
Bactericidal Properties of Human Cell-Free Lymphiai.asm.org/content/13/3/768.full.pdf · As yet,...
8
INFECTION AND IMMUNITY, Mar. 1976, p. 768-775 Copyright © 1976 American Society for Microbiology Vol. 13, No. 3 Printed in USA. Bactericidal Properties of Human Cell-Free Lymph RAFAEL J. MARTINEZ Department of Bacteriology, University of California, Los Angeles, California 90024 Received for publication 20 November 1975 The bactericidal properties of human cell-free lymph have been investigated. Gram-negative and gram-positive bacteria are susceptible to the killing action of lymph fluid. The colony-forming ability of Escherichia coli and Bacillus subtilis was reduced by 3 logs after 20 min of incubation with 10% (vol/vol) lymph. Salmonella and Listeria were less susceptible to killing. The presence of multiple killing factors in lymph showing different specificities for different bacterial cells was demonstrated. It is known that a wide variety of bacteria are susceptible to the bactericidal effects of mam- malian serum. It is also generally accepted that for gram-negative bacteria the bactericidal ac- tion of serum is mediated by the antibody-com- plement system. A serum bactericidal agent effective against gram-positive bacteria has been termed G-lysin (8). It appears to be a com- plement-independent and nonspecific system which kills a wide array of gram-positive cells (2). There is suggestive evidence for the exist- ence of more than one 8-lysin-like system in serum exhibiting some specificity for different bacteria (4). As yet, the biochemical mode of action of these bactericides, either for gram- positive or gram-negative bacteria, has not been elucidated. There are suggestions that both the antibody-complement system and /8- lysin act on the cytoplasmic membrane, but the initial lesion(s) induced by these systems has not been rigorously defined. Although some limited information exists on the bactericidal activity of serum, essentially no information is available about the compara- ble activity of lymph. The majority of poten- tially disease-producing bacteria, be they bona fide pathogens or opportunistic ones, upon in- vasion cf an animal host initially contact lymph rather than serum upon penetration of tissue barriers. It may be presumed, therefore, that activities of the humoral as well as the cellular components of lymph would be of significance in protection against infection. An investiga- tion of the bactericidal properties of lymph fluid, freed of all cellular constituents, was therefore undertaken. This communication reports the results of experiments using unfractionated cell-free hu- man lymph fluid. Data are presented demon- strating the existence of bactericidal agents ef- fective in killing both gram-negative and gram- positive bacteria. MATERIALS AND METHODS Organisms and growth conditions. Escherichia coli ML35 (lacI, lacY) is constitutive for 3-galacto- sidase and is galactoside permease negative. It is a typical smooth colony type and is not susceptible to several rough-specific phages (B. A. D. Stocker, per- sonal communication). It was grown overnight from a slant inoculum in nutrient broth (Difco) at 37 C with agitation. The following morning, a 1:100 dilu- tion was made into prewarmed nutrient broth and incubation was continued until the optical density of the culture, measured in a Zeiss PMQII spectropho- tometer at 540 nm (OD540), reached approximately 0.4. This is equivalent to 7 x 107 cells/ml. The cul- ture was then in the early stages of logarithmic growth. Salmonella typhimurium strain Keller and S. typhimurium SL4044 were grown on nutrient broth in an identical manner. S. typhimurium Keller is a wild-type strain used by Robson and Vas (9) and shown by them to kill mice of various suscep- tible lines on intraperitoneal injection of less than 10 bacteria. B.A. D. Stocker of Stanford University has obtained similar results (personal communication). It shows a typical smooth-phage sensitivity pattern. Strain SL4044 (11) is an LT 2 derivative showing slightly rough characteristics. A nonflagellate (fla-) subline of Bacillus subtilis 168 (trp-) was grown from a slant inoculum in Spizi- zen' minimal medium (10) containing glucose (Q% wt/vol) at 37 C with agitation. A 1:10 dilution was made in the morning into prewarmed medium of the same composition and incubation continued until the OD540 reached 0.35, which is equivalent to 7 x 107 cells/ml. The culture was then in early logarith- mic growth. Listeria monocytogenes 44-144 was grown in Brucella broth (BBL) as above, using a 1:50 dilution to an OD540 at harvest at 0.45. At the prescribed times the cultures were har- vested by centrifugation at 12,000 x g for 5 min at 4 C. The cell pellet was suspended to a density of 101 cells/ml in 0.05 M potassium phosphate buffer, pH 7.4, containing 5 x 10-4 M Mg2+, 1.5 x 10-4 M Ca2+, and 1% glucose (glucose-phosphate buffer). The cell suspensions were chilled in ice and used immedi- ately. Control experiments with all oganisms used revealed that storage in ice for as long as 3 h did not 768 on July 16, 2018 by guest http://iai.asm.org/ Downloaded from
-
Upload
truongkiet -
Category
Documents
-
view
213 -
download
0
Transcript of Bactericidal Properties of Human Cell-Free Lymphiai.asm.org/content/13/3/768.full.pdf · As yet,...

INFECTION AND IMMUNITY, Mar. 1976, p. 768-775Copyright © 1976 American Society for Microbiology
Vol. 13, No. 3Printed in USA.
Bactericidal Properties of Human Cell-Free LymphRAFAEL J. MARTINEZ
Department of Bacteriology, University of California, Los Angeles, California 90024
Received for publication 20 November 1975
The bactericidal properties of human cell-free lymph have been investigated.Gram-negative and gram-positive bacteria are susceptible to the killing actionof lymph fluid. The colony-forming ability of Escherichia coli and Bacillussubtilis was reduced by 3 logs after 20 min of incubation with 10% (vol/vol)lymph. Salmonella and Listeria were less susceptible to killing. The presence ofmultiple killing factors in lymph showing different specificities for differentbacterial cells was demonstrated.
