
Bioacoustic analysis of frog calls from northeast India
13
J. Biosci., Vol. 18, Number 3, September 1993, pp 381-393. © Printed in India. Bioacoustic analysis of frog calls from northeast India DEBJANI ROY and ANDREAS ELEPFANDT* Institute of Self Organising Systems and Biophysics, North Eastern Hill University, Mawkynroh-Umshing, Mawlai, Shillong 793 022, India *Department of Biology, University of Konstanz, D-78434 Konstanz, Germany MS received 28 December 1992; revised 30 June 1993 Abstract. Mating calls of three frog species abundant in northeast India Rana tigerina, Rana cyanophlyctis and Rana limnocharis were recorded in the fields of Assam and Meghalaya during their breeding season (July-August, 1991). The calls were analysed for their temporal and spectral characters. They were species specific, with distinct call duration and call period, number of pulses per call and interpulse interval, and dominant frequency and frequency domain. A comparison of the mating calls of Rana cyanophlyctis with those of the sibling Rana ehrenbergi from Yemen showed differences in their temporal and spectral characters, supporting the suggestion that these two species are distinct species, rather than subspecies of the same species. Differences in the temporal and spectral pattern were found in the mating calls of morphologically alike specimens of Rana limnocharis, indicating that the present morphotype Rana limnocharis in northeast India is composed of several species. Keywords. Indian frogs; frog call; bioacoustic analysis; taxonomy. 1. Introduction The anuran mating or advertisement call is now widely considered an important character for species identification, more distinctive than morphological characters. Its function of attracting homospecific females makes the mating call the primary species specific isolating mechanism in anuran speciation (Blair 1964; Littlejohn 1977; Wells 1977; Drewry and Rand 1983; Zimmerman and Bogart 1988). Thus, frogs that, on the basis of morphological criteria, had been considered as belonging to one species may, by analysis of their mating call, turn out to belong to different species. Among recent examples are the well known European lake frog, Rana ridibunda, which now has been shown to be composed of three species (Schneider and Sinsch 1992) and for the water frogs of Greece (Schneider et al 1993). Anuran vocalization is also a valuable tool for studying the ecology and distribution of frogs, once the call of the species under consideration has been determined. Frog calls can be heard from a considerable distance, and often the species can be identified easily by simple listening. One therefore need not catch the individuals which might disturb their population structure and often is also impossible due to the inaccessable terrain. Without interfering with the animals and by merely acoustically localizing the animals one thus can learn a lot about the habitat preference, general behavioural biology and ecology of the animal (Ryan 1986). These advantages made analysis of frog calls a central tool in the study of anuran biology, which now is well established for years all over the world, as in America (Gerhardt 1982; Ryan 1985; Schwartz and Wells 1985; Zelik et al 1991); in Europe (Nevo and Schneider 1976; Schneider and Sinsch 1992; Schneider et al 1993); in 381
-
Upload
debjani-roy -
Category
Documents
-
view
218 -
download
2
Transcript of Bioacoustic analysis of frog calls from northeast India

J. Biosci., Vol. 18, Number 3, September 1993, pp 381-393. © Printed in India. Bioacoustic analysis of frog calls from northeast India
DEBJANI ROY and ANDREAS ELEPFANDT*Institute of Self Organising Systems and Biophysics, North Eastern Hill University, Mawkynroh-Umshing, Mawlai, Shillong 793 022, India *Department of Biology, University of Konstanz, D-78434 Konstanz, Germany MS received 28 December 1992; revised 30 June 1993 Abstract. Mating calls of three frog species abundant in northeast India Rana tigerina, Rana cyanophlyctis and Rana limnocharis were recorded in the fields of Assam and Meghalaya during their breeding season (July-August, 1991). The calls were analysed for their temporal and spectral characters. They were species specific, with distinct callduration and call period, number of pulses per call and interpulse interval, and dominant frequency and frequency domain. A comparison of the mating calls of Rana cyanophlyctis with those of the sibling Rana ehrenbergi from Yemen showed differences in their temporal and spectral characters, supporting the suggestion that these two species are distinct species, rather than subspecies of the same species. Differences in the temporal and spectral pattern were found in the mating calls of morphologically alike specimens of Rana limnocharis, indicating that the present morphotype Rana limnocharis in northeast India is composed of several species. Keywords. Indian frogs; frog call; bioacoustic analysis; taxonomy.
