
BIOACTIVE PRINCIPLES FROM PANAMANIAN MEDICINAL PLANTS
250
BIOACTIVE PRINCIPLES FROM PANAMANIAN MEDICINAL PLANTS Thesis presented by Pablo Narclso Solis Gonzalez for the degree of Doctor of Philosophy In the Faculty of Medicine of the University of London Department of Pharmacognosy The School of Pharmacy University of London 1994
Transcript of BIOACTIVE PRINCIPLES FROM PANAMANIAN MEDICINAL PLANTS

BIOACTIVE PRINCIPLES FROM PANAMANIAN
MEDICINAL PLANTS
Thesis presented by
Pablo Narclso Solis Gonzalez
for the degree of
Doctor of Philosophy
In the Faculty of Medicine
of the University of London
Department of Pharmacognosy
The School of Pharmacy
University of London
1994

ProQuest Number: 10105140
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10105140
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States Code.
Microform Edition © ProQuest LLC.
ProQuest LLC 789 East Eisenhower Parkway
P.O. Box 1346 Ann Arbor, Ml 48106-1346

A tribute to my Grand Parents,
Pablo and Conce
and to my Parents,
Nato and Gladys
my models of fortitude.

ABSTRACT
Plants are widely used in Panama and Central America to cure different
diseases, including malaria and amoebic dysentery. Cephaëlis ipecacuanha of
the famiiy Rubiaceae contains emetine, which is used in the treatment of
severe amoebic dysentery. Species of Rubiaceae, Simaroubaceae, Meliaceae
and Menispermaceae have been examined, they are known to contain
isoquinoline alkaloids, quassinoids, limonoids and bisbenzylisoquinoline (bbiq)
alkaloids, respectively, with activity against Plasmodium falciparum.
In order to obtain antiprotozoal natural products such as quassinoids,
limonoids, bbiq’s and emetine reiated alkaloids it was decided to investigate the
following plants: Guarea macropetaia, G. rhopalocarpa, Ruagea glabra
(Meliaceae); Abuta dwyerana (Menispermaceae); Cephaëlis camponutans, 0.
dichroa, 0. dimorphandrloldes, 0. glomerulata, Laslanthus panamensis
(Rubiaceae); PIcramnIa antldesma subsp. fessonia, P. teapensis
(Simaroubaceae).
Extracts from different parts of the plants were fractionated and tested
against brine shrimp and KB ceils. Four plants were chemically studied and the
isoiated compounds were characterized by mass spectra, infrared, ultraviolet,
and NMR, also, two dimensional NMR experiments such as COSY-45,
HMQC, HMBC and ROESY. Circular dichroism was used to establish the
absolute configuration of some of the compounds.
C. camponutans yielded benz[g]isoquinoline-5,10-dione, isolated for the
first time from the plant kingdom, and a the new 1-OH-
benzoisochromanquinone. Both compounds showed activity against brine
shrimp, KB ceils and Plasmodium falciparum, in vitro. C. dichroa yielded six
indole monoterpenoid aikaloids, including strictosidine, strictosamide, angustine,
vallesiachotamine, iso-vallesiachotamine and the novel vallesiachotamine
lactone. In contrast, 0. glomerulata yielded three new quinoline alkaloids.

named, glomerulatine A, B and C. The taxonomic position of Cephaëlis is
unclear and more detailed information on the constituent alkaloids may prove
valuable in unravelling this problem.
The butanolic fraction from de leaves of P. antldesma subsp. fessonia,
showed strong activity against KB cells and yielded two known anthraquinones
(aloe-emodin, aloe-emodinanthrone) and three new C-glycoside of aloe-emodin,
named picramnioside A, B and C. The C-glycosides showed activity against
KB cells.
A cytotoxic microwell test using Anemia salina (brine shrimp) was
developed in order to overcome some of the disadvantages of the previously
described method. The test was predictive of cytotoxicity and compounds with
activity on the nervous system were inactive. The activity of the quassinoids
against KB cell and P. falciparum parallel the activity against brine shrimp.

ACKNOWLEDGEMENTS
I would like to thank my supervisor Professor J. David Phillipson for his
precious help and friendship without which this work would not have been
possible. I am also indebted to Professor Mahabir P. Gupta, Departamento de
Quimica Medicinal y Farmacognosia, Facultad de Farmacia, Universidad de
Panama for his encouragement and guidance.
I would like to express my appreciation and thanks to Professors Don
Antonio Gonzalez and Don Angel Gutierrez Ravelo and colleagues in the
Institute de Productos Maturates Organicos, Universidad de La Laguna,
Tenerife, Spain, where part of this work was carried out.
Also, I am indebted to Professor Mireya Correa and Mrs Carmen Galdames
for their valuable help in collection, identification and plant preparation. I am
grateful to Mr. Rutilio Paredes, Research Coordinator of Program of Ecology
and Management of Wild Areas of Kuna Yala (PEMASKY), who very kindly
allowed access for plant collection. I thank Mrs. J. Hawkes for running the
NMR experiments in the AMX400 and WM250 NMR, University of London
Intercollegiate Research Service at King’s College and Dr. K. Welham and his
colleagues for running the mass spectra at the Mass Spectrometry Unit at The
School of Pharmacy, University of London.
I am grateful to my colleges in the Department of Pharmacognosy, The
School of Pharmacy with whom I have collaborated in various aspects of this
work, particularly Miss Maria Camacho, Miss Caroline Lang’at and Dr. Colin
Wright. I also wish to thank Drs. D.C. Warhurst and G.C. Kirby, Department
of Medical Parasitology, The London School of Hygiene and Tropical Medicine
for running the antimalarial activity of some of the isolated compounds.
I am grateful to Dr. R.T. Brown, University of Manchester for generous gifts
of strictosidine, vincosidine and their acetylated derivatives and for their

NMR spectra. Also, I would like to thank Mrs. M. Pickett for technical help and
Mrs A. Cavanagh for excellent graphical work in the preparation on slides and
posters for use at conferences.
Undoubtedly, I am eternally indebted to my wife and lover, Nilka, and my
children, Nilkita, Pablito and Paola, for their daily encouragement,
comprehension and patience during these three difficult years of my life.
I am grateful to the Commission of the European Community for the
provision of a research scholarship (B/CI1*-913064).

CONTENTS
Page
Abstract 3
Acknowledgement 5
List of Abbreviations 12
List of Figures 15
List of Tables 16
SECTION 1 INTRODUCTION 18
1.1 Health care in Panama 19
1.1.1 The country 19
1.1.2 Treatment of diseases 20
1.1.2.1 Health care coverage in Panama 20
1.1.2.2 Major health problems in Panama 21
1.1.3 Ethnomedicine in Panama 25
1.1.4 Natural resources and conservation 28
1.2 Selection of plants for chemical and biological
investigation 36
1.2.1 Traditional medicine 36
1.2.2 Random selection of plants for chemical investigation 38
1.2.3 Chemotaxonomy 39
1.2.3.1 Chemotaxonomy background 41
1.3 Biological testing of plant extracts 42
1.3.1 Advantages and disadvantages of in-vivo and
in-vitro testing 43
1.3.2 Simple bioassays 45
1.3.2.1 Brine shrimp assay 45
1.3.2.2 KB cells assay 46
1.3.2.3 P. falciparum assay 47
1.4 Aims of this study 49

SECTION 2. PLANTS SELECTED FOR INVESTIGATION 50
2.1 Meliaceous plants 51
2.2 Menispermaceous plants 57
2.3 Rubiaceous plants 58
2.4 Simaroubaceous plants 67
SECTION 3 TESTING FOR BIOLOGICAL A CTIVITY 72
3.1 Materials and methods 73
3.1.1 New microwell brine shrimp assay 73
3.1.2 Brine shrimp assay 74
3.1.3 KB cells assay 74
3.1.4 P. falciparum àssay 76
3.2 Results and discussion 76
3.2.1 Evaluation of brine shrimp assay 76
3.2.2 Activity of the plant crude extracts against
brine shrimp and KB cells 80
SECTION 4 PHYTOCHEMISTRY 89
4.1 Material and general methods 90
4.1.1 Plant material 90
4.1.2 Solvents 90
4.1.3 Chromatography techniques 92
4.1.4 Spectrometric methods 92
4.1.4.1 Ultraviolet spectroscopy (UV) 92
4.1.4.2 Infrared spectroscopy (IR) 92
4.1.4.3 Mass spectrometry (MS) 93
4.1.4.4 Nuclear magnetic resonance (NMR) 93
4.1.4.5 Polarimetry 93
4.1.4.6 Circular dichroism (CD) 94
4.1.5 Preparation of acetyl derivatives 94
4.2.1 Preparation of crude plant extracts for
biological test 94
8

4.2.1.1 Results 94
4.3 Isolation and identification of bioactive compounds
from Cephaëlis camponutans {Psychotria camponutans) 98
4.3.1 Extraction procedure 98
4.3.2 Identification of the isolated compounds 98
4.3.3.1 1 -Hydroxybenzoisochromanquinone 98
4.3.3.1.1 Spectral data 98
4.3.3.1.2 1 - Acety Ibenzoisochromanquinone 99
4.3.3.1.2.1 Spectral data 99
4.3.3.2 Benz[g]isoquinoline-5J0 dione 99
4.3.3.2.1 Spectral data 99
4.3.4 Discussion 1 0 2
4.3.5 Biological activity of the isolates 108
4.3.5.1 Results and discussion 108
4.4 Isolation of alkaloids from Cephaëlis dichroa 1 1 0
4.4.1 Extraction and separation procedure 1 1 0
4.4.2 Identification of isolated compounds 1 1 0
4.4.2.1 Vallesiachotamine lactone 1 1 0
4.4.2.1.1 Spectral data 1 1 0
4.4.2.2 Vallesiachotamine 1 1 1
4.4.2.2.1 Spectral data 1 1 1
4.4.2.3 Strictosidine 1 1 2
4.4.2.3.1 Spectral data 1 1 2
4.4.2.4 Strictosidine lactam 113
4.4.2.4.1 Spectral data 113
4.4.2.4.2 Acetyl strictosidine lactam 114
4.4.2.4.2.1 Spectral data 114
4.4.2.5 Angustine 115
4.4.2.5.1 Spectral data 115
4.4.3 Discussion 117

4.5 Isolation and identification of alkaloids from
Cephaëlis glomerulata (Psychotria glomerulata) 121
4.5.1 Extraction and separation procedure 121
4.5.2 Identification of the isolated compounds 121
4.5.2.1 Glomerulatine A 121
4.5.2.1 . 1 Spectral data 121
4.5.2.2 Glomerulatine B 122
4.5.2.2.1 Spectral data 122
4.5.2.5 Glomerulatine C 123
4.5.2.3.1 Spectral data 123
4.5.3 Discussion 124
4.6 Isolation and identification of bioactive compounds
from Picramnia antldesma var. fessonia 131
4.6.1 Extraction and separation procedure 131
4.6.2 Identification of the isolated compounds 132
4.6.2.1 Picramnioside A 132
4.6.2.1 . 1 Spectral data 132
4.6.2.2 Picramnioside B 133
4.6.2.2.1 Spectral data 133
4.6.2.3 Picramnioside C 135
4.6.2.3.1 Spectral data 135
4.6.2.4 Aloe-emodin 136
4.6.2.4.1 Spectral data 136
4.6.2.5 Aloe-emodinanthrone 136
4.6.2.5.1 Spectral data 136
4.6.3 Discussion 137
4.6.4 Biological activity of the isolated compounds 144
SECTION 5 GENERAL DISCUSSION 145
5.1 Introduction 146
10

5.2 Plants selected for this study 147
5.3 Testing for biological activity 148
5.3.1 Brine shrimp assay 148
5.3.2 Biological activity of plant extracts 149
5.4 Phytochemistry 149
5.4.1 Cephaëlis species 149
5.4.2 Picramnia antidesma subsp. fessonia 156
5.5 Are plants predictable in their chemotaxonomy? 157
SECTION 6 CONCLUSIONS 159
SECTION 7 REFERENCES 162
APPENDIX I SPECTRA 185
APPENDIX n LIST OF SCIENTIFIC PAPERS 247
II

List of Abbreviations
pCi
pM
Dept
'H
2D
BBIQ
BuOH
br
QD,
CC
CD
CD3 OD
CDCI3
CIMS
cm'
COSY-45
d
DCCC
dd
ddd
DMSO-Dg
EDgo
EIMS
EtOAc
EtOH
eV
Microcurie
Micromolar
Distortionless enhancement by polarization transfer (differentiation
between CH, CHj, and CH 3 using the improved sensitivity of
polarisation transfer)
Carbon thirteen
Proton
Two dimensional
Bisbenzylisoquinoline alkaloid
n-Butanol
Broad
Centigrade
Deuterobenzene
Column chromatography
Circular dichroism
Deuteromethanol
Deuterochloroform
Chemical ionization mass spectrometry
Centimetre to the minus one
Two dimensional correlation spectroscopy
Doublet
Droplet counter current chromatography
Double doublet
Double double doublet
Deuterodimethylsulphoxide
Effective dose fifty
Electron impact mass spectrometry
Ethylacetate
Ethanol
Electron volt = 94.487 kJ/mol = 23.06 kcal/mol
12

FABMS Fast atom bombardment mass spectrometry
FDMS Field desorption mass spectrometry
FAO Food and Agricultural Organization
g Gram
GC Gas chromatography
HETCOR Heterocorrelation
HMBC Heteronuclear multiple bond connectivity
HMQC Heteronuclear multiple quantum coherence
HPLC High performance liquid chromatography
HRMS High resolution mass spectrometry
hrs Hours
Hz Hertz
IR Infrared
J Coupling constant
Kg Kilogram
Km Square kilometre
1 Litre
LC 5 0 Liquid concentration fifty
m Multiplet
M Molar
N Normal
NCI National Cancer Institute
max Maxima
mg Milligram
ml M illilitre
miz The mass of the ion in dalton divided by its charge
MegCO Acetone
MeOH Methanol
MHz Mega Hertz
MNOBA Mononitro-ortho-benzoic acid
MS Mass spectrometry/spectrum
nm Nanometre
13

nM Nanomolar
NMR Nuclear magnetic resonance
nOe Nuclear Overhauser effect
NOESY Two dimensional nuclear Overhauser effect spectroscopy
ppm Part per million
pTLC Preparative thin-layer chromatography
ROESY Rotating frame nuclear Overhauser effect spectroscopy
s Singlet
sd Standard deviation
sh Shoulder
t Triplet
TEA Trifluoroacetic acid
TLC Thin-layer chromatography
TMS Tetramethylsilane
UV Ultraviolet
WHO World Health Organization
14

LIST OF FIGURES
Page
Figure 1.1 Distribution of the Indigenous population in Panama 27
Figure 1.2 Different vegetation zones in Panama 32
Figure 1.3 Natural parks and protected areas in Panama, 1985. 35
Figure 4.1 Extraction and fractionation procedure 95
Figure 4.2 Compounds isolated from Cephaëlis camponutans and the
acetyl derivative of 1-hydroxibenzoisochromanquinone 103
Figure 4.3 Putative mass spectral fragmentation of
1 -hydroxybenzoisochromanquinone 106
Figure 4.4 Putative mass spectral fragmentation of
benz[g]isoquinoline-5,10-dione 107
Figure 4.5 Alkaloids isolated from C. dichroa showing their
possible biosynthetic relationship 116
Figure 4.6 Putative mass spectral fragmentation of
vallesiachotamine lactone 118
Figure 4.7 Possible structures of the alkaloids isolated from
Cephaëlis glomerulata 128
Figure 4.8 Anthraquinones isolated from
P. antidesma subsp. fessonia 138
Figure 4.9 C-Glycoside isolated from P. antidesma subsp. fessonia
showing the most relevant ROESY effects 139
Figure 4.10 Picramniosides B and C isolated from P. antidesma subsp.
fessonia showing the most relevant ROESY effects 140
Figure 4.11 Putative mass spectra fragmentation of
picramniosides A and C 142
Figure 5.1 Biosynthetic pathway of indole and isoquinoline
monoterpenoid alkaloids 151
Figure 5.2 Putative biosynthetic pathways of non-iridoid alkaloids
of the quinoline- and polyindolenine-type typical
of the tribe Psychotriae 153
Figure 5.3 Biosynthetic relationship of the alkaloids produced in
sub-genera Heteropsychotria and Psychotria 155
15

LIST OF TABLES
Table 1.1 Health care coverage in Panama, in 1991
Table 1.2 Health indicators in urban and rural areas
in Panama, 1990
Table 1.3 Number of cases of some life threatening diseases
in Panama
Table 1.4 Major life forms of the vegetation in Panama
Table 1.5 General information on the Panamanian flora
Table 1.6 Geographic distribution of plant species in Panama
Table 1.7 Plant families in Panama with more than 80 species
Table 1.8 Advantages and disadvantages of in vivo and in viti'o
bioassays
Table 1.9 Panamanian medicinal plants selected for chemical and
biological studies
Table 2.1 Chemical compounds isolated from the genus Guarea
Table 2.2 Chemical compounds isolated from Abuta species
Table 2.3 Chemical compounds from Psychotria species
Table 2.4 Compounds isolated from species of Picramnia
Table 3.1 Comparison of the values obtained in the brine
shrimp micro-technique with previously reported data
Table 3.2 Comparison of brine shrimp toxicity against in vitro
KB cell cytotoxicity of compounds with different
mechanisms of action
Table 3.3 Activities of quassinoids against brine shrimps,
KB cell and P falciparum in vitro
Table 3.4 Activity against brine shrimp of the extracts from
Guarea macropetaia
Table 3.5 Activity against brine shrimp of the extracts from
Guarea rhopalocarpa
Table 3.6 Activity against brine shrimp of the extracts from
Ruagea glabra
Table 3.7 Activity against brine shrimp of the extracts
from Abuta dwyerana
Page
22
23
24
29
30
31
33
44
49
52
59
65
68
78
79
81
82
82
83
83
16

Table 3.8 Activity against brine shrimp of the extracts from
Cephaëlis camponutans 84
Table 3.9 Activity against brine shrimp of the extracts from
Cephaëlis dichroa 84
Table 3.10 Activity against brine shrimp of the extracts from
Cephaëlis dimorphatidrioides 85
Table 3.11 Activity against brine shrimp of the extracts from
Cephaëlis glomerulata 85
Table 3.12 Activity against brine shrimp of the extracts from
Laslanthus panamensis 86
Table 3.13 Activity against brine shrimp of the extracts from
Picramnia antidesma subsp. fessonia 86
Table 3.14 Activity against brine shrimp of the extracts from
Picramnia teapensis 87
Table 3.15 Activity against KB cells of extracts selected on
the basis of previously activity against brine shrimp 88
Table 4.1 Plants collected in Panama 91
Table 4.2 Crude extracts for biological test 96-97
Table 4.3 NMR Spectral data of compounds isolated from
C. camponutans 100
Table 4.4 NMR spectral data of the compounds isolated from
C. camponutans 101
Table 4.5 Activity of the compounds isolated from P. camponutans
against brine shrimps, KB cells and P. falciparum 108
Table 4.6 Comparison of the Spectral Data of
Calycanthine-Type Alkaloids 126
Table 4.7 ^ C NMR spectral data of the compounds isolated from
P. antidesma subsp. fessonia 134
Table 4.8 Activity of the compounds isolated from
P. antidesma subsp. fessonia against KB cells 144
Table 5.1 Compounds expected and those found in the plants
selected for this study 158
17

I. INTRODUCTION.

1.1 HEALTH CARE IN PANAMA.
1.1.1 The Country
The Republic of Panama is in the northern hemisphere between the
coordinates 7°12’07” of latitude north, and the coordinates 77°09’24” and
83®03’07” of longitude west, in the intertropical zone next to the equator.
Panama limits with the Caribbean sea in the north, eastward with the Republic
of Colombia, in the south with the Pacific ocean and the Republic of Costa Rica
in the west. It is isthmus that joins South America to the rest of the American
continent.
The total land surfaces of the Republic is 75,517 Km , including 1023
islands in the Caribbean sea and 495 in the Pacific side. The climate is
characterized by moderately high and constant temperatures around the year
(22-31 °C) with abundant rainfall during the year, averaging 1.7 m/year.
Panama is recognized for its extraordinarily rich diversity of plants and
animals (see section 1.1.4) and serves as a biological bridge in the migration
of species between north and south America. Moreover, Panama has been an
important point in the evolution of the species, for example, there are more
species of birds, mammals, reptiles and plants species in Panama than in
Canada, United States and north Mexico together (Anonymous, 1993a).
There is a population of 2.3 millions inhabitants, with a density of 30.8
inhabitants/Km^ (Panama city capital 76.0 inhabitants/Km^, rest of the country
18.0 inhabitants/Km^), about 46.3 % of the Panamanian population lives in rural
areas. The Panamanian population is a mixture of Indians, Africans and
Spanish. In 1991, it was estimated that the 10.7 % of the population were
illiterate.
19

The gross national product per capita was US $ 2,240 in 1991, while the
inflation reached only 2.8% in 1991 with respect to 1987. However, according
to the Ministerio de Planificacion y Politica Economica, the level of poverty of
the population is estimated at 33.6 %. The percentage of unemployed in 1991
was 16.1 (19.5 % in Panama city capital, 11.4 rest of the country) (Anonymous,
1992).
1.1.2 Treatment of Diseases.
1.1.2.1 Health Care Coverage In Panama.
The health care coverage in Panama is a government priority with US
$370.7 millions assigned in 1993, for health care, corresponding to 7.4 % of the
gross national product. More than US $ 64.97 millions were spent on drugs,
representing a drug consumption per capita of approximately US $ 26
(Anonymous, 1993b)
In the last 20 years the health care coverage has considerably increased,
the country has 56 hospitals, 181 health centres and clinics and 433 health
subcentres. In 1973, 68.9 % of births were assisted by physicians, whereas,
in 1991 this number increased to 88.6 % (urban 99.6 %, rural area 78.6 %).
Higher priority is given to the vaccination programs and, in 1991, 70 % of the
children were vaccinated against polio myelitis, DPT (diphtheria, pertussis and
tetanus) and BCG (Bacilli Calmette Guerini) (Anonymous, 1992).
The health sector is composed of public and private sectors; within the
public sector the Ministry of Health is financed by the government and the
social security is financed by employee and employer. Social security covered
48.7 % of the population, in the year 1991, while, the private sector covered
only 15 % (Anonymous, 1992). Table 1.1 (p.22) shows the health care
coverage in Panama, in 1991.
20

While, in Panama city, between 1979 and 1986, the total population
covered by Ministry of Health and social security was 75.2 %, in Darien this
number only reached 10.8 %. In Panama city, in 1991, there was an average
of 10.7 physicians per 10,000 inhabitants and 5.2 beds in hospitals per 1,000
inhabitants, whilst in Darien these numbers were 4.0 and 2.7, respectively.
These statistics illustrate the difference between the more and less developed
areas in the country (Anonymous, 1992).
1.1.2.2 Major Health Problems In Panama.
The death rate, in 1990, was 4.1 per 1,000 inhabitants and the child death
rate was 18.9 per 1,000 live birth, whereas the child birth rate was 24.8 per
1,000 inhabitants and the male expectation of life, reached 72.2 years. Table
1.2 (p.23) shows these health indicators during 1990 and compares the urban
and rural areas (Anonymous, 1992).
The main causes of death in 1990 were: malignant tumours (15.5 %),
accidents and other violent deaths (14.1 %), cerebrovascular diseases (10.8
%), acute myocardial failures (8.2 %), other heart diseases (5.1 %), pulmonary
circulation and related diseases (5.8 %), diabetes mellitus (3.1 %), pneumonia
(3.1 %) and congenital abnormalities (2.9 %). Ten percent of child deaths,
between 1 and 4 years old, was caused by malnutrition and anaemia. The
major causes of death in children under one year old include intestinal
infections, anaemia, pneumonia and congenital abnormalities.
Panama has being threatened by many infectious diseases in recent years.
Diseases such as haemorrhagic dengue and yellow fever, which are viral
diseases transmitted by the mosquito Aedes aegypti. The haemorrhagic
dengue is causing disasters in neighbouring countries such as Colombia and
more recently in Costa Rica. In Panama, both diseases, have been effectively
controlled, although the infestation of mosquitoes, in Panama city, is above the
minimum levels needed to spread these diseases.
21
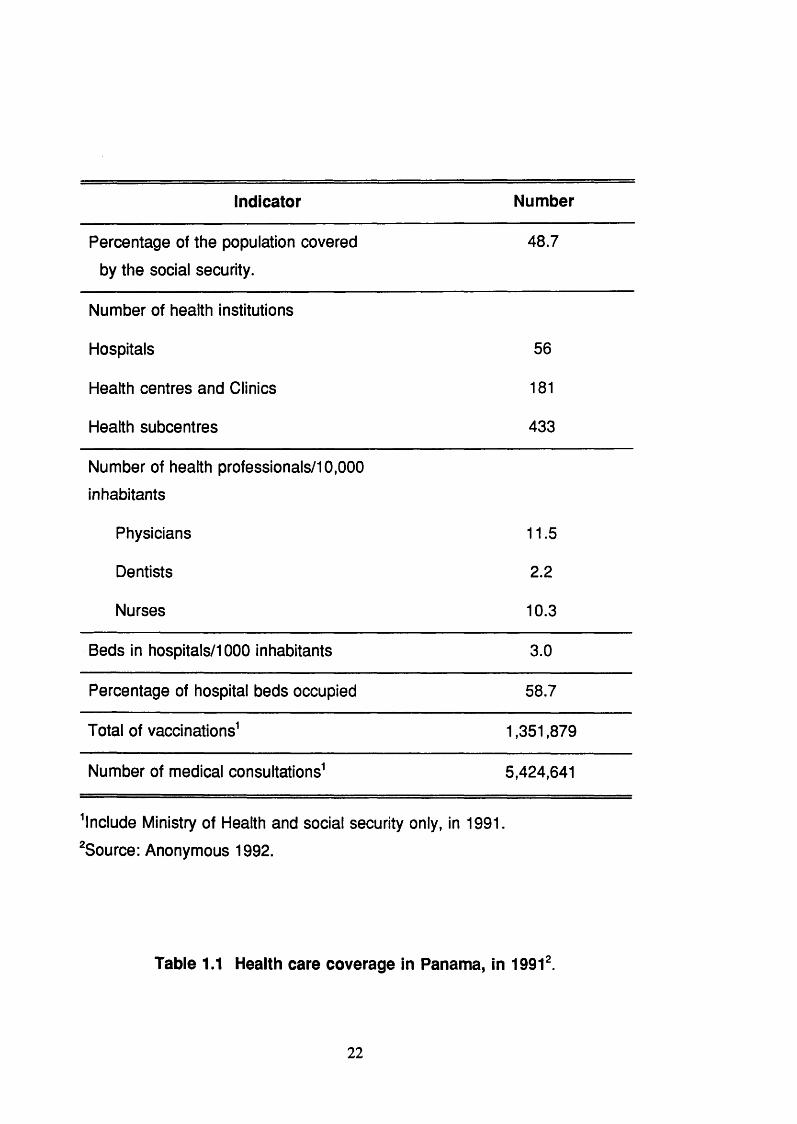
indicator Number
Percentage of the population covered 48.7
by the social security.
Number of health institutions
Hospitals 56
Health centres and Clinics 181
Health subcentres 433
Number of health professionals/10,000
inhabitants
Physicians 11.5
Dentists 2.2
Nurses 10.3
Beds in hospitals/1000 inhabitants 3.0
Percentage of hospital beds occupied 58.7
Total of vaccinations^ 1,351,879
Number of medical consultations^ 5,424,641
^Include Ministry of Health and social security only, in 1991.
^Source: Anonymous 1992.
Table 1.1 Health care coverage in Panama, in 1991‘
22

Health Indicator Total Urban Rural
Child birth rate/1,000 inhabitants 24.8 21.4 28.6
Death rate/1,000 inhabitants 4.1 4.1 4.0
Child death
rate/1,000 live births 18.9 17.9 19.7
Maternal death
rate/1,000 lives birth 0.5 0.3 0.8
Professional assistance during
the child birth, in percentage 86.3 99.3 75.4
Expectation of life in years (male) 72.4 74.1 69.9
Source: Anonymous 1992.
Table 1.2 Health Indicators In Urban and Rural Areas in Panama, 1990\
In January, 1991, epidemic choiera emerged in Peru and was spread to
northern countries such as Colombia and imported to Central American
countries such as El Salvador and Nicaragua (Glass et al., 1991). The first
case of cholera in Panama, was reported in September 1991 and it reached
a total of 1180 cases, in the same year, including 29 deaths, this number was
increased to 2,418 cases in 1992; whereas in 1993 the total was 42 with 4
deaths and the last case was reported in May of the same year (Anonymous,
1993c). This case may reflect the capacity of the health authorities in Panama
with respect to the neighbouring countries, where cholera is still a major
problem.
Levels of infestation of the mosquito Anopheles, the malaria transmitter, is
23

low in the urban areas in Panama. Two third of the malaria cases, in the
Republic, occur in the Province of Darien and the Indigenous reservation in San
Bias, both bordering with Colombia, where malaria is an endemic disease.
Since 1984, when AIDS (Acquired Immune Deficiency Syndrome) appeared
in Panama, a total of 606 cases and 352 deaths has been reported until the
end of 1993. Two percent of the cases have been transmitted by blood
transfusion, 3.0 % by perinatal transmission, 4.1 % by illegal drug users sharing
their syringes and 83.2 % by sexual contact.
Tuberculosis, is one of the diseases growing throughout the world, and in
Panama, the rate of cases seems to be constant, with about 30 cases per
100,000 inhabitants, in the last three years. The number of cases of some
infectious threatening diseases is presented in Table 1.3 (p.24).
Diseases
Year
1989 1990 1991 1992 1993
AIDS 77 64 70 N.A 78'
Cholera N.C. N.C. 1180 2418 42
Malaria 427 381 1115 727 300'
Tuberculosis 676 799 824 714 731'
N.A.= Data not available. N.C.= No reported cases.
^Data until October 1993.
^Source: Anonymous 1992. and Anonymous 1993c.
Table 1.3 Number of Cases of Some Life Threatening Diseases in
Panama^
24

1.1.3 Ethnomedicine in Panama.
In Panama, traditional healers are prohibited by law to practice their healing
arts. They are, nevertheless, active in both rural and urban areas and the
Government has no plans to include folk healers in health services. Traditional
medicine is more frequently used by the poor section of the Amerindian
population, and the rural populace and some herbs are sold regularly in the
public market and in the drug stores. Despite the World Health Organization
policy urging countries to utilize their traditional system of medicine in order to
achieve the policy of health for all by the year 2000 (Akerele, 1991) there is no
general policy or guidance on the use of traditional medicine, the cultivation,
sale or use of medicinal plants.
Today in Panama there are five Amerindian groups (Torres de Arauz 1980):
1.- Kuna live in the Archipelago of San Bias, and around Bayano,
Chucunaque and Tuira rivers. They are some 54,000 Amerindians
(Guionneau-Sinclair, 1989) who practice horticulture, fishing, hunting,
and collecting fruits, wild leaves and medicinal plants.
2.- Embera and Waunana are descendants of Colombian Chocoes and they
are concentrated around the borders of Darien Rivers, with a total of
17,264 Amerindians.
3.- Ngawbes or Guaymi proper is the most important group settled in the
Provinces of Chiriqui, Bocas del Toro and Veraguas. This is the most
numerous group with a total of 123,626 and they are the poorest
indigenous group.
4.- Bokotas live in the Provinces of Bocas del Toro and Veraguas, with only
3,784 Indians.
5.- Teribes live on the banks of Rio Teribe and San Durvi in the Province
of Bocas del Toro and in the Costarican side. In Panama there are
2,194 Indians.
The total population of the five Amerindian groups is approximately 9% of
25

the total population of the country. Although the Government of Panama has
made efforts to provide health care through health centres, health posts and
hospitals, these ethnic groups continue to use a variety of medicinal plants for
curing their ailments. Figure 1.1 (p. 27) shows the distribution of the different
indigenous groups in Panama.
Ethnobotanical and ethno-ecological studies have been done with the
human groups in eastern Panama, especially the Kuna and Embera Indians,
as part of the sea level canal studies (Duke, 1967; Duke, 1968; Duke, 1972;
Torres de Arauz, 1972). Folk herbal remedies of the Afro-AntiMeans in Colon
have been reported (Angermuller, 1968). Similarly, there have been
ethnobotanical and phytochemical evaluations of other human groups in
Panama, especially mestizo groups of the central provinces (Gupta et al., 1979;
Joly, 1981 ; Gupta et al., 1986).
More recently, two major ethnobotanical inventories have been done with
the Guaymi group in western Panama (Joly et al.,1987; Joly et al., 1990) and
the Kuna Indians in San Bias, eastern Panama (Gupta et al., 1993a), using the
questionnaire recommended by World Health Organization.
The ethnobotanical inventory with the Guaymi (Joly et al.,1987; Joly et al.,
1990) was carried out in collaboration with four "curanderos" (witch doctors).
The "curanderos" guided the daily collection trips. Interviews were taken in the
Guaymi language and translated into Spanish. A total of 104 plants was
identified and their uses and preparation were reported.
In the ethnobotanical study of the Kuna Indians (Gupta et.al., 1993a), 90
species were reported. In this study 54.4% of the plants were used topically,
26.7% used internally and 18.9% were used externally and internally. The
common illnesses for which these medicinal plants are used, include muscle
and joint aches, fever, skin infections, eye infections, asthma, wound healing,
snakebite, childbirth, acne, colds and general weakness.
26
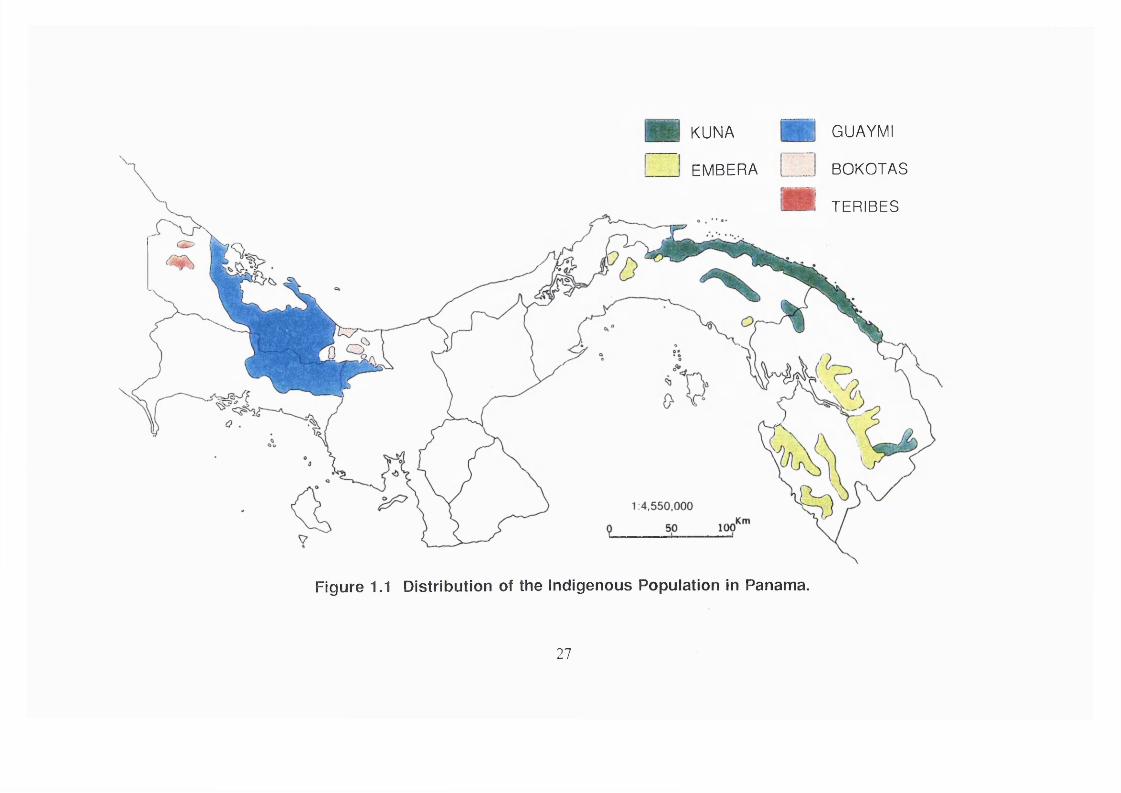
KUNA
EMBERA
GUAYMI
BOKOTAS
TERIBES
Figure 1.1 Distribution of the Indigenous Population in Panama.
27

1.1.4 Natural Resources and Conservation.
The Flora of Panama is one of the richest in the world (Schultes 1972), as
plants from North and South America can be found there. About 750 botanists
or naturalists have collected plants in Panama for herbarium collections.
According to Dwyer (1964; 1985), there are about one half million specimens,
excluding duplicate materials, of Panamanian plants in herbaria throughout the
world. The Missouri Botanical Garden has the largest collection. There have
been three distinct periods of plant collection in Panama: 1700-1914, 1915 to
1957 and 1958-1981. During the first period, Europeans dominated the scene.
Berthold Seemann wrote the first flora of Panama and Henry Pittier’s collections
are noteworthy (Dwyer, 1985). During the second period, Paul Standley made
a significant contribution and wrote Flora of Barro Colorado Island (Standley
1927) and Flora of the Canal Zone (Standley 1928). During the second and
third period the Missouri Botanical Garden has been the most active. Although
publication of the Flora of Panama has been completed, new species are still
being found and many areas remain to be explored. The Flora of Panama
recognized 5314 species and a total of 5597 taxa of species rank or below, by
1987 this number was increased to 7345 species and included 195 families
(D'Arcy, 1987).
The total number of vascular species in Panama is estimated between
8,000 and 10,000 species, of which 17.3% are endemic. There are also large
numbers of non vascular species in Panama, but they have not been
investigated fully. Flowering plant-richness and biodiversity of Panama ranks
fourth in North and Central America (World Conservation Monitoring Centre,
1992). In Panama there are over 891 species of ferns alone and about 1000
species of orchids, of which 10% are endemic. The epiphytes, the climbers,
and lianas are a major component of the Panamanian tropical forest. Mosses
abound in moist forests as well as other parts of Panama. Mangroves are
characteristic plant formations of tropical seacoasts. Tables 1.4 -1 .7 (p.29, 30,
31, 33) provide general information on the Panamanian Flora including major
28

life forms, major plant families and geographic distribution. Figure 1 . 2 (p.32)
shows the different vegetation zones in Panama.
The Isthmus of Panama constitutes the narrowest territorial portion of the
American continent where North and South America join. Panama, situated in
biogeographic regions of the Neotropics, plays an important role as a biological
bridge between the northern and southern lobes of the Continent. Indeed,
three of the four principal routes of bird migration converge in its territories
(Cobos-Moran, 1992).
Number Habit Percentage^
895 Climber 12.3
195 Epiphyte 2.7
2 Hemiepiphyte 0.03
3,126 Herb 42.9
1 1 Parasite 0 . 2
1,952 Tree 26.8
53 Treelet 0.7
56 Without Habit (not available
from present data)
'Taken from D’Arcy, (1987).
^Do not total to 1 0 0 % because of multiple categories
Table 1.4 Major life forms of the vegetation In Panama'.
29

NUMBER OF
SPECIES"
Percentage"
Monocotyledons 2,246 27.6
Dicotyledons 5,099 62.6
Total of native species 7,123 87.4
Endemic species 1,230 17.3
Introduced species 2 2 2 2.7
Species of flowering plants 7,345 90.2
Pteridophytes species 800 9.8
Total number of vascular plants 8,145 1 0 0
^Taken from D’Arcy, (1987).
Arcy, W. G. (1987), "Since 1981 over 330 species of flowering plants have been newly described from material collected in Panama, and no diminution of this rate of discovery has been seen to date. Thus, the total flora of Panama may include 8,500-9000 flowering plants and 900 pteridophytes. These numbers will decrease as habitats are eliminated from Panama and species are extirpated or extinguished".
t o not total 1 0 0 % because of multiple categories.
Table 1.5 General Information on the Panamanian flora\
30

Region Number of Species Percentage^
Canal Area 2,545 (35)
Chiriqui 3,057 (42)
Panama 3,246 (44)
Western Panama (Bocas del Toro, Chiriqui)
3,921 (53)
Central Provinces (Canal Area, Codé, Colon, Herrera, Los Santos, Panama, Veraguas)
5,146 (70)
Azuero Peninsula (Herrera, Los Santos)
801 (1 1 )
Eastern Panama (Darien, San Bias)
2,563 (35)
All Panama 7,345 (1 0 0 )
^Taken from D’Arcy (1987).
^Do not total 1 0 0 % because plant species had been collected in different regions.
Table 1.6 Geographic distribution of plant species in Panama^
31

P rem o n lan e ram forest
H um id low m o n tan e forest
Very tiumid low m o ntane forest Low m ontane rain forest
Very tium id m o n tan e forest I — I M o n tan e rain forest
e m u D ry tropical forest
I I H um id tropical forest
i m c l Very fium id tropical forest
i I D ry p rem o n tan e torest
S B H um id p re m o n tan e forest [ » e a V ery tium id p rem o n tan e forest
1. 2 ,000,000
Figure 1.2 Different Vegetation Zones In Panama.
32

FAMILY NATIVE SPECIES ENDEMIC SPECIES^
MONOCOTYLEDONSORCHIDACEAE 891 113GRAMINEAE 344 1 1
ARACEAE 234 95CYPERACEAE 164 1
BROMELIACEAE 136 1 2
PALMAE 95 26SUBTOTAL 1,864 (85%)' 258 (85%)'
DICOTYLEDONSRUBIACEAE 431 194LEGUMINOSAE 430 35COMPOSITAE 272 42MELASTOMATACEAE 244 43PIPERACEAE 213 80GESNERIACEAE 159 63SOLANACEAE 138 27MYRSINACEAE 134 95EUPHORBIACEAE 134 7ACANTHACEAE 1 1 0 15MORACEAE 96 4SAPINDACEAE 85 23VERBENAGEAE 81 5ERICACEAE 81 41BIGNONIACEAE 80 4
SUBTOTAL 2 , 6 8 8 (53%)' 678 (69%)'TOTAL IN THIS GROUP 4,552 (64%)' 936 (72%)'
^Taken from D’Arcy, (1987).^The number of endemic species is higher than that reported in the Flora of Panama because of the inclusion of 60 varieties.^The figures in parenthesis represent the percentage with respect to the total number of species in the Flora of Panama.
Table 1.7 Plant families In Panama with more than 80 species^
33

Panama] comprises a terrestrial bridge of extreme biological importance.
Presently, throughout the Isthmus, there is an amalgam of biotas of the North
and of the South. A consequence of this mixing is a vast plant and animal
diversity. This diversity is supported by a unique collection of environmental
factors, which influence and allow the coexistence of so many species (Cobos-
Moran, 1993). Therefore, its integrity as a biological bridge is necessary for the
continued existence and evolution of the species of this important region of the
planet.
In Panama, in 1947, 70% of the territory, excluding the Panama Canal
Area, was covered by forests. In 1980, primary forests covered a land area of
3,549,700 hectares. More recently, the National Institute of Renewable Natural
Resources of Panama (INRENARE) estimated that this number in 1991 was
reduced to only 40% and it is estimated that by the year 2000 this percentage
will be reduced to 33%. The annual rate of forest loss is estimated at 65,000
hectares (Cobos-Moran, 1992). Therefore, it is vital to study this flora before
the species become extinct.
There are two major institutions responsible for conservation in Panama; the
National Institute of Renewable Resources (INRENARE) on the Panamanian
Government behalf and a private group called Ancon. INRENARE and Ancon
are managing fourteen national parks that cover a land surface of 1,389,463
hectares that represent 18.4 % of the national territory; seven forest reserves;
six wild life refugees and seven other minor categories of management. The
total land surfaces under conservation by law reach 27.5 % (2,077,914
hectares) of the total surface of the country. This number does not include the
indigenous reserves that are conservationist by nature; noteworthy are the
Kuna Indians recognized by the WWF (World Wild Foundation) as an example
of how mankind can live in harmony with the wild life. Figure 1.3 (p.35) shows
the protected areas in Panama during the year 1985, whilst Figure 1 . 2 (p.32)
shows the distribution of the indigenous population.
34
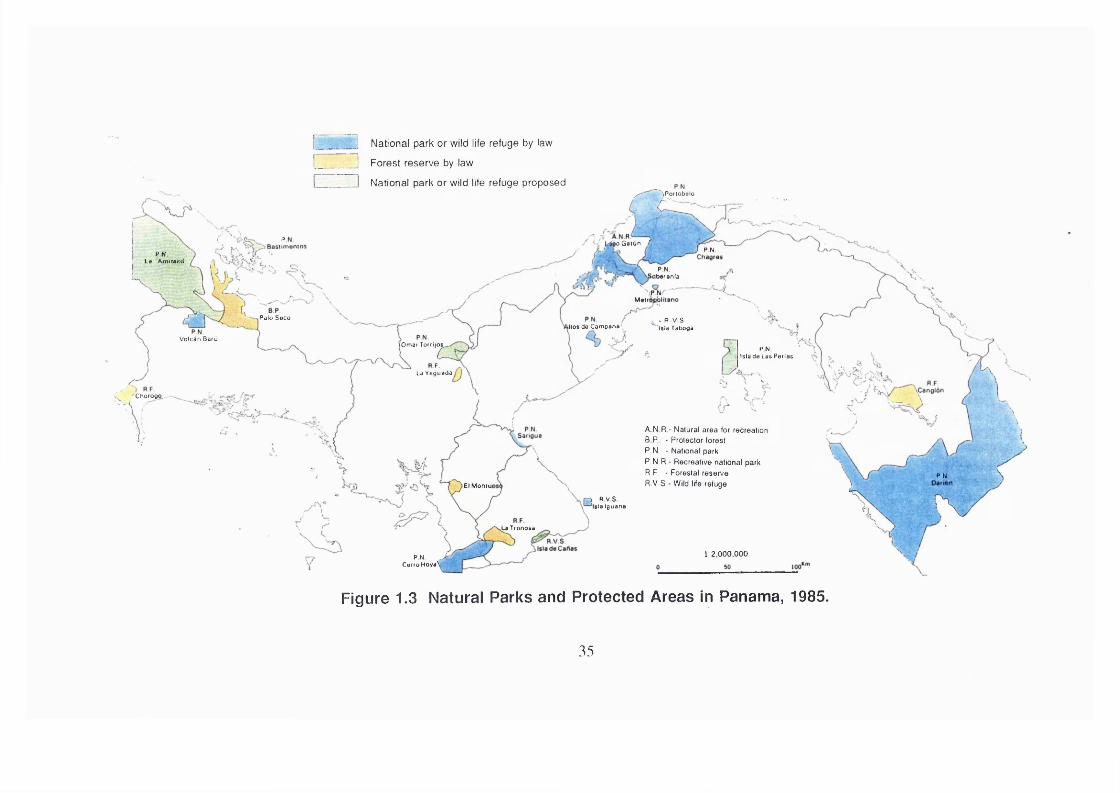
I National park or wild life refuge by law
! Forest reserve by law
N ational park or wild life refuge proposed^ ~ ^P oflobalo
g o G a lun f .( I} PN
P N Ls Amisied
PN>barsnla
",P N /- 'Main
Palo Seco - R V S Isia Tabogakilo* da Campana'
Voicàn BarùlOmar Torrijo^
PNIslade Lae Perlae
la Veguada
Chorogo.. —
A N R - N atu ra l a rea for recreation
B P - P rotector forest
P .N . - N ational park
P .N .R .- R ecrea tive national park
R .F . - Forestal reserve
R .V .S - W ild life refugePN
El Moniui
I R V S 'Id a Iguana
^ T io n o s a
l: 2,000,000P N > CarroHoyaV
Figure 1.3 Natural Parks and Protected Areas In Panama, 1985.
35

1.2 Selection of Plants for Chemical and Biological Investigation.
Some of the most important drugs of the past 50 years or so, which have
revolutionized modern medical practices, have first been isolated from plants,
and often from plants that for one purpose or another have been employed in
primitive or ancient society (Schultes, 1986). These drugs include, vinca
alkaloids used to treat leukaemia, quinine that is effective against malaria and
served as prototype in the development of synthetic antimalarial drugs,
podophyllotoxin in the development of synthetic anticancer drugs, morphine
alkaloids used as analgesic and antitussive drugs and in the study of the
biochemistry of pain, digoxin and other cardioactive glycosides, to mention
some examples. More recently, taxol, another drug obtained from plants is
used in the treatment of colon and breast cancer. Furthermore, in 1973, in
United State 25.2 % of the prescriptions contained one or more active
constituents obtained from higher plants, including 76 different chemical
compounds of known structure derived from higher plants (Farnsworth and
Bingel, 1977). Therefore plants should be considered as a primary source of
drugs to treat different diseases, of chemical templates to build up more
effective drugs and of compounds which may be used as pharmacological tools
to give a better understanding of biological processes.
There are three principal approaches used to select plants for chemical and
biological investigations, namely, the use of plants in traditional medicine,
random selection of plants for massive studies and the chemical relationship
of the plant species, chemotaxonomy.
1.2.1 Traditional Medicine.
Plants have been used by man kind since early times and it seems that
Neanderthals used plants as medicine (Solecki, 1975). Also, a large number
of plants have been used for more than 3,000 years in the Chinese traditional
medicine. Ayurvedic medicine and Unami medicine (Farnsworth and Soejarto,
36

