

Professional Development For Science Education October 2014 For Science Education October 2014.

BINDURA UNIVERSITY OF SCIENCE EDUCATION
DEPARTMENT OF ENVIRONMENTAL SCIENCE
RESPONSE OF BLUE DUIKERS TO UNFAMILIAR CONSPECIFICS’
FAECAL PELLET DEPOSITS OF VARIOUS AGES .
REJOICE NYABASA
(B0621730)
A DISSERTATION SUBMITTED IN PARTIAL FULFILLMENT OF THE
REQUIREMENTS OF THE BACHELOR OF ENVIRONMENTAL SCIENCE
HONOURS DEGREE IN WILDLIFE AND RANGELAND MANAGEMENT.
(JANUARY 2010)

i
DEDICATION
To my dad who loves animals.

ii
ACKNOWLEDGEMENTS
I would like to thank my industrial supervisor Miss N. Lunt, for all her guidance,
advice and support. Surely you are a superb tutor and I believe I am now equipped to
pursue any kind of research specifically in the field of wildlife conservation. To my
academic supervisors Mr J. Muvengwi and Mr V. Muposhi, I am ever grateful for
constant advice given throughout the attachment period. I would also like to express
gratitude to S. Williams and A. Mudungwe for knowledge and encouragement given
while I was at Marwell Zimbabwe Trust. The mention of Ms V. Bowman (MZT
director), Mrs. A Edwards (logistics manager) and Mr C. Davison (station manager) is
inevitable for their boundless support during my stay at MZT.
A warm thanks to my friend, Mr I. Mavhurere, for inspiring me and making my ten
months stay at DAMBARI Field Station joyous and worthwhile. I will always be
grateful for valuable academic discussions we did together. To my colleague, Miss N.
Sithole, it was my greatest pleasure to work and stay with you during the attachment
period. I also wish to sincerely thank the general staff at Marwell Zimbabwe Trust, I
am ever grateful that I had an opportunity to work with you.
My special and sincere thanks proceed to my family for standing as a pillar of
strength and motivation in all aspects of my life. With your support, surely the sky is
my limit!
Forever, I thank God the Almighty for everything.
Nyabasa Rejoice.

iii
ABSTRACT
Blue duikers (Cephalophus monticola) are territorial animals and both males and
females mark out their territory continually using urine, faecal pellets and preorbital
gland secretions. The aim of the research was to assess the behavioural response of
blue duiker to unfamiliar conspecific’s faecal pellet deposits of varying ages in its
territory. To determine the intensity and type of blue duikers’ responses to foreign
conspecifics’ faecal pellets deposits, the following parameters were recorded for each
recipient: dung age, recipient sex and donor sex. Captive adult blue duikers kept in a
semi-natural environment at DAMBARI Field Station were used for the research
experimental observations. Experimental treatments consisted of dung piles from
donor animals that had aged outdoors to 1 hr, 10 hrs, 34 hrs, and 58 hrs. Two types of
controls were used: one comprised stones (“dummy” dung) similar in appearance to
faecal pellets while the second involved faecal piles from the recipient enclosure that
were aged in the same manner as donor faeces and reintroduced to the same
enclosure. Faecal pellets from donor animal enclosures were aged in the same natural
environment and later deposited in recipient pens for observations. To determine how
strongly (i. e. intensity) and in what ways (i. e. type) blue duikers responded to
foreign conspecifics' faecal pellets the following measurements and observations were
recorded for each recipient: response lag time (minutes), number of times the deposit
was approached, time spent investigating introduced pellets (seconds), type of
response exhibited and frequency of scent marking. The main type of response to
foreign faecal pellets deposits was sniffing and preorbital scent marking. Consistent
trends were detected in response intensity whereby fresh to intermediate dung were
investigated more than old dung. The results indicate that dung scent retains a high
degree of efficacy for some time (up to 34 hours) and afterward the scent signal starts
to fade out. A slight increase was noted when recipient animals investigated dung
from the same sex donor animals, most probably due to perceived threat to mates. The
results from this study suggest that the interaction of donor sex, dung age and
recipient sex had a significant effect on the intensity of response by blue duiker
recipient animal.

iv
TABLE OF CONTENTS
DEDICATION ......................................................................................................................................... I
ACKNOWLEDGEMENTS ................................................................................................................... II
ABSTRACT .......................................................................................................................................... III
TABLE OF CONTENTS ..................................................................................................................... IV
LIST OF FIGURES .............................................................................................................................. VI
LIST OF TABLES .............................................................................................................................. VII
LIST OF ACRONYMS AND ABBREVIATIONS ........................................................................... VIII
APPENDICES ...................................................................................................................................... IX CHAPTER 1: INTRODUCTION........................................................................................................... 1
1.0 BACKGROUND TO THE STUDY .......................................................................................... 1 1.1 PROBLEM STATEMENT .......................................................................................................... 1 1.2 JUSTIFICATION ...................................................................................................................... 2 1.3 AIM ....................................................................................................................................... 2 1.4 OBJECTIVES .......................................................................................................................... 2 1.5 RESEARCH HYPOTHESES ....................................................................................................... 3
CHAPTER 2: LITERATURE REVIEW ............................................................................................... 4
2.0 INTRODUCTION..................................................................................................................... 4 2.1 INTERNATIONAL AND LOCAL DISTRIBUTION OF DUIKERS ..................................................... 4 2.2 IMPORTANCE OF DUIKERS ..................................................................................................... 5 2.3 THE CONSERVATION STATUS OF DUIKERS ............................................................................ 6 2.4 ECOLOGY AND BIOLOGY ASPECTS OF BLUE DUIKERS ......................................................... 10
CHAPTER 3: RESEARCH METHODOLOGY.................................................................................. 17
3.0 STUDY AREA ........................................................................................................................ 17 3.1 RESEARCH METHOD ............................................................................................................ 17 3.2 DATA ANALYSIS................................................................................................................... 19 3.3 LIMITATIONS OF THE STUDY ............................................................................................... 19
CHAPTER 4: RESULTS ...................................................................................................................... 21
4.0 TYPE OF RESPONSE............................................................................................................ 21 4.1 RESPONSE OF RECIPIENT ANIMALS TO DONOR ANIMAL DUNG ........................................... 23 4.2 THE EFFECT OF DUNG AGE, RECIPIENT SEX AND DONOR SEX TO RECIPIENT ANIMALS’ RESPONSE ......................................................................................................................................... 23 4.3 TRENDS IN BLUE DUIKER RESPONSE (LAG) TIME TO INTRODUCED TREATMENTS............... 25 4.4 TIME SPENT BY BLUE DUIKERS INVESTIGATING INTRODUCED TREATMENTS ..................... 26 4.5 PREORBITAL SCENT MARKING BY BLUE DUIKERS TO INTRODUCED TREATMENTS IN THEIR TERRITORIES .......................................................................................................................... 26 4.6 INVESTIGATION FREQUENCY BY BLUE DUIKERS TO TREATMENTS INTRODUCED INTO THEIR TERRITORIES .......................................................................................................................... 28
5.0 DISCUSSION .......................................................................................................................... 29

v
CHAPTER 6: CONCLUSION AND RECOMMENDATIONS .......................................................... 31
6.0 CONCLUSION ....................................................................................................................... 31 6.1 RECOMMENDATIONS ........................................................................................................... 32
REFERENCES ..................................................................................................................................... 33

vi
LIST OF FIGURES FIGURE 1: BLUE DUIKER URINATION SITE WITHIN ENCLOSURE ................................................................... 21 FIGURE 2: BLUE DUIKER DEFECATION SITE ADJACENT TO NEIGHBORING ENCLOSURE ............................ 22 FIGURE 3: MARKS OF PREORBITAL GLAND SECRETION LEFT BY A TERRITORIAL BLUE DUIKER ON THE
POLE. ............................................................................................................................................. 22 FIGURE 4: TRENDS IN MALE RECIPIENTS RESPONSE TIMES TO INTRODUCED TREATMENTS ........................... 25 FIGURE 5: TRENDS IN FEMALE RECIPIENTS RESPONSE TIMES TO INTRODUCED TREATMENTS...................... 25 FIGURE 6: MEAN TIME TAKEN BY MALE RECIPIENTS SNIFFING INTRODUCED TREATMENTS .......................... 26 FIGURE 7: MEAN TIME TAKEN BY FEMALE RECIPIENTS SNIFFING INTRODUCED TREATMENTS ...................... 26 FIGURE 8: TRENDS IN PREORBITAL SCENT MARKING BY MALE RECIPIENTS ................................................. 27 FIGURE 9: TRENDS IN PREORBITAL SCENT MARKING BY FEMALE RECIPIENTS .............................................. 27 FIGURE 10: FREQUENCY OF INVESTIGATION BY MALE RECIPIENTS TO INTRODUCED TREATMENTS ............... 28 FIGURE 11: FREQUENCY OF INVESTIGATION BY FEMALE RECIPIENTS TO INTRODUCED TREATMENTS............ 28

