature shift in suspension culture of Taxus chinensis”, Enzyme and Microbial Technology, Vol. 27,
2000b, pp. 593–598.
[192] Kim SI, Choi HK, Kim JH, Lee HS, and Hong SS, “Effect of osmotic pressure on paclitaxel production in
suspension cell cultures of Taxus chinensis”, Enzyme and Microbial Technology, Vol. 28, 2001, pp. 202–
209.
[193] Christen AA, Gibson DM, and Bland J, “Production of taxol or taxol-like compounds with Taxus brevifolia
callus culture”, US Patent 5,019,504, 28 May 1991.
[194] Ciddi V, Srinivasan V, and Shuler ML, “Elicitation of Taxus sp. cell cultures for production of taxol”,
Biotechnology Letters, Vol. 17, 1995, pp. 1343–1346.
[195] Wang C, Wu J, and Mei X “Enhancement of taxol production and excretion in Taxus chinensis cell culture by
fungal elicitation and medium renewal”, Applied Microbiology and Biotechnology, Vol. 55, 2001, pp. 404–
410.
[196] Cusido RM, Palazon J, Bonfill M, Navia-Osorio A, Morales C, and Pinol MT, “Improved paclitaxel and
baccatin III production in suspension cultures of Taxus media”, Biotechnology Progress, Vol. 18, 2002, pp.
418–423.
[197] Zhang CH, and Wu JY, “Ethylene inhibitors enhance elicitor-induced paclitaxel production in suspension
cultures of Taxus spp. Cells”, Enzyme and Microbial Technology, Vol. 32, 2003, pp. 71–77.
[198] Yu LJ, Lan WZ, Qin WM, and Xu HB, “Effects of salicylic acid on fungal-elicitor induced membrane-lipid
peroxidation and taxol production in cell suspension cultures of Taxus chinensis”, Process Biochemistry,
Vol. 37, 2001, pp. 477–482.
[199] Srinivasan V, Ciddi V, Bringi V, and Shuler ML “Metabolic inhibitors, elicitors and precursors as tools for
probing yield limitation in taxane production by Taxus chinensis cell cultures”, Biotechnology Progress,
Vol. 12, 1996, pp. 457–465.
[200] Zhang CH, Mei XG, Liu L, and Yu LG, “Enhanced paclitaxel production induced by the combination of
elicitors in cell suspension cultures of Taxus chinensis”, Biotechnology Letters, Vol. 22, 2000, pp. 1561–
1564.

e Journal of Biological Sciences www.ejarr.com
68
[201] Linden JC, and Phisalaphong M, “Oligosaccharides potentiate methyl jasmonate-induced production of
paclitaxel in Taxus Canadensis”, Plant Science, Vol. 158, 2000, pp. 41–51.
[202] Wu JY, Wang CG, and Mei XG, “Stimulation of taxol production and excretion in Taxus spp. cell cultures by
rare earth chemical lanthanum”, Journal of Biotechnology, Vol. 85, 2001, pp. 67–73.
[203] Zang X, Mei XG, Zhang CH, Lu, CT, and Ke T, “Improved paclitaxel accumulation in cell suspension
cultures of Taxus chinensis by brassinolide”, Biotechnology Letters, Vol. 23, 2001, pp. 1047–1049.
[204] Clinton CS, Erik BGF, Angela H, Abbey H, and Amanda K, “„Hicksii‟ yews as a sustainable source of
anticancer compounds”http://www.cropinfo.net/AnnualReports/1997/yew.alt.html, 1998
[205] Luo J, Liu L, and Wu CD, “Enhancement of paclitaxel production by abscisic acid in cell suspension cultures
of Taxus chinensis”, Biotechnology Letters, Vol. 23, 2001, pp.1345–1348.
[206] Fett-Neto AG, Melanson SJ, Nicholson SA, Pennington JJ, and Dicosmo F, “Improved taxol yield by aromatic
carboxylic acid and amino acid feeding to cell cultures of Taxus cuspidata”, Biotechnology and
Bioengineering, Vol. 44, 1994, pp. 967–971.
[207] Jha S, Sanyal D, Ghosh B, and Jha TB “Improved taxol yield in cell suspension culture of Taxus wallichiana
(Himalayan yew)”, Planta Medica, Vol. 64, 1998, pp. 270–272.
[208] Veeresham C, Mamatha R, Babu CP, Srisilam K, and Kokate CK, “Production of taxol and its analogues from
cell cultures of Taxus wallichiana”, Pharmaceutical Biology, Vol. 41, 2003, pp. 426–430.
[209] Expósito O, Bonfill M, Onrubia M, Jané A, Moyano E, Cusidó RM, Palazón J, and M. Teresa Piñol, “Effect of
taxol feeding on taxol and related taxane production in Taxus baccata suspension cultures”, New
Biotechnology,Vol. 25, 2009, pp. 252–259.
[210] Li YC, Tao WY, and Cheng L, “Paclitaxel production using co-culture of Taxus suspension cells and
paclitaxel-producing endophytic fungi in a co-bioreactor”, Applied Microbiology and Biotechnology, Vol.
83, 2009, pp. 233–239.
[211] Han KH, Fleming P, Walker K, Loper M, Chilton WS, Mocek U, Gordon MP, and Floss HG, “Genetic
transformation of mature Taxus: an approach to genetically control the in vitro production of the anticancer
drug, taxol”, Plant Science, Vol. 95, 1994, pp.187–196.
