
BIOEN 482 Senior Capstone Thesis
49
Three-Dimensional Modeling of the Osteocyte Network by Chao Huang Advisor: Ted S. Gross, Ph.D. University of Washington Department of Bioengineering Senior Capstone Project 6 June 2008
-
Upload
chaocharliehuang -
Category
Documents
-
view
218 -
download
0
Transcript of BIOEN 482 Senior Capstone Thesis

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 1/49
Three-Dimensional Modeling
of the Osteocyte Network
by
Chao Huang
Advisor: Ted S. Gross, Ph.D.
University of Washington
Department of Bioengineering
Senior Capstone Project
6 June 2008

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 2/49
Table of Contents
ABSTRACT ................................................................................................................................ 2
INTRODUCTION...................................................................................................................... 3
DEFINITION OF PROJECT ............................................................................................................... 3MEDICAL AND SCIENTIFIC SIGNIFICANCE ..................................................................................... 4SOCIAL, ETHICAL, AND ECONOMIC ISSUES.................................................................................... 5TECHNICAL BACKGROUND ........................................................................................................... 7
Theory ..................................................................................................................................... 7 Review of literature................................................................................................................. 9 Previous relevant work in advisor’s laboratory ................................................................... 14 Outstanding technical issues at outset of project ................................................................. 17
INITIAL DESIGN OF EXPERIMENTS, TOOLS, AND DEVICES........................ 19 MATERIALS AND METHODS ........................................................................................................ 19COSTS ........................................................................................................................................ 26I NITIAL RESEARCH PLAN ............................................................................................................ 27
RESULTS.................................................................................................................................. 29
FINAL TIMELINE ......................................................................................................................... 29DATA ......................................................................................................................................... 30EXPERIMENTAL/DESIGN DECISIONS............................................................................................ 38A NALYSIS AND CONCLUSIONS.................................................................................................... 39SUGGESTIONS FOR FUTURE WORK .............................................................................................. 43
ACKNOWLEDGEMENTS.................................................................................................... 45
REFERENCES......................................................................................................................... 46
1

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 3/49
Abstract
Mechanical loading stimulates enhanced bone formation, and intercellular communication
through the gap junctions that join osteocytes (the mechanotransducers in bone) may play an
important role in bone’s response to external stimuli. Osteocytes are believed to be responsible
for maintaining the bone matrix by transmitting chemical signals induced by exogenous forces.
Assuming that such signals can travel in both transverse and longitudinal directions within bone,
I hypothesized that the three-dimensional (3-D) organization of the osteocyte network must be
heterogeneous in space in order to steer these signals in specific directions. To investigate this
theory, a MATLAB program that uses serial cross-sectional light microscope images of
hematoxylin and eosin-stained murine tibia cortical bone was designed to quantify the spatial
locations of the osteocytes and produce a 3-D representation of the osteocyte network
morphology. Each cross-sectional plane is oriented such that the positive x-direction runs lateral-
medial and the positive y-direction runs anterior-posterior; the positive z-direction runs distal-
proximal. The program automatically detects the xy-coordinates of cells in each image, and then
compares the coordinates in successive images to identify matching cells present in multiple
layers. By determining which cells are identical and which are unique in each layer, the z-
coordinates of the cells can be extrapolated, thus allowing the 3-D osteocyte network model to be
constructed. This model has shown that on average, osteocytes are closer to cells in a different
serial layer than the layer it currently resides in. Because osteocytes that are closer together are
more likely to be connected and transmit signals, my preliminary results suggest that the
longitudinal component of the 3-D osteocyte network may play a vital role in determining the
direction of signaling pathways.
2

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 4/49
Introduction
Definition of project
There is a need to develop a simple and fast method for identifying the three-dimensional (3-D)
locations of osteocytes within a piece of bone. I have designed a MATLAB program that uses
serial transverse images to automatically construct a 3-D model of the osteocyte network in
murine cortical bone. The program takes the images as inputs and automatically detects the
osteocyte locations in each cross-section through filtering means. A graphical user interface is
provided to allow the user to manually correct any mistakes that the program has made in
detecting cell locations. The user enters the current layer number for each cross-section in the
ordered series of images, and saves this information along with the xy-coordinates as separate
MATLAB files that can be loaded and accessed later. Once the necessary coordinate information
has been collected for each image, the user may then choose the desired files to be used to
construct the network model (the files must be contiguous cross-sections and in order).
The model is a 3-D scatterplot that shows the spatial locations of the osteocytes in the
user-selected layers and may be rotated in any direction to provide many different views. Upon
saving the model, the user may select a number of parameters to be calculated, including the
number of cells in each layer, the number cells in predefined sectors of each cross-section (to
determine uniformity), and the minimum distance between neighboring cells for both planar and
z-directions. Being able to perform morphological analysis by quantifying the 3-D organization
of osteocytes within the section of bone is an important step for tracing the direction of their
signaling pathways.
3

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 5/49
Medical and scientific significance
In humans, cortical (outer layer) and trabecular (spongy bone within the joint ends) bone begin to
degrade after peak bone mass is reached at approximately 30 years of age [6]. This degradation
is the result of elevated bone resorption (breakdown of bone and release of minerals) and
decreased bone formation [3]. By age 40, bone loss is estimated to be 0.3% to 0.5% per year, and
in women, menopause may elevate annual bone loss to up to 3% [6]. Based on these estimates,
an average 70-year-old woman may experience a decrease of 25% to up to 40% in bone mass
from her peak bone mass while a like-aged man may expect a decline of 10% to 15% [7]. In
addition, diseases such as osteoporosis may further reduce the bone mineral density, therefore
increasing the risk of bone fracture and posing a significant health concern for senior citizens [8].
Given these concerns, it is essential to develop novel techniques for sustaining or augmenting
bone mass throughout an individual’s lifetime.
One of the signals for bone cells to form new bones is mechanical stress, such as that
experienced during exercise [9]. Previous studies have shown that bone mass increases with
exercise, and so it is apparent that mechanical components of bone’s functional environment
have an effect on bone mass and morphology [10]. By studying how loads are anabolic to bone
and identifying specific signals responsible for inducing new bone formation, non-
pharmacologic methods using mechanical loading may be developed to inhibit osteoporosis and
promote osteogenesis [11].
A model of the osteocyte network in bone must be constructed in order to visualize where
bone cells are in relation to each other and understand how their organization dictates the
transmission of mechanical signals to induce bone formation. As will be highlighted in the
proceeding literature review, previous methods for analyzing signaling pathways in osteocyte
4

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 6/49
networks have either focused on osteocytes in a two-dimensional plane or used complicated and
expensive techniques and equipment to construct a three-dimensional (3-D) model. Thus, there is
a need to design a simple, easy, and fast computer program that will find the 3-D locations of
osteocytes fairly accurately and perform basic morphological analysis in a short amount of time.
Although there are already 3-D reconstruction programs available (e.g. ImageJ), such programs
do not offer the same capabilities of a program designed specifically for analysis of a network of
cells within bone. For example, traditional imaging methods would require the user to manually
locate where cells are, which is both laborious and time-consuming. The program that I have
designed automatically finds the locations of cells, constructs the 3-D model, and then performs
calculations related to the organization of the network of cells. Gathering such data will yield
substantial insight into how osteocytes are organized in bone and how the architecture of these
networks might play a role in the complex process of bone formation in response to mechanical
loading. Constructing an accurate model of the osteocyte network is therefore essential for
tracing the mechanotransduction signaling pathways that induce bone growth.
Social, ethical, and economic issues
Bone formation induced by mechanical stress is studied with the ultimate goal of adapting and
using mechanical loading in a clinical setting. If the ways in which bone perceives and responds
to mechanical stimuli are well understood, then such information may be used to design novel
exercise regimens that provoke a similar response in bone as mechanical loading. Exercise is a
socially accepted activity in all cultures of the world that is widely considered to promote good
health. Therefore, it is not expected that introducing a new exercise regimen specially designed
to induce bone growth will engender substantial fear or mistrust among the public. While the
public may be much more hesitant to use a new device or undergo a surgical procedure, a novel
5

