
Biol 724 - Biosystemsfishermj/local_html/biol724-metabolic-pathways.pdf · Biol 724 - Biosystems...
23
Biol 724 - Biosystems Control of Enzymes in Metabolic Pathways
Transcript of Biol 724 - Biosystemsfishermj/local_html/biol724-metabolic-pathways.pdf · Biol 724 - Biosystems...

Biol 724 - Biosystems
Control of Enzymes in Metabolic Pathways

Feedback Inhibition
The first example of feedback inhibition identified in ametabolic pathway concerned the inhibition of the firstenzyme in the biosynthetic pathway to the amino acidisoleucine.
Threonine deaminase is specifically and strongly inhibited byisoleucine.
The structurally similar amino acids leucine and valine arenot effective inhibitors of the enzyme.
Feedback loops on the first enzyme of a pathway are acommon structural feature of metabolism.
Substrate Intermediate-1 Intermediate-2 Intermediate-3 ProductEnzyme-1 Enzyme-2 Enzyme-3 Enzyme-4
Inhibit ion

Biosynthetic pathways are often branched - the early part ofthe pathway leads ultimately to two or more end products:
The regulation of branched pathways has to cope withdifferential variations in the net demand for the different endproducts. There are two basic models for feedback control insuch branched pathways - nested and sequential feedback.
Substrate Intermediate-1
Intermediate-2 Intermediate-3 Product-1
Intermediate-4 Intermediate-5 Product-2
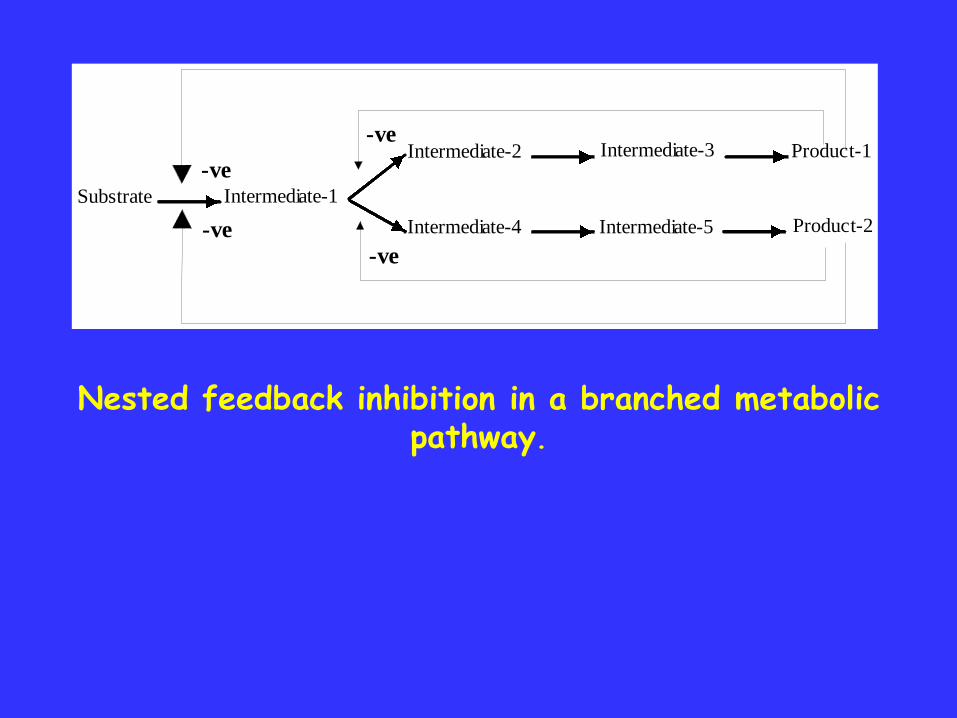
Substrate Intermediate-1
Intermediate-2 Intermediate-3 Product-1
Intermediate-4 Intermediate-5 Product-2
-ve
-ve
-ve
-ve
Nested feedback inhibition in a branched metabolic pathway.