It is known that a wide variety of bacteria aresusceptible to the bactericidal effects of mam-malian serum. It is also generally accepted thatfor gram-negative bacteria the bactericidal ac-tion of serum is mediated by the antibody-com-plement system. A serum bactericidal agenteffective against gram-positive bacteria hasbeen termed G-lysin (8). It appears to be a com-plement-independent and nonspecific systemwhich kills a wide array of gram-positive cells(2). There is suggestive evidence for the exist-ence of more than one 8-lysin-like system inserum exhibiting some specificity for differentbacteria (4). As yet, the biochemical mode ofaction of these bactericides, either for gram-positive or gram-negative bacteria, has notbeen elucidated. There are suggestions thatboth the antibody-complement system and /8-lysin act on the cytoplasmic membrane, but theinitial lesion(s) induced by these systems hasnot been rigorously defined.Although some limited information exists on
the bactericidal activity of serum, essentiallyno information is available about the compara-ble activity of lymph. The majority of poten-tially disease-producing bacteria, be they bonafide pathogens or opportunistic ones, upon in-vasion cfan animal host initially contact lymphrather than serum upon penetration of tissuebarriers. It may be presumed, therefore, thatactivities of the humoral as well as the cellularcomponents of lymph would be of significancein protection against infection. An investiga-tion of the bactericidal properties of lymphfluid, freed of all cellular constituents, wastherefore undertaken.This communication reports the results of
experiments using unfractionated cell-free hu-man lymph fluid. Data are presented demon-strating the existence of bactericidal agents ef-fective in killing both gram-negative and gram-positive bacteria.
MATERIALS AND METHODSOrganisms and growth conditions. Escherichia
coli ML35 (lacI, lacY) is constitutive for 3-galacto-sidase and is galactoside permease negative. It is atypical smooth colony type and is not susceptible toseveral rough-specific phages (B. A. D. Stocker, per-sonal communication). It was grown overnight froma slant inoculum in nutrient broth (Difco) at 37 Cwith agitation. The following morning, a 1:100 dilu-tion was made into prewarmed nutrient broth andincubation was continued until the optical density ofthe culture, measured in a Zeiss PMQII spectropho-tometer at 540 nm (OD540), reached approximately0.4. This is equivalent to 7 x 107 cells/ml. The cul-ture was then in the early stages of logarithmicgrowth. Salmonella typhimurium strain Keller andS. typhimurium SL4044 were grown on nutrientbroth in an identical manner. S. typhimuriumKeller is a wild-type strain used by Robson and Vas(9) and shown by them to kill mice of various suscep-tible lines on intraperitoneal injection of less than 10bacteria. B.A. D. Stocker of Stanford University hasobtained similar results (personal communication).It shows a typical smooth-phage sensitivity pattern.Strain SL4044 (11) is an LT 2 derivative showingslightly rough characteristics.A nonflagellate (fla-) subline of Bacillus subtilis
168 (trp-) was grown from a slant inoculum in Spizi-zen' minimal medium (10) containing glucose (Q%wt/vol) at 37 C with agitation. A 1:10 dilution wasmade in the morning into prewarmed medium of thesame composition and incubation continued untilthe OD540 reached 0.35, which is equivalent to 7 x107 cells/ml. The culture was then in early logarith-mic growth. Listeria monocytogenes 44-144 wasgrown in Brucella broth (BBL) as above, using a 1:50dilution to an OD540 at harvest at 0.45.
At the prescribed times the cultures were har-vested by centrifugation at 12,000 x g for 5 min at4 C. The cell pellet was suspended to a density of 101cells/ml in 0.05 M potassium phosphate buffer, pH7.4, containing 5 x 10-4 M Mg2+, 1.5 x 10-4 M Ca2+,and 1% glucose (glucose-phosphate buffer). The cellsuspensions were chilled in ice and used immedi-ately. Control experiments with all oganisms usedrevealed that storage in ice for as long as 3 h did not
768
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
BACTERICIDAL PROPERTIES OF HUMAN LYMPH 769
affect the viability of the cell suspensions, nor theirsusceptibility (or resistance) to the bactericidal ac-tion oflymph. The addition of glucose to the dilutionbuffer is essential to prevent autolysis ofB. subtilis.It is not essential for the maintenance of viable E.coli, Salmonella, or Listeria in buffer.
Bactericidal assay. The incubation mixture con-sisted of approximately 10' cells/ml, cell-free lymphfluid at the desired concentration (vol/vol), and glu-cose-phosphate buffer. The lymph-buffer mixtureswere prewarmed to 37 C for 2 min before initiatingthe reaction by addition of bacteria. Incubation wasat 37 C. Samples were removed at timed intervalsand serially diluted in glucose-phosphate buffer,and appropriate dilutions were plated on nutrientagar (Difco) (Brucella agar for Listeria) by the soft-agar overlay technique. The plates were incubatedat 37 C for 18 to 24 h and the resulting bacterialcolonies were counted with the aid of a New Bruns-wick colony counter. Each experiment was per-formed at least three times. The rate of reduction ofcolony-forming units (CFU) in these experiments isused as the measure of the kinetics of killing.
