
Biome reconstruction from pollen and plant macrofossil ...
21
Journal of Biogeography (1998) 25, 1007–1027 Biome reconstruction from pollen and plant macrofossil data for Africa and the Arabian peninsula at 0 and 6000 years D J 1 * , I. C P 1 ,R B 2 ,A B 3 ,M B 4 , P B 4 ,G B 2 ,D B 5 ,J -P C 6 ,R C 7 , T E 8 ,H E 2 ,S E 2 ,J G 7 ,F L 9 ,H L 10 , A -M L 6 ,J M 4 ,M M 11 ,O P 7 ,M R 7 , I R -F 4 ,G R 2 ,J C. R 12 ,E R 13 ,L S 14 , I S 15 ,H S 16 ,M U 17 ,E V C 18 , S V 19 ,A V 2 and M W 20 1 Global Systems Group, Department of Plant Ecology, Lund University, Ecology Building, So ¨lvegatan 37, S-223 62 Lund, Sweden; 2 Laboratoire de Ge ´ologie du Quaternaire, CNRS, CEREGE, BP80, 13545 Aix-en-Provence, Cedex 04, France; 3 Universite ´ d’Angers, Departement de Ge ´ographie, 35 Rue de la Barre, F-49000 Angers, France; 4 Laboratoire de Palynologie, U.S.T.L., 34095 Montpellier Cedex 5, France; 5 Department of Biological Sciences, Fordham University, Bronx, New York, 10458 U.S.A.; 6 De ´partement de Ge ´ologie Se ´dimentaire, Universite ´ Pierre et Marie Curie, 4 Place Jussieu, F-75252 Paris cedex 5, France; 7 Laboratoire de Botanique Historique et Palynologie, Case 451, Faculte ´ des Sciences de St Je ´rome, CNRS UA 1152, F-13397 Marseille Cedex 13, France; 8 F.D.S., U.B., De ´partement de Botanique, BP1515, Lome ´, Togo; 9 Dynamic Palaeoclimatology Group, Lund University, Box 117, S-221 00 Lund, Sweden; 10 Department of Geography, University of Wales, Penglais, Aberystwyth, DYFED, SY23 3DB Wales, U.K.; 11 UNILU, De ´partement de Ge ´ographie, BP1825, Lubumbashi, Zaı ¨re; 12 Pebbledash Cottage, Corfe, Taunton, Somerset, TA3 7AJ England, U.K.; 13 Laboratoire de Palynologie, Muse ´e Royal de l’Afrique Centrale, B-3080, Tervuren, Belgique; 14 Botany Department, University of Orange Free State, PO Box 339, Bloemfontein, 9301, Republic of South Africa; 15 Department of Geology, Makerere University, PO Box 7062, Kampala, Uganda; 16 Botanisches Institut, Neue Universita ¨t, Olshausenstrasse 40–60, D-24098 Kiel, Germany; 17 Geology Department, Addis Ababa University, PO Box 3434, Addis Ababa, Ethiopia; 18 Laboratoire d’Ecologie Terrestre, UMR 5552 (CNRS-UPS), 13 Avenue du Colonel Roche, BP 4403, 31405 Toulouse cedex 4, France; 19 Pale ´ontologie, Universite ´ de Lie `ge, Place du XX Aout, 4000 Lie `ge, Belgium; 20 School of Geography, Kingston University, Penrhyn Road, Kingston upon Thames, Surrey, KT1 2EE England. Abstract. Biome reconstruction from pollen and plant Madagascar, eastern, southern and central Africa show only macrofossil data provides an objective method to minor changes in terms of biomes, compared to present. reconstruct past vegetation. Biomes for Africa and the Major changes in biome distributions occur north of 15°N, Arabian peninsula have been mapped for 6000 years and with steppe in many low-elevation sites that are now desert, provide a new standard for the evaluation of simulated and temperate xerophytic woods/scrub and warm mixed palaeovegetation distributions. A test using modern pollen forest in the Saharan mountains. These shifts in biome data shows the robustness of the biomization method, which distributions imply significant changes in climate, especially is able to predict the major vegetation types with a high precipitation, between 6000 years and present, reflecting a confidence level. The application of the procedure to the change in monsoon extent combined with a southward 6000 years data set (pollen and plant macrofossil analyses) expansion of Mediterranean influence. shows systematic di erences from the present that are Key words. Biome, plant functional type, pollen, plant consistent with the numerous previous regional and macrofossil, Africa, Arabian peninsula, Madagascar, continental interpretations, while providing a more extensive and more objective basis for such interpretations. 6000 years . the outputs from Atmospheric General Circulation Models INTRODUCTION (AGCMs) into maps of potential vegetation distribution Global vegetation models (e.g. Prentice et al., 1992; for present (Claussen & Esch, 1994; Foley et al., 1996), past Haxeltine & Prentice, 1996) provide a way to translate (Prentice et al., 1993; Claussen, 1994; Foley, 1994; Harrison et al., 1995; de Noblet et al., 1996; Kutzbach et al., 1996; Correspondence: Dr Dominique Jolly, Department of Plant Ecology, Lund TEMPO Members, 1996), and future climate scenarios University, Ecology Building, So ¨lvegatan 37, S-223 62 Lund, Sweden; (Prentice & Sykes, 1994; VEMAP Members, 1995; Mellilo e-mail: [email protected]. 1998 Blackwell Science Ltd 1007
Transcript of Biome reconstruction from pollen and plant macrofossil ...

Journal of Biogeography (1998) 25, 1007–1027
Biome reconstruction from pollen and plant macrofossil data forAfrica and the Arabian peninsula at 0 and 6000 years
D J1∗ , I. C P1, R B2, A B3, M B4,P B4, G B2, D B5, J-P C6, R C7,T E8, H E2, S E2, J G7, F L9, H L10,A-M L6, J M4, M M11, O P7, M R7,I R-F4, G R 2, J C. R 12, E R13, L S14,I S15, H S16, M U17, E V C18,S V19, A V2 and M W20 1Global Systems Group, Department of PlantEcology, Lund University, Ecology Building, Solvegatan 37, S-223 62 Lund, Sweden; 2Laboratoire de Geologie duQuaternaire, CNRS, CEREGE, BP80, 13545 Aix-en-Provence, Cedex 04, France; 3Universite d’Angers, Departement deGeographie, 35 Rue de la Barre, F-49000 Angers, France; 4Laboratoire de Palynologie, U.S.T.L., 34095 Montpellier Cedex 5,France; 5Department of Biological Sciences, Fordham University, Bronx, New York, 10458 U.S.A.; 6Departement de GeologieSedimentaire, Universite Pierre et Marie Curie, 4 Place Jussieu, F-75252 Paris cedex 5, France; 7Laboratoire de BotaniqueHistorique et Palynologie, Case 451, Faculte des Sciences de St Jerome, CNRS UA 1152, F-13397 Marseille Cedex 13,France; 8F.D.S., U.B., Departement de Botanique, BP1515, Lome, Togo; 9Dynamic Palaeoclimatology Group, LundUniversity, Box 117, S-221 00 Lund, Sweden; 10Department of Geography, University of Wales, Penglais, Aberystwyth,DYFED, SY23 3DB Wales, U.K.; 11UNILU, Departement de Geographie, BP1825, Lubumbashi, Zaıre; 12PebbledashCottage, Corfe, Taunton, Somerset, TA3 7AJ England, U.K.; 13Laboratoire de Palynologie, Musee Royal de l’AfriqueCentrale, B-3080, Tervuren, Belgique; 14Botany Department, University of Orange Free State, PO Box 339, Bloemfontein,9301, Republic of South Africa; 15Department of Geology, Makerere University, PO Box 7062, Kampala, Uganda;16Botanisches Institut, Neue Universitat, Olshausenstrasse 40–60, D-24098 Kiel, Germany; 17Geology Department, AddisAbaba University, PO Box 3434, Addis Ababa, Ethiopia; 18Laboratoire d’Ecologie Terrestre, UMR 5552 (CNRS-UPS), 13Avenue du Colonel Roche, BP 4403, 31 405 Toulouse cedex 4, France; 19Paleontologie, Universite de Liege, Place du XXAout, 4000 Liege, Belgium; 20School of Geography, Kingston University, Penrhyn Road, Kingston upon Thames, Surrey, KT12EE England.
Abstract. Biome reconstruction from pollen and plant Madagascar, eastern, southern and central Africa show onlymacrofossil data provides an objective method to minor changes in terms of biomes, compared to present.reconstruct past vegetation. Biomes for Africa and the Major changes in biome distributions occur north of 15°N,Arabian peninsula have been mapped for 6000 years and with steppe in many low-elevation sites that are now desert,provide a new standard for the evaluation of simulated and temperate xerophytic woods/scrub and warm mixedpalaeovegetation distributions. A test using modern pollen forest in the Saharan mountains. These shifts in biomedata shows the robustness of the biomization method, which distributions imply significant changes in climate, especiallyis able to predict the major vegetation types with a high precipitation, between 6000 years and present, reflecting aconfidence level. The application of the procedure to the change in monsoon extent combined with a southward6000 years data set (pollen and plant macrofossil analyses) expansion of Mediterranean influence.shows systematic differences from the present that are
Key words. Biome, plant functional type, pollen, plantconsistent with the numerous previous regional andmacrofossil, Africa, Arabian peninsula, Madagascar,continental interpretations, while providing a more extensive
and more objective basis for such interpretations. 6000 years .
the outputs from Atmospheric General Circulation ModelsINTRODUCTION(AGCMs) into maps of potential vegetation distributionGlobal vegetation models (e.g. Prentice et al., 1992;for present (Claussen & Esch, 1994; Foley et al., 1996), pastHaxeltine & Prentice, 1996) provide a way to translate(Prentice et al., 1993; Claussen, 1994; Foley, 1994; Harrisonet al., 1995; de Noblet et al., 1996; Kutzbach et al., 1996;
Correspondence: Dr Dominique Jolly, Department of Plant Ecology, Lund TEMPO Members, 1996), and future climate scenariosUniversity, Ecology Building, Solvegatan 37, S-223 62 Lund, Sweden; (Prentice & Sykes, 1994; VEMAP Members, 1995; Melliloe-mail: [email protected].