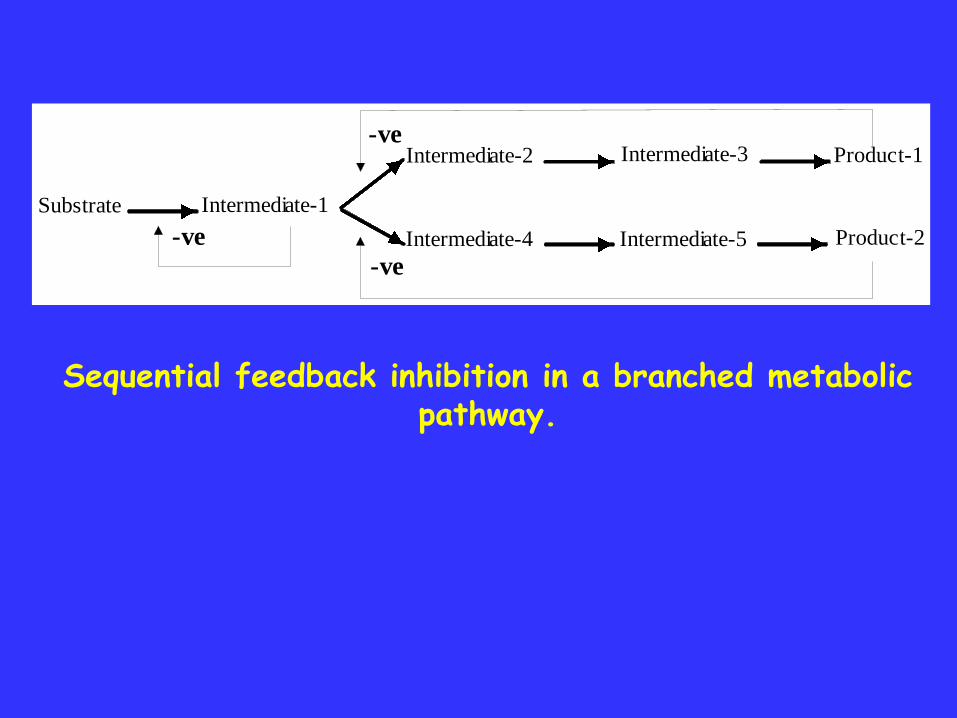
Substrate Intermediate-1
Intermediate-2 Intermediate-3 Product-1
Intermediate-4 Intermediate-5 Product-2
-ve
-ve
-ve
Sequential feedback inhibition in a branched metabolic pathway.
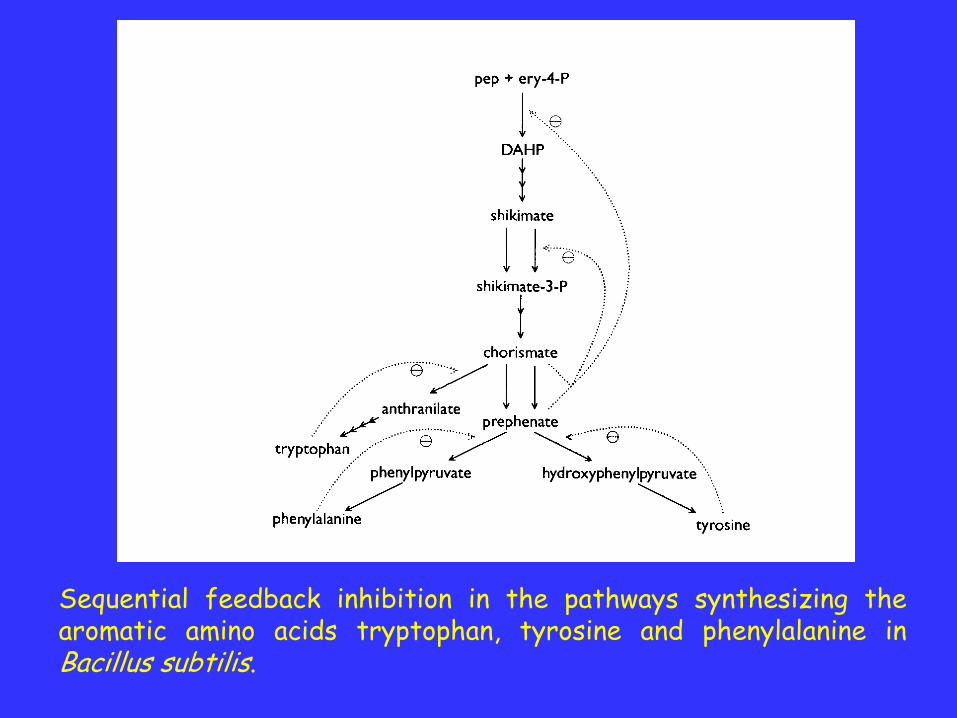
Sequential feedback inhibition in the pathways synthesizing thearomatic amino acids tryptophan, tyrosine and phenylalanine inBacillus subtilis.