1991).
The World Health Organization (Penso, 1983) has attempted to identify all
plants used in medicine around the world. More than 20,000 species have
been listed providing the latin name of the plants and the country where the
plants were used. More recently, a data base, named NAPRALERT, has been
established at the University of Illinois, where ca of 9,200 species have been
documented (Farnsworth, 1983; Farnsworth and Soejarto, 1991). This data
base includes the scientific name of the plants, synonyms, the use, how they
are used including the dose in some cases, vernacular names, country where
the species are used and the reference where the statement was published.
It is estimated that 74% of the 121 biologically active plant-derived
compounds presently in use worldwide, have been discovered by studies that
based the plant selection on ethnomedical information (Farnsworth et a i 1985).
There is a great deal of information regarding the medicinal use of plants and
the reliability of the information should be taken in account to select a plant for
chemical investigations. How this information was collected, who is the
informant and if a particular species or genus is used for different population
groups to treat a particular disease. Also important is the interpretation of the
data, as usually a symptom is reported rather than a disease; for example, the
term intermittent fever may suggest malaria whereas the term backache could
suggest either renal disease or muscular distention.
The way in which the information is collected is crucial in order to have
valuable information. Some foreigner researchers go to live with aborigine
populations, to gain their confidence and a few months later a paper is
submitted to a scientific or medical journal. Most aborigines are reluctant to
give information and although, sometimes, they are paid for it, the information
lacks value because it is not possible to buy their confidence. In the last two
ethnobotanical inventories in Panama (Joly et al., 1987; Joly et al., 1990; Gupta
et al., 1993a) a descendant from the community to be studied was chosen
37

among the final year students at the University of Panama, to overcome the
confidentiality task and the language limitations. Many differences were found
between the work carried out by Duke (1975) and that more recently reported
by Gupta (1993a) with the Kuna Indians. Similarly, Joly (1990) found some
differences between her work and that from Hazlett (1986) with the Guaymi
Indians.
The information on medicinal plants may be obtained from a variety of
forms. Among the firsts to write about healing properties of plants were the
Chinese and Ayurvedic "doctors", which received long training before they are
allowed to prescribe. Then, there are the Shaman in Peru and Sukias and
Inadulet in Panama, who received some training before they were allowed to
prescribe. Also there is information on those practitioners that receive little or
no instruction and the knowledge which they receive has been handed down
from the father to the son.
1.2.2 Random Selection of Plant for Chemical Investigation.
Perhaps, the random selection approach is the most expensive and
unscientific way to select plants for chemical and biological evaluations. The
limitations of random selection of plants have been described (Spjut, 1985).
Nevertheless, some promising antitumours agents have been discovered
through this approach under the program of the National Cancer Institute. This
program, started to screen plant extracts in 1960, twenty years later, in 1980,
114,045 extracts had been screened and 4.3% of the extracts showed activity.
This massive program yielded seven compounds, which were in clinical trial by
1980 (Suffness and Douros, 1982).
These compounds included bruceantin and maytansine that showed weak
activity and they were not developed further (Suffness and Douros, 1982).
Although, indicine N-oxide, in phase II of the clinical trial, showed activity in
acute leukaemia; it is ineffective in the treatment of osteosarcoma.
38

neuroblastoma and paedriatic brain tumours, and hepatotoxicity has been
shown at therapeutic doses (Miser et al., 1992; Miser et al., 1991).
Homoharringtonine, a cephalotaxine alkaloid, is safe and effective for patients
with acute myelogenous leukaemia (Feldman et al., 1992). Phyllantoside,
isolated in 1977 (Kupchan, et al., 1977) is active against the B16 melanoma.
4p-Hydroxywithanolide E with an a oriented side chain at C-17 was undergoing
formulation studies for toxicology (Suffness and Douros, 1982). Finally, taxol
isolated from Taxus brevifolia (Wani, et al. 1971 ) has remarkable anti neoplastic
qualities against ovarian cancer, melanoma and colon cancer (Edington, 1991 ).
1.2.3 Chemotaxonomy
Secondary metabolites present in plant cells represent not only a chemical
entity, but are the manifestation of a whole series of enzymes, which in turn are
the genetic expression. Hence, chemical characters could show the
relationship between plant individuals and their evolution. Since the presence
of secondary metabolites in plants could predict the relationship between plant
species, this approach could be used to select plants for chemical and
biological investigations. It is mainly useful to find new or more rich sources of
a particular compound, or to find more active or selective structural analogues.
The foxglove (Digitalis purpurea) was introduced into modern medicine on
the basis of its use as heart stimulant in folk medicine, but the active principle,
digitoxin, has short latent periods of action and low cumulative effects.
Synthetic variants of digitoxin have proved not to be as effective as natural
variants of the drug in related species of Digitalis. Thus, D. lanata, a species
native to southeastern Europe, was found to contain three to five fold greater
concentration of active principles than the foxglove (Hansel, 1972) and one of
the active principles, digoxin, has relatively better pharmacokinetics properties
than digitoxin.
The first report of the relationship between chemical components and races
39

of plant was written in 1673 by Nehemiah Grew, who used medicinal plants in
his examples. The second, James Petiver, an eminent London apothecary,
who wrote, in 1699, "some attempts made to prove that herbs of the same
make or class for the generality, have the like virtue and tendency to work the
same effects" (Gibbs, 1963).
However, it was not until the beginning of this century when Gresshoff, in
1909, demanded that every accurate description of a genus or of a new species
should be accompanied by short chemical description of the plant (Gibbs,
1963). Later, McNair attempted to apply comparative chemistry generally to
taxonomy utilizing the fats and oils content of the plants (McNair, 1929). Also,
McNair, paid attention to other groups of compounds including the alkaloids of
aconite (McNair, 1935). There have been a number of treatises on
chemotaxonomy including Manske and Holmes (1950-1958), Hutchinson (1959)
and Hegnauer (1962-1994). The chemotaxonomy of the Papaveraceae
(Santavy, 1970) and Leguminosae (Harborne, etal., 1971) have been reviewed.
Modern techniques for the isolation of natural products, such as
chromatography (TLC, CC, HPLC, GC, DCCC, etc.), and physical methods for
structure elucidation, such as mass spectrometry (EIMS, FABMS, CIMS,
FDMS, etc.) and nuclear magnetic resonance ( H, 2D experiments such as
COSY 45, HMQC, HMBO, NOESY, etc.) now make it possible to fully
characterize small amounts of compounds present in plants. In addition, the
readily available literature on natural products allows for correlations to be
made between the chemical composition and evolution within the plant
kingdom.
Plants are widely used in Panama and Central America to cure different
diseases, including malaria and amoebic dysentery (Morton, 1981 ; Joly et al.,
1987; Joly et al., 1990; Gupta et al., 1986). Species of Simaroubaceae,
Meliaceae and Menispermaceae are known to contain quassinoids, limonoids
and bisbenzylisoquinoline (bbiq) alkaloids, respectively, with activity against
40

Plasmodium falciparum. In addition, Cephaëlis ipecacuanha is known to
contain emetine, which is used in the treatment of severe amoebic dysentery
(Tyler et al., 1988). Interest in antiprotozoal natural products and the possibility
of obtaining quassinoids, limonoids, bbiq’s and emetine related alkaloids led to
the decision to investigate the plants listed in Table 1.9 (p.49).
1.2.3.1 Chemotaxonomy background
Taxonomy is a study aimed at producing a system of classification of
organisms, which best reflects the totality of their similarities and differences
(Cronquist, 1968) and chemotaxonomy incorporates the principles and
procedures involved in the use of chemical evidence for classification purposes.
Features used in a classification are termed taxonomic characters. All sources
of taxonomic evidence are scanned in the search for taxonomic characters and
among the richest have been the fields of morphology, anatomy, cytology,
ecology and genetic. The use of chemical data as taxonomic characters marks
a recent extension of the range of recognized sources of taxonomic evidence
(Smith, 1976).
Taxonomic characters should be easy to assay and the unambiguity of
modern chemical analytical procedures enables accurate identification of
individual compounds. Also, the taxonomic characters, should be consistently
present in a taxon, bearing in mind the nature of natural grouping, which allows
exceptions to any generalization. To affirm that a characteristic is consistently
present in a taxon implies that it is to be expected in a high proportion of its
members. In addition, a taxonomic character, should not be affected by
environmental factors, unless the taxonomist is equipped to detect the property
of variability itself and use it taxonomically (Heslop-Harrison, 1963). The
chemotaxonomic character, in turn, should be easily detected or identifiable,
with limited distribution i.e. within a genus of a family, the biosynthetic pathway
should be fully understood and its presence should not be seasonal or climatic-
condition dependant.
41

1.3 Biological Testing of Plant Extracts.
Unfortunately, there was, and still is, little communication between
phytochemists and pharmacologists. New compounds are isolated from plants
and their structures elucidated, but they lie dormant on the phytochemist’s
shelf. Usually, minute quantities of the compounds are isolated and then is not
enough for biological activity testing (McLaughlin, 1991). Pharmacologists, on
the other hand, are reluctant to test tarry plant extracts.
Isolation of active compounds from plants by means of bioactivity guided
fractionation are time consuming and expensive. Nevertheless, it is the way to
obtain meaningful and significant results. Using a bioactivity-guided
fractionation in the isolation of a natural product not always ends with a novel
compound, but, perhaps, with known compounds with a novel application in
medicine.
Many scheme fractionations have been proposed to follow up the activity
of plant extracts ( e.g. Ferrigni et al., 1984; Samuelsson et al., 1985; O’Neill et
al., 1987). Although, in traditional medicine most of the remedies are prepared
by extraction with water, either by boiling the plant parts or by soaking them in
cold water, most of the researchers use polar or moderately polar organic
solvents. Aqueous extracts tend to be avoided due to their complexity and
difficulties in developing suitable work up procedures (Samuelsson et al., 1985).
Also, strategies for pharmacological evaluations of crude drugs prescribed in
traditional medicine have been reported (e.g. Kyerematen and Ogunlana, 1987).
However, the most important is to biologically test all the fractions at each step
and all the isolated compounds should be tested in different bioassays.
Therefore, the assays designed to guide the fractionation should be simple,
rapid, reliable and inexpensive.
The bioassay should be highly sensitive because most natural products are
present in the crude extract at dilutions of 1 :1 , 0 0 0 or more even up to
42

1:1,000,000. It is highly likely that in vivo screening alone is going to miss
compounds that may be quite active but are not potent enough of not
concentrated enough in a crude extract for detection. That is one of the main
reason why in vitro methods are preferred to in vivo screens (Suffness and
Douros, 1982). However, the bioassay used in screening crude plant extracts
must be insensitive to the many compounds or ubiquitous compounds,
which give false actives. The National Cancer Institute, for example, dropped
the Walker 256 screen from use in the plant programme because it was highly
sensitivity to tannins (Suffness and Douros, 1982). Apart from the general
requirements of any good assay, such as validity, predictability, correlation,
reproducibility and reasonableness of cost, the following considerations should
be taken in account in bioassays used to screen plant crude extracts (Suffness
and Douros, 1982):
1 .- Selectivity: The assay must be selective enough to limit the number
of false positives.
2 .- Sensitivity: The assay must be very sensitive in order to detect active
compounds in low concentrations.
3.- Methodology: The assay must be adaptable to materials that are
highly coloured, tarry, poorly soluble in water and chemically complex.
1.3.1 Advantages and Disadvantages of in vivo and in vitroTesting.
Table 1 . 8 (p.44) summarize the advantages and disadvantages of the in
vitro and in vivo bioassays. Research with experimental animals does not
provide the only major route to advances in biological understanding and
delivery of medical benefits, but it does provide an approach that is very
important, such as pharmacokinetic and bioavailability, and one that is likely to
remain so for the foreseeable future. Animal usage might become,
progressively, superseded in the preparation of antibody-like molecules.
43

however they have to be made by immunisation of animals (Rees, 1992) and
in the search of drugs targeting specific peptide receptors such as analgesic
drugs acting on bradykinin receptors, but still isolated from rat uterus (Snell and
Snell, 1989).
Advantages Disadvantages
In vivo Activity data Long turn over
Expensive
Often less sensitive
Relatively large sample
needed
In vitro Speed In vitro data only
Less costly Activity may not
Sensitivity correspond to in vivo
Small sample size activity.
Table 1.8 Advantages and Disadvantages of in vivo and In vitro
Bloassays.
A criticism often repeated by opponents of animal experimentation is that
"animal and human diseases are rarely, if ever, identical. Using an animal
model that is not identical to human disease is a logically flawed process"
(Anonymous, 1991 ). There are metabolic and toxicological differences between
mouse and man and the data from mice has been found misleading, specially
in cytotoxic anticancer drugs. It is now considered desirable to carry out such
studies as soon as possible in human cancer patients following the minimum
of animal studies to ensure some degree of relative efficacy/safety of the drugs
for man (Parke, 1983).
44

Similar anomalies in rodent studies of the carcinogenicity of chlorinated
hydrocarbon pesticides led the Joint Meeting of FAQ and WHO on Pesticides
Residues to reject carcinogenicity tests in the mouse as being predictive of
potential hazard to man (Anonymous, 1981 ). The lack of relevance of the long
term animal carcinogenicity to man and the need for more rapid and less
expensive assays has led to the development, and acceptance by regulatory
authorities, of a number of short-term in vitro tests (Parke, 1983). Among
these, the Ames test (Ames et al., 1975) is the most popular and successful
and has been adopted by the Committee on Safety of Medicine as an
alternative to in vivo carcinogenicity studies in experimental animals (Parke,
1983).
In vitro assays may require the presence of mammalian metabolic
activation preparations to reproduce some of the aspects of whole animal
metabolism of foreign compounds. The procedures for such activation mixes
with mammalian ceils are far from optimal and their presence often leads to
cellular toxicity (Scott, 1982).
1.3.2 Simple Bloassays.
1.3.2.1 Brine Shrimp Assay.
Artemia salina Leach (brine shrimp) is a small shrimp commercially used
as a food for tropical fish and it has been used at different life stages in
experimental researches. Brine shrimp has been used in the analysis of
pesticides residues (Tarpley, 1958), mycotoxins (Reiss, 1972; Tanaka et al.,
1982; Hoke et al., 1987), metals (McRae and Pandey, 1991), protein
biosynthesis inhibitor (Robyn and irvin, 1980), cocarcinogenic phorboi esters
(Kinghorn et ai., 1976), anaesthetics (Robinson et ai., 1965) and dinofiagellate
toxins (Granade et al., 1976).
45

Also brine shrimp has been used to guide fractionation and isolation of
mycotoxins (Eppley and Bailey, 1973), plant neurotoxins (Greig et al., 1980)
and antibiotics (Hamill et al., 1969). It has been used in field evaluation of
traditional medicine (Trotter et al., 1983; Beioz, 1992) and placed in the second
level of evaluation of traditional materia medica (Kyerematen and Ogunlana,
1987). Furthermore, Artemia salina has been subjected extensively to
comparative biochemical and anatomical studies ( McRae et al., 1989; Warner
et al., 1988).
In 1982 a simple method, using brine shrimp, was developed for screening
and fractionation of active materials from higher plants (Meyer et al., 1982) and
demonstrations of the use of brine shrimp bioassay in the isolation of bioactive
natural products have been reviewed (McLaughlin et al., 1993). The use of
potato disc and brine shrimp bioassays to detect activity and isolate
antileukaemic natural products have been reported (Ferrigni et al., 1984). In
a blind comparison of brine shrimp and human tumour cell cytotoxicities as
antitumour prescreens, brine shrimp prove to be superior or equally accurate
as the in vitro human solid tumour cell lines (Anderson et al., 1991a). A
convenient bioassay to detect antiparasitic avermectin analogues has been
reported (Blizzard et al., 1989).
As the brine shrimp test (Meyer et al., 1982) requires relatively large
quantities of material (20 mg for crude extracts and 4 mg for pure compounds)
and the preparation of dilutions is time-consuming thus limiting the number of
samples and dilutions that can be tested in one experiment, it will be attempted
to develop a microdilution technique to overcome the above disadvantages.
1.3.2.2 KB Cells Assay.
Since 1960 the in vitro KB assay has been used by the National Cancer
Institute (NCI) as a preliminary screen for cytotoxicity and for fractionating plant
samples before carrying out assays for in vivo activity. This in vitro system is
46

an excellent bioassay but it is a poor screen because of the sensitivity of the
cells to cytotoxic substances that are devoid of in vivo activity (Suffness and
Douros, 1982).
The KB cell in vitro test was initially described by Eagle in 1955 and Oyama
and Eagle in 1956. The assay has since been standardised by the NCI (Geran
and Greenberg, 1972) and later modified by Wall et al. (1987). More recently,
a microdilution technique was developed for the assessment of in vitro
cytotoxicity against KB cells derived from a human epidermoid carcinoma of the
nasopharynx (Anderson et al., 1991b).
1.3.2.3 Plasmodium falciparum Assay.
Malaria in humans is caused by four species of protozoal parasites of the
genus Plasmodium (P. falciparum, P. malariae, P. ovale and P. vivax). It is
characterized by fever and other symptoms at the time when the merozoites
are released from ruptured red cells so that intermittent fever is produced.
Anaemia occurs due to haemolysis and other factors. P. falciparum infection
is particularly dangerous as cerebral malaria may occur (Cattani, 1993).
The prophylaxis and treatment of malaria have become increasingly
complex and difficult because of the widespread resistance of P. falciparum to
drugs (Payne, 1987). Multidrug resistance to antimalarials, such as chloroquine
and quinine, and adverse effects of drugs, such as pyrimethamine-sulphadoxine
combination, have severely limited available therapy (Peters, 1985). New drugs
are urgently needed as resistance has occurred to the more recently introduced
mefloquine into clinical use. Resistance has been produced in the laboratory
against artemisinin and other antimalarials under development, so that new
drugs such as halofanthrine (Horton, 1988) may not have a long-term future
unless their use is strictly controlled (Warhurst, 1985).
47

The antiplasmodial in vitro test described by O’Neill et al (1985), with
modifications described by Ekong et al (1990), is based on the method of
Desjardins et al (1979). Cultures of P. falciparum are maintained in vitro in
human erythrocytes by a method described by Trager and Jensen (1976) and
later modified by Fairlamb et al. (1985). The technique measures the
incorporation of ^H-hypoxanthine into drug-treated infected red blood cells
compared to untreated infected red blood cells. The in vitro testing of plants
extracts has been reviewed (Phillipson et al., 1993, Phillipson et al., 1994).
More recently, a colorimetric assay has been described measuring the parasite
lactate dehydrogenase activity, which is distinguishable from the host lactate
dehydrogenase activity using the 3-acetyl pyridine adenine dinucleotide
analogues of nicotinamide adenine dinucleotide (Makler et al., 1993).
48

1.4 Aims of this Study.
The aims of this study are:
1 .- To investigate a small selected number of Panamanian plants. The chosen
plants are showed in Table 1.9 (p.49).
2.- To use bioassay guided fractionation procedures.
3.- To develop a brine shrimp microwell assay
4 .- To isolate and identify known compounds as well as to characterise and to
determine the chemical structures of novel compounds.
Plant Family Plant species
Meliaceae Guarea macropetala Pennington
Guarea rhopalocarpa Radlkofer
Ruagea glabra Triana & Planchon
Menispermaceae Abuta dwyerana Kruk. & Barneby
Rubiaceae Cephaëlis camponutans Dwyer & Hayden
Cephaëlis dichroa (Standley) Standley
Cephaëlis dimorphandrioides Dwyer
Cephaëlis glomerulata J. Donnel Smith
Lasianthus panamensis Dwyer
Simaroubaceae Picramnia antidesma
subsp. fessonia (DC) W.Thomas
Picramnia teapensis Tul
Tabie 1.9 Panamanian Medicinai Riants Seiected for Chemical and
Biological Studies.
49

Section 2. Plants Selected for Investigation

2.1 Meliaceous Plants.
jpredominantlyA tropical and subtropical family/of the Old World, the Meliaceae comprises
50 genera and more than 1000 species of herbs, shrubs and trees. It has been
divided into five subfamilies, based on characters of the stamens and seeds
(Schultes and Raffauf, 1990).
The family Meliaceae is known to contains limonoids (Connolly, 1983),
tetranortriterpenoids (Banerji and Nigam, 1984) and the chemistry of the family as
a whole has been reviewed (Taylor, 1983). Limonoids are a group of oxidized
triterpenes closely related to the quassinoids which occur in species of
Simaroubaceae (Connolly, 1983; Taylor, 1983; Banerji and Nigam, 1984) and
several showed moderate antimalarial activity, in vitro (Bray et al., 1990). The
most active of these, gedunin, had activity around three times higher than that
of chloroquine (Khalid et al., 1986; Bray et al., 1990).
The genus Guarea has 150 species of trees and shrubs in tropical America
and 20 in Africa (Schultes and Raffauf, 1990); 8 of them occur in Panama
(D’Arcy, 1987). Table 2 . 1 (p.52) shows the species of Guarea that have been
investigated chemically.
Biological activity for extracts of Guarea species: A water extract of the
leaves of G. guidonia showed no activity on guinea pig atrium (Carbajal et al.,
1991), whereas, an ethanolic extract of the seed showed antiinflammatory
activity in rats (Oga et al., 1981). Different extracts and fractions of the fruits,
stem bark and stem wood of G. multiflora showed no activity against
Plasmodium falciparum, in vitro (Bray et al., 1990). A fluid extract from the
bark of G. rusbyi was active against Mycobacterium tuberculosis, in vitro
(Fitzpatrick, 1954). The ether extract of the flower of G. sepium showed activity
against various species of helminths including Strongiloides stercoralis,
Ancylostoma caninum and A duodenale (Gilbert et al., 1972). The aqueous
extract of the bark of G. thompsonii showed a LC5 0 150 mg/Kg when
51
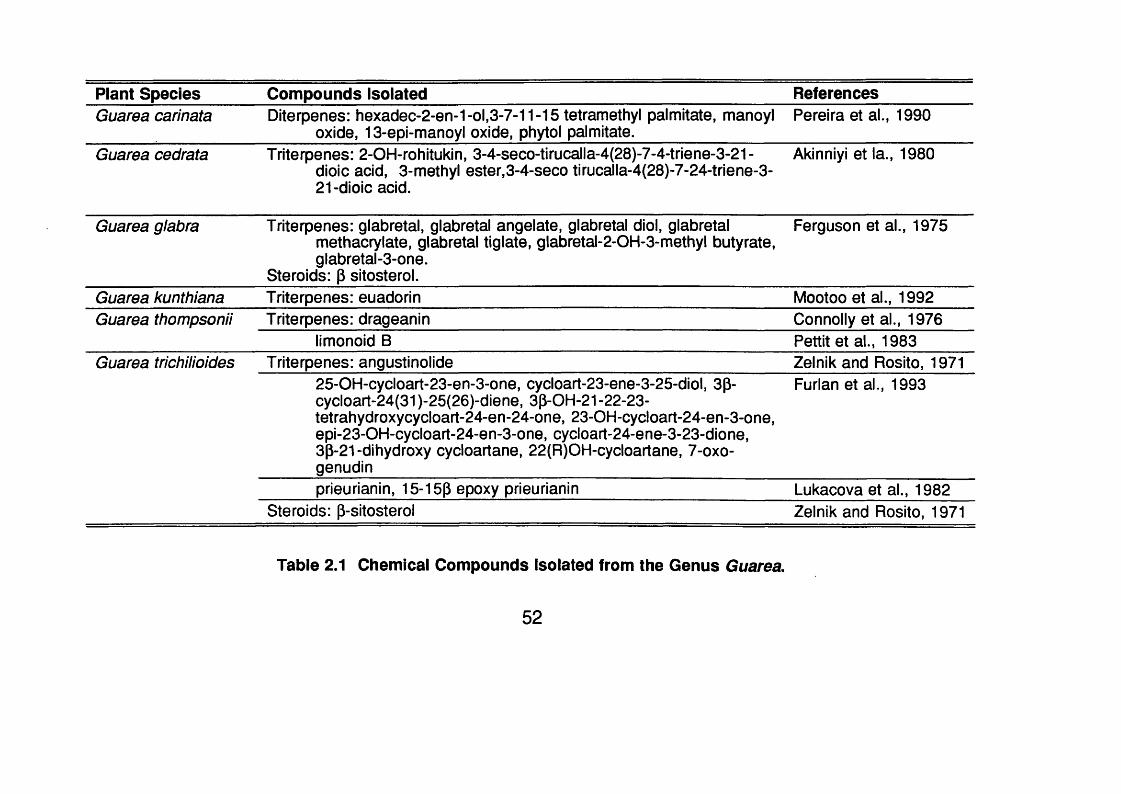
Plant Species Compounds Isolated ReferencesGuarea carinata Diterpenes: h0xadec-2-en-1-ol,3-7-11-15 tetramethyl palmitate, manoyl
oxide, 13-epi-manoyl oxide, phytol palmitate.Pereira et al., 1990
Guarea cedrata Triterpenes: 2-OH-rohitukin, 3-4-seco-tirucalla-4(28)-7-4-triene-3-21 - dioic acid, 3-methyl ester,3-4-seco tirucalla-4{28)-7-24-triene-3- 2 1 -dioic acid.
Akinniyi et la., 1980
Guarea glabra Triterpenes: glabretal, glabretal angelate, glabretal did, glabretalmethacrylate, glabretal tiglate, glabretal-2-OH-3-methyl butyrate, glabretal-3-one.
Steroids: p sitosterol.
Ferguson et al., 1975
Guarea kunthiana Triterpenes: euadorin Mootoo et al., 1992Guarea thompsonii Triterpenes: drageanin Connolly et al., 1976
limonoid B Pettit et al., 1983Guarea trichilioides Triterpenes: angustinolide Zelnik and Rosito, 1971
25-OH-cycloart-23-en-3-one, cycloart-23-ene-3-25-diol, 3p- cycloart-24(31 )-25(26)-diene, 3P-OH-21 -22-23- tetrahydroxycycloart-24-en-24-one, 23-OH-cycloart-24-en-3-one, epi-23-OH-cycloart-24-en-3-one, cycloart-24-ene-3-23-dione, 3p-21-dihydroxy cycloartane, 22(R)OH-cycloartane, 7-oxo- genudin
Furlan et al., 1993
prieurianin, 15-15P epoxy prieurianin Lukacova et al., 1982Steroids: p-sitosterol Zelnik and Rosito, 1971
Table 2.1 Chemical Compounds Isolated from the Genus Guarea.
52

administered intraperitoneaiiy in rats (Sandberg and Cronlund, 1977). An
aqueous extract of the fruits of G. trichilioides Vi/as inactive against Plasmodium
gallinaceum, in chicken (Spencer et al., 1947) and the chloroformic extract of
the root bark showed an ED5 0 0.29 pg/ml against LEUK-P388 cell culture
(Lukacova et al., 1982).
Guarea macropetala Pennington: There is no previous chemical or
biological study of this plant. It is a tree of wet evergreen lowland and lower
montane forest, known only from scattered collections in Panama (Pennington
et al., 1981).
Botanical description:
"Rami noveili graciies, aureo-tomentosi usque villose, tandem pallide
cinereo-aibi, glabri, interdum lenticellati, non suberosi. Folia pinnata,
usque ad 50 cm ionga, gemmula terminali incremento intermittente;
petioius semiteres; rachis teres vel quadranguiaris, primo aureo-
tomentosa; petioiulus 3-6 mm. Foliola usque ad 8 -juga, late obionga vel
oblanceolata, apice obtuse cuspidate, attenuate vel acuminata, basi
breviter et anguste attenuate, chartacea, 14.5-22.2 [17.4] cm Ionga, 6-9
[7] cm lata; costa supera et nervi secundarii dense pubescentes, lamina
glabra sed crebris punctulis exstantibus conspersa; costa infera nervique
dense et grosse pubescentes, lamina sparse pubescens usque glabra,
nec glanduloso-punctata nec-striata; venatio eucamptodroma, costa
impressa; nervi secundarii 11-14 utroque costae latere, adscendentes,
arcuati, paralleli, intersecundarii nulli tertiarii oblique, spissi, paralleli.
Inflorescentia cauligena, solitaria vel geminate, 5.5-21 cm Ionga, in
racemum vel paniculam gracilem ramulos latérales prope basin
gerentem disposita, dense aureo-puberula; pedicellus iatus, 1 - 2 mm.
Calyx late cyathiformis, 6-7 mm longus, in alabastro clausus, tunc
irregulariter in 3-4 lobes late ovatos, acutos vel obtusos, 2-6 mm longos
53

dehiscens, extus adpresse puberulus. Petala 4, valvata vel paullum
imbricata, 15-18 mm Ionga, 4-5 mm lata, lingulata vel anguste elliptica,
apice acuta vel obtusa, extus dense aureo-sericea, intus glabra. Tubus
stamineus 10-13 mm longus, 3-5 mm Iatus, margine undulatus, glaber;
antherae 10-13,2-2.2 mm longae. Nectarium breviter stipitiforme, 0.25-
1.5 mm longum, apice paullum expansum, glabrum. Ovarium dense
strigosum, 7-9-loculare, loculis 2 ovula superposita continentibus; stylus
strigosus. Capsula (statu immaturo) pyriformis, apice truncate, sensim
and basin attenuate, obscure longitrorsus striata, dense papillose, ca.
4.6 cm Ionga, ca. 3.4 cm lata, 8 -valvata, valvis 2 semina superposita
tenentibus; pericarpium 5-7 mm crassum. Semen non visum"
(Pennington et al., 1981).
Guarea rhopalocarpa Radlkofer;
Synonymous: Guarea tuisiana. There is no previous chemical or biological
studies on this plant. It is a tree of lowland tropical rain forest and montane rain
forest in Costa Rica and Western and Central Panama (Pennington et al.,
1981).
Botanical description:
"Young branches minutely appressed puberulous less frequently
pubescent, soon glabrous, greyish-white. Leaves pinnate with a terminal
bud showing intermittent growth, to 55 cm long; petiole semiterete,
rachis terete or quadrangular, minutely puberulous or less frequently
pubescent at first, soon glabrous; petiolule 7-12 mm long. Leaflets 4-5
pairs, broadly attenuate, often decurrent into petiolule, chartaceous,
12.5-21 [16.4] cm long, 4.8-8.7[6.2] cm broad, upper surface glabrous,
but with numerous minute raised dots, lower surface usually glabrous,
less frequently finely puberulous on midrib and veins, usually glandular-
punctate and -striate; venation eucamptodromous, midrib flat or slightly
54

sunken; secondaries 9-12 on either side of midrib, ascending, usually
arcuate and slightly convergent, rarely straight and parallel;
intersecondaries absent, tertiaries oblique. Flowers unisexual, plants
dioecious; inflorescences mainly cauliflorous and ramiflorous, with a few
from leaf axils, 10-30 cm long, a densely-flowered pendulous spike,
minutely puberulous, flowers ± sessile. Calyx usually patelliform, rarely
short cyathiform, 2-3 mm long, with 3-4 irregular, ovate, acute or obtuse
lobes 0.5-3 mm long, indumentum of minute scattered appressed hairs.
Petals 4-6, valvate, 9.5-14 mm long, 1.5-3.5 mm broad, strap-shaped,
apex acute of obtuse, minutely appressed puberulous on outer surface,
glabrous inside. Staminal tube 8-12.5 mm long, 2-3.5 mm broad, margin
orenulate, glabrous; anthers 8-9,1-1.5 mm long; antherodes similar, not
dehisced, without pollen. Nectary a short stipe expanded at apex to
form a small annulus below ovary, 0.5-1 mm long, stipe glabrous,
expanded portion puberulous. Ovary 4-5(-6)-locular, loculi with 2
superposed ovules, appressed puberulous to pubescent; style
puberulous of short pubescent; pistillode similar, with a longer and more
slender style, containing well-developed, non-functional ovules. Capsule
pyriform to ellipsoid, apex acute, rounded or emarginate, base long
tapering, densely papillose-puberulous, 4-7.5 cm long, 3-4 cm broad, 4-5
valved, valves broadly and shallowly 3-ribbed, ribs sometimes torulose,
valves with 2 superposed seeds; pericarp 6 - 1 0 mm thick. Seed truncate
at base or apex, 1.5-2 cm long, 0.6-1 cm broad, surrounded by a thick
fleshy sarcotesta; seed coat thin, cartilaginous; hilum narrow, extending
length of seed. Embryo with plano-convex, superposed cotyledons;
radicle abaxial, extending to surface"(Pennington et al., 1981).
Ruagea glabra Triana & Ptanchon:
Synonymous: Guarea trianae, G, weberbaueri, Ruagea jelskiana, R.
augusti, R. weberbaueri, R. trisperma, R. floribunda.
55

There is no previous chemical or biological studies on this genus and it is
represented by five species confined largely to montane rain forest and clouds
forest from Guatemala to Panama, and Andean South America as far south as
Peru (Pennington et al., 1981).
Botanical description:
"Young branches usually slender, minutely appressed puberulous when
young soon glabrous, pale greyish-brown, usualiy with a few lenticels.
Bud scales absent. Leaves imparipinnate or paripinnate, with or without
some limited apicai growth, 8-55 cm long; petiole semiterete, rachis
often canaiiculate above, sometimes narrowiy winged, puberulous of
glabrous; petiolule of lateral leaflets 1-7 mm long, petiolule of terminal
leaflet 1 0 - 1 2 mm long. Leaflets opposite or alternate, (5-)8-15(17),
elliptic, oblong of oblanceolate, apex usually short, narrowly attenuate,
lees frequently acute to obtuse rarely rounded, base acute, cuneate or
attenuate, margin rarely revolute, usually chartaceous, 8-20(-32)[14.6]
cm long, 3-9(-16)[5.9] cm broad, upper midrib sometimes puberulous,
lower surface glabrous or with sparse to dense soft short pubescence
on midrib and veins and scattered hairs on lamina,sometimes mixed with
a few red papillae; venation eucamtodromous or rarely brochidodromous;
secondaries (8-)10-14(-18) on either side of midrib, broadiy spreading,
arcuate at tip, parallel or slightly convergent; intersecondaries often
rather iong; tertiaries usuaily obscure. Flowers unisexual, plants
dioecious; inflorescence axillary, 5-30(-60) cm long, usually a slender
panicle with short basal branches less frequently with widely spreading
branches to 1 0 cm long, flowers in smali ciusters or racemose, minuteiy
puberulous or subglabrous; pedicel 0-1 mm long. Calyx patelliform,
sepals 0.75-2 mm long, ovate to orbicular, sparsely puberulous or
glabrous, dilate. Petals 5-7(-8) mm long, 1.5-2.5 mm broad, elliptic,
oblong or spathulate, apex rounded, glabrous. Staminal tube cyathiform
56

or short cylindrical, 2-6 mm long, 1.5-3 mm broad, margin crenulate or
with emarginate lobes, glabrous or sparsely barbate inside; anthers (8 -
10(-11 ), 0.75-1 (-1.1) mm long, sometimes with a short basal appendage,
glabrous; antherodes narrower, without pollen not dehiscing. Nectary a
short stout stipe, sometimes swollen below the ovary, 0.25-1 (-2) mm
long, glabrous. Ovary (2-)3(-4)-locular, loculi with (1-)2 superposed
ovules, glabrous or sparsely pubescent near the base; style stout,
glabrous; style stout, glabrous; style-head discoid; pistilloide similar with
well-developed non-functional ovules. Capsule globose or short
pyriform, smooth, dark brown (when dry) with large pale lenticels,
glabrous, 2-3 cm diam., 3-valved, valves 1-seeded; pericarp 0.5-2 mm
thick. Seed ca. 1.5 cm long, 0.7 cm broad, ± ellipsoid, with a thick,
fleshy, basal sarcotesta; seed coat thin, membraneous. Embryo with
plano-convex, collateral cotyledons; radicle apical, extending to surface"
(Pennington et al., 1981).
2.2 Menispermaceous Plants.
The Menispermaceae comprises 65 genera and approximately 400 species
native to the tropics and subtropics of both hemispheres. They are mostly
lianas, sometimes small trees or shrubs and occasionally perennial herbs and
the family is divided into eight tribes based on characters of the fruits and
seeds (Schultes and Raffauf, 1990). The major alkaloids in Menispermaceae
are the bbiq alkaloids and some have been found to exhibit a variety of
pharmacological activities (Schiff, 1983; Schiff, 1987). Bbiq alkaloids showed
to be more active against P. falciparum than against Entamoeba histolytica an6
KB cell, in vitro (Marshall et al., 1994).
Phaeanthine, a bbiq alkaloid, isolated from the woody part of Triclisia
57

patens was found to be twice as potent against a chloroquine-resistant P.
falcipamm strain (K1) as against a chloroquine-sensitive clone (Ekong et al.,
1991). Tetrandrine, the enantiomer of phaenthine, was more active against
chloroquine-resistant than against a choloroquine-sensitive strains of P.
falcipamm (Ye and Van Dyke, 1989); also, tetrandrine and structurally related
alkaloids have shown to be effective in reversing resistance in chloroquine-
resistant P. falciparum and when used in combination with artemisinin,
tetrandrine is said to provide a long-acting and synergistic activity (Van Dyke,
1991). Similarly, Shiraishi et al. (1987) have shown that some bbiq alkaloids
are more active against a multidrug-resistant cancer line than they are against
a drug-sensitive line.
The species of Abuta are shrubs, small trees or woody lianas natives to
tropical South America and comprises some 35 species (Schultes and Raffauf,
1990). The genus contains alkaloids characteristic of the family and the
chemical components isolated from this group of plants are presented in Table
2.2 (p.59).
Abuta dwyerana Kruk. & Barneby: There is no previous chemical or
biological studies on this plant and there are three Abuta species occurring in
Panama.
2.3 Rubiaceous Plants.
One of the largest of the plant families, the Rubiaceae has 7000 species
distributed in 500 genera. Although, it is cosmopolitan most of the species are
tropical and is a very natural group, but distinction between some of the genera
is frequently difficult and controversial. The family has been divided into three
subfamilies and 29 tribes based of a variety of characters (Schultes and
Raffauf, 1990).
58

Plant species Isolated compounds ReferenceA. bullata Isoquinoline alkaloids: saulatine, dihydro saulatine. Hocquemiller and
Cave, 1984A. grandifolia Isoquinoline alkaloids: grandirubrine Menachery and Cava,
1980A. grisebachii Isoquinoline alkaloids: grisabine Ahmad and Cava,
1977macolide, macoline, peinamine, 7-0-demethyl-peinamine, 7-0-demethyl-N- methyl-peinamine
Galeffi et al., 1977
imelutine, imenine, imerubine, homo-moschatoline, rufescine, nor-rufescine. Cava et al., 1975aA. pahni Isoquinoline alkaloids: (+) coclaurine, (-)daurisoline, 2’-N-nor-daurisoline, (-)
lindoldhamine, 2'-N-methyl-lindoldhamine, dimethyl-lindoldhamine, stepharine, thalifoline
Dute et al., 1987
A. panurensis Isoquinoline alkaloids: panurensine, nor-panurensine Cava et al., 1975bA. racemosa Triterpenes: dammara-20-25-dien-3p-24a diol Hammond et al., 1990A. rufescens Isoquinoline alkaloids:imeluteine, imenine, imerubine, rufescine, nor-rufescine Cava et al., 1975a
homo-moschatoline, splendidine Skiles et al., 1979A. splendida Isoquinoline alkaloids: aromoline, homo-aromoline, krukovine Saa et al., 1976A. velutina Triterpenes: abutasterone Pinheiro et al., 1983
Table 2.2 Chemical Compounds Isolated from Abuta species.
59

This is the largest family of dicotyledons in Panama with about 431 species
in 87 genera (D’Arcy, 1987). The vast majority of Rubiaceae in Panama are
trees or shrubs with the genus Psychotria representing almost one quarter of
the species. Subshrubs or rarely herbs like Borreria, Diodia and Spermacoce
are particularly difficult to identify as to species (Dwyer, 1980).
The family is rich in chemical constituents of potential in medicine and
industrial uses, including iridoids, several type of alkaloids, triterpenes, sterols,
quinones, naphthalene derivatives, polyphenols and tannins. The most
important medicinally compounds of the family including quinine from Cinchona
species, emetine from Cephaëlis species and the widely distributed quinones.
The three major type of alkaloids from the family are all derived from the iridoid
precursor, ioganin, and contain either indole, tetrahydroisoquinoline or quinoline
moieties, however, non iridoid alkaloids are present. The alkaloids of
Rubiaceae have been reviewed (Hemingway and Phiilipson, 1980).
There are some 2 0 0 species of Cephaëlis occurring throughout the tropics,
of which 21 occur in Panama (Dwyer, 1980). The taxonomic position of
Cephaëlis species is unclear. The study of Cephaëlis Swartz and Psychotria
Linnaeus of the Northeastern South America (Steyermark, 1972) and
Venezuela (Steyermark, 1974) demonstrated that Cephaëlis as generally
circumscribed in South America is polyphyletic and represents no more than a
convergent assortment of species and species groups, each of them more
closely related to species of Psychotria than to other species of Cephaëlis
(Taylor et ai., 1991). in Central America, where fewer intermediate taxa grow
than in South America, Cephaëlis has been maintained as a genus (Dwyer,
1980; Standley and Williams, 1975). More recently however, in the preparation
of a monograph on the Rubiaceae in Costa Rica (Taylor, 1991), Cephaëlis
species have been included in Psychotria, subgenus Heteropsychotria. The
genus Psychotria is constituted of two principal subgenera, Psychotria and
Heteropsychotria, and they have been combined mostly because both of them
60

have tiny white flowers and the species of both are numerous and difficult to
separate (Taylor, 1993).
As not all the new names for the species selected for this study have been
published, the Cephaëlis name will be maintained, however, all the
synonymous are showed under each plant description. In addition, and to
further our knowledge of this group of plants, a review of the chemical
compounds present in Psychotria species is presented in this section.
Most treatises of pharmacognosy, ie. Tyler et al. (1988) state that the
Panamanian ipecac is Cephaëlis acuminata Karsten, however, it has been
shown that this name appears to be a nomen nudum, ie. it has never been
validly published from a taxonomic point of view. Then, the correct name for
the commercial source of emetine be C. ipecacuanha (Broth.) Rich. (Hatfield
et al., 1981) therefore, all the scientific reports about C. acuminata should be
considered as C. ipecacuanha.
Some 24 monoterpenoid isoquinoline alkaloids have been isolated from C.
ipecacuanha (Wiegrebe, et al., 1984; Itoh, et al., 1989; Itoh, et al., 1991 ;
Nagakura, et al. 1993). Emetine is used in the treatment of amoebic dysentery
and the extract of the roots as expectorant in cough syrup and in the treatment
of poisonings (Tyler et al., 1988). The simple indole alkaloid gramine has been
isolated from C. stipulacea (Yulianti and Djamal, 1991). None of the other
Cephaëlis species have been investigated for its chemical constituents or
biological activities.
Cephaëlis camponutans Dwyer & Hayden.
The accepted name of this species is Psychotria camponutans (Dwyer &
Hayden) Hammel and occurs in Costa Rica and Panama (Taylor et al, 1991).
Botanical description:
"Erect, glabrous to pu be ru lent, somewhat succulent, suffruticose herbs
61

0.5-1.25 m tall, unbranched; stems quadrate with rounded corners. Leaf
blades oblong to obovoid-oblong, 18-25 cm long, 7-9 cm wide, acute at
apex, acute to attenuate at base, coriaceous, dark green above, paler
and below; secondary veins 9-11 pairs, not looping to interconnect,
indistinct but midrib keeled above, prominulous below, without domatia;
petioles 25-55 mm long; stipules deciduous, connate interpetiolarly,
triangular, succulent, divergent, 4-5 mm long minutely bilobed.
Inflorescences pseudoaxillary, with cymes capitate, green, puberulent,
5-10 mm in diameter, subsessile; involucral bracts green, ovate, 10 mm
long, floral bracts lanceolate, 6 - 8 mm long, acuminate; flowers sessile,
5-merous; calyx limb ca.1 mm long, lobed to base; corolla white,
funnelform, glabrous, tube ca. 3 mm long, lobes 1-1.5 mm long. Fruits
obovoid to ellipsoid, 7-8 mm long, succulent, red; pyrenes planoconvex,
6-7 mm long, 4-5-angled on back, smooth with a median sulcus
ventrally." (Taylor, 1991).
Cephaëlis dichroa (Standley) Standley.
This species is known only from Panama and Evea dichroa has been used
as synonymous (Dwyer, 1980).
Botanical description:
"Glabrous shrubs, to 2 m tall. Leaves elliptic, 8.5-13.0 cm long, to ca. 3.2
cm wide, acuminate at the apex, cuneate at the base, the lateral veins
ca. 1 2 , stiffly papyraceous, concolorous, often drying yellowish, glabrous;
petioles to 3 cm long, wiry, often flexuous; stipules connate, ca. 3.5 mm
long, the body compressed cylindrical, each part with 2 widely spaced,
erect, obtuse lobes about as long as the sheath. Inflorescences
terminal, the peduncle 5-8(-9) cm long, terminated by 3 heads of flowers,
the trio subtended by 2 large obovate bracts, to 2 . 2 cm long, ca. 1 . 8 cm
wide. Fruits subrotund to 0.8 cm wide." (Dwyer, 1980).
62

Cephaëlis dimorphandrioides Dwyer.This species is known only from Panama and is distinguished from all other
Cephaëlis by the cylindrical spikelike inflorescence which bears a resemblance
to the inflorescence of the legume genus Dimorphandra (Dwyer, 1980).
Botanical description:
"Shrubs \o 1.4 m tall. Leaves elliptic oblong, often falcate oblong, 17-30
cm long, 6-14 cm wide, deltoid or obtuse toward the apex, acute or
abruptly acute toward the base, the costa slender above, prominent
beneath, the lateral veins 13-17, arcuate, often with one smaller and
shorter vein between 2 lateral veins, the intervenal areas spreading
reticulate, rigidly chartaceous, discolours, glabrate above, minutely
puberulent beneath; petioles lignose, to 4.5 cm long, twisted, puberulent;
stipules (only one pair seen at tip of twig) oblong rotund, to 0.7 mm long,
deeply 2 -lobed, coriaceous, the lobes rounded, farinose, glabrous within.
/n//orescencesterminal, sessile, cylindrical, 5-12 cm long, to 4 cm wide,
obtuse at the apex, black when dry, the flowers more than 1 0 0 . Flowers
perhaps sessile, the hypanthium short, swollen, puberulent, the calycine
cup short, ca. 5 mm long, the teeth subulate, to 6.5 mm long, unequal,
acute, puberulent outside, glabrous inside, carnose, with 3 longitudinal
veins, with 2-4 oblong glands, to 0.3 mm long, at the sinuses; corolla red
outside, the tube narrowly cylindrical, 10-14 mm long, puberulent
outside, glabrous within, subcarnose, the lobes 5, yellow within, lance
oblong, to 3 mm long, carnose, puberulent outside; stamens 5 , the
anthers narrow oblong, 2,8-3.0 long, minutely apiculate, slender,
attached below the middle of the tube; ovarian disc 4 lobed, to 0.8 mm
long, the style linear, 9-14 mm long, the stigmas 2, reflexed, linear
oblong, to 0 . 8 mm long. Fruits (immature) oblong, to 5 mm long, to 0.3
wide, truncate, each pyrene smooth, black, 1 0 -ribbed, the hairs weak,
white, arachnoid." (Dwyer, 1980)
63

Cephaëlis glomerulata J.Donnell Smith.The proper name of this species is Psychotria glomerulata (J. Donnell Smith)
Steyermark and occurs from Guatemala to Panama, in wet forests (Taylor,
1991).
Botanical description:
"Erect glabrous shrubs 0.5-1 (-2 ) m tall, much-branched; stems quadrate.
Lea/ blades narrowly elliptic to oblong, (5-)10-13 cm long, (15-)25-55
mm wide, acuminate at apex, attenuate at base, chartaceous, green on
both surfaces; secondary veins 12-16(-20) pairs, looping to interconnect
near margins, prominulous above and below, without domatia; petioles
5-15 mm long; stipules persistent, membranaceous, appressed, connate
interpetiolarly and intrapetiolarly, truncate, 1.5-3 mm long, with two
groups of 6-12 caducous awns 0.5-1 mm long. Inflorescences terminal,
with cymes capitate, ovoid, pale green to purple, 1.5-2.5 cm long,
glabrous; peduncles 0-15 mm long; involucral bracts broadly ovate, 10-
15 mm long, rounded, floral bracts ovate, 8-10 mm long; flowers
distylous, 5-merous; calyx limb 0.5-1 mm long, partially lobed; corolla
white, funnelform, glabrous, tube 10-15 mm long, lobes 2.5-3 mm, long.
Fruits ovoid to cylindrical, 10-15 mm long, ca. 5-6 mm in diameter,
succulent, bright blue; pyrenes planoconvex, 3-5 mm long, 5-ribbed
dorsally, smooth with a median sulcus ventrally". (Taylor, 1991).
Table 2.3 (p.65) shows the chemical compounds isolated from the genus
Psychotria, however, some early reports (Wehmer, 1931; Willaman and
Schubert, 1961) of the presence of emetine and related alkaloids in this genus
should be treated with prudence.
64

Plant Species Compounds Isolated ReferencesP. adenophylla Triterpenes: a-amyrin, baurenol, baurenol acetate, betuiin, betulinic
acid, friedenn, ursolic acid.Steroids: 0-sitosterol
Dan and Dan, 1986
P. bacteriophila Steroids: 0-sitosterol (from callus tissues) Dohnal et al., 1989P. duoarrei Inorganics: nickel Kersten et al., 1980P. emetica Alkaloids: cephaeline, psychotrine Wehmer, 1931P. forsteriana Polyindolenines alkaloids: chimonanthine, hodgkinsine, psychotridine,
iso-psychotridine, quadrigemine A, quadrigemine BAdjibade et al., 1989
meso-chimonanthine Adiibade et al., 1992iso-psychotridine 0 Roth et al., 1985
Quinoline alkaloids: calycanthine Adjibade et al., 1991(-) calycanthine, iso-calycanthine Adiibade et al., 1992
P. granadensis Isoquinoline alkaloids: (-) emetine Willaman and Schubert, 1961P. manillensis Monoterpenes: asperuloside Inouye et al., 1988P. oleoides Polyindolenine alkaloids: hodgkinsine, psychotridine, iso-psychotridine
A, iso-psychotridine B, quadrigemine CLibot et al., 1987
psycholeine Gueritte-Voegelein et al., 1992P. rubra Monoterpenes: asperuloside Inouye et al., 1988
Naphtoquinones: psychorubin sesquiterpenes: helenalin
Hayashi et al., 1987
Steroids: 0-sitosterol Hui and Yee, 1967P. serpens Monoterpenoids: asperuloside Inouye et al., 1988
Steroids: 0-sitosterol, stigmasterol Hui and Yee, 1967Triterpenes: ursolic acid Lee et al., 1988
P. tomentosa Isoquinoline alkaloids: (-) emetine Wehmer, 1931
Table 2.3 Chemical Compounds from Psychotria Species.
65