1998 Blackwell Science Ltd 1007

1008 Dominique Jolly et al.
et al., 1996). In order to check the ability of the AGCMs DATAto simulate the changes in regional climates under different
Modern pollen databoundary conditions for key periods in the past (COHMAP,1988; Wright et al., 1993), the palaeodata have to be A modern pollen data set including 966 samples wascompiled in a global data set in a uniform manner for a compiled for Africa, south of the Sahara, and for thedirect comparison with the outputs of coupled atmosphere- Arabian peninsula. All the data are on file as raw pollenterrestrial biosphere models. This is the primary aim of the counts. From this data set, 269 samples (West and Centralinternational project BIOME 6000 (Prentice & Webb, in Africa) have already been used to test the biomizationpress). Associated with the data synthesis effort, an objective method at a subcontinental scale (Jolly et al., in press),method to translate the palaeovegetation data in terms of including sixty-six samples from Senegal, five frombiomes is required in order to make data-model comparisons Mauritania and three from Mali (Lezine, 1987), 125 samplesin a direct sense (Prentice & Jolly, in press). from Togo (Edorh, 1986), six from Chad (Maley, 1981),
An ‘objective biomization’ method, based on the concept four from Ghana (Maley & Livingstone, 1983; Maley,of assigning taxa to one or more plant functional types, unpublished), sixteen from Gabon (Jolly et al., 1996),was initially developed for Europe (Prentice et al., 1996) twenty-seven from Congo (Elenga, 1992), thirteen fromallowing the first attempt at such a data-model comparison Cameroon (Maley & Brenac, 1987; Brenac, 1988; Brenac &for the European continent at 6000 14C years (Prentice Maley, unpublished) and four from Nigeria (Agwu, 1986).et al., 1998). A similar approach was used in a preliminary To these data we now add 101 pollen samples from Southattempt for Africa, but here the palaeovegetation data set Africa and Namibia (Scott, 1982, 1987a, 1989; unpublished;was not yet available and a more subjective assignment Cooremans, 1989; Scott & Bousman, 1990; Scott &procedure had to be used (Jolly et al., 1998). Even this Cooremans, 1990, 1992; Scott & Brink, 1991), eighty-ninerepresented a clear improvement compared to standard samples collected in Madagascar and in the Mascarenesqualitative data-model comparisons (COHMAP, 1988; (Straka, 1991), three collected in Qatar (Bonnefille & Riollet,Street-Perrott & Perrott, 1993) because it allowed the 1988a), five in Yemen (Lezine, unpublished), fifty-one inmagnitude as well as the direction of the simulated climate Jordan, Iraq, Kuwait and Saudi Arabia (El-Moslimany,changes to be evaluated. These data-model comparison 1983). To this compilation we have added 451 modernexercises for Europe and Africa gave for the first time an pollen spectra included in the East African pollen data setobjective and precise diagnostic for the performance of an (ninety-four pollen taxa) used to reconstructAGCM (CCM1; Kutzbach et al., 1998) when used to palaeotemperatures and precipitation in East Africasimulate palaeoclimate. (Bonnefille et al., 1992; Vincens et al., 1993).
In order to extend the application of the objectivebiomization method to tropical areas, a preliminary test of
6000 year pollen and plant macrofossil datathe method was performed on a modern pollen data setfor West and Central Africa (Jolly et al., in press). The We compiled a data set of ninety-nine pollen records forbiomization procedure was able to predict the potential 6000±500 14C years ago. These data come from differentnatural vegetation in a situation with far higher taxonomic sources: eighty-five pollen spectra are primary pollen countsdiversity than Europe and with extreme differences of pollen from published or unpublished sources while fourteen pollenproduction among anemophilous and entomophilous spectra were digitized from published pollen diagramsplants. These results demonstrate the applicability of the (Table 1). We compiled a separate plant macrofossil databiomization method in tropical areas. set of six charcoal (carbonized plant fragment) records for
The African continent is a major piece of the global 6000 years, which provide a valuable complement to pollenpuzzle because it lies between 30°S and 30°N, and because data in the most arid regions. Pollen percentages for all thethe vegetation distribution is forced by two gradients (one data (modern and fossil) were calculated on the basis of thein latitude, and one in elevation for East Africa) giving the total sum minus the pollen and spores of (1) aquatic taxa,possibility to independently estimate changes in temperature such as Cyperaceae, Nymphaea or Rhizophora, (2) exoticand precipitation amount or seasonality. Africa is also a taxa (Casuarina equisetifolia, Tectona grandis . . .), and (3)key region to study changes in monsoon strength and taxa present only once with one pollen grain (e.g. Homalium,extension through time. The 6000 year time slice has been Ammannia∗ and Hernandiaceae). The number of taxa usedadopted as being a key period to study the influence of for the biome reconstructions is much greater [364] thanchanges in the seasonal distribution of the insolation due was used in Europe [41] (Prentice et al., 1996), in the Formerto changes in the earth’s orbital parameters (Kutzbach & Soviet Union and Mongolia [94] (Tarasov et al., 1998, thisWebb, 1993) and a current major focus of the palaeoclimate issue), and in the initial biome reconstruction made formodelling community (COHMAP, 1988; Joussaume & China [68] (Yu et al., 1998, this issue). This differenceTaylor, 1995; TEMPO Members, 1996). reflects the very high taxonomic diversity of the tropical
The aims of this paper are (1) to present the results of and subtropical African flora.the compilation of modern and 6000 year data sets forAfrica, (2) to test the biomization scheme at the scale of
METHODthe entire African continent (including Arabian peninsula),and (3) to reconstruct biome distribution at the discrete The ‘biomization’ (objective biome assignment) method for
pollen data was developed and applied to modern and 6000locations of the palaeoecological data sites at 6000 years.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1009
TABLE 1. Summary of pollen and plant macrofossil records at 6000 years for Africa and the Arabian peninsula.
Site Name Country Latitude Longitude Elevation D.C. Biome Reference(dec.) (dec.) (m) 6000 years 6000 years
La chataigneraie Algeria 36.80 4.57 1225 2D WAMF Salamani (1991, 1993)Sidi Bou Rhaba Morocco 34.25 −6.67 0 2D XERO Reille (1997)Tigalmamine Morocco 32.90 −5.35 1626 1C WAMF Lamb et al. (1989, 1995)Tighaslant Morocco 31.43 −7.47 2197 1C∗ XERO Bernard & Reille (1987)Saoura Algeria 30.08 −2.15 ≈500 2D STEP Beucher (1971)Izriten Morocco 28.50 −11.80 ≈120 1D STEP Beucher (1979)Khor Qatar 26.00 51.50 ≈10 2D STEP Bonnefille & Roillet
(1988a)Ti-n tora Lybia 25.80 10.58 ≈750 n/a STEP Schulz, unpublishedAbu Minqar (c) Egypt 25.77 27.80 ≈250 1D DESE Neumann (1989b)Uan Muhuggiag Lybia 24.83 10.48 ≈1000 n/a STEP Schulz, unpublishedTibesti Chad 23.50 17.50 ≈500 3D STEP Schulz, unpublishedWadi el Akhdar (c) Egypt 23.20 26.02 ≈1100 2D DESE Neumann (1989b)Wadi Bakht (c) Egypt 23.20 26.28 ≈950 1D STEP Neumann (1989b)Taoudenni Mali 22.50 –4.00 120 2D STEP Cour & Duzer (1976)Agorgott Mali 22.47 –4.03 133 3D TXWS Schulz (1991)Mouskorbe Chad 22.37 18.53 2600 n/a STEP Maley (1981, 1983)Enneri Chad 21.50 17.13 1100 1D WAMF Schulz (in Gabriel, 1977),
Schulz (1980)Selima Sudan 21.37 29.32 200 1D STEP Ritchie & Haynes (1987)Morzouba Mauritania 21.00 –16.20 ≈10 1D STEP Beucher (1979)Burg et Tuyur (c) Sudan 20.92 27.68 ≈450 1D STEP Neumann (1989b)Wadi Shaw (c) Sudan 20.53 27.50 ≈500 2D STEP Neumann (1989b)Tjolumi Chad 20.52 17.18 (1900 1D XERO Schulz (in Gabriel, 1977)Rub’al Khali Saudi Arabia 20.48 46.62 ≈550 2D STEP El-Moslimany (1983)Oyo Sudan 19.27 26.18 510 1C STEP Ritchie et al. (1985)Achenouma Niger 19.12 12.90 ≈450 n/a STEP Schulz, unpublishedArrigui Niger 19.05 12.90 ≈450 n/a STEP Schulz, unpublishedBilma Niger 18.67 12.93 ≈450 n/a STEP Schulz, unpublishedEl Atrun Sudan 18.17 26.65 510 2D TXWS Ritchie & Haynes (1987)LGS3 Senegal 16.28 –15.83 1 1D TXWS Lezine (1988a)LGS2 Senegal 16.12 –15.92 0 1D SAVA Lezine (1988a, 1989a, b)Termit/Ouest Niger 16.20 11.07 ≈450 n/a TXWS Schulz, unpublishedPotou Senegal 15.75 –16.50 12 3C TSFO Lezine (1988b)Lompoul Senegal 15.42 –16.72 3 3C TSFO Lezine (1988b)Diogo Senegal 15.27 –16.80 8 2C TSFO Lezine (1988b, 1989a, b)Touba N’Diaye Senegal 15.17 –16.87 6 1C SAVA Lezine (1988b)Tanma Senegal 14.92 –17.08 ≈250 1D TXWS Medus (1984)Oursi Burkina Faso14.65 0.48 290 3C TXWS Ballouche & Neumann
(1995)Ari Koukouri Niger 13.93 13.10 270 n/a STEP Schulz, unpublishedTjeri Chad 13.73 16.50 300 1C TXWS Maley (1981, 1983, 1989)Bougdouma Nigeria 13.32 11.67 ≈300 4C SAVA Lezine and Gasse,
unpublishedBal Oasis Nigeria 13.31 10.95 ≈330 1C SAVA Waller, unpublishedKajemarum Oasis Nigeria 13.31 11.03 ≈330 2C SAVA Waller, unpublishedKonduga (c) Nigeria 11.63 13.43 220 1C TDFO Ballouche & Neumann,
(1995)Badda Ethiopia 7.87 39.37 4040 2C XERO Hamilton (1982);
Bonnefille & Hamilton(1986)
Abiyata Ethiopia 7.70 38.60 1578 6D WAMF Lezine & Bonnefille (1982);Lezine (19820
Langeno3 Ethiopia 7.67 38.73 1585 1D WAMF Bonnefille, unpublishedDanka Ethiopia 6.97 38.65 3830 4C XERO Hamilton ( 1982);
Bonnefille & Hamilton(1986)
Bosumtwi Ghana 6.53 –1.33 100 2C TSFO Maley & Livingstone(1983); Talbot et al. (1984)
continued
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1010 Dominique Jolly et al.
TABLE 1. Summary of pollen and plant macrofossil records at 6000 years for Africa and the Arabian peninsula.—continued
Site Name Country Latitude Longitude Elevation D.C. Biome Reference(dec.) (dec.) (m) 6000 years 6000 years
Barombi Mbo Cameroon 4.67 9.40 300 2C TSFO Brenac (1988); Maley et al.(1990); Maley (1991);Giresse et al. (1991)
Ossa Cameroon 3.83 10.00 8 2D WAMF Reynaud-Farrera (1995)Shanga Central Afr. 3.18 16.10 570 3D TSFO Bonnefille, Riollet and
Rep. Buchet, unpublishedAlbert Zaıre- 1.52 30.57 619 4C∗ WAMF Ssemmanda & Vincens
Uganda (1993)Koitoboss Kenya 1.13 34.57 3940 2D WAMF Hamilton (1982)Laboot Kenya 1.12 34.58 2880 1C XERO Hamilton (1982)Kimilili Kenya 1.10 34.57 4150 1C XERO Hamilton (1982)Kaisungor Kenya 1.00 35.47 2900 7C WAMF Coetzee (1967)Kitandara Uganda 0.35 29.88 3990 4D WAMF Livingstone (1967)Mahoma Uganda 0.35 29.97 2960 5D WAMF Livingstone (1967)Victoria Uganda 0.30 33.33 1134 1C WAMF Kendall (1969)Bogoria1 Kenya 0.18 36.17 990 1C∗ STEP Tiercelin, Vincens et al.