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 7/49
exercise regimen should not be a difficult medical technique to encourage the public to try.
Previous studies have shown that athletes who play racquet sports have enhanced bone mass in
their serving arms [12], and these visible and tangible results should encourage subjects to
choose exercise over other pharmacological means of inducing bone formation.
Because exercise regimens present a non-pharmacological intervention for bone loss, it is
also expected that such treatments will not be as difficult to gain FDA approval for as
pharmaceutical treatments such as estrogens and bisphophonates. In addition, when the
biochemistry and mechanics of bone formation are better understood, exercise regimens will be
specially designed for the elderly to induce maximal bone formation from minimal strain
magnitudes, lessening any ethical concern for causing injury during clinical trials. Furthermore,
it may be more beneficial socially and ethically for the public to adopt a non-pharmacological
technique for enhancing bone formation since it is a tangible activity that can be controlled by
the individual, unlike reading directions and blindly taking medication without knowing what is
happening inside one’s body.
A final advantage of mechanically induced bone formation over pharmacological
treatments is that, beyond experimental and clinical testing, there is little to no cost involved. In
1995, the national healthcare cost of osteoporosis was estimated to be $13.8 billion and may
increase to as much as $240 billion over the next 50 years [13]. Most of these costs can be
attributed to either medication, which may be avoided if the exercise alternative is chosen, or
surgery, which may also be avoided if individuals begin their exercise regimens earlier in life.
With the potential social, ethical, and economic benefits of a possible low-magnitude exercise
regimen, mechanical loading holds the potential to be an effective and safe technique for
inducing new bone formation to counter the effects of aging.
6

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 8/49
Technical background
Theory
The primary mechanical function of bone is to provide support for muscles to act against and
hold the body in an upright position [14]. Bone is constantly subjected to a dynamic loading
environment through a person’s daily movements, and therefore must adapt responsively to
maintain its structure and withstand physical activity [15]. The basis for bone’s adaptation to
exogenous forces is through bone remodeling, which consists of bone resorption and new bone
formation. Bone resorption is the breaking down of bone into its minerals (such as calcium) by
osteoclasts, resulting in the transfer of such minerals and proteins to a different location via
blood [16]. New bone is then formed by osteoblasts that synthesize osteoid, an unmineralized
bone matrix composed of organic components. Some osteoblasts will become trapped within the
matrix that they lay down and differentiate into osteocytes, the most abundant cells in bone (each
spanning approximately 10 μm) [17]. In addition to being responsible for maintaining the bone
matrix, osteocytes are believed to be the mechanosensors and mechanotransducers in bone [18].
Bone adaptation requires cellular mechanotransduction, and osteocytes are affected by a
variety of mechanical factors generated by loading. These factors include strain generated across
the cells’ substrate, pressure within intramedullary cavities, and shear forces through the
canaliculi that connect the cells [11]. The conversion of these forces into a cellular response
consists of four phases: mechanocoupling, biochemical coupling, transmission of signaling, and
effector cell response. During mechanocoupling, the applied mechanical forces are transduced
into local mechanical signals perceived by osteocytes [14]. Such signals may be sensed by a
number of mechanoreceptors including mechanosensitive channels that induce membrane hyper
or depolarization, integrin proteins that span the membrane to couple the cell to its extracellular
environment, connexins that form regulated channels allowing direct exchange of small
7

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 9/49
molecules between adjacent cells, and membrane structure proteins that facilitate transmembrane
communication as well as provide docking positions for signaling complexes [11]. After these
local mechanical signals are sensed, biochemical coupling occurs, when the mechanical signals
are transduced into biochemical signals that lead to gene expression or protein activation [14].
Biochemical signaling may occur through G-proteins, calcium transients, MAPK activation, and
release of nitric oxide [11].
Cell-to-cell transmission of biochemical signals occurs intracellularly through the
processes that join osteocytes as well as through the extracellular fluid in which osteocytes are
immersed [19]. Gap junctions at the tip of the cell processes that contain hemichannels provide a
direct and efficient mechanism for intracellular signaling pathways [20], although osteocytes also
remain in contact via their common environment in the contiguous bone fluid space. Such fluid
acts as a coupling medium through which mechanochemical signals may still be transmitted by
hydraulic conductivity, pressure and osmotic gradients, as well as electromechanical and
acoustic energy effects [15]. The combination of the intracellular and extracellular transmission
of signals induces an effector cell response at the tissue level, at which point bone remodeling
commences [14].
The degree of bone adaptation in response to external loading depends on strain
magnitude, distribution, duration, frequency, bone history, and type of stress created
(compression, tension, or shear). Over the last century, numerous experiments have been
conducted to test each of these factors’ effect on bone adaptation and many common threads
have emerged [21]. The following literature review highlights some of these findings and ends
with a summary of the research on bone adaptation being carried out in my current advisor’s
laboratory.
8

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 10/49
Review of literature
The notion that the form and function of bone is produced and maintained by mechanical forces
was first popularized by anatomist Julius Wolff in his 1892 treatise, The Law of Bone
Remodeling [22]. In what has now become known as Wolff’s Law, Wolff states that “Every
change in the form and function of bone […] is followed by certain definite changes in their
internal architecture, and equally definite alteration in their external conformation, in accordance
with mathematical laws [23].” Through demonstrating a definite link between trabecular bone
architecture and the functional stresses placed upon it, Wolff was able to assert that the stresses
surrounding bone forces the architecture inside living bone to continuously adapt through
remodeling. Later, embryologist Wilhelm Roux generalized the notion of functional adaptation,
suggesting that mechanical stimuli govern effector cells that regulate bone’s formation and
adaptation locally in a self-organizational process [22]. In the early 20th Century, biologist
D’arcy Thompson proposed that the condition of strain, caused by the stress induced by
mechanical forces, is a direct stimulus for bone growth and thus is the source of functional
adaptation. In the 1960’s, orthopedist Harold Frost asserted that not only was mechanical strain
the primary determinant of bone adaptation, but that a minimum strain threshold must be reached
before bone adaptation occurs [21]. The combination of all these findings provided a foundation
for other investigators to study the relationship between bone’s mechanical environment and its
effect on bone’s development of mass and architecture.
Early investigations that have used approaches such as exercise, disuse, and stress
protection employed loading regimens superimposed on existing effects of normal load-bearing,
and thus an accurate, discernable remodeling response to the newly applied mechanical situation
could not be extracted [1]. Such difficulties led to the design of novel experimental setups that
could elucidate bone’s functional adaptation to mechanical forces. A new technique in which
9

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 11/49
bones were externally loaded in vivo over a period
of weeks was first used by Hert et al. in early
1970’s studies with rabbits. Hert found that dynamic
strains increased the amount of bone formation in
the rabbits while static strains did not, and therefore
suggested that dynamic strains are primarily
responsible for bone adaptation [21]. In 1984,
Rubin and Lanyon refined Hert’s loading approach and confirmed his findings by functionally
isolating the external ulnae in turkeys and applying known intermittent loads over a period of six
weeks. As Figure 1 illustrates, a template was clamped and pinned to the ulna, allowing two
parallel transverse osteotomies to be performed and leaving the entire midshaft of the ulna
undisturbed. The ends of the pins were connected to a loading apparatus that imposed a 0.5-Hz
intermittent compressive load cycle to engender strains in the bone. The number of consecutive
load cycles that was applied each day was varied to determine the bone’s response to different
load magnitudes. The strains were measured at the surface of the bone using strain gauges and
could be directly correlated to the remodeling induced by the loading. Photon-absorption
densitometry and postmortem histological methods were used to comparatively assess bone
mineral content and determine the course of remodeling in response to the strain stimulus [1].
Figure 1: Schematic of template used for parallel osteotomies and holding the ulna in place as loads were administered [1].
In comparisons of unloaded, control bone and dynamically loaded bone with a variable
number of consecutive load cycles, it was found that bone mass decreased in ulnae that were not
subjected to any external loading, which was the predicted response to disuse. As Figure 2
shows, ulnae subjected to four loading cycles a day showed a slight increase in bone mineral
content, and as the number of consecutive cycles was increased to 36 cycles of an identical strain
regimen, substantial subperiosteal and endosteal new-bone formation was measured [1].
10