vii
LIST OF TABLES TABLE 1: SUMMARY OF THE CONSERVATION STATUS OF SUB-SAHARAN AFRICAN ANTELOPE ...................... 6 TABLE 2: SOURCES OF VARIATION FOR TOTAL SNIFF DURATION SHOWING THAT MALE RECIPIENTS
SNIFFED FOR LONGER DURATION 10HR FEMALE DUNG COMPARED TO 1HR, 34HR, AND 58HR AGED DUNG FROM CONTROL, MALE AND FEMALE DONORS......................................................................... 24
TABLE 3: SOURCES OF VARIATION FOR INVESTIGATION FREQUENCY. MALE RECIPIENTS INVESTIGATED MORE 10HR FEMALE DUNG COMPARED TO 1HR AND 58HR AGED DUNG FROM CONTROL AND FEMALE DONOR. ............................................................................................................................. 24
TABLE 4: SOURCES OF VARIATION FOR LAG TIME RESPONSE VARIABLE SHOWING THAT FEMALE RECIPIENTS TOOK LONGER TIME TO START INVESTIGATING INTRODUCED 58HR FEMALE DUNG COMPARED TO 1HR AGED DUNG. ..................................................................................................... 25

viii
LIST OF ACRONYMS AND ABBREVIATIONS
CAMPFIRE Communal Areas Management Programme For Indigenous Resources
DAMBARI Duikers and Mini-antelope Breeding and Research Institute
MZT Marwell Zimbabwe Trust
ODA Overseas Development Administration

ix
APPENDICES APPENDIX 1: SAMPLE DATA RECORD SHEET ............................................................................................ 39

1
CHAPTER 1: INTRODUCTION
1.0 Background to the Study
Scent marking is a common form of signaling in mammals and it usually occurs in
complex spatial patterns, often involving several marks that are regularly replenished
in marking and remarking sessions by the signaler. According to Ralls (1971), most
mammals have a highly developed olfactory sense, which they employ in social
communication using signals originating in urine, faeces or scent glands. Several
territorial mammals apply scented secretions and excretions to areas or objects in their
environment as a way of demarcating their territories or home ranges (ibid).
Scent marks are a form of status signal, advertising territory ownership, and recent
evidence suggests that they may also be condition-dependent signals of quality and
competitive ability (Gosling et al., in press). A unique characteristic of scent marks as
a social signal is that receivers often encounter the signal in the absence of the
signaler. Studies which have examined the distribution of scent marks within a
territory have usually found that they are either clustered at the territory boundary or
near its centre (Roberts, 1992).
Blue duikers are the smallest antelopes found in the southern African subregion
(Smithers, 1983). The average height of adult males is generally 35.5 cm with a mass
of about 4, 5 kg, while females are slightly larger with an average height of 36.2 cm
with a mass of 5.4 kg (Best and Best, 1977). Blue duikers are territorial (Bowland and
Perrin, 1995) and not easily driven from their territories. Direct interactions between
neighbours are rare, but territorial occupancy is continually advertised by visual,
olfactory and sometimes by auditory means (Estes, 1997). They scent mark with a
pair of preorbital glands, urine or faecal pellets to demarcate their territory boundary
and pedal glands mark their movements in their forest habitats (Smithers, 1983).
1.1 Problem Statement
Scent marking is a widespread but poorly understood form of communication in

2
duikers (Johnson, 1973; Stoddart, 1980; Brown and Macdonald, 1985). In their
natural environment, blue duikers’ territorial behaviour is very difficult to observe and
study in detail due to the reason that they are shy and secretive (Wilson, 2005). In
comparison to other aspects of scent marking and territoriality in mammals, the
strength of behavioural response of blue duikers to the introduction of unfamiliar
conspecifics’ faecal pellet deposits of varying ages has received relatively little
attention.
1.2 Justification
Little has been done to investigate on the response of blue duikers to the introduction
of unfamiliar scent of varying ages, therefore this study will enable a better
understanding of this species’ territorial behaviour. The study will also facilitate the
provision of detailed information on blue duiker territorial behaviour for present and
future academic purposes especially in wildlife related research work. Territoriality is
one of the most important behavioural traits affecting the spatial organization of
animal populations (Mizutani, 1993). As a result, successful re-introductions of blue
duiker species require the general knowledge of the animal’s territorial behaviour so
as to be able to facilitate the maintenance of social organization, space use patterns,
resource utilization and also reduce energy demanding direct encounters among
animals of the same species. The availability of captive blue duikers in semi-natural
enclosures at DAMBARI Field Station helped in providing a conducive environment
for clear observations of individual animal territorial behaviour thus accurate results
and conclusions can be deduced.
1.3 Aim
To determine the responses of blue duikers to unfamiliar conspecifics’ faecal pellets
in their territories.
1.4 Objectives
i) To determine how intensity and type of response by recipient animals varies with
sex of donor, dung age and sex of recipient.

3
1.5 Research Hypotheses
H0: The intensity and type of response by recipients do not vary with sex of donor.
H0: The intensity and type of response by blue duikers do not vary with sex of
recipient.
H0: The intensity and type of response by recipients do not vary with dung age.

4
CHAPTER 2: LITERATURE REVIEW
2.0 Introduction
The life history of duikers is relatively unstudied in natural conditions due to the fact
that they are secretive and shy thus difficult to observe for long periods in forest
environments (Newing, 1994). However, they follow the general patterns of forest
animals including small body size, short horns, frugivorous diet and small group size
(Dubost, 1983). According to Estes (1997), observations of captive blue duikers
maintained under semi-natural conditions confirm that pair bonds are probably life
long.
All duikers defend their territories against intruders thus they are classified under
territorial animals (Kingdon, 1997). They scent mark with a pair of preorbital glands,
urine or faecal pellets to demarcate their territory boundary and pedal glands mark
their movements in their forest habitats (Smithers, 1983). Usually there is no joint or
family defense of the territory, but each resident adult and subudult selectively
excludes interlopers of its own sex (Estes, 1997).
This chapter outlines the ecological aspects of duikers including habitat preference,
social organizations and also feeding, breeding and territorial behaviors. It also gives
an overview of the general distribution of duikers internationally and locally and also
the ecological, economic and social importance of duikers. The conservation status of
duikers in Africa is also summarized incorporating the present survival threats and the
conservation strategies in use to reduce continuing decline of duiker populations.
2.1 International and Local Distribution of Duikers
There are 16 species of duikers found in Africa (Wilson, 2005). Nearly every African
forest and woodland is occupied by at least one species of duiker (Kingdon, 1997) and
the majority of duiker species live in the Equatorial Rain Forest (Estes, 1997). The
blue duiker has a very wide distribution in Africa south of the Sahara desert with its
overall range extending from the east bank of the Cross River in Nigeria stretching

5
through Central Africa to the coastal areas of the Cape Province of South Africa
(Wilson, 2005). In Zimbabwe, blue duikers are confined to the evergreen forests in
the eastern highlands specifically areas like Chirinda forest, Chipete forest, Vumba
Botanical Garden, Bunga forest, Bridal Veil Falls forest and also Nyanga and Honde
valley (Wilson, 2005). However, the largest population of blue duiker in Zimbabwe
occurs in the Chirinda evergreen forest on Mt. Selinda (ibid).
2.2 Importance of Duikers
Conservationists have long recognized the necessity of examining not only the
ecological but also the economic and social importance of wildlife to communities (Fa
et al., 1995; Fitzgibbon et al., 1995). Duikers are generally shy and secretive animals
thus difficult to study in their natural environment. Due to this reason, they have been
assigned minor roles in many ecological studies or even ignored. However, they play
important roles in many ecosystems including structuring understorey vegetation,
recycling nutrients, seed dispersal and forming a major part of the prey base for a
wide range of predators (N. Lunt, pers. comm.).
Ecologically, duikers play an important role in the diet of certain tropical forest
predator species including leopard (Panthera pardus) and golden cat (Felis aurata)
(Hart et al., 1996). The threat to Africa’s predator populations caused by dramatic
declines in their prey base is likely to be significant without duikers (ibid).
Duikers are primarily frugivorous although they also browse selectively and
occasionally take animal matter (Feer, 1995). About 150 different species of fruit
were found among the rumen contents of forest duikers, which suggest that they may
play an important role as seed dispersal agents (ibid).
Rural communities are highly dependent on forest resources for both nutritional and
economic subsistence (Lahm, 1993; Noss, 1995). Traditionally, duikers are hunted by
humans for consumption as a primary source of protein. The majority of bushmeat
studies conducted to date involve market surveys, many of which indicate that duikers
comprise the majority (40-80%) of meat sold in bushmeat markets throughout Central
Africa (Chardonnet and Fritz, 1995; Bowen–Jones, 1998).