1. Introduction The anuran mating or advertisement call is now widely considered an important character for species identification, more distinctive than morphological characters. Its function of attracting homospecific females makes the mating call the primary species specific isolating mechanism in anuran speciation (Blair 1964; Littlejohn 1977; Wells 1977; Drewry and Rand 1983; Zimmerman and Bogart 1988). Thus, frogs that, on the basis of morphological criteria, had been considered as belonging to one species may, by analysis of their mating call, turn out to belong to different species. Among recent examples are the well known European lake frog, Rana ridibunda, which now has been shown to be composed of three species (Schneider and Sinsch 1992) and for the water frogs of Greece (Schneider et al 1993).
Anuran vocalization is also a valuable tool for studying the ecology and distribution of frogs, once the call of the species under consideration has beendetermined. Frog calls can be heard from a considerable distance, and often the species can be identified easily by simple listening. One therefore need not catch the individuals which might disturb their population structure and often is also impossible due to the inaccessable terrain. Without interfering with the animals and by merely acoustically localizing the animals one thus can learn a lot about the habitat preference, general behavioural biology and ecology of the animal (Ryan 1986).
These advantages made analysis of frog calls a central tool in the study of anuran biology, which now is well established for years all over the world, as in America (Gerhardt 1982; Ryan 1985; Schwartz and Wells 1985; Zelik et al 1991); in Europe (Nevo and Schneider 1976; Schneider and Sinsch 1992; Schneider et al 1993); in
381

382 Debjani Roy and Andreas Elepfandt Australia (Loftus-Hills and Littlejohn 1971) and in Africa (Passmore and Carruthers 1979; Vigny 1979; Passmore 1981). Unlike these areas, very little is known about vocalization characteristics for the Indian species (Kanamadi et al 1992). In view of this obvious importance of bioacoustical studies of anuran vocalization and the abundant distribution of the anurans in the northeastern states (Pillai and Chanda 1976; Inger and Dutta 1986), a survey and analysis of acoustical characters of the calls of the most abundant species Rana tigerina, Rana cyanophlyctis and Rana limnocharis was done. The function of this preliminary report is to demonstrate in the three abundant species, the method and its immense importance for studying the biology and taxonomy of the Indian frogs. The Indian bullfrog, R. tigerina and the skipper frog, R. cyanophlyctis had once been abundant in India but their population has declined as a consequence of intensive capture and selling (Daniels 1991). Knowledge of its place of occurrance and ecology is therefore of importance for its protection. R. cyanophlyctis, is found from India up to Iran, with a sibling semispecies R. cyanophlyctis ehrenbergi living in South Arabia. Lately, however, it has been argued that the South Arabian population should be considered a separate species, R. ehrenbergi (Dubois 1981; Balletto et al 1985). Comparing calls from both the population may shed some additional light on this debate. The Indian cricket frog, R. limnocharis exhibits intraspecific variation (Pillai and Chanda 1979) and is also a very polymorphic species in India (Chanda 1986). In Nepal, the previously supposed R. limnocharis has been shown to be a mixture of four species, only one of them being R. limnocharis sensu strictu (Dubois 1975). Thus, the question arises whether a similar heterogeneity might exist in the Indian R. limnocharis. 2. Materials and methods Calls of R. tigerina, R. cyanophlyctis and R. limnocharis were recorded from Guwahati in Assam and Shillong in Meghalaya during monsoon season (July–August, 1991) coinciding with their breeding period. The average ambient temperature was 28° C at Guwahati and 22° C at Shillong.
The calls were recorded with a directional probe measuring Sennheiser MKH 816 microphone held about 20 to 40 cm from the calling male. The recorded calls were stored on magnetic tapes with a professional HiFi SONY WM-D6C cassette recorder. At the end of each record, the specimen from which the calls were recorded was caught. All these specimens were brought to the Zoological Survey of India, Calcutta, where their species identification was done based on their morphological characters. Acoustical analysis was done on a subsequent visit of DR to the Department of Biology, University of Konstanz, Germany. Oscillogram, sonagram and power spectrum were prepared with a computerized FFT analysis system (Transpek 800) and printed with a laser printer.