(1987)Sacred Kenya 0.03 37.47 2400 4C WAMF Coetzee (1967); Van
Zinderen Bakker & van &Coetzee (1972)
Rutundu Kenya –0.17 37.32 3140 1D XERO Coetzee (1967); VanZinderen Bakker &Coetzee (1972)
Naivasha Kenya –0.75 36.33 1890 2C STEP Maitima (1991)Rugezi1 Rwanda –1.40 29.83 2050 3D WAMF Roche, unpublishedGatovu 1 Burundi –2.53 30.05 1350 1C WAMF Elmoutaki, Bonnefille,
Riollet and Buchet,unpublished
Ndurumu2 Burundi –2.72 29.93 1363 2C WAMF Jolly & Bonnefille (1992)Buyongwe2 Burundi –2.80 29.92 1366 1D WAMF Bonnefille, Riollet and
Buchet, unpublishedBuyongwe3 Burundi –2.80 29.92 1366 1C TSFO Bonnefille, Riollet and
Buchet, unpublishedNyamuswaga1 Burundi –2.90 29.98 1546 5C WAMF Jolly (1993)Mukibongo Burundi –3.15 30.58 1540 4C WAMF Jolly (1993)Rusaka3 Burundi –3.43 29.62 2070 1C WAMF Bonnefille et al. (1995)Ijenda1 Burundi –3.45 29.60 2150 1D WAMF Roche & Bikwemu,(1989)Kashiru1 Burundi –3.47 29.57 2240 1C WAMF Bonnefille (1987);
Bonnefille & Riollet(1988b)
Bilanko Congo –3.52 15.35 700 7D TSFO Elenga et al. (1991)Tanganyika Bur17 Burundi –3.53 29.18 773 1D WAMF Vincens, unpublishedKuruyange 1 Burundi –3.58 29.68 2000 1C WAMF Bonnefille et al. (1991)Kuruyange 2 Burundi –3.58 29.68 2000 1C WAMF Jolly & Bonnefille (1991);
Jolly et al. (1994)Tanganyika Sd14 Burundi –4.03 29.33 773 1D WAMF Vincens (1989), (1993)Ngamakala Congo –4.07 15.38 400 4C TSFO Elenga (1992); Elenga et al.
(1994)Kitina Congo –4.27 12.00 100 3D TSFO Elenga, unpublishedTanganyika Sd24 Tanzania –4.58 29.33 773 4D SAVA Vincens (1989), (1993)Songolo Congo –4.77 11.87 5 1D TSFO Elenga, unpublishedTanganyika Sd36 Zambia/ –4.97 29.37 773 2D WAMF Vincens (1989), (1993)
TanzaniaTanganyika Mpu3 Zambia/ –8.50 30.62 773 1D SAVA Vincens, unpublished
TanzaniaLupembashi R.D. Congo –11.12 27.13 1150 2D TDFO Mbenza Muaka & Roche
(1980)Itasy Madagascar –19 46.50 1230 4C WAMF Straka (1993)Torotorofotsy Madagascar –19 48.50 956 7D WAMF Straka (1993)Tritrivakely1 Madagascar –19.78 46.92 1778 2C XERO Burney (1987)
continued
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1011
TABLE 1. Summary of pollen and plant macrofossil records at 6000 years for Africa and the Arabian peninsula.
Site Name Country Latitude Longitude Elevation D.C. Biome Reference(dec.) (dec.) (m) 6000 years 6000 years
Tritrivakely2 Madagascar –19.78 46.92 1778 1C XERO Gasse et al. (1994); VanCampo, unpublished
Drotsky Botswana –20.02 21.40 ≈950 1C TXWS Burney et al. (1994)Windhoek Namibia –22.38 17.50 1700 1C STEP Scott et al. (1991)Tate Vondo South Africa –22.88 30.33 1100 2D STEP Scott (1987a)Wondercrater South Africa –24.43 28.75 1100 2D TXWS Scott (1982)Moreletta South Africa –25.44 28.18 1310 3D SAVA Scott (1983)Saltpan South Africa –25.57 28.08 1100 1C TXWS Partridge et al. (1993)Rietvlei South Africa –25.83 28.33 1480 3D STEP Scott & Vogel (1983)Equus Cave South Africa –27.27 24.37 1250 2C XERO Scott (1987b)Wonderwerk South Africa –27.85 23.55 1665 3D STEP Van Zinderen Bakker
(1982)Teza South Africa –28.48 32.17 ≈30 1D TDFO Scott, unpublishedCraigrossie South Africa –28.53 28.47 1735 4C STEP Scott (1986, 1989)Blydefontein South Africa –31.09 25.04 1700 1C STEP Bousman et al. (1988)Pakhuis South Africa –32.06 19.04 600 2C XERO Scott (1994)Groenvlei South Africa –33.80 22.87 0 4C XERO Martin (1968)Hangklip South Africa –34.33 18.90 45 3C XERO Schalke (1973)
∗ Pollen and/or stratigraphic correlations with a nearby radiocarbon-dated site.– Co-ordinates are given for precise location of the pollen record.
(c), charcoal record.n/a, not available.
Codes for biome: DESE, Desert: STEP, Steppe: TXWS, Tropical xerophytic woods/scrub: SAVA, Savanna: TDFO, Tropical dry forest:TSFO, Tropical seasonal forest: TRFO, Tropical rain forest: WAMF, Warm mixed forest: XERO, Temperate xerophytic woods/scrub.Codes for dating control: For continuous records (C): 1C, bracketing dates within a 2000-yr interval about the time being assessed: 2C,bracketing dates, 1 within 2000 years, the second within 4000 years of the time being assessed: 3C, bracketing dates within a 4000-yr intervalabout the time being assessed: 4C, bracketing dates, 1 within 4000 years, the second within 6000 years of the time being assessed: 5C,:bracketing dates within a 6000-yr interval about the time being assessed: 6C, bracketing dates, 1 within 6000 years, the second within8000 years of the time being assessed: 7C,: poorly dated.For discontinuous records (D): 1D, date within 250 years of the time being assessed: 2D, date within 500 years of the time being assessed:3D, date within 750 years of the time being assessed: 4D, date within 1000 years of the time being assessed: 5D, date within 1500 years ofthe time being assessed: 6D, date within 2000 years of the time being assessed: 7D, poorly dated.)
year pollen data from Europe by Prentice et al. (1996). The (Prentice et al., 1996), we used a value of 0.5% for thepollen data, in order to reduce the noise due to occasionalmethod is briefly as follows. First each taxon is assigned to
one or more plant functional types (PFTs), on the basis of long-transport pollen grains. We have not used anythreshold percentage (hj=0) for plant macrofossil datathe known biology and biogeography of the plant species
it includes. The product of this step is a PFT (taxon matrix. because we assume they have a local source.The second step consists of the assignment of each PFT toone or more biomes, resulting in a biome (PFT matrix. A
IMPLEMENTATION FOR AFRICA ANDbiome (taxon matrix is then created using the two previousARABIAN PENINSULAones, illustrating which taxon occurs in each biome. The
affinity scores for each biome are then calculated for all The assignment of pollen taxa to the plant functional typespollen samples by (PFTs) was performed taking account of (1) the PFT
definitions following Prentice et al. (1992) and Prentice &Aik=Rj dij {max [0, (pjk – hj)]}1/2
Jolly (in press), and (2) the known biology of the plantsfrom several floras (Flore d’Afrique Centrale, 1972–85;where Aik is the affinity of pollen sample k for biome i;
summation is over all taxa j; dij is the entry in the biome× Flore du Congo Belge et du Ruanda-Urundi, 1948–63; Floredu Congo et al., 1967–71; Hutchinson & Dalziel, 1954–72),taxon matrix for biome i and taxon j; pjk are the pollen
percentages, and hj is a threshold pollen percentage. Each and botanical and palynological studies (Trochain, 1940;Quezel, 1965; Letouzey, 1968; Hamilton, 1972; Hall &pollen sample is assigned to the biome having the highest
affinity score. If two biomes have exactly the same affinity Swaine, 1976; Schnell, 1977; Huntley & Walker, 1982; White,1983; Ake-Assi, 1984; Walter, 1985; Schulz & Whitney,score, the sample is assigned to the biome that is defined
by the smaller number of taxa. 1986; Neumann, 1987). For pollen data collected inMadagascar, the assignment of the taxa to the differentHere, we used exactly the same procedure to reconstruct
the biomes both from pollen and from plant macrofossil PFTs was done using the comprehensive floristic andbiogeographic data provided by Straka (1991). The list ofdata. The only difference is in the value of hj. As in Europe
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1012 Dominique Jolly et al.