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 12/49
Interestingly, ulnae subjected to 360 and
1800 cycles of loading did not have
significantly higher bone mineral content that
those subjected 36 cycles, demonstrating that
an increased duration of loading does not
yield a proportional increase in bone mass,
but rather causes the bone formation
response to saturate [21]. A likely
explanation for this behavior is that bone
cells are triggered to form new bone by a
strain-specific stimulus instead of responding
to a non-specific (reparative) effect, and a
small exposure to this stimulus appears to be
enough to induce engender new bone formation. This result was important, especially clinically,
as it suggested that the adaptive response of bone can be engendered by short, infrequent loads
rather than requiring a long period of repetitive ac
Figure 2: Percentage change in bone mineral content produced by a variable number of loading cycles per day[1].
tivity [1].
In a later study, Rubin and Lanyon modified their experiment to apply 100 consecutive 1-
Hz load cycles daily over eight weeks to engender variable strain magnitudes within the ulna
bone. By comparing the cross-sectional areas of the loaded and unloaded ulna midshafts, it was
found that newly formed bone area was proportional to the applied strain magnitude, as
illustrated by Figure 3. These results suggested that the main function of bone’s adaptive
remodeling is to produce a mass and orientation of bone tissue that is optimal for withstanding
the strain that the bone is subjected to. Mechanical strain therefore appears to be a natural
stimulus that induces an increase in bone mass not normally countered by bone resorption
11

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 13/49
activity [2]. In 1994, Rubin and McLeod then
studied the effect that strain frequency has on
bone adaptation by employing three different
loading protocols over eight weeks: (1) no
loading (disuse), (2) disuse followed by 100
seconds of 1-Hz low-magnitude mechanical
stimulus daily, and (3) disuse followed by 100
seconds of 20-Hz mechanical stimulus at a
similar strain magnitude. It was found that
disuse alone caused a mean 8.3% loss of bone
as there was no longer a stimulus to induce bone formation to negate resorption activity. The
daily 1-Hz low-amplitude mechanical strain stimulation, however, caused a mean 28% increase
in area, and importantly, the 20-Hz protocol caused bone growth to increase to 69%. These
results suggested that not only can a brief and low-magnitude stimulus induce bone formation,
but also that bone adaptation and formation are sensitive to loading frequency [1]. The dynamic
loading experiments conducted by Rubin et al. to elucidate the effects that strain rate, frequency,
and magnitude have on inducing new bone formation paved the way for others to study the
physiology and biochemistry behind bone’s adaptive response to mechanical stimuli.
Figure 3: Change in cross-sectional area of the ulnamidshaft from comparison between loaded and unloaded bone [2].
The most important aspect of studying bone’s adaptive response is to understand how
osteocytes communicate with each other to induce new bone formation, and the most
fundamental need for achieving this goal is to image where osteocytes are in relation each other.
However, osteocytes are encased in a hard, mineralized matrix in the inner cavity of bone, thus
making the task of imaging them and describing cell-to-cell signaling pathways in three-
dimensions (3-D) extremely difficult [24]. Therefore, previous methods of imaging have only
12

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 14/49
been capable of showing the osteocyte distribution in a two-dimensional (2-D) plane. For
example, Krempien et al. revealed the non-viable cell network in 2-D by staining lacunae (small
spaces in the bone that contain one osteocyte each) with fuchsin stain [25]. Such a network
model is not sufficient for analyzing the direction of signaling pathways as the longitudinal
component along the length of the bone is not included.
In efforts to move towards 3-D imaging, Palumbo et al.
were able to gather morphological data of the 3-D structure of
osteocytes by cutting transverse sections of bone serially in thin
slices and visualizing with a transmission electron microscope
[26]. Although such methods accurately depict the locations of
osteocytes, they do not offer the means to perform morphometric
analysis on osteocyte processes, the primary means of intercellular
communication in bone. Recently, Sugawara et al. conducted
extensive analyses of the osteocyte network morphology by using
3-D reconstructed fluorescent images [4]. As shown in Figure 4,
the osteocytes were stained with Texas-Red® phalloidin, which
binds to the actin filaments present in osteocyte processes [27].
The 3-D osteocyte structure was imaged on a confocal laser
scanner microscope (CLSM), which allowed for the construction
of a model to calculate the length of processes, volume, and surface area of osteocytes. This was
the first time that 3-D morphology and morphometry analysis was performed on bone [4].
However, given the amount of time needed to prepare the bone sections for staining and imaging
with an expensive and complex piece of machinery such as the CLSM, in addition to the
Figure 4: 3-D reconstruction of osteocyte network shown from periosteal side of bone. Top:arrows show osteocytes, bar = 20
μm. Bottom: zoomed-view, bar =
10 μm [4].
13

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 15/49
laborious task of manually collecting morphological data, it is evident that there is a need to
develop a simpler and faster way of analyzing the 3-D osteocyte network.
Previous relevant work in advisor’s laboratory
The laboratory group that I have joined, the
Orthopaedics Science Laboratories (OSL)
directed by Professor Ted S. Gross, examines
how bone responds to physical stimuli. Many of
the aforementioned methods for loading and
imaging have been adapted for investigation and
modeling of real-time cell-to-cell signaling in
response to mechanical loading.
Figure 5: Schematic of the noninvasive murine loadingdevice. The mouse’s right tibia is secured at themetaphysis by a gripping cup attached to the adjustablemedial support. A computer-controlled linear forceactuator applies small forces to the distal tibialmetaphysic [3].
To study the effects of loading, a novel, Gross et al. designed a noninvasive device that
allows for controlled external loads of various waveforms to be applied to murine tibia (Figure
5). In previous experiments, the tibia of the mouse was positioned against the fixed lateral
support, and the shielded linear electromagnetic actuator applied a load waveform via a digital
analog interface that enables programmable waveforms. After the mice were euthanized using
carbon dioxide, cross-sections of the loaded bone (approximately 30 μm thick) were used to
examine how the tibia’s cross-sectional area had changed.
Results have shown that the loading elicits a significant
.7% increase in cortical width for a loaded right tibia
ompared to the unloaded left tibia from the same mouse. In
augmentation of the bone’s midshaft area. Figure 6 shows a
33
c
Figure 6: Cross-sections of anunloaded tibia (left) and amechanically loaded tibia (right).The arrow indicates a 33.7%increase in cortical width of theloaded tibia [3].
14

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 16/49
addition, Gross et al. confirmed Rubin and Lanyon’s observation that dynamic loading induced
substantial new bone formation whereas static loading produced an absence of response. It was
concluded that the mechanical loading device is capable of noninvasively inducing an anabolic
response to mechanical load consistent with aforementioned models of bone adaptation [3].
More recent investigations of the OSL have
focused
not have enough time to recover from inertial damping
at a non-loaded 10-
on using low-magnitude mechanical loading
with rest intervals inserted between each load cycle. As
was explained previously, it is hypothesized that the
fluid flow near osteocytes underlies mechano-
transduction in bone and that this flow past the cell
bodies and processes is highly viscuous. If bone is
repetitively loaded, then osteocyte level fluid flow may
effects between each load cycle, and therefore the
effects of loading-induced bone formation may be
reduced after the first few cycles of repetitive loading. It w
second rest interval inserted between each loading cycle (Figure 7) would allow sufficient
recovery from inertial damping effects and elevate fluid flow near osteocytes, thus increasing the
osteogenic potential of a low-magnitude mechanical loading regimen. To verify this hypothesis,
three loading regimens were tested: (1) a low-magnitude 1-Hz waveform, (2) a high-magnitude
1-Hz waveform (twice as large as the low-magnitude waveform), and (3) a low-magnitude 1-Hz
waveform with a 10-second interval of rest inserted between each load cycle. Results showed
that while low-magnitude loading induced minimal bone-forming activity, the bone formation
rate was significantly enhanced when a 10-second rest interval was inserted between each load
Figure 7: First 12 s of (A) standard loadingwaveform and (B) rest-inserted waveformwith identical magnitude and total cyclenumber, differing only by the 10-s pauseinserted between each loading cycle of the rest
inserted waveform.[5]
as hypothesized th
15