Specimen of a F2-population of R. ehrenbergi were obtained from a colony raised from individuals caught in Yemen and maintained in the animal house of the University of Konstanz. The calls were recorded in tanks of the animal house and analysed as mentioned above. Temperature at recording had been raised with a heating lamp to approximately 22° C. The characters analysed were: (i) Call duration (ms) : Duration from the beginning of a call to its end.

Bioacoustic analysis of frog calls from northeast India 383 (ii) Call period (ms) : Duration from the beginning of a call to the
beginning of the next call. (iii) Pulse number : The number of individual components of each
call. (iv) Interpulse interval (ms) : Time between the pulses of a call. (v) Dominant frequency (Hz) : The frequency with maximum intensity (vi) Frequency domain (Hz) : The range of frequencies that differ by less than
10 dB intensity from the dominant frequency. 3. Results The frogs emit mating calls only after dusk. However there was absolutely nocalling if there was no cloud cover or atleast sporadic rainfall during the day. 3.1 R. tigerina R. tigerina were found sitting on land on the edge of swamps. Its call is low pitched and lasts approximately 300 ms, it is produced with a large call period of nearly 1·5 s (figure 1; table 1). The call is very regular, it is composed of about 16 pulses with only 18 ms interpulse interval, so that it sounds as one single sound for the human listener. A smooth increase and then decrease of pulse intensity from the begin to the end of the call gives the call, despite its considerable loudness, a soft sound like "oong" (Daniel 1975). The dominant frequency is at 520 Hz, with a third harmonic of about equal amplitude at about 1650 Hz. The soft and low pitched character of the call makes it clearly distinguishable from other calls. 3.2 R. cyanophlyctis R. cyanophlyctis were found in abundance. They are aquatic, found mostly submerged or afloat 20 cm deep. Calls were emitted while hovering at water surface. These frogs were the most nervous types amongst all other species studied. Approach of the observer, sometimes other frogs or even sounds that were abiotically caused, such as splashing of water or rustling of leaves, usually made them to escape several meters away. Calls lasted somewhat more than half a second, about 615 ms and were emitted at 2–3 s interval (figure 2; table 1). The call was composed of 6–8 pulses with quite constant long interpulse interval of 85 ms, giving the call a rattle-like sound. Frequency distribution is bimodal with dominant frequencies at about 1650 Hz and its second harmonic.