The final common intermediate of this pathway is amixture of chorismate and prephenate. Theseintermediates can inhibit the first step in their synthesis,the joining of phosphoenolpyruvate and the 4-carbon sugarerythrose 4-phosphate.
In contrast, none of the aromatic amino acids inhibit thisstep. Each of them specifically inhibits the first step of itsown branch.
Sequential systems like this are regarded as beingreliable in that they generally behave as required when thesource and end product metabolites vary.
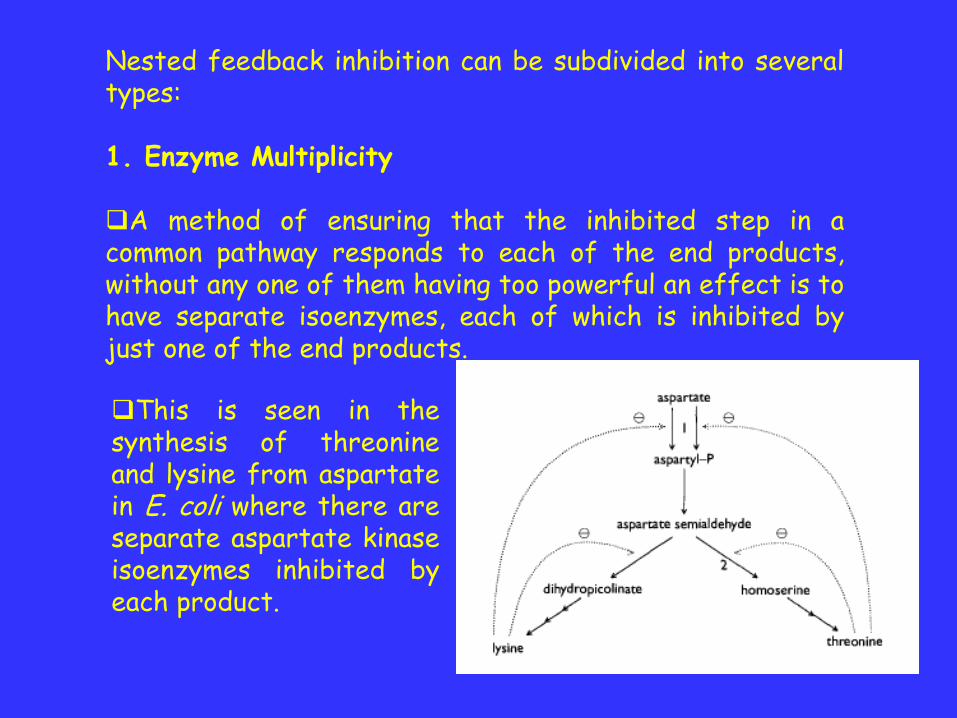
Nested feedback inhibition can be subdivided into severaltypes:
1. Enzyme Multiplicity
A method of ensuring that the inhibited step in acommon pathway responds to each of the end products,without any one of them having too powerful an effect is tohave separate isoenzymes, each of which is inhibited byjust one of the end products.
This is seen in thesynthesis of threonineand lysine from aspartatein E. coli where there areseparate aspartate kinaseisoenzymes inhibited byeach product.