Lysis of cells in the incubation mixture was moni-tored in a Zeiss PMQII spectrophotometer at 540 nmat a chamber temperature of 37 C. Bacterial lysisand clumping were also monitored by counting cellsand clumps using a Petroff-Hauser counting cham-ber under phase optics.Human cell-free lymph was kindly provided by
Harold Paulus, UCLA Center for Health Sciences. Itwas obtained by thoracic duct drainage of patientsas part of treatment for rheumatoid arthritis (7).The lymph samples used in this study were obtainedfrom patients not undergoing chemotherapy. Thelymph was stored at -70 C in small aliquots. Insome experiments the cell-free lymph was cleared oflipids by high-speed centrifugation. No significantdifference in killing activity was observed betweennormal lymph and lipid-cleared lymph.
RESULTSPreliminary experiments using E. coli as test
organism and 10% cell-free lymph as bacteri-cidal agent demonstrated a marked reductionin CFU as a function of incubation time. Thesensitivity of the bacterial cells to killing bylymph was found to be profoundly affected bythe physiological age ofthe culture. Cultures inthe early stages of logarithmic growth weremost sensitive, as has been reported for sensi-tivity to serum (6). Cultures in the early stagesof logarithmic growth were, therefore, used infurther studies.
In published studies on the bactericidal prop-erties of serum and other tissue bactericidalagents, the starting cell densities used in differ-ent laboratories have varied from approxi-mately 10:1 to more than 108 cells/ml. A startingdensity of 107 cells/ml was selected after controlexperiments showed that killing kinetics didnot fluctuate significantly as a function of start-
ing cell densities (from 105 to 1081/ml). Further,agglutination ofE. coli and B. subtilis in lymphdid not occur until the density exceeded 2 x 108to 3 x 108 cells/ml. This starting cell densityallowed adequate dilution of the killing agentbefore plating for CFU.The possibility that the bactericidal potency
of lymph decays, even though it was stored at-70 C, was examined. No decrease in killingability was observed during a 6-month storagetime. Control experiments revealed no loss ofkilling ability with up to six freeze-thaw cycles.With these variables controlled, the kinetics
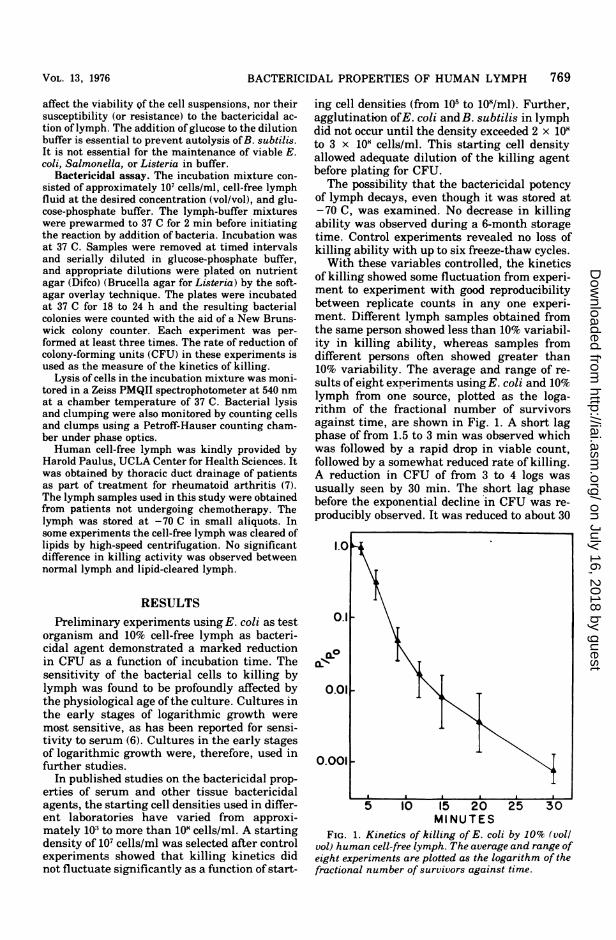
of killing showed some fluctuation from experi-ment to experiment with good reproducibilitybetween replicate counts in any one experi-ment. Different lymph samples obtained fromthe same person showed less than 10% variabil-ity in killing ability, whereas samples fromdifferent persons often showed greater than10% variability. The average and range of re-sults of eight experiments using E. coli and 10%lymph from one source, plotted as the loga-rithm of the fractional number of survivorsagainst time, are shown in Fig. 1. A short lagphase of from 1.5 to 3 min was observed whichwas followed by a rapid drop in viable count,followed by a somewhat reduced rate of killing.A reduction in CFU of from 3 to 4 logs wasusually seen by 30 min. The short lag phasebefore the exponential decline in CFU was re-producibly observed. It was reduced to about 30
1.0
0.I
a>
0.01-
0.001 1
MINUTESFIG. 1. Kinetics of killing of E. coli by 10% (voll
vol) human cell-free lymph. The average and range ofeight experiments are plotted as the logarithm of thefractional number of survivors against time.