6
2.3 The Conservation Status of Duikers
Despite their widespread range, nearly all species of duiker are currently projected to
decline considerably in the near future while several species are currently considered
on the point of extinction if immediate protection measures are not put into effect
(East and IUCN/SSC/ASG, 1999).
The antelope data presented in Table 2.1 is completely taken from the African
Antelope Database 1998 (AAD 1998) compiled by Rod East of the IUCN/SSC
Antelope Specialist Group. However, the information given in AAD 1998 is not
complete due to lack of detailed information available from many parts of Africa. The
term threatened in this case is used specifically to refer to those species classified as
vulnerable, endangered or critically endangered (ibid).
According to East and IUCN/SSC/ASG (1999), the percentage of small antelope that
are currently threatened is approximately the same as that for large antelope. However
within the next 15 years the percentage of small antelope forms that will be threatened
will more than double, whereas other threatened antelope will only increase by 38%.
Small antelope will become more threatened than the average African antelope.
Perhaps the most worrying of all is that an astounding 92% of small antelope are
currently undergoing population declines or show a tendency to decline compared to
66% of other antelope.
Table 1: Summary of the conservation status of sub-Saharan African antelope
Small Antelope Other Antelope Total Antelope
Threatened 10(27%) 23(32%) 33(30%)
Threatened by 2025 22(59%) 37(51%) 59(54%)

7
Decreasing or showing a
tendency to decrease
34(92%) 47(66%) 81(74%)
Source: African Antelope Database, 1998
Eves (1996) states that one possible reason for the difference in population trend is
that the majority of other antelope are larger savannah species. These species have
already experienced dramatic declines from historical population sizes and are now
stabilizing at reasonably low levels in protected areas and areas that still have low
human densities. In contrast a large number of small antelope are forest dwellers. By
virtue of their habitat, these species have been less exposed to widespread human
influences until more recent times when human population continued to increase
(Bowen-Jones, 1998).
In Africa, the blue duiker is still widespread and common in spite of heavy hunting
pressure all over its range and is still regarded as non-endangered species (Kingdon,
1997). However, in Zimbabwe blue duikers are in danger of extinction and unlikely to
survive if the present population decline contributing factors continue operating
(Wilson, 2005).
2.3.1 Survival Threats of Duikers
Many populations of duikers are at risk of local or ecological extinction due to
excessively high off take rates and unnecessary wastage (Hart, 2000), whereby some
of the animals caught in snares are lost to scavengers and decomposition and some
escape with dreadful injuries. The major threats facing most of these species are bush
meat hunting and habitat loss or degradation (ibid). Both problems are complex and
cannot be solved easily and will require considerable funding, research and education
initiatives.
2.3.2 Commercial and Traditional Hunting for Bush Meat
Hunting of duikers for traditional markets has been undertaken for considerable time
and is economically, culturally and socially important to local communities (Eves,
1996). Due to demographic changes and better road network between the source and

8
potential markets, hunting for commercial markets in towns and cities has also
increased dramatically in recent years. It is possible to hunt for these species at a
sustainable level but much more data are needed on most species to be able to
determine what the sustainable yields may be (Hart, 2000). There will be need for
education and capacity building to convince hunters that it is for their own advantage
to abide by quotas, low wastage hunting methods and proper policing and protection
to monitor hunting effort (ibid).
2.3.3 Habitat Loss and Degradation
In some areas particularly in East Africa, habitat loss or degradation may be an equal
or even greater threat to duikers than hunting (Masoud, 2000). This is largely due to
clearance for agriculture and commercial wood harvesting. Many duiker populations
are still outside of protected areas and are therefore at high risk of increased habitat
loss as human populations continue to expand in the future (Bowen-Jones, 1998).
2. 3. 2 Conservation Strategies for Small Antelopes
The survival of most species of duiker in the future significantly depends on increased
conservation efforts presently and in the coming years. Masoud (2000) states that
participation by local and international stakeholders is greatly helpful in allowing
successful implementation of conservation measures.
2.3.4 Law Enforcement
In the past, law enforcement was the major conservation tool whereby governments
would allocate enough resources to employ as many guards as required to deter
people from hunting (Masoud, 2000). However, changes in socio-economic
conditions of people, population pressure and lack of adequate financial resources
have created a more difficult environment for the governments to rely on law
enforcement alone (ibid.). Therefore, this strategy is often implemented through
community participation programmes like CAMPFIRE and ODA.
2.3.5 Community Management Programmes

9
The community wildlife programmes started as a result of human-environment
interaction work done through research (Masoud, 2000). During this period, local
people expressed their desire to be involved in the management of natural resources
especially antelope since they believe that they are an important alternative protein
source. Community participation allow locals to plan and set aside their forestlands
for various purposes to include conservation and sustainable exploitation areas and in
return they gain revenue from hunting fees to support conservation and development
work in respective villages.
A case study of Zanzibar reveals a successful community management programme
whereby a new Forestry legislation was passed. It empowers the community to enter
into agreement with the Forest Administrator to manage their wildlife resources on
presentation of a management plan and by-laws. This has been done very successfully
with 12 village communities on southern Unguja, whereby through technical backup
from national and international staff, the by-laws where prepared approved and are
currently in force (ibid).
2.3.6 Translocation
Western and Pearl (1989) states that translocation is believed to be one of the
effective ways of saving a population from complete extinction within its bio-
geographical region. Translocations have been done in Zanzibar as rescue
programmes. Aders’ duikers were first translocated into Funzi Island in Pemba but the
population became locally extinct due to uncontrolled hunting. The second
translocation was a community managed initiative where suni antelope were first
introduced into Juja islet at Ukongoroni, Zanzibar. The community, except for socio-
cultural functions involving the entire community, strictly forbids hunting. This
translocation has been successful and the population of suni still exists at Juja. This
technique is common in New Zealand and has been in use with greater success (ibid).
2.3.7 Captive Breeding Programme
Captive breeding can be an important strategy to prevent species from extinction and
has grown enormously in recent years (Snyder et al., 1996). However, there are

10
numerous drawbacks such as high costs involved in establishing conducive captive
breeding facilities and loss of heterozygosis in poorly managed captive populations
(Masoud, 2000).
2.4 Ecology and Biology Aspects of Blue Duikers
2.4.1 Habitat Preferences
Duikers generally flourish in a wide range of habitats in which requirements prescribe
an obligate balance between food resources and shelter (Bowland and Perrin, 1995),
of which a deficiency of either reduces habitat quality. Blue duiker habitats include
lowland and montane rainforests and also riverine and moist thickets (Kingdon,
1997). They mostly occupy areas with comparatively little undergrowth to minimise
obstruction of its movements or vision (Estes, 1997).
2.4.2 Activity Periods and Social Organization
All duikers are thought to be sedentary and territorial (Estes, 1997). Some species are
solitary, some live in pairs and others are occasionally found in groups with one adult
male and a number of adult females (Feer, 1989). At any rate, pairs of blue and
Maxwell’s duikers (Cephalophus maxwellii) apparently associate more closely than
most other duikers because they are more vulnerable to predators than larger species
that live in denser cover in more spacious home ranges. Blue and Maxwell’s duikers
are known to live in monogamous pairs within small territories that are jointly marked
and defended against foreign members of the same sex (Estes, 1997). In blue duikers
pair bonding is reinforced mainly by pressing the preorbital glands together and social
coat licking.
Activity periods of duikers vary with species. Some species such as the blue duiker
and the Maxwell’s duiker are active during the day (Newing, 1994). Others such as
the bay duiker (Cephalophus dorsalis) are nocturnal and the larger species like
yellow-backed duiker (Cephalophus sylvicultor), Abbott’s duiker (Cephalophus
spadix) and Jentink’s duiker (Cephalophus jentinki) are active both by day and by
night (ibid).