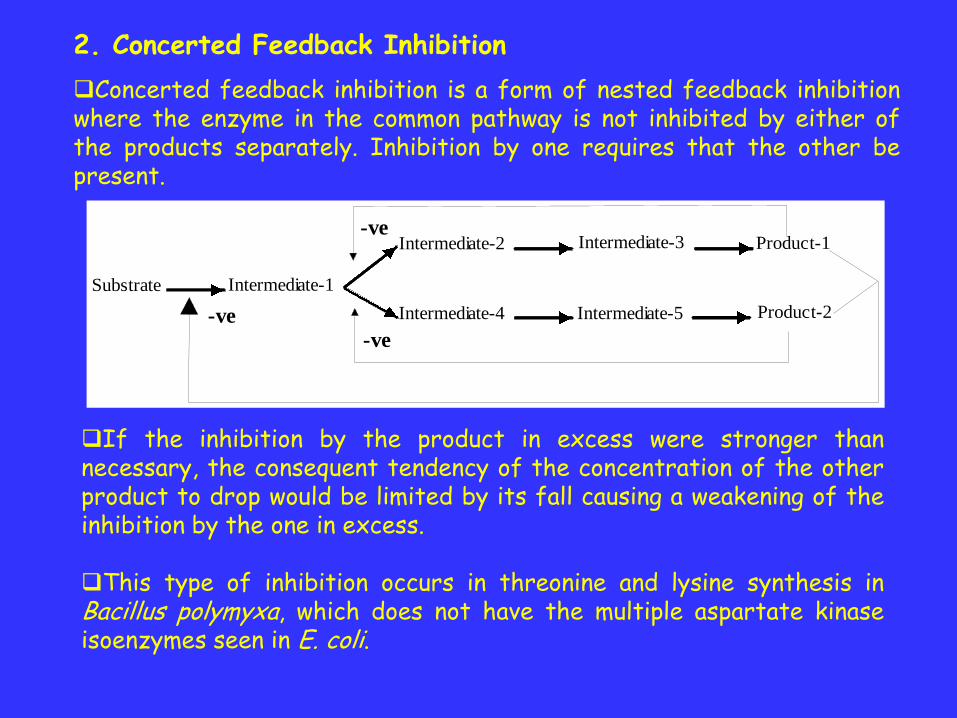
2. Concerted Feedback Inhibition
Concerted feedback inhibition is a form of nested feedback inhibitionwhere the enzyme in the common pathway is not inhibited by either ofthe products separately. Inhibition by one requires that the other bepresent.
Substrate Intermediate-1
Intermediate-2 Intermediate-3 Product-1
Intermediate-4 Intermediate-5 Product-2
-ve
-ve
-ve
If the inhibition by the product in excess were stronger thannecessary, the consequent tendency of the concentration of the otherproduct to drop would be limited by its fall causing a weakening of theinhibition by the one in excess.
This type of inhibition occurs in threonine and lysine synthesis inBacillus polymyxa, which does not have the multiple aspartate kinaseisoenzymes seen in E. coli.

3. Cumulative Feedback Inhibition
This is nested feedback inhibition where the inhibition of thecommon enzyme in the presence of both end products is greaterthan that caused by either separately; however, this greaterinhibition is exactly that expected from the combination of theeffects of each when they act independently.
This is the case for glutamine synthase from E. coli. Glutamineis the common nitrogen-containing metabolite of a number ofdiverging pathways. Each of the end products of these pathwayshas a separate inhibition site on the enzyme, each of whichseparately causes only partial inhibition even at saturatingconcentrations of the inhibitor. However, together the effects arecumulative causing almost complete inhibition.

4. Synergistic Feedback Inhibition
Synergistic feedback inhibition is intermediate betweenconcerted and cumulative feedback inhibition.
Each end product exhibits some partial inhibition on its own, butthe inhibition by both end products together is much greater thanwould be expected for cumulative inhibition i.e. there is interactionbetween the separate inhibition sites.
Synergistic inhibition is observed in purine biosynthesis on thefirst enzyme of the common pathway to AMP and GMP. Thisallosteric enzyme has separate inhibitory sites for AMP and GMP,but both of them together interact to cause greater inhibition thanwould be expected.