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 17/49
cycle, transforming the ineffective low-magnitude regimen into a potent anabolic stimulus. As
Figure 8 illustrates, the rest-inserted loading significantly enhanced bone formation despite a 10-
fold reduction in the number of load cycles daily, and
was also statistically comparable to the induced bone
formation of rate of the high-magnitude loading
regimen that was 10-fold greater in cycle number and
2-fold greater in load magnitude and rate. These
results suggest great promise for rest-inserted loading
to be used as a potential strategy to build bone mass
via exercise [5].
anotransduction that occurs during induced bone
formation, the OS
Figure 8: Periosteal bone formation rates for control (intact) and experimental bone induced
by low-, high-, and rest-inserted mechanicalloading [5].
To better understand the mech
L has developed a computer simulation of real-time cellular signaling induced
by mec
nged in an idealized
hanical stimuli using agent based modeling (ABM). ABM is a technique used to explore
the wide range of consequences of introducing a stimulus to the agent or local level. Through a
set of user-specified rules, ABM provides a method for multi-scale modeling of complex global
behaviors. The OSL’s current model is governed by three cellular functions: (1) threshold levels
of bone tissue strain (the change in length of the bone due to a bending stimulus) required to
activate osteocytes, (2) propagated activity to neighboring cells for strains above threshold, (3)
modulated activity dependent on rates at which molecular stores are replenished––caused by
insertion of unloaded rest between load cycles. The model has confirmed that rest-inserted
stimuli can induce enhanced and sustain signaling within the osteocyte network by enabling
molecular stores to be replenished before the cells are activated again [28].
The OSL is now focused on fine-tuning and adding more depth to the ABM’s
capabilities. So far, the model has assumed that osteocytes are spatially arra
16

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 18/49
networ
hnical issues at outset of pro ject
my original capstone proposal, I aimed to not only find the 3-D locations of the osteocyte
er processes. I proposed to use a
k within cortical bone [28]. Thus, the model would be greatly improved if a more accurate
description of where exactly osteocytes are located within a volume of bone was provided. My
project makes it possible for the 3-D locations of osteocytes to be found automatically and
quickly, therefore introducing the opportunity of incorporating this 3-D network of cells into the
ABM simulations.
Outstanding tec
In
bodies, but also show the interconnectivity of their long, slend
confocal laser scanning microscope to image bone sections stained with the fluorescent
phallotoxin phalloidin that binds at the interface between F-actin subunits in the osteocyte
processes [29], much like Sugawara et al.’s method described previously [4]. However, when
using the Differential Interference Contrast mode on the confocal laser scanning microscope
(CLSM), I found the fluorescent stain was not concentrated in the osteocyte cell bodies, but
rather was spread out throughout all regions of the imaging plane. I concluded that this result
must suggest that the phalloidin was not binding specifically to the actin in osteocytes, and
proceeded to spend the majority of the Summer and Autumn quarters of 2007 altering and testing
my staining protocol. After employing many changes that still did not produce improved images,
I contacted Invitrogen (the company that supplied the phalloidin) for product support. Invitrogen
suggested that the reason why the phalloidin was non-specifically binding the osteocytes is
because the actin filaments in the processes were being denatured by the incubation step in my
procedure, and recommended that I try cryo-sectioning instead. After considering the amount of
work left to finish and the decreasing amount of time left, I decided that attempting to learn, use,
and potentially still be unsuccessful with cryo-sectioning was not an idea worth pursuing.
17

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 19/49
Therefore, I settled on using light microscopy to image my bone sections with hematoxylin and
eosin (H&E) staining, a simpler and faster protocol that the OSL had had success with in the
past.
Hematoxylin is a dye that colors basophilic structures containing nucleic acids blue while
the alcohol-based acidic eosin is a dye that colors eosinophilic structures composed of
intracellular or extracellular protein pink. Thus, in a bone-cross section, the nuclei of the
osteocytes would appear as blue while its surrounding regions would appear pink, allowing the
locations of the cells to be easily seen [30]. The disadvantage of using H&E stain is that it is not
fluorescent and thus incompatible with the CLSM. Using the z-motor feature of the CLSM, serial
transverse images could be taken at predetermined intervals of a thick section of bone stained
with phalloidin. With H&E and light microscopy, however, these individual thin transverse
sections must be physically cut in order to be stained and imaged. Such a procedure creates an
image registration problem, as each imaged section will be misaligned relative to others, thus
creating difficulties in comparing the coordinates of osteocyte locations between sections.
Addressing this technical issue was the main challenge in writing my MATLAB program for
creating the 3-D osteocyte network model, and the design process is detailed in the proceeding
Methods section.
18

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 20/49
Initial Design of Experiments, Tools, and Devices
Materials and methods
A section of bone taken from the midshaft of an unloaded (control) left murine tibia was
decalcified and embedded in paraffin blocks. Five serial transverse sections, each 5 μm thick,
were machine-cut from the block, mounted on glass slides, and stained with a commercially
available hematoxylin dye and a prepared eosin solution following the proceeding protocol. A
stock solution of eosin was made from 1 g Eosin Y dye, 20 mL deionized water, and 80 mL 95%
ethanol (EtOH), and the working solution of eosin was made from 25 mL stock solution, 75 mL
80% EtOH, and 0.5 mL Glacial Acetic Acid.
H&E Staining Protocol for Paraffin-Embedded Sections
1. De-paraffinization and rehydrationa. Wash in 3 changes of xylene (5 min. each). Blot excess xylene. b. Wash in 100% EtOH (5 min.)c. Wash in 95% EtOH (3 min.)
d.
Wash in 80% EtOH (3 min.)e. Wash in 70% EtOH (3 min.)f. Wash in deionized water (5 min.)
2. Hematoxylin staininga. Stain with hematoxylin (5 min.) b. Wash in tap water to allow stain to develop (5 min.)c. Wash in deionized water (5 min.). Blot excess water.
3. Eosin staining and dehydrationa. Stain with eosin working solution (30 sec.) b. Wash in 70% EtOH (5 min.)c. Wash in 2 changes of 95% EtOH (1 min. each)d. Wash in 2 changes of 100% EtOH (3 min. each). Blot excess EtOH.e. Wash in xylene (15 min.)
4. Coverslip slides using Permount (xylene-based).
19
7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 21/49
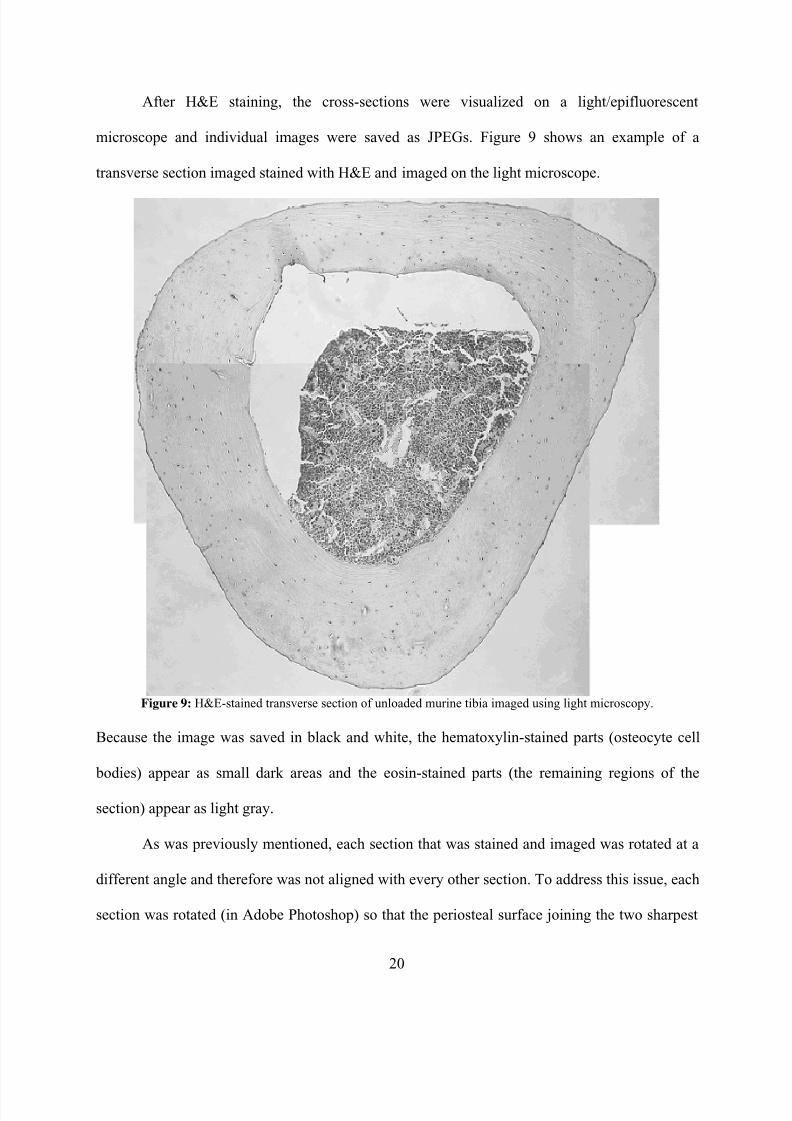
After H&E staining, the cross-sections were visualized on a light/epifluorescent
microscope and individual images were saved as JPEGs. Figure 9 shows an example of a
transverse section imaged stained with H&E and imaged on the light microscope.
Figure 9: H&E-stained transverse section of unloaded murine tibia imaged using light microscopy.
Because the image was saved in black and white, the hematoxylin-stained parts (osteocyte cell
bodies) appear as small dark areas and the eosin-stained parts (the remaining regions of the
section) appear as light gray.
As was previously mentioned, each section that was stained and imaged was rotated at a
different angle and therefore was not aligned with every other section. To address this issue, each
section was rotated (in Adobe Photoshop) so that the periosteal surface joining the two sharpest
20