In contrast to these regular calls of R. cyanophlyctis from India, the calls from the Yemenite R. ehrenbergi were more irregular and differed in many respects (figure 3; table 1). First, each call lasted about 100 ms and was composed of about 10–12 pulses with short interpulse interval. Due to reflections, the exact number of pulses and their intervals could not be determined exactly, but the difference from R. cyanophlyctis is clearly evident. Second, calls are emitted at highly varying rates, from a single call per second over small groups of calls with about 150 ms call period up to long series of calls that last more than 6–8 s with the same short call period, so that one may doubt which unit should be named a "call". Finally, the
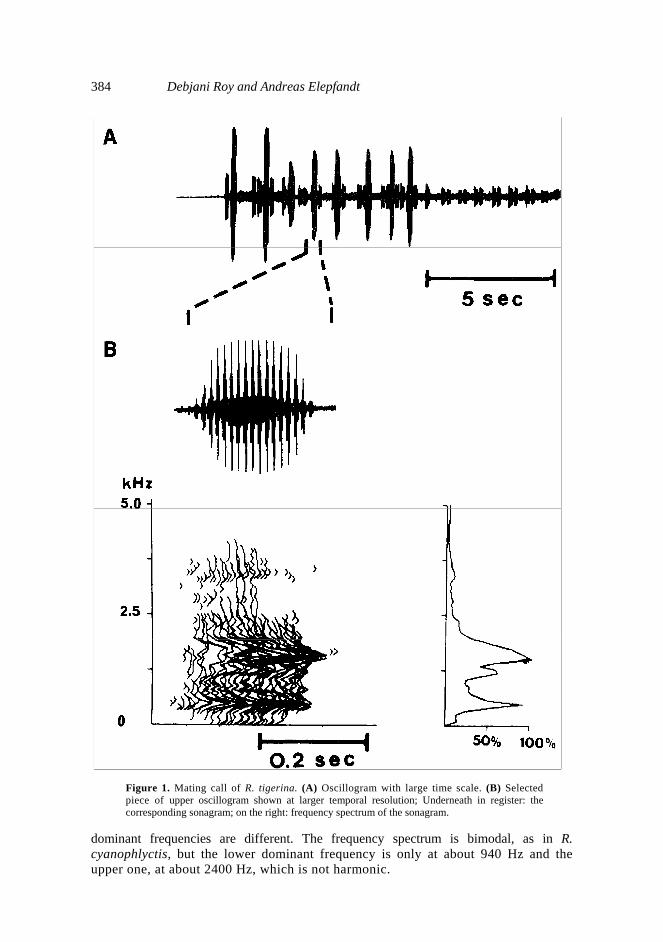
384 Debjani Roy and Andreas Elepfandt
Figure 1. Mating call of R. tigerina. (A) Oscillogram with large time scale. (B) Selected piece of upper oscillogram shown at larger temporal resolution; Underneath in register: the corresponding sonagram; on the right: frequency spectrum of the sonagram.
dominant frequencies are different. The frequency spectrum is bimodal, as in R. cyanophlyctis, but the lower dominant frequency is only at about 940 Hz and the upper one, at about 2400 Hz, which is not harmonic.

Bioacoustic analysis of frog calls from northeast India 385
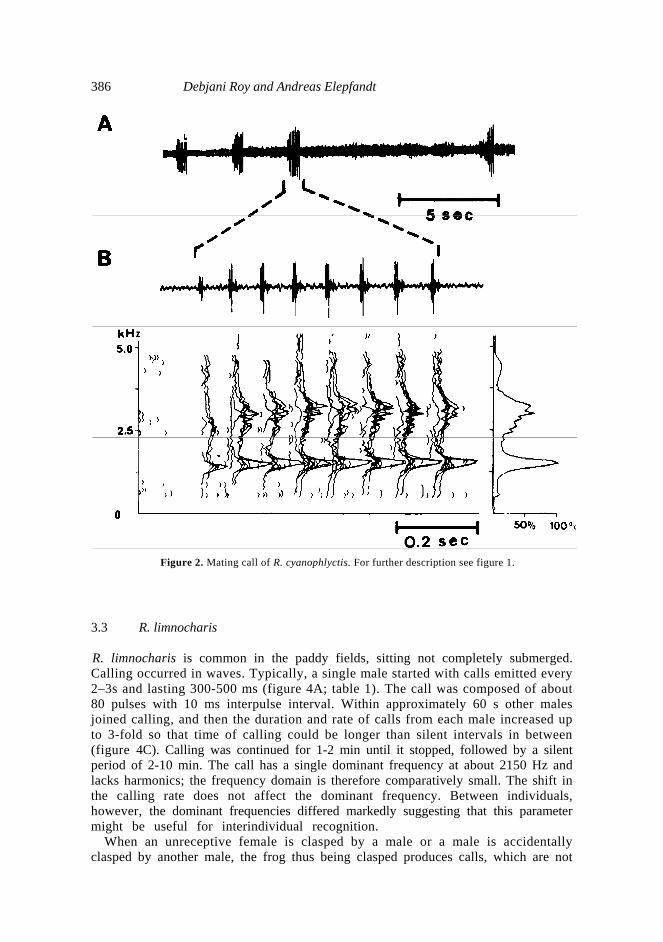
386 Debjani Roy and Andreas Elepfandt
Figure 2. Mating call of R. cyanophlyctis. For further description see figure 1. 3.3 R. limnocharis R. limnocharis is common in the paddy fields, sitting not completely submerged. Calling occurred in waves. Typically, a single male started with calls emitted every 2–3s and lasting 300-500 ms (figure 4A; table 1). The call was composed of about 80 pulses with 10 ms interpulse interval. Within approximately 60 s other males joined calling, and then the duration and rate of calls from each male increased up to 3-fold so that time of calling could be longer than silent intervals in between (figure 4C). Calling was continued for 1-2 min until it stopped, followed by a silent period of 2-10 min. The call has a single dominant frequency at about 2150 Hz and lacks harmonics; the frequency domain is therefore comparatively small. The shift in the calling rate does not affect the dominant frequency. Between individuals, however, the dominant frequencies differed markedly suggesting that this parameter might be useful for interindividual recognition.