taxa and their assignment to PFTs is presented in Table 2. of the pollen taxa such as Artemisia are equally characteristicof both biomes.By our definition, the PFTs occurring in Africa are as
(7) Desert forb/shrub (df). Again ‘warm’ and ‘cool’ desertfollows.plants are not separated. This PFT includes drought(1) Tropical evergreen (Te). According to Prentice et al.resistant taxa such as Heliotropium strigosum∗, Ephedra and(1992), tropical evergreen trees require a mean temperatureCruciferae.of the coldest month (Tc) > 15.5 °C and values of the ratio
All arboreal pollen taxa present only in the circum-(a) of actual to equilibrium evapotranspiration > 0.80. TeMediterranean region were assigned to the PFTs (borealwas subdivided into (1) ‘wet’ tropical evergreen trees (Te1,summer green [bs], eurythermic conifer [ec], temperateincluding taxa such as Pausinystalia∗ macroceras orsummer green [ts], cool-temperate summer green [ts1],Calpocalyx∗ that are characteristic of the rain forest;warm-temperate summer green [ts2], warm-temperatea > 0.95), and ‘dry’ tropical evergreen trees (Te2, e.g.sclerophyll shrub [wte2]) as in Prentice et al. (1996), whoDacryodes, Crudia gabonensis, that are also found in tropicalused more than 600 modern samples collected south ofseasonal forest; a < 0.95).40°N to check these assignments. Empirically, Gramineae(2) Tropical raingreen (Tr). Tropical raingreen trees havewas assigned to all the nonarboreal plant functional types,temperature constraints similar to tropical evergreen trees,except desert (never > 40% in the pollen samples collectedbut can withstand a longer dry season by shedding theirin the Namib, Sahara and Rub al Khali deserts). Contraryleaves. This PFT was subdivided into three groups: Tr1 (e.g.to the European scheme, we also allowed Gramineae toChaetacme or Tiliacora∗ funifera) which occur in seasonalcharacterize temperate xerophytic woods/scrub, because thisforests, Tr2 (e.g. Brachystegia or Cochlospermum)taxon is always present (Hedberg, 1954; White, 1983) withcharacteristic of dry forests, and Tr3 (e.g. Hallea∗high percentages in the high-elevation xerophytic vegetationrubrostipulata) confined to the dryest woodlands andin the intertropical area (Hamilton, 1972) and in the fynbossavannas. Tr3 helps to identify the savanna biome (Table 3).vegetation of Cape Province (Schalke, 1973).(3) The tropical sclerophyll/succulent PFT (Tss) defined
With these PFT definitions, biomes occurring in Africaprovisionally in Jolly et al. (in press) was found not tocan be defined as very simple combinations of PFTs, ascorrespond accurately with any of the taxa present in ourindicated in Table 3.modern pollen data set. Succulents possess Crassulacean
Acid Metabolism (CAM) (Larcher, 1995), a photosyntheticsystem that allows them to thrive in environments subject RESULTSto periodic extreme drought and therefore allows them tooccur across a range of biomes from dry savanna to desert Present-day pollen-derived biome reconstruction(White, 1983). Twenty-five families (Euphorbiaceae,
A visual comparison of the pollen-derived biome mapRubiaceae, Asteraceae, Liliaceae . . .) include members that
(Fig. 1) with the modern biome distribution (Olson et al.,use CAM and many of these are present in Africa (Griffiths, 1983; Fig. 2) shows a good agreement. A numerical1989), but in our data set only Aizoaceae and Crassulaceae comparison (Table 4) also shows the following.are unambiguously succulents. These two families grow For almost all of the biomes, the number of correctpreferentially in the steppe and desert biomes, where they assignments exceeds the number of incorrect assignments.employ CAM in response to drought or salt stress (Larcher, Steppe, temperate xerophytic woods/scrub, tropical1995). Some other sclerophyll taxa also belong in the steppe xerophytic woods/scrub, savanna, warm mixed forest,forb (sf) or in the Tr3 plant functional types. tropical rain forest and tropical seasonal forest are predicted
(4) Warm-temperate evergreen (wte). This PFT includes with high confidence.taxa of the montane forests of the Maghreb, East Africa, When incorrect assignments occur, the method usuallyMadagascar and South Africa, regions where Tc is in the indicates a biome that is contiguous in bioclimatic space.5–15.5 °C range and a > 0.65. This PFT includes both broad- For example, steppe samples are sometimes assigned to(Macaranga, Syzygium, Entandrophragma excelsa . . .) and savanna (ten times), tropical xerophytic woods/scrub (nineneedle-leaved taxa such as the warm temperate conifers times) or temperate xerophytic woods/scrub (six times)(Podocarpus, Juniperus procera). instead of steppe (correctly predicted 171 times).
(5) Temperate sclerophyll/succulent (tss): this PFT Table 4 shows that the biomization method applied toincludes both the taxa occurring in the circum- modern pollen data from Africa and the Arabian peninsulaMediterranean (maquis and garrigue; Olea, Phillyrea, is successful at a large scale, but two biomes present a lowerPistacia, Cistus . . .) and Cape (fynbos; Restionaceae) percentage of correct assignments using the modern pollenregions and the taxa occurring in the drier parts of the data set currently available: namely desert and tropical drymontane and high plateaux in the intertropical area forest.(Ericaceae, Protea . . .), at the margins of the forested The sixteen modern samples assigned to desert are indeedbiomes. from the desert biome, but twenty-eight other samples from
(6) Steppe forb/shrub (sf): as for the other parts of the the desert biome are assigned to steppe. The most probableglobe where the biome reconstruction was attempted, we cause of this underestimate of desert comes from the naturehave grouped the two initial PFTs ‘warm’ and ‘cool’ grasses/ of the vegetation sampled. It is extremely difficult to collectshrubs sensu Prentice et al. (1992), because they could not modern pollen samples in desertic areas. Most often, the
samples are collected in unusual locations where samplingbe distinguished among our modern pollen data set. Many
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1013
TABLE 2. Plant functional types and the pollen and plant macrofossil taxa assigned to them.
continued
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1014 Dominique Jolly et al.
TABLE 2. Plant functional types and the pollen and plant macrofossil taxa assigned to them.—continued
continued
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1015
TABLE 2. Plant functional types and the pollen and plant macrofossil taxa assigned to them.—continued
continued
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1016 Dominique Jolly et al.
TABLE 2. Plant functional types and the pollen and plant macrofossil taxa assigned to them.—continued
continued
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1017
TABLE 2. Plant functional types and the pollen and plant macrofossil taxa assigned to them.—continued
continued
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1018 Dominique Jolly et al.
TABLE 2. Plant functional types and the pollen and plant macrofossil taxa assigned to them.—continued
TABLE 3. African biomes and their characteristic plant functional rain forest or tropical seasonal forests (three plus ten times),types (PFTs). Abbreviations for PFTs as in Table 2. temperate xerophytic woods/scrub (six times) or warm
mixed forest (six times) for some modern samples from theBiome Plant functional typetropical dry forest biome. This might suggest a problem
Tropical rain forest Te1+Te2 with the assignment of taxa to the critical PFT Tr2, butTropical seasonal forest Te2+Tr1 this is unlikely because the miombo (African tropical dryTropical dry forest Tr2 forest) is one of the best known of African biomes from aWarm mixed forest wte+ec+bs+ts+ts1+ts2
floristic and ecological point of view (Lawton, 1978;Temperate xerophytic woods/ tss+ecMalaisse, 1978; Chidumayo, 1987; Ernst, 1988; Munyanzizascrub& Oldeman, 1996). It seems more probable that the surfaceTropical xerophytic woods/scrub Tss
Savanna Tr3 sample set is somewhat unrepresentative geographically,Steppe sf and indeed many of the misallocated samples are near theDesert df biome margin while the core area of the miombo in southern
Africa has not been sampled at all.
is possible. The vegetation at these locations is normally6000 year biome reconstruction
closer to steppe than desert. Thus this tendency tounderestimate desert should not affect the interpretation of The biomization method was applied to the 6000 year data
set, including the ninety-nine pollen records and the six6000 year samples derived from lake sediments or charcoalfinds. charcoal records, across Africa and the Arabian peninsula.
The resulting reconstruction (Fig. 3) shows coherentTropical dry forest was identified incorrectly for threesurface samples from the savanna biome, one surface sample patterns of biome distribution. There are clear spatial shifts
of several biomes in comparison with their modernfrom the steppe biome and one surface sample from thewarm mixed forest biome. Note however, that with one distributions. The main features revealed by the comparison
of Figs 1 and 3 can be summarized as follows.exception only, these misidentified samples were collectedin ‘wooded savanna’ according to the authors. Conversely, In the Maghreb, warm mixed forest was present at high
elevations (over 1200 m) while temperate xerophytic woods/twenty-six samples from the tropical dry forest were assignedto savanna. Definitions of savanna vary considerably scrub occurred in the lowlands.
The desert that now defines the Sahara was extremely(White, 1983; Eiten, 1986; Scholes & Walker, 1993), andthe ‘natural’ definition in terms of PFT abundances may reduced. In the 6000 year data set, only two sites in southern
Egypt are assigned to desert. All the other Saharan siteswell differ from that of Olson et al. (1983).A greater concern is the incorrect identification of tropical that are now desert were either steppe, at low elevation, or
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1019
TABLE 4. Numerical comparison of each site between pollen-derived (‘p’) and observed (‘o’) biomes at 0 ka for Africa and the Arabianpeninsula (DESE=desert; STEP=steppe; TXWS=tropical xerophytic woods/scrub; SAVA=savanna; XERO=temperate xerophyticwoods/scrub; WAMF=warm mixed forest; TDFO=tropical dry forest; TSFO=tropical seasonal forest; TRFO=tropical rain forest.
DESEo STEPo SAVAo XEROo WAMFo TDFOo TSFOo TRFOo
DESEp 5 0 0 0 0 0 0 0STEPp 7 126 27 2 2 3 0 0SAVAp 0 25 206 3 2 41 2 0XEROp 1 14 25 98 54 8 0 0WAMFp 0 0 3 6 140 7 4 0TDFOp 0 1 7 0 1 21 0 0TSFOp 0 0 3 0 0 8 32 8TRFOp 0 0 0 0 0 2 0 13
temperate xerophytic woods/scrub, or even warm mixed reduction in the representation of desert plants along thenorth–south gradient.forest in the Saharan mountains.
Tropical seasonal forest was present in all the sites from Examination of the biome scores also provides usefulinformation about the nature of the vegetation recorded incentral Africa, except for one site from Cameroon where
there was warm mixed (tropical montane) forest. Tropical the pollen data. For example, a group of 6000 year pollensites from Senegal are assigned either to savanna (two times)rain forest was not present at any of the sampling sites in
this region. However, the density of 6000 year sites in the and tropical xerophytic woods/scrub (two times) or tropicalseasonal forest (three times). The inconsistency may have aequatorial region is very low so it is impossible to infer the
true extent of the tropical rain forest biome at 6000 years. simple explanation. At the three pollen sites identified toforest, the second largest affinity score is for savanna,In eastern and southern Africa (including Madagascar)
the biome distribution was generally similar to today, with suggesting that these sites may be recording a local (riparian)forest within a regional savanna vegetation matrix. Thiswarm mixed forest on the mountains, and temperate
xerophytic woods/scrub on the high-plateaux and in the interpretation implies an azonal northward migration ofraingreen taxa with Guinean affinity along rivers, and solvesCape province. Only one pollen record documents the
continental biome distribution between 15 and 20°S. It the problem posed by the apparent disagreement in theinterpretation of the pollen data at similar latitudes fromindicates the occurrence of tropical dry forest in southern
Zaıre and savanna in northern Botswana, as today. the western and the central Sahel, discussed by Ballouche& Newmann (1995).However, warm mixed forest is not shown in South Africa,
even for pollen sites close to the mountains such as theDrakensberg: here the data show temperate xerophytic
DISCUSSIONwoods/scrub at 6000 years.We now examine some local-scale features in more depth, The main features of the pollen-and plant macrofossil-
by looking at the affinity scores of some 6000 year samples derived biome distribution of Africa at 6000 years arefor different biomes, in order to add more information that consistent with previous interpretations based on a morecan be obtained from assignments. qualitative interpretation of the data at a continental scale
At the two sites in southern Egypt (Abu Minqar and (Street-Perrott & Perrott, 1993; Jolly et al., 1998). TheWadi el Akhdar) where desert is shown for 6000 years, the palaeoclimatic significance of the patterns shown in ouraffinity scores are in fact identical for desert and steppe. biome reconstruction remains to be discussed.These sites are defined by the occurrence of charcoals ofTamarix, which can be found either in the steppe or in the 1. Our reconstruction of temperate xerophytic woods/scrub
in the lowlands of the Maghreb is in good agreementdesert, particularly in the wadis (White, 1983). Steppe isencountered at the nearest 6000 year site further south with pollen data from marine cores in the Gulf of
Gabes. These data indicate the establishment of the Oleo-(Wadi Bakht), suggesting that the steppe/desert ecotone islocated somewhere in southern Egypt ≈23°N. Lentiscetum (Mediterranean sclerophyll) association by
8600 years (Brun, 1979, 1983). This pattern parallelsTwo sites in the Sudan (Oyo and El Atrun) similarlyprovide an objective estimate of the 6000 year location of that found in the northern Mediterranean region where
temperate forests of the early to mid-Holocene (Prenticethe southern limit of the sahelian steppe in the easternSahara, at 19°N. The ratio R=Adesert/(Adesert + Asteppe) was et al., 1996) were eventually replaced by xerophytic
vegetation, suggesting conditions wetter than presentalso calculated at Selima, Oyo and El Atrun. These arepollen sites analysed by the same palynologist, located at (Cheddadi et al., 1997).
