
Biochem1 2014 Recitation Chapter 11 – Lipids/Membrane ... · Chapter 11 – Lipids/Membrane...
15
Biochem1 2014 Recitation Chapter 11 – Lipids/Membrane Structure
Transcript of Biochem1 2014 Recitation Chapter 11 – Lipids/Membrane ... · Chapter 11 – Lipids/Membrane...

Biochem1 2014 Recitation
Chapter 11 – Lipids/Membrane
Structure

What are membranes? What is the function of the plasma membrane?
• Complex lipid-based structures that form pliable
sheets
• Composed of a variety of lipids and proteins
• Some membrane lipids and proteins are
glycosylated; glycolipids, glycoproteins &
proteoglycans
• cell membrane, which separates the cell from its
surrounding
• Eukaryotes have various internal membranes that
divide the internal space into compartments

Common Features of Membranes • Sheet-like flexible structure, 30–100 Å (3–10 nm) thick
• Main structure is composed of two leaflets of lipids (bilayer)
– With the exception of archaebacteria: monolayer of bifunctional lipids
• Form spontaneously in aqueous solution and are stabilized by noncovalent forces, especially hydrophobic effect
• Protein molecules span the lipid bilayer
• Asymmetric
– Some lipids are found preferably “inside”
– Some lipids are found preferably “outside”
– Carbohydrate moieties are always outside the cell
– Electrically polarized (inside negative ~ –60mV)
• Fluid structures: two-dimensional solution of oriented lipids

The Fluid Mosaic Model: Details
Fluid mosaic model for plasma membrane structure. The fatty acyl chains
in the interior of the membrane form a fluid, hydrophobic region. Integral proteins float in this sea of lipid, held by hydrophobic interactions with their nonpolar amino acid side chains. Both proteins and lipids are free to move laterally in the plane of the bilayer, but movement of either from one leaflet of the bilayer to the other is restricted. The carbohydrate moieties attached to some proteins and lipids of the plasma membrane are exposed on the extracellular surface.

Membrane composition is highly variable in different organisms
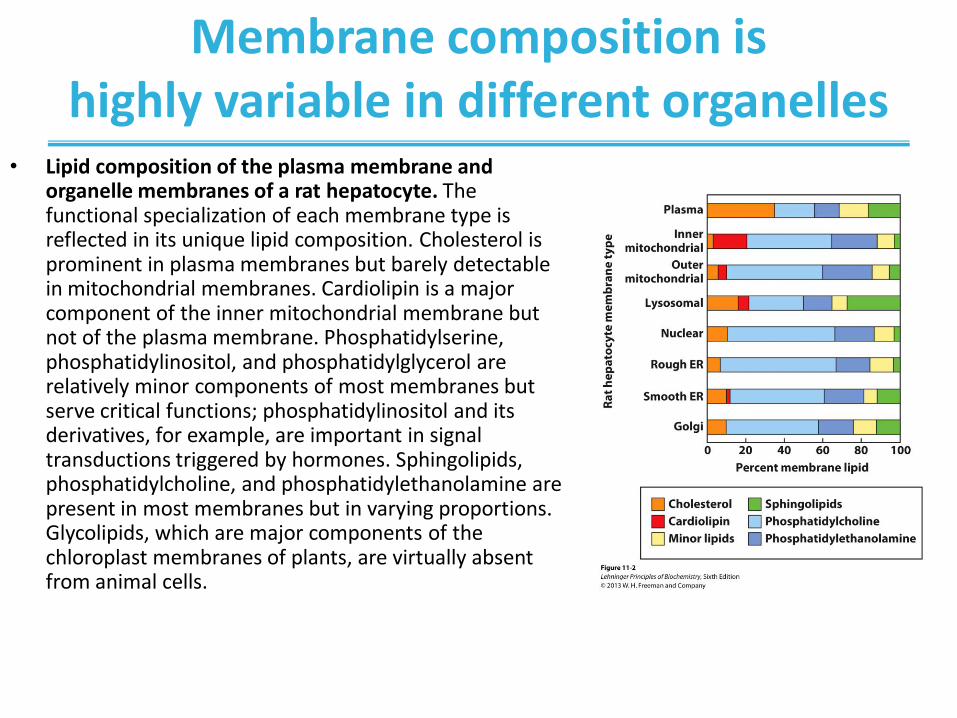
Membrane composition is highly variable in different organelles
• Lipid composition of the plasma membrane and organelle membranes of a rat hepatocyte. The functional specialization of each membrane type is reflected in its unique lipid composition. Cholesterol is prominent in plasma membranes but barely detectable in mitochondrial membranes. Cardiolipin is a major component of the inner mitochondrial membrane but not of the plasma membrane. Phosphatidylserine, phosphatidylinositol, and phosphatidylglycerol are relatively minor components of most membranes but serve critical functions; phosphatidylinositol and its derivatives, for example, are important in signal transductions triggered by hormones. Sphingolipids, phosphatidylcholine, and phosphatidylethanolamine are present in most membranes but in varying proportions. Glycolipids, which are major components of the chloroplast membranes of plants, are virtually absent from animal cells.