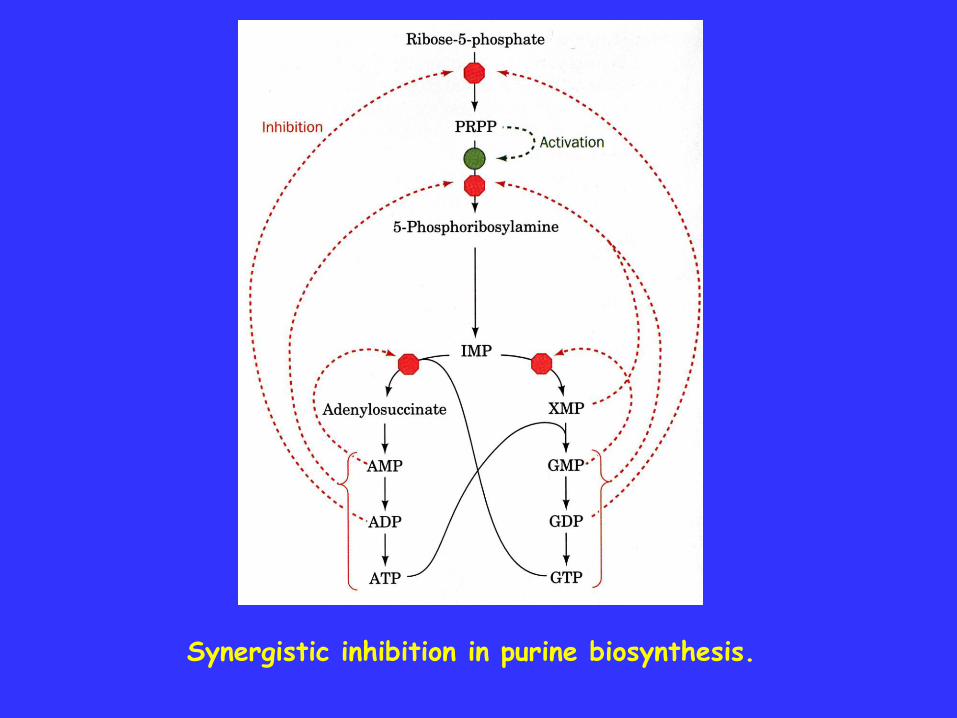
Synergistic inhibition in purine biosynthesis.
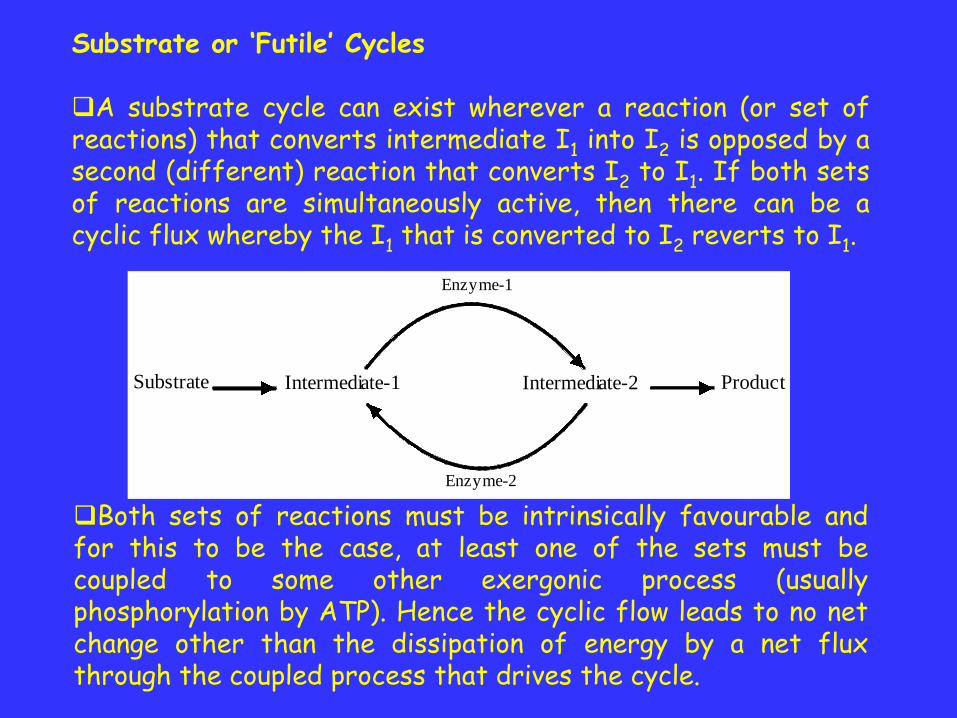
Substrate or ‘Futile’ Cycles
A substrate cycle can exist wherever a reaction (or set ofreactions) that converts intermediate I1 into I2 is opposed by asecond (different) reaction that converts I2 to I1. If both setsof reactions are simultaneously active, then there can be acyclic flux whereby the I1 that is converted to I2 reverts to I1.
Substrate Intermediate-1 Intermediate-2 Product
Enzyme-1
Enzyme-2
Both sets of reactions must be intrinsically favourable andfor this to be the case, at least one of the sets must becoupled to some other exergonic process (usuallyphosphorylation by ATP). Hence the cyclic flow leads to no netchange other than the dissipation of energy by a net fluxthrough the coupled process that drives the cycle.