2. The apparent absence of warm mixed forest in the easterndifferent latitudes. R is largest at the northernmost siteSelima (21.37°N; R=0.33), intermediate at Oyo (19.27°N; part of South Africa, where xerophytic vegetation was
present instead, may indicate drier than presentR=0.24), and smallest at the southernmost site El Atrun(18.17°N; R=0.08). These figures illustrate a gradual conditions in this region at 6000 years. This result is in
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027
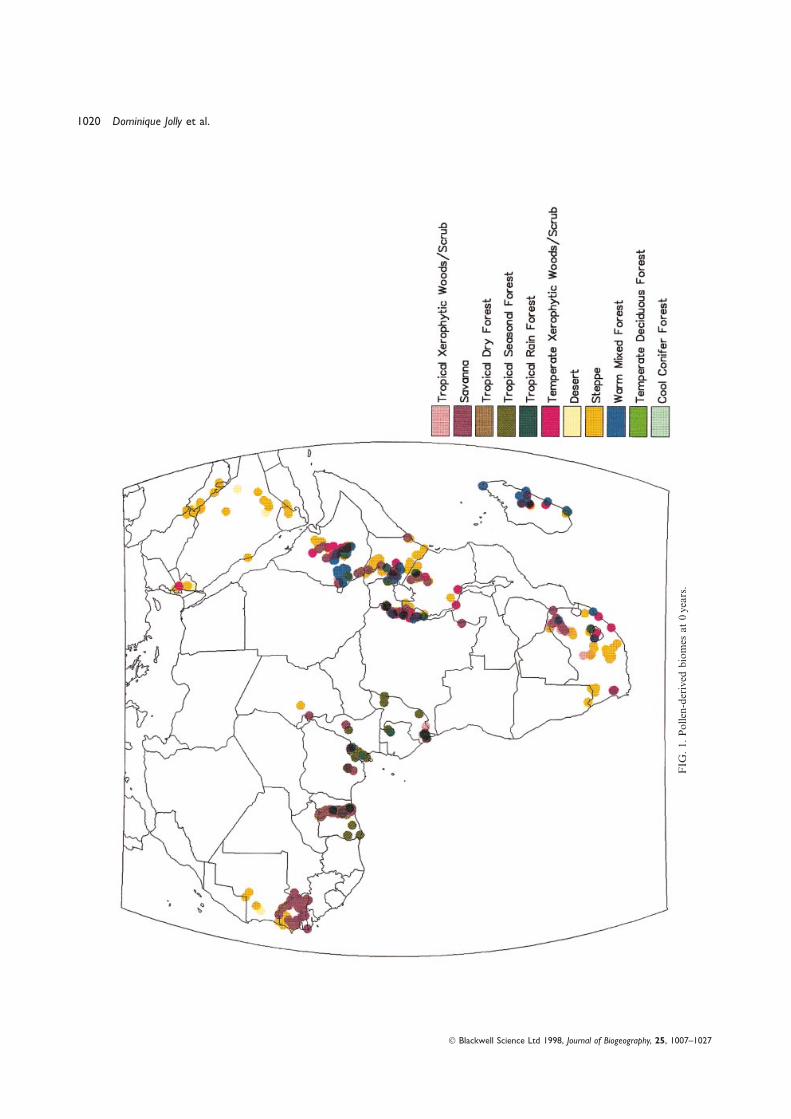
1020 Dominique Jolly et al.
FIG
.1.
Pol
len-
deri
ved
biom
esat
0ye
ars.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1021
FIG
.2.
Mod
ern
biom
edi
stri
buti
onfr
omO
lson
etal
.(1
983)
.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1022 Dominique Jolly et al.
FIG
.3.
Pol
len
and
plan
tm
acro
foss
il-de
rive
dbi
omes
at60
00ye
ars.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1023
ACKNOWLEDGMENTSagreement with palynological investigations from twomarine cores in the Mozambique Channel (Elmoutaki
Financial support for this project was provided by theet al., 1992; Elmoutaki, 1994), characterized by high
European Community (EV5 V-CT95–0075), the U.S.percentages of Podocarpus and raingreen taxa (Tr2 and National Science Fundation (TEMPO: Testing EarthTr3) and suggesting drier than present conditions in the System Models with Palaeoenvironmental Observations),Zambezi Basin. and by the US Environmental Protection Agency via a
3. The biome reconstruction shows an extreme reduction subcontract with the University of New Hampshire. Thisof the Sahara desert at 6000 years, while more moisture- is a contribution to the IGBP-GAIM (Global Analysis,demanding biomes (steppe at low elevation, xerophytic Interpretation and Modelling) ‘6000 years BP experiment’,vegetation and warm mixed forest at higher elevation) to the core research of IGBP’s project on Global Changewere found in the present-day desert region. This and Terrestrial Ecosystems (GCTE), and to PMIPreconstruction is in full agreement with previous (Palaeoclimate Modelling Intercomparison Project); PMIPsyntheses (Wickens, 1975; Schulz, 1987; Neumann, is sponsored by the IGBP and WCRP. We are grateful to1989a,b; Petit-Maire, 1992; Street-Perrott & Perrott, E. Schulz for providing us with raw pollen counts from1993; Lioubimtseva, 1995; Jolly et al., 1998). The numerous published and unpublished sites from the Centralidentification of steppe at almost all of the Saharan sites Sahara, and to F. Gasse who allowed us to use thecannot be explained away by a ‘wadi effect’ (runoff from unpublished pollen counts from Bougdouma. The datathe ergs and accumulation of the water in the wadis) management and all the calculations were performed usingbecause many of the 6000 year sites are not located in the software PPPBASE (J. Guiot and C. Goeury, LBHP,wadis. Furthermore, independent data suggest the former Marseille, France)occurrence of steppe over large parts of the Sahara [weeds(Wasylikowa, 1992) and faunal data (Wendorf & Schild,
REFERENCES1980; Schild & Wendorf, 1982; Guerin & Faure, 1983;Peters, 1988; Vernet & Striedter, 1992)]. Almost all ofthe lakes in the Sahara were high at 6000 years (Street- Agwu, C.O.C. (1986) History of climate and vegetation of eastPerrott et al., 1989; Jolly et al., 1998), and the lakes and central Nigeria as deduced from pollen analysis: preliminarywetlands occupied≈7.5% of the total area of the Sahara investigations. Bull. ASEQUA, N° Special Symp INQUA-
ASEQUA, June 1986, 5–8.(Hoelzmann et al., 1998) as compared with 0.5% todayAke-Assi, L. (1984) Flore de la Cote d’Ivoire: etude descriptive et(Cogley, 1991). A large increase in precipitation is needed
biogeographique, avec quelques notes ethnobotaniques.to explain these findings, and such an increase couldUnpublished thesis, University of Abidjan.probably only be achieved by a strong enhancement of
Ballouche, A. & Neumann, K. (1995) A new contribution to thethe African monsoon (Pachur & Braun, 1980; KutzbachHolocene vegetation history of the West African Sahel: pollen
& Street-Perrott, 1985; Petit-Maire, 1992). Estimates of from Oursi, Burkina Faso, and charcoal from three sites inthe total amount of precipitation during the early to northeast Nigeria. Veg. Hist. Archaeobo. 4, 31–39.mid-Holocene range between 100 mm/yr (Kropelin, 1987) Bernard, J. & Reille, M. (1987) Nouvelles analyses polliniques dansand 200 mm/yr (Haynes & Haas, 1980) in the eastern l’Atlas de Marrakech, Maroc. Pollen Spores, 29, 225–240.
Beucher, F. (1971) Etude palynologique des formations neogenesSahara and between 200 mm/yr (Petit-Maire & Riser,et quaternaires au Sahara nord-occidental. Unpublished thesis,1988) and 500 mm/yr (Riser, 1989) in the western Sahara.University of Paris.Some authors have advanced the hypothesis that the
Beucher, F. (1979) Flore: contexte palynologique. Le Sahararegion experienced a Mediterranean-type winter rainfallAtlantique a l’Holocene. Peuplement et ecologie (ed. by N. Petit-
regime (Butzer & Hansen, 1968; Petit-Maire, 1989), but Maire), pp. 235–237. CRAPE (mem. 28), Algiers.our biome reconstruction does not allow us to confirm Bonnefille, R. (1987) Evolution forestiere et climatique au Burundior reject this hypothesis. durant les quarante derniers milliers d’annees. C. R. Acad. Sci.
305, 1021–1026.Bonnefille, R., Chalie, F., Guiot, J. & Vincens, A. (1992)The biome reconstructions from equatorial Africa show
Quantitative estimates of full glacial temperatures in equatorialat one site (in Cameroon) the presence of a tropicalAfrica from palynological data. Clim. Dyn. 6, 251–257.
deciduous forest at 6000 years, in place of the modern Bonnefille, R. & Hamilton, A.C. (1986) Quaternary and Latetropical rain forest. This reconstruction is based on the Tertiary history of Ethiopian vegetation. Acta Univ. Upsa. Symb.increase of the relative proportion of deciduous trees Bot. 26, 48–63.(raingreen, Tr1) implying an increase in the length of the Bonnefille, R. & Riollet, G. (1988a) Palynologie des sediments
holocenes de sites archeologiques du Qatar. Prehistoire a Qatardry season. This result is in complete agreement with much(ed. by M.-L. Inizan), pp 137–145. Edns Recherche sur lesevidence in pollen diagrams from equatorial East Africa forCivilisations, Paris.relatively high abundance of deciduous taxa such as Celtis
Bonnefille, R. & Riollet, G. (1988b) The Kashiru pollen sequencewithin tropical montane forest at 6000 years (Livingstone,(Burundi). Palaeoclimatic implications for the last 40,000 yr B.P.
1967; Kendall, 1969; Taylor, 1990, 1993; Ssemmanda & in tropical Africa. Quat. Res. 30, 19–35.Vincens, 1993; Jolly et al., 1994). Thus our findings for the Bonnefille, R., Riollet, G. & Buchet, G. (1991) Nouvelle sequenceequatorial region are coherent with the hypothesis of a pollinique d’une tourbiere de la crte Zaıre-Nil (Burundi). Rev.northward migration (not simply an expansion) of the Palaeobot. Palynol. 67, 315–330.
Bonnefille, R., Riollet, G., Buchet, G., Icole, M., Lafont, R.,African monsoon.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1024 Dominique Jolly et al.