Membrane bilayers are asymmetric Asymmetric distribution of
phospholipids between the inner and outer monolayers of the erythrocyte plasma membrane. The distribution of a specific phospholipid is determined by treating the intact cell with phospholipase C, which cannot reach lipids in the inner monolayer (leaflet) but removes the head groups of lipids in the outer monolayer. The proportion of each head group released provides an estimate of the fraction of each lipid in the outer monolayer.

Three Types of Membrane Proteins • Peripheral, integral, and amphitropic proteins.
Membrane proteins can be operationally distinguished by the conditions required to release them from the membrane. Most peripheral proteins are released by changes in pH or ionic strength, removal of Ca2+ by a chelating agent, or addition of urea or carbonate. Integral proteins are extractable with detergents, which disrupt the hydrophobic interactions with the lipid bilayer and form micelle-like clusters around individual protein molecules. Integral proteins covalently attached to a membrane lipid, such as a glycosyl phosphatidylinositol (GPI; see Fig. 11–15), can be released by treatment with phospholipase C. Amphitropic proteins are sometimes associated with membranes and sometimes not, depending on some type of regulatory process such as reversible palmitoylation.

Six Types of Integral Membrane Proteins
For known proteins, the spatial relationships of protein domains
to the lipid bilayer fall into six categories. Types I and II have a
single transmembrane helix; the amino-terminal domain is
outside the cell in type I proteins and inside in type II. Type III
proteins have multiple transmembrane helices in a single
polypeptide. In type IV proteins, transmembrane domains of
several different polypeptides assemble to form a channel through
the membrane. Type V proteins are held to the bilayer primarily
by covalently linked lipids, and type VI proteins have both
transmembrane helices and lipid
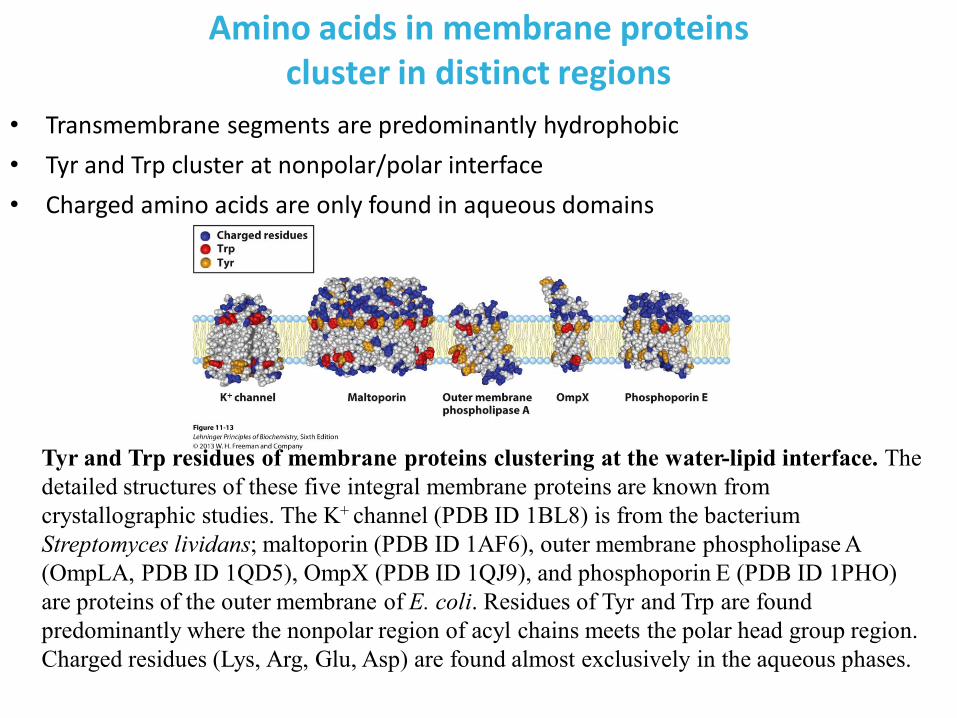
Amino acids in membrane proteins cluster in distinct regions
• Transmembrane segments are predominantly hydrophobic
• Tyr and Trp cluster at nonpolar/polar interface
• Charged amino acids are only found in aqueous domains
Tyr and Trp residues of membrane proteins clustering at the water-lipid interface. The
detailed structures of these five integral membrane proteins are known from
crystallographic studies. The K+ channel (PDB ID 1BL8) is from the bacterium
Streptomyces lividans; maltoporin (PDB ID 1AF6), outer membrane phospholipase A
(OmpLA, PDB ID 1QD5), OmpX (PDB ID 1QJ9), and phosphoporin E (PDB ID 1PHO)
are proteins of the outer membrane of E. coli. Residues of Tyr and Trp are found
predominantly where the nonpolar region of acyl chains meets the polar head group region.
Charged residues (Lys, Arg, Glu, Asp) are found almost exclusively in the aqueous phases.