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 22/49
corners was 15° off of the vertical plane, as Figure 10 illustrates. This aligns each cross-section
in an anatomically oriented position and makes consistent analysis possible. In addition, the
background of the image was removed and replaced with a solid black background to isolate the
bone and facilitate image-filtering steps later.
Figure 10: Transverse section rotated 15° from the vertical plane.
Next, a solid white mask of the bone area was created in Photoshop for each section. The H&E-
stained image and the mask for each section were then loaded and saved in MATLAB as
intensity image matrices. Using the ‘regionprops’ function, the center of mass of each of the
masks could be calculated, as shown in Figure 11. Because each section was only 5 μm apart, the
bone morphology should not have changed significantly between sections, and therefore each
section should have had a similar center of mass relative to the dimensions of its own image. To
verify that all sections had approximately the same shape (and thus center of mass), the moments
21

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 23/49
of area were calculated for each of the five sections and are listed in Table 1. The moment about
the x axis was calculated as and the moment about the y axis was calculated as
. The moments were similar for all sections, with the coefficient of variation (mean
divided by standard deviation) for I x being 0.80% and the coefficient of variation for I y being
1.90%. This confirmed that each transverse section had a similar shape and was rotated correctly
in alignment with the other sections.
∫= dA y I x2
∫= dA x I
y
2
Center of mass
Figure 11: Solid mask of transverse section showing the center of mass of its area (green dot).
Table 1: Moments of area of each transverse section.
Layer Ix (μm4) Iy (μm4)
1 4.0262E+11 4.9522E+11
2 4.0704E+11 5.1149E+11
3 4.0264E+11 4.9051E+11
4 4.0490E+11 5.0130E+11
5 3.9834E+11 5.1173E+11
Mean 4.0311E+11 5.0205E+11
SD 3.2351E+09 9.5289E+09
CV 0.80% 1.90%
22

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 24/49
To automatically find the locations of osteocyte cell bodies in the H&E-stained images,
each image was put through a series of filters. First, a threshold value was determined for the
color (shade of black) differentiating the cell bodies (darkened nuclei) and the rest of the
intracellular and extracellular protein (lighter gray colors). The edges of the cell bodies were then
detected using the threshold value and the area of each osteocyte body was delineated. Finally,
each outline of an osteocyte body was dilated and filled in, thus enlarging the osteocyte body
area and facilitating the process of locating these areas. Figure 12 shows an example of the
image after the final filtering step.
Figure 12: Transverse section after filtering steps to facilitate the automaticdetection of its osteocyte locations. Each little area within the section represents thelocation of an osteocyte cell body.
23

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 25/49
The center of mass of each of the osteocyte body areas was then automatically found, after which
the user was allowed to manually double-check and fix any mistakes that the automatic cell-
location detector may have made. Figure 13 shows the found cell locations each marked by a
single red dot within each osteocyte lacuna.
Figure 13: Automatically detected osteocyte locations, each marked by red dots within the individual lacunae.
The x and y coordinates of these locations were saved after subtracting the respective x and y
coordinates of the particular section’s calculated center of mass. Saving the coordinates with
respect to the section’s center of mass ensured that the coordinates from each section were
aligned.
24

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 26/49
Once the x and y coordinates of the osteocyte locations were found for each transverse
section, the last remaining step in creating the 3-D osteocyte network model was to find the z
coordinates. Given that osteocytes are approximately 10 μm thick and the transverse sections are
each 5 μm thick, it was assumed that any osteocyte would appear in at most two contiguous
transverse section images. If such is the case, then the z coordinate of the osteocyte is, in reality,
somewhere in between the two layers. Based on this assumption, the z coordinates of the
osteocyte locations could be approximated (extrapolated) by viewing two contiguous images at a
time, finding the osteocytes that appear in both images (defined as within a distance threshold of
30 μm), and moving these osteocyte locations into a layer that was halfway between the two
current layers. This was repeated for all subsequent groups of contiguous layers. At the end, the
topmost and bottommost layers out of the entire group were removed because they could have
contained cells that were in the previous or the next layer, which were not imaged. Figure 14
illustrates the entire procedure of automatically extrapolating the z coordinate of the cell
locations.
Figure 14: (A) Start with 5 layers; (B) find the osteocytes that appear in two contiguous layers,remove them from those layers, and place them in a layer halfway in between; (C) remove thetopmost and bottommost layers to end with a total of 7 layers.
Once all three coordinates of the osteocyte locations were found, the 3-D osteocyte network
model could be constructed by simply creating a scatterplot of the coordinates. Such a model
25

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 27/49
provided the basis for morphological analysis to be performed and this data is presented in the
proceeding Results section.
Costs
Table 2 summarizes the approximate cost of materials used for this project. Some equipment
such as the light microscope or software such as Photoshop and MATLAB were already present
in the laboratory and free to use, so their costs are not included.
Table 2: Approximate cost of materials.
Solutions used for current project: $100- H&E stain
- Alcohols
- Xylenes
- Other misc. solutions
Solutions previously used: $400
- Texas-Red phalloidin
- Mouse-on-mouse blocking reagent
Lab materials: $300
- Gloves
- Pipettes
- Glass slides
- Expenses for bone sectioning
- Other misc. materials
Total: $800
26

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 28/49
Initial research plan
My capstone is a design-based project and therefore involves very little research elements.
Rather, the MATLAB program that I have designed to construct a model of the 3-D osteocyte
network in bone is a tool that can be used for research purposes in the future. For example, the
model may be used to compare and contrast the osteocyte network of different regions of bone or
bones that underwent different loading regimens, and such possibilities are discussed later in the
“Suggestions for future work” portion of the Results section.
Despite the lack of research focus in my project, I designed my MATLAB program to
meet three general criteria: accuracy, usability, and efficiency. The model must identify the 3-D
locations of osteocytes in a section of bone as accurately as possible, and this required me to
develop a method to not only find the transverse ( x, y) coordinates of the cells in each section,
but also to extrapolate a longitudinal ( z) coordinate for each cell. I decided to write my program
using five transverse section images for a total thickness of 25 μm as this made up a substantial
portion of bone while still comprising a small number of sections for easier testing and
troubleshooting. However, the final program allows for more transverse sections to be added and
included in the model, therefore meeting my second criteria of usability. A graphical user
interface (Figure 15) was also implemented in the program for ease of use, insuring that all users
may be able to construct the 3-D model without being proficient in MATLAB. Finally, the use of
simple H&E-stained images and almost fully-automated processes of the program creates a quick
and efficient technique for visualizing the 3-D locations of osteocytes. The establishment of
these spatial locations and the added capability of performing simple morphological analysis on
the network model also improve the usability and efficiency of the program, and provide the
basis for collecting data for future research studies.
27

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 29/49
Figure 15: Graphical user interface for constructing the 3-D osteocyte network model. The user may load the H&E-stained image and solid mask, find and save its osteocyte location coordinates, and pick the contiguous layers toinclude in the network.
28

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 30/49
Results
Final timeline
29

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 31/49
Data
Figure 16 shows the constructed 3-D osteocyte network model with its seven distinctive layers as
described in the Methods section. Each scatterplot point represents the spatial location of an
ostoecyte in the section of bone.
Figure 16: 3-D osteocyte network model showing seven layers constructed from five transverse section images. Theaxes of the model are in micrometers.
30