11
2.4.3 Diet Selection
The common accepted view of duikers is that they are high concentrate feeders
(Hofmann, 1973 and Wilson, 2005). The requirement for easily assimilated and
highly nutritious foods is linked with the high metabolic demands of a small body
(Bell, 1971; Milton, 1982; Van Soest, 1983). The Jarman-Bell principle illustrates that
small mammalian ruminants feed on small, energy-rich, dispersed food sources at
relatively low density and low in plant fibre (Jarman, 1974; Spinage, 1986;
McNaughton, 1987).
Blue duikers are primarily frugivorous but also browse selectively (i.e. leaves, shoots,
and buds) and occasionally take animal matter in the form of insects (Bodmer, 1990;
Hanekom and Wilson, 1991). In areas where fruiting trees and shrubs are abundant, it
was found that upto 80% of blue duiker diet may be fruits with little of foliage,
flowers, gum and animal (Kingdon, 1997). Preferred fruits are the abundant small
berries or seeds that are often typical of dominant trees in secondary or colonizing
forest (ibid). Blue duikers have been seen to follow below monkeys and birds to pick
up fallen fruits. Probably duikers gain enough water from their food and rarely have
to drink, but are occasionally seen licking rainwater off foliage (Estes, 1997).

12
2.4.4 Breeding
According to Estes (1997), all African bovids bear one young and breed at least once
a year (small species often twice), giving birth to well developed young after gestation
periods of about six months (small antelopes and gazelles) to eight or nine months in
medium and large species. Wilson (2005), states that female blue duiker age of sexual
maturity ranges from eight to twelve months whilst for males ranges from eleven to
fourteen months. A female’s maximum lifetime offspring production has been found
to be about 14 young ones for both small and large species. Most species have
extended but definite breeding seasons during the rainy period of the year and in
equatorial regions with two rainy seasons, many have a secondary calving peak
(Estes, 1997). However, records at DAMBARI Field Station indicate year round
breeding in blue duikers with a slight peak in the wet season (N. Lunt, unpub. data).
2.4.5 Scent Marking and Territoriality
Scent marking is a form of animal communication which is common in territorial
species (Roberts, 1997). It refers to the application of scented secretions and
excretions by an animal to areas or objects in its environment (Wells and Bekoff,
1981). A territory is a specific area within a home range in which an animal is
aggressive and supremely dominant with respect to certain categories of intruder
(Anderson, 1983).
2.4.6 Types of Scent Marks
Scent marks occur in complex patterns often involving several marks that are
regularly replenished in active marking and remarking sessions (Gosling and McKay,
1990). Many species demarcate territories using chemical signals originating from
urine, faeces and cutaneous scent glands (Ralls, 1971).
Several mammal species including antelopes use faeces for territory demarcation (Asa
et al, 1985). Faeces serve as substrate for glandular secretions and as visual
‘signposts’ to intruders. The use of faeces may be ideal for scent marking because
they have a minimal energetic cost to the signaler (Gosling, 1981) since they are
waste from metabolism. However, limited food sources may affect faecal pellets

13
production thus limiting marking rates in species that use them to advertise territory
borders (Komers, 1996).
The use of gland secretions is also common with many animal species whereby
different scent glands secretions are used through varying marking methods. Many
duiker species scent mark using secretion from preorbital gland located just below
each eye (Ralls, 1971). Both males and females frequently rub the gland on objects in
their territory leaving a small, visible deposit of secretion (Roberts, 1994). Some
mammals like rabbits have two main scent gland secretions which are associated with
social dominance and territoriality (Ralls, 1971). These are submandibular or chin
gland secretion and anal gland secretion which is usually produced together with
faeces (ibid).
Urine serves a variety of functions besides territory demarcation, depending on the
species and the environment in which it is expressed (Brown, 1985). Bowyer and
Kitchen (1987) mentioned that scent urination is believed to contribute in information
transmission whereby it could act as a visual display and also as an olfactory signal.
Scent urination is commonly performed by males whereby they urinate onto
themselves and others urinate onto the ground and sometimes then wallow in the
urine.
2.4.7 Significance of Scent Marks
Scent marking may serve as a mechanism for advertising territory ownership and also
serve as a challenge ritual to potential intruders thus reducing the frequency of
fighting over territorial rights (Roberts, 1997). Scent marks provide an opportunity for
intruders to identify resource holders and then to modify their behaviour in a way that
takes account of the benefits of utilizing the defended resource and the costs of
meeting the resource holder (Gosling, 1990). A receiver could examine deposited
scent marks and it would be able to determine if the opponent was of high rank and,
depending on its own relative competitive ability and the value of the marked
resource, it could withdraw from the encounter at an appropriate stage (Gosling,
1982). As a result, resource holders can avoid or reduce the costs of contests with a
large proportion of opponents (ibid).

14
Scent marks also contribute in sex recognition and also serve as an indicator of sexual
condition and maturity thus helping in mate selection (Wells and Bekoff, 1981).
During the breeding season, many male ungulates use urine as a medium to transmit
information to conspecifics (ibid). Female white- tailed deer investigate and scent
mark at male marking sites when they are in estrus suggesting that males use marking
sites to assess potential mates (Sawyer et al., 1989). Female mice also use male mice
odor originating from urine to distinguish parasitized and unparasitized males thus
avoiding mating with infected males (Gosling, 1985).
Scent marks also help in providing internal information to orient members of the same
resident group (Wells and Bekoff, 1981). Estes (1997) states that scent trails left by
the pedal glands and the preorbital gland secretion left on trees probably help family
members keep track of one another. Blue duikers also perform mutual gland pressing
whereby mate pairs scent mark each other or their offspring on projections such as the
forehead and tail base as a social behaviour (ibid).
2.4.8 Temporal Variation in Scent Mark Efficacy
Scent marks are placed in the environment often in the absence of a potential receiver
and only detected some time later often when the signaler is no longer present
(Gosling and Roberts, 2000). The two crucial parameters of temporal variation for
signalers are the rise time, which is the interval between deposition and achievement
of maximal detection and the mark’s persistence time which is interval between
deposition and the time when the mark can no longer be detected (Ferkin et al., 1995).
The efficacy of animal scent marks is not constant but varies with time after
deposition (Alberts 1992). Recipient animals use the relative proportions of volatile
sub-components of the scent to estimate the age of the scent mark (Johnston and
Schmidt 1979; Apps et al. 1989; Ferkin et al. 1995). Information about the age of
scent marks may allow receivers to modulate their response to scent encountered in
proportion to the perceived likelihood of encountering the signaler (Roberts, 1998).
Owners encountering foreign scent marks in their territory may take evasive action if
the scent is very fresh compared to old scent perhaps to avoid physical conflict since
the probability that the signaler will be in the vicinity is high (Maynard Smith, 1974).

15
It can be expected to over mark its area with own scent to maximise scent matching
when the intruder assess the area again (Gosling, 1985).
A receiver can examine deposited scent marks and it will be able to determine if the
opponent was of high rank (i.e. physical and sexual condition) and, depending on its
own relative competitive ability and the value of the marked resource, it can withdraw
from the encounter at an appropriate stage (Gosling, 1982). Several studies conducted
before revealed that when receivers approach scent marks, they often sniff and lick
the scent deposit (Brown and MacDonald, 1985; Idris, 1994; Roberts, 1998) or they
over mark on or around the deposited marks (Melemis and Falls, 1982).
2.4.9 Territoriality in Blue Duikers
Territoriality is one of the most important behavioural traits affecting the spatial
organization of animal populations (Mizutani, 1993). Particularly in blue duikers,
direct interactions between neighbours are rare thus territorial occupancy is
continually advertised by visual, olfactory and probably also by auditory means
(Estes, 1997). Odendaal and Bigalke (1979) indicated that the blue duiker is strongly
territorial and both males and females mark out their home range continuously using
urine, faecal pellets and preorbital gland secretion.
Rubbing the preorbital glands on tree trunks and other objects, wherever the duiker
goes, is the main form of demarcation (Estes, 1997). The scent oozes from glands just
below their eyes. The preorbital gland is equally productive in both sexes and the
secretion is an inhomogeneous mixture of a clear, colourless liquid and white mucous
material which in females, tends to have a bluish-grey colour (Kingdon, 1997).
According to Estes (1997), the other form of territorial demarcation is tree horning, in
which both sexes engage from the time their horns emerge. Permanent, visible marks
are made on target trees, chiefly young saplings with stems small enough to fit into
the fork between the horns. The duikers always horn at shoulder level after first
sniffing and licking and sometimes scraping the spot with the incisors. Audible alarm
signals may also serve as an auditory reminder that a territory is occupied (ibid).