Membrane proteins also contain -sheets
Membrane proteins with -barrel structure. Three proteins of the E.
coli outer membrane are shown, viewed in the plane of the membrane.
FepA (PDB ID 1FEP), involved in iron uptake, has 22 membrane
spanning β strands. OmpLA (derived from PDB ID 1QD5), a
phospholipase, is a 12-stranded β barrel that exists as a dimer in the
membrane. Maltoporin (derived from PDB ID 1MAL), a maltose
transporter, is a trimer; each monomer consists of 16 β strands.

Membrane Rafts Lipid distribution in a single leaflet is not random or uniform • Lipid rafts
– contain clusters of glycosphingolipids with longer-than-usual tails – are more ordered – contain specific doubly or triply acylated proteins – allow segregation of proteins in the membrane
• Membrane microdomains (rafts). Stable associations of sphingolipids and cholesterol in the outer leaflet produce a microdomain, slightly thicker than other membrane regions, that is enriched with specific types of membrane proteins. GPI-anchored proteins are prominent in the outer leaflet of these rafts, and proteins with one or several covalently attached long-chain acyl groups are common in the inner leaflet. Inwardly curved rafts called caveolae are especially enriched in the protein caveolin. Proteins with attached prenyl groups (such as Ras) tend to be excluded from rafts.