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 32/49
From the initial five layers of transverse sections, the mean surface area of a layer was
calculated to be 0.579 ± 0.006 mm2. The number of osteocytes present in each of the seven
constructed layers is listed in Table 3 along with their respective osteocyte densities. There was a
variable range in the number of osteocytes for each layer, with the first (bottom) layer containing
significantly more osteocytes than the other layers. Therefore, the standards of deviation for
number of osteocytes and osteocyte density that were calculated in each layer were relatively
high.
Table 3: Number of osteocytes and osteocyte density in each layer of the model.
Layer # of Osteocytes Density (osteocytes/mm
2
)1 319 550.7
2 119 205.4
3 149 257.2
4 152 262.4
5 262 452.3
6 137 236.5
7 187 322.8
Mean 189.3 ±73.9 326.8 ±127.7
Simple morphological analysis was performed on the constructed 3-D osteocyte network
model to get a better understanding of the network’s architecture. For each osteocyte in each
layer, the distance to its closest neighboring cell was calculated, taking into account only cells
that were in the same layer as the current one. Figure 17 shows a histogram of these closest-
neighbor distances for each of the seven layers in the network. The mean and the median
distances were calculated for each layer, and the mean distance across all layers was calculated
as well.
31

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 33/49
Figure 17: Histograms for the distance of the closest neighboring cell calculated for each
osteocyte in each of the seven layers. The distances were calculated taking into account onlyother cells in the same layer as the current one. Mean 2-D distance for all layers = 31.9 ±
14.9 μm.
32

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 34/49
The closest neighboring cell distances were then calculated again, this time taking into
account cells in the two layers directly above and below the current layer as well. Figure 18
shows histograms of these new distances for each of the seven layers, with the mean and median
distances calculated for each layer as well as the mean distance across all layers.
Figure 18: Histograms for the distance of the closest neighboring cell calculated for eachosteocyte in each of the seven layers. The distances were calculated taking into accountcells in the current layer as well as those in the layers directly above and below the current
one. Mean 3-D distance for all layers = 21.1 ± 8.1 μm.
33

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 35/49
The results from Figures 17 and 18 suggest that an osteocyte, on average, is closer to
cells in a different layer than cells in its current layer because of the discernibly smaller 3-D
distance (21.1 μm) compared to the 2-D distance (31.9 μm). To verify that this was a consistent
trend throughout all regions of the bone section, each transverse section was divided into eight
equal sectors as shown in Figure 19. Morphological analysis was performed for each of the
separate sectors to compare and contrast any differences between them that may possibly be
skewing the preceding data.
Figure 19: Transverse bone section divided into eight equal sectors.
34

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 36/49
The mean number of osteocytes per sector was calculated across all layers and is shown
in a bar graph in Figure 20. On average, some sectors had more osteocytes than others, so it was
essential to compare the architecture of each sector.
Figure 20: Mean number of osteocytes per sector across all layers of the network = 23.9± 5.8 osteocytes.
To examine the architecture of each sector, the distance to the closest neighboring cell
was again calculated for each osteocyte in each layer, first taking into account only cells in the
current layer. Figure 21 shows a bar graph of the mean closest-neighbor distance calculated for
all sectors in each of the seven layers. Although there was some variability in distances among
individual layers, when the mean distance was calculated for each sector across all layers, the red
bar graph in Figure 21 shows that the closest-neighbor distances were fairly consistent in each
sector. The mean distance across all sectors was calculated to be 33.6 μm, a very similar distance
to the 31.9 μm distance calculated in Figure 17.
35

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 37/49
Figure 21: Bar graphs of the mean closest-neighbor distances calculated in each sector for each of the seven layers in the network. The distances were calculated taking into accountonly other cells in the same layer as the current one. The red bar graph shows the mean
distance across all layers. The mean distance across all sectors was 33.6 ± 1.6 μm.
The closest neighboring cell distances per sector were then calculated taking into account
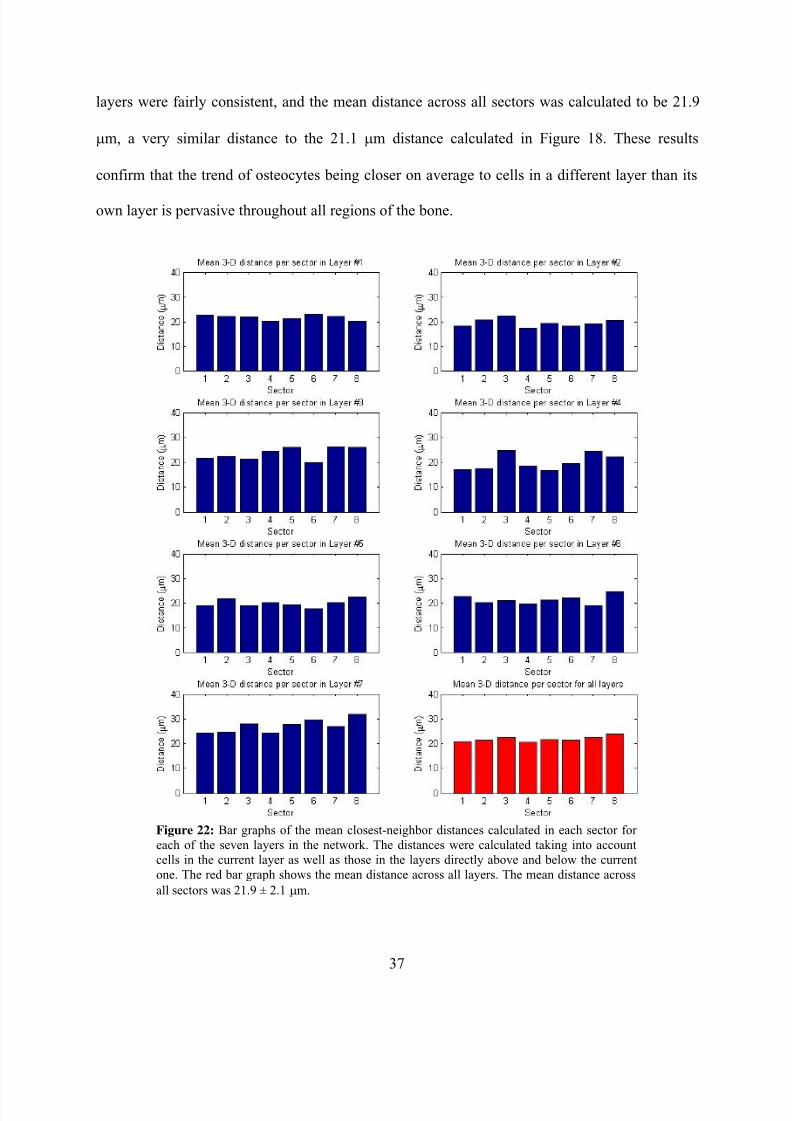
cells in the two layers directly above and below the current layer as well. Figure 22 shows a bar
graph of the new mean closest-neighbor distance calculated for all sectors in each of the seven
layers. The red bar graph once again shows that the mean nearest-neighbor distances across all
36
7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 38/49
layers were fairly consistent, and the mean distance across all sectors was calculated to be 21.9
μm, a very similar distance to the 21.1 μm distance calculated in Figure 18. These results
confirm that the trend of osteocytes being closer on average to cells in a different layer than its
own layer is pervasive throughout all regions of the bone.
Figure 22: Bar graphs of the mean closest-neighbor distances calculated in each sector for each of the seven layers in the network. The distances were calculated taking into accountcells in the current layer as well as those in the layers directly above and below the currentone. The red bar graph shows the mean distance across all layers. The mean distance across
all sectors was 21.9 ± 2.1 μm.
37

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 39/49
Experimental/design decisions
As previously mentioned, an experimental decision was made to forego phalloidin staining in
favor of H&E as nearly two quarters worth of testing the phalloidin and confocal laser scanning
microscopy (CLSM) method proved to be unsuccessful. It was determined that an easier and
faster histology protocol would better serve the purposes of the project, and therefore the H&E
protocol was used. If images were obtained from CLSM, then the computer programming
portion of the project would have been less complex since there would no longer be a need to
account for the misalignment of bone transverse sections. However, it was decided that the faster
and simpler method of constructing a 3-D osteocyte network model using H&E-stained images
was worth the tradeoff of having to design an additional technique/algorithm to align the images
before constructing the model.
The method used to make the 3-D osteocyte network model was also simple in design,
therefore improving the usability of the computer program as it only requires users to have a
minimal amount of computer/programming skills and engineering background knowledge. The
concepts of center of mass and moment of area (second moment of inertia) are relatively
straightforward and easily grasped by most users with any basic mathematics background, so
they may use the computer program with a good understanding of how it operates instead of as a
black box.
Despite the simplicity of the program, it has proven to be fairly accurate. Calculating the
moments of area of the transverse sections (Table 1) showed that they were correctly aligned
with little error, and there was also agreement between the morphological data generated by the
model and those in the literature, as will be discussed in the next section. Thus, the decision to
employ simple experimental/design techniques for this project has provided a usable and
efficient method for constructing a 3-D osteocyte network model without sacrificing accuracy.
38