Arnold, M. & Jolly, D. (1995) Glacial/Interglacial record from Elmoutaki, S. (1994) Transition glaciaire-interglaciaire et YoungerDryas dans l’Hemisphere Sud (1°-20° Sud): analyse palynologiqueintertropical Africa, high resolution pollen and carbon data at
Rusaka, Burundi. Quat. Sci. Rev. 14, 917–936. a haute resolution de sondages marin et continentaux (lac etmarecage) 130 pp. Unpublished thesis, University of Aix-Bousman, C.B., Partridge, T.C., Scott, L., Metcalfe, S.E., Vogel,
J.C., Seaman, M. & Brink, J.S. (1988) Palaeoenvironmental Marseille III.Elmoutaki, S., Lezine, A.M. & Thomassin, B.A. (1992) Mayotteimplications of Late Pleistocene and Holocene valley fills in
Blydefontein Basin, Noupoort, C.P., South Africa. Palaeoecol. (canal de Mozambique). Evolution de la vegetation et du climatau cours de la derniere transition glaciaire-interglaciaire et deAfrica, 19, 43–67.
Brenac, P. (1988) Evolution de la vegetation et du climat dans l’Holocene. C. R. Acad. Sci. 314, 237–244.Ernst, W.H.O. (1988) Seed and seedling ecology of Brachystegial’Ouest-Cameroun entre 25 000 et 11 000 ans B.P. Institut Francais
Pondichery, Trav. Sec. Sci. Techn. 25, 91–103. spiciformis, a predominant tree component in miombowoodlands in South Central Africa. For. Ecol. Mgmnt, 25,Brun, A. (1979) Recherches palynologiques sur les sediments marins
du Golfe de Gabes: resultats preliminaires. Geol. Mediter. 4, 195–210.Flore d’Afrique Centrale (Zaıre & R.w.a.n.d.a., Burundi) (1972–85)247–264.
Brun, A. (1983) Etude palynologique des sediments marins Jard. Bot. Nat. Belgique, Bruxelles.Flore du Congo Belge et du Ruanda-Urundi (1948–63) Publ. Inst.Holocenes de 5000 B.P. a l’actuel dans le Golfe de Gabes (Mer
Pelagienne). Pollen Spores, 25, 437–460. Nat. Et. Agron. Congo Belge, Bruxelles, 10 volumes.Flore du Congo, du Rwanda et du Burundi (1967–71) Jard. Bot.Burney, D.A. (1987) Pre-settlement vegetation changes at Lake
Tritrivakely, Madagascar. Palaeoecol. Africa, 18, 357–381. Nat. Belgique, Bruxelles.Foley, J.A. (1994) The sensitivity of the terrestrial biosphere toBurney, D.A., Brook, G.A. & Cowart, J.B. (1994) A Holocene
pollen record for the Kalahari Desert of Botswana from a U- climatic change: a simuation of the middle Holocene. GlobalBiogeochem. Cycles, 8, 505–525.series dated speleothem. The Holocene, 4, 225–232.
Butzer, K.W. & Hansen, C.L. (1968) Desert and river in Nubia: Foley, J.A., Prentice, I.C., Ramankutty, N., Levis, S., Pollard, D.,Sitch, S. & Haxeltine, A. (1996) An integrated biosphere model ofgeomorphology and prehistoric environments at the Aswan
reservoir, 562 pp. University of Wisconsin Press, Madison. land surface processes, terrestrial carbon balance, and vegetationdynamics. Global Biogeochem. Cycles, 10, 603–628.Cheddadi, R., Yu, G., Guiot, J., Harrison, S.P. & Prentice, I.C.
(1997) The climate of Europe 6000 years ago. Clim. Dyn. 13, Gabriel, B. (1977) Zum okologischen Wandel im Neolithikum derostlichen Zentralsahara. Berl. Geogr. Abh. 27, 1–70.1–9.
Chidumayo, E.N. (1987) A survey of wood stocks for charcoal Gasse, F., Cortijo, E., Disnar, J.R., Ferry, L., Gibert, E., Kissel,C., Laggoun-Defarge, F., Lallier-Verges, E., Miskovsky, J.C.,production in the miombo woodlands of Zambia. For. Ecol.
Mgmnt, 20, 105–115. Ratsimbazafy, B., Ranaivo, F., Robison, L., Tucholka, P., Saos,J.L., Siffedine, A., Taieb, M., Van Campo, E. & Williamson, D.Claussen, M. (1994) On coupling global biome models with climate
models. Clim. Res. 4, 203–221. (1994) A 36 ka environmental record in the southern tropics:Lake Tritrivakely (Madagascar). C. R. Acad. Sci. 318, 1513–1519.Claussen, M. & Esch, M. (1994) Biomes computed from simulated
climatologies. Clim. Dyn. 9, 235–243. Giresse, P., Maley, J. & Kelts, K. (1991) Sedimentation andpalaeoenvironment in crater lake Barombi Mbo, Cameroon,Coetzee, J.A. (1967) Pollen analytical studies in East and southern
Africa. Palaeoecol. Africa, 3, 1–146. during the last 25,000 years. Sedim. Geol. 71, 151–175.Griffiths, H. (1989) Carbon dioxide concentrating mechanisms andCogley, G.C. (1991) GGHYDRO Global Hydrological Data,
Release 2.0. Trent Climate Note, 91, 1–11. the evolution of CAM in vascular epiphytes. Vascular plants asepiphytes. Evolution and ecophysiology (ed. by U. Luttge), pp.COHMAP Members (1988) Climatic changes of the last 18000
years: observations and model simulations. Science, 241, 42–86. Springer, Berlin.Guerin, C. & Faure, M. (1983) Mammiferes. Sahara ou Sahel?1043–1052.
Cooremans, B. (1989) Pollen production in central southern Africa. Quaternaire recent du bassin de Taoudenni (Mali) (ed. by N.Petit-Maire and J. Riser), pp. 239–272. Marseille: ImprimeriePollen Spores, 31, 61–78.
Cour, P. & Duzer, D. (1976) Persistance d’un climat hyperaride au Lamy.Hall, J.B. & Swaine, M.D. (1976) Classification and ecology ofSahara central et meridional au cours de l’Holocene. Rev. Geogr.
Phys. Geol. Dyn. 18, 175–198. closed-canopy tropical forest in Ghana. J. Ecol. 64, 913–952.Hamilton, A.C. (1972) The interpretation of pollen diagrams fromEdorh, M.T. (1986) Vegetation et pluie pollinique actuelles au Togo,
146 pp. Unpublished thesis, University of Aix-Marseille II. Highland Uganda. Palaeoecol. Africa, 7, 45–149.Hamilton, A.C. (1982) Environmental history of East Africa: a studyEiten, G. (1986) The use of the term ‘savanna’. Trop. Ecol. 27,
10–23. of the Quaternary. Academic Press, New York.Harrison, S.P., Kutzbach, J.E., Prentice, I.C., Behling, P.J. & Sykes,Elenga, H. (1992) Vegetation et climat du Congo depuis 24 000 ans
B.P. Analyse palynologique de sequences sedimentaires du Pays M. (1995) The response of Northern Hemisphere extratropicalclimate and vegetation to orbitally induced changes in insolationBateke et du littoral, 238 pp. Unpublished thesis, University of
Aix-Marseille III. during the last interglaciation. Quat. Res. 43, 174–184.Haxeltine, A. & Prentice, I.C. (1996) BIOME3: an equilibriumElenga, H., Schwartz, D. & Vincens, A. (1994) Pollen evidence of late
Quaternary vegetation and inferred climate changes in Congo. terrestrial biosphere model based on ecophysiological constraints,resource availability, and competition among plant functionalPalaeogeogr. Palaeoclim. Palaeoecol. 109, 345–356.
Elenga, H., Vincens, A. & Schwartz, D. (1991) Presence d’elements types. Global Biogeochem. Cycles, 10, 693–709.Haynes, C.V. & Haas, H. (1980) Radiocarbon evidence for Holoceneforestiers montagnards sur les Plateaux Bateke (Congo) au
Pleistocene superieur. Nouvelles donnees palynologiques. recharge of groundwater, Western Desert, Egypt. Radiocarbon,22, 705–717.Palaeoecol. Africa, 22, 239–252.
El-Moslimany, A. (1983) History of climate and vegetation in the Hedberg, O. (1954) A pollen-analytical reconnaissance in tropicalEast Africa. Oikos, 5, 137–166.Eastern Mediterranean and the Middle East from the Pleniglacial
to the mid-Holocene, 229 pp. Unpublished thesis, University of Hoelzmann, P., Jolly, D., Harrison, S.P., Laarif, F., Bonnefille, R.& Pachur, H.-J. (1998) Mid-Holocene land-surface conditions inWashington.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1025
northern Africa and Arabian peninsula: a data set for AGCM Letouzey, R. (1968) Etude phytogeographique du Cameroun. P.Lechevalier, Paris.sensitivity experiments. Global Biogeochem. Cycles, 12, 35–51.
Huntley, B.J. & Walker, B.H. (1982) Ecology of tropical savannas. Lezine, A.-M. (1982) Etude palynologique des sedimentsQuaternaires du lac Abiyata (Ethiopie). Palaeoecol. Africa, 14,Ecological Studies 42, Springer-Verlag, Berlin.
Hutchinson, J. & Dalziel, J.M. (1954) 1958, 1963, 1972) Flora of 93–98.Lezine, A.-M. (1987) Paleoenvironnements vegetaux d’Afrique nordWest Tropical Africa, Vols 1–4. Whitefriars Press, London.
Jolly, D. (1993) Evolution et dynamique des ecosystemes du Burundi. tropicale depuis 12000 ans B.P. Analyse pollinique de seriessedimentaires continentales (Senegal-Mauritanie). These dePollen et statistique. Unpublished thesis, University of Aix-
Marseille II. doctorat, Universite Aix-Marseille II.Lezine, A.M. (1988a) New pollen data from the Sahel, Senegal.Jolly, D. & Bonnefille, R. (1991) Diagramme pollinique d’un
sondage Holocene de la Kuruyange (Burundi, Afrique Centrale). Rev. Paleobot. Palynol. 55, 141–154.Lezine, A.M. (1988b) Les variations de la couverture forestierePalaeoecol. Africa, 22, 265–274.
Jolly, D. & Bonnefille, R. (1992) Histoire et dynamique du marecage mesophile d’Afrique occidentale au cours de l’Holocene. C. R.Acad. Sci. 307, 439–445.tropical de Ndurumu (Burundi), donnees polliniques. Rev.
Palaeobot. Palynol. 75, 133–151. Lezine, A.M. (1989a) Le Sahel: 20 000 ans d’histoire de lavegetation. Bull. La Soc. Geol. France, 5, 35–42.Jolly, D., Bonnefille, R. & Roux, M. (1994) Numerical interpretation
of a high resolution Holocene pollen record from Burundi. Lezine, A.M. (1989b) Late Quaternary Vegetation and Climate ofthe Sahel. Quaternary Res. 32, 317–334.Palaeogeogr. Palaeoclim. Palaeoecol. 109, 357–370.