Lasianthus panamensis Dwyer.
This plant belongs to the tribe Psychotriae and is the only species of this
genus in Panama, formerly named Dressleriopsis panamensis (D’Arcy, 1987).
Botanical description:
"Subshrubs to 1 m tall, often with a pair of slender, rigid, opposite
branches. Leaves oblong, to 19 cm long, to 9 cm wide, deltoid to
subobtuse, short acuminate, basally cordate, truncate, or auriculate, the
lateral veins ca. 15, arcuate, forming a conspicuous submarginal vein,
the intervenal areas, spreading reticulate, rigidly chartaceous,
discolorous, pilose petioles to 6 cm long; stipules triangular, free, ca. 7
mm long, ca. 6 mm wide, carnose, glabrous inside, diffusely hirsute
outside. Inflorescences axillary, the flowers congested into sessile
globose heads 2 cm in diam.; bracteoles subulate, to 2 mm long.
Flowers subsessile; hypanthium subrotund, to 2 mm long hirsute, the
hairs septate, lobes 4, to 4 mm, ca. 1. 6 mm wide, basally attenuate
glandular inside at the base; corolla white, cylindrical, ca. 3.5 cm long,
glabrous within, the lobes 5, often horizontally expanded, oblong, ca. 4
mm long, villose, attenuate; upwards anthers 5, slightly exserted,
subsessile, oblong, ca. 1 mm long, glabrous; disc cushion shaped, ca.
0.5 mm wide, the locules 8 , a single linear oblong axile ovule filling each
locule, the style narrowly, to 5 mm long, the stigmas 8 , radiate. Fruits
(?immature) berrylike, globose, to 1 . 1 cm in diam., not ribbed,
compressed at the apex, the internal pulp with abundant raphides, the
pyrenes 8 , pear shaped or somewhat hemispherical, ca. 3 mm long,
hard, orange, with an oblong scar on the concave side". (Dwyer, 1980).
Chemical compounds isolated fr tm species of Lasianthus: L. curtisii,
L. cyanocarpus, L. fordii, L. obliquinervis and L. plagiophyllus contain
asperuloside (monoterpenoid) (Inouye et al., 1988). Betuiin (triterpene),
damacanthal (quinone), (3-sitosterol and stigmasterol (steroid) have been
66

isolated from L chinensis (Hui et a!., 1967). The flavonoids eriodictyol and
eriodictyol-7-O-p-D-glucoside and the coumarin scopeletin have been isolated
from L japonica (Ishikura et al., 1979).
2.4 Simaroubaceous Plants.
This family of tropical and subtropical trees has about 30 genera and 2 0 0
species (Schultes and Raffauf, 1990). Antimalarial activity has been reported
for quassinoids (Trager and Polonsky, 1981; Guru et al., 1983; O’Neill et al.,
1987; Chan et al., 1986; Bray et al., 1987; O’Neill et al., 1988) and for indole
alkaloids (Kardono et al., 1991 ; Pavanand et al., 1988) isolated from
Simaroubaceous species, but it is the former group of compounds which is the
more potent.
The in vitro testing of 40 individual quassinoids against Plasmodium
falciparum (K1 ), a chloroquine-resistant strain, has been reported (O’Neill et al.,
1986). Also, quassinoids have been shown to have in vitro activity against
Entamoeba histolytica (Gillin et al., 1982; Wright et al., 1988) and Leishmania
donovani (Robert-Gero et al., 1985).
There are some 60 species of Picramnia in Central and South America and
the West Indies (Schultes and Raffauf, 1990), of which six occur in Panama
(Porter, 1973). Quassinoids have not been found, so far, in species of
Picramnia. Table 2.4 (p.6 8 ) shows the chemical compounds isolated from this
genus.
Biological activity for extracts of Picrmania species: The chloroformic
extract of the branches of P. antidesma was active against Plasmodium
gallinaceum in vivo, but the aqueous extract was inactive (Spencer et al.,
1947), and the methanolic extract of the leaves were inactive in plant
germination inhibition (Dominguez and Alcorn, 1985). The chloroformic and
67

aqueous extracts of P. carpinterae (P. teapensis), P. locupes, P. longissima, P.
pentandra, P. sellowii and P. xalapensis were inactive against P. gallinaceum,
in vivo (Spencer et a!., 1947). An aqueous extract of the leaves of P.
macrostachys was active against Staphylococcus aureus and Streptococcus sp.
(Arana and Juica, 1986). The ethanolic extract of the leaves of P. pentandra
showed spasmogenic activity in isolated ileum of guinea pig and vasodilator
activity in isolated hind quarter of rat (Feng et al., 1962).
Plant species Compounds Isolated References
P. antidesma Steroids: p-sitosterol Dominguez and
Alcorn, 1985.
P. macrostachys Quinones: chrysophanic acid,
physcion
Arana and JuIca,
1986
P. parvifolia Quinones: aloe-emodin,
chrysophanic acid,
emodin, physcion, rhein
Popinigis et al,
1980
P. pentandra Triterpenes: 3-epi-betulinic acid Herz et al., 1972
P. sellowii Quinones: chrysophanic acid,
emodin, physcion.
Triterpene: betulinic acid, 13-epi-
betulinic acid
Leon, 1975
Table 2.4 Compounds Isolated from Species of Picramnia.
Picramnia antidesma subsp. fessonia (DC) W. Thomas:
68

Synonymous; P. fessonia, P. antidesma var. pubescens, P. andicoia, P.
bonplandiana, P. tetramera, P. quaternaria, P. seemannaina, P.
brachybotryosa, P. pistaciaefolia, P. locuples, P. velutina, P. aiienii.
This species is the most common and variable taxon of Picramnia in Mexico
and Central America. This variability, particularly in flower merosity, leaflet
number and shape, and features of the indûment, is responsible for the long list
of synonymous (Thomas, 1988). The very bitter, liquorice-flavoured bark and
leaves have been much used in Central America and the West Indies as
remedies for malaria and stomach and intestinal ailments (Standley and
Steyermark, 1958). The bark was formerly exported to Europe for use in
treating erysipelas and venereal diseases (Morton, 1981).
Botanical description:
"Shrubs or small trees, 1-7 m high; branch lets yellowish appressed-
tomentulose, becoming glabrate. Leaves alternate, pinnate, 18-24,5 cm
long; petiole and the rachis yellowish appressed-tomentulose, becoming
almost glabrate; leaflets 1 2 -2 0 , mainly and even number, alternate to
lowermost subopposite, ovate to elliptic, narrowly acuminate apically,
rounded and markedly inequilateral basally, the margins entire, revolute,
chartaceous, yellowish appressed-tomentulose above and beneath on
the main vein and marginally, the blade sparingly pubescent, 22-75 mm
long and 8-23 mm wide, the terminal leaflets largest, the lowermost
smallest and reflexed, the petiolules yellowish appressed-tomentulose,
2 - 3 mm long. Staminate racemes subterminal, becoming axillary,
aggregated to solitary, simple or rarely once-branched, densely-flowered,
yellowish appressed-tomentulose, to 38 cm long. Staminate flowers, 3 -
merous, 1 -several aggregated in clusters, white or greenish, the
spreading, yellowish appressed-tomentulose, to 3 mm long, sepals 3,
ovate, apiculate, appressed-pubescent, ca. 15 mm long and 1 mm wide,
petals 3, narrowly elliptic, apiculate, glabrous to rarely sparingly
69

pubescent, longer than the sepals, ca. 1.5 mm long; stamens 3,
exserted, the filaments filiform, glabrous, ca. 2.5 mm long, ovary absent.
Carpellated racemes solitary or in pairs, yellowish appressed-
tomentulose, to 28 cm long. Carpellate flowers 3-merous, the pedicels
appressed-tomentulose, articulated basally, 1-1.5 cm long in fruit; sepals
3, broadly triangular, appressed-pubescent, persistent and spreading in
fruit, ca. 2 mm long and 1 mm wide; style bilobed, the lobes recurved
and persisting in fruit. Berries red or orange, ellipsoid to obovoid,
appressed-pubescent to glabrate, to 1.5 cm long; seeds 1". (Porter,
1973).
Picramnia teapensis TulSynonymous: P. carpinterae, P. antidesma var. carpinterae.
This species is found in cool moist forest from Mexico to Venezuela and it
is most common in Costa Rica and Panama (Thomas, 1988). In Costa Rica,
a decoction of the bitter bark of the trunk and roots is taken as a tonic to
stimulate the appetite and digestion, and as a febrifuge (Nunez-Melendez,
1975).
Botanical description:
"Sma// trees, 5-12 m high, the crown flat; branch lets yellowish-villous.
Leaves alternate, pinnate, to 20 cm long; petiole and the rachis
yellowish-villous; leaflets 9-11, alternate to subopposite, obtuse or acute
and acuminate apically, more or less inequilateral, the uppermost and
middle leaflets elliptic to ovate and cuneate basally, the lowermost ovate
and cordate basally, paler beneath, the margins repand, revolute,
subcoriaceous, villous marginally and beneath, especially on the veins,
to 11 cm long and 5.5 cm wide, the lowermost pair much smaller than
the others, the petiolules villous, 1 - 2 mm long. Staminate racemes
subterminal, becoming axillary, solitary, simple, yellowish-villous, 1 2 - 2 2
70

cm long. Staminate flowers 3-merous, numerous, 1 -several aggregated
in clusters, the pedicels glabrous, 1-3 mm long, sepals 3, obovate,
sparingly pubescent, 1.5 mm long and 1 mm wide, petals 3, narrowly
obovate, ca. 1 mm long; disc 3-4 lobed; stamens 3(-4), exserted, the
filaments subulate, glabrous, ca. 2.5 mm long, ovary rudimentary,
conical, villous apically, the style and stigma absent. Carpellate racemes
solitary, yellowish-villous, 23-27 cm long in flower, to 30 cm long in fruit.
Carpellate flowers 3-merous, white, the pedicels villous to glabrate, 2-4
mm long in flower, 4-12 mm long in fruit, articulated basally; sepals 3,
broadly triangular, densely villous, persistent and spreading in fruit, ca.
1.5 mm long and 1.5 mm wide; petals 3, narrowly obovate, apiculate,
spreading, ca. 2 mm long; staminodes 3, opposite the petals; disc 3-
lobed, glabrous; ovary ovoid, densely pubescent, 1 - 2 mm high, 2 -
loculed, the ovules 2 per locule, the style bilobed, the lobes recurved
and persisting in fruit. Berries red, ellipsoid, sparingly pubescent; seeds
1-2". (Porter, 1973).
71

SECTION 3. TESTING FOR BIOLOGICAL ACTIVITY.

3.1 Materials and Methods
3.1.1 New microwell brine shrimp assay
Brine shrimp (Artemia salina) eggs obtained locally (Interpet Ltd. Dorking,
England) were hatched in artificial sea water prepared from sea salt (Sigma
Chemical Co., U.K.) 40 g/l of distilled water supplemented with 6 mg/l dried
yeast and oxygenated with an aquarium pump. After 48 hours incubation in a
warm room (22-29°C) nauplii were collected with a pasteur pipette after
attracting the organisms to one side of the vessel with a light source. Nauplii
were separated from the eggs by pipetting them 2-3 times in small beakers
containing fresh sea water.
Samples for testing (0 . 6 mg) were made up to 1 mg/ml in artificial sea water
except for water insoluble compounds which were dissolved in 50 pi DMSO
prior to adding sea water.
Serial dilutions were made in 96-well microplates (ICN Flow, U.K.) in
triplicate in 100 pi sea water. Control wells with DMSO were included in each
experiment. The nauplii were place in a small beaker and air was pumped to
oxygenate and keep a homogenous suspension of larvae, 1 0 0 pi containing
10-15 organisms was added to each well and the covered plate incubated at
22-29°C for 24 hours. Plates were then examined under a binocular
microscope (x12.5) and the number of dead (non-motile) nauplii in each well
were counted. 1 0 0 pi methanol was then added to each well to kill all the
shrimps and after 15 minutes the total numbers of organisms in each well were
counted. LCgo values were then calculated by Probit analysis (Finney, 1971).
Test compounds: Bruceines A, C and D and brusatol were isolated as
previously described (O’Neill et al., 1987). Villastonine was isolated from
Alstonia angustifolia as previously reported (Wright et al., 1992).
Homoharringtonine, isoharringtonine and cephalotaxine were kindly supplied by
Dr. Powell, United States Department of Agriculture, Illinois, and quassin was
73

a gift from Bush, Boake and Allen Ltd. U.K., thymol was obtained from May and
Baker Ltd, U.K., actinomycin D, from Aldrich Chemical Co. U.K., and atropine
sulphate, chloramphenicol, quinidine sulphate and quinine hydrochloride were
supplied by BDH, U.K. All other compounds were obtained from Sigma
Chemical Co. U.K.
3.1.2 Brine shrimp assay
Brine shrimp assay was carried out in test tube as previously reported
(Meyer et al., 1982). The eggs of Artemia salina were hatched as described
in Section 3.1.1 and samples for testing were made up to lOrng/m/ in artificial
sea water or in methanol for water insoluble compounds. Appropriate amounts
of solution were transferred into test tubes to make a final concentration of
1000, 100 and 10 pg/ml of compound for a total volume of 5 ml. Three
replicates were prepared for each concentration. When methanol was used to
dissolve the samples it was evaporated under a stream of nitrogen and kept
under vacuum overnight. Control vials were prepared using only the solvent.
Ten nauplii were transferred to each sample test tube and artificial sea
water was added to make 5 ml. The mixtures were incubated in a warm room
(22-29 °C) and after 24 hrs survivors were counted and the percent of deaths
at each concentration and control were determined. The LCgg values were
calculated by Probit analysis (Finney, 1971).
3.1.3 KB cells assay
The microdilution technique for KB cells assay was carried out as
previously described by Anderson et al. (1991b). Human nasopharyngeal KB
cells were maintained in Eagles Minimum Essential medium with Earle's salts
and sodium bicarbonate (0.85 g/l), supplemented with 1% foetal bovine serum,
L-glutamine (2 mM) (ICN Flow, UK.), penicillin (50 units/ml) and streptomycin
(50 pg/ml) (ICN Flow, UK.).
74

When the monolayer became fully confluent the cells were subcultured
using 0.25 % trypsin solution (ICN Flow, UK.). The detached cells were
transferred to sterile centrifuge tubes (10 ml) and spun at 400 rpm for
approximately 15 seconds. After twice washing with complete medium, the
cells were resuspended in 1 ml of complete medium and mixed thoroughly to
break down any cell aggregates. The cells were counted using a
haemocytometer and the appropriate number of cells added to fresh culture
vessels containing complete medium to give a final concentration of 1 0 ®
cells/ml.
A 50 pi aliquot of cells suspension (1x10® cells/ml) was mixed with 50 pi
aliquot of serial dilution of compounds, in triplicate each time. In each
experiment two controls were used, one without drug as negative control and
emetine HCI (Sigma Chem. Co. UK.) as positive control. Incubation for 48 hrs
at 37 °C was done in atmosphere of 5 % of COg in air. After incubation, the
cells were fixed by adding 6 6 pi of 25 % trichloroacetic acid (Sigma Chem. Co.
UK.) at 4 °C to each well and refrigerated at the same temperature for one
hour. The plates were then rinsed with water 5 times and left to dry. 1 0 0 pi
of 1 % aqueous eosin B stain (Sigma Chem. Co., UK.) was added to each well,
after an hour the plate was washed 5 times with 1 % acetic acid.
The plates were dried overnight before 200 pi of 5 mM sodium hydroxide
solution was added to each well and kept for 2 0 minutes to release the dye.
The optical density of the solution in each well was determined at 490 nm by
a microplate reader, model MR-700 (Dynatech Labs Inc. UK.). The percentage
inhibition of KB cell growth was calculated by comparison of the absorbance of
the control and test samples. The ICgo was calculated by a linear regression
of the percentage of inhibition against the logarithms of the concentration of
sample being tested, using a computer program (Minitab, statistical software,
PC version, release 8,1991. Quickset Inc. Rosemont, PA, USA).
75

3.1.4 Plasmodium falciparum assay.
This was carried out as described previously (Ekong et a!., 1990) by Drs.
D.C. Warhurst, G.C. Kirby, Department of Medical Parasitology, the London
School of Hygiene and Tropical Medicine, and Miss C. Lang'at. Department of
Pharmacognosy, The School of Pharmacy, University of London.
Cultures of P. falciparum (chloroquine-resistant strain K-1 ) were maintained
in human erythrocytes, suspended in RPMI 1640 medium supplemented with
D-glucose, NaHCOg, TES and human serum. Two fold dilutions of the sample
were prepared in 96-microwell microtitre plates and 50 pi of a suspension of
infected human red blood cells (1% parasitaemia) was added to each well. The
mixture was incubated in a 3% Og, 4% COg and 93% N atmosphere at 37 °C
for 24 hrs before adding 5 pi of [^H]-hypoxanthine (40 pCi/ml) and continuing
incubation for a further 18-24 hrs. Chloroquine diphosphate was tested
simultaneously to monitor the sensitivity of the parasite strain. Two series of
controls were performed: one with infected red blood cells in the absence of
drug and the other with uninfected red blood cells.
3.2 Results and Discussion
3.2.1 Evaluation of brine shrimp assay
The method described above (3.1.1) provides a simple and inexpensive
screening test for cytotoxic compounds. It has the advantages of requiring only
small amounts (0.6 mg) of compounds, in contrast with 50 to 20 mg in the test
tube method (3.1.2), and the employment of microplate technology facilitates
the testing of large number of samples and dilutions. The use of methanol to
kill live shrimps at the end of the experiment enables rapid counting of the total
numbers of shrimps.
The results obtained with a number of pharmacologically active compounds,
some of which are cytotoxic, are comparable with results reported using a
76

previous method in which test tubes were used (Meyer et a!., 1982); with the
exception of thymol, LC5 0 values determined by the two methods are similar.
However, when thymol was tested in our laboratory using the test tube method
a LC5 0 value of 46.7 ± 12.0 pM was obtained which is in close agreement with
that found using the new microplate test.
The test was predictive of cytotoxicity for all the compounds listed in Table
3 . 1 (p.78) except for cyclophosphamide which is known to require metabolic
activation in the liver for activity in man. Since >4. salina does not posses the
enzymes required for this process, cyclophosphamide would not be expected
to be toxic to this organism.
Table 3.2 (p.79) shows the LC5 0 values determined for a number of
biologically active compounds which have various modes of action together with
their reported ED5 0 values against KB cells (human nasopharyngeal carcinoma).
Of the six compounds active against KB cells, five were also toxic to nauplii.
However, there was little correlation observed in the degree of toxicities seen
between the two methods. Emetine was more toxic to KB cells than
villastonine but against the brine shrimp the converse was found to be the
case. Similarly, although podophyllotoxin and actinomycin D were equitoxic to
KB cells, the brine shrimp was >50,000 times more sensitive to the latter than
to the former. One cytotoxic compound, 6 -mercaptopurine was not toxic to A.
salina and this is explicable as its purine metabolism is markedly different from
that in mammalian cells (Van Debos and Finamore, 1974) and it is possible that
the intracellular activation which occurs in man may not take place in A. salina.
Quassinoids are constituents of the Simaroubaceae and have potent in vitro
activity against P. falciparum (O’Neill et al., 1987). These compounds inhibit
protein synthesis in both mammalian cells and malaria parasites (Kirby et al.,
1989) and, as shown in Table 3.3 (p.81) were found to be toxic to the brine
shrimp. Quassin which does not possess cytotoxic or antiplasmodial activity
was, as expected, devoid of toxicity. These results suggest that for this group
77

DRUGS MICRO-TECHNIQUE
LCso(pM) ±sd
REPORTED VALUES
pM
ANISOMICIN 35.2 ±14.2 (2 )* -
ATROPINE SULPHATE >1 0 0 0 . (3) 987.3"
BERBERINE CHLORIDE 25.2 ± 2.9 (3) 60.5"
CAFFEINE >1 0 0 0 . (3) > 1 0 0 0 ."
CEPHALOTAXINE 549.9 ± 83.7 (3) -
CYCLOPHOSPHAMIDE >1 0 0 0 . (3) -
EPHEDRINE >1 0 0 0 . (3) > 1 0 0 0 ."
HOMOHARRINGTONINE 7.4 ± 4.7 (3) 7.5°
ISOHARRINGTONINE 13.9 ±6.2 (2 ) -
NICOTINE >1 0 0 0 . (2 ) >1 0 0 0 .°
PODOPHYLLOTOXIN 15.0 ±2.1 (2 ) 5.8"
QUINIDINE SULPHATE 518.8 ± 126.9 (3) 287.8"
THYMOL 56.3 ± 10.6 (3) >1000."; 46.7 ±
1 2 .0 "
^Indicates number of experiments performed in triplicate, using the method
described in section 3 .1 . 1
‘'Data obtained from Meyer et a!.,(1982)
®Data obtained from Anderson et a!., (1991a)
the previously reported data were transformed to pM for comparison
purpose only.
Value obtained in our laboratory using the test tube method (3.1.2).
Table 3.1 Comparison of the Values Obtained in the Brine Shrimp Micro-
Technique with Previously Reported Data.
78
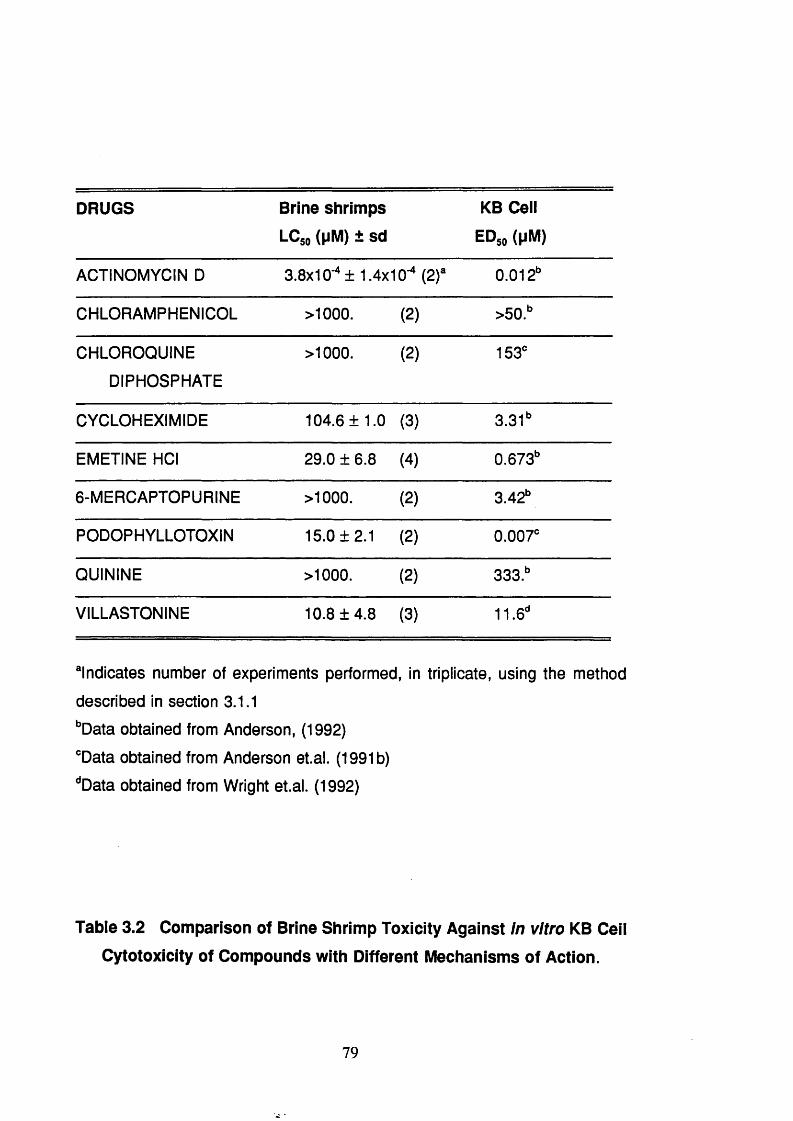
DRUGS Brine shrimps
LCso (pM) ± sd
KB Ceil
EDso (pM)
ACTINOMYCIN D 3.8x10-^ ± 1.4x10- (2 )“ 0 .0 1 2 "
CHLORAMPHENICOL >1 0 0 0 . (2 ) >50."
CHLOROQUINE
DIPHOSPHATE
>1 0 0 0 . (2 ) 153*
CYCLOHEXIMIDE 104.6 ± 1.0 (3) 3.31"
EMETINE HCI 29.0 ± 6 . 8 (4) 0.673"
6 -MERCAPTOPURINE >1 0 0 0 . (2 ) 3.42"
PODOPHYLLOTOXIN 15.0 ±2.1 (2 ) 0.007*
QUININE >1 0 0 0 . (2 ) 333."
VILLASTONINE 10.8 ±4.8 (3) 1 1 .6 "
“Indicates number of experiments performed, in triplicate, using the method
described in section 3.1.1
‘’Data obtained from Anderson, (1992)
"Data obtained from Anderson et.al. (1991b)
‘'Data obtained from Wright et.al. (1992)
Table 3.2 Comparison of Brine Shrimp Toxicity Against in vitro KB Ceil
Cytotoxicity of Compounds with Different Mechanisms of Action.
79

of compounds the brine shrimp test is suitable for the screening of plant
extracts and fractionation of cytotoxic quassinoids, thus obviating the need for
the difficult and expensive antiplasmodial test at every stage in the isolation
procedure. Similarly, the brine shrimp test has been used as a convenient
assay for the detection of antifilarial activity of avermectin analogs (Blizzard et
al., 1989).
This approach may be useful for other types of biologically active
compounds where the brine shrimp responds similarly to the corresponding
mammalian systems. For example the DNA-dependent RNA polymerases of
A. salina have been shown to be similar to the mammalian type (Birndorf et al.,
1975) and the organism has a ouabaine-sensitive Na and dependent
ATPase (Ewing et al., 1974), so that compounds or extracts acting on these
systems would be expected to be detected in this assay. Although the assay
did not detect those compounds which require metabolic activation in man, it
may be conveniently used where, as in the case of the quassinoids, the brine
shrimp is a reliable detector of biological activity.
3.2.3 Activity of the plant crude extracts against brine shrimp and KB
ceils
Tables 3.4 - 3.14 (p.82-87) show the activity of 135 extracts and fractions
obtained from the eleven Panamanian medicinal plants selected for this study.
The value represents the average of two experiments in triplicate. Section
4.1.4 shows the extraction and fractionation procedures. Table 3.15 (p.8 8 )
shows the activity against KB cells of extracts selected on the basis of
previously activity against brine shrimp.
80

QUASSiNOiD BRINE SHRIMP
LCjo (pM) ± sd
KB CELL
EDso (pM)»
P. falciparum
iCso MM
BRUCEINE A 0.887 ± 0.24 (2)“ 0.188 0 .0 2 1 "
BRUCEINE C 0.798 ±0.17 (2) 0.037 0.009"
BRUCEINE D 4.47 ±0.19 (2) 2.82 0.037"
BRUSATOL 1.69 ±0.137 (2) 0.196 0.006"
QUASSIN > 1 0 0 0 . >50. 247"
“Indicates number of tests performed in triplicate, using method described in
section 3.1.1
Data obtained from Anderson et.al.(1989)
"O'Neill et.al.(1987).
''Data obtained from Anderson (1992)
Table 3.3 Activities of quassinoids against brine shrimps, KB ceil and
P. falciparum in vitro.
81

PLANT PART (Code N°)
EXTRACT BRINE SHRIMP LC5 0 pg/ml
MeOH 93.6
CHCI3 89.3
BranchBuOH 395.8
HgO-FRAC. > 1 0 0 0 .(3a) H2 O-EXT. > 1 0 0 0 .
MeOH 154.3
CHCI3 141.5
LeavesBuOH >1 0 0 0 .
H.O-FRAC. > 1 0 0 0 .(3b) H 2 O-EXT. > 1 0 0 0 .
PLANT PART EXTRACT BRINE SHRIMP(Code N°) LC5 0 pg/ml
MeOH 22.2
CHCI3 23.8
BuOH >1000.Branches HgO-FRAC. >1000.
(la) HgO-EXT. >1000.MeOH 5.8CHCI3 8.2
Wood BuOH >1000.H2O-FRAC. >1000.
(1b) H2O-EXT. >1000.MeOH N/TCHCI3 39.6
Leaves BuOH >1000.HgO-FRAC. >1000.
(1c) HgO-EXT. >1000.
Table 3.4 Activity Against Brine Shrimp of the
Extracts from Guarea macropetalaTabie 3.5 Activity Against Brine Shrimp of the
Extracts from Guarea rhopalocarpa
82

PLANT PART (code N*)
EXTRACT BRINE SHRIMP LC5 0 pg/mi
MeOH 324.4
CHCI3 197.1BuOH >1000.
Leaves H2O-FRAC. >1000.
(2 a) HgO-EXT. >1000.
MeOH 228.8CHCI3 44.9BuOH >1000.
Stem-bark H 2 O-FRAC. >1000.
(2 b) H.O-EXT. >1000.
PLANT PART (Code N°)
EXTRACT BRINE SHRIMP LC« pg/mi
MeOH 313.9CHOU 411.5
Leaves BuOH 486.6(4a) H9 O-FRAC. > 1 0 0 0 .
H,0-EXT. > 1 0 0 0 .MeOH > 1 0 0 0 .
CHOU 911.Stems BuOH > 1 0 0 0 .(4b) H.O-FRAC. > 1 0 0 0 .
H.O-EXT. > 1 0 0 0 .
MeOH 755.CHOU 263.
Roots BuOH > 1 0 0 0 .(4c) H,0-FRAC. > 1 0 0 0 .
HgO-EXT. > 1 0 0 0 .
Table 3.6 Activity Against Brine Shrimp of the
Extracts from Ruagea glabraTabie 3.7 Activity Against Brine Shrimp, of theExtracts from Abuta dwyerana
83

PLANT PART (Code N°)
EXTRACT BRINE SHRIMP LC5 0 pg/mi
MeOH > 1 0 0 0 .
CHCI3 579.Aerial BuOH > 1 0 0 0 .parts
H2 O-FRAC. > 1 0 0 0 .(1 0 a) H2 O-EXT. > 1 0 0 0 .
MeOH 275.
CHCig 62.1Roots BuOH > 1 0 0 0 .
(1 0 b) H2O-FRAC. > 1 0 0 0 .
H.O-EXT. > 1 0 0 0 .
PLANT PART (Code N°)
EXTRACT BRINE SHRIMP LC5 0 pg/mi
MeOH >1 0 0 0 .
CHCI3 >1 0 0 0 .Aerial BuOH >1 0 0 0 .parts
H2O-FRAC. > 1 0 0 0 .(8a) H2O-EXT. > 1 0 0 0 .
MeOH >1 0 0 0 .
CHCI3 >1 0 0 0 .
BuOH > 1 0 0 0 .Roots
H2O-FRAC. > 1 0 0 0 .(8b) H2O-EXT. > 1 0 0 0 .
Table 3.8 Activity Against Brine Shrimp of theExtracts from Cephaelis camponutans
Tabie 3.9 Activity Against Brine Shrimp of theExtracts from Cephaëlis dichroa
84

PLANT PART (Code N°)
EXTRACT BRINE SHRIMP LC^ pg/ml
MeOH > 1 0 0 0 .
CHCI3 605.
Leaves BuOH 514.
(9a) H2O-FRAC. > 1 0 0 0 .
HgO-EXT. > 1 0 0 0 .
MeOH > 1 0 0 0 .CHCI3 > 1 0 0 0 .BuOH >1000.
Stems H 2 O-FRAC. >1000.
(9b) H 2 O -E X T. >1000.
MeOH > 1 0 0 0 .CHOU 876.
Roots BuOH > 1 0 0 0 .H,0-FRAC. > 1 0 0 0 .
(9c) HpO-EXT. > 1 0 0 0 .
PLANT PART (Code N®)
EXTRACT BRINE SHRIMP LC5 0 pg/ml
MeOH >1 0 0 0 .
CHCI3 > 1 0 0 0 .
BuOH > 1 0 0 0 .Aerial parts
H2 O-FRAC. > 1 0 0 0 .(1 1 a) H2 O-EXT. > 1 0 0 0 .
MeOH > 1 0 0 0 .
CHCI3 772.8
BuOH >1000.Roots
H 2 O-FRAC. >1000.
(1 1 b) H 2 O -E X T. >1000.
Table 3.10 Activity Against Brine Shrimp of
the Extracts from Cephaëlis dimorphandrioidesTable 3.11 Activity Against Brine Shrimp of
the Extracts from Cephaëlis glomerulata
85

PLANT PART {Code N°)
EXTRACT BRINE SHRIMP LC5 0 pg/mi
MeOH >1000.
CHCI3 >1000.
Aerial BuOH >1000.parts
HgO-FRAC. >1000.(7a) H2O-EXT. >1000.
MeOH >1000.
CHCI3 >1000.
BuOH >1000.Roots
H 2 O-FRAC. >1000.(7b) H 2 O -E X T . >1000.
PLANT PART (Code N®)
EXTRACT BRINE SHRIMP LC5 0 pg/mi
MeOH 511.6
CHCI3 > 1 0 0 0 .
BuOH 485.Leaves
HgO-FRAC. > 1 0 0 0
(5a) H2O-EXT. > 1 0 0 0
MeOH 664.
CHCI3 109.7
BuOH > 1 0 0 0 .Stems
H 2 O-FRAC. > 1 0 0 0 .(5b) H 2 O -E X T. > 1 0 0 0 .
Table 3.12 Activity Against Brine Shrimp of the
Extracts from Lasianthus panamensisTabie 3.13 Activity Against Brine Shrimp of the
Extracts from Picramnia antidesma subsp. fessonia
86

PLANT PART (Code N°)
EXTRACT BRINE SHRIMP LC5 0 pg/mi
MeOH > 1 0 0 0 .
CHCI3 > 1 0 0 0 .
LeavesBuOH > 1 0 0 0 .
HgO-FRAC. > 1 0 0 0 .(6 a) HgO-EXT. > 1 0 0 0 .
MeOH > 1 0 0 0 .
CHCI3 736.2
RootsBuOH > 1 0 0 0 .
H2 O-FRAC. > 1 0 0 0 .(6 b) H2 O-EXT. > 1 0 0 0 .
MeOH > 1 0 0 0 .
CHCI3 395.9
BranchesBuOH > 1 0 0 0 .
H2 O-FRAC > 1 0 0 0 .(6 0 ) H2 O-EXT. > 1 0 0 0 .
MeOH > 1 0 0 0 .
CHCI3 416.2
StemsBuOH > 1 0 0 0 .
H2 O-FRAC. > 1 0 0 0 .(6 d) H2 O-EXT. > 1 0 0 0 .
Table 3.14 Activity Against Brine Shrimp of the Extracts from Picramnia teapensis
87

Plant name Plant part ExtractKB ceil
LCso(pg/ml)
Branch MeOH 87.4
Branch BuOH 402.3
Branch CHCI3 38.2
Guarea macropetala Leaves MeOH 96.9
Leaves CHCI3 54.9
Branch MeOH 86.08
Branch CHCI3 38.5
Guarea rhopalocarpaWood MeOH 50.4
Wood CHCI3 29.1
Leaves MeOH 89.6
Leaves CHCI3 32.6
Ruagea glabraLeaves CHCI3 81.6
Stem bark CHCI3 78.7
Leaves MeOH >500
Abuta dwyeranaLeaves BuOH 320
Leaves CHCI3 338.8
Roots CHCI3 54.8
Cephaelis camponutans Roots CHCI3 27.54
Picramnia antidesma Leaves BuOH 8.7subsp. fessonia
Stems CHCI3 127.4
Table 3.15 Activity Against KB Ceils of Extracts Selected on the Basis
of Previously Activity Against Brine Shrimp.
88

Section 4. Phytochemistry

4.1 Material and General Methods
4.1.1 Plant material
More than 30 plants species were selected for collection either because of
their traditional use or because of chemotaxonomic considerations. The
botanical descriptions and the voucher specimens of each plant were carefully
studied at the Herbarium of the University of Panama before each trip
collection. The trips were organized by Mrs Carmen Galdames, a final year
student of botany and who is working at the Centre for Pharmacognostic
Research (CIFLORPAN) in the Faculty of Pharmacy, University of Panama.
The collections were made, mainly, in three different places including Western,
Middle and Eastern Panama.
Eleven plants were collected and a voucher specimen was prepared for
each plant each time collected. Sample were deposited at the Herbarium of the
University of Panama and authenticated by Prof. Mireya Correa. The plants
were separated into different part ie. leaves, roots, stembark, branches, etc.,
air dried, chopped and powdered (1 mm) using a Wiley mill (Thomas Scientific,
USA) before despatch to the School of Pharmacy, University of London. Table
4.1 (p.91) shows the date, place of collection, voucher number, plant part
collected and the amount of dried powdered plant collected; the botanical
description of each plant has been presented in Section 3.
4.1.2 Solvents
Methanol was purchased in bulk and distilled before use. All other solvents
were reagent grade and obtained from May & Baker or BDH.
90
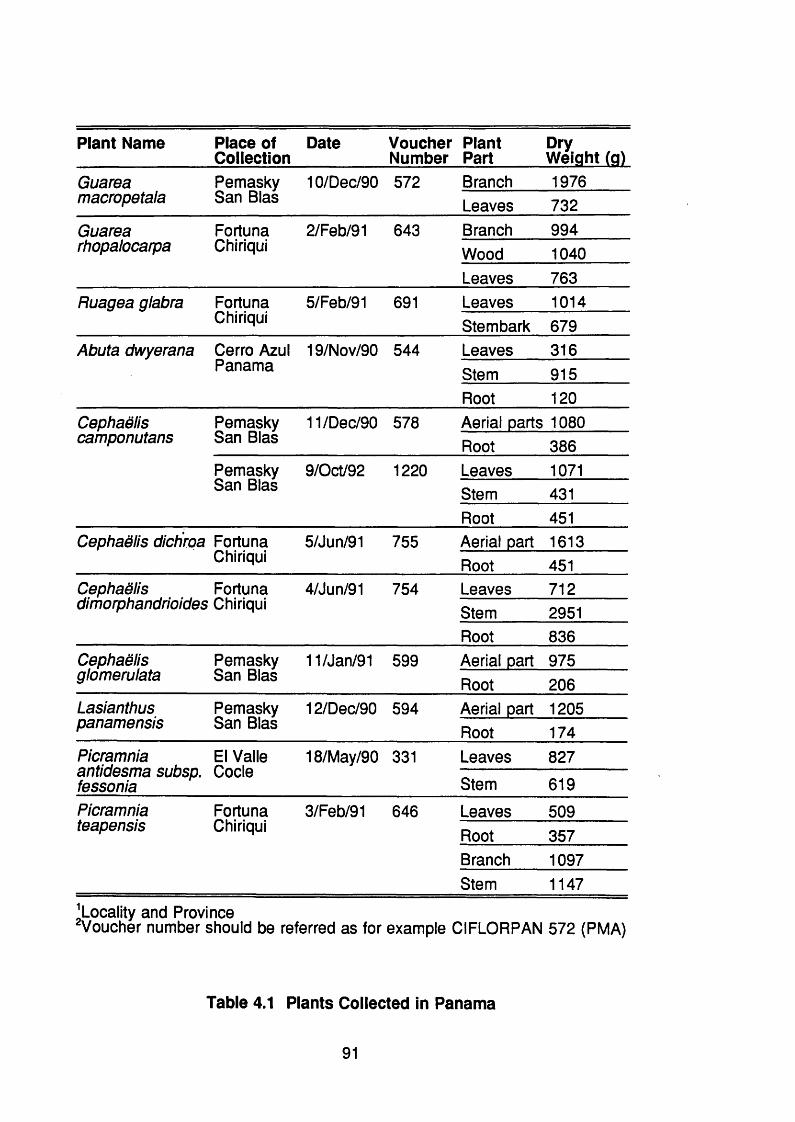
Plant Name Place of Collection
Date VoucherNumber
PlantPart
DryWeight (g)
Guarea Pemasky 10/Dec/90 572 Branch 1976macropetala San Bias Leaves 732Guarea Fortune 2/Feb/91 643 Branch 994rhopalocarpa Chiriqui Wood 1040
Leaves 763Ruagea glabra Fortune 5/Feb/91 691 Leaves 1014
Chiriqui Stembark 679Abuta dwyerana Cerro Azul 19/Nov/90 544 Leaves 316
Panama Stem 915Root 1 2 0
Cephaëliscamponutans
Pemasky San Bias
11/Dec/90 578 Aerial parts 1080 Root 386
Pemasky 9/Oct/92 1 2 2 0 Leaves 1071San Bias Stem 431
Root 451Cephaëlis dichroa Fortune
Chiriqui5/Jun/91 755 Aerial part
Root1613451
Cephaëlis Fortune 4/Jun/91 754 Leaves 712dimorphandrioides Chiriqui Stem 2951
Root 836Cephaëlisglomerulata
Pemasky San Bias
11/Jan/91 599 Aerial part Root
975206
Lasianthus Pemasky 12/Dec/90 594 Aerial part 1205panamensis San Bias Root 174Picramnia antidesma subsp. fessonia
El Valle Code
18/May/90 331 Leaves 827
Stem 619Picramnia Fortune 3/Feb/91 646 Leaves 509teapensis Chiriqui Root 357
Branch 1097Stem 1147
^Locality and Province^Voucher number should be referred as for example CIFLORPAN 572 (PMA)
Table 4.1 Plants Collected in Panama
91

4.1.3 Chromatography techniques
Thin layer chromatography (TLC) was carried out on pre-coated silica gel
GF-254 aluminum backed plates (Merck Ltd). Column chromatography (00)
was performed on silica gel 60, 70-230 mesh (Merck Ltd). Preparative TLO
(pTLO) plates were prepared with silica gel GF-254 (Merck Ltd) using a Oamag
silica gel spreader adjusted to 0.75 mm; plates were activated by heating at
1 1 0 ° 0 overnight, washed by running in acetone and air dried before use.
Visualization, of both TLO and pTLO, was by means of short (254 nm) and
long (365 nm) ultra-violet light and spray reagents were also used.
Dragendorff’s reagent, iodoplatinate (Stahl, 1969) and ferric chloride in
perchloric acid (Jork et al., 1990) were used to detect alkaloids. Kedde’s
reagent, to detect a-p unsaturated five membered lactone, and KOH in
methanol, to detect anthraquinones were prepared as previously described
(Stahl, 1969).
4.1.4 Spectroscopic methods4.1.4.1 Ultraviolet spectroscopy (UV)
Spectra were recorded in MeOH using a Perkin Elmer model 402 double
beam spectrometer. The alkaloids from Cephaëlis glomerulata were recorded
on a Perkin Elmer model 550 SE double beam spectrometer.
4.1.4.2 Infrared spectroscopy (IR)
Spectra were recorded in KBr pellets or preparing a thin film on NaCI cells
using a Perkin Elmer model 841 infrared spectrometer. The alkaloids isolated
from C. glomerulata \vere recorded on a Perkin Elmer model 1600 FT Fourier
transformed infrared spectrometer.
92

4.1.4.3 Mass spectrometry (MS)
Electron-impact mass spectra (EIMS) and accurate mass determination
were made on a VG-Analytical instrument ZABI F mass spectrometer at 70 eV
with an inlet temperature of 180 - 200 °C unless otherwise stated.
Chemical ionization mass spectra (CIMS) were recorded on a VG massiab
12-250 quadrupole instrument using ammonia as ionising gas unless otherwise
stated.
Fast atom bombardment mass spectra (FABMS) were recorded on a VG
analytical ZABSE spectrometer with xenon as the atom source at 8 eV using
the matrix indicated on each spectrum.
4.1.4.4 Nuclear magnetic resonance (NMR)
For the alkaloids isolated from C. dichroa the NMR spectra were
recorded at 250 MHz and the ^^0 NMR at 62.9 MHz on a Bruker WM250 in the
solvent indicated with IMS (tetramethylsilane) as internal standard. H-C
correlation and COSY 45 spectra were also recorded on a Bruker WM250.
All other ^H NMR spectra were recorded at 400 MHz and the NMR at
100.6 MHz on a Bruker AMX-400 in the indicated solvent with IMS as internal
standard. Two dimensional experiments were also recorded on an AMX-400
nuclear magnetic resonance spectrometer.
4.1.4.5 Polar! metry
The [a]° were recorded at 2 0 °C in the solvent and concentration indicated
on a Perkin Elmer model 241 polarimeter using a 10 cm cell.
93

4.1.4.6 Circular dichroism (CD)
The circular dichroism spectra were recorded on a Jasco J-600
spectropolarimeter in the solvent and concentration indicated.
4.1.5 Preparation of acetyl derivatives
Acétylation was carried out by taking up the compound in pyridine and
acetic anhydride and keeping the mixture in the dark overnight. The mixture
reaction was dried in a rotary evaporator by azeotroping with toluene to lower
the boiling point of the mixture.
4.2.1 Preparation of crude plant extracts for biological test
Twenty grams of each plant part were extracted twice, successively, by
maceration with petrol ether 24 hrs each time. The solvent was removed by
filtration under vacuum and both extracts were combined and evaporated in
rotary evaporator. The plant material was air dried before further extraction with
methanol (MeOH) using the procedure as described above and ultimately the
plant material was extracted with distilled water (HgO-Ext.) using the same
method.
A portion of the methanolic extract was partitioned between chloroform
(CHCy and water (HgO-Frac.); the aqueous layer was , then, extracted with
butanol (BuOH). The aqueous extract (HgO-Ext.) and the fraction (HgO-Frac.)
were lyophilised. Figure 4.1 (p.95) shows the scheme of the extraction
procedure.
4.2.1 . 1 Results
A total of 162 extracts were obtained and the amount of each extract is
presented in Table 4.2 (p.96-97).
94

D o u b l e macer at i on • w i t h pet r ol et her
t or 24 hr s
M or c
D ou bl e mocer et i on w i t h M eO H fo r 24 hrs
M a r cMeOH
Doubl e macer at i on w i t h H 2 O fo r 24 h rs
E vapor at i on
R esi du e
Ex t r actP a r u t i o n between C H C I 3 / H 2 O 1 /5 I i ophyl i zed
CH Cl
/ B u O H
BuOH IH 2O F r ac
P e t ro l et h er
D r i e d P l a n t M a t e r i a l
Figure 4.1 Extraction and Fractionation Procedure.
95
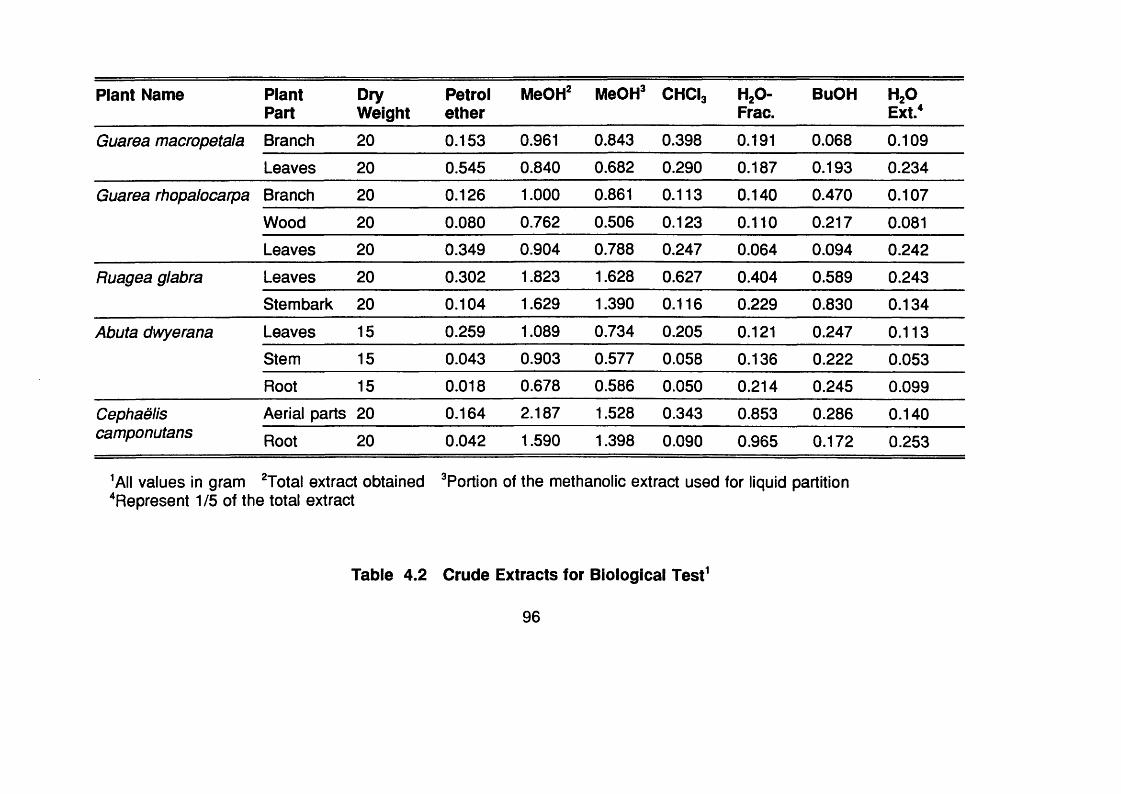
Plant Name PlantPart
DryWeight
Petrolether
MeOH: MeOH: CHCI3 HjO-Frac.
BuOH HjOExt*
Guarea macropetala Branch 2 0 0.153 0.961 0.843 0.398 0.191 0.068 0.109
Leaves 2 0 0.545 0.840 0.682 0.290 0.187 0.193 0.234
Guarea rhopalocarpa Branch 2 0 0.126 1 . 0 0 0 0.861 0.113 0.140 0.470 0.107
Wood 2 0 0.080 0.762 0.506 0.123 0 . 1 1 0 0.217 0.081
Leaves 2 0 0.349 0.904 0.788 0.247 0.064 0.094 0.242
Ruagea glabra Leaves 2 0 0.302 1.823 1.628 0.627 0.404 0.589 0.243
Stembark 2 0 0.104 1.629 1.390 0.116 0.229 0.830 0.134
Abuta dwyerana Leaves 15 0.259 1.089 0.734 0.205 0 . 1 2 1 0.247 0.113
Stem 15 0.043 0.903 0.577 0.058 0.136 0 . 2 2 2 0.053
Root 15 0.018 0.678 0.586 0.050 0.214 0.245 0.099
Cephaëlis Aerial parts 20 0.164 2.187 1.528 0.343 0.853 0.286 0.140camponutans Root 2 0 0.042 1.590 1.398 0.090 0.965 0.172 0.253
^All values in gram ^Total extract obtained ^Portion of the methanolic extract used for liquid partition ^Represent 1/5 of the total extract
Table 4.2 Crude Extracts for Biological Test^
96