When an unreceptive female is clasped by a male or a male is accidentally clasped by another male, the frog thus being clasped produces calls, which are not

Bioacoustic analysis of frog calls from northeast India 387
Figure 3 Mating call of R. ehrenbergi. For further description see figure 1.
in any respect similar to the mating calls (figure 5; table 1) They are obviously the release calls. The calls were emitted in less than 1 s interval and each call lasted about 100 ms, composed of about 10 pulses with 11 ms interpulse interval. It has a wide frequency domain from about 320 to 2400 Hz, showing no dominant frequency in their sonagram.
3.4 Other frogs morphologically considered as R. limnocharis In ponds near Guwahati, two more distinct advertisement calls were recorded from unknown frogs. Specimens of the calling male were identified morphologically as R. limnocharis. However the calls of these frogs strongly differ from the advertisement calls of R. limnocharis described earlier.
The calls of the first kind of frog, named sp. 1 is as follows: There were 3–4 calls per second, with each call lasting approximately 100 ms. Each call has 5–6 pulses, with 15–20 ms interpulse interval. It has a wide frequency domain. The dominant frequency is at about 1000 Hz with a second harmonic of equal amplitude at about 2000 Hz (figure 6).

388 Debjani Roy and Andreas Elepfandt
Figure 4. Mating call of R. limnocharis. For further description see figure 1.
The calls of the second kind of frog, named sp. 2 is as follows: There were 3–4 calls per second with each call lasting approximately 150 ms. It has 9 pulses with 20 ms

Bioacoustic analysis of frog calls from northeast India
Figure 5. Release call of R. limnocharis. For further description see figure 1.
interpulse interval. Dominant frequency is at about 1050 Hz, with a secondharmonic at about 2200 Hz of about equal amplitude (figure 7).
The most obvious and common difference of these calls from that of R. limnocharis, is the harmonic frequency distribution with peaks at about 1000 and 2000 Hz for sp. 1 and peaks at about 1050 and 2200 Hz for sp. 2, while R. limnocharis has only one frequency at about 2150 Hz. Also the calls are emitted much more regularly than in R. limnocharis, no chorusing reaction was observed.
389

390 Debjani Roy and Andreas Elepfandt
Figure 6. Mating call of a specimen of R. limnocharis morphotype called sp. 1 in text. Forfurther description see figure 1.
The number of pulses in a call is small and the interpulse interval more than twice than in R. limnocharis. Whilst these differences from R. limnocharis are common for both species, their calls differ from each other by the number of pulses per call, which is 5–6 in sp. 1 and 9 in sp. 2. Also the call period is relatively stable in sp. 2and less so in sp. 1. These differences were clearly heard. 4. Discussion In the present paper, acoustic characters of calls from three species abundant in the northeast India are discussed. The most distinctive call is that of R. tigerina. It is very low pitched, more than an octave lower than from any other frog in this area. In addition, it is, despite its loudness, a soft call ("oong"). The calls of all other frogs are much sharper like clicks and rattles. Thus, it is extremely easy to identify this species on the basis of its call and use its call for its habitat determination.

Bioacoustic analysis of frog calls from northeast India 391
Figure 7. Mating call of a specimen of R. limnocharis morphotype called sp. 2 in text. Forfurther description see figure 1.