Jolly, D., Burcq, S., Bonnefille, R. & Roux, M. (1996) Lezine, A.M. & Bonnefille, R. (1982) Diagramme polliniqueHolocene d’un sondage du lac Abiyata (Ethiopie, 7°42Nord).Representation pollinique de la fort dense humide du Gabon,
tests statistiques. C. R. Acad. Sci. 322, 63–69. Pollen Spore, 24, 463–480.Lioubimtseva, E.U. (1995) Landscape changes in the Saharo-Jolly, D., Harrison, S.P., Damnati, B. & Bonnefille, R. (1998)
Simulated climate and biomes of Africa during the Late Arabian area during the last glacial cycle. J. Arid Environ. 30,1–17.Quaternary: comparison with pollen and lake status data. Quat.
Sci. Rev. 17, 629–657. Livingstone, D.A. (1967) Postglacial vegetation of the RuwenzoriMountains in Equatorial Africa. Ecol. Monogr. 37, 25–52.Jolly, D., Prentice, I.C., Guiot, J., Bengo, M., Bonnefille, R.,
Brenac, P., Buchet, G., Edorh, T., Elenga, H., Lezine, A.M., Maitima, J.M. (1991) Vegetation response to climatic change inCentral Rift Valley, Kenya. Quat. Res. 35, 234–245.Maley, J., Reynaud-Farrera, I., Riollet, G. & Vincens, A. (in
press) Reconstitution de biomes a partir des donnees polliniques Malaisse, F. (1978) The miombo ecosystem. Tropical forestecosystems. Natural Resources Research XIV, UNESCO, Paris.actuelles d’Afrique de l’Ouest. Geogr. Physique Quat.
Joussaume, S. & Taylor, K.E. (1995) Status of the Paleoclimate Maley, J. (1981) Etudes palynologiques dans le bassin du Tchad etpaleoclimatologie de l’Afrique nord-tropicale de, 30, 000 ans aModeling Intercomparison Project (PMIP). WCRP Report, 92,
425–430. l’epoque actuelle. Thesis, Universite des Sciences et Techniquesdu Languedoc. Travaux et documents 127. Edns de l’ORSTOM.Kendall, R.L. (1969) An ecological history of the Lake Victoria
basin. Ecol. Monogr. 39, 121–176. Paris.Maley, J. (1983) Histoire de la vegetation et du climat de l’AfriqueKropelin, S. (1987) Palaeoecological evidence from early to mid-
Holocene playas in the Gilf Kebir (SW Egypt). Palaeoecol. Africa, nord-tropicale au Quaternaire recent. Bothalia, 14, 377–389.Maley, J. (1989) Les climats. 20.000 ans d’evolution des climats du18, 189–208.
Kutzbach, J.E., Bonan, G., Foley, J. & Harrison, S.P. (1996) Sahara Central aux Savanes tropicales humides. Sud Sahara-Sahel Nord, pp. 34–52. Publ. Centre Cult. Francais Abidjan, pp.Vegetation and soil feedbacks on the response of the African
monsoon to orbital forcing in the early to middle Holocene. 589–606.Maley, J. (1991) The African rain forest vegetation andNature, 384, 623–626.
Kutzbach, J.E., Gallimore, R., Harrison, S.P., Behling, P., Selin, palaeoenvironments during Late Quaternary. Climatic Change,19, 79–98.R. & Laarif, F. (1998) Climate simulations for the past 21,000
years. Quat. Sci. Rev. 17, 473–506. Maley, J. & Brenac, P. (1987) Analyses polliniques preliminairesdu Quaternaire recent de l’Ouest Cameroun: mise en evidenceKutzbach, J.E. & Street-Perrott, F.A. (1985) Milankovitch forcing
of fluctuations in the level of tropical lakes from 18 to 0 kyr B.P. de refuges forestiers et discussion des problemes paleoclimatiques.Mem. Trav. Ecole Pratiq. Hautes Etudes Inst. Montpellier, 17,Nature, 317, 130–134.
Kutzbach, J.E. & Webb, T. III (1993) Conceptual basis for 129–142.Maley, J. & Livingstone, D.A. (1983) Extension d’un elementunderstanding Late-Quaternary climates. Global climates since
the Last Glacial Maximum (ed. by H.E. Wright Jr, J.E. Kutzbach, montagnard dans le sud du Ghana (Afrique de l’Ouest) auPleistocene et a l’Holocene inferieur: premieres donneesT. Webb III, W.F. Ruddiman, F.A. Street-Perrott, and P.J.
Bartlein), pp. 5–11. University of Minnesota Press, Minneapolis, polliniques. C. R. Acad. Sci. 296, 1287–1292.Maley, J., Livingstone, D.A., Giresse, P., Thouveny, N., Brenac,London.
Lamb, H.F., Eicher, U. & Switsur, V.R. (1989) An 18,000-year P., Kelts, K., Kling, G., Stager, C., Haag, M., Fournier, M.,Bandet, Y. & Zogning, A. (1990) Lithostratigraphy, volcanism,record of vegetation, lake-level and climatic change from
Tigalmamine, Middle Atlas, Morocco. J. Biogeogr. 16, 65–74. paleomagnetism and palynology of Quaternary lacustrinedeposits from Barombi Mbo (West Cameroon): preliminaryLamb, H.F., Gasse, F., Benkaddour, A., El Hamouti, N., van der
Kaars, S., Perkins, W.T., Pearce, N.J. & Roberts, C.N. (1995) results. J. Volcanic Geothermal Res. 42, 319–335.Martin, A.R.H. (1968) Pollen analysis of Groenvlei lake sediments,Relation between century-scale Holocene arid intervals in tropical
and temperate zones. Nature, 373, 134–137. Knysna (South Africa). Rev. Palaeobot. Palynol. 7, 107–144.Mbenza Muaka and Roche, E. (1980) Exemple d’evolutionLarcher, W. (1995) Physiological plant ecology. Ecophysiology and
tree physiology of functional groups, 3rd edn. Springer, Berlin. paleoclimatique au Pleistocene terminal et a l’Holocene au Shaba(Zaıre). Mem. Mus. Nat. Hist. Nat., Paris, Serie B, Bot. 27,Lawton, R.M. (1978) A study of the dynamic ecology of Zambian
vegetation. J. Ecol. 66, 175–198. 137–148.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

1026 Dominique Jolly et al.
Medus, J. (1984) Analyse polliniques des sediments holocenes du Woodwell), pp. 304–312. Oxford University Press, New York.Prentice, I.C., Sykes, M.T., Lautenschlager, M., Harrison, S.P.,lac Tanma, Senegal, Palaeoecol. Africa, 16, 255–264.
Mellilo, J., Prentice, I.C., Schulze, E.-D., Farquhar, G. & Sala, O. Denissenko, O. & Bartlein, P.J. (1993) Modelling globalvegetation patterns and terrestrial carbon storage at the last(1996) Terrestrial biotic responses to environmental change and
feedbacks to climate. Climate Change 1995: the science of climate glacial maximum. Global Ecol. Biogeogr. Letts, 3, 67–76.Prentice, I.C. & Webb III, T. (in press) Global palaeovegetationchange (ed. by J.T. Houghton, L.G. Meira Filho, B.A. Callender,
N. Harris, A. Kattenberg and K. Maskell), pp445–482. data for climate-biosphere model evaluation. GAIM Report, inpress.Cambridge University Press, Cambridge.
Munyanziza, E. & Oldeman, R.A.A. (1996) Miombo trees: Quezel, P. (1965) La vegetation du Sahara du Tchad a la Mauritanie.Masson, Paris.ecological strategies, sylviculture and management. Ambio, 25,
454–458. Reille, M. (1979) Analyse pollinique du lac de Sidi Bou Rhaba,littoral atlantique (Maroc). Ecologia Med. 4, 61–65.Neumann, K. (1987) Jebel Tageru. A contribution to the flora of
the southern Libyan Desert. J. Arid Environ. 12, 27–39. Reynaud-Farrera, I. (1995) Histoire des paleoenvironnementsforestiers du Sud Cameroun a partir d’analyses palynologiques etNeumann, K. (1989a) Holocene vegetation of the eastern Sahara:
charcoal from prehistoric sites. Afr. Archaeol Rev. 7, 97–116. statistiques de depots Holocenes et actuels. Unpublished thesis,University of Montpellier II.Neumann, K. (1989b) Zur Vegetationsgeschichte der Ostsahara im
Holozan. Holzkohlen aus prahistorischen Fundstellen (Mit einem Riser, J. (1989) Modeles Quaternaires et bilans d’erosion dans deuxplaines du bassin de Taoudenni. Abstracts of IPCG 252, Jerba,Exkurs uber die Holzkohlen von Fachi-Dogonboulo /Niger).
Afrika Praehistorica, 2, 13–181. Tunisia.Ritchie, J.C., Eyles, C.H. & Haynes, C.V. (1985) Sediment andde Noblet, N., Braconnot, P., Joussaume, S. & Masson, V. (1996)
Sensitivity of simulated Asian and African summer monsoons pollen evidence for an early to mid-Holocene humid period inthe eastern Sahara. Nature, 314, 352–355.to orbitally induced variations in insolation 126, 115 and 6 kBP.
Clim. Dyn. 12, 589–603. Ritchie, J.C. & Haynes, C.V. (1987) Holocene vegetation zonationin the eastern Sahara. Nature, 330, 645–647.Olson, J.S., Watts, J.A. & Allison, L.J. (1983) Carbon in live
vegetation of Major world ecosystems. ORNL-5862, Oak Ridge Roche, E. & Bikwemu, G. (1989) Paleoenvironmental change onthe Zaıre-Nile ridge in Burundi; the last 20 000 years: anNational Laboratory, Oak Ridge.
Pachur, H. & Braun, G. (1980) The palaeoclimate of the Central interpretation of palynological data from the Kashiru core,Ijenda, Burundi. Quaternary environmental research on EastSahara, Libya, and the Libyan Desert. Palaeoecol. Africa, 12,
351–363. African Mountains (ed. by W.C. Mahaney), pp. 231–242.Balkema, Rotterdam.Partridge, T.C., Kerr, S.J., Meltcafe, S.E., Scott, L., Talma, A.S. &
Vogel, J.C. (1993) The Pretoria Saltpan: a 200,000 year Southern Salamani, M. (1991) Premieres donnees palynologiques sur l’histoireHolocene du massif de l’Akfadou (Grande-Kabylie, Algerie).African lacustrine sequence. Palaeogeogr. Palaeoclim. Palaeoecol.
101, 317–337. Ecologia Med. 17, 145–159.Salamani, M. (1993) Premieres donnees paleophytogeographiquesPeters, J. (1988) The palaeoenvironment of the Gilf Kebir-Jebel
Uweinat area during the first half of the Holocene: The latest du Cedre de l’Atlas (Cedrus atlantica) dans la region de grandeKabylie (NE Algerie). Palynosciences, 2, 147–155.evidence. Sahara, 1, 73–76.
Petit-Maire, N. (1989) Interglacial environments in presently Schalke, H.J.W.G. (1973) The upper Quaternary of the Cape Flatsarea (Cape Province, South Africa). Scripta Geologica, 15, 1–57.hyperarid Sahara: palaeoclimatic implications. Paleoclimatology
and paleometeorology. Modern and past patterns of global Schild, R. & Wendorf, F. (1982) Recent research on the Neolithicof the Western Desert (Egypt). Africana Bull. 31, 89–92.atmospheric transport (ed. by M. Leinen and M. Sarnthein), pp.