Plant Name PlantPart
DryWeight
Petrolether
MeOH^ MeOH* CHCI3 HjO-Frac.
BuOH H2 OExt/
Cephaëlis dichroa Aerial part 2 0 0 . 0 2 0 1.323 0.895 0.305 0.171 0.277 0.207
Root 2 0 0.014 1.107 0.616 0 . 1 1 2 0.205 0.167 0.146Cephaëlis Leaves 2 0 0.257 1.143 1.018 0.626 0.141 0.139 0.275dimorphandrioides Stem 2 0 0.031 1.008 0.608 0.109 0.260 0.174 0.145
Root 2 0 0.024 0.466 0.306 0.063 0.078 0.083 0.103Cephaëlis glomerulata Aerial part 2 0 0.063 1.142 0.539 0 . 2 0 1 0.118 0.192 0.154
Root 2 0 0.024 0.752 0.596 0.098 0.230 0.140 0 . 1 2 1
Lasianthus Aerial part 2 0 0.070 2.685 1.758 0.370 0.940 0.350 0.213panamensis Root 2 0 0.019 1.483 0.961 0.127 0.550 0.143 0.259Picramnia antidesma Leaves 2 0 0.109 3.610 2.335 0.558 0.466 1.238 0.158subsp. fessonia Stem 2 0 0.009 1.133 0.834 0.083 0.183 0.569 0.172Picramnia teapensis Leaves 2 0 0.041 1.795 1.177 0.555 0.260 0.261 0.237
Root 2 0 0.014 1.403 1.089 0.062 0.160 0.790 0.084Branch 2 0 0.027 1.345 0.881 0.058 0.157 0.556 0.095Stem 2 0 0.007 1.036 0.829 0.077 0.318 0.388 0.152
Table 4.2 Continued.
97

4.3 Isolation and Identification of bloactlve compounds from
Cephaëlis camponutans
4.3.1 Extraction procedure
The dried and powdered woody parts (600 g) of Psychotria camponutans
were extracted with MeOH by percolation. The extract was filtered and
concentrated in vacuo, the residue (60 g) was partitioned between CHCI3 and
HgO. The CHCI3 fraction (3.2 g) showed activity against brine shrimps (see
Table 3.8, p.84), and was submitted to a bioactivity-guided fractionation by
repeated column chromatography (1.5 x 20 cm), using silica gel 60 (70-230
mesh) and CHCl3 -EtOAc (9:1) as eluent. Two major compounds were
obtained, 1-hydroxybenzoisochromanquinone (87 mg) and benz[g]isoquinoline-
5,10-dione(6img^he structure elucidation was achieved by MS, UV, IR, ^H,
and HETCOR NMR spectroscopy.
More recently collected plant material (October 1992) (see Table 4.1, p.91)
was carefully cleaned and entirely dried, in order to determine whether the
isolates were from a specific part of the plant or from fungal contamination.
Sabouraud plates were inoculated with the plant material and incubated for 72
hrs at 25 °C, the resulting fungal mass was extracted with MeOH, filtered and
concentrated, the residue was compared on TLC (CHCl3 -EtOAc 9:1). The root
bark was carefully separated from the wood of 25 g of roots, and both root bark
and wood were, separately, extracted with MeOH-CHCl3 (1 :1 ), the extracts
were compared on TLC (CHCl3 -EtOAc 9:1).
4.3.2 Identification of the isoiated compounds
4.3.3. 1 1-Hydroxy benzoisochromanquinone4.3.3.1.1 Spectrai data
UV
X(MeOH) max. nm: 211, 245, 271 sh
98

IRD (KBr) cm ’ : 3450, 1660, 1595
Elemental analysisFound: 68.37 % (C), 4.43 % (H), 27.22 % (O); calculated values for C1 3 H1 0 O4 :
67.81 % (C), 4.38 % (H), 27.81% (O)
EIMS (see spectrum 1 in Appendix I)
m/z (relative intensity %): [M]" 230 (10), 212 (70), 201 (15), 184 (100), 173
(15), 156 (35), 128 (85), 115 (24), 104 (32), 76 (45)
HRMSm/z calculated [M]+ 230.0579 (C1 3 H1 0 O4 ); found 230.0572. m/z calculated [M-
H2OY 212.0473 (C1 3 H8 O3 ); found 212.0464
NMR
and NMR assignment are given in Table 4.3 (p. 100) and 4.4 (p.1 0 1 )
respectively. HETCOR was used to achieve this assignment. Appendix
1 shows the and NMR of 1-hydroxybenzoisochromanquinone (Spectra
2 and 3, respectively).
4.3.3.1. 2 1-Acetylbenzolsochromanquinone4.3.3.1.2.1 Spectral data
NMR
(see Spectrum 4 in Appendix I) and NMR are shown in Table 4.3 (p. 100)
and 4.4, respectively.
4.3.3.2 Benz[g]lsoqulnollne-5,10-dlone
4.2.3.2.1 Spectral data
UV
X (MeOH) max. nm: 209, 249 IR
x> (KBr) cm"*: 1680, 1580, 1300,700
99

Compounds
PROTON 1-Hydroxybenzoisochroman
quinone‘
1 -Acetylbenzoisochroman
qulnone^
Benz[g]isoquinollne-
5,10-dione‘'
H-1 5.3 (1H)t (3.6) 6.43 dd (1H) (2.02,4.0) 9.57 (1H) s
H-3 4.84-4.63 (2H) ddt (3.0, 18.5) 4.62-4.78 (2H) ddd (1.56,2.3,18.8) 9.12 d (1H) (5.0)
H-4 2.84-2.64 (2H) ddq (3.0,18.9) 2.78 - 2.91 (2H) m 8.08 d (1H) (5.0)
H-6.9 7.76-7.72 (2H) m 7.73-7.77 (2H) m 8.34-8.30 (2H) m
H-7,8 8.10-8.03 (2H) m 8.13-8.06 (2H) m 7.90-7.83 (2H) m
Me—
2.09 (3H) s—
^Chemical shifts are in 5 values (ppm) with coupling constants J in Hz. ‘’Measured in CDCI3 + MeOD.‘'Measured in CDCI3.
Table 4.3 NMR Spectral Data" of Compounds Isolated from C. camponutans.
100

CompoundsCarbon
la° 2 °
C- 1 90.49 (CH)"* 89.19 (CH) 155.36 (CH)
C-3 57.93 (CHg) 57.95 (CHg) 149.65 (CH)
C-4 28.44 (CHg) 26.11 (CH;) 118.93 (CH)
C-4a 139.97 (C) 138.26 (C) 126.11 (C)
C-5 183.93 (C) 182.70 (C) 182.39 (C)
C-5a 131.87 (C) 131.63 (C) 132.92 (C)“
C- 6 134.04 (CH) 133.87 (CH) 135.01 (CH)
C-7 126.25 (CH) 126.18 (CH) 127.27 (CH)
C" 8 126.56 (CH) 126.46 (CH) 127.32 (CH)
C-9 133.99 (CH) 133.92 (CH) 138.33 (CH)
C-9a 132.12 (C) 131.78 (C) 132.92 (C)
C-10 183.44 (C) 183.05 (C) 182.47 (C)
C-lOa 141.74 (C) 140.66 (C) 138.33 (C)
C- 1 (C=0) - 169.32 (C) -
C-1(COMe)
- 20.90 (CH;) -
Chemical shifts are in 6 -values in ppm.3'‘ Measured in CDCIj-MeOD.
""Measured in CDCI3."^Multiplicity from Dept ^^0 NMR experiment.®This signal integrates for two quaternary carbons in an inverse gated decoupled NMR experiment, giving almost quantitative intensities.Compound 1 , 1-hydroxybenzoisochromanquinone; 1 a, 1 -acetylbenzoisochromanquinone; 2, benz[g]isoquinoline-5,10-dione
Table 4.4 NMR Spectral Data‘ of the Compounds Isolated from C. camponutans
101

EIMS (see Spectrum 5 in Appendix I)
m/z (relative intensity %): [M]" 209 (100), 181 (55), 153 (65), 126 (40), 83 (20),
76 (24), 50 (27)
HRMSm/z: calculated [M]" 209.0477 (CigH^NOg); found, 209.0474. m/z calculated
[M-28r, 181.0528 (C gHyNO); found, 181.0521. m/z calculated [M-56]",
153.0578 (C1 1 H7 N); found 153.0574.
NMRand NMR are shown in Table 4.3 (p. 100) and 4.4 (p. 101) respectively.
Spectra 6 and 7 in appendix I shows the and NMR of
benz[g]isoquinoline-5,10-dione.
4.3.4 Discussion
Two benzoquinones have been isolated from the woody parts of C.
camponutans, as a result of bio-activity guided fractionation using brine
shrimps. Figure 4.2 (p. 103) shows the chemical structures of the compounds
isolated from this plant and the acetyl derivative of 1 -
hydroxibenzoisochromanquinone.
The UV spectrum of 1-hydroxibenzoisochromanquinone showed a
conjugated ketone band at 245 nm and the IR spectrum showed absorption
typical of C=0 (1660 cm'^). The HRMS showed a [M] peak at m/z 230.0572
corresponding to the molecular formula C1 3 H1 0 O4 . The EIMS spectrum showed
a weak [M] m/z 230 (10 %), loss of water [M-18r m/z 212 (70 %) and
consecutive loss of two C=0 moieties, typical for quinones (Budzikiewicz et al.,
1967), as shown by peaks at m/z 184 (100 %) and 156 (35 %). A portion of
the total amount of compound analyzed lost the two C=0 moieties without
losing the water, as demonstrated by peaks at m/z 2 0 1 (15 %) \ [M-29r and
[M-56]^ m/z 173 (15%). Figure 4.3 (p.106) shows the mass spectral
fragmentation of 1 -hydroxybenzoisochromanquinone.
102

8
1 -OH-Benzoisochromanquinone
8
1 -Aoetylbenzolsochromanquinone
Benz[g]lsoqulnollne-5,10-cllone
Figure 4.2 Compounds Isolated from Cephaëlls camponutans and
the Acetyl Derivative of 1-Hydroxlbenzolsochromanquinone.
103

The NMR spectrum (CDCI3, 400 MHz) showed four aromatic protons
typical of a 1,2 substituted benzene ring and a fine triplet at 5 5.3 ppm assigned
for the proton on C-1. A double doublet at Ô 4.84-4.63 ppm (J=3.01 and 18.5
Hz) showing geminal coupling constant, was assigned to protons on 0-3 and
the double doublet at Ô 2.84-2.64 ppm, that showed similar coupling constant
to the latter, was assigned to the protons on 0-4. ^^0 NMR showed a quasi
symmetric molecule in the aromatic region, including two very similar carbonyl
signals. The signal at 6 90.49 ppm typical of a carbon supporting two oxygens,
and the ^^0 DEPT experiment showed a OH signal for this carbon. Furthermore,
the acétylation of 1 -hydroxibenzoisochromanquinone confirmed the presence
of an OH group, showing in ^H NMR of 1 -acetylbenzoisochromanquinone a
three protons singlet at S 2.09 ppm signal for the methyl of the acetyl group and
the triplet, that in 1 -hydroxibenzoisochromanquinone is at 6 5.3 ppm, being
shifted 1.1 ppm down field, indicating the position of the OH group. This is the
first report of the occurrence of 1 -hydroxybenzoisochromanquinone from natural
or synthetic sources.
The UV of benz[g]isoquinoline-5,10-dione showed absorption at 249 nm for
a conjugated ketone and the IR showed carbonyl band at 1680 cm'\ The
HRMS spectrum of benz[g]isoquinoline-5,10-dione showed a strong [M] peak
at 209.0474 corresponding to the formula C1 3 H7 NO2 . EIMS spectrum showed
a strong [M]" m/z 209 (1 0 0 ), consecutive loss of two C=0 moieties m/z 181
(55) and 153 (65), and loss of HCN m/z 126 (40); the mass fragmentation is
showed in Figure 4.4 (p. 107). The ^H NMR showed seven aromatic protons,
a singlet at 5 9.57 ppm was assigned to the proton on C-1 , two doublets
coupling to each other at Ô 9.12 ppm (J=4.95 Hz) and 5 8.08 (J=5.0 Hz) were
assigned to protons on 0-3 and 0-4 respectively; the other two signals
integrating for two protons were characteristic of a 1 ,2 -substituted benzene ring.
NMR showed a quasi symmetric molecule, including two identical
quaternary carbons at 5 132.92 assigned to 0-5a and 0-9a; there were also
two very similar signal for carbonyl groups and two pairs of OH for the benzene
ring. The isolated compound was identical on TLO (OHOl3 -EtOAc 9:1) to a
104

commercial sample of benz[g]isoquinoline-5,10-dione, obtained from Aldrich
Chemical Co. U.K.; also, the NMR spectra were superimposable.
Benz[g]isoquinoline-5,10-dione does not react with 2% KOH in MeOH, as
the other quinones, was not sensitive to Dragendorff’s reagent (only reacting
at high concentrations), and only reacted with iodoplatinate when the plate was
previously sprayed with 5% H2 SO4 .
Benz[g]isoquinoline-5,10-dione has been isolated as an impurity from
commercial acridine (Walton et al., 1983), but this is the first report of its
occurrence in the plant kingdom. However, the skeleton, but not the
benz[g]isoquinoline-5,10-dione itself has been found in Nectria haematococca,
a fungus (Parisot et al., 1990). In order to ascertain that benz[g]quinone-5,10-
dione was a constituent of C. camponutans, and not from a fungal contaminant,
the powdered plant material was incubated to produce fungal biomass.
Extraction obtained from the sabouraud plates was compared on TLC and
showed that 1-hydroxibenzoisochromanquinone and benz[g]isoquinoline-5,10-
dione were not present. When the extraction of separated root bark and wood
were compared on TLC the results showed that 1 -
hydroxybenzoisochromanquinone was mainly in the root bark, while
benz[g]isoquinoline-5,10-dione was only detected in the root wood. It is reported
that anhydrofusarubin lactol, a compound with a similar skeleton to 1 -
hydroxibenzoisochromanquinone can react with ammonia at room temperature
to yield bostrycoidin (Parisot et al., 1989), a compound with similar skeleton of
benz[g]isoquinoline-5,10-dione.
In order to establish that compound benz[g]isoquinoline-5,10-dione was not
an artifact, 1-hydroxibenzoisochromanquinone was suspended in MeOH and
concentrated ammonia (1:1) and allowed to stand for 72 hours at ambient
temperature. TLC showed that benz[g]isoquinoline-5,10-dione was absent,
even after warming the mixture at 100 °C for 2 hrs. Therefore, the possibility
that benz[g]isoquinoline-5,10-dione is an artifact formed during extraction or for
105

OH
-H , G
m/z 230.0579
CO-H
m/c 212.0473
-CO
m fi 201 mlz 184
-CO
OH
;CC^Hntfz 173
-CO
mfz 156
m fi 128
Figure 4.3 Putative Mass Spectral Fragmentation of 1 -Hydroxybenzoisochromanquinone
106

O
mfz 209.OS74C 13 H7 N O2
mfz 181.0528 C 12 N O
-co
a :
mà 153.0574 C „ H7 N,
CNH
_ n :
mfz 125.0446CiQ Hg
Figure 4.4 Putative Mass Spectral Fragmentation of
Benz[g]lsoqulnollne-5,10-dlone.
107

the presence of ammonia in nature may be excluded, as well as, the possibility
that it is due to fungal contamination.
4.3.5 Biological activity of the isolates
4.3.5.1 Results and discussion
Table 4.5 (p. 108) shows the activity of 1 -hydroxibenzoisochromanquinone,
1-acetylbenzoisochromanquinone and benz[g]isoquinoline-5,10-dione against
brine shrimps, KB cells and P. falciparum. Benz[g]isoquinoline-5,10-dione was
three times as active, against brine shrimps, than 1 -
hydroxibenzoisochromanquinone. In KB cells the activity of 1-
hydroxibenzoisochromanquinone and benz[g]isoquinoline-5,10-dione was
similar, but 1 -acetylbenzoisochromanquinone was less active.
CompoundsBrine Shrimps
LCso±sd (pM/mi)KB ceils
LCso±sd (pM/mi)P. falciparum
iCgotsd (pM/mi)
1 31.83±4.30 (2)“ 8.06±0.30 (2) 11.56±2.96 (2)
la 17.24±0.92 (2) 11.80±3.93 (2) 2 2 .1 2 ± 0 . 6 6 (2 )
2 11.72±1.48 (3) 7.75±1.44 (2) 4.02±1.53 (2)
Control'* 36.35±3.42 (3) 1.50±0.56 (3) 0.61±0.01 (2 )
“Number of test performed, in triplicate
'’Emetine was used for Brine shrimps and KB cell tests, Chloroquine
diphosphate for P. falciparum
Compound 1 , 1 -hydroxybenzoisochromanquinone, 1a, 1 -
acetylbenzoisochromanquinone, 2, benz[g]isoquinoline-5,10-dione.
Table 4.5 Activity of the Compounds isolated from P. camponutans
Against Brine Shrimps, KB Ceils and P. falciparum.
108

Benz[g]isoquinoline-5,10-dione was some three times more active than 1-
hydroxibenzoisochromanquinone and some 5 times more active than 1-
acetylbenzoisochromanquinone against multi-drug resistant P. falciparum (K1 ).
1-Hydroxibenzoisochromanquinone was four times more active against KB
cells and P. falciparum than against brine shrimps, while its acetylated
derivative appears to have the same value in the three bio-assays. 1 -
acetylbenzoisochromanquinone was less active against both, KB cells and P.
falciparum, than the starting compound 1 -hydroxibenzoisochromanquinone, an
increase in lipophilicity, apparently, decreased its activity, but the converse was
observed against brine shrimps. Although, benz[g]isoquinoline-5,10-dione has
been reported as mutagenic against insects (Walton et al., 1983), the results
obtained against the malaria parasite are promising for further studies of this
kind of compounds.
109

4.4 Isolation of Alkaloids from Cephaëlis dichroa.
4.4.1 Extraction and separation procedure
Dried and powdered aerial parts (900g) of C. dichroa (Standley) Standley
were extracted with MeOH by percolation. The extract was filtered and
concentrated to a gum (70.5g) under vacuum and then partitioned between HgO
and CHCI3 ; the aqueous layer was extracted with BuOH, the extract was
concentrated under vacuum and the residue (11.2g) was taken up in MeOH
and precipitated with MegCO. The soluble fraction was submitted to CC using
EtOAc-EtOH-HgO (2:1:2 upper layer) as eluent, strictosidine (2.9 g) was
obtained and strictosidine lactam was also detected. The remaining aqueous
fraction was basified to pH ~9 with NH4 OH and extracted with CHCI3 , the CHCI3
extracts were dried on anhydrous NagSO and concentrated under vacuum, this
fraction (0.676g) was submitted to CC using CHCl3 -MeOH (9:1) and EtOAc-
EtOH-HgO (2 :1 : 2 upper layer) as eluent, vallesiachotamine (59.3 mg),
strictosidine lactam (134.1 mg) were obtained, strictosidine and angustine were
also detected.
The CHCI3 fraction was extracted with 2 N HCI, filtered, basified to pH-9
using NH4 OH and re-extracted with CHCI3 ; extracts were dried on Na2 S0 4 and
concentrated under vacuum. This fraction (0.401 g) was submitted to CC using
CHCI3 , CHCl3 -MeOH (9:1 and 8 :2 ) as eluents. Vallesiachotamine (38.7 mg)
and angustine (3.4 mg) were isolated, and vallesiachotamine-lactone (1.5 mg)
was further purified by p-TLC using CHCl3 -EtOAc (1 :1 ) as solvent system.
4.4.2 Identification of Isolated compounds
4.4.2.1 Vallesiachotamine lactone
4.4.2.1.1 Spectral data
Chemical reactions on TLC
The isolated alkaloid gave a positive reaction with Dragendorff’s reagent as well
a giving a pink-purplish colour with Kedde’s reagent, which is typical of a-p five
110

membered lactone (Stahl, 1969).
UV(MeOH) X,max. 225, 285 (sh), 291 nm.
IR
(KBr) D max. nm \ 3430(N-H), 1750, 1740 and 1730 (C=0), 1620 (C=C).
EIMS (Spectrum 8 , Appendix I)
m/z (Relative intensity %) [M]" 364(55), 349(10), 333(22), 319(5), 305(100),
221 (11) 209(25), 169(20), 143(15), 130(15), 115(15).
HRMS
m/z calculated value: 364.1423 (CgiHgoNgOJ; found: 364.1435.
H NMR (Spectrum 9, Appendix I)
(CDCI3, 250MHz, 5): 1.85-1.92 (m,1 H, H-14a), 2.70-2.99 (m.3H, H-5,H-14p),
3.68 (8 , 3H,COOMe-16),3.48-3.75 (m,2 H, H-6 ), 3.87 (m, 1 H, H-15), 4.28 (dd,
J=12.6, 1H, H-3), 4.87 (m, 2H, H-18), 7.09-7.22 (m, 3H, H-10, H-1 1 , H-19),
7.35 (d, J=7.64, 1H, H-12), 7.48 (d, J=7.29, 1H, H-9), 7.74 (s, 1 H, H-17), 8.14
(br 8 , 1 H, N-H). A ^H-^H COSY-45 (Spectrum 10, Appendix I) was used to
confirm this assignment.
4.4.2.2 Vallesiachotamine
4.4.2.2.1 Spectral data
UV
(MeOH) Xmax. 222, 285 sh, 291
IR
(KBr) D max. cm '\ 3250, 2840, 2910, 2950, 1680 (0=0), 1660 (0=0), 1635,
1620, 1425, 1310, 1230, 1190, 750.
EIMS (Spectrum 1 1 , Appendix I)
m/z (Relative intensity %) 350 (39), 335 (2 0 ), 322 (38), 307 (28), 291 (63), 279
(100), 263 (6 ), 249 (27), 221 (67), 209 (27), 184 (22), 170 (32), 169 (28), 156
(22), 154 (21), 144 (16), 143 (16), 143 (15), 128 (22).
I l l

HRMSm/2 calculated value; 350.1630 (CjiHjaNjOj); found: 350.1641.
'H-NMR (Spectrum 12, Appendix I)
(CDCI3 . 250 MHz, 5): 1.93 (m, 1H, H-14a), 2.10 (d, J„.„=7.28,3H, H-18), 2.17
(m, 1 H, H-14P), 2.86 (m, 2 H, H-6 ), 3.64 (s, 3H, COOCHj-16), 3.73 (m, 2 H, H-
5), 4.02 (dd, J=4.57,1H, H-15), 4.48 (dd, J=11.7,1H, H-3), 6 . 6 8 ( q, J,s.„=7.30,
1 H, H-19), 7.08-7.20 (m, J=1.26, 7.24, 2 H, H-10, H-11), 7.30 (dd,J=1.48, 7.13,
1H, H-12), 7.49 (dd,J=7.25, 1H, H-9), 7.68 (s, 1H, H-17), 7.95 (br S, 1H, N-H),
9.37 (S , 1H, H-21).
"C NMR
(62.9 MHz, CDCy 5 132.5 (C-2 ), 49.4 (C-3), 50.7 (0-5), 2 2 . 1 (0-6), 108.0 (0-
7), 126.8 (0-8) 118.1 (0-9), 119.8 (0-10), 122.1 (0-11), 111.1 (0-12), 136.3(0-
13), 34.1 (0-14), 30.9 (0-15), 94.2 (0-16),168.4 (000-16), 51.1 (-OMe-16),
147.5 (0-17), 15.0 (0-18), 152.8 (0-19), 146.4 (0-20), 195.9 (0-21). 'H-” 0
heterocorrelatlon was used to achieve this assignment.
4.4.2.3 Strictosidine
4.4.2.3.1 Spectral data
UV
(MeOH) Xmax. 227, 283, 289.
IR
(KBr) T) max. cm ’ : 2900,1660, 1630,1450,1325, 1090.
FARMS (Spectrum 13, Appendix I)
(MNOBA + Na matrix) m/z (relative intensity %) 531 (4), 329 (16), 236 (20), 176
(100), 154(25), 136(24).
’H-NMR (Spectrum 14, Appendix i)
(OD3OD, 400 MHz, 8): 1.90-2.08 (m, 2 H, H-14), 2.64-2.73 (m, 2H, H-6 , H-20),
2.79-2.82 (m, 1 H, H-6 ), 2.91-2.96 (m, 1 H, H-5), 3.00-3.05 (m, 1 H, H-15), 3.23-
3.45 (m, 5H, H-2',H-3', H-4’, H-5', H-5), 3.62-3.69 (m, 1 H, H-6 ’), 3.75 (s, 3 H, H-
23), 3.95-3.98 (m, 2H, H-6 ’, H-3), 4.80 (d, J=7.8, 1H, H-1 ’), 5.19-5.32 (ddd,
J=0.9, 11.0, 17.4, 2H, H-18), 5.79-5.88 (m, 2 H, H-2 1 , H-19), 6.94-6.97 (ddd.
112

J=0.9,7.2,7.6,1 H, H-10), 7.01 -7.05 (ddd, J=0.9, 6.9, 8.1,1H, H-11), 7.24-7.38
(dd, J=0.9, 7.6, 1H, H-12), 7.37 (d, 1H, H-9), 7.67 (s, 1H, H-17).
” C NMR(100.6 MHz, CD3OD) 5 136.54 (C-2 ), 52.01 (C-3), 43.66 (C-5), 22.72 (C-6 ),
108.72 (C-7), 128.73 (C-8 ), 118.92 (C-9), 120.00 (C-1 0 ), 122.29 (C-11), 112.14
(C-1 2), 137.93 (C-13), 37.38 (C-14), 33.04 (C-15), 110.96 (C-16), 155.71 (C-
17), 119.55 (C-18), 136.26 (C-19), 46.16 (C-20), 97.83 (C-2 1 ), 170.47 (C-2 2 ),
52.59 (C-23), 100.55 (C-1'), 74.89 (C-2 ’), 78.91 (C-3’), 71.97 (C-4’), 78.23 (C-
5’), 63.26 (C-6 ’). COSY-45 (Spectrum 15, Appendix I) and 'H-'^C
heterocorrelatlon (Spectrum 16, Appendix I) NMR experiments were used to
confirm these assignments.
4.4.2.4 Strictosidine iactam4.4.2.4.1 Spectrai data
UV
(MeOH) Xmax. 225, 283, 291
iR
(KBr) ucm ’ : 3400, 2920,1650 (C=0), 1590, 1080, 745
EiMS (Spectrum 17, Appendix I)
m/z (relative Intensity %) [MJ* 498 (35), 336 (1 2 ), 335 (1 2 ), 319 (13), 265 (92),
235 (100), 221 (1 1 ), 206 (50), 169 (58), 143 (32), 127 (24), 115 (33), 85 (33),
73 (44)
CiMS
(NH3 gas) m/z (relative Intensity): [M+1 ]" 499 (25), 337 (10), 267 (52), 250
(19), 237 (14), 198 (10), 180 (100), 160 (27), 145 (40), 127 (31), 115 (20), 85
(37), 69 (70).
HRMS
m/z calculated value 498.2002 (CjeHjoNjOe); found 498.2016
’H NMR (Spectrum 18, Appendix I)
(CD3 OD, 250MHz, 5): 1.97-2.10 (m, 1 H, H-14a), 2.43-3.24 (m, 1 OH, H-14P, H-5,
113

H-6 , H-15, H-2 ', H-3', H-4’, H-5’), 3.60-3.67 (d, J=11.7,1H, H-6 ’), 3.83-3.88 (dd,
J=2.3,11.7,1H, H- 6 ), 4.61 (d, J=7.9,1H, H-1’), 4.92-4.90 (m, 1H, H-20), 5.06
(m, 1H, H-3), 5.30-5.41 (m, 3H, H-18, H-21), 5.57-5.72 (m, J=9.8,17.2,1H, H-
19), 6.97 (m, 2 H, H-10, H-1 1 ), 7.32-7.40 (m, 3 H, H-9, H-1 2 , H-23).
” C NMR(62.9 MHz, CDCy 8 137.70 (C-2 ), 54.22 (0-3), 43.75 (C-5), 27.20 (C-6 ),
109.14(0-7), 128.66 (C-8 ), 118.80 (C-9), 122.32 (C-10), 122.04 (C-1 1 ), 112.21
(C-1 2 ), 134.57 (C-13), 21.64 (C-14), 24.68 (C-15), 110.33 (C-16), 165.14 (17),
120.36 (C-18), 133.35 (C-19), 44.56 (C-2 0 ), 97.82 (C-2 1 ), 148.02 (C-23),
100.51 (C-1 ’), 74.12 (C-2 ’), 77.93 (C-3 ), 71.34 (C-4 ), 77.93 (C-5’), 62.74 (C-6 ’).
4.4.Z4.2 Acetyl strictosidine lactam4.4.2.4.2.1 Spectral data
UV
(MeOH) W ax. 227, 283, 291
IR
(KBr) Dcm' : 3400. 2940, 1760, 1660, 1498, 1220, 1065, 1040, 965.
EIMS (Spectrum 19, Appendix I)
m/z (relative intensity %) [M]* 6 6 6 (1 0 0 ), 624 (8 ), 564 (1 2 ), 379 (23), 331 (92),
319 (100).
FABMS
(MNOBA + Na matrix) m/z (relative intensity %) [M+Na]^ 689 (100), [M]^ 6 6 6
(22), 319 (6 ), 265 (15), 169 (46), 109 (34).
HRMS
m/z calculated value 666.2425 (C3 4 H3 8 N2 O1 2 ) found: 666.2434.
H NMR (Spectrum 20, Appendix I)
(CDCI3, 250MHz, Ô): 1.21, 1.88, 1.99, 2.07 (s,3H each,Me-Ac),2.03-2.19
(m,2H,H-14), 2.58-2.71 (m, 3H, H-5, H-15),2.99-3.04 (m,2H,H-6), 3.65-3.72 (m,
1H, H-3'), 4.06-4.29 (dd, 2H, H-6') 4.74-4.82 (m, 2H, H-4', H-21 ) 4.94-5.02 (m,
3H, H-3, H-20, H-5'),5.08-5.16 (m,1 H,H-2'), 5.27-5.36 (m, 3H, H-18, H-1 '), 5.53-
114

5.68 (quint, 1 H. J=9.58, 7.8, 13.5, H-19), 7.06-7.12 (t, 1 H, J=1.0, 7.0, H-10),
7.15-7.21 (t, 1H, J=1.0, 7.0, H-11), 7.36 (dd, 1H, H-12), 7.42 (S, 1H, H-17),
7.43 (dd, 1H, H-9), 8.26 (br s, 1 H, N-H). COSY-45 and 'H-” C
heterocorrelatlon (Spectrum 22, Appendix I) NMR experiments were used to
confirm these assignments.
"C NMR(62.9 MHz, CDCy S 132.9 (C-2), 53.4 (C-3), 43.7 (C-5), 21.0 (C-6 ), 108.5 (C-
7), 127.5 (C-8 ), 118.0 (C-9), 119.8 (C-1 0 ), 122.2 (C-11), 111.3(C-12,136.1 (C-
13), 26.5 (C-14), 23.9 (C-15), 110.5 (C-16), 164.7 (C-16a), 120.8 (C-18), 132.1
(C-19), 42.8 (C-20), 94.9 (C-21), 146.9 (C-17), 94.8 (C-1), 72.0 (C-2 '), 72.1 (C-
3 ), 70.0 (C-4’), 68.3 (C-5’), 61.7 (C- 6 ), 169.1, 169.5, 1699.9, 170.6 (COO
Acetyl), 19.2, 20.5, 20.6, 20.7 (CH, Acetyl).
4.4.2.S Angustine4.4.2.5.1 Spectral data
UV
(MeOH) Xmax. 214, 376, 396.
EIMS (Spectrum 2 2 , Appendix I)
m/z (relative Intensity %) [M]* 313 (99), 298 (28), 282 (9), 255 (13), 169 (5),
157 (12), 141 (11), 128 (18), 115 (17), 94 (35), 44 (43), 28 (100).
HRMS
m/z calculated value 313.1215 (C^gHi^NgO); found 313.1219
'H NMR
(DMSO-Dg, 250MHz, 5): 3.14 (t, 2H, J= 6 .6 , H-6 ), 4.41 (t, 2H, J=6 .6 , H-5), 5.63
(dd, 1H, J= 1.0,11.8 , H-18), 6.07 (dd, 1H, J=1.0,17.5, H-18), 7.10-7.36 (m, 4H,
H-10, H-1 1 , H-14, H-19), 7.47 (dd, 1 H, J= 2.4, 8 .1 , H-12), 7.63 (d, 1 H, J= 7.8,
H-9), 8 . 8 8 (S, 1H, H-21), 9.24 (s, 1H, H-17), 11.82 (s, 1H, H-1).
115
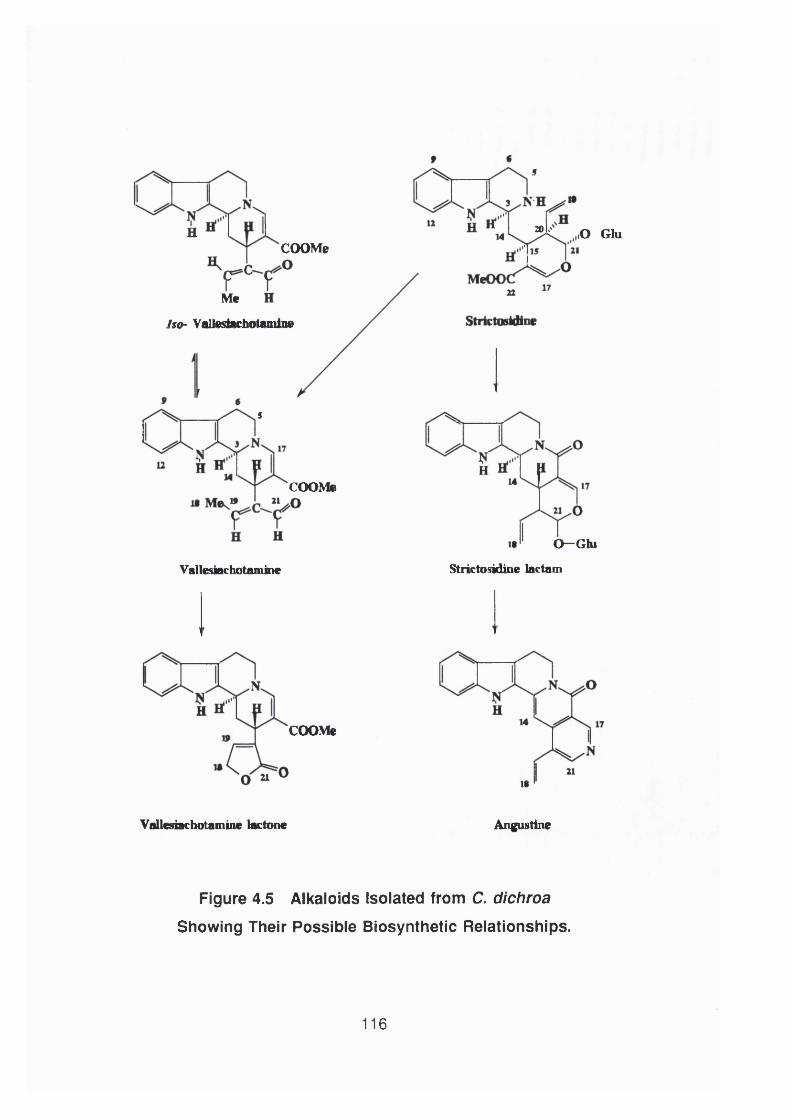
20jk' O GhiCOOMe
Me
Is<h ValWmcholmmine
s
COOMe
O -G lu
Strictosidine lactamVallesiachotamine
COOMe
Vallesiachotamine lactone Angustine
Figure 4.5 Alkaloids Isolated from C. dichroa
Showing Their Possible Biosynthetic Relationships.
116

4.4.3 Discussion
The structures of the isolated compounds are showed in Figure 4.5 (p.116),
unexpectedly all of them are indole derivatives instead of the anticipated
emetine-related alkaloids which are found in C. ipecacuanha. This figure also
shows the possible biosynthetic relationship of the alkaloids isolated from C.
dichroa.
The UV spectrum of vallesiachotamine lactone was characteristic for
alkaloids of the vallesiachotamine type (Djerassi et al., 1966) and the IR
spectrum showed absorptions typical of NH, C= 0 of ester and five membered
lactone ring. The EIMS showed a molecular ion m/z 364 (55) with fragments
for the loss of CHg m/z 349 (10), CH3O m/z 333 (22), COOCH3 m/z 305 (100)
and the loss of the lactone ring and the ester function giving arise to m/z 2 2 1
(1 1 ), Figure 4.6 (p.118) shows the mass fragmentation. The HRMS spectrum
of vallesiachotamine lactone showed a [M]" peak at m/z 364.1435
corresponding to the molecular formula C2 1 H2 0 N2 O4 .
The NMR spectrum (CDCl3,250 MHz,) was found to be closely related to
that of iso-vallesiachotamine (Waterman and Shouming Zhong, 1982) and
signals were observed for the proton at C-3 and it has an a-
configuration (Ô 4.28). A two-proton broad singlet at 5 4.87 was assigned to the
0-18 and one proton in the aromatic region (5 7.09 - 7.22 m,3H) was assigned
to the proton on 0-19. Furthermore, ^H-^H OOSY (Spectrum 1 0 , Appendix I)
clearly showed long range coupling both between 6 3.87 (0-15H) and the
multiplet at Ô 7.09 - 7.22 (0-19H) and between 5 3.87(0-15H) and S 4.87
(2H,0-18H) supporting a lactone ring for this alkaloid attached to position 15.
In addition, Kedde reagent gave a clear pink-purplish colour (Stahl, 1969)
indicating the presence of the five membered a-p unsaturated lactone; while no
colour was given by a mixture of vallesiachotamine and isovallesiachotamine.
The spectral data of vallesiachotamine agreed closely with that previously
117

co o
mfz 349miz 364.1435
m /l 305mfz 333
m/z 221
Figure 4.6 Putative Mass Spectral Fragmentation of
Vallesiachotamine lactone.
118

published (Waterman and Shouming Zhong, 1982) except that the
Heterocorrelation experiment indicated that the multiplet at 5 2.86 should be
assigned to the C- 6 protons and the multiplet at 5 3.73 to protons on 0-5.
The spectra clearly resolve the 0-14 proton into multiplets at Ô1.93 and 5 2.17
corresponding to the a and p protons respectively. In addition, it was
observed that the isolated sample of vallesiachotamine after two months in
storage was converted to a 1 : 1 mixture of vallesiachotamine and iso
vallesiachotamine.
Vallesiachotamine was originally isolated from Vallesia dichotoma (Djerassi
et al., 1966), and subsequently from Rhazya orientalis and R. stricta all of
which are members of the family Apocynaceae (Evans et al., 1968); more
recently it has been reported from Strychnos trycalysiodes
(Loganiaceae)(Waterman and Shouming Zhong, 1982). Vallesiachotamine has
not been reported in any taxa of the Rubiaceae.
The spectral data of strictosidine agreed closely with that previously
published (Battersby et al., 1969; Smith, 1968), which is the putative precursor
of the vallesiachotaman type alkaloids. Strictosidine was lactamized by adding
NagOOj (Battersby et al., 1969), the NMR and mass spectra of the product
were similar to that obtained for strictosidine lactam, confirming the identity of
the isolated compound as strictosidine. The and NMR were fully
assigned by COSY-45 and ^H-^% heterocorrelation.
The UV, IR, and NMR of strictosidine lactam agreed with those
recently published (Atta-Ur-Rahman et al., 1991); it has previously been
isolated from Nauclea latifolia (Rubiaceae) (Brown et al., 1977; Hotellier et al.,
1975) and from Rhazya stricta (Apocynaceae) (Warren, 1970). In the present
study, the strictosidine lactam was acetylated and its and ^^0 NMR fully
assigned by Heterocorrelation, which showed that the C-3 (S 53.4)
previously reported as Ô 53.6 (Cordell, 1981) clearly correlated with the multiplet
at 5 4.94-5.02 (3H) rather than with the signal at 5 2.58-2.71 previously reported
119

(Atta-Ur-Rahman et al., 1991). The isolated alkaloid and its acetylated
derivative were found to be identical on TLC with an authentic sample of
strictosidine lactam, kindly supplied by Dr. R.T. Brown, University of
Manchester, and their ^H-NMR were superimposable.
The spectral (UV, EIMS, NMR) data of a yellow fluorescent compound
agreed closely with that observed previously for angustine, which occurs widely
throughout the Rubiaceae and Loganiaceae (Phillipson et al., 1974; Au et al.,
1973). The isolated compound was found to be identical on TLC with an
authentic sample of angustine available in the laboratory.
120

4.5 Isolation and Identification of Alkaloids from Cephaëlis
glomerulata,4.5.1 Extraction and separation procedures
Dried and powdered aerial parts (792 g) of Cephaëlis glomerulata J.Don.
Smith were extracted with MeOH by percolation. The extract was filtered and
concentrated to a gum (49.4 g) under vacuum and partitioned between 2 M HCI
and CHCI3. The acidic fraction was filtered, basified to pH~9 using NH4 OH and
re-extracted with CHCI3. The CHCI3 fraction was dried on NagSO and
concentrated under vacuum and the residue (1.08 g) was submitted to 0 0
using OHOl3 -MeOH + NH4 OH (98:2 + 0.1 %) as solvent system. A total of 50
(10 ml) fractions were obtained, from fractions 9-11 47 mg glomerulatine A
were obtained; fractions 13-27 containing glomerulatine A and 0 were
chromatographed on pTLO using EtOAc-MOgOO + NH4 OH (80:20 + 0.1 %) as
solvent system, 5 mg of glomerulatine 0 were, thus, obtained. Fractions 32-38
were chromatographed on pTLO and developed in the same conditions,
yielding 4 mg of glomerulatine B.
4.5.2 Identification of the Isolated compounds
4.5.2.1 Glomerulatine A
4.5.2.1.1 Spectral data
UV
X max. (EtOH)nm: 274, 304 shoulder IR
NaCI \) cm ': 2919, 2860, 1625 (C=N), 1590 (C=C), 1466, 1407, 1272, 1219,
755
[a]“20°C
(0=0.36, GHCy -466.1
CD (Spectrum 23, Appendix I)
(CH3CN, 0= 105 nM) X nm (A e): 310.3 (-13.49), 296.0 (-13.11), 280.6 (+0.04),
2.68.8 (+12.66), 246.6 (+2.99), 214.7 (+12.17).
121

EIMS (Spectrum 24, Appendix I)
m/z (relative intensity %) 342 (100), 327 (3), 313 (10), 298 (11), 284 (20), 270
(14), 256 (16), 255(16), 242 (12), 230 (7), 209 (14), 128 (10). 115 (12)
HRMSm/z calculated: 342.18445 (CggHggNJ found:342.18565
NMR (Spectrum 25, Appendix I)
(CeDg , 400 MHz) Ô 1.35-1.39 (dd, J=6.2, 1 H, H-3), 1.90-1.98 (m, 1H, H-3),
2.61 {t, J=8.98, 1H, H-2), 2.92 (s, 3H, N-1 Me), 2.96-3.09 (m. 1H, H-2), 6.62-
6.67 (m, 1H, H-5), 6.76-6.78 (dd, J=7.7, 1.5, 1 H, H-4), 6.93-6.98 (m, 1H, H-6 ),
7.46-7.48 (dd, 1H, H-7)
NMR (Spectrum 26, Appendix I)
( CgDe, 100.6 MHz): 5 30.26 (C-3), 30.87 (N- 1 Me), 48.15 (0-2), 49.09 (C-3a),
122.28 (C-5), 123.68 (C-4), 125.04 (C-7), 126.41 (C-4a), 128.90 (C-6 ), 147.32
(C-7a), 165.11 (C-8 a). COSY 45, ^H- ^C heterocorrelation (Spectrum 27,
Appendix I), ^H- ^C long range heterocorrelation (Spectrum 28, Appendix I) and
ROESY (Spectrum 29, Appendix I) NMR experiments were used to confirm
these assignments.
4.5 2.2 Glomerulatine B4.5.2.2.1 Spectral data
UV
X max.(CH3 CN) nm: 279
CD
(CHgCN, C= 116 nM) X nm (A e): 305.5 (-12.97), 219.9 (-13.11), 274.3 (+0.098),
262.9 (+12.19), 242.2 (+8.09), 213.9 (+12.27)
EIMS (Spectrum 30, Appendix I)
m/z (relative intensity %): 328 (100), 313 (6 ), 299 (12), 284 (9), 270 (8 ), 243
(5), 229 (4), 132 (17), 113 (11), 50 (27)
HRMS
m/z calculated: 328.16880 (CgiHgg NJ found 328.16957
122

'H NMR (Spectrum 31, Appendix I)
(CgDg, 400 MHz): 5 1.30-1.34 (m, IN, H-3), 1.46-1.49 (m. 1 H, H-3), 1.92-2.03
(m, 2H, H-3 and 3 ), 2.58 (t, J=8.98,1H, H-2), 2.90 (s, 3H, N-1 Me), 2.93-2.95
(m, 1H, H-2), 3.17-3.19 (m, 1H, H-2 ), 3.29-3.33 (m, 1 H, H-2 ’), 6.58-6.62 (m,
1 H, H-5’), 6.64-6.68 (dd, 1 H, H-5), 6.77 (d. J=7.51, 1H, H-4), 6.88-6.92 (dd,
J=7.63, 7.39,1H, H- 6 ), 6.94-6.98 (dd, J=7.51,1H, H-6 ), 7.10-7.12 (m, J=7.63,
6.15, 2H, H-7’ and 4 ), 7.47 (d, J=7.63, 1H, H-7)
"C NMR (Spectrum 32, Appendix i)
(CgDg, 1 0 0 . 6 MHz): 8 30.28 (C-3 and 3’), 30.79 (N- 1 Me), 48.09 (C- 2 and 2 ’),
48.62 (C-3a and 3’a), 120.85 (C-4), 1 2 2 .2 1 “ (C-5), 122.59“ (C-5 ), 124.03" (C-
4 ), 124.37" (C-7’), 125.15" (C-7), 125.42" (C-4’a), 126.14" (C-4a), 128.78" (C-
6 ’), 128.97" (C-6 ), 147.13 (C-7a and 7’a), 164.68 (C-8 a and 8 ’a), the
superscript indicates interchangeable assignment within the same letter. COSY
45, 'H-"C heterocorrelatlon, 'H-"C long range heterocorrelatlon and "C DEPT
NMR experiments were used to confirmed these assignments.
4.5.23 Glomerulatine 04.5.2.3.1 Spectral data
UV
X max. (EtOH) nm: 274, 304 shoulder
IR
NaCI u cm ’ : 2952, 2857, 2352, 1629 (C=N), 1588 (C=C), 1470, 1407.
CD
(CHjCN, C= 65 nM) X nm (A e): 305.1 (-12.08), 287.2 (-12.32), 258.6 (+0.1),
247.9 (+12.47), 228.4 (+9.40), 216.4 (+13.35)
EIMS (Spectrum 33, Appendix i)
m/z (relative intensity %) 344 (23), 343 (6 8 ), 273 (23), 256 (57), 219 (20), 197
(23), 169 (25), 153 (22), 135 (19),130 (41), 127 (22), 115 (30)
HRMS
m/z [M-1]* calculated 343.19227 (Cg^Hg^NJ found 343.19248
123