The substrate cycles of glycolysis/gluconeogenesis
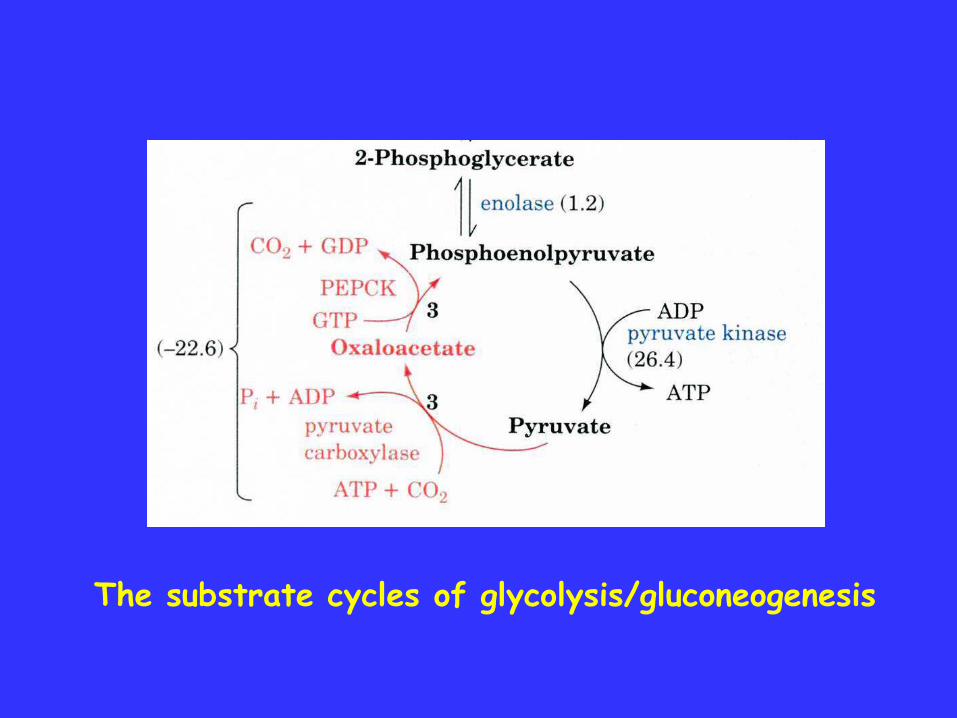
The substrate cycles of glycolysis/gluconeogenesis

Flight muscle of bumble bees. The fructose 6-phosphate:fructose 1,6-bisphosphate cycle is used by bumble bees toraise the temperature of their flight muscles in cold weather.
Insect PFK (mmol/min/g)
F 1,6 bisPase (mmol/min/g)
Bumble Bee:
Worker 56 73
Male 40 77
Queen 26 25
Honey Bee:
Worker 30 0.05
Cuckoo Bumble Bee:
Male 16 1.8
Queen 16 2.2
Sensitivity of Control
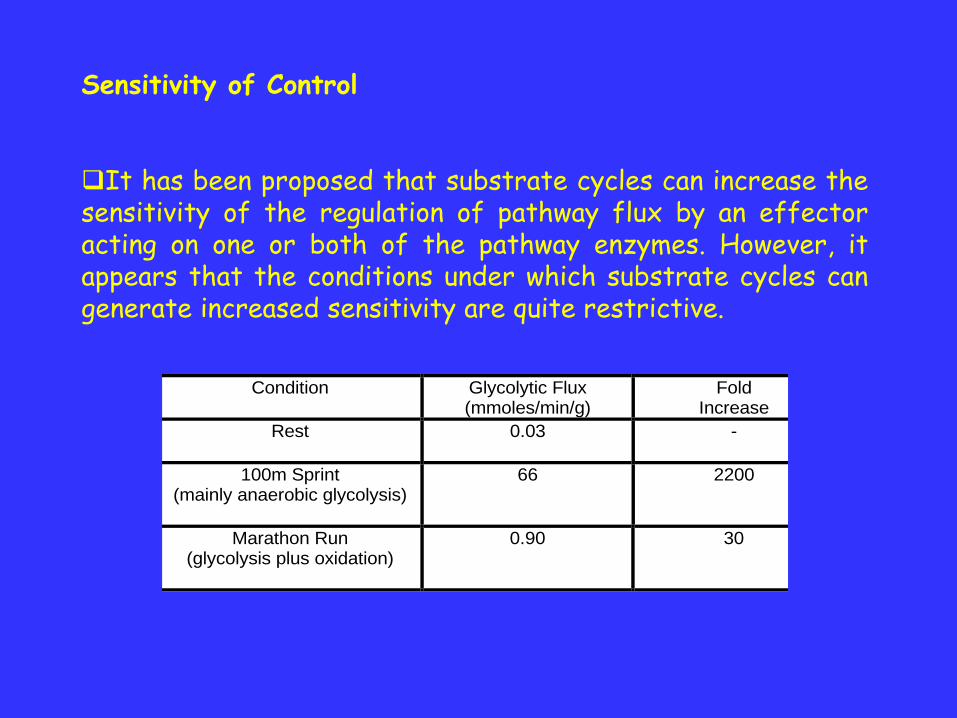
It has been proposed that substrate cycles can increase thesensitivity of the regulation of pathway flux by an effectoracting on one or both of the pathway enzymes. However, itappears that the conditions under which substrate cycles cangenerate increased sensitivity are quite restrictive.
Condition Glycolytic Flux (mmoles/min/g)
Fold Increase
Rest
0.03 -
100m Sprint (mainly anaerobic glycolysis)
66 2200
Marathon Run (glycolysis plus oxidation)
0.90 30

If a substrate cycle exists this may explain the dramaticincrease in glycolytic flux seen in the sprinter.
If flux = 0.03 at rest then:
In the sprinter at rest
Vf = 0.3 and Vr = 0.27 (i.e. net flux = 0.03)
In the marathon runner at rest
Vf = 0.04 and Vr = 0.01 (i.e. net flux = 0.03)
If at the start of a sprint Vr ~ 0 then.
For a sprinter the new net flux = 0.3
For a marathon runner the new net flux = 0.04
i.e. If there is a high rate of cycling at rest then a massivechange in flux can occur at the start of a sprint. Marathonrunners do not need this massive change in flux.

Covalent Modification of Enzymes
Irreversible cascades
There are many types of post-translational modifications ofenzymes. Some of these are irreversible and not primarilymechanisms of control e.g. proteolytic cleavage of inactiveprecursors (zymogens) is a mechanism for synthesizing and storingpotentially hazardous products such as digestive enzymes or bloodclotting factors.

Activation of blood clotting involves a cascade of zymogensin which each factor, upon activation, proteolytically cleavesthe next zymogen in the sequence, until prothrombin iscleaved to thrombin, which cleaves soluble fibrinogen to givethe clot-forming fibrin fibres.
The sequence shows amplification through using catalyststhat create catalysts, and convert the minute initiating signalgenerated following tissue injury into a major responsewithin a few seconds.
However, for the enzymes involved, it is a once-onlymechanism as the clotting can only be stopped by theirproteolytic inactivation.