[212] Furmanowa M, and Syklowska-Baranek K, “Hairy root cultures of Taxus x media var. Hicksii Rehd. as a new
source of paclitaxel and 10-deacetylbaccatin III”, Biotechnology Letters, Vol. 22, 2000, pp. 683-686.
[213] Zhong JJ, Wang SJ, and Wang HQ, “A new centrifugal impeller reactor and its application to plant cell
cultures”, In: Horizon of Biochemical Engineering. The Society of Chemical Engineers, Japan, Tokyo,
1997, pp. 175–179.
[214] Srinivasan V, Pestchanker L, Moser S, Hirasuna TJ, Taticek RA, and Shuler ML, “Taxol production in
bioreactors: kinetics of biomass accumulation, nutrient uptake and taxol production by cell suspensions of
Taxus baccata”, Biotechnology and Bioengineering, Vol. 47, 1995, pp. 666–676.
[215] Navia-Osorio A, Garden H, Cusido RM, Palazón J, Alfermann AW, and Piñol MT, “Taxol (R) and baccatin
III production in suspension cultures of Taxus baccata and Taxus wallichiana in an airlift bioreactor”,
Journal of Plant Physiology, Vol. 159, 2002, pp. 97–102.
[216] Son SH, Choi SM, Lee YH, Choi KB, Yun SR, Kim JK, Park HJ, Kwon OW, Noh EW, Seon JH, and Park
YG, “Large-scale growth and taxane production in cell cultures of Taxus cuspidata (Japanese yew) using a
novel bioreactor”, Plant Cell Reports, Vol. 19, 2000, pp. 628–633.
[217] Yukimune Y, Hara Y, Nomura E, Seto H, and Yoshida S, “The configuration of methyl jasmonate affects
paclitaxel and baccatin III production in Taxus cells”, Phytochemistry Vol. 54, 2000, pp. 13–17.
[218] Choi HK, Son JS, Na GH et al “Mass production of paclitaxel by plant cell culture”, Korean J Plant
Biotechnol Vol. 29, 2002, pp. 59–62.
[219] Giri A, Giri CC, Dhingra V, and Narasu ML, “Podophyllum hexandrum Royle: a potential source for
production of clinically useful anticancer drugs”, In: Role of biotechnology in medicinal and aromatic
plants, Vol. 4. Ukaaz publication, India, 2001b , pp. 80–88.
[220] Moss GP, “Nomenclature of lignans and neolignans”, Pure and Applied Chemistry, Vol. 72, 2000, pp. 1493–
1523.
[221] Smollny T, Wichers H, Kalenberg S, Shahsavari A, Petersen M, and Alfermann AW, “Accumulation of
podophyllotoxin and related lignans in cell suspension cultures of Linum album”, Phytochemistry, Vol. 48,
1998, pp. 575–579.
[222] Konuklugil B, Schimdt TJ, and Alfermann AW, “Accumulation of aryltetralin lactone lignans in cell
suspension cultures of Linum nodiflorum”, Planta Medica, Vol. 65, 1999, pp. 587–588.

e Journal of Biological Sciences www.ejarr.com
69
[223] Konuklugil B, Schimdt TJ, and Alfermann AW, “Accumulation of lignans in suspension cultures of Linum
mucronatum spp. armenum (Bordz.) Davis”, Zeitschrift für Naturforschung, Vol. 56c, 2001, pp. 1164–
1165.
[224] Chattopadhyay S, Srivastava AK, Bhojwani SS, and Bisaria VS, “Development of suspension culture of
Podophyllum hexandrum for the production of podophyllotoxin”, Biotechnology Letters, Vol. 23, 2001, pp.
2063–2066.
[225] Kutney JP, Arimoto H, Hewitt GM, Jarvis TC, and Sakata K, “Studies with plant cell cultures of Podophyllum
peltatum L. I. Production of podophyllotoxin, deoxypodophyllotoxin, podophyllotoxone and 4'-
demethylpodophyllotoxin”, Heterocycles, Vol. 32, 1991, pp. 2305–2309.
[226] Kutney JP, Chen YP, Gao S, Hewitt GM, Kuri-Brena F, Milanova RK, and Stoynov NM, “Studies with plant
cell cultures of Podophyllum peltatum L. II. Biotransformation of dibenzylbutanolides to lignans.
Development of a „biological factory‟ for lignan synthesis”, Heterocycles Vol. 36, 1993, pp. 13–20. [227] Choi HK, Kim SI, Son JS, Hong SS, Lee HS, Chung IS, and Lee HJ, “Intermittent maltose feeding enhances
paclitaxel production in suspension culture of Taxus chinensis cells”, Biotechnology Letters, Vol. 22,
2000a, pp. 1793–1796.
[228] Linden JC, Haigh JR, Mirjalili N, and Phisaphalong M, “Gas concentration effects on secondary metabolite
production by plant cell cultures”, Advanced Biochemical Engineering/Biotechnology, Vol. 72, 2001, pp.
28–62.
[229] Bringi V, and Kadkade PG, “Enhanced production of taxol and taxanes by cell cultures of Taxus species”, WO
Patent No. 93/17121, 1993.


