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 40/49
Analysis and conclusions
Overall, my computer program for constructing a 3-D osteocyte network model using H&E-
stained transverse section images performed well and met my three general criteria of efficiency,
usability, and accuracy. The H&E staining protocol only took approximately half the time to
execute as the phalloidin protocol, and the light microscope imaging system was also much more
straightforward to use than the confocal laser scanning microscope. Although using the light
microscope introduced the problem of misalignment between images, the images were re-aligned
in Photoshop and checked in MATLAB by calculating and comparing the moment of area of
each section. As Table 1 shows, the error in alignment (coefficient of variation) was found to be
less than 2%, and thus it was concluded that my manual re-alignment method was acceptably
accurate.
With no user manipulation, the program took less than one minute in total running time to
go from loading the five individual transverse section images to constructing the full 3-D model.
However, as was expected for a simple automated program, there were several errors in the
found osteocyte locations (usually around 50 errors per image). Therefore, to improve the
accuracy of the model, the user had to manually double-check and fix these mistakes that were
made by the automatic cell-locating program, which took up to 10 minutes per image. Although
this method is significantly less laborious than locating every cell by hand, the amount of time it
takes to fix the mistakes generated by the automated program can still be reduced if the image-
filtering algorithm can be improved in the future. This would in turn improve the efficiency and
accuracy of the program and model.
The 3-D osteocyte network shown in Figure 16 has significantly more osteocytes in its
first (bottom) layer compared to its other layers. This discrepancy is unlikely a systematic error
as all layers were put through the same network-construction program to extrapolate the z
39

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 41/49
coordinates of the osteocytes. Therefore, it is likely that there were simply more osteocytes
towards that particular end of the bone, and this could be verified by cutting, imaging, and
analyzing more contiguous transverse sections from that end. Despite the variability in the
number of osteocytes per layer in the network, the average osteocyte density (326.8 ± 127.7
osteocytes/mm2) calculated in Table 3 is within the same range as the density found by other
investigators (an estimated 400-500 osteocytes/mm2 by Power et al. [31]), albeit a little lower.
More transverse sections need to be imaged and analyzed to obtain a better comparison of
osteocyte density, although this preliminary agreement in values between my model’s
calculations and those found in the literature suggests that the imaging and cell-locating
techniques used in this project were fairly accurate.
For each osteocyte in each layer in the model, the distance to its nearest neighboring cell
calculated taking into account only cells in the same layer as the current one (referred to as the 2-
D distance) averaged to be 31.9 ± 14.9 μm, as shown in Figure 17. The distribution of distances
around this mean was generally normal, although there were some layers that had almost as
many distances near the minimum and maximum as around the mean. However, the mean and
median distances were very similar (the largest difference being 15.7%), indicating that the
values at the extremes were not significantly skewing the mean value.
When the nearest neighboring cell distances were calculated also taking into account cells
in the two layers directly above and below the current one (referred to as the 3-D distance), the
mean distance was calculated to be 21.1 ± 8.1 μm as shown in Figure 18. In addition to having a
smaller standard deviation than that of the 2-D distance, the 3-D distances also show a much
more normal distribution around the mean. The differences between the mean and median 3-D
distance in each layer were also smaller on average than those for the 2-D distance, with the
largest difference being only 6.7%.
40

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 42/49
The results in Figures 17 and 18 suggest that, on average, osteocytes are more likely to be
closer to cells in a different layer than its own layer. Not only was the mean 3-D distance shorter
than the 2-D distance, the 3-D standard deviations and differences between mean and median
were also smaller, and the 3-D distance distributions were more normal in shape than that of the
2-D distances. This indicates that there was less variability in the calculated 3-D distances
compared to the 2-D distances, thus reinforcing the perceived trend of osteocytes being closer to
cells in a different layer than its own layer.
To verify that such a trend was present in all regions, the transverse sections were divided
into eight sectors as shown in Figure 19. The mean number of osteocytes per sector calculated
across all layers was 23.9 ± 5.8 osteocytes, indicating that there was some variability between
layers, especially between Sectors 7 and 8 (a 200% difference). To determine if this relatively
large variability had an effect on the previously calculated nearest-neighboring cell distances, the
2-D and 3-D distances were calculated in each separate sector of each layer. As Figures 21 and
22 show, there was some variability among the sectors in different layers for both the 2-D and 3-
D distances. However, when averaged across all sectors and layers, the distances showed much
less variability, with a mean 2-D distance of 33.6 ± 1.6 μm and a mean 3-D distance of 21.9 ±
2.1 μm. The mean 2-D and 3-D distances averaged across sectors were very similar to the
distances initially calculated from Figures 17 and 18 (a 5.1% difference for 2-D distance and a
3.8% difference for 3-D distance), suggesting that the nearest neighboring cell distance
calculations were not significantly different between the different sectors of the transverse
section. Therefore, these results verify that the tendency for an osteocyte to be closer to cells in a
different layer than its own layer is true throughout all regions of the transverse section.
To check the accuracy of the calculations made by my computer program, the mean 2-D
and 3-D distances were compared to similar data found in the literature. Chakkalakal et al.
41

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 43/49
estimated that the length of canaliculi linking adjacent osteocytes in the same transverse plane
was approximately 30 to 40 μm [32], which is in the same range as my program’s calculated
mean 2-D distance of 31.9 ± 13.9 μm. Sugawara et al. were able to use their CLSM-generated
osteocyte network to measure the point-to-point 3-D distance between the centers of mass of
adjacent osteocytes and reported a mean distance of 24.1 ± 2.8 μm [4]. These values are also in
the same range as my program’s calculated nearest neighboring cell 3-D distance of 21.1 ± 8.1
μm. The approximate agreement in morphological data between those previously measured and
those generated by my model not only verifies the accuracy of my program, but also
demonstrates its efficiency. My technique for constructing a 3-D osteocyte network model is
therefore advantageous in that it uses significantly simpler (H&E staining and light microscopy)
and faster (automatic cell detection and model construction) methods than those previously used
to produce equally accurate data.
An interesting result that was obtained from constructing the 3-D osteocyte network
model is that osteocytes are seemingly closer to cells in layers different than their own layer.
Because osteocytes that are closer together are more likely to be connected via their processes
and transmitting signals to each other [33], this result suggests that the longitudinal component
of cell-to-cell communication may have a larger influence on the direction of signaling pathways
than previously recognized. Whether or not osteocytes’ signaling pathways tend to travel in the
longitudinal direction rather than in the transverse direction remains to be validated in future
investigations. The 3-D bone morphology in different parts along the bone’s long axis needs to
be examined to determine the true complex nature of osteocyte signaling pathway directions.
Such analysis may be performed using sections that are cut transversely as well as longitudinally
to verify the accuracy of the z-dimension data. In addition, more detailed imaging methods such
as those employed by Sugawara et al. [4] may be used to determine the exact connections that
42

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 44/49
each osteocyte has to its neighbors in both the transverse and longitudinal directions. These
possible investigations are beyond the scope of this capstone project; however, this project has
demonstrated that obtaining a solid understanding of the direction of signaling pathways requires
the study of bone’s 3-D osteocyte network architecture and specifically the influence of the
longitudinal component of cell-to-cell signaling.
Suggestions for future work
Although my computer program has proven to be quite efficient compared to previously used
methods for analyzing osteocyte networks, improvements may still be made to increase its
efficiency and accuracy. Specifically, if more time was devoted to understanding how to
properly stain the bone with the fluorescent antibody phalloidin, then the images obtained
through CLSM should not only show the spatial location of osteocytes, but also the processes
that connect them. Visualizing exactly how osteocytes are connected is vital to understanding the
direction of their signaling pathways, and is also much more accurate than only knowing the
osteocytes’ spatial locations and inferring their activity based on their distances to each other. In
addition, using phalloidin staining may possibly eliminate the need to cut serial transverse
sections of the bone (and later re-align them) as CLSM is capable of taking serial transverse
images within thick sections of bone. Despite these advantages of phalloidin staining and CLSM,
there is a tradeoff between using complex and expensive techniques to construct a more detailed
model and using faster and simpler techniques to construct a fairly accurate model. Such a
decision must be carefully considered and will depend on the needs of future experiments.
As my capstone project stands now, the technique designed to automatically construct a
3-D osteocyte network model provides the opportunity to analyze the morphology of any given
section of bone in a quick, easy, yet accurate manner. The computer program may be used to
43