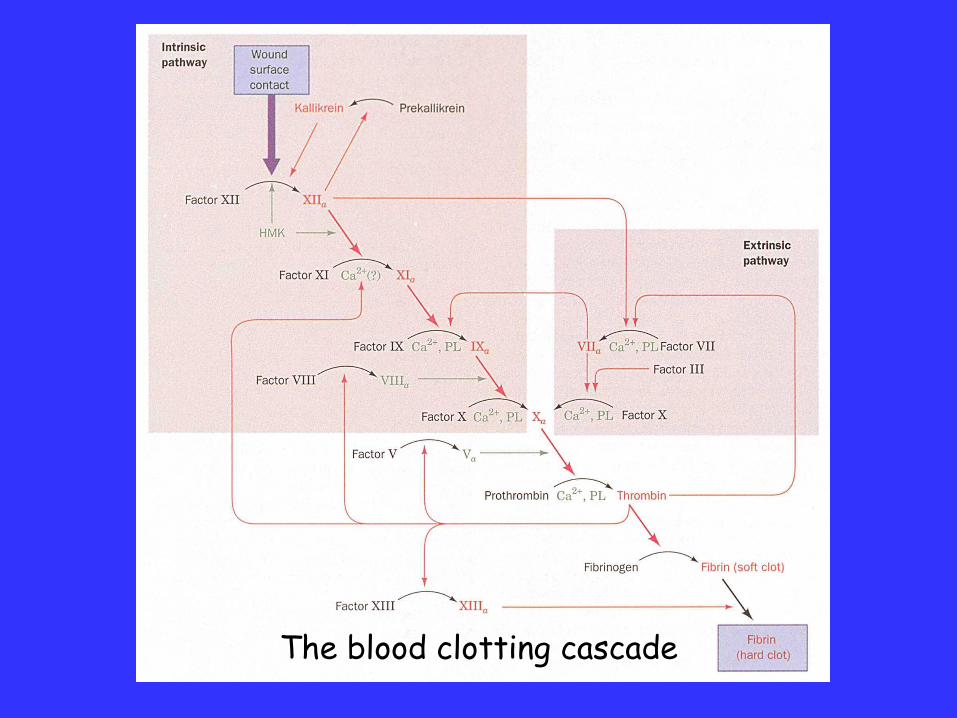
The blood clotting cascade
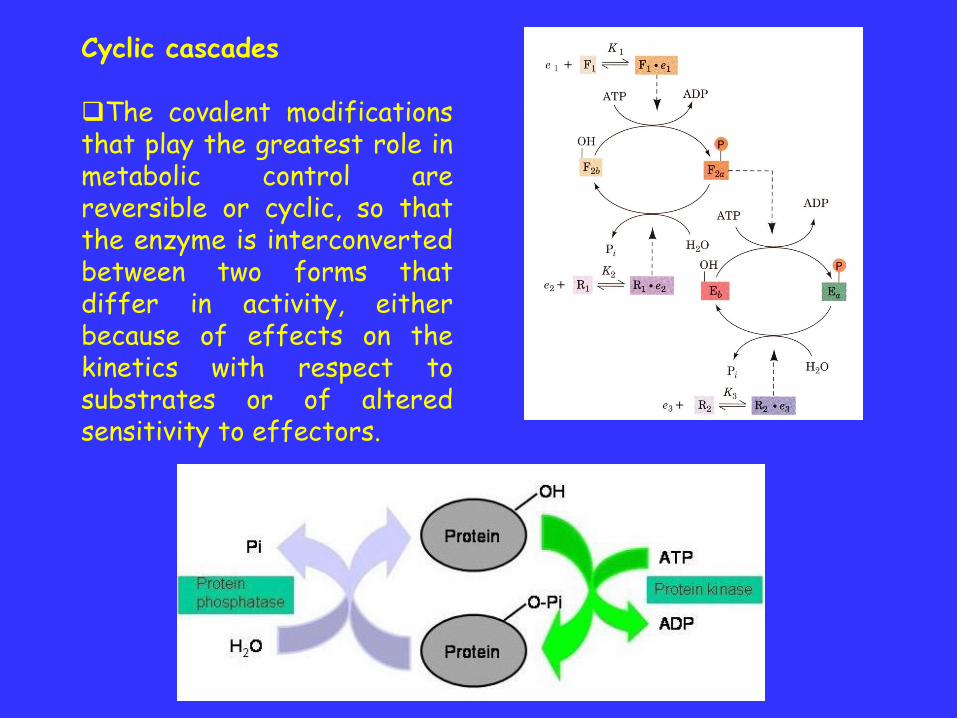
Cyclic cascades
The covalent modificationsthat play the greatest role inmetabolic control arereversible or cyclic, so thatthe enzyme is interconvertedbetween two forms thatdiffer in activity, eitherbecause of effects on thekinetics with respect tosubstrates or of alteredsensitivity to effectors.

Summary
Enzymes operate in metabolic pathways and overall control ofthese metabolic pathways can depend upon feedback inhibitionof selected enzymes be specific metabolites.
Feedback inhibition can be sequential or nested in nature.Nested feedback may be concerted, cumulative or synergistic .
The occurrence of ‘substrate’ or ‘futile’ cycles, withinmetabolic pathways, may have important implications for theoverall responsiveness of metabolic pathways to changingdemands.
Enzyme activity may also be changed by irreversible (e.g.partial proteolysis) or reversible (e.g. phosphorylation/dephosphorylation) covalent modification.


