R. cyanophlyctis possesses a relatively constant call, consisting of a few pulses with long interpulse intervals, giving the call a rattle like character. The interpulse interval was by far the longest of all species. Call period is 2 s or longer. The call from the Yemenite R. ehrenbergi, on the other hand, is strongly different. Its call may occur seperately, in small groups or in large sequences with the call period being typically only 1/7 s, the interpulse interval is short, and the dominant frequencies different. These characters make the call completely different from that of R. cyanophlyctis. With regard to the differences in the dominant frequencies one has to take the different recording situation into account. The records from R. ehrenbergi were possibly due to the recordings done from older specimens than the records from R. cyanophlyctis, and this factor is known to decrease the dominantfrequency in the frogs. However, the difference of nearly an octave appears too large to be accounted for by these differences in the recording condition. Also the ratio of the dominant frequencies, which was harmonic in R. cyanophlyctis and non-harmonic in R. ehrenbergi, should also not be affected by the differences in recording. As a conclusion, the difference of the advertisement calls in R. cyanophlyctis and in R. ehrenbergi strongly support the seperation of R. enrenbergias a species on its own from the Indian R. cyanophlyctis.

392 Debjani Roy and Andreas Elepfandt Three different advertisement calls were recorded from frogs, all of which were
identified morphologically as belonging to R. limnocharis. These differences suggest that the so called R. limnocharis in India is not one species, but a composition of several species of similar morphological appearance. Our data clearly suggests the necessity of re-examination of R. limnocharis in north east India with a combined bioacoustic, as well as morphological study. It was well known that R. limnocharis is polymorphic (Chanda 1986), but our data show that this morphological variability can atleast in part be contributed to the existence of several species.
It is interesting to note that also in Nepal, which is neighbouring northeast India, the previous R. limnocharis has been subdivided into four species, namely R. limnocharis proper, R. syhadrensis, R. nepalensis R. pierrei, all of them belonging to one R. limnocharis group (Dubois 1975). Their seperation has been done on the basis of the species mating calls. Unfortunately, the data presented in the short note of Dubois allow only a very limited comparison with our results. The R. limnocharis referred by Dubois shows the same harmonic frequencies as our sp. 1 and sp. 2, and the number of pulses per call is given as 5–6 like in sp. 1. The calls from the other three species have no similarity to our records. This might mean that even the short recordings of Dubois (1975) and ours immediately reveal the existence of 6–7 species, all originally considered to be R. limnocharis.
A common feature of Nepal and of northeast India is the existence of mountains in close proximity to lowlands. Thus, both areas comprise very different climatic zones, changing with altitude, and zones with similar climate may be seperated on close range, by zones of different climate. These areas have been found favourable for speciation. The anurans make up 91 % species of Indian amphibians, which is just slightly more than their percentage on a world wide basis (Inger and Dutta 1986). The abundance of species of amphibians is very uneven across India. The highest concentration of species and genera are in the northeast and in the western ghats of the West Peninsular region. In the north-eastern region alone 54 anuran species have been reported on the basis of purely morphological criteria, and the species show strong differences in their altitudinal distribution (Pillai and Chanda 1976; Chanda 1990). Inclusion of call analysis will certainly reveal an even higher number of species. This marks northeast India as a "hot spot" for diverse frog speciation within close range to each other and as a favourable area for the study of the process of speciation. Acknowledgements