NMR (Spectrum 34, Appendix I)
(CgDe, 400 MHz) 5: 1.52-1.58 (m, IN, H-3'), 1.83-1.87 (m, 1H, H-3), 1.91-1.98
(m, 1H,H-3’), 2.14 (s. 3H, N-1’ Me), 2.17-2.20 (m, 1 H, H-2 ’), 2.30-2.34 (m, 1 H,
H-3), 2.63-2.70 (m, 2H, H-2 and 2’), 2.97 (s, 3H, N-1 Me), 2.95-3.02 (m, 1H,
H-2), 3.78 (d, J=4.52,1H, H-8 ’a), 3.86 (s broad, 1 H,H-8 ’) 6.19-6.21 {dd, J=0.88,
7.48, 1H, H-7’), 6.45-6.49 (ddd, J= 1.12, 7.36, 7.52, 1 H, H-5’), 6.74-6.86 (m,
3 H, H-5, 4’ and 6 ), 6.96-7.00 (dd, J=7.68, 7.44,1 H, H-6 ) 7.01-7.04 (dd, J=1.2,
7.72, 1H, H-4), 7.49-7.51 (dd, J=1.24, 7.76, 1H, H-7)
NMR (Spectrum 35, Appendix I)
(CgDe, 100.6 MHz) 6 : 30.73 (N- 1 Me), 31.71 (C-3), 34.02 (0-3’), 39.57 (N-1 ’
Me), 45.30 (3’a), 48.51 (0-2), 49.15 (0-3a), 50.45 (2’), 76.51 (0-8’a), 114.37
(0-7’), 117.53 (0-4’), 122.11 (0-5), 122.26 (0-4’a), 123.00 (0-4), 124.94 (0-5’),
125.2 (0-7), 127.12(0-6 ), 128.80 (0-6), 129.52 (0-4a), 142.10 (0-7’a), 148.62
(0-7a), 166.49 (0-8a). OOSY-45 (Spectrum 36, Appendix I), DEPT, ^H-'^O
long range Heterocorrelation (spectrum 37, Appendix I) and ROESY (Spectrum
38, Appendix I) NMR experiments were used to confirm these assignments.
4.5.3 DiscussionThe UV spectrum of glomerulatine A showed a band at 274 nm of a
conjugated quinoline moiety and the IR spectrum showed adsorption for 0=N
(1625 cm'^) and 0=0 (1590 cm'^). The HR mass spectrum of glomerulatine A
showed a [M]^ peak at m/z 342.18565 corresponding to the molecular formula
O2 2 H2 2 N4 . While the EIMS showed a strong [M]*" with 1 00% relative abundance,
indicative of calycanthine-type alkaloids instead of chimonanthine-type
(Adjibade et al., 1992). However, the peak at m/z 231 has no significant
abundance suggesting that the iso-calycanthine-form be more likely for this
compound (Adjibade et al., 1992).
However, 1 the spectroscopic techniques used, in the present work,
in the structure elucidation which include EIMS, ^H, NMR, 2D NMR
experiments, [a]° and 0 1 ^ failed to distinguish between the calycanthine- or iso-
calycanthine-type. Hence, glomerulatine A is either the 8 -8 a,8 ’-8 ’a
124

tetradehydro(-)calycanthine or the 8-8a,8'-8'a tetrad0 hydro(-)iso-calycanthine
(Figure 4.7, p. 128).
Table 4.6 (p. 126) shows a comparison of the spectroscopic data of
caiycanthine and iso-calycanthine previously published and glomerulatine A.
It is important to note that the chemical structure of caiycanthine has been
established by X-ray diffraction (Hamor et al., 1960). In a more recently
published work, Adjibade et al. (1992) characterized the first five membered
ring quinoline alkaloid as iso-calycanthine. Their argument were based mainly
on slight differences in and NMR, differences in relative abundance in
EIMS and the major difference being the values, when comparing iso-
calycanthine with caiycanthine.
Furthermore, Adjibade et al (1992) did not find difference in the
displacements of the signals in NMR of the C- 2 and C-3, although, the
signals in NMR for a five membered ring are shifted down field when
compared with a six membered ring. For example, the C-2 and 0-3 in
chimonanthine forming a five member ring has shown signals a 5 52.6 and 5
35.7 respectively, whereas, in caiycanthine, as part of a six member ring, they
show signals at 5 46.6 and 5 31.7, respectively (Libot et al., 1988). The
structure of iso-calycanthine has not been confirmed by X-ray diffraction
analysis.
The NMR (CgDg, 400 MHz) spectrum of glomerulatine A showed only 1 1
protons and the ^^0 NMR showed 11 carbons, indicating that the dimeric nature
of the alkaloid. There were signals for four aromatic protons, from 7.48 to 6.62
ppm, characteristic of a 1 ,2 -disubstituted aromatic ring; four protons signals
from 3.09 to 1.35 ppm, clearly separated from each other, were assigned to
protons on 0-2 and 0-3 using OOSY 45 and ^H-^^0 heterocorrelation NMR
experiments.
125
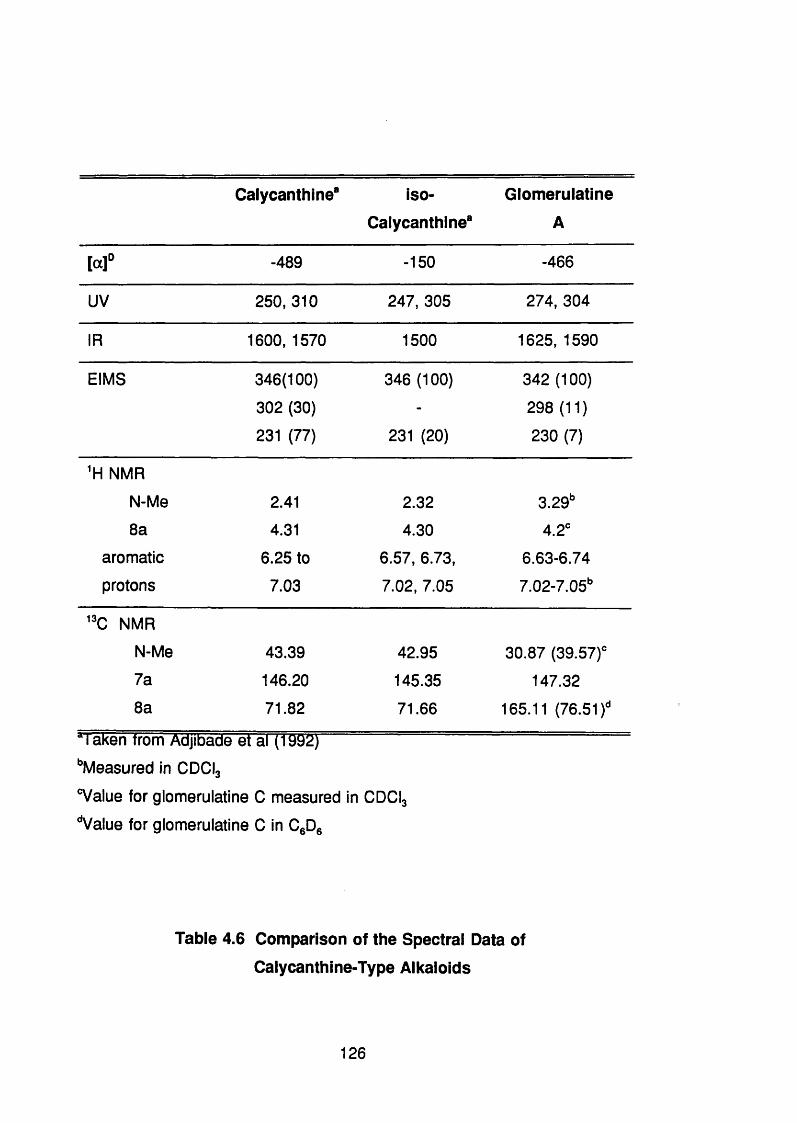
Caiycanthine* iso-
Calycanthlne*
Glomerulatine
A
[aŸ -489 -150 -466
uv 250,310 247, 305 274, 304
IR 1600, 1570 1500 1625, 1590
EIMS 346(100) 346 (100) 342 (100)
302 (30) - 298 (11)
231 (77) 231 (20) 230 (7)
'H NMR
N-Me 2.41 2.32 3.2Sf
8 a 4.31 4.30 4.2'
aromatic 6.25 to 6.57, 6.73, 6.63-6.74
protons 7.03 7.02, 7.05 7.02-7.05"
” C NMR
N-Me 43.39 42.95 30.87 (39.57)'
7a 146.20 145.35 147.32
8 aa r r -z .T T Z i~ ï^ J .___ ■ - i ü l
71.82 71.66 165.11 (76.51)'
‘'Measured in CDCI3
®Value for glomerulatine C measured in CDCI3
‘Value for glomerulatine C in C Dg
Table 4.6 Comparison of the Spectral Data of
Calycanthine-Type Alkaloids
126

Also, the NMR showed a singlet integrating for three protons at 2.92 ppm
for a N-methyl, and long range heterocorrelation NMR experiment
showed a correlation between the signal of the N-methyl and the carbon
signals at 165.11 (C-8 a) and 48.15 ppm (C-2 ), establishing its position on N-1 .
The G-3a shows a signal at 5 49.09 shifted to lower field because it is adjacent
to an additional conjugated carbon (C-8 a), rather than to a methine group as
in caiycanthine, which displays a signal at 5 30-40 characteristic of a
piperidinoquinoline ring (Libot et al.,1988). Whilst, a ROESY effect was shown
between the N-methyl and the proton on 0-4’, and the proton a on C- 2 and C-
4’, which are more feasible in the /so-calycanthine form than in caiycanthine,
where the methyl and the protons a on C- 2 are farther from the protons on C-
4’, however, it is possible as well.
The showed that glomerulatine A is levogyre with a value of (-)466, close
to that reported for (-) caiycanthine (- 489) (Adjibade et al., 1992), whereas, for
/so(-)calycanthine a value of (-)150 has been reported (Adjibade et al., 1992).
The CD spectrum indicated a S-configuration for glomerulatine A. Hence, the
isolated compound may have a configuration similar to caiycanthine, however,
none of the spectroscopic techniques used to elucidate this structure showed
further evidence to distinguish between the caiycanthine or iso-calycanthine
isomers. Only X-ray crystallography could unravel this problem and, although,
the isolated alkaloid was crystallized the crystals were not good enough to
complete the experiment. This plant should be collected again in order to
isolate more of these compounds to establish the configuration. Figure 4 . 7
(p. 128) shows both possible isomers of the isolated alkaloids from C.
glomerulata.
Glomerulatine B showed similar UV and IR spectra to glomerulatine A.
The HR mass spectrum showed a [M] peak at m/z 328.16957 corresponding
to the molecular formula C2 1 H2 0 N4 . EIMS showed a strong [M]^ with 100%
relative abundance, fourteen mass units less than glomerulatine A suggesting
that this alkaloid is the structural N-demethyl isomer.
127

HjCk
1 CH)
o r
5
G l o m e r u l a t i n e A
o r
G l o m e r u l a t i n e B
" ^ n j I
o rCH, N
CH,
G l o m e r u l a t i n e C
Figure 4.7 Possible Structures of the Alkaloids Isolated from
Cephaëlis glomerulata.
128

NMR showed an unsymmetrical molecule and only one signal integrating
for three protons at 5 2.90 for a N-methyl group similar to glomerulatine A.
The signal for the protons on C-2 and 0-3 were spread from 5 1.30 to 3.33,
whereas, the aromatic part of the spectrum showed eight protons indicating two
orï/70-disubstituted aromatic rings. Furthermore, NMR in CDCI3 showed a
very broad singlet at ô 2.55 that disappeared after the addition of DgO,
indicating the NH at position 1 '. The assignment of this spectrum was achieved
by comparison with glomerulatine A, since one monomeric moiety is identical,
also for 2D NMR experiments, such as COSY 45, ^H- ^C heterocorrelation and
^H- ^C long range heterocorrelation confirmed assignments made for the
demethyl analogue.
^ C NMR showed sixteen signals indicating that some of them were
overlapped. In the upper field, the signals for C-8 a and C-7a were quite similar
to those for glomerulatine A. While, the aromatic CH and the C-4a were split
showing ten separated signals, but, resembling the values showed for
glomerulatine A.
Glomerulatine C showed an UV and IR spectra similar to glomerulatine A
and the HR mass spectrum showed a [M-1T at m/z 343.19227 indicating
to the molecular formula C2 2 H2 3 N4 . EIMS showed a relatively weak [M]^ peak
with only 23% of relative abundance, but a stronger [M-l]'" with 6 8 % of
relative abundance, two mass units less than glomerulatine A, suggesting that
this alkaloid has loss one double bond. NMR, as in glomerulatine B,
showed an unsymmetrical molecule including two singlets integrating for three
protons each, one of them at 5 2.97 as in glomerulatine A and B and the
other at Ô 2.14 (52.40 in CDCy as reported for caiycanthine (Libot et al., 1988).
Also, a doublet at 5 3.78 and a broad singlet at 5 3.86 for a proton on the
carbon between the two nitrogens and for N-H respectively. This data
suggested that one of the monomers is similar to glomerulatine A and the
other similar to caiycanthine.
129

Furthermore, NMR showed twenty-two carbons, a CH signal at Ô 76.51
for the methine (0-8) and a quaternary carbon at 5166.49, which are in favour
of the first hypothesis. long range heterocorrelation NMR experiment
showed a correlation between the singlet at 5 2.4 (in ODOy and the carbon at
5 76.5 and 50.45, establishing that this methyl group is on the N-1 '. In addition, the methyl, displaying a signal at 5 2.97, showed the same correlation
observed in glomerulatine A. ROESY NMR experiment showed, apart from the
signals characteristic of the monomer similar to glomerulatine A a correlation
between the doublet at 5 3.78, for the proton on C-8 'a, and the multiplet at 5
7.01 -7.04, for the proton on 0-4 indicating that this proton is in position a to the
six members ring. Therefore it can be concluded that the isolated compound
is the 8 '-8 'a dihydro glomerulatine A, named as glomerulatine 0.
It was decided that ^H-^% long range heterocorrelation of glomerulatine 0
may help to distinguish between the two possible isomers, caiycanthine or iso-
calycanthine. Since, the proton on 0-8’a (5 3.78) may show a correlation to
three bonds to the 0-3 (5 31.7) if the alkaloid is of the calycanthine-type or to
the C-3’(5 34.02) if it is of the iso-calycanthine-type. The long range
heterocorrelation NMR experiment failed to show this correlation, but it is highly
probable that this was due to the fact that only 5 mg of alkaloid was available
for the experiment.
130

4.6 Isolation and Identification of Bioactive Compounds fromPIcramnia antidesma subsp. fessonia.
4.6.1 Extraction and separation procedure
Dried and powdered leaves (600 g) of P. antidesma subsp. fessonia were
extracted with MeOH by percolation. The extract was filtered and concentrated
to a gum (127 g) under vacuum, and partitioned between HgO and CHCI3, the
aqueous layer was subsequently extracted with BuOH.
The butanolic fraction (59 g) was submitted to CO (silica gel 60, 0.063-0.2
pm, Merck) using a glass column (4.5 x 75 cm) and CHCI3 , CHCl3 -MeOH (9:1,
6 :2 , 7:3) as eluent. Sixty fractions (~ 125 ml) were obtained, fractions 7 to 1 0
were washed with CHCI3 and crystallized from CHCI3 yielding 0.83 g of aloe-
emodin; fractions 18 to 27 were submitted to GO using CHCI3 , CHCI3 -M0 OH
(95:5, 9:1, 8:2) as eluent, yielding, after crystallization from CHCl3 -MeOH (9:1)
63 mg of picramnioside A; fractions 28 to 30 were submitted to GO using the
same eluent as for picramnioside A, yielding, after precipitation from MeOH-
GHGI3 (9:1), forming 0.43 g of amorphous powder of picramnioside B; fractions
31 to 35 were washed with MeOH and a residue soluble only in DMSO was
obtained, after repeated precipitations from DMSO by adding GHGI3 yielded
0.490 g of picramnioside G; whereas from fractions 36-41 after preparative TLG
using GHGl3 -EtOAc (1 :1 ) 2 mg of aloe-emodin anthrone were obtained.
Oxidative hydrolysis: 10 mg of picramnioside B and picramnioside G were
separately dissolved in 25 ml of 4N HGI containing 1 mg of FeG^, and heated
at 100 °G for 4 hrs. The cooled solution was extracted with GHGI3 ; the
chloroformic fraction was submitted to pTLG using GHGl3 -MeOH (9:1) as
solvent system. The isolated compound was identical to aloe-emodin on TLG
(GHGl3 -MeOH 9:1) and their NMR spectra were superimposable.
131

4.6.2 Identification of the isoiated compounds
4.6.2.1 Picramnioside A
4.6.2.1.1 Spectral data
UVX max (MeOH)nm; 221, 273, 302, 382 IR
t max (KBr) cm'^: 3440 (OH), 2950, 1725, 1640, 1620, 1600, 1300
FDMS
m/z (relative intensity %): [M+ Na]* 531 (1 00), [M]'’ 508 (12 ), [M-benzoate+ Na]'’
409 (18), benzoate 122 (7)
FABMS (Spectrum 39, Appendix I)
(glycerol/thioglycerol/TFA matrix) m/z (relative intensity %) [M+1]* 509 (17), 387
(54), 256 (46), 239 (26), 207 (40), 181 (53), 165 (37), 149 (34), 131 (34)
HRMS (FABMS)[M-benzoate] m/z calculated 387.0716 (CgoHigOg) found 387.0724. The
molecular ion was too small to be measured.
CD (Spectrum 40, Appendix I)
(MeOH, C= 86.5 pM) X nm (A e): 342.8 (+1.68), 314.2 (-0.027), 295.0 (-6.36),
239.0 (+1.70), 230.6 (-3.89), 221.2 (+14.4)
H NMR (Spectrum 41, Appendix I)
(DMSG-Dg, 400 MHz): 5 3.45-3.53 (m, 1 H, H-2 '), 3.67 (m, 2 H, H-3' and 4"),
3.85 (dd, Ji..io= 2.1, J i 2 .= 9.9 1H, H-1"), 4.03 (of, 2H, H-11), 4.65 (d, 1H, H-10),
4.91 (d, 1H, H-3'OH), 5.01 (d,1H, 4'OH), 5.20 (t, 1H, H- 1 1 OH), 5.42 (d, 1H, H-
20H), 5.62 (s, 1H, H-5'), 6.62 (s, 1H, H-2), 6.83 (s, 1H, H-4), 6.93 (d, 1H, H-7),
7.15 (d, 1H, H-5), 7.50-7.53 (dd, 2H, H-3"), 7.57-7.61 (dd, 1 H, H-6 ), 7.65-7.69
(dd, 1H, H-4"), 7.71-7.73 (dd, 2H, H-2"), 11.83 (s, 1H, H-1 ), 11.96 (s, 1 H, H-8 ).
long range heterocorrelation NMR experiment (Spectrum 43,
Appendix I)
proton (carbons) 2 (1a,11), 3 (2, 3), 4 (la, 2, 4a, 10, 11), 5 (6 , 7, 8 a, 10), 6
(5a), 7 (5, 6 ), 10 (la, 4, 4a, 5a, 8 a, 1'), 1' (4a), 5’ (V, 3', 5'C=0), 2" (3", 4",
5'C=0), 3" (1")
132

4.6.2.2 Picramnioside B
4.6.2.2.1 Spectral data
UVX max (MeOH) nm: 225, 260, 270, 300, 365 X max (MeOH + NaOH) 230, 265, 375, 390, 455 (sh)IR
0 KBr (cm'^): 3350 (OH), 2730, 1760, 1640, 1620, 1600, 1590, 1295, 1230
FABMS (Spectrum 45, Appendix I)
(glycerol/thioglycerol/TFA matrix) m/z (relative intensity %) [M +l^ 447 (48),
387 (16), 351 (46), 256 (100), 239 (39)
HRMS (FABMS)
lh^+^Y m/z calculated 447.1291 (C2 2 H2 2 O1 0 + 1 ) found 447.1299. [M-acetyl] m/z
calculated 387.0716 (C2 0 H1 9 O8 ) found 387.0722.
CD (Spectrum 46, Appendix I)
(MeOH, 0= 180 |iM) X nm (A e): 355.0 (+0.27), 336.2 (-0.02), 321.6 (-0.58),
308.6 (-0.004), 294.6 (+1.25), 272.0 (-0.003), 266.0 (-0.94), 240.4 (+0.11),
226.6 (-2.27), 210.8 (+2.43)
NMR (Spectrum 47, Appendix I)
(DMSO-6 , 400 m/z MHz) 61.17 (s, 3H, H-S'CH^), 3.34- 3.41 (m, 1 H, H-2 '), 3.46-
3.48 (m, 2H, H-3’ and 4"), 3.64-3.67 (dd, Jvio=2.17, Ji,2 =9 .7 , 1 H, H-1’), 4.58-
4.62 (m, 3H, H-11 and H-10), 4.85 ( , 1 H, H-3’0H), 4.94 ( , 1 H, H-4'OH), 5.32
(d, 1H, H-2’0H), 5.42 (s, 1H, H-5 ), 5.46 (t, 1 H, H-11 OH), 6 . 8 6 (d, 1H, H-7),
6.89 (s, 1H. H-2), 6.99 (d, 1 H, H-5), 7.07 (s, 1 H, H-4), 7.53-7.57 (dd, 1H, H-6 ),
11.85 (s, 1H, H- 1 OH), 11.89 (s, 1H. H- 8 OH). COSY-45 (Spectrum 48,
Appendix I), ^H- ^C heterocorrelation (Spectrum 49, Appendix I), ^H- ^C long
range heterocorrelation and ROESY (Spectrum 50, Appendix I) were used to
confirm these assignments.
133

CarbonPicramnioside
A B c
C- 1 161.38 (C)‘* 161.23 (O f 161.17 [C f
C-la 115.58 (C) 115.74 (C) 115.44(C)
C- 2 112.17 (CH) 113.03 (CH) 112.09 (CH)
C-3 152.64 (C) 151.84 (C) 152.29 (C)C-4 115.69 (CH) 118.41 (CH) 115.54 (CH)
C-4a 141.26 (C) 141.24 (C) 141.07 (C)
C-5 120.65 (CH) 118.21 (CH) 120.40 (CH)
C-5a 145.97 (C) 146.07 (C) 145.81 (C)
C- 6 135.8 (CH) 136.27 (CH) 135.43 (CH)
0-7 116.12 (CH) 115.74 (CH) 115.79 (CH)
C- 8 161.51 (C) 161.64 (C) 161.19 (C)C-8 a 117.17 (C) 117.15 (C) 116.90 (C)C-9 193.46 (C) 193.50 (C) 193.21 (C)C- 1 0 42.44 (CH) 42.74 (CH) 42.30 (CH)C- 1 1 62.12 (CHg) 62.59 (CHg) 62.25 (CHj)C-1 ' 81.56 (CH) 80.87 (CH) 80.65 (CH)c - 2 ’ 66.83 (CH) 66.72 (CH) 66.48 (CH)C-3’ 71.91 (CH) 71.73 (CH) 71.42 (CH)C-4’ 69.39 (CH) 69.30 (CH) 69.01 (CH)C-5’ 94.74 (CH) 93.74 (CH) 93.35 (CH)C-5’ (C=0 ) 163.36 (C) 167.96 (C) 167.71 (C)C-5’(CH3) - 20.46 (CHg) 2 0 . 1 0
C-1 ” 129.17 (C) - -
C-2 ” 129.22 (CH)® - -
C-3” 128.89 (C)® - -
C-4” 133.71 (CH) - -
“Chemical shifts are in 6 -values in ppm, measured in DMSO-Dg.‘ Multiplicity from Dept ^^0 NMR experiment.H"he intensity for this signal was twice that of the other CH signals.
Table 4.7 NMR Spectral Data° of the Compounds Isolated from P. antidesma subsp. fessonia.
134

4.6.2.S Picramnioside C
4.6.2.3.1 Spectrai data
UV
X max (MeOH) nm: 225, 261 (sh), 270, 300, 365 iR
ù (KBr) cm ': 3500, 3390, 2990, 2920, 1725, 1640, 1620, 1600, 1300, 1250.
FABMS (Spectrum 51, Appendix I)
(glycerol/thioglycerol/TFA matrix) m/z (relative intensity %) [M+1]* 447 (63),
387 (48), 351 (26), 256 (100), 239 (40)
HRMS
[M+1]* m/z calcuiated 447.1291 (CjjHjjO,» +1) found 447.1302. [M-acetyl] m/z
calculated 387.0716 (CgoH^O,) found 387.0712.
CD (Spectrum 52, Appendix I)
(MeOH, 0= 85 pM) X nm (A e): 351.4 (+0.78), 317.0 (-0.019), 294.0 (-5.37),
268.8 (+0.009), 253.0 (+1.1), 230.0 (-3.13), 210.6 (+6.47)
'H NMR (Spectrum 53, Appendix I)
(DMSO-Dg, 400 MHz) 5 1.72 (s, 3H, H-5'CH,), 3.38-3.44 (m, 1 H, H-2'), 3.54-
3.57 (m, 2H, H-3’ and 4'), 3.67-3.70 {dd, J,.,o=2.0, J,..2=9.85, 1H, H-1 '), 4.51-
4.63 (m, 3H, H-10 and H-11), 4.85 (d, 1H, H-3'OH), 4.93 (d, 1 H, H-4'OH), 5.38
(d 1H, H-2'OH), 5.40 (s, 1 H, H-5 ), 5.48 (f, 1 H, H- 1 1 OH), 6.80 (s, 1H, H-2),
6.91 (d 1H, H-7), 6.98 (s, 1 H, H-4), 7.13 (d 1 H, H-5), 7.55-7.59 (dd, 1H, H-6 ),
11.84 (s, 1H, H- 1 OH), 11.93 (s, 1 H, H- 8 OH)
'H-'^C long range heterocorrelation NMR experiment (Spectrum 55,
Appendix I)
proton (carbons) 1 OH (1 , 2 ), 2 (1, la, 4, 1 1 ), 4 (2, la , 1 0 , 1 1 ), 5 (1 0 ), 6 (8 ,
5a), 7 (5, 6 , 8 , 8 a ) , 8 OH (8 , 8 a) 10 (1, la, 4a, 5a, 8 a, 1'), 11 (2, 3), T (2 '), 2'
(3 ), 5' (V, 3'), 5'CH, (5'C=0)
135

4.6.2.4 Aloe-emodin
4.6.2.4.1 Spectral data
UV
X max (MeOH) nm: 224, 256, 277, 287 X max (MeOH+NaOH) nm: 232, 247, 292 (sh), 330 EIMS (Spectrum 57, Appendix I)
m/z (relative intensity %) [M]" 270 (1 0 0 ), 214 (85), 224 (9), 213 (1 1 ), 196 (6 ),
168 (10), 139 (22), 128 (10), 121 (15)
HRMSm/z calculated 270.0528 (CigHioOg); found 270.0536
NMR (Spectrum 58, Appendix I)
(DMSO-6 , 400 MHz) 5 4.59 (s, 2H, H-1 1 ), 5.59 (br s, 1 H, OH-1 1 ), 7.19 (d,
J=0.9,1H, H-2), 7.28-7.30 (dd, J=8 .2 , 0.86, 1 H, H-7), 7.56 (d, J=0.9, 1H, H-4),
7.56-7.60 (dd, J=7.6, 0.86, 1 H, H-5), 7.73-7.74 (dd,J= 8 .2 , 7.6, 1 H, H-6 ), 11.83
(br s, 2H, H-1, H-8 )
'"C NMR
(DMSO-6 , 400 MHz) Ô 161.31 (C-1 ), 114.13 (C- 1 a), 117.03 (C-2 ), 153.69 (C-3),
119.26 (C-4), 132.87 (C-4a), 120.57 (C-5), 133.07 (C-5a), 137.27 (C-6 ), 124.30
(C-7), 161.61 (C-8 ), 115.59 (C-8 a), 191.45 (C-9), 181.09 (C-1 0 ), 62.04 (C-1 1 )
4.62.5 Aloe-emodinanthrone
4.6.2.5.1 Spectral data
EIMS (Spectrum 59, Appendix I)
m/z (relative intensity %) [M]+ 256 (100), 238 (32), 226 (31), 210 (32), 197
(24), 181 (12), 165 (7), 152 (16), 139 (15), 127 (16), 115 (10)
'H NMR
(CD3OD, 400 MHz) Ô 4.44-4.55 (q, J=14.7,11.2,1H, H-11), 4.64 (s, 2H, H-10),
6.06 (s, 1H, H-2), 6.77 (d, J=7.4,1H, H-7), 6.84 (s, 1 H, H-4), 6.89-6.91 (dd, J=
8.3, 0.7, 1 H, H-5), 7.48-7.52 (dd, J= 8.3, 7.4, 1 H, H-6 )
136

NMR
(CD3OD, 400 MHz) 5 163.15 (C-1), 116.66 (C-1 a), 114.90 (C-2), 152.35 (C-3),
117.93 (C-4), 142.12 (C-4a), 119.44 (C-5), 143.72 (C-5a), 137.35 (C-6 ), 121.42
(C-7), 163.70 (C-8 ), 118.55 (C-8 a), 193.47 (C-9), 57.52 (C-10), 64.38 (C-1 1 )
4.6.3 Discussion
A bioactivity guided fractionation of the methanolic extract from P.
antidesma yielded two known anthraquinones, aloe-emodin and aloe-emodin
anthrone. Figure 4.8 (p. 138), and three new aloe-emodin C-glycosides, named
picramnioside A, Figure 4.9 (p. 139), picramnioside B and picramnioside C,
Figure 4.10 (p. 140). This plant was selected because of its activity against P.
gallinaceum (Spencer et al., 1947) and from the chemotaxonomic stand point
the presence of quassinoids was predictable. Only the butanolic fraction (see
Table 3.15, p.8 8 ) proved to be active against KB cell (LC5 0 8.7 pg/ml), however,
this fraction showed no significant activity against brine shrimp (LC5 0 485 ng/ml)
(see Table 3.13, p.8 6 ).
The UV spectrum of picramnioside A showed four bands (221, 273, 302,
and 382 nm) characteristic of highly conjugated system, such as
anthraquinones and the IR spectrum showed a band at 1725 cm '' for a ketone
group. The FDMS showed a strong peak at m/z 531 (100) [M+Na]^ and m/z
508 (12) for [M] , and FABMS (glycerol/thioglycerol/TFA matrix) showed a weak
[M+1 ] peak at 509 (17). The peaks at m/z 387 (54) indicated the loss of a
benzoate moiety [M-1 22]'’ and at m/z 256 indicated the aglycone. Figure 4.11
(p. 142) shows the mass spectra fragmentation of picramniosides A and C.
HRMS (FABMS) showed a [M-benzoate]"^ m/z 387.0724 corresponding to the
molecular formula CgoHigOg, the molecular ion at 509 was too small to be
measured using this technique.
^H NMR of picramnioside A showed two singlets at S 11.96 and 11.83
137
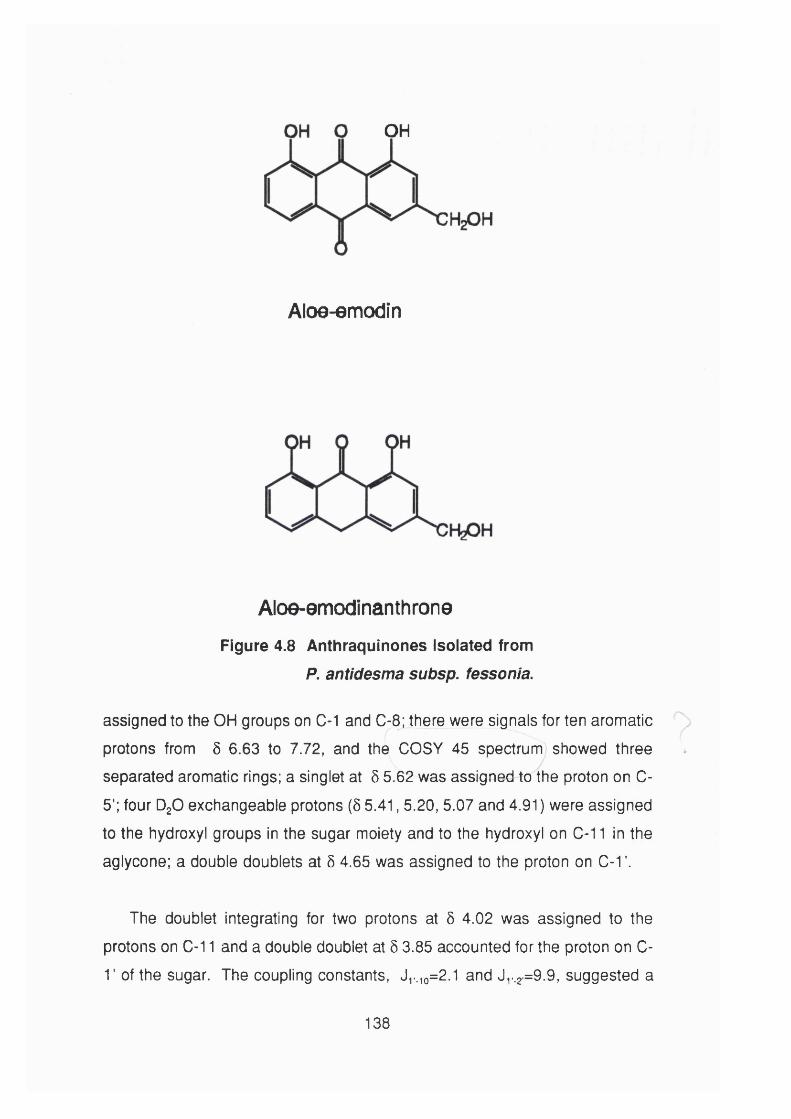
OH
Aloe-emodin
Aloe-emodinanthroneFigure 4.8 Anthraquinones Isolated from
P. antidesma subsp. fessonia.
assigned to the OH groups on C-1 and C-8 ; there were signals for ten aromatic
protons from 5 6.63 to 7.72, and the COSY 45 spectrum showed three
separated aromatic rings; a singlet at 5 5.62 was assigned to the proton on C-
5’; four DgO exchangeable protons (5 5.41, 5.20, 5.07 and 4.91) were assigned
to the hydroxyl groups in the sugar moiety and to the hydroxyl on C- 1 1 in the
aglycone; a double doublets at 5 4.65 was assigned to the proton on C- 1 ’.
The doublet integrating for two protons at 5 4.02 was assigned to the
protons on C- 1 1 and a double doublet at 5 3.85 accounted for the proton on C-
T of the sugar. The coupling constants, Ji._io=2.1 and Jy.2 =9 .9 , suggested a
138

Picramnioside A
Figure 4.9 C-Glycoside Isolated from P. antidesma subsp. fessonia
Showing the Most Relevant ROESY Effects.
p configuration for the carbon of the sugar bound to the aglycone, close to those reported for aloins (Manitto et al., 1990): the two proton singlet-shaped
multiplet at Ô 2.90 was assigned to the protons on C-3’ and 0-4’ and a multiplet
at Ô 3.53-3.44 accounted for the protons on C-2 ’.
The ^ C nmr assignment is presented in Table 4.7 (p. 134), and there are 25
signals. Two CH signals at 5 129.22 and 133.7 showing a double intensity
characteristic of a monosubstituted benzene ring, an interesting feature of this
C-glycoside in 1 NMR is the low field position of the signal for C-5’ (Ô 94.74),
which may indicate the presence of an 0-glycoside. In fact, this sugar residue
has the same configuration of xylose indicated by ROESY NMR experiment,
but, it has the aloe-emodinanthrone moiety attached to the C- 1 and theI
benzoate is attached to C-5 of the xylose moiety.
2D NMR experiments such as COSY 45, ^H- ^C heterocorrelation, ^H- ^C long
range heterocorrelation and ROESY were used to confirm these assignments.
139

OH
OH
8 OH
Picramnioside B
OH
OH
OHlO H
Picramnioside CFigure 4.10 Picramniosides B and C isoiated from P. antidesma
subsp. fessonia Showing the Most Reievant ROESY Effects.
140

The long range correlation NMR experiment showed that the singlet at
Ô 5.62 corresponding to the proton on C-5' and the double doublet at 5 7.73-
7.65 assigned to the proton on C-2” , were correlating to the carbon at 5 163.36
assigned to the carbonyl of the ester, thus establishing the position of the
benzoate on C-5’. Also, the signal at 5 4.65 for the proton on C- 1 0 showed a
correlation to the carbon at Ô 81.56 for the C-1 ’ corroborating the position of
attachment between the aglycone and the sugar moiety.
In addition, the ROESY NMR experiment showed a correlation between the
signal at 5 5.62 for the proton on C-5’ and the signals at 5 3.85, 3.67 and 5.01
for the protons on C-1’, C-3’ and the OH on C-4’, respectively; indicating that
all the substituents on the sugar moiety are in the equatorial position.
Moreover, the proton on C -1 ’ ( 6 3.85) and the signal for the proton on C-4 (5
6.83) showed correlation, and the multiplet at Ô 3.45-3.53 (H-2’) and the doublet
at 5 7.15 (H-5). Therefore the configuration of the C- 1 0 is R, as previously
reported for aloins (Manitto et al., 1990) and cascarosides (Manitto et al.,
1993). Figure 4.9 (p. 139) shows the ROESY effects displayed for
picramnioside A, B and C and their absolute configuration. Also, the Circular
dichroism agreed with that previously reported for (10R) aloin (Manitto et al.,
1990).
The UV spectra of picramnioside B and picramnioside C were similar and
the IR spectra showed little differences the carbonyl band which appear at 1725
cm* in the spectrum of picramnioside B and at 1760 cm' for picramnioside C.
Oxidative hydrolysis (Fairbairn and Simic, 1963) of both compounds, yielded
aloe-emodin, which was identified by comparison on TLC and ^H NMR with a
reference sample. FABMS (glycerol/thyoglycerol/TFA matrix) showed, for both
picramnioside A and picramnioside B, a [M+1]^ peak at m/z 447, a peak at m/z
387 indicating the loss of the acetate of the sugar and at m/z 256 for the
aglycone. HRMS showed a [M+1]* at m/z 447.1299 for picramnioside B, and
at m/z 447.1302 for picramnioside C, both corresponding to the formula
C2 2 H2 2 O1 0 .
141
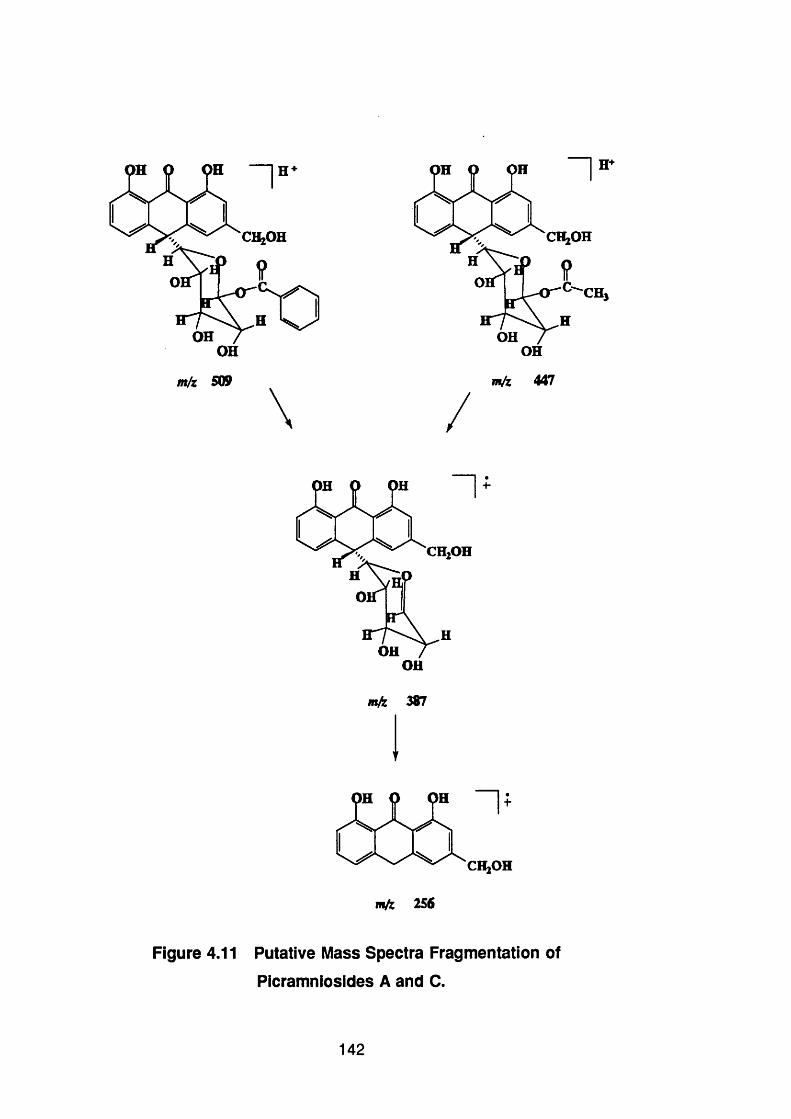
H |H +
C&OH
tnlz 509
Cft
OHOH
mji 447
OHOH
mfi 387
H n :
Cl^OH
mil 256
Figure 4.11 Putative Mass Spectra Fragmentation of
Picramniosides A and C.
142

NMR showed, in both cases, only two aromatic systems indicating a
difference with respect to picramnioside A; instead, both picramnioside B and
picramnioside C showed a methyl group signal at 6 1.7 for an acetyl group,
suggesting that picramnioside B and picramnioside C are isomers and have an
acetyl substituent at the 5’ instead of the benzoate substituent as in
picramnioside A. The NMR of picramnioside B and picramnioside C were
only differentiated in the aromatic pattern and the displacement of the signals;
picramnioside B showed, from 5 6.86-7.07, a doublet-singlet-doublet-singlet
pattern, while, picramnioside C showed, from 5 6.80-7.13, a singlet-doublet-
singlet-doublet pattern.
NMR showed little differences between these two compounds,
suggesting that they are the C- 1 0 isomers; and this was apparent when, both
compounds, after a few hours in solution (DMSO-g) were converted into each
other. ^H-^% long range heterocorrelation NMR experiments, for both
compounds, showed, as important features, a correlation of the signal at 6 1.7
(methyl) with the carbon at 5 167 for the carbonyl group; the signal at 6 4.6 for
the proton on 0-10 correlated to the signal for the protons on C-1 ’, C-la, C-4,
C-4a, C-5, C-5a and C-8 a. Furthermore, ROESY NMR experiment, as in
picramnioside A, indicated the same sugar configuration and, as previously
reported for aloins and cascarosides (Manitto et al., 1990; Manitto et al., 1993).
The nOe effect showed by the proton on C-1 ’ and C-2 ’ helps to differentiate
between the R and 8 isomers; picramnioside B showed an interaction between
the proton on C- 1 ’ (5 3.64-3.67) and the doublet at 6 6.99 for the proton on C-5 ,
whereas the proton on C-2 ’ (Ô 3.34-3.41) interacted with the singlet at 5 7.07
for the proton on C-4, therefore the absolute configuration of the C-10 in
picramnioside B is S. On the other hand, picramnioside C showed, in the
ROESY NMR experiment, a correlation between the signal at 5 3.67-3.70 (C- 1 ’)
and the singlet at Ô 6.98 for the proton on C-4, whereas, the signal proton at
Ô 3.38-3.44 (C-2 ’) correlated to the doublet at 5 7.13 for the proton on C-5 ,
showing an R configuration at C- 1 0 in picramnioside C.
143

The circular dichroism spectra, of picramnioside B and picramnioside C
showed that they were similar to that reported for (1 0 S) aloin and (10R) aloin
respectively, establishing, thus, the absolute configuration of picramnioside B
as (10S) and of picramnioside C as (10R).
Aloe-emodin was identified for its spectroscopic properties (UV, HREIMS,
and NMR) and by comparison of the NMR with a commercial sample
(Apin Chemical, UK). Aloe-emodinanthrone was characterized using EIMS,
and NMR and by comparison of previously reported values (Rychener and
Steiger, 1989).
4.6.4 Biological Activity of the isolated Compounds
Picramniosides A, B, and 0 were tested against KB cells and the result is
shown in Table 4.8 (p. 144). Picramnioside A was some four times less active
than the acetyl isomers, picramniosides B and 0 and, although, picramniosides
B and 0 are chiral isomers appear to have the same value, may be, because
they are converted into each other in solution.
CompoundsKB ceils
LCso±sd (pM/ml)
Picramnioside A 35.7±0.10 (2)®
Picramnioside B 8.74±2.40 (3)
Picramnioside 0 8.32±2.47 (3)
Emetine 1.50±0.56 (3)
^Number of tests performed, in triplicate.
Table 4.8 Activity of the Compounds Isoiated from
P. antidesma subsp. fessonia Against KB Cells.
144

SECTION 5 GENERAL DISCUSSION

5.1 Introduction
Panama is a small Central American country, with a level of poverty
estimated at 33.6 % and with a disparate health care coverage between the
rural and urban area. The Panamanian government spent US $ 64.97 millions
In drugs, In 1993, and all of these drugs were imported from developed
countries. Panama has no plans to Include folk medicine in health services,
as the World Health Organisation has urged developing countries to do.
Nonetheless, herbs are sold In public markets and In drug stores without any
general policy, guidance on the use or quality control of herbal medicines.
There are more than 200,000 (about 9 % of the total population)
Amerindians, today In Panama, distributed In five groups, which live in the
forest using plants as the major. If not the unique, source of medicine for curing
their ailments. Although, there have been two major attempts to compile the
ethnobotanlcal Information amongst these Amerindian groups, more surveys are
urgently needed to rescue all their knowledge before it become extinct, since
the youngsters are moving to the cities and leaving their culture behind.
More than 200 plants species have been reported to be used as medicines
(Joly et al., 1987; Joly et al., 1990; Gupta et al., 1993a) amongst the Guaymi
and Kuna Indians. Nevertheless, little or nothing has been done to scientifically
evaluate these plant species, which may yield effective drugs that can be
Introduced Into the Panamanian health care system.
The flora of Panama Is one of the richest In the world (Schultes, 1972) with
more than 8 , 0 0 0 plant species and more than one half million voucher
specimens being available worldwide (Dwyer, 1964; Dwyer, 1985). However,
little Is known about the chemistry and biological activity of these plant species.
Although 27.5 % (2,077,914 hectares) of the Panamanian land surface Is
under conservation by law, the annual rate of deforestation is estimated at
146

65,000 hectares. Therefore, it is vital to study this flora before the species
become extinct.
5.2 Plants selected for this study
The plants investigated in this study were selected using chemotaxonomic
criteria at different taxonomic ranks to limit the range of plants. For the
Meliaceous, Menispermaceous and Simaroubaceous plants the family rank was
used. Thus, Guarea and Ruagea belongs to the family Meliaceae which is
known to contain limonoids (Connolly, 1983) and some of them are active
against P. falciparum (Khalid et al., 1986; Bray et al., 1990).
Abuîa belongs to the family Menispermaceae which produces bbiq alkaloids
with a variety of pharmacological activities (Schiff, 1983; Schiff, 1987). Some
of these alkaloids show more activity against drug resistant strains of P.
falciparum (Ye and Van Dyke, 1989) and multidrug resistant cancer cell lines
(Shiraishi et al., 1987) than against the drug sensitive strains. The genus
Picramnia belongs to the family Simaroubaceae, which is known to contain a
bitter group of compounds, named quassinoids, some having activity against
the malaria parasite (e.g. O’Neill et al., 1986). Both, P. antidesma subsp.
fessonia and P. teapensis have bitter tastes, suggesting the presence of
quassinoids in these species.
On the other hand, for the species of Cepfiaëlis, investigated in this study,
the rank of genus was used. Cephaëlis ipecacuanha is the commercial source
of emetine, which is effective in the treatment of severe amoebic dysentery.
Until recently, knowledge of the chemistry of Cephaëlis species was limited to
C. ipecacuanhavfh\ch contains monoterpenoid isoquinoline alkaloids (Wiegrebe
et al., 1984; Itoh et al., 1989; Itoh, et al., 1991 ; Nagakura, et al., 1993) and to
C. stipuiacea which contains gramine (Yulianti and Djamal, 1991), an indole
alkaloid. Since this study started two Cephaëlis species from Panama have
been studied in coordinated work by the Centre for Pharmacognostic Research
147