VOL. 13, 1976
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
770 MARTINEZ
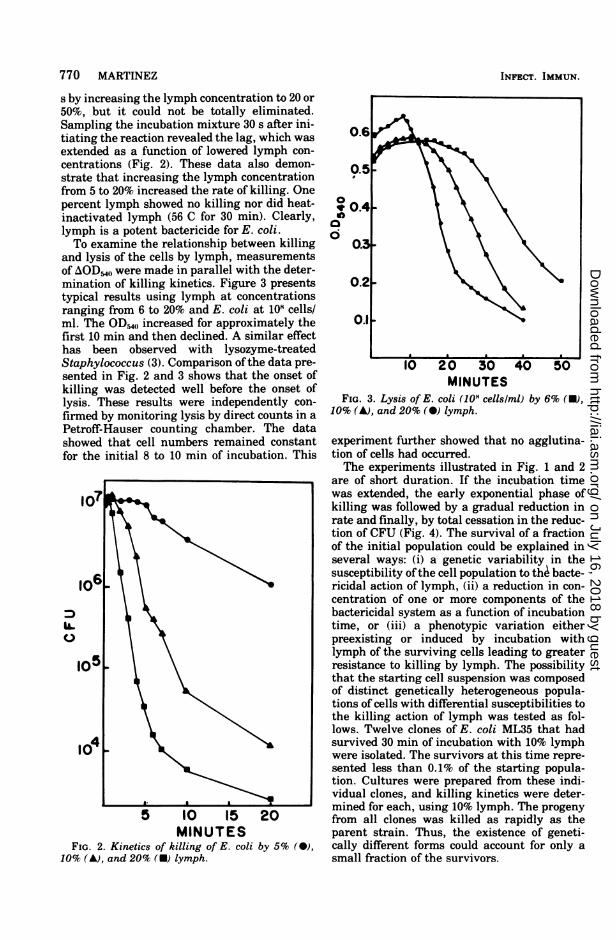
s by increasing the lymph concentration to 20 or50%, but it could not be totally eliminated.Sampling the incubation mixture 30 s after ini-tiating the reaction revealed the lag, which wasextended as a function of lowered lymph con-centrations (Fig. 2). These data also demon-strate that increasing the lymph concentrationfrom 5 to 20% increased the rate of killing. Onepercent lymph showed no killing nor did heat-inactivated lymph (56 C for 30 min). Clearly,lymph is a potent bactericide for E. coli.To examine the relationship between killing
and lysis of the cells by lymph, measurementsof AODM,,( were made in parallel with the deter-mination of killing kinetics. Figure 3 presentstypical results using lymph at concentrationsranging from 6 to 20% and E. coli at 101 cells/ml. The OD,40 increased for approximately thefirst 10 min and then declined. A similar effecthas been observed with lysozyme-treatedStaphylococcus (3). Comparison ofthe data pre-sented in Fig. 2 and 3 shows that the onset ofkilling was detected well before the onset oflysis. These results were independently con-firmed by monitoring lysis by direct counts in aPetroff-Hauser counting chamber. The datashowed that cell numbers remained constantfor the initial 8 to 10 min of incubation. This
107
MINUTESFIG. 2. Kinetics of killing of E. coli by 5% (0),
10% (A), and 20% (U) lymph.
0
CX00i
MINUTESFIG. 3. Lysis of E. coli (108 cellslml) by 6% (U),
10% (A), and 20% (0) lymph.
experiment further showed that no agglutina-tion of cells had occurred.The experiments illustrated in Fig. 1 and 2
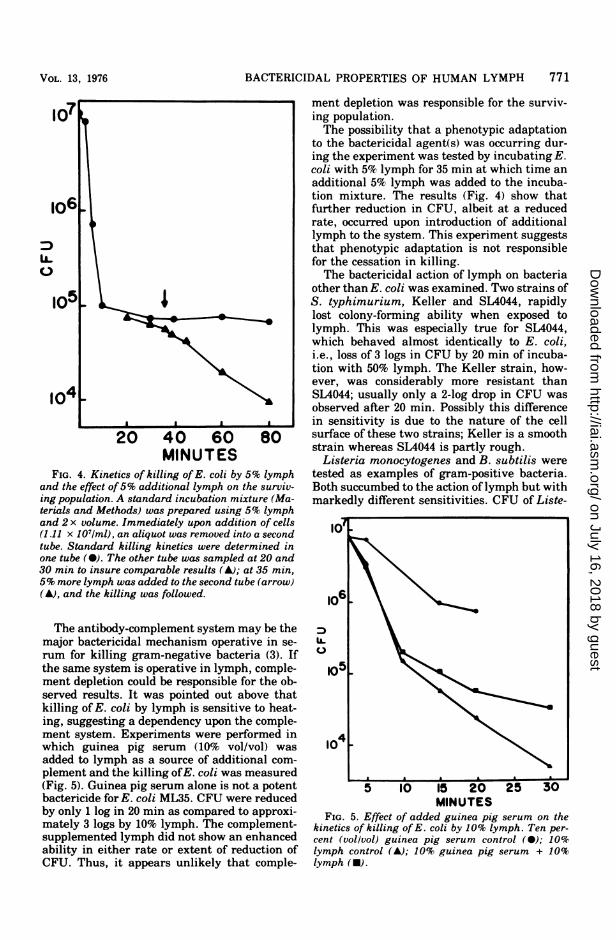
are of short duration. If the incubation timewas extended, the early exponential phase ofkilling was followed by a gradual reduction inrate and finally, by total cessation in the reduc-tion of CFU (Fig. 4). The survival of a fractionof the initial population could be explained inseveral ways: (i) a genetic variability in thesusceptibility ofthe cell population to the bacte-ricidal action of lymph, (ii) a reduction in con-centration of one or more components of thebactericidal system as a function of incubationtime, or (iii) a phenotypic variation eitherpreexisting or induced by incubation withlymph of the surviving cells leading to greaterresistance to killing by lymph. The possibilitythat the starting cell suspension was composedof distinct genetically heterogeneous popula-tions of cells with differential susceptibilities tothe killing action of lymph was tested as fol-lows. Twelve clones of E. coli ML35 that hadsurvived 30 min of incubation with 10% lymphwere isolated. The survivors at this time repre-sented less than 0.1% of the starting popula-tion. Cultures were prepared from these indi-vidual clones, and killing kinetics were deter-mined for each, using 10% lymph. The progenyfrom all clones was killed as rapidly as theparent strain. Thus, the existence of geneti-cally different forms could account for only asmall fraction of the survivors.