Caveolin forces membrane curvature
• Caveolin forces inward curvature of a membrane. Caveolae are small invaginations in the plasma membrane, as seen in (a) an electron micrograph of an adipocyte that is surface-labeled with an electron-dense marker. (b) Cartoon showing the location and role of caveolin in causing inward membrane curvature. Each caveolin monomer has a central hydrophobic domain and three long-chain acyl groups (red), which hold the molecule to the inside of the plasma membrane. When several caveolin dimers are concentrated in a small region (a raft), they force a curvature in the lipid bilayer, forming a caveola. Cholesterol molecules in the bilayer are shown in orange.
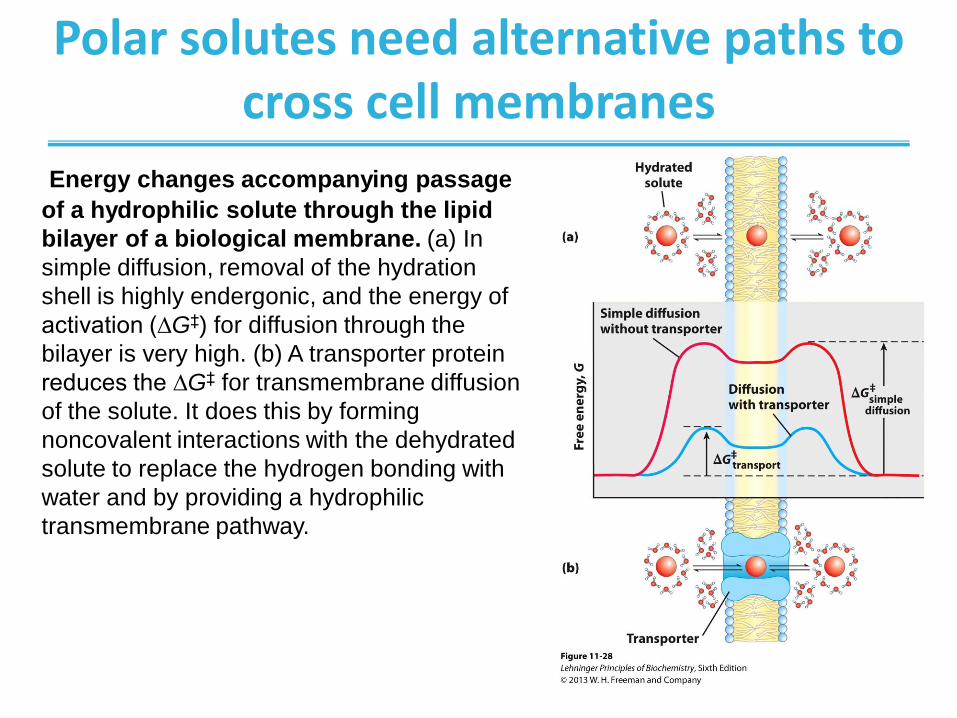
Polar solutes need alternative paths to cross cell membranes
Energy changes accompanying passage
of a hydrophilic solute through the lipid
bilayer of a biological membrane. (a) In
simple diffusion, removal of the hydration
shell is highly endergonic, and the energy of
activation (∆G‡) for diffusion through the
bilayer is very high. (b) A transporter protein
reduces the ∆G‡ for transmembrane diffusion
of the solute. It does this by forming
noncovalent interactions with the dehydrated
solute to replace the hydrogen bonding with
water and by providing a hydrophilic
transmembrane pathway.
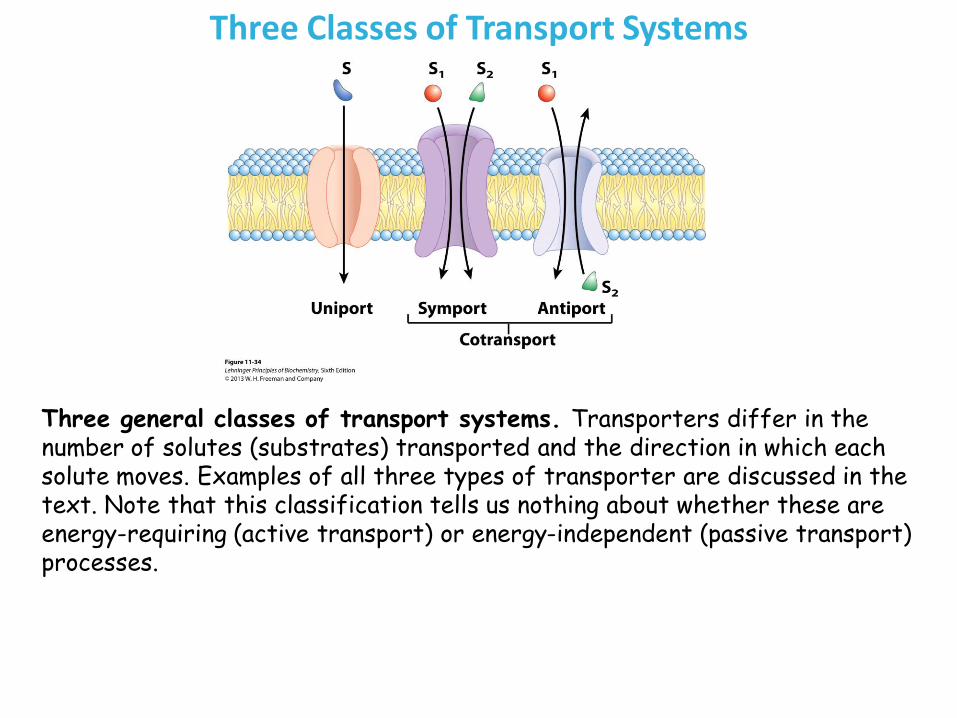
Three Classes of Transport Systems
Three general classes of transport systems. Transporters differ in the number of solutes (substrates) transported and the direction in which each solute moves. Examples of all three types of transporter are discussed in the text. Note that this classification tells us nothing about whether these are energy-requiring (active transport) or energy-independent (passive transport) processes.


