17
CHAPTER 3: RESEARCH METHODOLOGY
3.0 Study Area
DAMBARI Field Station is a research institution situated 25 km along Bulawayo-
Gwanda road southeast of Bulawayo, Zimbabwe. The station property covers a total
area of 25 ha, situated at an altitude of 1050 m. The area receives an annual average
rainfall of 600 mm, which normally falls between November and March. The
vegetation type is mainly Acacia woodland interspersed with shrubs. The land slopes
down to a small seasonal river called Umguza (Bowman & Plowman, 2002), and the
vlei area near the river is dominated by Hyparrhenia species.
3.1 Research Method
Captive blue duikers (Cephalophus monticola) were observed at DAMBARI Field
Station between November 2008 and April 2009. Eight adult blue duikers (six
females and two males) in their prime were maintained either singly or in pairs in six
150 m2 fenced, semi-natural pens that are arranged in a hexagonal pattern around a
double-storey observation platform. Minimization of interactions between animals in
adjacent enclosures was facilitated by the presence of black opaque plastic sheeting
attached to fences to a height of 1 m.
Donor animals were maintained singly in pens located at least 40 m from the pens of
recipient animals. They were composed of four adult blue duikers (two females and
two males) in their prime, which where also kept in a similar semi-natural
environment to recipient animals. Faecal pellets were collected when fresh, and aged
in the same natural environment or immediately deposited in recipient pens,
depending on the treatment requirement. Clean polythene bags were used to collect
faeces, to prevent scent contamination by the researcher.
Experimental treatments consisted of dung piles from donor animals that had aged
outdoors to 0-1 hr, 9-10 hr, 33-34 hr or 57-58 hr. Two types of controls were used:
one comprised stones (“dummy” dung) similar in appearance to faecal pellets while
the second involved faecal piles from the recipient enclosure that were aged in the

18
same manner as donor faeces, and reintroduced to the same enclosure. The dummy
dung control, which was scent-neutral, was used to determine if animals used visual
cue to identify unfamiliar objects in their pens.
Treatments were given in a randomized fashion in a repeated measures design such
that each enclosure received each treatment once. An interval of at least 24 hours was
allowed before introducing a different treatment to the same enclosure to ensure that
there was no temporal autocorrelation in recipient responses.
Treatments were introduced to experimental enclosures at 1630 hrs, deposited at a site
away from the animals’ usual latrine site/s but readily visible to the observer.
Observation periods were restricted to times when duikers were most active (Bowman
and Plowman 2002; pers. obs.), i.e. from 1630 hrs to 2000 hrs. During observation
periods, territory-defence related activities occurring in one pen were observed and
recorded.
3.1.1 Territorial Behaviour Assessment
The number of times a recipient individual approached an introduced unfamiliar
conspecifics’ faecal deposit was recorded, with the type of response stated.
Responses consisted of investigation (sniffing) and pre-orbital scent marking within
the range of 1 m from the introduced deposit. To determine how strongly (i. e.
intensity) and in what ways (i. e. type) blue duikers responded to unfamiliar
conspecifics’ faecal pellets, the following measurements and observations were
recorded for each individual:
1) Response lag time (number of minutes from faecal introduction to first
investigation).
2) Type of response exhibited (for example approach and sniff; approach and
scent-mark).
3) Time (in seconds) spent investigating faecal pellets on each occasion the pile
was approached.
4) Frequency of scent marking on each occasion the pile was approached.
Other important factors like name of donor and recipient animal, weather conditions,

19
starting and ending time of observations were also recorded.
3.2 Data Analysis
Data collected were entered into Microsoft Excel (Microsoft Corporation, 2003)
spreadsheets and pivot tables were used to summarize data. Data from the pivot table
were then analyzed using Minitab 15 (Minitab Inc., 2006). Response variables were
(i) lag time, (ii) total sniff duration (sum of seconds spent sniffing an introduced
deposit), (iii) mean sniff duration (mean number of seconds spent sniffing a deposit)
and (iv) scent-mark frequency (total number of occasions that pre-orbital scent-marks
were applied to structures within 1 m of the introduced deposit). Predictor variables
were dung age, recipient sex and donor sex.
To assess the validity of using ANOVA for analysis, Levene’s test and normal
probability distributions were run for each response variable against predictors. Data
were non-normal (Anderson-Darling tests, p < 0.05), but variances were not
significantly different (Levene’s test, p > 0.05 for all tests). To be conservative,
permutational multivariate analysis (which is not affected by non-normal data) was
used to test the effects of donor sex, recipient sex and dung age on each response
variable with DISTLM (Anderson, 1999). To ensure that responses to donors were
random, Sign tests were run for each response. Line plots of responses against dung
age for each donor/recipient group were run using SigmaPlot 8.0 (Brannan et al.,
2002).
3.3 Limitations of the Study
Variation in weather conditions could have affected the results when experiments
were conducted during bad weather conditions (i. e. cold and wet). During cold and
wet weather, recipients tend to become inactive, so the intensity of response to
introduced faecal deposits is expected to be underestimated. Inaccurate conclusions

20
can also be deduced if donor females in estrus were involved, because recipient males
may have responded more to faecal pellets of females in estrus resulting in an
overestimate of response intensity.

21
CHAPTER 4: RESULTS
4.0 Type of Response
Sniffing and scent marking with preorbital gland secretion were the only responses
noticed for both male and female recipients. Recipient animals sniffed at or around
the deposited foreign faecal pellets then sometimes followed by deposition of
preorbital gland secretions on nearby tree and grass stems, poles and rocks. Urine and
faeces were also deposited during observation periods, but this was never directly
associated with the introduced deposits; rather, faeces and urine were deposited at
fixed locations along borders shared with other animals and sometimes randomly
within the pen (Fig. 1 and 2). Animals also did not scent mark exclusively at
introduced pellet sites, but continued to mark their usual marking sites such as
perimeter poles (Fig. 3).
Figure 1: Blue duiker urination site within enclosure

22
Figure 2: Blue duiker defecation site adjacent to neighboring enclosure
Figure 3: Marks of preorbital gland secretion left by a territorial blue duiker on the pole.

23
4.1 Response of Recipient Animals to Donor Animal Dung
Recipient animal responses were not significantly linked with specific individuals
within the same sex category (Sign test; p > 0. 05 for all tests), indicating that there
was no preferential response to one donor over another for example male Nyathi over
male Secret.
4.2 The Effect of Dung Age, Recipient Sex and Donor Sex to Recipient
Animals’ Response
The interaction of donor sex, dung age and recipient sex had a significant effect on the
intensity of response by recipient animals (permutation ANOVA, d.f = 3, 164;
p<0.05) for all response variables tested.
To precisely determine the source of variation, Tukey’s HSD tests were run, and the
significant results were recorded (tables 4.1 – 4.3). Male recipients sniffed for longer
duration (table 4.1) and investigated more (table 4.2) 10 hr female dung compared to
1 hr, 34 hr and 58 hr aged dung. Female recipients reacted faster and sniffed longer to
fresh female dung than to old dung from either male or female donor (table 4.1 and
4.3).
The tables below represent sources of variation derived from Tukey’s HSD tests for
specific response variables showing experimental combinations with significant
different results. Combinations which were not significantly different were excluded
from the results listed in tables below.