We thank Μ Κ Chandrashekaran for helpful comments on the manuscript, andΚ Ρ Sinha for supporting the work. Thanks are also due to S Κ Chanda for morphological identification of the specimens which was crucial to this study. Weare grateful to Ε Zimmermann for providing FFT analysis system, Ρ Elepfandt for assistance in sound recording, and I Eistetter for support in preparing the graphs. The work was supported by the INSA-DFG Bilateral Exchange Programme. References Balletto E, Cherchi Μ A and Gasperetti J 1985 Amphibians of the Arabian peninsula; Fauna Saudi
Arabia 7 319–392

Bioacoustic analysis of frog calls from northeast India 393
Blair W F 1964 Isolating mechanisms and interspecies interactions in anuran amphibians; Q. Rev. Biol. 39 333-344
Chanda S Κ 1986 A study of anuran (Amphibia) fauna of northeast India, Ph.D. thesis, Kalyani University, Kalyani
Chanda S Κ 1990 Amphibian (anura) species and their altitudinal distribution in northeast India; J. Bombay Nat. Hist. Soc. 87 157-158
Daniel J C 1975 Field guide to the amphibians of western India; J. Bombay Nat. Hist. Soc. 72 506-522 Daniels R J R 1991 The problem of conserving amphibians in the Western Ghats, India; Curr. Sci. 60
630-632 Drewry G Ε and Rand A S 1983 Characteristics of an acoustic community; Puerto Rican frogs of the
genus Eleutherodactylus; Copeia 1984 941-953 Dubois A 1975 Un nouveau complexe d'especes jumelles distinguees par le chant: les grenouilles du
Nepal voisines de Rana limnocharis Boie (amphibiens, anoures); C. R. Acad. Sci. Paris D281 1717-1720 Dubois A 1981 Liste des genres et sous-genres nominaux de ranoidea (amphibiens, anoures) du monde,
avec identification de leurs espece-types: consequences nomenclaturales; Monit. Zool. Ital Suppl. 15 226-284
Gerhardt Η C 1982 Sound pattern recognition in some North American tree frogs (Anura: Hylidae) for mate choice; Ann. Zool. 22 581-595
Inger R F and Dutta S Κ 1986 An overview of the amphibian fauna of India; J. Bombay Nat. Hist. Soc. (Suppl.) 83 135-146
Kanamadi R D, Hiremath C R and Schneider Η 1992 Vocalization and breeding in South Indian tropical frog Rana crassa (Jerdon); Zool. Anz. 228 26-31
Littlejohn Μ J 1977 Long-range acoustic communication in anurans: an integrated and evolutionary approach; in The reproductive biology of amphibians (eds) D Η Taylor and Guttman S I (New York: Plenum Press) pp 263-294
Loftus-Hills J J and Littlejohn Μ J 1971 Mating call sound intensities of anuran amphibians; J. Acoust. Soc. Am. 49 1327-1329
Nevo Ε and Schneider Η 1976 Mating pattern of green toads in Israel and its ecological correlates; J. Zool. London 178 133-145
Passmore Ν I 1981 The relevance of the specific mate recognition concept to anuran reproductive biology; Μon. Zool. Ital. 6 93-108
Passmore Ν I and Carruthers V C 1979 South African frogs (Johannesburg: Witwatersrand Univ. Press)Pillai R S and Chanda S Κ 1976 The distribution pattern of amphibia in northeast India; J. Assam Sci.
Soc. 19 53-56 Pillai R S and Chanda S Κ 1979 Amphibian fauna of Khasi Hills, Meghalaya; Rec. Zool. Surv. India 75
383-395 Ryan Μ J 1985 The tungara frog: a study of sexual selection and communication (Chicago: Chicago Univ.,
Press). Ryan Μ J 1986 Factors influencing the evolution of acoustic communication: biological constraints: Brain
Behav. Evol. 28 70–82 Schneider Η and Sinsch U 1992 Mating call variation in Lake Frogs referred to as Rana ridibunda
Pallas, 1771-Taxonomic implications; Z. Zool. Syst. Evolutionsforsch. 30 297–310 Schneider Η, Sinsch U and Sofianidou T S 1993 The water frogs of Greece — Bioacoustic evidence for a
new species; Z. Zool. Syst. Evolutionsforsch. 31 47-63 Schwartz J J and Wells Κ D 1985 Intra- and interspecific vocal behavior of the neotropical treefrog Hyla
microcephala; Copeia 1985 27-38 Vigny C 1979 The mating call of 12 species and sub-species of the genus Xenopus (Amphibia, Anura); J.
Zool., London 188 103–122 Wells Κ D 1977 The social behavior of anuran amphibians; Anim. Behav. 25 666-693 Zelick R G, Rose G and Rand AS 1991 Differential response to frequency modulation rate and direction
by the neotropical frog, Physalaemus pustulosus; Anim. Behav. 42 413–421 Zimmerman Β L and Bogart J Ρ 1988 Ecology and calls of four species of Amazonian forest frogs; J
Herpetol. 22 97–108


