INFECT. IMMUJN.
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
BACTERICIDAL PROPERTIES OF HUMAN LYMPH 771
20 40 60 80
MINUTESFIG. 4. Kinetics of killing of E. coli by 5% lymph
and the effect of5% additional lymph on the surviv-ing population. A standard incubation mixture (Ma-terials and Methods) was prepared using 5% lymphand 2 x volume. Immediately upon addition of cells(1.11 x 10'/ml), an aliquot was removed into a secondtube. Standard killing kinetics were determined inone tube (0). The other tube was sampled at 20 and30 min to insure comparable results (A); at 35 min,5% more lymph was added to the second tube (arrow)(A), and the killing was followed.
The antibody-complement system may be themajor bactericidal mechanism operative in se-
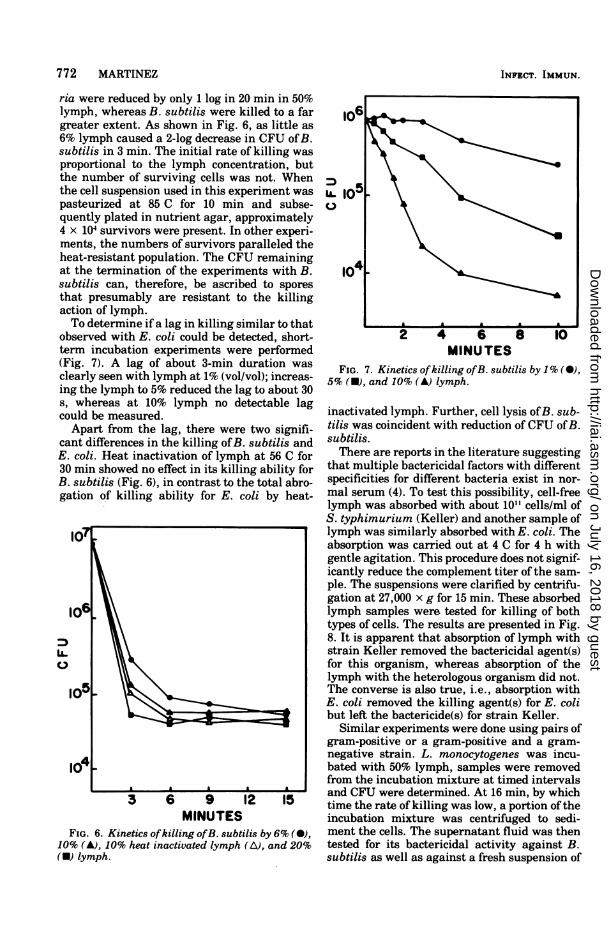
rum for killing gram-negative bacteria (3). Ifthe same system is operative in lymph, comple-ment depletion could be responsible for the ob-served results. It was pointed out above thatkilling of E. coli by lymph is sensitive to heat-ing, suggesting a dependency upon the comple-ment system. Experiments were performed inwhich guinea pig serum (10% vol/vol) wasadded to lymph as a source of additional com-plement and the killing ofE. coli was measured(Fig. 5). Guinea pig serum alone is not a potentbactericide for E. coli ML35. CFU were reducedby only 1 log in 20 min as compared to approxi-mately 3 logs by 10% lymph. The complement-supplemented lymph did not show an enhancedability in either rate or extent of reduction ofCFU. Thus, it appears unlikely that comple-
ment depletion was responsible for the surviv-ing population.The possibility that a phenotypic adaptation
to the bactericidal agent(s) was occurring dur-ing the experiment was tested by incubating E.coli with 5% lymph for 35 min at which time anadditional 5% lymph was added to the incuba-tion mixture. The results (Fig. 4) show thatfurther reduction in CFU, albeit at a reducedrate, occurred upon introduction of additionallymph to the system. This experiment suggeststhat phenotypic adaptation is not responsiblefor the cessation in killing.The bactericidal action of lymph on bacteria
other than E. coli was examined. Two strains ofS. typhimurium, Keller and SL4044, rapidlylost colony-forming ability when exposed tolymph. This was especially true for SL4044,which behaved almost identically to E. coli,i.e., loss of 3 logs in CFU by 20 min of incuba-tion with 50% lymph. The Keller strain, how-ever, was considerably more resistant thanSL4044; usually only a 2-log drop in CFU wasobserved after 20 min. Possibly this differencein sensitivity is due to the nature of the cellsurface of these two strains; Keller is a smoothstrain whereas SL4044 is partly rough.Listeria monocytogenes and B. subtilis were
tested as examples of gram-positive bacteria.Both succumbed to the action oflymph but withmarkedly different sensitivities. CFU of Liste-
o6
U.)
MINUTESFIG. 5. Effect of added guinea pig serum on the
kinetics of killing of E. coli by 10% lymph. Ten per-cent (vollvol) guinea pig serum control (0); 10%lymph control (A); 10% guinea pig serum + 10%lymph (U).