24
Table 2: Source of variation for total sniff duration showing that male recipients
sniffed for longer duration 10hr female dung compared to 1hr, 34hr, and 58hr aged
dung from control, male and female donors.
Donor Recipient Sign Donor Recipient P-Value
Female Female (1 hr) > Female Female (58 hr) 0.004
Female Female (1 hr) > Male Female (58 hr) 0.045
Female Male (10 hr) > Male Female (1 hr) 0.013
Female Male (10 hr) > Control Female (1 hr) 0.016
Female Male (10 hr) > Control Female (10 hr) 0.038
Female Male (10 hr) > Female Female (34 hr) 0.046
Female Male (10 hr) > Male Female (34 hr) 0.042
Female Male (10 hr) > Female Female (58 hr) 0.001
Female Male (10 hr) > Male Female (58 hr) 0.009
Female Male (10 hr) > Control Female (58 hr) 0.038
Table 3: Source of variation for investigation frequency. Male recipients investigated more 10hr female dung compared to 1hr and 58hr aged dung from control and female donors.
Donor Recipient Sign Donor Recipient P-Value
Female Male (10 hr) > Control Female (1 hr) 0.036
Female Male (10 hr) > Female Female (58 hr) 0.005

25
Table 4: Source of variation for lag time response variable showing that female recipients took longer time to start investigating introduced 58hr female dung compared to 1hr aged dung. Donor Recipient Sign Donor Recipient P-Value
Female Female (58 hr) > Female Female (1 hr) 0.015
4.3 Trends in Blue Duiker Response (lag) Time to Introduced Treatments Consistent trends were detected in response whereby both male and females took
longer to start investigating fresh (1 hr) to intermediate (10 hr and 34 hr) control dung
compared with foreign dung. Both recipient sexes investigated fresh foreign dung
faster than old foreign dung (58 hr) from same sex donor animals.
Recipient male
Dung age (hrs)
0 20 40 60 80 100
Lag
time
(min
s)
0
20
40
60
80
100
120
140
160
180
200
Donor male, Recipient male Recipient male, Donor female Control, Recipient male Stones, Recipient male
Recipient female
Dung age (hrs)
0 20 40 60 80 100
Lag
time
(min
s)
0
20
40
60
80
100
120
140
160
180
200
Recipient female, Donor female Donor male, Recipient female Control, Recipient female Stones, Recipient female
Figure 4: Trends in male recipients response times to introduced treatments
Figure 5: Trends in female recipients response times to introduced treatments

26
4.4 Time Spent by Blue Duikers Investigating Introduced Treatments
Male and female recipients sniffed control dung less than foreign dung. Very fresh
dung from same sex donor animals was sniffed more compared to that from opposite
sex donor animals. However, the longest sniff duration recorded for both sexes was
for introduced stones. Recipient Male
Dung age (hrs)
0 20 40 60 80 100
Mea
n sn
iff d
urat
ion
(sec
)
0
2
4
6
8
Donor male, Recipient male Control, Recipient male Recipient male, Donor female Stones, Recipient male
Recipient female
Dung age (hrs)
0 20 40 60 80 100
Mea
n sn
iff d
urat
ion
(sec
)
0
2
4
6
8
Recipient female, Donor female Donor male, Recipient female Control, Recipient female Stones, Recipient female
Figure 6: Mean time taken by male recipients sniffing introduced treatments
Figure 7: Mean time taken by female recipients
sniffing introduced treatments
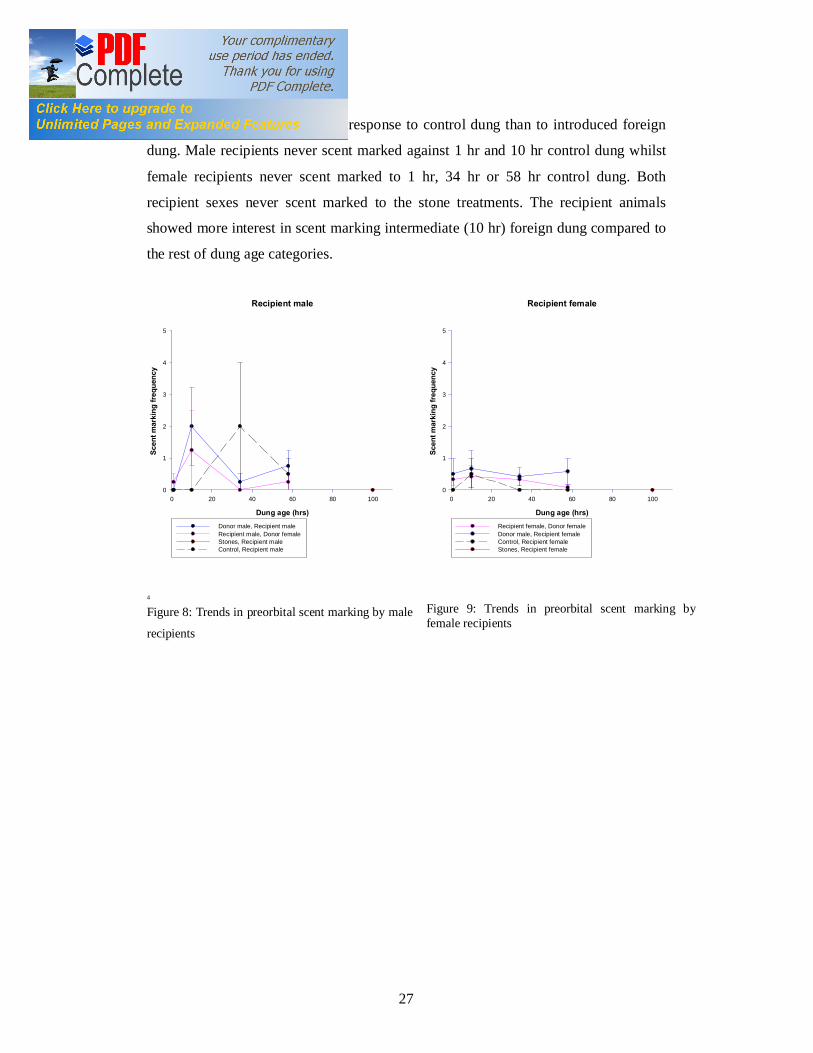
4.5 Preorbital Scent Marking by Blue Duikers to Introduced Treatments in
their Territories
27
Both sexes scent marked less in response to control dung than to introduced foreign
dung. Male recipients never scent marked against 1 hr and 10 hr control dung whilst
female recipients never scent marked to 1 hr, 34 hr or 58 hr control dung. Both
recipient sexes never scent marked to the stone treatments. The recipient animals
showed more interest in scent marking intermediate (10 hr) foreign dung compared to
the rest of dung age categories.
Recipient male
Dung age (hrs)
0 20 40 60 80 100
Scen
t mar
king
freq
uenc
y
0
1
2
3
4
5
Donor male, Recipient male Recipient male, Donor female Stones, Recipient male Control, Recipient male
Recipient female
Dung age (hrs)
0 20 40 60 80 100
Scen
t mar
king
freq
uenc
y
0
1
2
3
4
5
Recipient female, Donor female Donor male, Recipient female Control, Recipient female Stones, Recipient female
4
Figure 8: Trends in preorbital scent marking by male
recipients
Figure 9: Trends in preorbital scent marking by female recipients
28
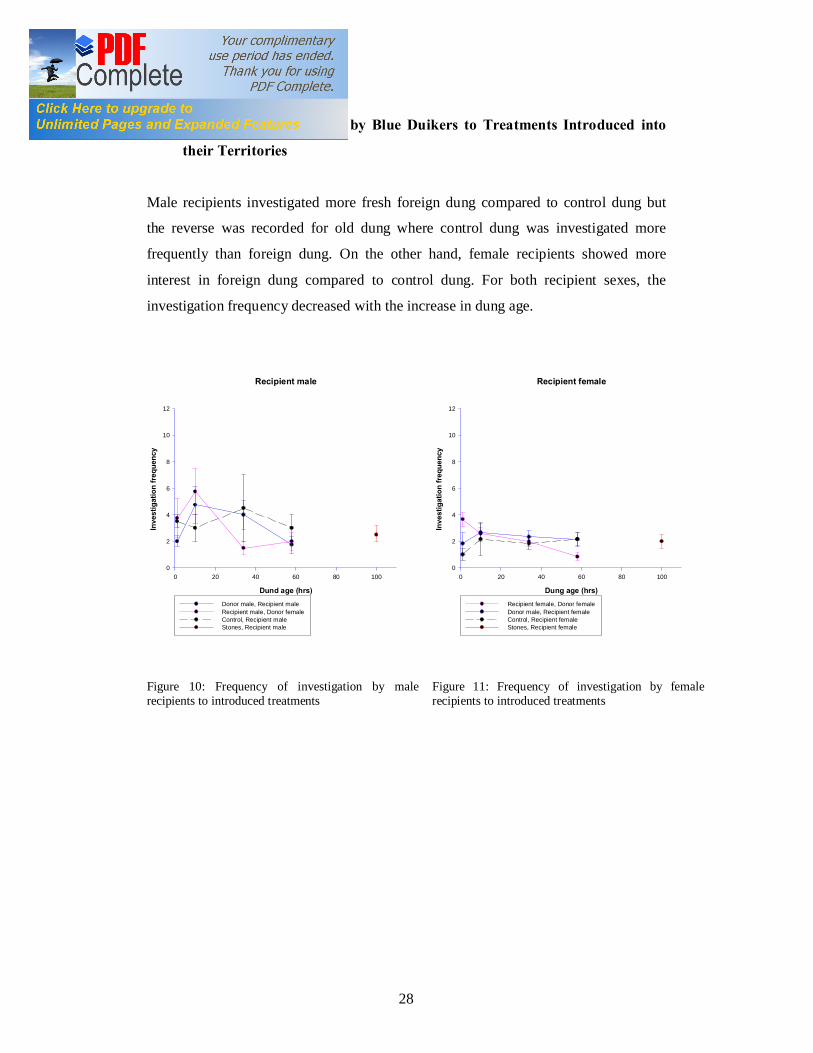
4.6 Investigation Frequency by Blue Duikers to Treatments Introduced into
their Territories
Male recipients investigated more fresh foreign dung compared to control dung but
the reverse was recorded for old dung where control dung was investigated more
frequently than foreign dung. On the other hand, female recipients showed more
interest in foreign dung compared to control dung. For both recipient sexes, the
investigation frequency decreased with the increase in dung age.
Recipient male
Dund age (hrs)
0 20 40 60 80 100
Inve
stig
atio
n fr
eque
ncy
0
2
4
6
8
10
12
Donor male, Recipient male Recipient male, Donor female Control, Recipient male Stones, Recipient male
Recipient female
Dung age (hrs)
0 20 40 60 80 100
Inve
stig
atio
n fr
eque
ncy
0
2
4
6
8
10
12
Recipient female, Donor female Donor male, Recipient female Control, Recipient female Stones, Recipient female
Figure 10: Frequency of investigation by male recipients to introduced treatments
Figure 11: Frequency of investigation by female recipients to introduced treatments