(CIFLORPAN) at the University of Panama. C. correae yielded the known iso-
dolichantoside, three new indole monoterpenoid alkaloids, and one isoquinoline
monoterpenoid glucoside alkaloid (Achenbach et al .,1993) and C. axillaris
yielded two known indole monoterpenoid alkaloids, vallesiachotamine and 6 ’-
trans-feruloyl-lyaloside (Gupta et al., 1993b).
5.3 Testing for biological activity
5.3.1 Brine shrimp assay
A new microwell method for brine shrimp testing has been developed in the
present work and this overcomes some of the disadvantages of the test tube
method, previously reported (Meyer et al., 1982). Requiring smaller amount of
sample and the employment of microplate technology facilitates the testing of
larger number of samples and dilutions.
Although, the use of brine shrimp has been proposed as a general
bioassay, detecting a wide variety of pharmacological activities (McLaughlin,
1991), compounds with only cytotoxic activity were detected in this work. In
general, drugs acting on the nervous system, such as atropine, caffeine,
ephedrine and nicotine were not active. It has been suggested that a brine
shrimp test could be used in the evaluation of traditional medicines
(Kyerematen and Ogunlana, 1967), nonetheless, the present study clearly
indicates that this assay is predictive of cytotoxicity, therefore, medicinal plants
containing only cytotoxic compounds would be detected. Plants containing
compounds with different pharmacological activities would not be evaluated
properly by a brine shrimp assay.
Brine shrimp may be useful to guide the fractionation of plant extracts
containing a group of compounds that is known to be active in this bioassay.
This was demonstrated using quassinoids, which were active against KB cell
as well as against P. falciparum. Thus, plant extracts from species of
Simaroubaceae showing activity against brine shrimp are likely to contain
148

cytotoxic quassinoids with activity against P. falciparum.
5.3.2 Biological activity of plant crude extracts
Extract from Guarea macropetala, G. rhopalocarpa and Ruagea glabra
(Meliaceae) showed strong activity against both brine shrimps and KB cells,
while extracts from Abuta dwyerana (Menispermaceae) showed only weak
activity in these two bioassays. Amongst the Cephaëlis species, the
chloroformic extract from the roots 0. camponutans showed strong activity
against brine shrimps and KB cells. Based on this result it was selected for
bioassay guided fractionation in order to isolate the active principles.
Although some extracts from P. antidesma subsp. fessonia showed weak
activity against brine shrimp, the butanolic fraction from the leaves was the
most active of all the extracts tested against KB cell and was submitted to a
bioassay-guided fractionation using KB cell assay.
5.4 Phytochemistry
5.4.1 Cephaëlis species
A bioassay-guided fractionation of the woody part of 0. camponutans,
(accepted name Psychotria camponutans) yielded a novel
benzoisochromanquinone and a 2 -aza-anthraquinone isolated for the first time
from the plant kingdom. C. dichroa extracts showed strong reaction with
Dragendorff’s reagents and five indole monoterpenoid alkaloids were isolated,
including a novel vallesiachotamine lactone, whereas, C. glomerulata, (accepted
name Psychotria glomerulata) yielded three new non iridoid quinoline alkaloids,
named, glomeruiatine A, B and C. Interestingly the expected emetine-type
alkaloids were not found in these three species.
The major alkaloids of the Rubiaceae are the indole monoterpenoid
149

alkaloids, derived via condensation of tryptamine and secologanin to yield
strictosidine, which is the precursor of the whole range of indole monoterpenoid
alkaloids. One exception to this biosynthetic pathway occurs in C. ipecacuanha
where dopamine condenses with secologanin to yield isoquinoline alkaloids of
the emetine type. Figure 5.1 (p.151) summarises the biosynthetic pathway of
indole and isoquinoline monoterpenoid alkaloids. The findings, in the present
work, clearly indicate that these biosynthetic pathways are not necessarily a
characteristic of the genus as a whole, since the three Cephaëlis species
studied do not contain isoquinoline-iridoid alkaloids, and only one of the three
species contained tryptamine-iridoid alkaloids.
The genus Cephaëlis belongs to the tribe Psychotriae and the major
alkaloids found throughout this tribe are of the polyindolenine-type, which are
derived via polymerisation of tryptamine. Also non-iridoid quinoline alkaloids of
the calycanthine-type have been found In Psychotha species (Adjibade et al.,
1992), being derived from two molecules of tryptamine. Figure 5.2 (p. 153)
shows the biosynthetic pathway of polyindolenine and calycanthine-type
alkaloids.
The taxonomic position of Cephaëlis species is unclear. Steyermark, in
1972 and 1974, grouped the Cephaëlis species within Psychotha.
Nevertheless, Dwyer in 1980 and Standley in 1985 retained Cephaëlis as a
genus. More recently, however, Cephaëlis species have been included in
Psychotha, subgenus Heteropsychotria (Taylor et al., 1994) and they have been
combined mostly because both of them have tiny white flowers and the species
of both are numerous and difficult to separate (Taylor, 1993).
Cephaëlis, which comprises some 2 0 0 species (Dwyer, 1980), was
originally distinguished by its capitate inflorescences and relatively large floral
and inflorescence bracts (Taylor, 1991 ). These plants are not clearly separable
from Psychotha, however, and in addition apparently represent a widely
polymorphic assortment of species with similar inflorescence structures
150
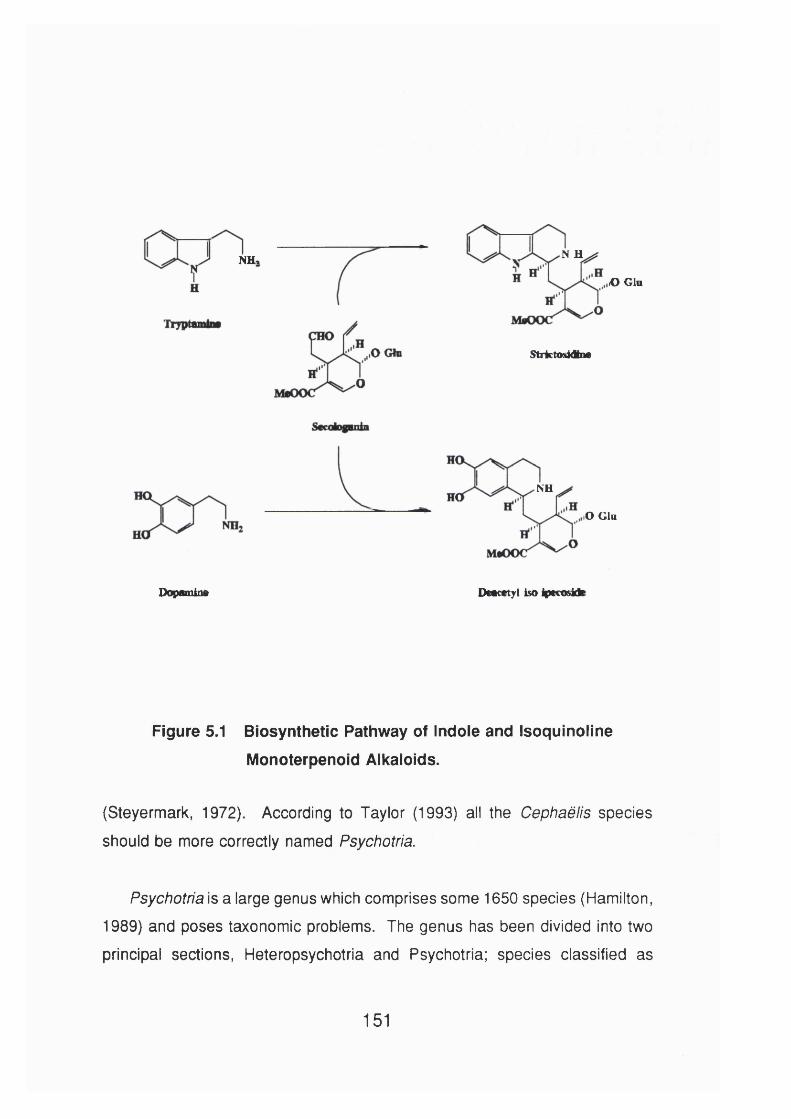
NH, NH
O Gla
Strktoüidine
NH
lO Gin
Dopttmin» Dcacctyi iso iptcoside
Figure 5.1 Biosynthetic Pathway of indole and isoquinoiine
Monoterpenoid Alkaloids.
(Steyermark, 1972). According to Taylor (1993) all the Cephaëlis species
should be more correctly named Psychotria.
Psychotha is a large genus which comprises some 1650 species (Hamilton,
1989) and poses taxonomic problems. The genus has been divided into two
principal sections, Heteropsychotria and Psychotria; species classified as
151

Cephaëlis have been assigned to the subgenus Heteropsychotria (Taylor,
1993). The subgenus Psychotria is pantropical and includes most of the
African species and many Asian and Pacific species. These species are
separated by its brown or grey drying colour, usually red fruits and deciduous
stipules leaving a line of well-developed colleters (Taylor, 1994). Whilst, the
Heteropsychotria species are neotropical and are characterized by green or
grey-green drying colour, stipules usually persistent at least on the distal 3-6
nodes and bilobed or bidentate and not leaving a ring of colleters, and fruit blue
or black (sometime passing through a red or orange intermediate stage)
(Taylor, 1994). "In this group fall most (though not all) of the species formerly
placed in Cephaëlis" (Taylor, 1994).
If the alkaloid content of the both subgenera Heteropsychotria (former
genus Cephaëlis) and Psychotria (former genus Psychotria) is taken into
account as a chemotaxonomic character, two major groups may be clearly
separated (Figure 5.3, p. 155). The first group producing isoquinoline or indole
monoterpenoid alkaloids, including C. ipecacuanha, C. dichroa, C. correae and
C. axiiiaris. A second group producing non-iridoid alkaloids, such as
polyindolenine and quinoline alkaloids, including C. giomeruiata, P. forsteriana
and P. oieoides. Within the first group two sub-groups may be differentiated,
those producing isoquinoline alkaloids as C. ipecacuanha and C. correae, and
those producing indole alkaloids as C. dichroa, C. axiiiaris and C. correae. As
an interesting feature, C. correae produces both isoquinoline and indole
monoterpenoids alkaloids, therefore other taxonomic characters should be used
to establish the relationship with one of the sub-groups. Fig 5.3 shows the
biosynthetic relationships of the alkaloids produced in sub-genera
Heteropsychotria and Psychotria.
Although C.camponutans contains a nitrogen-containing anthraquinone, it
is not possible to situate it within this scheme, since the biosynthetic pathway
to build up this 2 -aza-anthraquinone is unknown, but this species produces
anthraquinones which are characteristic of the Rubiaceae as a whole.
152

Tycoon
L - Tryptophan
rCH,
N'-M«thyl Tryptamine Indolenh» r«dlcal
Ç'HpNHj
N -H
H-
CEf H
Iso- Calyuvithlnt CaiycamtMn# CMmomanthina
Figure 5.2 Putative Biosynthetic Pathways of Non-iridoid
Alkaloids of the Quinoline- and Poiyindoienine-Type
Typical of the Tribe Psychotriae.
153

On the other hand, it does not mean, necessarily, that C. camponutans is not
closely related to other species of the sub-genus Heteropsychotria or
Psychotria. Since the production of alkaloids needs a whole block of active
genes and if only one is affected the plant is not able to produce them.
Although, perhaps the other genes are identical. That is why the presence of
a taxonomic character is likely to be more important for taxonomic purpose than
its absence (Cronquist, 1968). Interestingly enough, is the fact that, M.
Nepokroeff (Department of Botany, University of Wisconsin), in a unpublished
work has shown that C. camponutans should be included in the subgenus
Notopleura within the genus Psychotria or maybe in a separate genus
Notopleura not closely related to Psychotria at all (Taylor, 1994). This group
of plant is recognized by a low, herbaceous, succulent, usually unbranched
habit, pseudoaxillaris inflorescences, and strange stipules that are succulent
and often deciduous (Taylor, 1994).
The presence of non-iridoid alkaloids in C. giomeruiata may indicate that
this species should be better placed in the subgenus Psychotria, unless other
taxonomic characters do not indicate so. Cephaëlis species produce different
types of alkaloids, however, little is known about the chemistry of this large
group of plants, now within Psychotha, and more work is needed to know if, a
posteriori, alkaloids are of value as taxonomic characters.
Since Cephaëlis species produce indole and isoquinoline alkaloids and not
the polyindolenine alkaloids characteristic of the tribe Psychotriae, it indicates
that Cephaëlis species are different from other genera of Psychotriae.
Nonetheless, genera should be defined in such way that they can be
recognised without recourse to technical characters not readily visible to the
naked eye (Cronquist, 1968). Consequently, since the alkaloidal content can
not be easily examined, Cephaëlis species may be kept as member of
Psychotria, in the sub-genus Heteropsychotria; unless a visible character is
correlated to this group of plants.
154

DTI.Dopamine
Me
Me
OMe
OMe
Isoquinoiine monoterpencid alkaloids as those produced by
C ip eca cuan ha , C, co rre a e...OGIu
MI2
Tryptamine
H3Q
CH3
Colycanthine alkaloids fhwn C. g lom erukU a , P . fo rs te r ia n a
> 3500 Indole-monoterpenoid alkaloids characteristic of Apocynaceae, Loganiaceae and Rnbiaceoe. C.dtchroa,C. axUaris, C. correae
C tt
Polyindobnine Alkaloids (non-iridoidaD
from Psychotria species
Figure 5.3 Biosynthetic Reiationship of the Aikaioids Produced in
Sub-Genera Heteropsychotria and Psychotria.
155

5.4.2 Picramnia antidesma subsp. fessonia
P. antidesma subsp. fessonia was selected for this study expecting to
isolate quassinoids with activity against P. falciparum. As the activity of the
quassinoids against P. falciparum and KB cell parallels the activity against brine
shrimp (see Table 3.3, p.8 8 ), it was decided to guide the fractionation using KB
cell and brine shrimp. Nevertheless, only the butanolic fraction proved to be
active against KB cell, but this fraction showed no significant activity against
brine shrimp.
The butanolic fraction was submitted to CC yielding two known
anthraquinones, aloe-emodin and aloe-emodinanthrone, and three new aloe-
emodin C-glycosides, named Picramnioside A, B and C. Previous work with
Picramnia (Leon, 1975; Popinigis et al., 1980; Arana and Juica, 1986) based
their search for quinones on oxidative hydrolysis, therefore missing these C-
glycosides.
An interesting feature of picramniosides is the that sugar, which has the
configuration of xylose, contains an acetate or benzoate attached to position 5’.
It is difficult to explain from the biosynthetic point of view, since a reactive group
may be expected on position 5' of the sugar to react with acetate or benzoate,
but, as far it is known, there is no known xylose substituted with reactive
groups, such as hydroxyl, on this position.
In addition, the picramniosides showed strong activity against KB cell,
whereas, the aglycone, aloe-emodin per se, is inactive against this cell line.
Hence, it is important to continue working on this group of plants to follow up
this rare group of C-glycosides and to evaluate them in different biological
assays.
156

5.5 Are Plants Predictable In their Chemotaxonomy ?
Chemotaxonomy is the use of chemical evidence for classification of plant
species and this is because a secondary metabolites present in plant cells
represent not only a chemical entity, but are the manifestation of a whole series
of enzymes which in turn are the genetic expression. The systematic position
of the species is of excellent predictive value concerning the existence of useful
chemicals (Gottlieb, 1982). Then related plant species may produce similar or
related secondary metabolites and this criteria has been used to select plants
for chemical studies (see Section 1 .2.3). Thus, species from Papaveraceae are
expected to contain isoquinoiine alkaloids, species from Rubiaceae are
expected to contain indole-monoterpenoid alkaloids and quinones, and species
from Menispermaceae are expected to contain bbiq alkaloids.
Cephaëlis species were selected for this study expecting emetine analogues
to be present and P. antidesma subsp. fessonia anticipating quassinoids.
Although, compounds with activity against brine shrimp, KB cell and P.
falcipamm were present none of the anticipated type of compounds were found.
Instead, each Cephaëlis species yielded different compounds, namely, indole
monoterpenoids alkaloids, aza-anthraquinone, benzoisochromanquinone and
quinoline alkaloids; and P. antidesma subsp. fessonia yielded three new C-
glycosides. Table 5.1 (p. 158) shows the compounds expected and those
actually isolated from the plants selected for this study.
Some researchers do not investigate particular plant species because they
are prejudged to contain compounds which are either too toxic or not very
active on the basis of their chemotaxonomic relationship to a species which has
previously been investigated. However, this result indicates that taxonomically
related plants may yield and offer new leads in the discovery of useful drugs.
It is estimated that only about 1 0 % of plants have been investigated for
pharmacologically active principles (Farnsworth and Morris, 1976), this figure
157

reflects that little is still known to predict with certainty what kind of compounds
or pharmacological activities may have some group of plants.
Plant species Anticipated Found
Cephaëlis Emetine No emetine
(Rubiaceae) analogues Indole alkaloids
Aza-anthraquinone
Quinoline alkaloids
Picramnia Quassinoids Anthraquinone
(Simaroubaceae) C-glycosides
Table 5.1 Compounds Expected and those Found In the Plants
Selected for this Study.
158

SECTION 6 CONCLUSION

1.- A new microwell method for brine shrimp has been developed that
overcomes some of he disadvantages of the previous method.
2 .- Only cytotoxic compounds appear to be active against brine shrimp,
while compounds acting on the nervous system are inactive.
3.- The activity of the quassinoids against Plasmodium falciparum and KB
cell parallel the activity against brine shrimp.
4.- There are some 200 species of Cephaëlis occurring throughout the
tropics of which 2 1 occur in Panama. Until recently, knowledge of the
chemistry of Cephaëlis was limited to C. ipecacuanha which contains
monoterpenoids isoquinoiine alkaloids and to C. stipulacea which
contains gramine.
5.- Cephaëlis camponutans yielded benz[g]isoquinoline-5,10 -dione for the
first time isolated from the plant kingdom and a new
1-hydroxibenzoisochromanquinone. Both compounds showed activity
against brine shrimp, KB cell and P. falciparum.
6 .- Cephaëlis dichroa yielded a new indole alkaloid, vallesiachotamine
lactone, together with four known indole alkaloids, vallesiachotamine,
angustine, strictosidine lactam and strictosidine.
7.- Cephaëlis giomeruiata yielded three new quinoline alkaloids, named
glomerulatine A, B and C, but none of the spectroscopic techniques
used in the structure elucidation distinguished between calycanthine or
iso-calycanthine-type. Only X-ray crystallography would unravel this
problem.
8 .- Since this study started two Cephaëlis species from Panama have been
studied in coordinated work by the Centre for Pharmacognostic
160

Research (CIFLORPAN) at the University of Panama. C. correae
yielded the known iso-dolichantoside, three new indole monoterpenoid
alkaloids, and one isoquinoiine monoterpenoid glucoside alkaloid and
C. axillaris yielded two known indole monoterpenoid alkaloids,
vallesiachotamine and 6 ’-trans-feruloyl-lyaloside.
9.- Picramnia antidesma subsp. fessonia yielded three new C-glycoside
anthraquinones, named picramniosides A, B and C, and two known
anthraquinone aloe-emodin and aloe-emodinanthrone.
10.- Previous work with Picramnia based their search for quinones on
oxidative hydrolysis, therefore missing these type of C-glycosides.
11.- The alkaloids prove to be a good chemotaxonomic character to unravel
the taxonomic position of the Cephaëlis species.
1 2 .- The plants studied were selected using the chemotaxonomic approach
and even though some compounds has shown activity against brine
shrimp, KB cell and P. falciparum, none of the expected compounds
were found.
161

SECTION 7 REFERENCES

Achenbach, H., Lottes, M., Waibel, R., Karikas, G.A., and Gupta, M.P.(1993) New alkaloids from Cephaëlis correae. Abstracts of the 41 Congress of the Gesellschaft fur Arzneipflanzenforschung, Society for Medicinal Plants Research. Dusseldorff, Germany. Poster # 50, p. 44.
Adjibade, Y, Weniger, B., Quirion, J.C., Kuballa, B., Cabalion, P. and Anton, R. (1992) Dimeric alkaloids from Psychotria forsteriana. Phytochemistry. 31, 317-319.
Adjibade, Y., Hue, B., Pelhate, M. and Anton, R. (1991) Action of calycanthine on nervous transmission in cockroach central nervous system. Planta Medlca. 57, 99-101.
Adjibade, Y., Kuballa, B., Cabalion, P., Jung, M.L., Beck, J.P. and Anton, R. (1989) Cytotoxicity on human leukaemic and rat hepatoma cell lines of alkaloid extracts of Psychotria forsteriana. Planta Medlca. 55, 567-568.
Ahmad, R. and Cava, M.P. (1977) Grisabine and grisabutine, new bisbenzylisoquinoline alkaloids from Abuta grisebachii. Journal of Organic Chemistry. 42,2271-2273.
Akerele, O. (1991) Medicinal plants: Policies and priorities. In: Conservation of Medicinal Plants. O. Akerele, V. Hey wood and H. Synge Editors. Cambridge University Press, pp. 3-11.
Akinniyi, J.A., Connolly, J.D., Rycroft, D.S., Sondengam, B.L. and Ifeadike, N.P. (1980) Tetranortriterpenoids and related compounds. Part 25. Two 3,4- secotirucallane derivatives and 2 ’-Hydroxyrohitukin from the bark of Guarea cedrata. Canadian Journal of Chemistry. 58, 1865-1868.
Ames, B.N., McCann, J. and Yamasaki, E. (1975) Methods for detecting carcinogens and mutagens with the salmonella mammalian-microsome mutagenicity test. Mutation Research. 31, 347-364.
Anderson, J.E., Goetz, C.M. and McLaughlin, J.L (1991a) A blind comparison of simple bench-top bioassays and human tumour cytotoxicities as antitumour prescreens. Phytochemical Analysis. 2,107-111.
Anderson, M.M. (1992) Cytotoxic and Antimalarial Natural Products. Ph.D. Thesis, University of London.
Anderson, M.M., O'Neill, M.J., Kirby, G.C., Phillipson, J.D. and Warhurst, D.C.(1989) A microdilution assay for the determination of in-vitro cytotoxicity of
163

potential antimalarial agents against KB cells. Journal of Pharmacy and Pharmacology. 41, suppl. 153.
Anderson, M.M., O’Neill, M.J., Phillipson, J.D. and Warhurst, D.C. (1991b) In vitro cytotoxicity of a series of quassinoids from Brucea javanica fruits against KB cells. Planta Medlca. 57, 62-64.
Angermuller, LA. (1968) Bush teas and folk remedies used by the West Indians population of Colon, Republic of Panama.Department of Anthropology, The Florida State University. Panamanian Anthropological notes-Notes In Anthropology 13, p.31-50.
Anonymous (1981) FAO Plant Production and Protection. Joint Meeting of PAO and WHO on Pesticides Residues. Report 1980. FAO, Rome. pp. 6-56.
Anonymous (1991) Advocates for Animal. 79th Annual Report, pp. 15.
Anonymous (1992) Panama en Cifras, Years 1987-1991. Direccion de Estadistica y Censo de la Contraloria General de la Republica. Panama. pp.250
Anonymous (1993a) Informe General Sobre Medldas Nacionales Adoptadas para la Conservaclon de la Diversidad Biologica. institute Nacional de Recursos Naturales Renovables. Ministerio de Desarrollo Agropecuario. Panama, pp. 10.
Anonymous (1993b) Departamento de Registres Medicos y Estadistica de Salud. Ministerio de Salud. Republic of Panama
Anonymous (1993c) Departamento de Estadistica, Division de epidemiologia. Ministerio de Salud. Republic of Panama.
Arana, C. and JuIca, B. (1986) Picramnia macrostchys Ami.Revlsta Peruana de Bloqulmlca. 8, 16-19.
Atta-Ur-Rahman, Zaman, K., Perveen, S., Habib-Ur-Rehman, Muzaffar, A., Choudhary, M. I. and Pervin, A. (1991) Alkaloids from Rhazya striata. Phytochemistry. 30, 1285-1293.
Au, T.Y., Cheung, H.T. and Sternhell, S. (1973) New corynanthe alkaloids from Strychnos angustiflora. Journal of Chemical Society. Perkin i. 13-16.
Banerji, B, and Nigam S.K. (1984) Wood constituents of Meliaceae: A review. FItoterapia. 55, 3-36.
Battersby, A.R., Burnett, A.R. and Parson, P.O. (1969) Alkaloid biosynthesis. Part IV. Partial synthesis and isolation of vincosidine and iso vincosidine: Biosynthesis of the three major classes of indole alkaloids from vincoside.
164

Journal of Chemical Society C. 1193-1200.
Beioz, A. (1992) Brine shrimp bioassay screening of two medicinal plants used by the Warao: Solanum straminifolium and Virola suranamensis. Journal of Ethnopharmacology. 37, 225-227.
Birndorf, H., D’Alessio, J. and Bagshaw, J. (1975) DNA-dependent RNA- polymerases from A. salina embryos: characterization of polymerases I and II from nauplius larvae. Developmental Biology. 45, 34-43.
Blizzard, T.A., Ruby, C.L., Mrozik, H., Preiser, F.A., Fisher, M.H. (1989) Brine shrimp (Artemia salina) as a convenient bioassay for avermectin analogs. The Journal of Antibiotics. 42,1304-1307.
Bray, D.H., Boardman, P., O’Neill, M.J., Chan, K.L, Phillipson, J.D., Warhurst,D.C. and Suffness, M. (1987) Plants as sources of antimalarial drugs. 5. Activities of Ailanthus altissima stem constituents and some related quassinoids. Phytotherapy Research. 1 , 22-24.
Bray, D.H., Warhurst, D.C., Connolly, J.D., O’Neill, M.J. and Phillipson, J.D.(1990) Plants as sources of antimalarial drugs. Part 7. Activity of some species of Meliaceae plants and their constituent limonoids. Phytotherapy Research. 4, 29-35.
Brown, R.T., Chappie, C.L. and Lashford, A.G. (1977) Isolation of strictosidine (isovincoside) lactam from Nauclea latifolia. Phytochemistry. 16, 1619-1920.
Budzikiewicz, H.; Djerassi, C. and Williams, D.H. (1967) Mass Spectrometry of Organic Compounds. Holden-Day, Inc. San Francisco, pp. 690.
Carbajal, D., Casaco, A., Arruzazabala, L, Gonzalez, R. and Fuentes, V.(1991) Pharmacological screening of plant decoctions commonly used in Cuban folk medicine. Journal of Ethnopharmacology. 33, 21-24.
Cattani, J. (1993) Malaria. In: Tropical Disease Research. Progress 1991- 1992. 11th Report of the UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases. World Health Organization, Geneva, p. 15-27.
Cava, M.P., Buck, K.T., Noguchi, I., Srinivasan, M., Rao, M.G. and Da Rocha, A.I (1975a) The alkaloids of Abuta imene and Abuta rubescens. Tetrahedron. 31, 1667-1669.
Cava, M.P., Saa, J.M., Lakshmikantham, M.V. and Mitchell, M.J. (1975b) Panurensine and nor-panurensine, new bisbenzylisoquinoline alkaloids from Abuta panurensis. Journai of Organic Chemistry. 40, 2647-2649.
165

Chan, K.L, O’Neill, M.J., Phillipson, J.D. and Warhurst, D.C. (1986) Plants as sources of antimalarial drugs. 3. Eurycoma longifolia. Planta Medlca. 52,105- 107.
Cobos-Moran, J.A. (1992) Los recursos naturales de Panama. Institute Nacional de Recursos Naturales Renovables. Ministerio de Desarrollo Agropecuarios. Panama, pp.25.
Cobos-Moran, J.A. (1993) Informe Nacional de la situacion forestal. Institute Nacional de Recursos Naturales Renovables. Ministerio de Desarrollo Agropecuario. Panama, pp.28.
Connoiiy, J.D. (1983) Chemistry of the limonoids of the Meliaceae and Cneoraceae. Annual Proceedings of the Phytochemical Society of Europe. 22, 175-213.
Connoiiy, J.D., Okorie, D.A., Dalene De Wit, L. and Tayior, D.A.H. (1976) Structure of dregeanin and rohitukin limonoids from the subfamily Melioideae of the family Meliaceae, an unusually high adsorption frequency for a six- membered lactone ring. Journal of Chemical Society, Chemicai Communications. 1976, 909-910.
Cordell, G. (1981) Introduction to Aikaioids. John Wiley & Son, New York,pp.662.
Cronquist, A. (1968) The Evoiution and Classification of Fiowering Plants.Nelson & Son Ltd. Great Britain, pp. 396.
D’Arcy, W.G., (1987) Flora of Panama. Checklist and index. Part I. Missouri Botanical Garden, Saint Louis, MO, USA.pp. 672.
Dan, S. and Dan, S.S. (1986) Phytochemical study of Adansonia digitata, Coccoloba excoriata, Psychotria adenophylla and Schleichera oleosa. Fitoterapla. 57, 445-446.
Desjardins, R.W., Canfield, C.J., Haynes, J.D. and Chulay J.D. (1979) Quantitative assessment of antimaiarial activity in vitro by a semiautomated microdilution technique. Antimicrobiai Agents and Chemotherapy. 16, 710- 718.
Djerassi, C., Monteiro, H. J., Walser, A. and Durham, L.J. (1966) Aikaloid studies. LVI. The constitution of vallesiachotamine. Journal of the American Chemical Society. 88,1792-1798.
Dohnal, B., Diak, J. and Stojakowska, A. (1989) Cultivation and preliminary phytochemical analysis of the Psychotria bacteriophiia callus tissue. Acta Poloniae Pharmaceutica. 46, 401-403.
166

Dominguez, X.A. and Alcorn, J.B. (1985) Screening of medicinal plants used by Huastec Mayans of Northeastern Mexico. Journal of Ethnopharmacology. 13, 139-156.
Duke, J.A. (1967) Darien Dictionary. Battelle Memorial Institute, Balboa, Panama, pp 82.
Duke, J.A. (1968) Darien Ethnobotanical Dictionary. Battelle Memorial Institute, Columbus, Ohio. pp. 131.
Duke, J.A. (1972) Isthmian Ethnobotanical Dictionary. Fulton, Maryland, pp. 96.
Duke, J.A. (1975) Ethnobotanical observations on the Cuna Indians. Economic Botany. 29, 278-293.
Dute, P., Weber, J.F., Fournet, A., Cave, A. and Bruneton, J. (1987) Bisbenzylisoquinoline alkaloids from Abuta pahni. Phytochemistry. 26,2136-2137.
Dwyer, J.D. (1964) Panama, plant collection and the Missouri Botanical Garden. Annals of Missouri Botanical Garden, 51, 109-17.
Dwyer, J.D. (1980) Family 179. Rubiaceae. Part I. In: Flora of Panama. Part IX. R.E. Woodson and W. Schery Editors. Annals of the Missouri Botanical Garden. St Louis, Missouri. 67(1): 59-82.
Dwyer, J.D. (1985) The history of plant collecting in Panama. In The Botany and Natural History of Panama. Eds. W.G. D’Arcy and M.D.A. Correa, Missouri Botanicai Garden, Saint Louis, Missouri, U.S.A. p. 179-83.
Edington, S.M. (1991) Taxoi out of the woods. Bio/Technology. 9, 933-938.
Ekong, R., Kirby, G.C., Patel, G.,Phillipson, J.D. and Warhurst, D.C. (1990) Comparison of the in vitro activities of quassinoids with activity against Piasmodium faiciparum, anisomicin and other inhibitors of protein synthesis. Biochemical Pharmacology. 40, 297-301.
Ekong, R., Partridge, S.J., Anderson, M.M., Kirby, G.C., Warhurst, D.C., Russell, P.P. and Phillipson, J.D. (1991) Piasmodium faiciparum: Effects of phaenthine, a naturally-occurring bisbenzylisoquinoline alkaloid, on chloroquine- resistant and -sensitive parasite in vitro, and its influence on chloroquine activity. Annals of Tropical Medicine and Parasitology. 85, 205-213.
Eppley, R.M. and Bailey, W.J. (1973) 12,13-Epoxy-A^-trichothecenes as the probable mycotoxin responsible for stachybotryotoxicosis. Science. 181, 758- 760.
167

Evans, D.A., Joule, J.A. and Smith, G.F. (1968) The alkaloids of Rhazya ohentalis. Phytochemistry. 7,1429-1431.
Ewing, R.D., Peterson, G.L and Conte, P.P. (1974) Larval salt gland of A salina nauplii: Localization and characterization of the sodium + potassium- activated adenosine triphosphatase. Journal of Comparative Physiology. 88, 217-234.
Fairbairn, J.W. and Simic, S. (1963) The quantitative conversion of barbaloin to aloe-emodin and its application to the evaluation of aloes. Journal of Pharmacy and Pharmacology. 15, 325-328.
Fairlamb, A.H., Warhurst, D.C. and Peters, W. (1985) An improved technique for the cultivation of Plasmodium falciparum in vitro without daily medium change. Annals of Tropical Medicine Parasitology. 79, 379-384.
Farnsworth, N.R. (1983) The Napralert data base as an information source for application to traditional medicine. In: Traditional Medicine and Health Care Coverage. R.H. Bannerman, J. Burton and C. Wenchieh Editors. WHO Publications, Geneva, pp. 96-101.
Farnsworth, N.R., Akerele, O., Bingel, A.S., Soejarto, D.D. and Guo, Z.G. (1985) Medicinal plants in therapy. Bulletin of the World Health Organization. 63, 965-981.
Farnsworth, N.R. and Bingel, A.S. (1977) Problems and prospects of discovering new drugs from higher plants by pharmacological screening. In: New Natural Products and Plant Drugs with Pharmacological, Biological or Therapeutical Activity. H. Wagner and P. Wolff Editors. Springer-verlag, Berlin, pp. 1-22.
Farnsworth, N.R. and Morris, R.W. (1976) Higher plants, the sleeping giant of drug development. American Journal of Pharmacy. March-April, 46-52.
Farnsworth, N.R. and Soejarto, D.D. (1991) Global importance of medicinal plants. In: Conservation of Medicinal Plants. Akerele, O., Heywood, V. and Synge, H. Eds.Cambridge University press, pp.25-51.
Feldman, E., Arlin, Z., Ahmed, T., Mittelman, A., Puccio, 0., Chun, H., Cook, P. and Baskind, P. (1992) Homoharringtonine is safe and effective for patients with acute myelogenous leukaemia. Leukaemia. 6, 1185-1188.
Feng, P.C., Haynes, L.J., Magnus, K.E., Plimmer, J.R. and Sherrat, H.S.A. (1962) Pharmacological screening of some West Indian medicinal plants. Journal of Pharmacy and Pharmacology. 14, 556-561.
Ferguson, G., Gunn, P.A., Marsh, W.C., McCrindle, R., Restivo, R., Connolly,
168

J.D., Fulke, J.W.B. and Henderson, M.S. (1975) Tetranortriterpenoids and related substances. XVII. A new skeletal class of triterpenoids from Guarea glabra (Meliaceae). Journal of the Chemical Society. Perkin Transaction i. 1975, 491-497.
Ferrigni, N.R., McLauchlin, J.L, Powell, R.G. and Smith, C.R. (1984) Use of Potato disc and brine shrimp bioassays to detect activity and isolate piceatannol as the antileukemic principle from the seeds of Euphorbia lagascae. Journai of Natural Products. 47, 347-352.
Finney, D.J. (1971) Probit Analysis. 3rd edition. Cambridge University Press. Cambridge, pp. 333.
Fitzpatrick, F.K. (1954) Plant substances active against Mycobacterium tuberculosis. Antibiotics and Chemotherapy. 4, 528-536.
Furlan, M., Roque, N.F. and Fihio, W.W. (1993) Cycloartane derivatives from Guarea trichilioides. Phytochemistry. 32, 1519-1522.
Galeffi, C., Scarpetti, P. and Marini-Bettolo, G.B. (1977) New curare alkaloids.II. New bisbenzylisoquinoline alkaloids from Abuta grisebachi (Menispermaceae). Farmaco Edizione Scientifica. 32, 853-865.
Geran R.l. and Greenberg, N.H. (1972) Cell culture technical procedures. KB cytotoxicity test-protocol 13. Cancer Chemotherapy Reports. 3,2.
Gibbs, R.D. (1963) History of chemical taxonomy. In: Chemical Plant Taxonomy. T.Swain Editor. Academic Press. London, p. 41-88.
Gilbert, B., Mors, W.B., Baker, P.M., Thomassini, T.C.B., Coulart, E.G., De Holanda, J.C., Ribeiro Da Costa, J.A. Lopes, J.N.G., Dos Santos FihIo, D., Sarti, S.J. and Turco, A.M.T. (1972) Anthelmintic activity of essential oils and their chemical components. Annals of the Brazilian Academy of Sciences. Supplement 44, 423-428.
Gillin, F.D., Reiner, D.S. and Suffness, M. (1982) Bruceantin, a potent amoebicide from a plant, Brucea antidysenterica. Antimicrobiai Agents and Chemotherapy. 22, 342-345.
Glass, R.I., Claeson, M., Blake, P.A., Waldman, R.J. and Pierce, N.F. (1991) Cholera in Africa: Lessons on transmission and control for Latin America. Lancet. 338, 791-795.
Gottlieb, O.R. (1982) Ethnopharmacology versus chemosystematic in the search for biologically active principles in plants. Journal of Ethnopharmacology. 6, 227-238.
169

Granade, H.R., Cheng, P.O. and Doorenbos, N.J. (1976) Ciguatera I: Brine shrimp (Artemia salina L) larval assay for ciguatera toxins. Journal of Pharmaceutical Sciences. 65, 1414-1415.
Greig, J.B., Say, S.J.E. and Bennets, R.J. (1980) A toxin from palmyra palm, Borassus flabellifer. Partial purification and effect in rats. Food and Cosmetic Toxicology. 18, 483-488.
Gueritte-Voegelein, P., Sevenet, T., Pusset, J., Adeline, M.T., Gillet, B., Beloeil, J.C., Guenard, D. and Potier, P. (1992) Alkaloids from Psychotria o/eo/cfes with activity on growth hormone release. Journal of Natural Products. 55, 923- 930.
Guionneau-Sinclair, F. (1989) Poblacion estimada y asentamientos Amerindios en Panama. Scientia (University of Panama). 4,37-46.
Gupta, M.P., Correa, M.A., Arias, T.D. and Lamba, S.S. (1979) Ethnopharmacognostic observation on Panamanian medicinal plants. Quarterly Journal of Crude Drug Research. 17,115-130.
Gupta, M.P., Correa, M.D., Solis, P.M., Jones, A., Galdames, C., Guionneau- Sinclair, P. (1993a) Medicinal plant inventory of Kuna Indians: Part I. Journal of Ethnopharmacology. 40,77-109.
Gupta, M.P., Gordaliza, M., Karikas, G.A., Miguel del Corral, J.M. and San Feliciano, A. (1993b) Major alkaloids from Cephaëlis axilaris. Poster presentation at Recontres Europeene Phytochimie. Anger, Prance.
Gupta, M.P., Solis, P.N., Soto, A.A., Cedeho, J.C., Correa, M.A., Esposito, M., Sanchez, C. (1986) Observaciones Etnofarmacognosticas sobre plantas médicinales en Panama. Parte II. Scientia (University of Panama) 1, 43-57.
Guru, P.Y., Warhurst, D.C., Harris, A. and Phillipson, J.D. (1983) Antimalarial activity of bruceantin in vitro. Annals of Tropical Medicine and Parasitology. 77, 433-435.
Hamill, R.L, Higgins, C.E. Boas, H.E. and Gorman, M. (1969) The structure of beauvericin, a new depsipeptide antibiotic toxin to Artemia salina. Tetrahedron Letters. 49, 4255-4258.
Hamilton, C.E. (1989) A revision of mesoamerican Psychotria subg. Psychotria (Rubiaceae). Part I. Annals of the Missouri Botanicai Garden. 76, 67-111.
Hammond, G.B., Baenziger, N.C. and Wiemer, D.P. (1990) Dammara-20-25- dien,3p,24a-diol: a natural repellent of Acromyrmex octospinosus. Phytochemistry. 29, 783-785.
170

Hamor, T.A., Monteath-Robertson, J., Shrivasava, H.N. and Silverton, J.V. (1960) The structure of calycanthine. Proceedings of the Chemical Society. 78-86.
Hansel, R. (1972) Medicinal plants and empirical drugs research. In: Plants in the Development of Modern Medicine. I . Swain Editor. Harvard University Press. USA. p. 161-174.
Harborne, J.B., Boulter, D. and Turner, B.L. (1971) Chemotaxonomy of the Leguminosae. Academic Press. London, pp. 612.
Hatfield, G.M., Arteaga, L. Dwyer, J.D., Arias, T.D. and Gupta, M.P. (1981) An investigation of Panamanian ipecac: Botanical source and alkaloid analysis. Journai of Natural Products. 44, 452-456.
Hayashi, T., Smith, F.T. and Lee, K.H (1987) Antitumor agents. 89. Psychorubrin, a new cytotoxic naphtoquinone from Psychotha rubra and its structure-activity relationships. Journal of Medicinal Chemistry. 30, 2005- 2008.
Hazlett, D.L. (1986) Ethnobotanical observation from Cabecar and Guaymi settlements in Central America. Economic Botany. 40, 339-352.
Hegnauer, R. (1962-1969) Chemotaxonomic de Pfianzen. Vols. 1-5. Birkhauser Verlag, Basel and Stuttgart.
Hemingway, S.R. and Phillipson, J.D. (1980) Alkaloids of the Rubiaceae. In: indole and Biogeneticaliy Related Aikaioids. J.D. Phillipson and M.H. Zenk Editors. Academic Press. London, p. 63-90.
Herz, W., Santhaman, P.S. and Wahlberg, I. (1972) 3-epi-betulinic acid, a new triterpenoid from Picramnia pentandra. Phytochemistry. 11, 3061-3063.
Heslop-Harrison, J. (1963) Species concepts: Theoretical aspects. IN: Chemical Plant Taxonomy. T.Swain Editor. Academic Press. London, p. 17- 40.
Hocquemiller, R. and Cave, A. (1984) Saulatine, A new isoquinoiine alkaloid isolated from Abuta builata. Journal of Natural Products. 47, 539-540.
Hoke, S.H., Carley, C.M., Johnson, T. and Broski, H. (1987) Use of solid phase extraction systems to improve the sensitivity of Artemia bioassay for trichothecene mycotoxin. Journai of Association of Official Analytical Chemistry. 70, 661-663.
Horton, R.J. (1988) Introduction of halofanthrine for malaria treatment. Parasitology Today. 4, 238-239.
171

Hotellier, F., Delaveau, P. and Pousset, J.L. (1975) Nauclefine et naucletine deux nouveaux alkaloides de type indoloquinolizidine isoles du Nauclea latifolia. Phytochemistry. 14,1407-1409.
Hui, W.H. and Yee, C.W. (1967) An examination of the Rubiaceae of Hong Kong. III. Phytochemistry. 6, 441-442.
Hutchinson, J. (1959) The Famiiies of Fiowering Piants. Vol. 1. Dicotyledons: Arranged according to a new system based on their probable phytogeny. 2nd Edition. Claredon Press. Oxford, pp. 510.
Inouye, H., Takeda, Y., Nishimura, H., Kanomi, A., Okuda, T. and Puff, C. (1988) Chemotaxonomic studies of Rubiaceous plants containing iridoid glycosides. Phytochemistry. 27, 2591-2598.
Ishikura, N., Sugahara, K. and Kurosawa, K. (1979) Eryodictyol-7-glucoside and other phenolics in the blue fruits of Lasianthus japonica. Z. Naturforsch. Ser. 0. 34, 628-629.
Itoh, A., Tanahashi, I . and Nagakura, N. (1989) Neoipecoside and 7- methylneoipecoside, new unusually cyclized tetrahydroisoquinoline monoterpene glucoside from Cephaelis ipecacuanha. Chemicai and Pharmaceuticai Buiietin. 37,1137-1139.
Itoh, A., Tanahashi, T. and Nagakura, N. (1991) Six tetrahydroisoquinoline monoterpene glucosides from Cephaelis ipecacuanha. Phytochemistry. 30, 3117-3123.
Joly, L.G. (1981) Feeding and trapping fish with Piper auritum. Economic Botany. 35, 383-390.
Joly, L.G., Guerra, S., Septimo, R, Solis, P.M., Correa, M.D., Gupta, M.P., Levy, S., Sandberg, F. and Perera, P. (1990) Ethnobotanical inventory of medicinal plants used by the Guaymi Indians in western Panama. Part II. Journai of Ethnopharmacoiogy. 28, 191-206.
Joly, L.G., Guerra, S., Septimo, R, Solis, P.M., Correa, M.D., Gupta, M.P., Levy, S. and Sandberg, F. (1987) Ethnobotanical inventory of medicinal plants used by the Guaymi Indians in western Panama. Part I. Journai of Ethnopharmacoiogy. 20, 145-171.
Jork, H., Funk, W., Fischer, W., Wimmer, H. (1990) Thin-iayer Chromatography. Reagents and Detection Methods. VCHverlagsgesellschaft mbH. Germany. Volume la, pp.464.
Kardono, L.B.S., Angerhofer, C.K., Tsauri, S., Padmawinata, K., Pezzuto, J.M. and Kinghorn, A.D. (1991) Cytotoxic and antimalarial constituents of the roots
172

of Eurycoma longifolia. Journal of Natural Products. 54, 1360-1367.
Kersten, W.J., Brooks, R.R., Reeves, R.D. and Jafre, T. (1980) Nature of nickel complexes in Psychotria dourrei and other nickel-accumulating plants. Phytochemistry. 19, 1963-1965.
Khalid, S.A., Farouk, A.F., Geary, T.G. and Jensen, J.B. (1986) Potential antimalarial candidates from African plants: an in vitro approach using Piasmodium falciparum. Journal of Ethnopharmacology. 15,201-209.
Kinghorn, A.D., Harjes, K.K. and Doorenbos, N.J. (1976) Screening procedure for phorbol esters using brine shrimp {Artemia salina) larvae. Journal of Pharmaceutical Sciences. 66,1362-1363.
Kirby, G.C., O’Neill, M.J., Phillipson, J.D. and Warhurst, D.C. (1989) In vitro studies on the mode of action of quassinoids with activity against chloroquine- resistant Plasmodium falciparum. Biochemistry and Pharmacology. 38,4367- 4374.
Kupchan, S.M., La voie, E.J. Branfman, A.R., Fei, B.Y., Bright, W.M. and Bryan, R.F. (1977) Phyllantocin, a novel bisabolane aglycone from antileukemic glycoside, phyllanthoside. Journal of American Chemical Society. 99, 3199- 3201.
Kyerematen, G.A. and Ogunlana, E.G. (1987) An integrated approach to the pharmacological evaluation of traditional materia medica. Journai of Ethnopharmacology. 20,191-207.
Lee, K.H., Lin, Y.M., Uw, T.S., Zhang, D.C., Yamagishi, T., Hall, I.H., Chang, J.J., Wu, R.Y. and Yang, T.H. (1988) The cytotoxic principles of Pruneiia vulgaris, Psychotria serpens and Hyptis capitata: ursolic acid related derivatives. Planta Medica. 54, 308-311.
Leon, J.J. (1975) Triterpenoids and anthraquinonic derivatives obtained from Picramnia seliowii (Simaroubaceae). Boietin de la Sociedad Quimica del Peru. 41,14-30.
Libot, F., Kunesch, N. and Poisson, J. E. (1988) Biomimetic transformation of hodgkinsine, a pyrrolidinoindoline alkaloid. Heterocycles. 27, 2381-2386.
Libot, P., Miet, C., Kunesch, N. and Poisson, J.E. (1987) Rubiaceae of Oceania: Alkaloids of Psychotria oieoides of new Caledonia and of Caiycodendron miinei from Vanuatu (new hybrids). Journal of Natural Products. 50, 468-473.
173

Lukacova, V., Polonsky, J., Moretti, C., Pettit, G.R. and Schmidt, J.M. (1982) Isolation and structure of 14,15-beta-epoxyprieurianin from the south American tree Guarea guidonia. Journal of Natural Products. 45, 288-294.
Makler, M.T., Ries, J.M., Williams, J.A., Bancroft, J.E., Piper, R.C., Gibbins, B.L. and Hinrichs, D.L. (1993) Parasite lactate dehydrogenase as an assay for Plasmodium falciparum drug sensitivity. American Journal of Tropical Medicine and Hygiene. 48, 739-741.
Manitto, P., Monti, D. and Speranza, G. (1990) Studies on Aloe. Part 6. Conformation and absolute configuration of aloins A and B and related 10-c- glucosyl-9-anthrones. Journal of Chemical Society. Perkin Transaction 1. 1297-1300.
Manitto, P., Monti, □., Speranza, G., Mulinacci, N., Vincieri, F.F., Griffini, A. and Pifferi, G. (1993) Conformational studies of natural products. Part 4. Conformation and absolute configuration of Cascarosides A, B, C, D. Journal of Chemical Society. Perkin 1 .1577-1580.
Manske, R.H.F. and Holmes, H. (1950-1958) The Alkaloids. Academic Press. New York.
Marshall, S.J., Russel, P.F., Wright, C.W., Anderson, M.M., Phillipson, J.D., Kirby, G.C., Warhurst, D.C. and Schiff, P.L. (1994) in vitro antiplasmodial, antiamoebic and cytotoxic activities of a series of bisbenzylisoquinoline alkaloids. Antimicrobial Agents and Chemotherapy. 38, 96-103.
McLaughlin, J.L. (1991) Crown gall tumours on potato discs and brine shrimp lethality: two simple bioassays for higher plant screening and fractionation. In: Assay for Bloactlvity. K. Hostettmann Ed. Vol.6 in the series Methods in plant biochemistry, P.M. Dey and J.B. Harborne, series editors. Academic Press. London, pp. 1-32.
McLaughlin, J.L., Chang, C.J. and Smith, D.L. (1993) Simple bench-top bioassays (brine shrimp and potato disks) for the discovery of plant antitumor compounds. Review of recent progress. American Chemical Society. Symposium series. 534,112-137.
McNair, J.B (1935) The taxonomic and climatic distribution of alkaloids. Bulletin Torrey Botany Club. 62, 832-841.
McNair, J.B. (1929) The taxonomic and climatic distribution of oils, fats and waxes in plants. American Journal of Botany. 16, 304-311.
McRae, T.H. and Pandey, A.S. (1991) Effects of metals on early life stages of the brine shrimp, Artemia: a developmental toxicity assay. Archives of Environmental contamination Toxicology. 20, 247-252.
174

McRae, T.H., Bagshaw, J.C. and Warner, A.H. (1989) Biochemical and Ceii Biology of Artemia. CRC Press Inc. Florida, pp. 264.
Menachery, M.D. and Cava, M.P. (1980) Grandirubrine, A new tropoloisoquinoline alkaloid. Heterocycies. 14, 943-945.
Meyer, B.N., Ferrigni, N.R., Putnam, J.E., Jacobsen, L.B., Nicholson, R.E., and McLaughlin, J.L. (1982) Brine shrimp: A convenient general bioassay for active plant constituents. Planta Medica. 45,31-34.
Miser, U.S., Smithson, W.A., Krivit, W. Hughes, C.H., Davis, D., Krailo, M.D. and Hammond, G.D. (1991) Phase II trial of indicine N-oxide in relapsed paediatric solid tumours. A report from the Children Cancer Study Group, investigational New Drugs. 9, 339-342.
Miser, J.S., Smithson, W.A., Krivit, W., Hughes, C.H., Davis, D., Krailo, M.D. and Hammond, G.D. (1992) Phase II trial of indicine N-oxide in relapsed acute leukaemia of childhood: A report from the Children Cancer Study Group. American Journal of Clinical Oncology. 15, 135-140.
Mootoo, B.S., Jativa, C., Tinto, W.F., Reynolds, W.F. and McLean, S. (1992) Euadorin, a novel tetranortriterpenoid of Guarea kunthiana: Structure elucidation by 2-DNMR spectroscopy. Canadian Journal of Chemistry. 70, 1260-1264.
Morton, J.F. (1981) Atlas of Medicinal Plants of Middle America. Thomas publisher. Illinois, pp. 1420.
Nagakura, N., Itoh, A., Tanahashi, I . (1993) Four tetrahydroisoquinoline monoterpene glucosides from Cephaelis ipecacuanha. Phytochemistry. 32, 761-765.
Nunez-Melendez, E. (1975) Plantas Médicinales de Costa Rica y su Foiciore. Ciudad Universitaria "Rodrigo Fado", San José. pp. 279.
Oga, S., Sertie, J.A., Brasile, A.C. and Hanada, S. (1981) Antiinflammatory effect of crude extract from Guarea guidonia. Planta Medica. 42, 310-312.
O’Neill, M.J., Bray, D.H., Boardman, P., Chan, K.L. Phillipson, J.D., Warhurst,D.C. and Peters, W. (1987) Plants as sources of antimalarial drugs. Part 4. Activity of Brucea javanica fruits against chloroquine-resistant Plasmodium falciparum in vitro and Plasmodium berghei in vivo. Journal of Natural Products. 50, 41-48.
O’Neill, M.J., Bray, D.H., Boardman, P., Phillipson, J.D. and Warhurst, D.C. (1985) Plants as antimalarial drugs. Part 1. In vitro test method for the evaluation of crude extracts from plants. Planta Medica. 5, 394-397.
175