VOL. 13, 1976
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
772 MARTINEZ
ria were reduced by only 1 log in 20 min in 50%lymph, whereas B. subtilis were killed to a fargreater extent. As shown in Fig. 6, as little as6% lymph caused a 2-log decrease in CFU ofB.subtilis in 3 min. The initial rate of killing wasproportional to the lymph concentration, butthe number of surviving cells was not. Whenthe cell suspension used in this experiment waspasteurized at 85 C for 10 min and subse-quently plated in nutrient agar, approximately4 x 104 survivors were present. In other experi-ments, the numbers of survivors paralleled theheat-resistant population. The CFU remainingat the termination of the experiments with B.subtilis can, therefore, be ascribed to sporesthat presumably are resistant to the killingaction of lymph.To determine if a lag in killing similar to that
observed with E. coli could be detected, short-term incubation experiments were performed(Fig. 7). A lag of about 3-min duration wasclearly seen with lymph at 1% (vol/vol); increas-ing the lymph to 5% reduced the lag to about 30s, whereas at 10% lymph no detectable lagcould be measured.Apart from the lag, there were two signifi-
cant differences in the killing ofB. subtilis andE. coli. Heat inactivation of lymph at 56 C for30 min showed no effect in its killing ability forB. subtilis (Fig. 6), in contrast to the total abro-gation of killing ability for E. coli by heat-
I0
I0
MINUTESFIG. 6. Kinetics ofkilling ofB. subtilis by 6% (0),
10% (A), 10% heat inactivated lymph (A), and 20%(U) lymph.
U.1
104 9
2 4 6 8 10MINUTES
FIG. 7. Kinetics ofkilling ofB. subtilis by 1% (M),5% (M), and 10% (A) lymph.
inactivated lymph. Further, cell lysis ofB. sub-tilis was coincident with reduction of CFU ofB.subtilis.There are reports in the literature suggesting
that multiple bactericidal factors with differentspecificities for different bacteria exist in nor-mal serum (4). To test this possibility, cell-freelymph was absorbed with about 1011 cells/ml ofS. typhimurium (Keller) and another sample oflymph was similarly absorbed with E. coli. Theabsorption was carried out at 4 C for 4 h withgentle agitation. This procedure does not signif-icantly reduce the complement titer of the sam-ple. The suspensions were clarified by centrifu-gation at 27,000 x g for 15 min. These absorbedlymph samples were tested for killing of bothtypes of cells. The results are presented in Fig.8. It is apparent that absorption of lymph withstrain Keller removed the bactericidal agent(s)for this organism, whereas absorption of thelymph with the heterologous organism did not.The converse is also true, i.e., absorption withE. coli removed the killing agent(s) for E. colibut left the bactericide(s) for strain Keller.
Similar experiments were done using pairs ofgram-positive or a gram-positive and a gram-negative strain. L. monocytogenes was incu-bated with 50% lymph, samples were removedfrom the incubation mixture at timed intervalsand CFU were determined. At 16 min, by whichtime the rate of killing was low, a portion oftheincubation mixture was centrifuged to sedi-ment the cells. The supernatant fluid was thentested for its bactericidal activity against B.subtilis as well as against a fresh suspension of
INFECT. IMMUN.
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
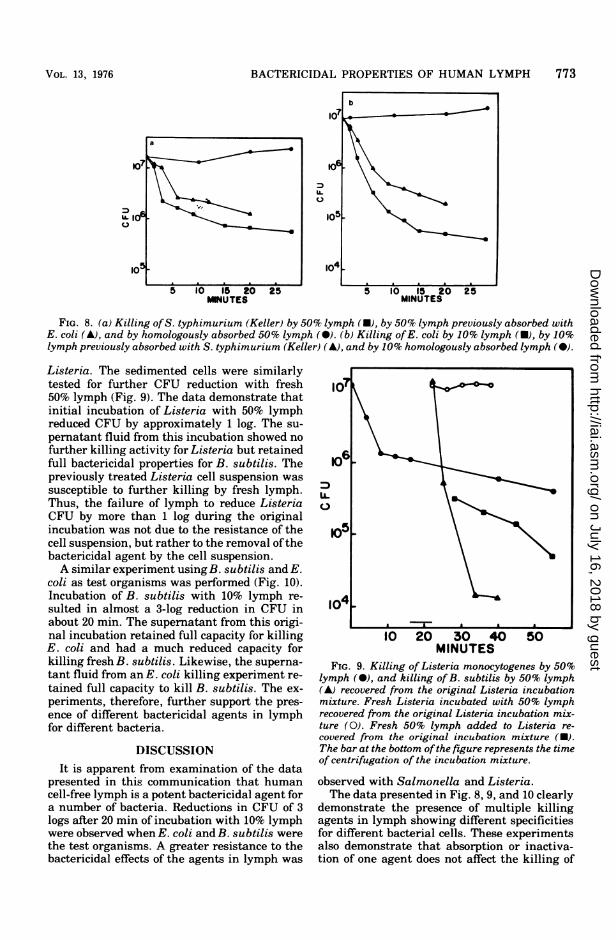
BACTERICIDAL PROPERTIES OF HUMAN LYMPH 773
MNUTES
FIG. 8. (a) Killing ofS. typhimurium (Keller) by 50% lymph (U), by 50% lymph previously absorbed withE. coli (A), and by homologously absorbed 50% lymph (0). (b) Killing ofE. coli by 10% lymph (U), by 10%lymph previously absorbed with S. typhimurium (Keller) (A), and by 10% homologously absorbed lymph (0).