637–661. Kluwer, Dordrecht. Schnell, R. (1977) La flore et la vegetation de l’Afrique tropicale.Gauthier-Villars, Paris.Petit-Maire, N. (1992) Environnements et climats de la ceinture
tropicale nord-africaine depuis 140 000 ans. Mem. Soc. Geol. Fr. Scholes, R.J. & Walker, B.H. (1993) An African savanna: synthesisof the Nylsvley study. Cambridge University Press.160, 27–34.
Petit-Maire, N. & Riser, J. (1988) Le Sahara a l’Holocene: Mali. Schulz, E. (1980) Zur Vegetation der ostlichen zentralen Saharaund zu ihrer Entwicklung im Holozan. Wurzburger Geogr. Arb.C.C.G.M., Paris, Institut Geographique National, 1 carte 1/
1 000 000. 51, 1–194.Schulz, E. (1987) Die Holozane Vegetation der zentralen Sahara.Prentice, I.C., Cramer, W., Harrison, S.P., Leemans, R., Monserud,
R.A. & Solomon, A.M. (1992) A global biome model based on Palaeoecol. Africa, 18, 143–161.Schulz, E. (1991) The Taoudenni-Agorgott pollen record and theplant physiology and dominance, soil properties and climate. J.
Biogeogr. 19, 117–134. Holocene vegetation history of the Central Sahara.Paleoenvironnements du Sahara. Lacs holocenes a TaoudenniPrentice, I.C., Guiot, J., Huntley, B., Jolly, D. & Cheddadi, R.
(1996) Reconstructing biomes from palaeoecological data: a (Mali) (ed by N. Petit-Maire), pp. 143–162. Edns du CNRS.Paris.general method and its application to European pollen data at
0 and 6 ka. Clim. Dyn. 12, 185–194. Schulz, E. & Whitney, J.W. (1986) Vegetation in north-central SaudiArabia. J. Arid Environ. 10, 175–186.Prentice, I.C., Harrison, S.P., Jolly, D. & Guiot, J. (1998) The
climate and biomes of Europe at 6000 yr BP: comparison of Scott, L. (1982) A late Quaternary pollen record from the TransvaalBushveld, South Africa. Quat. Res. 17, 339–370.model simulations and pollen-based reconstructions. Quat. Sci.
Rev. 17, 659–668. Scott, L. (1983) Late Quaternary palaeoenvironments in theTransvaal on the basis of palynological evidence. Sasqua IntPrentice, I.C. & Jolly, D. (in press) Biome modelling. Forward and
inverse modelling in palaeoclimatology (ed. by P.J. Bartlein, J. Symp/Swaziland/29 August-2 September 1983, 317–327.Scott, L. (1986) Pollen analysis and palaeoenvironmentalGuiot, S.P. Harrison and I.C. Prentice), NATO ASI Series, Series
I: Global Environ. Change. interpretation of late Quaternary sediment exposures in theeastern Orange Free State, South Africa. Palaeoecol. Africa, 17,Prentice, I.C. & Sykes, M.T. (1994). Vegetation geography and
carbon storage changes. Biotic feedbacks in the global climate 113–122.Scott, L. (1987a) Pollen analysis of hyena coprolites and sedimentssystem: will the warming speed the warming? (ed. by G.M.
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027

Biome reconstruction for Africa and the Arabian peninsula 1027
from Equus Cave, Taung, Southern Kalahari (South Africa). Uganda: a pollen record for the Holocene from AhakagyeziSwamp. The Holocene, 3, 324–332.Quat. Res. 28, 144–156.
TEMPO Members (1996) Potential role of vegetation feedback inScott, L. (1987b) Late Quaternary forest history in Venda, Southernthe climate sensitivity of high-latitude regions: a case study atAfrica. Rev. Palaeobot. Palynol. 53, 1–10.6000 years B.P. Global Biogeochem. Cycles, 10, 727–736.Scott, L. (1989) Late Quaternary vegetation history and climatic
Tiercelin, J.J., Vincens, A., (coord.), Barton, C., Carbonel, P.,change in the eastern Orange Free State, South Africa. S. Afr.Casanova, J., Delibrias, G., Gasse, F., Grosdidier, E., Herbin,J. Bot. 55, 107–116.J.P., Huc, A.Y., Jardine, S., Le Fournier, J., Melieres, F., Owen,Scott, L. (1994) Palynology of Late Pleistocene Hyrax middens,R.B., Page, P., Palacios, C., Paquet, H., Peniguel, G., Peypouquet,South-Western Cape Province, South Africa: a preliminaryJ.P., Raynaud, J.F., Renaut, R.W., De Reneville, P., Richert,report. Historical Biology, 9, 71–81.J.P., Riff, R., Robert, P., Seyve, C., Vandenbroucke, M. & Vidal,Scott, L. & Bousman, C.B. (1990) Palynological analysis of hyraxG. (1987) Le demi-Graben de Baringo-Bogoria, rift Gregory,middens from southern Africa. Palaeogeogr. Palaeoclim.Kenya. 30 000 ans d’histoire hydrologique et sedimentaire. Bull.Palaeoecol. 79, 367–379.Des Centres Recherche Exploration-Prod Elf-Aquitaine, 11,Scott, L. & Brink, J.S. (1991) Quaternary palaeoenvironments of249–540.pans in central South Africa: palynological and
Trochain, J. (1940) Contribution a l’etude de la vegetation du Senegal.palaeoenvironmental evidence. S. Afr. Geogr.Larose, Paris.Scott, L. & Cooremans, B. (1990) Late Quaternary pollen from a
VEMAP Members (1995) Vegetation/ecosystem modeling andhot spring in the upper Orange River Basin, South Africa. S.analysis project: Comparing biogeography and biogeochemistryAfr. J. Sci. 86, 156–165.models in a continental-scale study of terrestrial ecosystemScott, L. & Cooremans, B. (1992) Pollen in recent Procavia (hyrax),responses to climate change and CO2 doubling. GlobalPetromus (dassie rat) and bird dung in South Africa. J. Biogeogr.Biogeochem. Cycles, 9, 407–437.19, 205–215.
Vernet, R. & Striedter, K.H. (1992) La place du Djado dans leScott, L., Cooremans, B., de Wet, J.S. & Vogel, J.C. (1991).Sahara Central: Hommes et climats a l’Holocene, WurzburgerHolocene environmental changes in Namibia inferred from pollenGeogr. Arb. 84, 201–234.analysis of swamp and lake deposits.The Holocene, 1, 8–13.
Vincens, A. (1989) Paleoenvironnements du Bassin Nord-Scott, L. & Vogel, J.C. (1983) Late Quaternary pollen profile fromTanganyika (Zaıre, Burundi, Tanzanie) au cours des 13 derniersthe Transvaal Highveld, South Africa. South Afr. J. Sci. 79,mille ans: apport de la palynologie. Rev. Palaeobot. Palynol. 61,266–272.69–88.Ssemmanda, I. & Vincens, A. (1993) Vegetation et climat dans le
Vincens, A. (1993) Nouvelle sequence pollinique du LacBassin du lac Albert (Ouganda, Zaıre) depuis 13000 ans B.P.Tanganyika: 30,000 ans d’histoire botanique et climatique du
Apport de la palynologie. C. R. Acad. Sci. 316, 561–567.Bassin Nord. Rev. Palaeobot. Palynol. 78, 381–394.
Straka, H. (1991) Palynologia Madagassica et Mascarenica- 2emeVincens, A., Chalie, F., Bonnefille, R., Guiot, J. & Tiercelin, J.J.
partie, Teil 2. Tropische Subtropische Pflanzenwel,t 78, 5–43.(1993) Pollen derived rainfall and temperature estimates from
Straka, H. (1993) Beitrage zur Kenntnis der Vegetationsgeschichte lake Tanganyika and their implication for late Pleistocene watervon Madagaskar (Vorlaufige Mitteilung). Dissertationes Bot. 196, levels. Quat. Res. 40, 343–350.439–449. Walter, H. (1985) Vegetation of the Earth and Ecological Systems
Street-Perrott, F.A., Marchand, D.S., Roberts, N. & Harrison, S.P. of the Geo-biosphere, 3rd edn. Springer-Verlag, Berlin.(1989) Global lake-level variations from 18,000 to 0 years ago: Wasylikowa, K. (1992) Exploitation of wild plants by prehistorica palaeoclimatic analysis, Rep. U.S. Doe/Er/.60304–H1, TR:046. peoples in the Sahara: Wurzburger Geogr. Arb. 84, 247–262.U.S. Department of Energy Technical Report. Wendorf, F. & Schild, R. (1980) Prehistory of the Eastern Sahara,
Street-Perrott, F.A. & Perrott, R.A. (1993) Holocene vegetation, 414 pp. Academic Press, New York.lake levels and climate of Africa. Global climates since the Last White, F. (1983) The vegetation of Africa. A descriptive memoirGlacial Maximum (ed. by H.E.Wright Jr, J.E. Kutzbach, T. Webb to accompany the UNESCO/AETFAT/UNSO vegetation map ofIII, W.F. Ruddiman, F.A. Street-Perrott and P.J. Bartlein), pp. Africa. UNESCO, Paris.318–356. University of Minnesota Press. Wickens, G.E. (1975) Changes in the climate and vegetation of the
Talbot, M.R., Livingstone, D.A., Palmer, P.G., Maley, J., Melack, Sudan since 20 000 B.P. Boissiera, 24, 43–65.J.M., Delibrias, G. & Gulliksen, S. (1984) Preliminary results Wright, H.E. Jr, Kutzbach, J.E., Webb, T. III, Ruddiman, W.F.,from sediment cores from lake Bosumtwi, Ghana. Palaeoecol. Street-Perrott, F.A. & Bartlein, P.J. (1993) Global Climates sinceAfrica, 16, 173–192. the Last Glacial Maximum. University of Minnesota Press.
Tarasov, P.E. & 28 authors (1998) Present-day and mid-Holocene Yu, G., Prentice, I.C., Harrison, S.P. & Sun, X. (1998) Pollen-basedbiomes reconstructed from pollen and plant macrofossils from the biome reconstructions for China at 0 ka and 6 ka. J. Biogeogr.Former Soviet Union and Mongolia. J. Biogeogr. 25, 1029–1053. 25, 1055–1969.
Taylor, D.M. (1990) Late Quaternary pollen records from two Zinderen Bakker, E.M. (1982) Pollen analytical studies of theUgandan mires: evidence for environmental change in the Rukiga Wonderwerk cave, South Africa. Pollen et Spores, 24, 235–250.Highlands of Southwest Uganda. Palaeogeogr. Palaeoclim. Zinderen Bakker, E.M. & van and Coetzee, J.A. (1972) A re-Palaeoecol. 80, 283–300. appraisal of Late-Quaternary climatic evidence from Tropical
Africa. Palaeoecol. Africa, 7, 151–181.Taylor, D.M. (1993) Environmental change in montane southwest
Blackwell Science Ltd 1998, Journal of Biogeography, 25, 1007–1027


