O’Neill, M.J., Bray, D.H., Boardman, P., Phillipson, J.D., Warhurst, D.C., Peters, W. and Suffness, M. (1986) Plants as sources of antimalarial drugs. 2. In vitro antimalarial activities of some quassinoids. Antimicrobial Agents and Chemotherapy. 30,101-104.
O’Neill, M.J., Bray, D.H., Boardman, P., Wright, C.W., Phillipson, J.D., Warhurst, D.C., Gupta, M.P., Correa, M. and Solis, P.N. (1988) Plants as sources of antimalarial drugs. 6. Activities of Simarouba amara fruits. Journal of Ethnopharmacology. 22,183-190.
Parisot, D., Devys, M. and Barbier, M. (1989) Conversion of anhydro fusarubin lactol into the antibiotic bostrycoidin. The Journal of Antibiotics. 52, 1189- 1190.
Parisot, D., Devys, M. and Barbier, M. (1990) 6-0-Demethyl-5- deoxybostrycoidin, a 2-aza-anthraquinone produced by the fungus Nectaria haematococca. Phytochemistry. 29, 3364-3365.
Parke, D.V. (1983) A more scientific approach to the safety evaluation of chemicals. In: Animals in Scientific Research: An Effective Substitute for Man?. P. Turner Editor. Macmillan Press Ltd. London, pp. 7-28.
Pavanand, K., Yongvanitchit, K., Webster, H.K., Dechatiwongse, T., Nutakul, W., Jewvachdamrogkul, Y. and Bansiddhi, J. (1988) In vitro antimalarial activity of a Thai medicinal plant Picrasma javanica. Phytotherapy Research. 2, 33- 36.
Payne, D. (1987) Spread of chloroquine resistance in Plasmodium falciparum. Parasitology Today. 3, 241-246.
Pennington, T.D., Styles, B.T. and Taylor, D.A. (1981) Meiiaceae. Monograph N° 28. Organization for Flora Neotropica. New York Botanical Garden, New York, pp.472.
Penso, G. (1983) Index Piantarum Medicinalium Totius Mundi Eorumque Synonymorum. O.E.M.F., Milano, Italy, pp. 1026.
Pereira, O.L., Wolter Fihio, W., Da Rocha, A.F.I., Carvahio, M.G. and Braz Fihio, R. (1990) Diterpenes isolated from Guarea carinata. Quimica Nova. 13, 247-249.
Peters, W. (1985) Antiprotozoal agents. In: The Scientific Basis of Antimicrobial Chemotherapy. D. Greenwood and F. O’Grady Editors. Cambridge University Press, pp. 95-131.
Pettit, G.R., Barton, D.H.R., Herald, C.L., Polonsky, J. Schmidt, J.M. and Connolly, J.D. (1983) Antineoplastic agents. 97. Evaluation of limonoids against
176

the murine P388 leukaemia cell line. Journal of Natural Products. 46, 397- 390.
Phillipson, J.D., Hemingway, S.R., Bisset, N.G., Houghton, P.J. and Shellard, E.J. (1974) Angustine and related alkaloids from species of Mitragyna, Nauclea, Uncar/a and Strychnos. Phytochemistry. 13, 973-978.
Phillipson, J.D., Wright, C.W., Kirby, G.C. and Warhurst, D.C. (1994) Phytochemistry of some plants used in traditional medicine for the treatment of protozoal diseases. In: Phytochemistry of Plants Used In Traditional Medicine. K. Hostettmann and A. Marston. Oxford University Press, Oxford, in press.
Phillipson, J.D., Wright, C.W., Kirby, G.C. and Warhurst, D.C. (1993) Tropical Plants as Sources of Antiprotozoal Agents. In: Phytochemical Potential of Tropical Plants. K.R. Downey, J.T. Romeo and H.A. Stafford. Vol. 27 in the Series: Recent Advances in Phytochemistry. Plenum Press. New York-London. p. 1-40.
Pinheiro, M.L, Wolter Fihio, W., Da Rocha, A.I., Porter, B., Wenkert, E. (1983) Abutasterone, an ecdysone from Abuta velutina. Phytochemistry. 22, 2320- 2321.
Popinigis, I., Moreira, E.A., Nakashima, T., Krambeck, R. and Miguel, O.G. (1980) Pharmacognostic study of Picramnia parvifolia Engler. Tribuna Farmaceutlca. 1,24-43.
Porter, D.M. (1973) Family 90. Simaroubaceae. In: Flora of Panama. Part IV. R.E. Woodson, R.W. Schery and Collaborators. Annals of the Missouri Botanical Garden. St Louis Missouri. Vol. 60 (1): 23-39.
Rees, D. (1992) Animal experimentation in fundamental research. In: Animal Experimentation on the Future of Medical Research. J.H. Betting Editor. Portland Press Proceedings. London, p. 13-27.
Reiss, J. (1972) Comparing investigation on the toxicity of some mycotoxin to the larvae of the brine shrimp (Artemia salina). Erte. Abt. Orlg. Relhe. B. Hyg. Praev. Med. 155, 531-534.
Robert-Gero, M., Bachrach, U., Bhatnagar, S. and Polonsky, J. (1985) Inhibition in vitro de la croissance des promastigotes de Leishmania donovani par des quassinoids. C.R. Academy of Sciences, Paris. 300:11, 803-806.
Robinson, A.B., Manly, K.F. Anthony, M.P., Catchpool, J.F and Pauling, L. (1965) Anaesthesia of Artemia larvae: method for quantitative study. Science. 149, 1255-1258.
177

Robyn, R. and Irvin, J. (1980) A survey of selected plants for the presence of eukaryotic protein biosynthesis inhibitors. Texas Journal of Sciences. 32, 55- 58.
Roth, A., Kuballa, B. Cabalion, P. and Anton, R. (1985) Preliminary study of the alkaloids of Psychotria forsteriana. Planta Medica. 51, 289.
Rychener, M. and Steiger, W. (1989) Reindarstellung, Glucose-6-phosphat- dehydrogenase-hemmung und HPLC-analytik von vier 1,8-dihydroxyanthronen. Pharmaceutlca Acta Helvetica. 64, 9-11.
Saa, J.M., Lakshmikantham, M.V., Mitchell, M.J. and Cava, M.P. (1976) Krudovine, a new bisbenzylisoquinoline alkaloid from Abuta splendida. Journal of Organic Chemistry. 41, 317-319.
Samuelsson, G., Kyerematen, G., and Farah, M.H. (1985) Preliminary chemical characterization of pharmacological active compounds in aqueous plant extracts. Journal of Ethnopharmacology. 14,193-201.
Sandberg, F. and Cronlund, A. (1977) What can we still learn from traditional folklore medicine? Examples from the results of a biological screening of medicinal plants from equatorial Africa. Proceedings of the Third Asian Symposium of Medicinal Plants and Spices. Colombo, Sri Lanka. 3, 178- 197.
Santavy, F. (1970) Papaveraceae alkaloids. In: The Alkaloids. Vol. XII. R.H.F. Manske Editor. Academic Press. New York and London. 12, 333-454.
Schiff, P.L. (1983) Bisbenzylisoquinoline alkaloids. Journal of NaturalProducts. 46, 1-43.
Schiff, P.L. (1987) Bisbenzylisoquinoline alkaloids. Journal of naturalProducts. 50, 529-599.
Schultes, R.E. (1972) Preface. In Flora Tôxica de Panamà. N. Escobar editor, Editorial Universitaria, Panama, p. 7-8.
Schultes, R.E. (1986) Ethnopharmacological conservation: a key to progress in medicine. Opera Botanlca. 92, 217-224.
Schultes, R.E. and Raffauf, R.F. (1990) The healing forest. Medicinal and toxic plants of the Northwest Amazonia. In: Historical, Ethno and Economic Botany Series. Vol. 2. T.R. Dudley Editor. Dioscorides Press. Oregon, pp. 484.
Scott, 0.(1982) UKEMS Collaborative genotoxicity trial, cytogenetic tests of
178

4CMB, BC and 4HMB: Summary and appraisal. Mutation Research. 100, 313-331.
Shiraishi, N., Akiyama, S., Nakagawa, M., Kobayashi, M. and Kuwano, M. (1987) Effect of bisbenzylisoquinoline (bicoclaurine) alkaloids on multidrug resistance in KB human cancer cells. Cancer Research. 47, 2413-1416.
Skiles, J.W., Saa, J.M. and Cava, M.P. (1979) Splendidine, a new oxoaporphine alkaloid from Abuta rufescens Aublet. Canadian Journal of Chemistry. 57, 1642-1646.
Smith, G.N. (1968) Strictosidine: a key intermediate in the biogenesis of indole alkaloids. Journal of Chemical Society, Chemical Communications. 15,912- 914.
Smith, P.M. (1976) The Chemotxonomy of Plants. Edward Arnold Ltd. Bristol, England, pp. 313.
Snell, P.M. and Snell, C.R. (1989) Solubilization and characterization of the bradykinin receptor from rat uterus. Biochemical Society Transactions. 17, 569-570.
Solecki, R. (1975) Shanidar IV. a Neanderthal flower burial in northern Iraq. Science. 190, 880-881.
Spencer, C.F., Koniuszy, F.R., Rogers, E.F., Shavel, J., Easton, N.R., Kaczka,E.A., Kuehl, F.A., Phillips, R.F., Walti, A., Folkers. K., Malanga, C. and Seeler, A.O. (1947) Survey of plants for antimalarial activity. Lloydia. 10, 145-174.
Spjut, R.W. (1985) Limitations of a random screen: Search for new anticancer drugs in higher plants. Economic Botany. 39, 266-288.
Stahl, E. (1969) Thin-layer Chromatography. A Laboratory Handbook. 2ndEdition. Sprlnger-Verlag. Heilderberg. pp. 1041.
Standley, P.C. (1927)The flora of Barro Colorado Island. Smithsonian Miscellaneous Collections. 78,1-32.
Standley, P.C. (1928). Flora of the Panama Canal Zone. Contributions to U.S. National Herbarium, 27, 1-416.
Standley, P.C. and Steyermark, J. (1958) Flora of Guatemala. Chicago Natural History Museum, Chicago, 24(3), 478.
Standley, P.C. and Williams, L.O. (1975) Flora of Guatemala, Part XI (Rubiaceae). Fleldlana Botany. 24(11) 1-274.
179

Steyermark, J.A. (1972) Rubiaceae. In: The Botany of the Guayana Highlands. Part IX. B. Maguire and Collaborators Editors. Memorial New York Botanical Garden. 23, 407-717.
Steyermark, J.A. (1974) Rubiaceae. In: Flora de Venezuela. T. Lasser Editor. Instituto Botanico, Direccion de Recursos Naturales Renovables, Ministerio de Agriculture y Cria. Caracas, Venezuela. 9(1-3) 1-2070.
Suffness, M. and Douros, J. (1982) Current status of the NCI plant and animal product program. Journal of Natural Product. 45, 1-14.
Tanaka, K., Manabe, M. and Matssura, S. (1982) Biological tests using the brine shrimp (A. salina). 3. The toxicities of metal compounds and the synergistic effects of metal compounds and mycotoxin on the brine shrimp. Report of the National Food Research Institute, (Tokyo). 39, 58-63.
Tarpley, W.A. (1958) Studies on the use of the brine shrimp Artemia salina (Leach) as a test organism for bioassay. Journal of Economic Entomology. 51,780-783.
Taylor, C.M. (1991) The vascular flora of the la selva biological station, Costa Rica, Rubiaceae. Selbyana. 12, 141-190.
Taylor, C.M. (1993) Personal Communication.
Taylor, C.M. (1994) Personal Communication.
Taylor, C.M., Hammel, B.E. and Burger, W.C. (1991) New species, combinations, and records in Rubiaceae from the selva biological station, Costa Rica. Selbyana. 12,134-140.
Taylor, D.H. (1983) Biogenesis, distribution and systematic significance of limonoids in the Meiiaceae, Cneoraceae and allied taxa. Annual Proceedings of the Phytochemical Society of Europe. 22, 353-375.
Thomas, W.W. (1988) A conspectus of Mexican and Central American Picramnia (Simaroubaceae). Brittonla. 40, 89-105.
Torres de Arauz, R. (1972) Aspectos etno-ecologicos de los grupos humanos del Este de Panama. Acta del II Symposium Naclonal de Antropologla, Arqueologla y Etnohlstorla de Panama. Universidad de Panama e Instituto Nacional de Culture, Panama, p. 291-311.
Torres de Arauz, R. (1980) Panamà Indi'gena. Instituto Nacional de Culture, Panama, pp.383.
Trager, W. and Jensen, J.B. (1976) Human malaria parasites in continuous
180

culture. Science. 193, 673-675.
Trager, W., Polonsky, J. (1981) Antimalarial activity of quassinoids against chloroquine-resistant Plasmodium falciparum in vitro. American Journal Tropical Medicine and Hygiene. 30, 531-537.
Trotter, R.T., Logan, M.H., Rocha, J.M. and Boneta, J.L (1983) Ethnography and bioassay: Combined methods for a preliminary screen of home remedies for potential pharmacological activity. Journal of Ethnopharmacology. 8,113- 119.
Tyler, V.E., Brady, LR. and Robbers, J.E. (1988) Pharmacognosy. 9th Edition. Lea & Febiger, Philadelphia. pp.519.
Van Debos, G. and Finamore, F.J. (1974) An unusual pathway for the synthesis of adenosine triphosphate by the purine-requiring organism Artemia salina. The Journal of Biological Chemistry. 249, 2816-2818.
Van Dyke, K. (1991) Use of Tetrandrlne and Its Derivatives to Treat Malaria.US Patent 5,025,020.
Wall, M.E., Taylor, H. and Wani, M.G. (1987) Plant antitumour agents. 24. Rapid 9-KB assay. Journal of Natural Products. 50, 764-766.
Walton, B.T., Ho, C.H., Ma, C.Y., O’Neill, E.G. and Kao, G.L (1983) Benzoquinolinediones: activity as insect teratogens. Science. 22, 422-423.
Wani, M.G., Taylor, H.L., Wall, M.E., Goggon, P. and McPhail, A.T. (1971) Plants antitumours agents. VI. The Isolation and the structure of taxol. A novel antileukemic and antitumor agent from Taxus brevifolia. Journal of American Chemical Society. 93, 2325-2327.
Warhurst, D.G. (1985) New drugs and their potential use against drug-resistant malaria. Annals of the Institute Super Sanita. 21, 327-336.
Warner, A.M., McRae, T.H. and Bagshaw, J.G. (1988) Cell and Molecular Biology of Artemia development. Plenum Press. New York. pp. 453.
Warren, K.E.H. (1970) The Structure and Absolute Stereochemistry of Strictosidine. Ph.D. Thesis, Manchester.
Waterman, P.G. and Shouming Zhong (1982) Vallesiachotamine and iso vallesiachotamine from the seeds of Strychnos tricalysioides. Planta Medica. 45, 28-30.
Wehmer, G. (1931) Die Pfianzenstoffe. Vol. 2. Jena Verlag von Gustav Fisher. Germany, p. 641-1511.
181

Wiegrebe, W., Kramer, WJ. and Shamma, M. (1984) The emetine alkaloids. Journal of Natural Products. 47, 397-408.
Willaman, J.J. and Schubert, B.G. (1961) Alkaloids bearing plants and their contained alkaloids. United Stated Department of Agriculture, Technical Bulletin. Supt documents,Government print off, Washington DC. 1234,1 -203.
World Conservation Monitoring Centre. (1992). Global Biodiversity - Status of the Earth’s Living Resources. Chapmann & Hall, London.pp 84-86.
Wright, C.W., Allen, D., Cai, Y., Phillipson, J.D., Said, I.M., Kirby, G. and Warhurst, D.C. (1992) In vitro antiamoebic and antiplasmodial activities of alkaloids isolated from Alstonia angustifolia roots. Phytotherapy Research. 6, 121-124.
Wright, C.W., O’Neill, M.J., Phillipson, J.D. and Warhurst, D.C. (1988) Use of microdilution to assess in vitro antiamoebic activities of Brucea javanica fruits, Simarouba amara stem, and a number of quassinoids. Antimicrobial Agents and Chemotherapy. 32, 1725-1729.
Ye, Z.G. and Van Dyke, K. (1989) Selective antimalarial activity of tetrandrine against chloroquine-resistant Plasmodium falciparum. Biochemistry and Biophysic. Research Communications. 159, 242-248.
Yulianti, D.A. and Djamal, R. (1991) Isolation of gramine from Cephaelis stipulacea. ACGC Chemical Research Communication. 1, 22-24.
Zelnik, R. and Rosito, C. (1971) The isolation of angustifolide from Guarea trichilioides. Phytochemistry. 10,1166-1167.
182

other Useful References Consulted
Breitmaier, E. (1993) Structure Elucidation by NMR in Organic Chemistry.
A Practical Guide. John Wiley & Sons. New York, pp.265.
Ernst, R.R., Bodenhausen, G. and Workaun, A. (1987) Principles of Nuclear
Magnetic Resonance In One and Two Dimensions. Clarendon Press. Oxford.
pp. 610.
Furniss, B.S., Hannaford, A.J., Smith, P.W.G and Tatchell. A.R. (1989) Vogei's
Text Book of Practicai Organic Chemistry. 5th Edition. Longman Scientific
& Technical. London, pp. 1514.
Goodman-Gilman, A., Goodman, L.S., Rail, T.W. and Murad, P. (Editors) (1985)
Goodman and Gliman’s. The Pharmacoiogicai Basis of Therapeutics. 7th
Edition. McMillan Publishing Company. New York. pp. 1839.
Harada, N. and Nakanishi, K. (1983) Circular Dichroic Spectroscopy.
Exciton Coupling in Organic Stereochemistry. University Science Books.
Mill Valley, California, pp. 460.
Hostettmann, K. Hostettmann, M. and Marston, A. (1986) Preparative
Chromatography Techniques. Applications in Natural Product isolation.
Springer-Verlag, Berlin, pp. 139.
Jakoby, W.B. and Pastan, I.H. (Editors) (1979) Ceil Culture. Vol. LVIII in the
series: Methods in Enzymology, S.P. Colowick and Kaplan, N.O. Academic
Press Inc. San Diego, California, pp. 642.
Luckner, M. (1990) Secondary Metabolism in Microorganisms, Plants and
Animals. 3rd Edition. Springer-Verlag. London, pp. 563.
183

March, J. (1992) Advanced Organic Chemistry. Reactions Mechanisms, and
Structures. 4th Edition. John Wiley & Sons. New York. pp. 1495.
McLafferty, F.W. and Turecek, F. (1993) interpretation of Mass Spectra. 4th
Edition. University Science Books. Mill Valley, California, pp. 371.
Sanders, J.K.M. and Hunter, B.K. (1987) Modern NMR Spectroscopy. A
Guide for Chemists. Oxford University Press. Oxford, pp. 308.
184

APPENDIX I. SPECTRA

Spectrum # Page #
1 Mass spectrum of 1-hydroxybenzoisochromanquinone 188
2 NMR spectrum of 1-hydroxybenzoisochromanquinone 189
3 NMR spectrum of 1-hydroxybenzoisochromanquinone 190
4 NMR spectrum of 1-acetatebenzoisochromanquinone 191
5 Mass spectrum of benz[g]isoquinoline-5,10-dione 192
6 NMR spectrum of benz[g]isoquinoline-5,10-dione 193
7 NMR spectrum of benz[g]isoquinoline-5,10-dione 194
8 Mass spectrum of vallesiachotamine lactone 195
9 NMR spectrum of vallesiachotamine lactone 196
10 COSY-45 NMR spectrum of vallesiachotamine lactone 197
11 Mass spectrum of vallesiachotamine 198
12 NMR spectrum of vallesiachotamine 199
13 Mass spectrum of strictosidine 200
14 NMR spectrum of strictosidine 201
15 COSY-45 NMR spectrum of strictosidine 202
16 ^H- ®C NMR spectrum of strictosidine 203
17 Mass spectrum of strictosidine lactam 204
18 NMR spectrum of strictosidine lactam 205
19 Mass spectrum of tetraacetyl-strictosidine lactam 206
20 NMR spectrum of tetraacetyl-strictosidine lactam 207
21 NMR spectrum of tetraacetyl-strictosidine lactam 208
22 Mass spectrum of angustine 209
23 Circular dichroism of glomerulatine A 210
24 Mass spectrum of glomerulatine A 211
25 NMR spectrum of glomerulatine A 212
26 ^ C NMR spectrum of glomerulatine A 213
27 ^H-^% NMR spectrum of glomerulatine A 214
28 ^H- ^C long range correlation NMR spectrum of
glomerulatine A 215
29 ROESY NMR spectrum of glomerulatine A 216
30 Mass spectrum of glomerulatine B 217
186

31 NMR spectrum of glomerulatine B 218
32 NMR spectrum of glomerulatine B 219
33 Mass spectrum of glomerulatine C 220
34 NMR spectrum of glomerulatine C 221
35 NMR spectrum of glomerulatine C 222
36 COSY-45 NMR spectrum of glomerulatine 0 223
37 long range heterocorrelation NMR spectrum
of glomerulatine 0 224
38 ROESY NMR spectrum of glomerulatine 0 225
39 Mass spectrum of picramnioside A 226
40 Circular dichroism spectrum of picramnioside A 227
41 NMR spectrum of picramnioside A 228
42 NMR spectrum of picramnioside A 229
43 long range heterocorrelation spectrum of
picramnioside A 230
44 ROESY NMR spectrum of picramnioside A 231
45 Mass spectrum of picramnioside B 232
46 Circular dichroism spectrum of picramnioside B 233
47 NMR spectrum of picramnioside B 234
48 COSY-45 NMR spectrum of picramnioside B 235
49 heterocorrelation spectrum of picramnioside B 236
50 ROESY NMR spectrum of picramnioside B 237
51 Mass spectrum of picramnioside C 238
52 Circular dichroism of picramnioside C 239
53 NMR spectrum of picramnioside C 240
54 ®C NMR spectrum of picramnioside C 241
55 ^H-^% long range heterocorrelation NMR spectrum
of picramnioside C 242
56 ROESY NMR spectrum of picramnioside C 243
57 Mass spectrum of aloe-emodin 244
58 NMR spectrum of aloe-emodin 245
59 Mass spectrum of aloe-emodianthrone 246
187

10195.9085 J80757065605550454835302520
1510
5
93SE270tl xl Bgd=0 01-MRR-93 14 5*01 8pM=8 1=2.4v Hn=900 TIC=173988000 RVÇP4 El 190 DEGC
00 2R8-SE El*RcntULSOP SysDCI
PT=0® CalDRVCRL
58
ilk
128
76
6357
AM
104
83
68
91i i
98
115
U 1 ill till100 120
MRSS=15877000
164184
156
145
|L ui/iikL140
173
212
201230
160 180 200
22
220L 253 267
240 260
Spectrum 1. Mass spectrum of 1-hydroxybenzoisochromanquinone
188
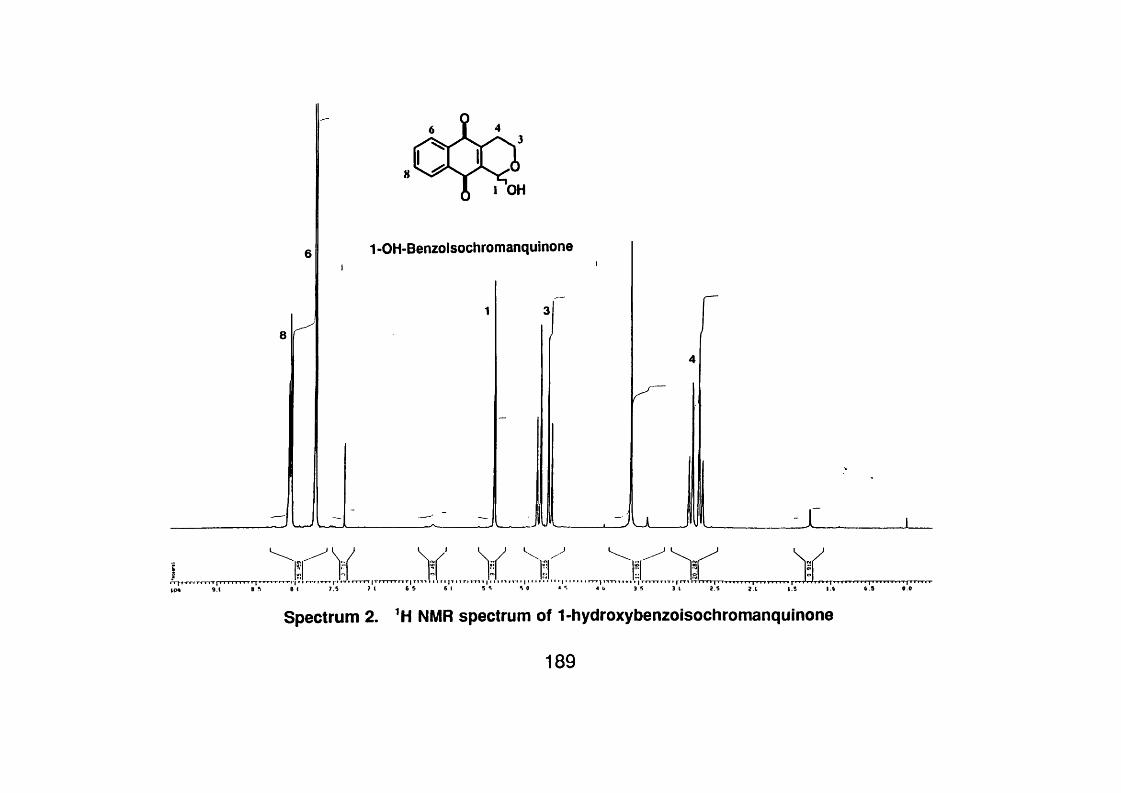
1 OH
1 -GH-Benzoisochromanquinone
_r
4 b 3 3 1 2.5 2 1 1.5 I t t 9 0 0
Spectrum 2. ’H NMR spectrum of 1-hydroxybenzoisochromanquinone
189

18 ÎR '
CC2 i n c n c i 1 t
V \ \ H v yR 5 5 S 5 5 S 5
I Y I
Spectrum 3. "C NMR spectrum of 1-hydroxybenzoisochromanquinone
190
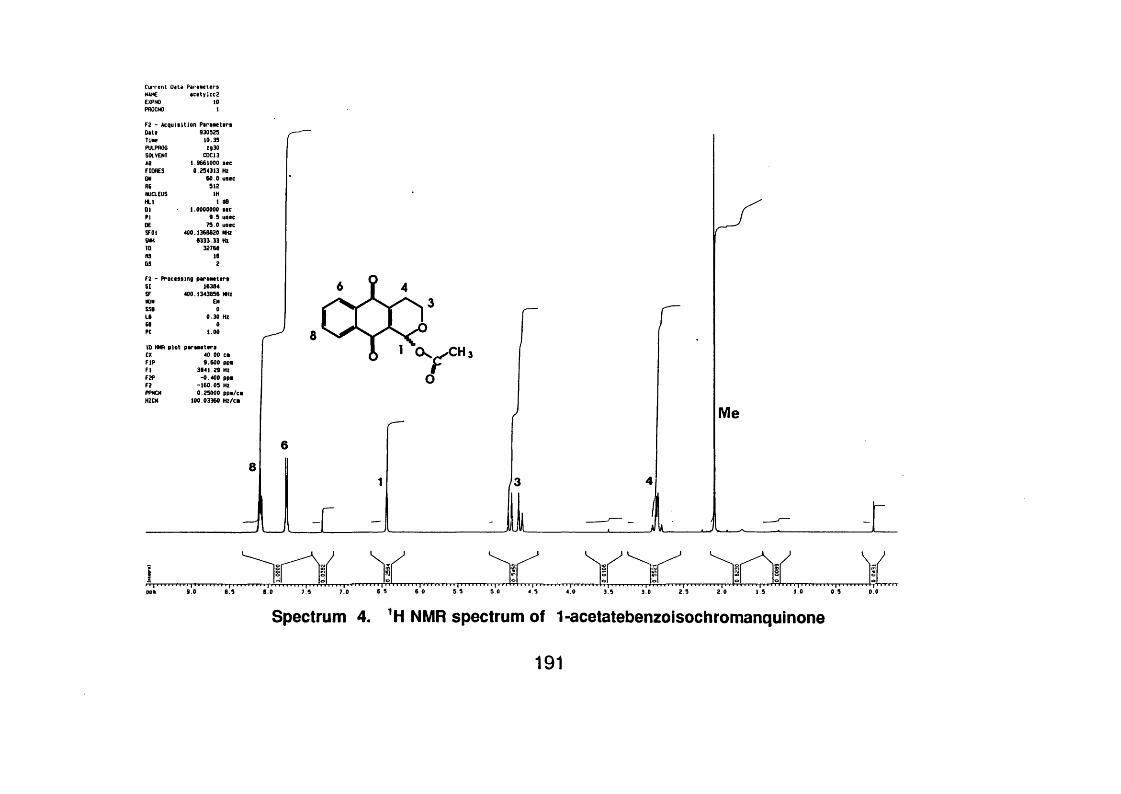
Current Oeta1 Parew tersNAME acety]cc2CXPNO 10pnocNO >
F2 - A cqu is ition P ir iK ts r s930525
T lw 10.35PULPflOG 2930SOLVENT CDC13AO 1.9661000 s tcFlOflES 0.254313 HzON 60.0 wsecAS 912NUCLEUS IHHLl 1 0801 1.0000000 stcFIOESFOl 400.1366620 MHZSMK 6333.33 HZ10 32766MS 16OS 2
SI 16364SF 400.1343656 MHzNON EMSSB 0L I 0.30 Hz66 0FC 1.00
ID NMR p lo t p o rs M ttrscxFIR 9.600 ppBFI 3641 29 HzF2P -0 400 ppmF2 -160.05 HZPPMCM 0.25000 ppm/cmHZCH 100.03360 Hz/cm
J i.
Me
r
Spectrum 4. *H NMR spectrum of 1-acetatebenzoisochromanquinone
191
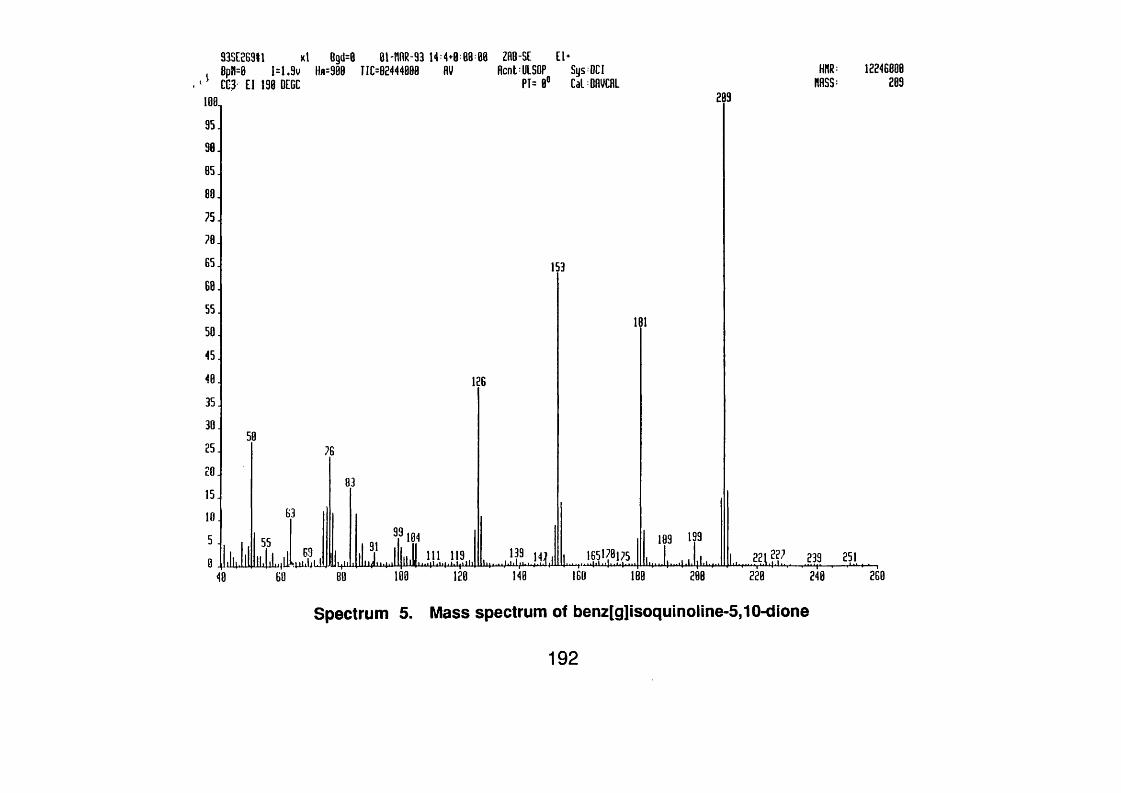
93SC2B9I1 xl Bgd=8 01'nnR-93 14 4*0 88 88 ZflB-SE El*Bpn=0 1=1.9v llfl=980 T1C=82444000 HV Acnt UlSOP Sys OCICC3 El 138 DEGC PT= 0® CaL BflVCflL MHSS:
12246080289
209188,95.90.85.
75.70.65.
25.
??' 239111 119.200 268
Spectrum 5. Mass spectrum of benz[g]isoquinoline-5,10-dione
192

J
J
8
u
Benz[g]isoquinoline-5,10-dione
Spectrum 6. NMR spectrum of benz[g]isoquinorme-5,10-dione
193

s s “i5 55r T «
CCI repeat of 13C using Inverse gated ilecoupiing to give almost q u an t i ta t ive spectrum
Spectrum 7. "C NMR spectrum of benz[g]isoquinoline-5,10-dione
194
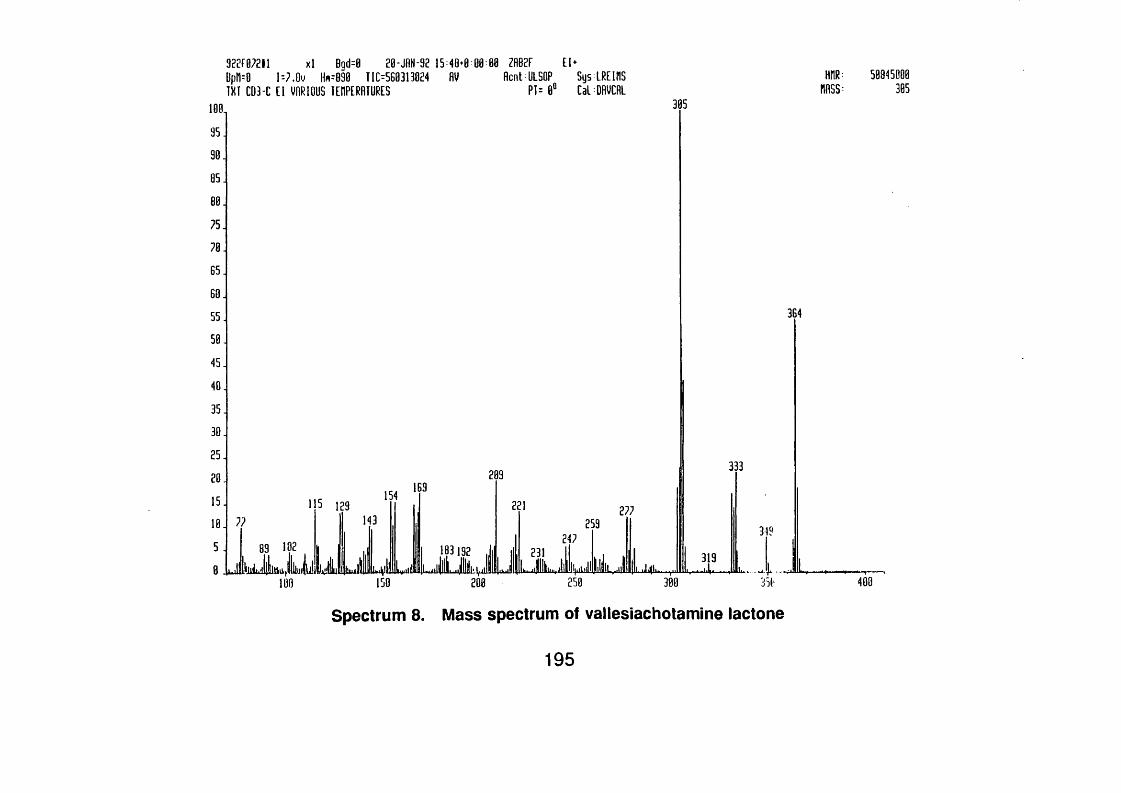
3 ? 2 f0 ? 2 l l x l Bqd=8 2 0 - jn N -9 2 15 48* 0pM=O U 7 .Û V Hm=890 T1C=5B0313024 flV1 X Ï CD3-C [ [ VARIOUS lEHPERAIURES
1 0 8 ZAB2F E l*flcn t ULSOP Sys ERE in s
P 1 = 0 “ CalDRVCRLHMR: 50845000
MASS: 305305100,
7 5 .
60.
4 5 .
3 0 .
3332092 0 .
27731?
. w ll ik llL .illilW lliilklijljlilimylilii hu jjljl L i\lil Lyall! 100 150
319
400300200
Spectrum 8. Mass spectrum of vallesiachotamine lactone
195

CD3-C IN C0CL3 *TMS. MM250 IH
COOMe
PPM/CM
17OMe
15;
14
Spectrum 9. NMR spectrum of vallesiachotamine lactone
196
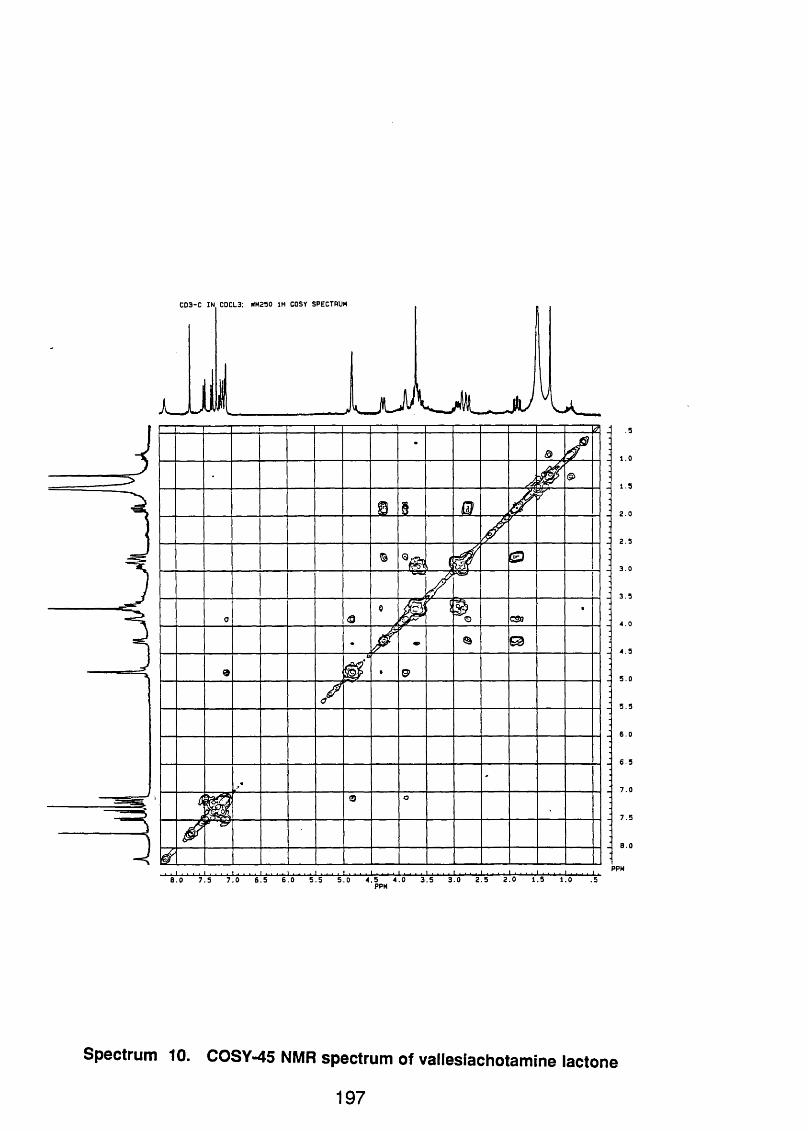
CD3-C IN C0CL3; HM250 IH COSY SPECTRUM
u
csa
. I . . . . I . . . I . . . . I I . . . . I ................ . I . . . . I . . . . I . . . . I . . . . I . ■ ■ ■ 1 . . I . . . . I . . . I .8 .0 7 .5 7 .0 6 .5 6 .0 5 .5 5 .0 4 .5 4 .0 3 .5 3 .0 2 .5 2 .0 1 .5 1 .0 .5
PPM
7 .5
J 8 . 0
PPM
Spectrum 10. COSY-45 NMR spectrum of vallesiachotamine lactone
1 9 7

9 2 2 F 0 4 5 I1 x l Bgd=0 H -J f lN -9 2 10 4 6 *0 Bpn=0 I= 7 .9 m Hn=090 TIC=38297?B944 AV[ 0 2 -H E l 190 DEGC
ZRB2F E l*Rcnt ULSOP Sys IRE 1RS
PT= 8“ Cal ORVCRLHMR: 51504008
MASS 279279100,
9 0 .
7 5 .
6 5 .
6 0 .
5 5 .249
5 0 .
4 5 . 350
307322
3 0 .
2 5 . 209
236
U lM i L> L4 II11
388200 250108 150
Spectrum 11. Mass spectrum of vallesiachotamine
198

C 02-C IN CDCL3 +IM S. NM250 IH
H Z/P I
“COOMe
OMe Me
19
ULi
Spectrum 12. 'H NMR spectrum of vallesiachotamine
199

92SCI72eil xl Bgd=0 18-N0V-92 I1=B>G BpH=8 1=8.5v Hm 28B8 TIC=352BB48B8 HV CDl HNQBfl > Nfl MATRIX
88 ZRB-SE FB*Rcnt-ULSOP SysLMLRFRB
PT= 8* CalFRBCRLHMR:
MASS:55455888
176108,
85.
75.
5 5 .
5 0 .
136 236329
187214128 258 289 307 352 373 309 413 553 604514 648
208 300 408 508
Spectrum 13. Mass spectrum of strictosidine
200

aO Glu
Me(X)C
F2 - A c q u is it io n ParametersDate 930324Tine B.59PULPROG zg30SOLVENT HeOHAQ 1.9660922 secFIORES 0.254314 HzON 60.0 useeRG 512NUCLEUS IHHLl 1 dB01 1.0000000 secPI 9 .5 useeDE 75.0 useeSFOl 400.13B4705 MHZSMH B333.37 HzTO 3276BNS BOS 2
F2 - Processing parametersSI 163B4SF 400.1359667 MHzWOWS5B 0LB 0.00 HZGB 0PC 1.00
10 NMR p lo t parametersCX 40.00 cmF IP 9.600 ppmFI 3B41.31 HZF2P -0.400 ppmF2 -160.05 HzPPMCH 0.25000 ppm/cmHZCH 100.03399 Hz/cm
Spectrum 14. NMR spectrum of strictosidine
201

CD-I COSY 45 spectrum
-8
ppm
Spectrum 15 COSY-45 NMR spectrum of strictosidine
202
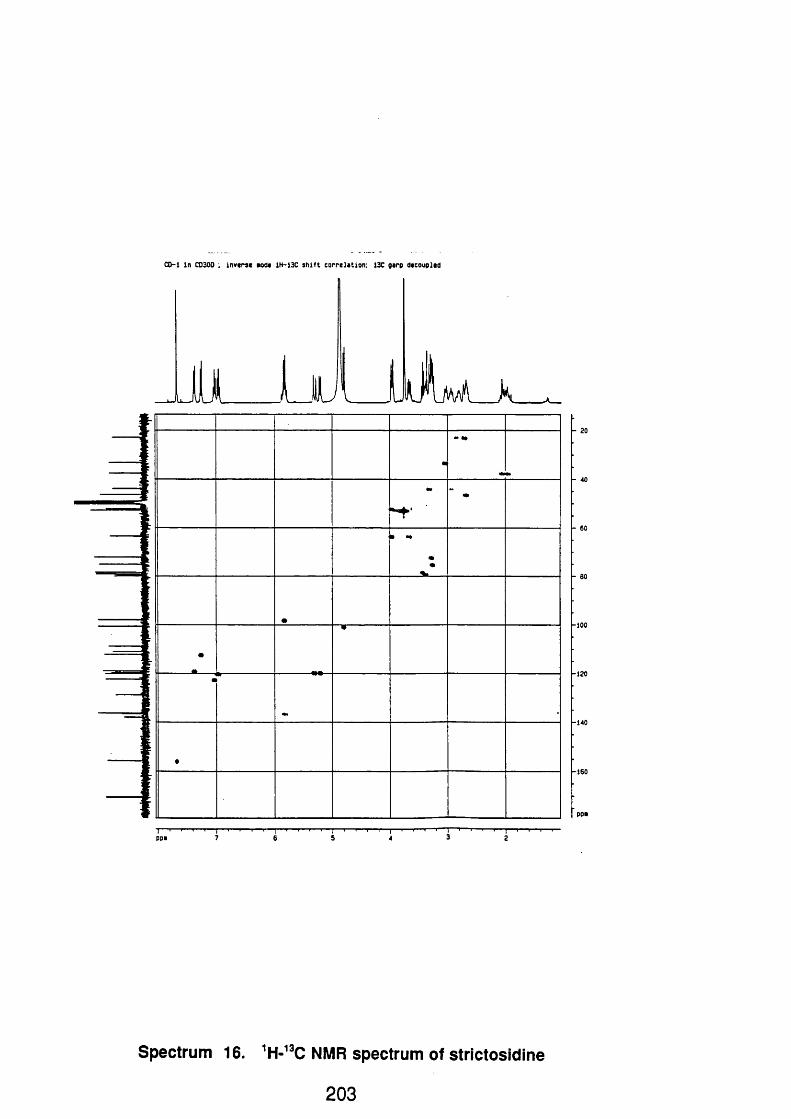
CO-1 in C0300 : inverse mode 1H-13C s h i f t c o rre la tio n : 13C garp decoupled
L
T—' ppm
Spectrum 16. 'H-"C NMR spectrum of strictosidine
203

9 2 2 fB 5 5 ll x l Bgd=0 16 -JR N -92 18 4 9 *0 l = 4 .9 v IU = a s a T IM 1 1 2 G 3 1 0 4 0 AV
C03-H E l VARIOUS TEMPERATURES 108, 235
88 ZAB2F E l*Aa.L ULSOP Sys LREIfIS
PT=8® CalBAVCAL;iUR: 32jJ[038
HASS: 235
498
4 6 7 4 8 0.A -*-288 250 300 358 488 458
Spectrum 17. Mass spectrum of strictosidine iactam
588 5!S
204

H367B8.001 DATE 1 0 -2 -9 2 TIME 12: 09
SF 2 50 .13 4
5 37 5 .0 00 16384 16384 3 75 9 .3 98
r .459SHH Z /P T
PHAOAQAGNSTE
2 .1 7 9200
FW 4700 02 0.1 OP 63L PO
LB
CXCY
4 0 .0 02 3 .0 0
9 .601P - 398P
6 2 .5 27.250
383 8.5 7
F2HZ/CMPPM/CM
18
I I ---------------1................... 1......... . . . I .......... i . . . . , . . . . i ............ . . . I 11 ■■111■«. 111■111. 1. 11. .1 .1. . . 11... . I. . . . 118 .0 0 7 .5 0 7 .0 0 6 .5 0 6 .0 0 5 .5 0 5 .0 0 4 .5 0 4 .0 0 3 .5 0 3 .0 0 - 2 .5 0 2 .0 0 1 .50 1 .0 0 .50 0 .0
PPM
Spectrum 18. 'H NMR spectrum of strictosidine lactam
205
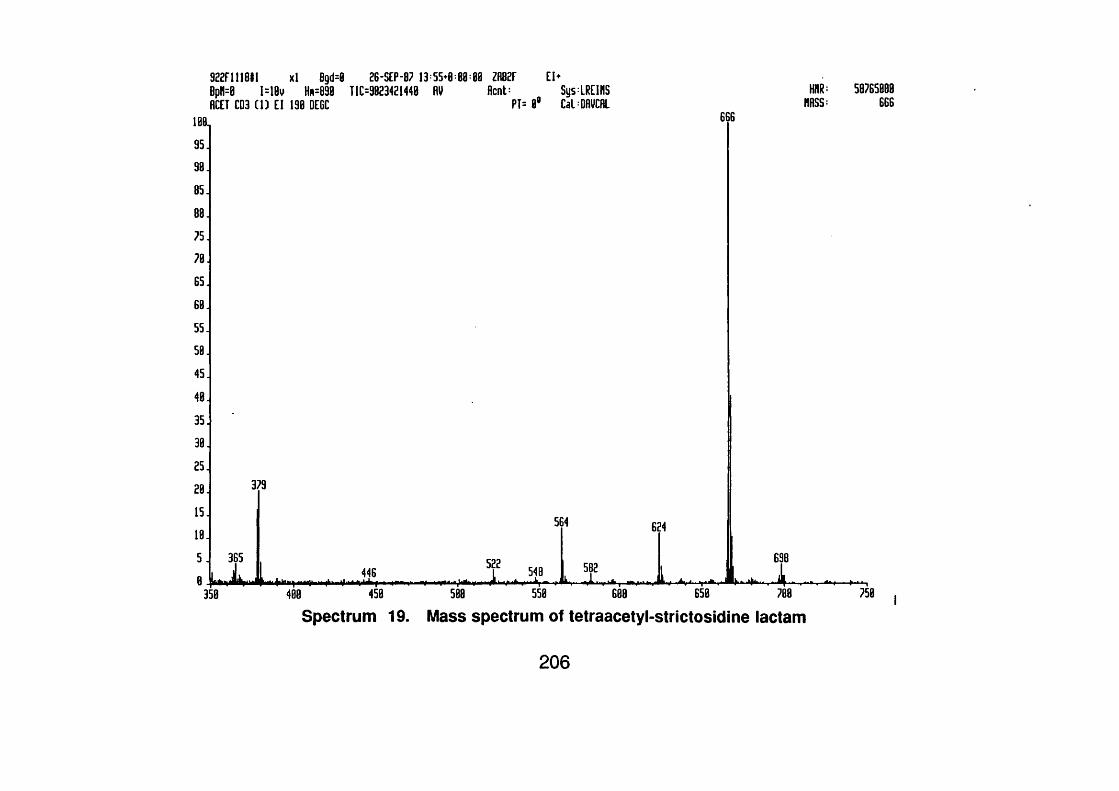
922F1118#1 x l Bgd= BpH=0 I= lB v Hn =890 RCET CD3 ( 1 ) E l 190 DEGC
26-S E P -87 13 55« T IC =9023421440 RV
101
9 5 .
9 0 .
8 5 .
8 8 .
7 5 .
7 0 .
8 5 .
60
55 J
58
4 5 .
4 0 .
3 5 .
3 0 .
2 5 .
2 0 .
1 5 .
1 8 .
5 .
379
1=88 ZR82F E hR en t: Sys:LREinS
P T = 0 ® CalDRVCRL
564
446 I X i l l
582
MRSS:58785800
666666
624
i698X
358 458 588 550 688 650 700
Spectrum 19. Mass spectrum of tetraacetyl-strictosidine lactam750
206