Listeria. The sedimented cells were similarlytested for further CFU reduction with fresh50% lymph (Fig. 9). The data demonstrate thatinitial incubation of Listeria with 50% lymphreduced CFU by approximately 1 log. The su-pernatant fluid from this incubation showed nofurther killing activity for Listeria but retainedfull bactericidal properties for B. subtilis. Thepreviously treated Listeria cell suspension wassusceptible to further killing by fresh lymph.Thus, the failure of lymph to reduce ListeriaCFU by more than 1 log during the originalincubation was not due to the resistance of thecell suspension, but rather to the removal ofthebactericidal agent by the cell suspension.A similar experiment using B. subtilis and E.
coli as test organisms was performed (Fig. 10).Incubation of B. subtilis with 10% lymph re-sulted in almost a 3-log reduction in CFU inabout 20 min. The supernatant from this origi-nal incubation retained full capacity for killingE. coli and had a much reduced capacity forkilling fresh B. subtilis. Likewise, the superna-tant fluid from an E. coli killing experiment re-tained full capacity to kill B. subtilis. The ex-periments, therefore, further support the pres-ence of different bactericidal agents in lymphfor different bacteria.
DISCUSSIONIt is apparent from examination of the data
presented in this communication that humancell-free lymph is a potent bactericidal agent fora number of bacteria. Reductions in CFU of 3logs after 20 min of incubation with 10% lymphwere observed when E. coli and B. subtilis werethe test organisms. A greater resistance to thebactericidal effects of the agents in lymph was
MINUTESFIG. 9. Killing of Listeria monocytogenes by 50%
lymph (0), and killing of B. subtilis by 50% lymph(A) recovered from the original Listeria incubationmixture. Fresh Listeria incubated with 50% lymphrecovered from the original Listeria incubation mix-ture (0). Fresh 50% lymph added to Listeria re-covered from the original incubation mixture (U).The bar at the bottom ofthe figure represents the timeof centrifugation of the incubation mixture.
observed with Salmonella and Listeria.The data presented in Fig. 8, 9, and 10 clearly
demonstrate the presence of multiple killingagents in lymph showing different specificitiesfor different bacterial cells. These experimentsalso demonstrate that absorption or inactiva-tion of one agent does not affect the killing of
VOL. 13, 1976
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
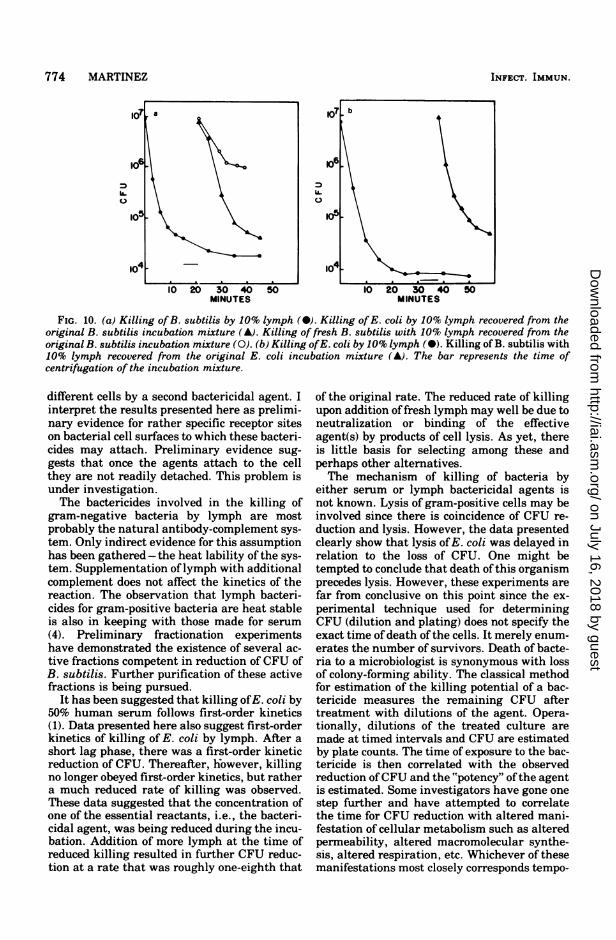
774 MARTINEZ
,5~
104'z_1 <10 20 30 40 50 10 20 30 40 50
MINUTES MINUTES
FIG. 10. (a) Killing of B. subtilis by 10% lymph (M). Killing ofE. coli by 10% lymph recovered from theoriginal B. subtilis incubation mixture (A). Killing of fresh B. subtilis with 10% lymph recovered from theoriginal B. subtilis incubation mixture (0). (b) Killing ofE. coli by 10% lymph (a). Killing of B. subtilis with10% lymph recovered from the original E. coli incubation mixture (A). The bar represents the time ofcentrifugation of the incubation mixture.
different cells by a second bactericidal agent. Iinterpret the results presented here as prelimi-nary evidence for rather specific receptor siteson bacterial cell surfaces to which these bacteri-cides may attach. Preliminary evidence sug-gests that once the agents attach to the cellthey are not readily detached. This problem isunder investigation.The bactericides involved in the killing of
gram-negative bacteria by lymph are mostprobably the natural antibody-complement sys-tem. Only indirect evidence for this assumptionhas been gathered - the heat lability of the sys-tem. Supplementation oflymph with additionalcomplement does not affect the kinetics of thereaction. The observation that lymph bacteri-cides for gram-positive bacteria are heat stableis also in keeping with those made for serum(4). Preliminary fractionation experimentshave demonstrated the existence of several ac-tive fractions competent in reduction of CFU ofB. subtilis. Further purification of these activefractions is being pursued.