29
CHAPTER 5: DISCUSSION
5.0 Discussion
Unlike other antelope species such as klipspringer Oreotragus oreotragus (Roberts
and Lowen, 1997) and oribi Ourebia ourebi (Brashares and Arcese, 1999), blue
duikers were never observed licking or over marking with their own faecal pellets on
deposited foreign dung. Sniffing and scent marking with preorbital gland secretion
were the only forms of response noticed from both male and female blue duiker
recipient animals. This observation concurred with Estes (1997) who specified that
rubbing the preorbital glands on tree trunks and other objects within its environment,
is the main form of territory demarcation by blue duikers. However, scent marking
was not carried out in response to all introduced treatments especially control dung
suggesting that territory owners are able to distinguish between familiar and foreign
scent through scent matching (Gosling and McKay, 1990).
The study revealed that both sexes play a role in territory defense related activities
which is in agreement with Odendaal and Bigalke (1979) who mentioned that the blue
duiker is strongly territorial and both males and females mark out their territory
continually using urine, faecal pellets and preorbital gland secretion. Recipient
animals were reacting faster and investigating more to introduced foreign dung than to
control dung which suggest that they are territorial, and they scent mark more in
response to immediate perceived threats.
Neither urine nor faecal pellets were deposited in immediate response to foreign
faecal pellets deposited in their territory. However, faecal pellets were occasionally
seen deposited in enclosures, but mostly at fixed locations along borders shared with
other animals, which may be border marking to intimidate potential intruders and
cause them to withdraw from the marked territory (Roberts and Lowen, 1997).
Considering that defecation was mostly along territory boundaries, the possible reason
might be that the placement of marks is prioritized in areas where they can easily be
detected by the intended receiver, thus maximizing their value for territory
advertisement (ibid).

30
The fact that fresh and intermediate dung were investigated more than old dung
indicates that dung age had a significant effect on the intensity of response by
recipients. Consistent trends of decreasing response with increasing dung age which
were detected from this study concur with Ferkin et al., (1995). The other possible
reason for recipients showing more interest in fresh and intermediate dung compared
to old dung could be for defense purpose. If an owner notices foreign scent which is
fresh in its territory, it is possible that the intruder is still in the vicinity.
Consequently, the territory resident is expected to increase its marking rate in order to
cover a greater area with its own scent marks to maximize the probability of scent
matching by the intruder (Gosling, 1982).
Little sniffing was observed in response to control dung compared to foreign dung as
suggested by Gosling and MacKay (1990) that territory residents have got the ability
to differentiate between foreign and familiar scent. Consequently, they modulate their
response by spending more time investigating foreign scent so that they extract any
useful information needed about the signaler identity.
Blue duikers sniffed introduced stones regardless of them being scent neutral,
indicating that they also make use of visual cue in addition to olfactory and auditory
stimuli in territory maintenance and defense (Estes, 1997). Animals did not scent
mark in response to stones, signifying that they did not regard them as a territorial
threat but rather as something novel in their environment. This suggests that blue
duikers regulate the usage of scent marks and prioritize the placement of marks to
circumstances and areas where their value is worthy.
The findings of this research highlighted a slight increase in response (reduced lag
time, increased sniffing duration, increased scent mark frequency) when recipient
animals investigated dung from the same sex donors, most probably due to perceived
threat for mates. Basing on the findings by Wells and Bekoff (1981), scent marks
were reported to contribute in sex recognition which may suggests that same sex
animals might not tolerate each other’s company to reduce competition over potential
mates.

31
CHAPTER 6: CONCLUSION AND RECOMMENDATIONS
6.0 Conclusion
Blue duikers are primarily monogamous animals in which pair bonding is probably
life long. Direct interactions between animals of different social groups are rare
indicating that they are territorial. They defend their territory against potential
intruders using urine, faecal pellets and preorbital gland secretion.
Sniffing on or around the deposited dung pellets occasionally followed by preorbital
scent marking is the main form of response by blue duiker territory residents. The
marking densities generally increase in areas close to the deposited foreign faecal
pellets since the territory owner will be trying to mark with own scent to maximize
scent-matching probability. Urine and faeces are also deposited either at fixed
locations or randomly within the recipient animal pens but is not done in immediate
counter response to foreign substances found within the territory.
The efficacy of animal scent marks is not constant but varies with time after
deposition thus recipient animals react more to fresher dung compared to old one.
Dung scent retains a high degree of efficacy for some time and afterward the scent
signal starts to fade out.
Sex of either recipient or donor affects response intensity to a lesser extent because
both males and females are almost equally involved in territory defense related
activities. A slight increase in response intensity was noted when fresh strange scent
was introduced to recipient animals of the same sex to donor animals compared to
scent from donors of the opposite sex.

32
6.1 Recommendations
It is recommended that more experimental studies specifically on blue duiker
territorial behaviour must be conducted in the natural wild in order to have more
detailed and reliable information. There is also need to conduct a similar study using
dung aged more than 58 hours to see how long it takes for dung scent to be
undetectable.

33
REFERENCES
Alberts, A. C. (1992). Constraints on the design of chemical communication systems
in terrestrial vertebrates. Anim. Nat. 139, 62-89.
Anderson, D. J. (1982). The Home range: a new nonparametric estimation technique.
Ecol 63: 103-112.
Apps, P. J., Viljoen, H. W., Richardson, P. R. K. and Pretorius, V. (1989), Volatile
components of anal gland secretion of aardwolf (Proteles cristatus). J. Chem. Ecol.
15: 1681-1688.
Asa, C. S., Mech, L. D. and Seal, U. S (1985), The use of faeces, urine and anal gland
secretions in scent marking by a captive wolf (Canis lupus) pack. Anim Behav, 33:
1034-1036.
Bell, R. H. V. (1971), A grazing system in the Serengeti. Scientific American 224:
86-93.
Bodmer, R. E. (1990), Ungulate frugivores and the browser-grazer continuum. Oikos
57: 319-325.
Bowen-Jones, E. (1998), A review of the commercial bushmeat trade with emphasis
on Central/West Africa and the Great Apes.Report for the Ape Alliance. Fauna and
Flora International, Cambridge, UK.
Bowland, A. E., Perrin, M. R. (1995), Temporal and spatial patterns of forest duikers
Philatomba monticola and Cephalophus natalensis. J. Zool. 237: 487-498.
Bowman, V & Plowman, A. 2002. Captive Duiker Management at the Duiker and
Mini Antelope Breeding and Research Institute (Dambari), Bulawayo, Zimbabwe.
Zoo Biol 21: 161-170.
Bowyer, R. T. and Kitchen, D. W. (1987), Significance of scent marking. J.
Mammalogy 62:574-582.
Brashares, J.S. and Arcese, P. 1999a: Scent marking in African antelope: II. The