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 45/49
construct models of different parts of a section of bone to examine how the osteocyte network
differs in these various sections. Alternatively, models could be constructed for bones that
underwent different loading regimens to compare and contrast how the osteocyte network has
changed in response to the loading. As long as there is a need to quickly examine the
morphology of any type of bone, the computer program may easily be modified according to the
bone’s specifications and then be used for analysis.
The automatically obtained spatial locations of osteocytes may also be incorporated into
the OSL’s ABM to improve the accuracy of its real-time simulations of osteocyte signaling
activity induced by artificially-introduced stimuli. Should it be decided that the ABM’s
simulations in 3-D need to be verified by in vivo experiments, then my computer program may
need to be modified to include the capability of constructing the osteocyte network model using
phalloidin-stained CLSM images that illustrate the physical interconnectivity of osteocytes. The
development of a program that can perform fast and accurate analysis of bone morphology in
addition to being able to show the detailed architecture of the osteocyte network may prove to be
an invaluable tool for investigators to understand mechanotransduction signaling pathways in
bone.
44

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 46/49
Acknowledgements
I would like to thank my advisor, Professor Ted S. Gross, for the opportunity to work at the OSL
and for his time and effort in guiding me through my capstone project. I would also like to thank
the rest of the OSL team, particularly DeWayne Threet for teaching me various immunohisto-
chemistry and imaging techniques and Brandon Ausk for aiding me with MATLAB. Finally, I
would like to acknowledge the additional help with programming that I received from my
classmates Jason Padvorac and Yung-Chun Chen. This capstone project was funded by the
Dean’s Undergraduate Research Award in the University of Washington College of Engineering
and the NIH AR45565.
45

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 47/49
References
1. Rubin, C.T. and L.E. Lanyon, Regulation of bone formation by applied dynamic loads. JBone Joint Surg Am, 1984. 66(3): p. 397-402.
2. Rubin, C.T. and L.E. Lanyon, Regulation of bone mass by mechanical strain magnitude. Calcif Tissue Int, 1985. 37(4): p. 411-7.
3. Gross, T.S., et al., Noninvasive loading of the murine tibia: an in vivo model for the study
of mechanotransduction. J Bone Miner Res, 2002. 17(3): p. 493-501.
4. Sugawara, Y., et al., Three-dimensional reconstruction of chick calvarial osteocytes and
their cell processes using confocal microscopy. Bone, 2005. 36(5): p. 877-83.
5. Srinivasan, S., et al., Low-magnitude mechanical loading becomes osteogenic when rest
is inserted between each load cycle. J Bone Miner Res, 2002. 17(9): p. 1613-20.
6. Riggs, B.L. and L.J. Melton, 3rd, Involutional osteoporosis. N Engl J Med, 1986.314(26): p. 1676-86.
7. Gross, T.S., et al., Why rest stimulates bone formation: a hypothesis based on complex
adaptive phenomenon. Exerc Sport Sci Rev, 2004. 32(1): p. 9-13.
8. Lewiecki, E.M., Prevention and treatment of postmenopausal osteoporosis. ObstetGynecol Clin North Am, 2008. 35(2): p. 301-15.
9. Knothe Tate, M.L. and U. Knothe, An ex vivo model to study transport processes and fluid flow in loaded bone. J Biomech, 2000. 33(2): p. 247-54.
10. Jones, H.H., et al., Humeral hypertrophy in response to exercise. J Bone Joint Surg Am,1977. 59(2): p. 204-8.
11. Rubin, J., C. Rubin, and C.R. Jacobs, Molecular pathways mediating mechanical
signaling in bone. Gene, 2006. 367: p. 1-16.
12. Haapasalo, H., et al., Effect of long-term unilateral activity on bone mineral density of
female junior tennis players. J Bone Miner Res, 1998. 13(2): p. 310-9.
13. American Association of Clinical Endocrinologists medical guidelines for clinical
practice for the prevention and treatment of postmenopausal osteoporosis: 2001 edition,
with selected updates for 2003. Endocr Pract, 2003. 9(6): p. 544-64.
14. Turner, C.H. and F.M. Pavalko, Mechanotransduction and functional response of the
skeleton to physical stress: the mechanisms and mechanics of bone adaptation. J OrthopSci, 1998. 3(6): p. 346-55.
46

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 48/49
15. Knothe Tate, M.L., "Whither flows the fluid in bone?" An osteocyte's perspective. JBiomech, 2003. 36(10): p. 1409-24.
16. Holtrop, M.E. and G.J. King, The ultrastructure of the osteoclast and its functional
implications. Clinical Orthopaedics and Related Research, 1977. 123: p. 177-196.
17. Reilly, G.C., et al., Investigation of the morphology of the lacunocanalicular system of
cortical bone using atomic force microscopy. Ann Biomed Eng, 2001. 29(12): p. 1074-81.
18. Knothe Tate, M.L., et al., The osteocyte. The International Journal of Biochemistry &Cell Biology, 2004. 36: p. 1-8.
19. Stains, J.P. and R. Civitelli, Gap junctions in skeletal development and function. BiochimBiophys Acta, 2005. 1719(1-2): p. 69-81.
20. Bonewald, L.F., Generation and function of osteocyte dendritic processes. JMusculoskelet Neuronal Interact, 2005. 5(4): p. 321-4.
21. Turner, C.H., Three rules for bone adaptation to mechanical stimuli. Bone, 1998. 23(5): p. 399-407.
22. Huiskes, R., If bone is the answer, then what is the question? J. Anat., 2000. 197: p. 145-56.
23. Wolff, J., The Law of Bone Remodeling. 1986, New York: Springer.
24. Kamioka, H., T. Honjo, and T. Takano-Yamamoto, A three-dimensional distribution of
osteocyte processes revealed by the combination of confocal laser scanning microscopy
and differential interference contrast microscopy. Bone, 2001. 28(2): p. 145-9.
25. Krempien, B., et al., The influence of immobilization on osteocyte morphology: osteocyte
differential count and electron microscopical studies. Virchows Arch A Pathol AnatHistol, 1976. 370(1): p. 55-68.
26. Palumbo, C., et al., Osteocyte differentiation in the tibia of newborn rabbit: an
ultrastructural study of the formation of cytoplasmic processes. Acta Anat (Basel), 1990.137(4): p. 350-8.
27. Cooper, J.A., Effects of cytochalasin and phalloidin on actin. J Cell Biol, 1987. 105(4): p.1473-8.
28. Ausk, B.J., T.S. Gross, and S. Srinivasan, An agent based model for real-time signaling
induced in osteocytic networks by mechanical stimuli. J Biomech, 2006. 39(14): p. 2638-46.
47

7/28/2019 BIOEN 482 Senior Capstone Thesis
http://slidepdf.com/reader/full/bioen-482-senior-capstone-thesis 49/49
29. Kamioka, H., et al., Terminal differentiation of osteoblasts to osteocytes is accompanied
by dramatic changes in the distribution of actin-binding proteins. J Bone Miner Res,2004. 19(3): p. 471-8.
30. Brown, H.S. Hematoxylin & Eosin (The Routine Stain). 2002 [cited 2008 May 16];
Available from: http://www.sigmaaldrich.com/img/assets/7361/Primer-H&Emay04.pdf.
31. Power, J., et al., Osteocyte density in aging subjects is enhanced in bone adjacent to
remodeling haversian systems. Bone, 2002. 30(6): p. 859-65.
32. Chakkalakal, D.A., Mechanoelectric transduction in bone. Journal of Materials Research,1989. 4: p. 1034-46.
33. Thi, M.M., et al., Fluid shear stress remodels expression and function of junctional proteins in cultured bone cells. Am J Physiol Cell Physiol, 2003. 284(2): p. C389-403.


