It has been suggested that killing ofE. coli by50% human serum follows first-order kinetics(1). Data presented here also suggest first-orderkinetics of killing of E. coli by lymph. After ashort lag phase, there was a first-order kineticreduction of CFU. Thereafter, however, killingno longer obeyed first-order kinetics, but rathera much reduced rate of killing was observed.These data suggested that the concentration ofone of the essential reactants, i.e., the bacteri-cidal agent, was being reduced during the incu-bation. Addition of more lymph at the time ofreduced killing resulted in further CFU reduc-tion at a rate that was roughly one-eighth that
of the original rate. The reduced rate of killingupon addition offresh lymph may well be due toneutralization or binding of the effectiveagent(s) by products of cell lysis. As yet, thereis little basis for selecting among these andperhaps other alternatives.The mechanism of killing of bacteria by
either serum or lymph bactericidal agents isnot known. Lysis of gram-positive cells may beinvolved since there is coincidence of CFU re-duction and lysis. However, the data presentedclearly show that lysis ofE. coli was delayed inrelation to the loss of CFU. One might betempted to conclude that death of this organismprecedes lysis. However, these experiments arefar from conclusive on this point since the ex-perimental technique used for determiningCFU (dilution and plating) does not specify theexact time ofdeath ofthe cells. It merely enum-erates the number of survivors. Death of bacte-ria to a microbiologist is synonymous with lossof colony-forming ability. The classical methodfor estimation of the killing potential of a bac-tericide measures the remaining CFU aftertreatment with dilutions of the agent. Opera-tionally, dilutions of the treated culture aremade at timed intervals and CFU are estimatedby plate counts. The time of exposure to the bac-tericide is then correlated with the observedreduction ofCFU and the "potency" ofthe agentis estimated. Some investigators have gone onestep further and have attempted to correlatethe time for CFU reduction with altered mani-festation of cellular metabolism such as alteredpermeability, altered macromolecular synthe-sis, altered respiration, etc. Whichever of thesemanifestations most closely corresponds tempo-
INFECT. IMMUN.
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BACTERICIDAL PROPERTIES OF HUMAN LYMPH 775
rally with CFU reduction is usually assumed tobe the cause of death of the cells. This approachmay have one significant fallacy- the treatedcells are almost invariably plated with the bac-tericide "attached"; i.e., no conscious effort ismade to remove the agent from the cells beforeplating except dilution, and from our data thismay not be adequate. Without removal of bac-tericides from treated cells before plating, ameaningful estimation of the time when cellsare "committed" or "irreversibly committed" todeath is virtually impossible. These two puta-tive stages may well be temporally, and thusmeasurably, distinct since "healing" of par-tially affected targets may occur in the platingmedium. Thus, it would be fallacious to corre-late the lysis and CFU reduction data pre-sented here since some of the cells plated after30 s of treatment with lymph may lyse duringthe 24-h incubation for CFU. Further, the timeof death of the plated organisms is not known,only that colonies do not appear after the 24-hincubation time.Work is in progress to purify and character-
ize the bactericidal factors from lymph and se-rum with the ultimate aim of determining thebiochemical mode of action of these agents.
ACKNOWLEDGMENTSThis investigation was supported in part by Public
Health Service grants AI 11241 and AI 12835 from theNational Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Davis, S. D., and R. J. Wedgewood. 1965. Kinetics ofthe bactericidal action of normal serum on Gram-negative bacteria. J. Immunol. 95:75-79.
2. Donaldson, D. M. 1973. ,-Lysin and host resistance, p.316-322. In W. Braun and J. Ungar (ed.), Non-specificfators influencing host resistance, S. Karger, NewYork.
3. Inoue, K. 1972. Immune bacteriolytic and bactericidalreactions, p. 177-222. In J. B. G. Kwapinski (ed.),Research in immunochemistry and immunobiology,vol. 1. University Park Press, Baltimore.
4. Johnson, F. B., and D. M. Donaldson. 1968. Purifica-tion of staphylocidal (8-lysin from rabbit serum. J.Bacteriol. 96:589-595.
5. Metcalf, R. H., and R. H. Deibel. 1969. Differentiallytic response of enterococci associated with additionorder of lysozyme and anions. J. Bacteriol. 99:674-680.
6. Michael, J. G., and W. Braun. 1959. Modification ofbactericidal effects of human serum. Proc. Soc. Exp.Biol. Med. 102:486-490.
7. Paulus, H. E., H. Machleder, R. Bangert, J. A. Strat-ton, L. Goldberg, M. W. Whitehouse, D. Yu, and C.M. Pearson. 1973. A case report: thoracic duct lym-phocyte drainage in rheumatoid arthritis. Clin. Im-munol. Immunopathol. 1:173-181.
8. Pettersson, A. 1924. Ueber die Thermostabilen Bakter-iolytischen substanzen des Normalserums. Z. Immu-nitatsforsch. 40:43-56.
9. Robson, H. G., and S. I. Vas. 1972. Resistance of inbredmice to Salmonella typhimurium. J. Infect. Dis.126:378-383.
10. Spizizen, J. 1958. Transformation of biochemically defi-cient strains of Bacillus subtilis by deoxyribonu-cleate. Proc. Natl. Acad. Sci. U.S.A. 44:1072-1078.
11. Vary, P. S., and B. A. D. Stocker. 1973. Nonsensemotility mutants in Salmonella typhimurium. Ge-netics 73:229-245.
VOL. 13, 1976
on July 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from


