A c e trJ -C D 3 IN C0CL3 t lH S . NN2S0 IH
H 3 9 t50 .00 t
HZ/PT
PPM/CM
I j
Spectrum 20. NMR spectrum of tetraacetyl-strictosidine lactam
207
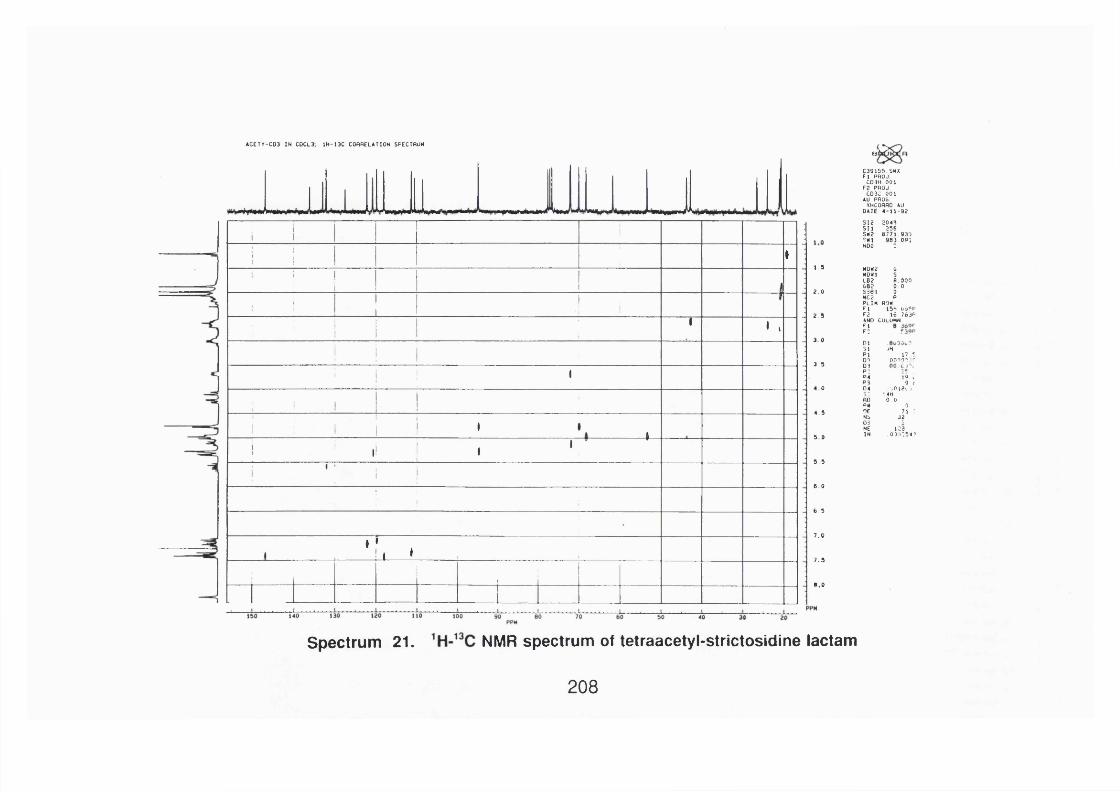
ACETY-C03 IN C0CL3. 1H-13C CORRELATION SPECTRUM
A . J1 1
!
1I
!i g
11
!1
1•; 1
■ i 11
1 ,
! 1 1
' 1 ' 1
1 I I I ' ,
1 ’ 1' 1 1 '
" i l
’
V , ,
1
B j
C39155 SHXF I PflOJ
C03H 001F2 PflOJ
C03C 001AU PflOG
XHCORflO AUDATE 4-11 -92
SI2 2049S Il 256SK2 6771 930? 0 l 983 09:NOG -
1 .5 M0N2 6NOXl SLB2 8.0006B2 0 .0
2 .0 S301 0MC2 pPLIM flowF I 159 t.ÿ^P
2 .5 F2 16 763PAND COLUMNF lF :
3 .0 DI31 iHPI 17 *01 00101.:'"
3 .5 03p :P4 Ï9 I
9 r04 vOiev J3: *.4HflO 0 0PM 0
4.5 DE 71 2Mb 3205ME 123
5 0 IN 0)1:54?
5 .5
6 0
6 .5
7 .0
7 .5
0 .0
Spectrum 21. NMR spectrum of tetraacetyl-strictosidine lactam
208

92Û 0E 24 I1 x l Bgd=0 0 5 -n flR -9 2 14=3*0=00=00 1 2 -250 E l *BpM=0 1 = 4 .2 v Hn=650 110=398292000 HV Rcnt-ULSOP Sys^STEnOEF C D l-8 IH E SCHOOL OF PHHRMHCY PT= fl® C a lU C H L
100. 20
HMR: 27694000MRSS: 28
55 G3 V
313
298
255
229282
150 200 250
Spectrum 22. Mass spectrum of angustine
L300 358
209
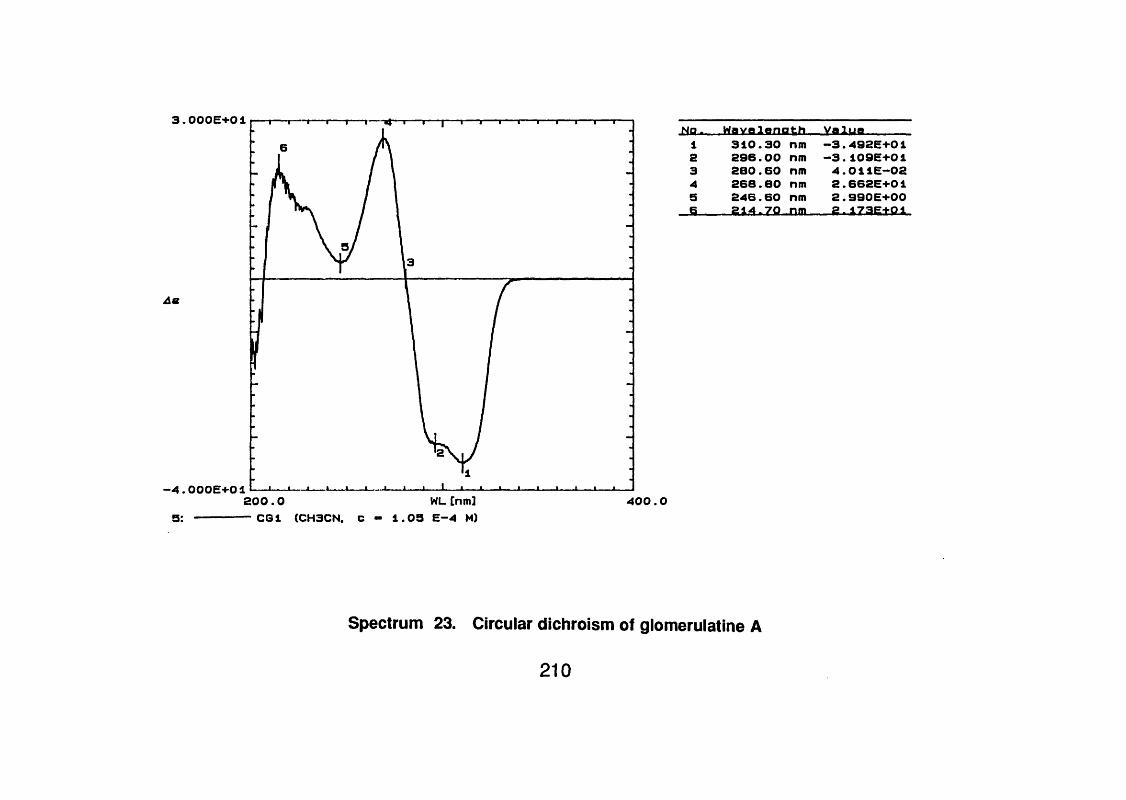
3.000E+01
4.000E+01
5:ZOO . O WL [nm]— CGI (CH3CN, C - 1.05 E-4 M)
N o . Wavalenath Valua1 310.30 nm -3.492E+012 296.00 nm - 3 . 109E+013 280.60 nm 4.011E-024 268.60 nm 2.662E+015 246.60 nm 2.990E+006 . .. 2 1 4 .7 0 .Jim .- 2 .1 7 a E t0 1
400.0
Spectrum 23. Circular dichroism of glomerulatlne A
210

scan 37 ( I . 2 45 mi n i of DPITM: B8 5 3 4 . D
-.1)aciTjTSC
Œ
C R L E D
5 0 1
2 0 91 6 7
3M a s s / C h a r g e
Spectrum 24. Mass spectrum of glomerulatlne A
211
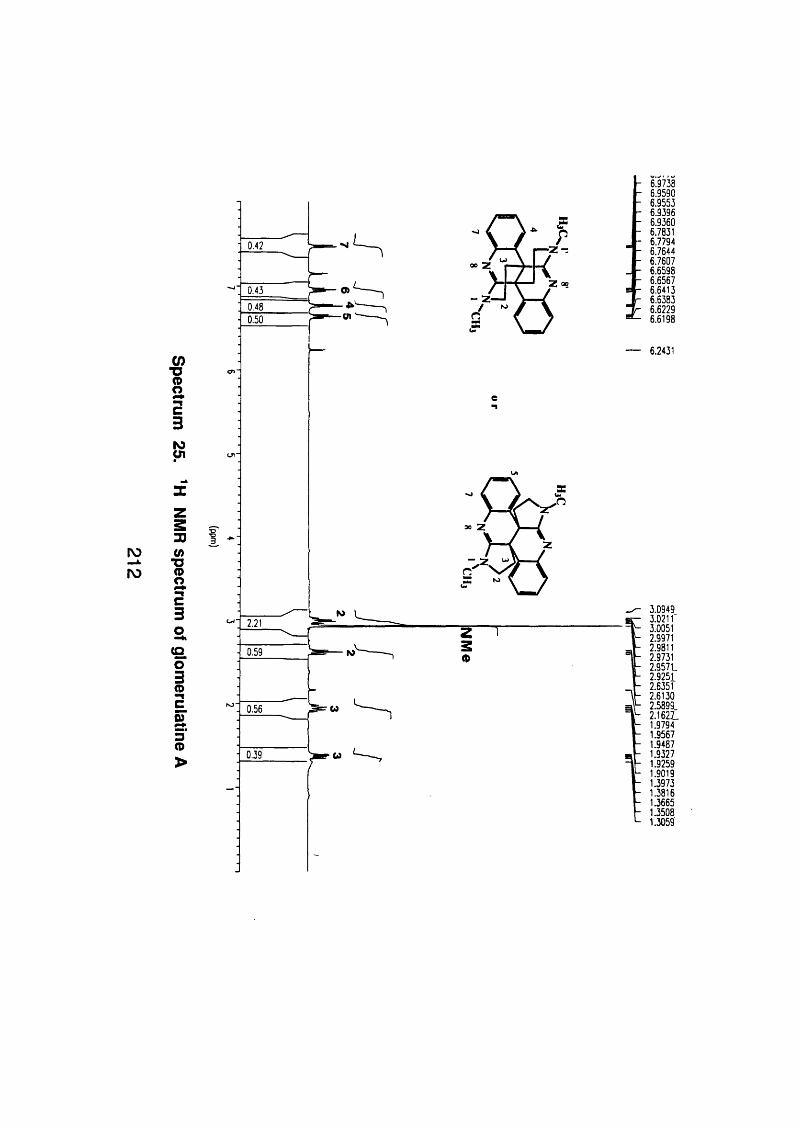
roN)
!23K
zSDUi
%sc3a(Q01cg3<D>
0.42
0.430.480.50
so Z
Î
2.21
0.59
0.56
6.97386.95906.95536.93966.93606.78316.77946.76446.76076.65986.65676.64136.63836.62296.6198
— 6.2431
3.09493.021 r3.00512.99712.98112.97312 .957 L2.92512.63512.61302.5899_2.16 2 L1.97941.95671.94871.93271.92591.90191.39731.38161.36651.35081.3059
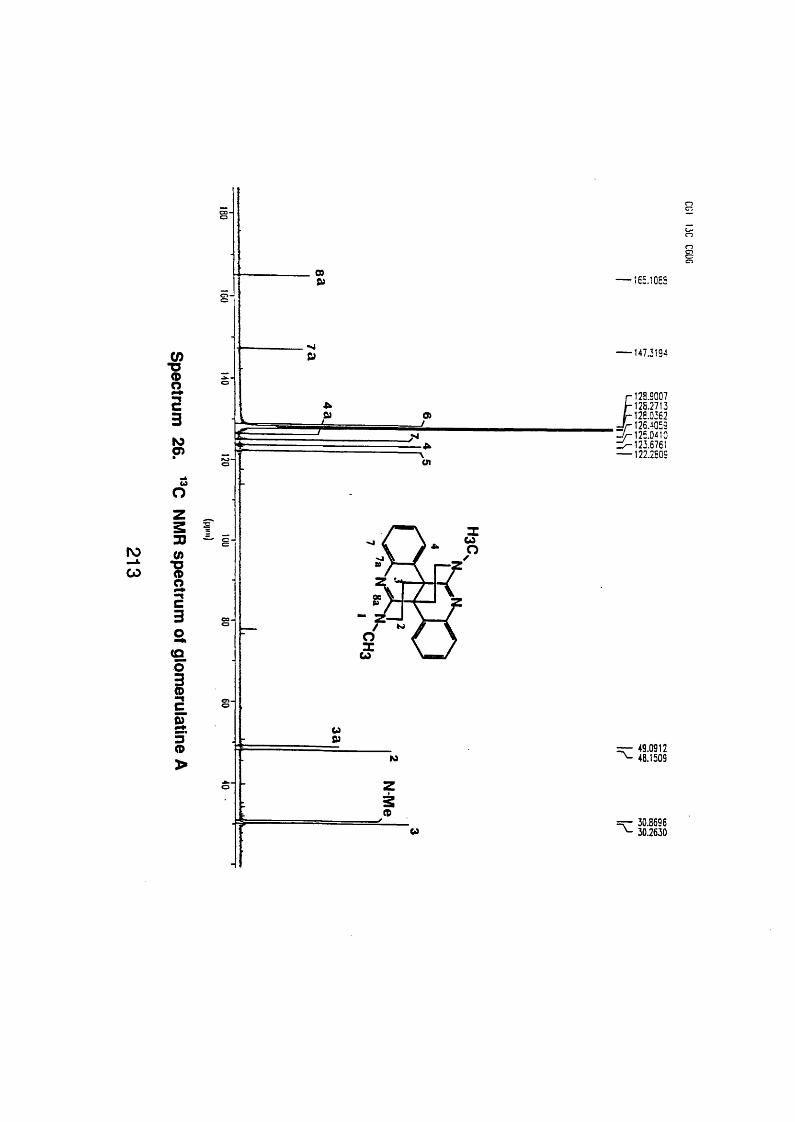
000) — 165.1055
I3toG)
B)
t i a>J
147.3194
J12S.9007 125.2713 125.0362 126.^059
' - ^ 1 2 5 . 0 4 1 0 1 ^ 1 2 3 .6 7 6 1 — 122.2509
roCO
0zs330)
1“tc32.(Q0
1I3<D
W
00
CJ
w&)to
49.091248.1509
ZÈ(D
30.869630.2630
L11 il LL
(ppm)
- -
0 1 «0
0 0
0 oO •0
-
(ppm)
Spectrum 27. 'H-"C NMR spectrum of glomerulatlne A
214
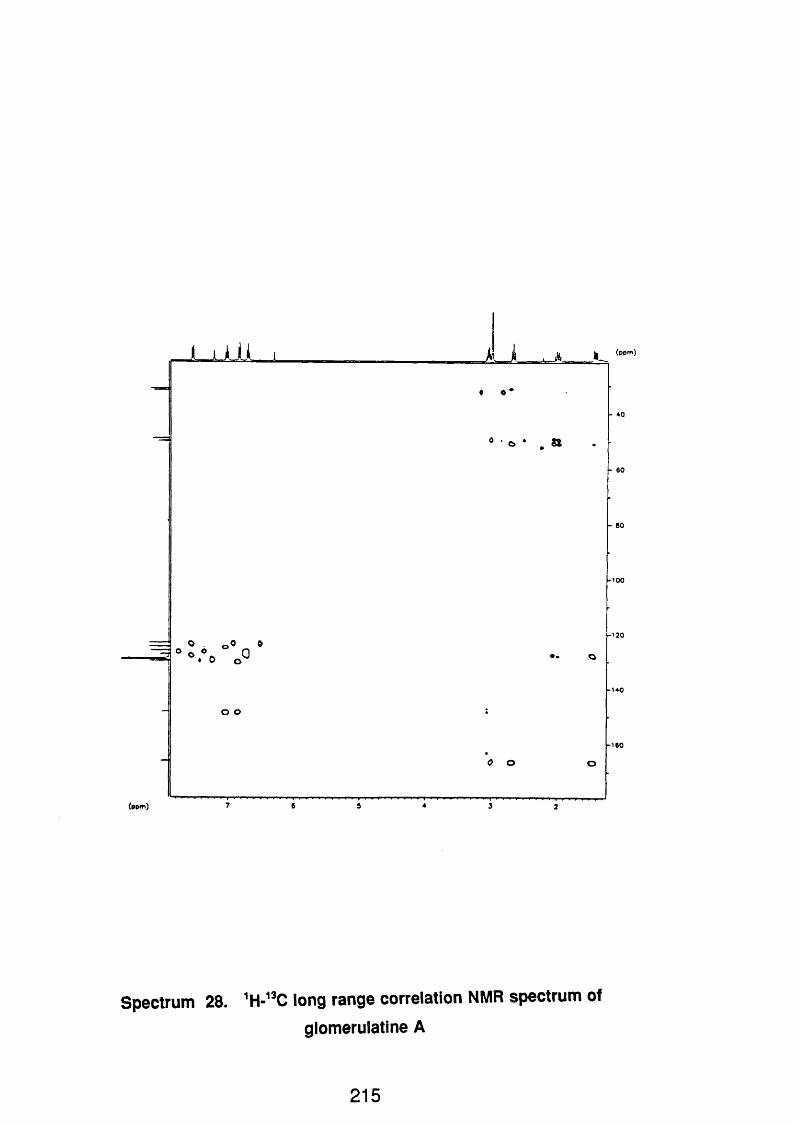
J ü J J -$ «
• o * a
(ppm)
Spectrum 28. long range correlation NMR spectrum of
glomerulatlne A
215

4
■LlIJJ. *___ I
* 0 ^
M • .7 ? °
o
c
0o
oâ1 r
, <91»
(ppm)
Spectrum 29. ROESY NMR spectrum of glomerulatlne A
216

f>e«criptior?: P a h \o i CÛ2 ) E / l . 7 0 ev Te*tp. F 20PC Date: 24-1 I -1993 Time: 13:59: 22
Scans 14 Pr 0:23 Pra<cs«l75 8ase: Mat$-328.16043, rm=l^28fl85
111, ■ J i . . l i l i i l .1' j. I'll piii'l350
tiass
Spectrum 30. Mass spectrum of glomerulatlne B
217

roœ
faC3w
%zs3(/)■OS
3a(O0
1I3(Dm
o o z
09Z
1
1.32
2,59
1.00
7.02746.91336.89456.87456.77986.77036.76106.75066.74266.73986.71806.71436.6992
3.67743.66023.63683.58763.57133.56303.54703.49633.47503.3007
2.52482.51682.50112.4934
1.80081.78361.77931.77041.75591.73931.7242

— 166.7668 8o
— 146.1073
IC3N
J 128.9094 128.8374 124.5720 124.1019
^ 123.6924 J r 122.9417 - ^ 1 2 2 .6 2 7 0 ^ 1 2 1 . 0 7 2 5
roCD
wo
(/)
1c3aCQ01■ncI3<DCD
— 48.9141
— 32.6717 31.5987
^ 30.4423

S c a n 1 0 4 ( 2 . 6 7 5 m i n ) o f D R T R : B 8 5 8 3 . D100:
90-
8 0 '
7 0 '
60 :
c(tj 50 :
3 40':Œ.
30 :
10:
57
1 3 0
100
SCALED
1691 9 7/
2 1 9y
257
d m M a s s - ' ' C h a r g
d m
Spectrum 33. Mass spectrum of glomerulatlne C
220
3 4 3
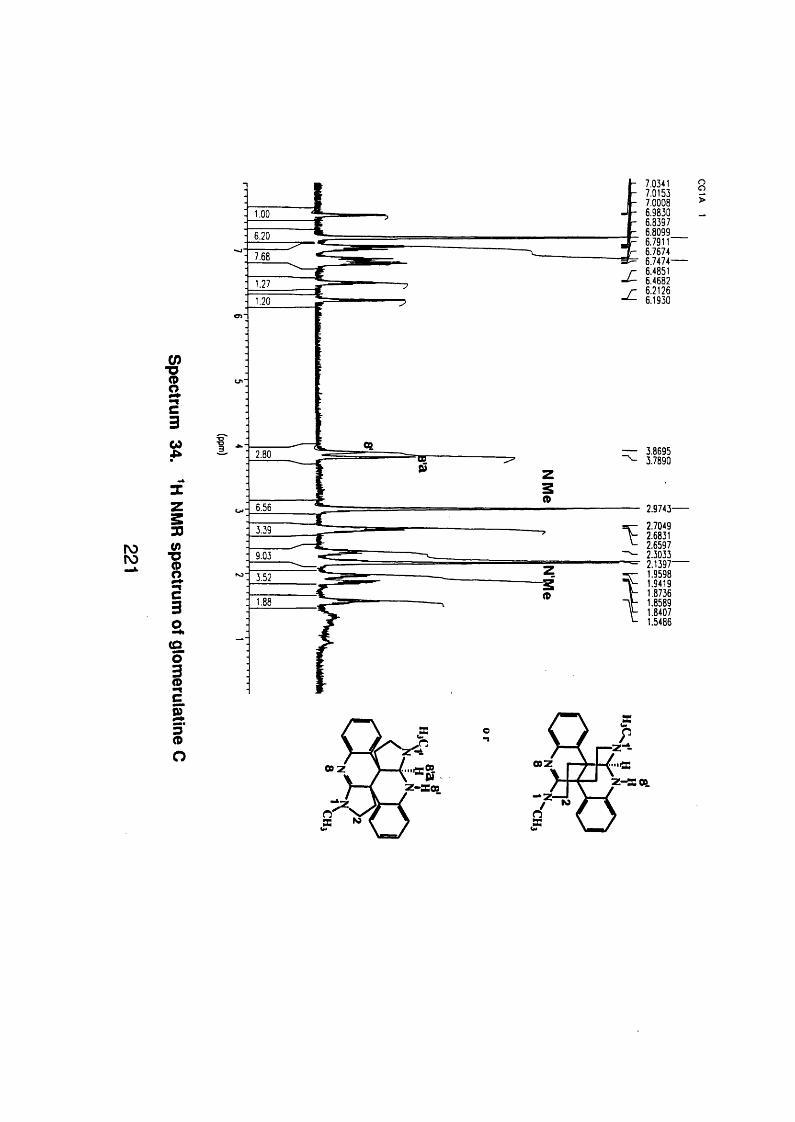
7.03417.01537.00086.98306.83976.80996.7911“6.76746.7474-6.48516.46826.21266.1930
IC3%
%z
0)
I3a(Q0
1
I.sO
— 3.8695 3.7890
2.9743-2.70492.68312.65972.30332.1397-1.95981.94191.87361.85891.84071.5486
Z-ΠO O

— 166.9790 S
I-nC3N
MO
148.1450
— 142.5783
r 129.9993 / r 125.6666
y j - 125.4031 J - 123.4761 T h 122.7226 = ^ 1 2 2 .5 8 7 0
— 118.0042 114.2469
33 10)
1■tc3a
(Q01C
I3(DO
— 76.9857
— 50.9244 49.6277 48.9860
^ 45.7690— 40.0392
34.4866— 32.1852 ^ 31.2052

j j A U i
3
f
(0 O
(
t
> § / ; jP caO
^ «
3 o
y
m
•
o oas o
fo ° °
0 1)
Spectrum 36. COSY-45 NMR spectrum of glomerulatlne C
223
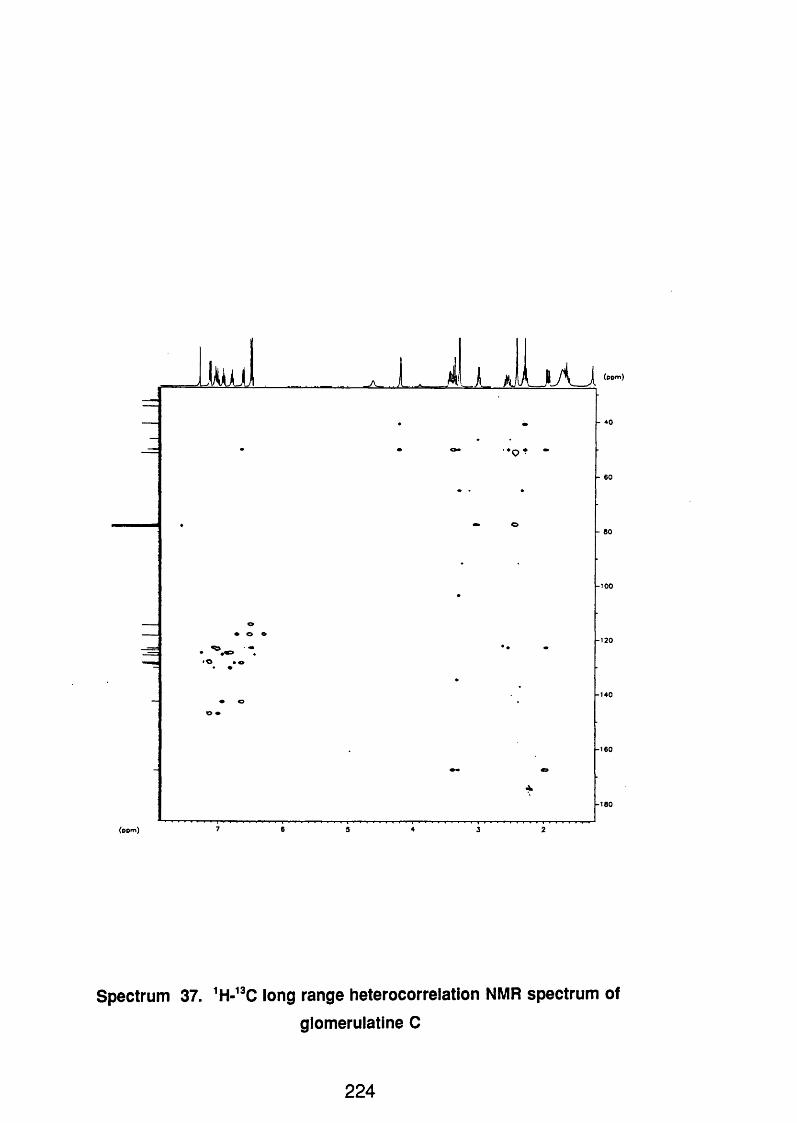
- A - J_I i k, jA J (ppm)
■ ♦ —
(ppm)
Spectrum 37. long range heterocorrelation NMR spectrum of
glomerulatlne C
224
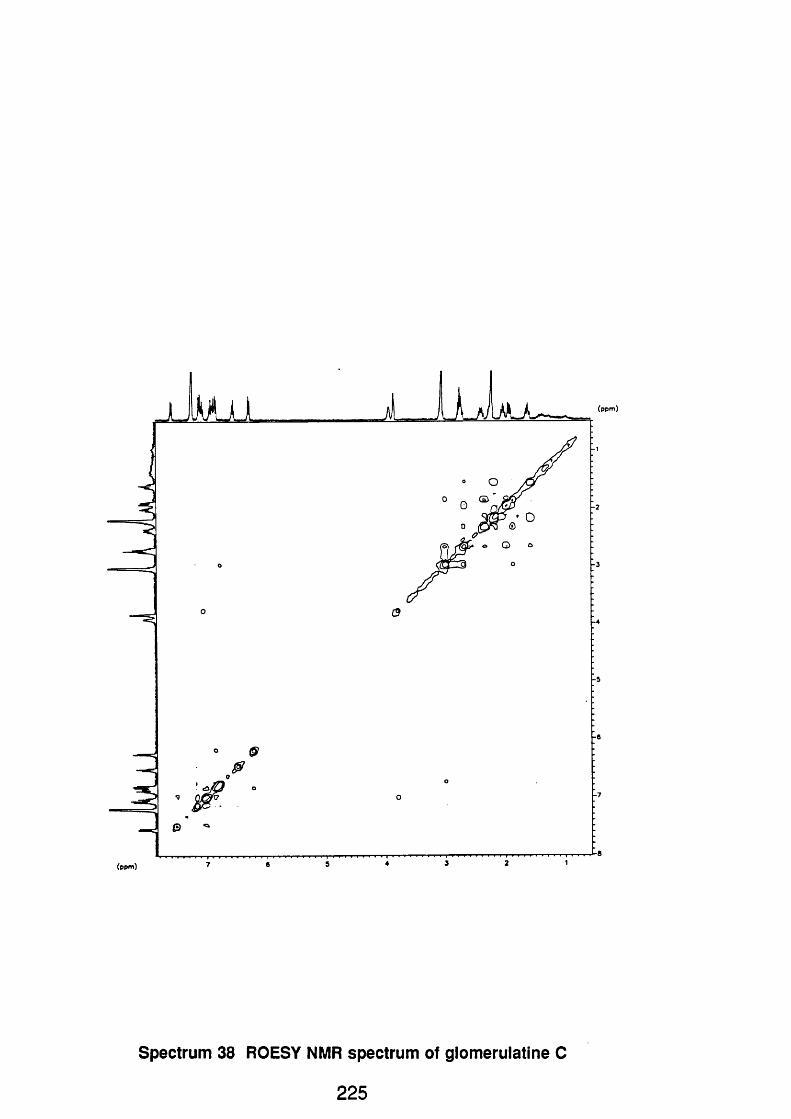
(ppm)
Spectrum 38 ROESY NMR spectrum of glomerulatine C
225

94S E 232I1 x l Bgd=l 14-FE B -94 14M 7>8=B B iflB ZflB-SE FB*BpM=75 I= 4 .5 v H«=1981 T IC=4G ?712932 RV SU R cnt ULSOP Sys LMFHB PRF-3 (flk e V Xe ♦FRB/HS) G L g c /Th logLyc /TFR n a t r L x . PT= 8® C a l FRCHL
I ' 9
I
589
433 459 475 4 M 531
MHSS=18358888
185
4JJ I Î 547iiHmMiHiiinii i I wiiii GB2
588 G88 788
Spectrum 39. Mass spectrum of picramnioside A
226

l.OOOE+Oi
Am
8.000E+00200.0 WL [nm] 000.0
No. WavBlenath Valua1 342.80 nm 1.682E+002 314.20 nm - 2 . 769E-023 290.00 nm -6.363E+004 239.00 nm 1.698E+000 230.80 nm -3.898E+00
. s ___2 2 1 .2 0 nm____l_ ^ 4 4 E t0 1 .
0: PAF3 (Matanol. c - 8 .60 E-O M)
Spectrum 40. Circular dichroism spectrum of picramnioside A
227

OHPAF3
F2 - AcquJilt:»n P a rau te rs OHzgao
I OH
HOH
10 rMR p lo t
J Lv V
Spectrum 41. NMR spectrum of picramnioside A
228
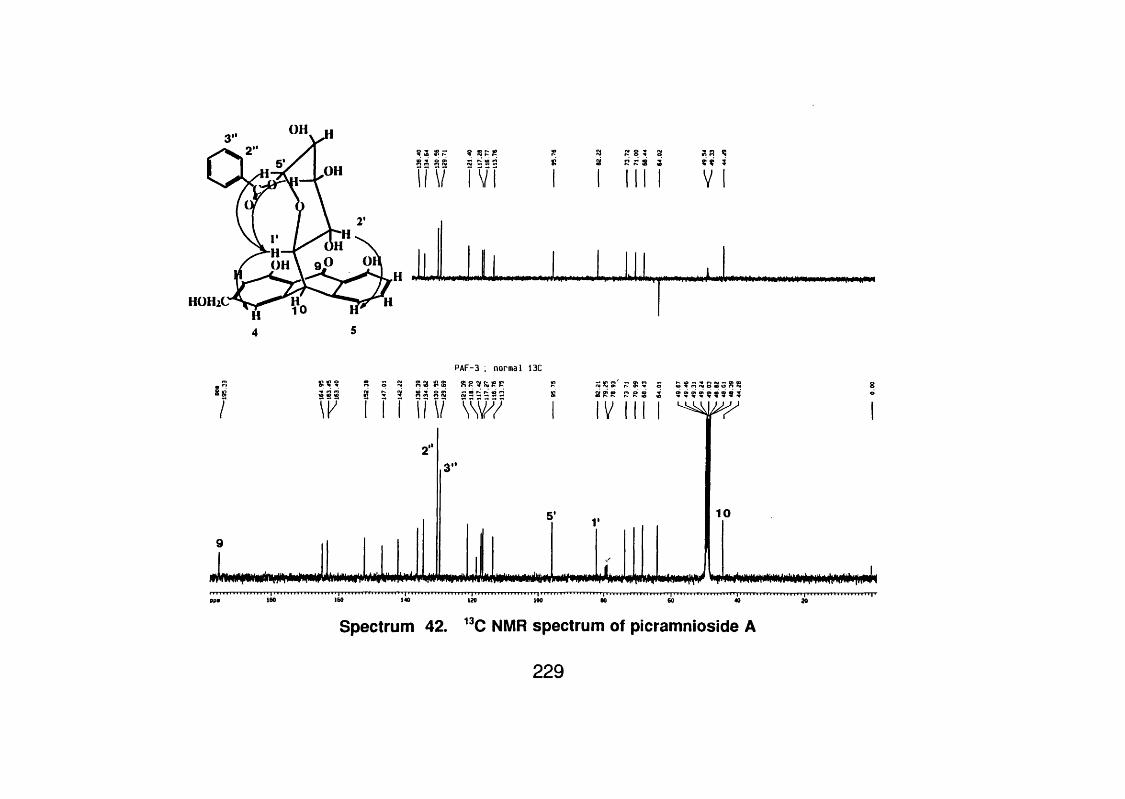
i ï v IVRI RI g 3 8ai RZ3 3
V
HOHiC
PAF-3 ; normal 13C
53 Si8» ;RR8 !î 9 8 P R 5 8 3 2 9 9 3 9 9 9 9 9 3
\ r I I 111 V \ w I IV III I
3"
5’ V 10
PP# 100 160 140 120 100 60 40
Spectrum 42. "C NMR spectrum of picramnioside A
229

PAF 3 ; long rango Invtrtt 1H-13C corrilitlon: 13C coupltd
u
I*,**:*
Spectrum 43. ’H-” C long range heterocorrelatlon spectrum of
picramnioside A
230

PAF3 ROESY spectrum; phase sensitive; positive peaks red;negative black
J
5.0 4.5 4.0 3.5
Spectrum 44. ROESY NMR spectrum of picramnioside A
231

94S E 218I1 x l Bgd=l 13 -FE B -94 IB 55*8=6 BpM=lB5 I= 4 .5 v Hm=1341 T 10=124392888 RV SU PRF-4 CBkeV Xe ♦F fiB /tlS ) G L y /T h L o g ly /T F fl m a t r ix .
256
=88 ZflB-SE FB*flcntULSOP Sys=LMFflB
PT=8® CalFflCflL
447
387
, I ■ I300 350 408 450 500 558
Spectrum 45. Mass spectrum of picramnioside B
HMR=MflSS= 258
658
232

3 . O O O E + 0 0
10
Am
*3, OOOEOO -I— 1.
200.0 W L [n m ] 6 0 0 . 0
No. WavBlenath Value1 399.00 n m 2.666E-012 336.20 n m -2 .314E -023 321.60 nm -9 .670E -014 306.60 n m -4 .969E -035 294.60 n m 1 .246E+006 272.00 n m -2 .644E -037 266.00 n m -9 .499E -01a 240.40 nm 1.066E-019 226.60 n m -2.271E+00
10 210.60 n m __2.4aiE±Q 0
6: P A F 4 ( M a t a n o l . c - 1 . 8 E - 4 M)
Spectrum 46. Circular dichroism spectrum of picramnioside B
233

..ren t D#t# Parvaeters name «U01693*EXPNO 1
F2 - A cqu is ition P s ra o e tir i
OH
OH
F2 - P roc ts ilng p ira a e t fn
Mepp«/4Mz/Cl
10
Spectrum 47. NMR spectrum of picramnioside B
234

PAF4 ; cosy 45
i
_jT V jl-1J
A ful l
00 ^ iB dg
m »'s>->
OGS ,S
3 /• '/Co >
a 1)S07.0 6.5 6.0 5 5 5.0 4.5 4.0 3.5
Spectrum 48. COSY-45 NMR spectrum of picramnioside B
235

.ppa 180 160 140 180 100 80 GO 40 20
Spectrum 49. 'H-"C heterocorrelation spectrum of picramnioside B
236

Ui
j______J
» ► 0|
0 «
_yJL
100
i .f
°0°
v_
5.0 4.5 4.0 3.5
Spectrum 50. ROESY NMR spectrum of picramnioside B
237

94S E 233I1 x l Bgd=l M -F E B 94 17=18^8 88 80 ZHB-SE FB*BpM=75 I=582m v H#=1343 T IC =73958888 RV SU R cnt ULSOP Sys LHFRB PRF-5 (DMSO) (B keV Xe ♦FRB/HS) G ly c /T h lo g ly c / ÎF R m a t r ix . PT= 8 * C a l FRCRL MRSS:
2281808256
2561889 5 .
9 8 .
4476 8 .
5 8 .
4 5 .
239
3 5 .
3 8 .
2 5 .
2 8 .
287 298 548438378283 632
469314
388
Spectrum 51. Mass spectrum of picramnioside C688588288
238

7 . O O O E + O O
G .O O O E + O OWL Inn]200.0
No . M avelanatn Vmlua1 391.40 nm 7.760E-012 317.00 nm -1.8G 0E-023 284.00 nm -5.373E+004 298.60 nm 8.001E -039 253.40 nm i.116E+006 230.00 nm - 3 . 132E+00.7 . . -210 .SO.jn n -___5..4ZOEfOO_
7: PAFO (M atsnol. c - 8 .5E -S M)
Spectrum 52. Circular dichroism of picramnioside 0
239

C urrin t Data Paranettrs N»H£ )n 2 « 3 .3EXPNO nPBOCNO J PAF5
F2 - A c q u ltit lo n Paraaetcrs
OH
OH
iPH OH'F2 - n-ocaastng p a ra a a ttr i
HOH 2'
10 p lo t parameters
Me140.04770 Hx/i i
10
I ..< ■ ■ I .: 3
Spectrum 53. 'H NMR spectrum of picramnioside C
240

V S w ' V Y N i l Y V Y
Ui
? A = R 8 : s â § 3 %R s % s  ofs ? Rx: « ; A s a s i & i i P §& z z z g Biz 9 ? s a s s s : ; £ s s s s s s s s s r s s % 3 g ai ai g a i a i g R a g g * * %%
î W M \ V V Y ST 111 Y \ V I
J##WWMm ill J L mW#wW#ppa 180
Spectrum 54. "C NMR spectrum of picramnioside C
241

PAF5, Ion* ronge 13C-1H co rre la tio n
" '• -
' ♦ » t.
f *•
: 5!i>
•
ppm 180 160 140 120 100 80 60 40 20
Spectrum 55. long range heterocorrelatlon NMR spectrum of
picramnioside C
242

PAF5; ROESr tppi
ÜJ. I. H iAj.- " i
01 • r
1
•M . / -,Vt'w : 4
.
r # \ • •
J
Spectrum 56. ROESY NMR spectrum of picramnioside 0
243

93028111 x l Bgd=0 16-NQ V-87 8 ^ 8 « 8 88 80 1 2 -2 5 8 E hBpH=8 I= 4 .2 v Hm=980 T IC = 150942800 RV Rcnt= SysSTEHDEFPRE 1 E l 1B0OEGC PT= 8 ° C a llC R L
HMR: 27732000HHSS: 278
9 5 .
9 8 .
8 5 .
7 8 .
6 5 .
6 8 .
5 8 .
4 5 .
3 5 .
3 8 .
2 5 .
2 0 .
213224
252
150 208 258
Spectrum 57. Mass spectrum of aloe-emodin
244

C u rrin t D t t i P i r iM t i r i NAME p i l lEXPNO 10PROCNO 1
p lo t p ii M i t i r i
12.600 ppa
0.32500 ppa/ca 130.04430 Hz/ca
Spectrum 58. 'H NMR spectrum of aloe-emodin
245
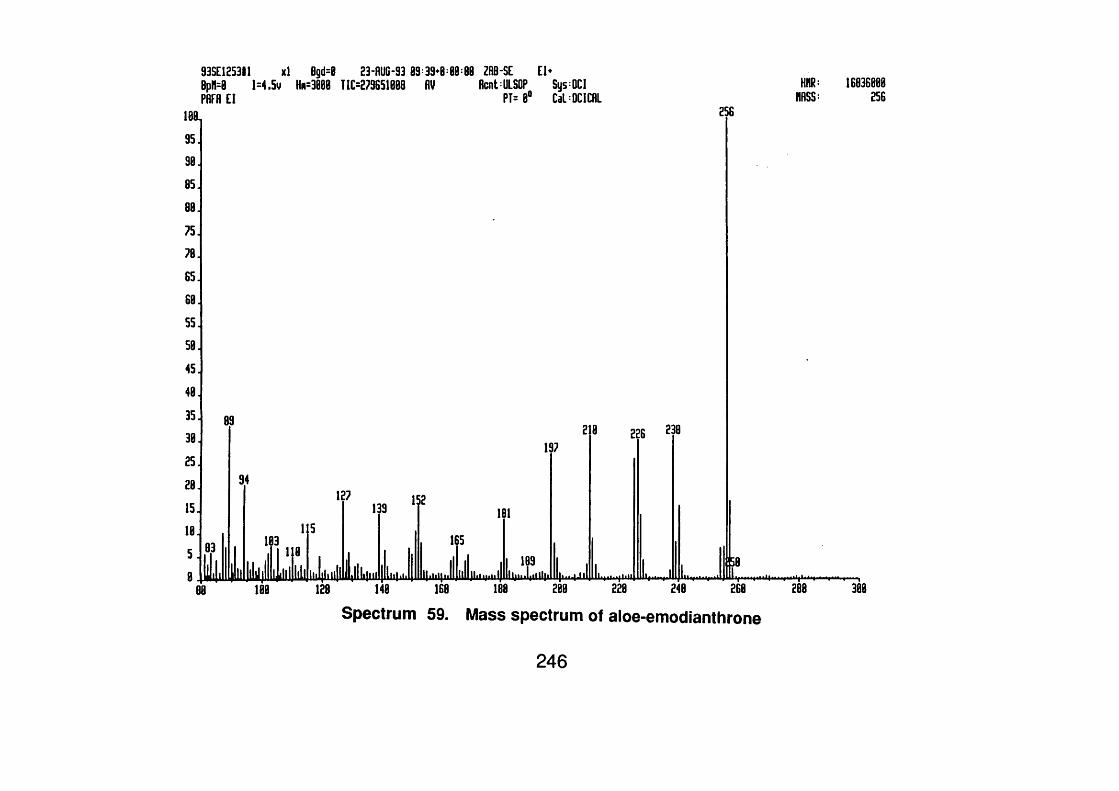
93S E 1253# ! x l BpH=0 1 = 4 ,5 v H#: PflFR E l
23-A U G -93 89 39^ T IC = 2 7 9 6 5 ie B 8 RV
1 8 1
9 5 .
9 8 .
8 5 .
88
75
78
6 5 .
68.5 5 .
5 8 .
4 5 .
4 8 .
3 5 .
3 8 .
2 5 .
2 8 .
1 5 .
1 8 .
5 .
8
89
83
llii
94
188 2R8-SE E l*RcntULSOP SysDCI
PT=8® CalBCICRL
197
i i# 4uiI L
181
165
HHR:HRSS:
16836088256
256
8 226 238
58
128 148 168 188 288 228 248 268 288
Spectrum 59. Mass spectrum of aloe-emodianthrone
308
246

APPENDIX II LIST OF SCIENTIFIC PAPERS

1.- Solis, P.N., Wright, C.W. and Anderson, M.M.: "Brine Shrimp: A Simple
Microwell Bench Top Bioassay". The Square Research Journal. January,
1993.
2.- Solis, P.N., Wright, C.W., Gupta, M.P. and Phillipson, J.D.:"Alkaloids from
Cephaelis dichroa. Phytochemistry. 33(5) 1117-1119, 1993.
3.- Solis, P.M., Wright, C.W., Anderson, M.M., Gupta, M.P. and Phillipson,
J.D.:" A Microwell Cytotoxicity Assay using Artemia salina (Brine Shrimp).
Planta Medica. 59(3), 250 - 252, 1993.
4.- Lang'at, C.G., Allen, D., Solis, P.N., Phillipson, J.D., Watt, R.A., Toth, I.,
Kirby, G.C. and Warhurst, D.C.i'The Activity of Quassinoid Analogues Against
Chloroquine Resistant Plasmodium falciparum in vitro. Proceedings of The 2P
Annual Conference of The U.K. Association of Pharmaceutical Scientists. April,
1993.
5.- Solis, P.N., Wright, C.W., Gupta, M.P. and Phillipson, J.D.: "Cephàelis
Species as Sources of Alkaloids". Proceedings of The Annual Conference
of The U.K. Association of Pharmaceutical Scientists. April, 1993.
6.- Solis, P.N., Gupta, M.P. and Phillipson, J.D.: "Cephaelis Species, A
Chemotaxonomic Dilemma". International Symposium of the Phytochemical
Society of Europe. Lausanne, Switzerland. 29-Sept-1-Oct., 1993.
7.-Lang'at, C.C., Solis, P.N., Watt, R.A., Phillipson, J.D., Kirby, G.C. and
Warhurst, D.C.:"Quassinoid Analogues: Their in v/Yro Antimalarial and Cytotoxic
Activity", international Symposium of the Phytochemicai Society of
Europe. Lausanne, Switzerland. 29-Sept-1-Oct., 1993.
8.- Solis, P.N., Lang'at, C., Gupta, M.P., Kirby, G.C., Warhurst, D.C., and
Phillipson, J.D."Bio-active Compounds From Psychotria camponutans".
248

Submitted to Planta Medica. 1994.
9.- Solis, P.N., Gutierrez-Ravelo, A., Gonzalez, A.G., Gupta, M.P. and
Phillipson, J.D."Bioactive Compounds from Picramnia antidesma subsp.
fessonia". Proceedings of The 3"* Annual Conference of The U.K. Association
of Pharmaceutical Scientists. March, 1994.
10.- Solis, P.M., Gutierrez-Ravelo, A., Gonzalez, A.G., Gupta, M.P. and
Phillipson, J.D."Bioactive Anthraquinone Glycosides from Picramnia antidesma
subsp. fessonia". Submitted to Phytochemistry. 1994.
11.- Solis, P.N., Lang’at, 0., Kirby, G.C., Warhurst, D.C. and Phillipson, J.D.
"Cephaelis Species, Diversity of Bioactive and Novel Compounds". Submitted
to the British Pharmaceutical Conference. 1994.
12.- Solis, P.M., Gutierrez-Ravelo, A., Gonzalez, A.G., Gupta, M.P. and
Phillipson, J.D "Quinoline Alkaloids from Psychotria glomerulata". In
preparation.
249










![International Journal of Research and Review · development of snake venom antagonists especially from plants sources. [6,7] Medicinal plants represent an important source of bioactive](https://static.fdocuments.us/doc/165x107/5e744b8e5142e90872257cf0/international-journal-of-research-and-review-development-of-snake-venom-antagonists.jpg)