34
economics of marking with faeces. Anim. Behav. 57, 11-17.
Brown, R. E. (1985), Effects of odours on reproductive physiology. Clarendon,
Oxford.
Chardonnet, P; Fritz, H. (1995), Chasse traditionelle. In: Faune Sauvage Africaine La
Ressource Oubliee. Commission Europeenne. Luxembourg: Office des publications
officielles des Communautes europpeennes: Tome 1 : 88-125.
Davison, B. (1999), Zimbabwe’s cheetah policy management plan (Unpublished
report).
Dubost, G. (1983), Le comportement de Cephalophus monticola Thunberg et C.
dorsalis Gray, et la place des cephalophes au sein des ruminants. Mammalia 47(3):
141-177.
East, R. (1999), African Antelope Database 1998. IUCN/SSC Antelope Specialist
Group.
Occasional Paper of the IUCN Species Suvival Commission No.21. IUCN, Gland,
Switzerland and Cambridge, UK.
Estes, R. D. (1997), Behavior Guide to African Mammals, UK.
Eves, H. E. (1996), Socioeconomic study 1996: Consultants report submitted to the
wildlife conservation society and The World Bank (Global environment society).
New York: WCS.
Fa, J. E., Juste, J., Del Val, J. P and Castroviejo, J. (1995), Impact of market hunting
on mammal species in Equatorial Guinea. Conservation Biology 9(5): 1107-1115.
Feer, F. (1989), Comparison of diets of frugivorous ruminants. Mammalia 53: 563-
601.

35
Feer, F. (1995), Seed dispersal in African forest ruminants. Jl of Tropical Ecol 11:
683-689.
Fergus, C. (1982), Blue duikers come to America. In: Research/Penn State. no. 2.
Ferkin, M. H, Burda, J., O’Connor, M. P. and Lee, C. J. (1995), Persistance of the
attractiveness of two sex specific scents in meadow voles, Ethol 101: 228-238.
Fitzgibbon, C. D.; Mogaka, H.; Fanshaw, J. H. (1995), Subsistance hunting in
Arabuko-Sokoke Forest, Kenya, and its effects on mammal populations. Cons Biol
9(5):1116-1126.
Gosling, L. M. (1981), Demarcation in a gerenak territory: an economic approach.
Zeitschrift fur Tierpsychologie 56: 305-322.
Gosling, L. M. (1982), A reassessment of the function of scent marking in territories.
Z. Tierpsychol 60: 89-118.
Gosling, L. M. (1985), The even toed ungulates: order Artiodactyla. Oxford, Oxford
University Press.
Gosling, L. M. (1990), Scent marking by resource holders. Oxford University Press,
U K.
Gosling, L. M. and Mckay, H. V. (1990), Competitor assessment by scent matching.
Behav. Ecol. Sociobiol. 26: 415-420.
Gosling, L. M. and Roberts, S. C. (2000), Life history costs of olfactory status
signaling in mice. Behav. Ecol. Sociobiol. 48: 328-332.
Gosling, L. M., Roberts, S. C, Thornton, E. A. and Andrew, M. J., Life history costs
of olfactory status signaling in mice, Behav. Ecol. Sociobio. (in press).
Hanekom, N.; Wilson, V. J. (1991), Blue duiker (Philatomba Monticola) densities in

36
the Tsitisikamma National Park and probable factors limiting these populations.
Koedoe 34(2):107-120.
Hart, J. A. (2000), Impact and suatainability of indigenous hunting in the Uturi
forest, Congo-Zaire, New York, Columbia University Press: 106-153.
Hart, J. A.; Katembo, M.; Punga, K. (1996), Diet, prey selection and ecological
relations of leopard and golden cat in the Uturi Forest, Zaire. African Journal of Ecol
34:364-379.
IUCN/SSC Antelope specialist group (1998), African Antelope Database 1998.
Compiler R. East. Occasional Paper of the IUCN Species Survival Commission No.
21. Gland Switzerland.
Jarman, P. J. (1974), The social organization of antelope in relation to their ecology.
Behav 48:215-266.
Johnston, R. E and Schmidt (1979), Response of hamsters to scent marks of different
ages. Behav. Neural Biol. 26: 64-75.
Kingdon, J. (1997), Field guide to African Mammals, Academic Press, London.
Komers, P. E. (1996), Conflicting territory use in females and males of a
monogamous ungulate, the Kirk’s dikdik. Ethol, 102: 568-579.
Lahm, S. A. (1993), Ecology and economics of human/wildlife Interaction in
Northeastern Gabon. Ph.D. Dissertation. New York University.
Marker, L., Kraus, D., Barnett, D. and Hurlbut, S. (1996), Cheetah survival on
Namibian farmlands. Cheetah Conservation Fund. Solitaire Press, Windhoek.
Masoud, T. (2000), Ecological values and conservation of Jozani-Chwaka Bay
Conservation Area, Zanzibar. Commission for Natural Resources, Zanzibar.

37
Maynard Smith, J. (1974), The theory of games and the evolution of wildlife
conflicts. J. Theo. Biol. 47: 209-221.
McNaughton, S. J. (1987), Adaptation of herbivores to seasonal changes in nutrient
supply, London: Academic Press.
Melemis, S. M. and Falls, J. B. (1982), The defence function: a measure of territorial
behaviour. Can. J. Zool. 60: 495-501.
Milton, K. (1982), Dietary quality and demographic regulation in a howler monkey
population, Washington: Smithsonian Institution Press.
Mizutani, F. (1993), Home range of leopards and their impact on livestock on Kenyan
ranches. Symp. Zool. Soc. Lond. 65: 425-439.
Newing, H. S. (1994), Behavioural ecology of duikers (Cephalophus spp.) in forest
and secondary growth. PhD Thesis, University of Stirling, Scotland.
Noss, A. J. (1995), Duikers, cables and nets: a cultural ecology of hunting in a
central African forest. Ph.D. Dissertation. University of Florida.
Odendaal, P. B. and Bigalke, R. C (1979), Home range and groupings of bushbuck in
the southern Cape. South African journal of Wildlife Research 9: 96-101.
Ortmann, R. (1960), Die analregion der Saugetiere. Handbuch der Zoologie, 3: 1-68.
Ralls. K. (1971). Mammalian Scent marking. Science, 171: 443-449.
Roberts, S. C. (1998), Behavioural responses to scent marks of increasing age in
klipspringer. Ethol. 104: 585-592.
Robinson, J.: Redfoed, K. (eds.) (1998), Neotropical wildlife use and conservation.
Chicago: The University of Chicago Press.

38
Rorberts, S. C. (1994), Machanics and function of territorial behaviour in
Klipspringer. PhD thesis, University of London.
Rorberts, S. C. (1997), Selection of Scent marking sites by klipspringers (Oreotragus
oreotragus). J. Zool. (Lond) 243: 555-564.
Rorberts, S. C. and Lowen, C. (1997), Optimal patterns of scent marks in klipspringer
(Oreotragus oreotragus) territories. J. Zool. (Lond) 243: 565-574.
Sawyer, T. G., Marchinton, R. L., and Miller, K. V. (1989), Response of female
white- tailed deer to scrapes and antler rubs J. Mam. 70: 431-433.
Smithers, R. H. N. (1983), Mammals of Southern African Subregion 1st Edition,
University of Pretoria, South Africa.
Snyder, N.; Derrickson, S.; Miller, B. (1996), Limitations of captive breeding in
Endangered Species Recovery. Cons Biol 10: 2.
Spinage, C. A. (1986), The natural history of antelopes. London: Croom Helm.
Van Soest, P. J. (1983), Nutritional ecology of the Ruminant, Corvallis, O and B
Books.
Wells, M. C. and Bekoff, M. (1981), An observational study of scent marking in
coyotes, Canis latrans. Anim. Behav., 29: 332-350.
Western, D.; Pearl, M. (eds.) (1989), Conservation for the twenty-first Century.
Wildlife Conservation International. New York.
Williams, S. (2007), Status of cheetahs in Zimbabwe. Cat News 3: 32-36.
Wilson, V. (2005), Duikers of Africa: Zambi Books, Pretoria, South Africa.

39
Appendix 1: Sample Data Record Sheet
DATE: ______________ TIME IN: __________ TIME OUT:
______________
TREATMENT: ________ PEN: ______________ WEATHER:
______________
Time Recipient
Animal
Sniff
duration(s)
Scent
marking
Notes
Type Frequency


















