Bacterial Cell Differentiation
66
Bacterial Cell Differentiation By: Naghmeh Poorinmohammad December 2014
Transcript of Bacterial Cell Differentiation
Bacterial Cell Differentiation
By: Naghmeh Poorinmohammad
December 2014

Outline
• Introduction
• Differentiation in bacteria
I. Single-cell Differentiations
II. Multicellular Differentiations
• Summary
• References

Introduction
• Living cells can change morphologically.
• Important reasons for differentiation:
1. Adaption to environmental conditions
2. Expressing different functions at different times in the life cycle
• Thus differentiation can be triggered by:
1. Environmental signal
2. No external signal inherent to the growth
• The specialized cell types is the outcome of complex regulatory pathways:
1. Altered gene expression
2. differential protein stability
3. differential protein localization
Chater, Keith F. "Bacterial Cell Differentiation." eLS (2005).
3

I. Single-Cell Differentiations
1. Dimorphic life cycle in Caulobacter crescentus
2. Endospore formation in Bacillus sibtilis
3. Actinomycetes spores
4. Cysts in Cyanobacteria
5. Heterocysts in Azotobacter
4

Dimorphic life cycle in
Caulobacter crescentus
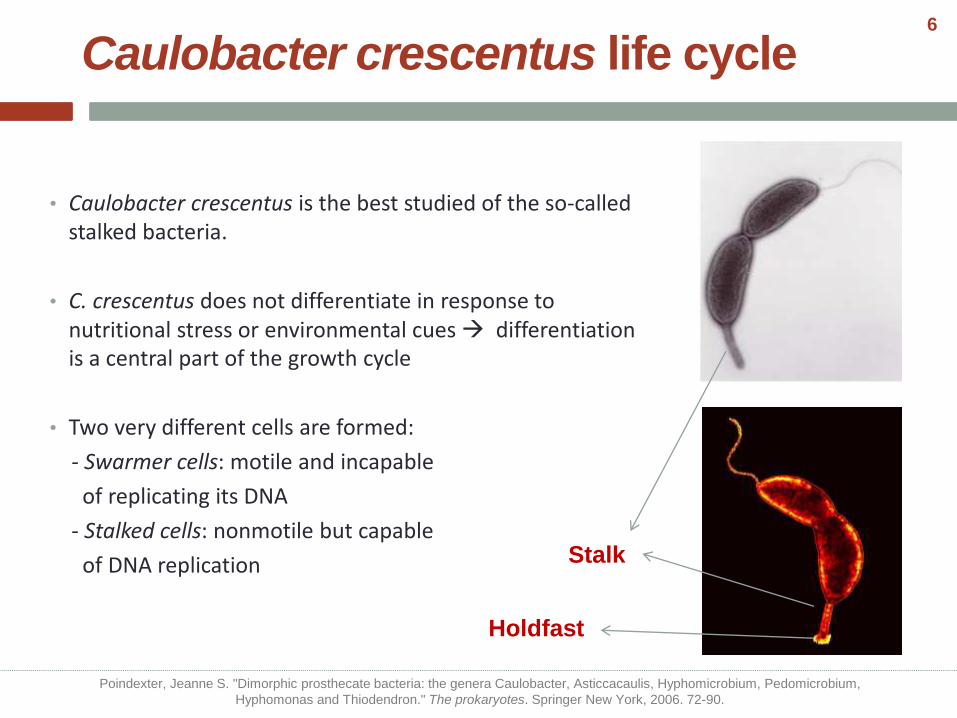
Caulobacter crescentus life cycle
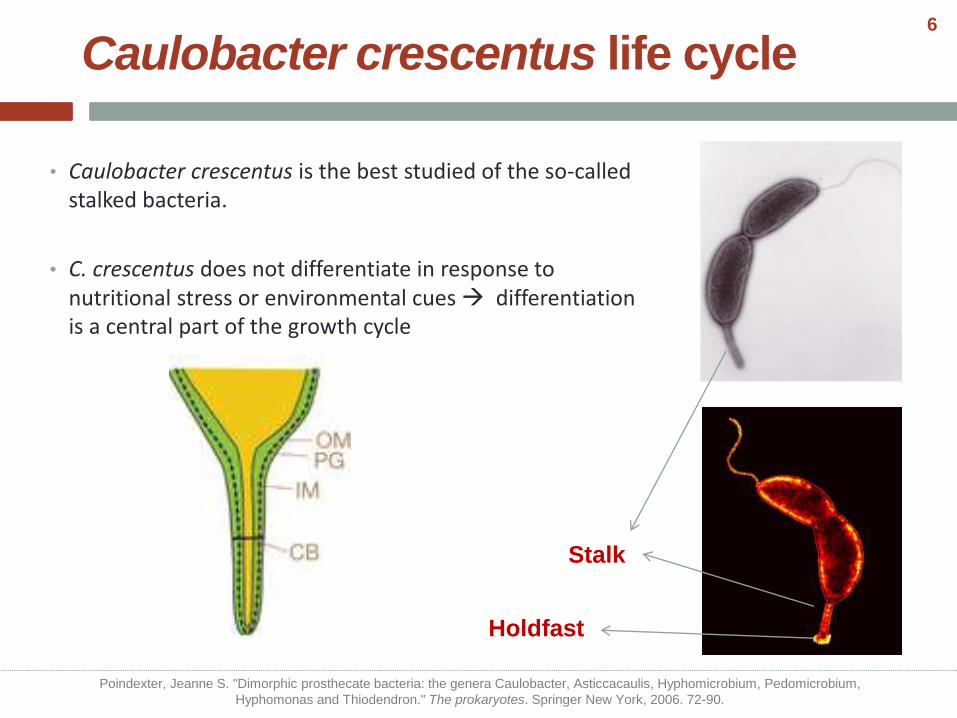
• Caulobacter crescentus is the best studied of the so-called stalked bacteria.
• C. crescentus does not differentiate in response to nutritional stress or environmental cues differentiation is a central part of the growth cycle
• Two very different cells are formed:
- Swarmer cells: motile and incapable
of replicating its DNA
- Stalked cells: nonmotile but capable
of DNA replication Stalk
Holdfast
Poindexter, Jeanne S. "Dimorphic prosthecate bacteria: the genera Caulobacter, Asticcacaulis, Hyphomicrobium, Pedomicrobium,
Hyphomonas and Thiodendron." The prokaryotes. Springer New York, 2006. 72-90.
6
Caulobacter crescentus life cycle
• Caulobacter crescentus is the best studied of the so-called stalked bacteria.
• C. crescentus does not differentiate in response to nutritional stress or environmental cues differentiation is a central part of the growth cycle
Stalk
Holdfast
Poindexter, Jeanne S. "Dimorphic prosthecate bacteria: the genera Caulobacter, Asticcacaulis, Hyphomicrobium, Pedomicrobium,
Hyphomonas and Thiodendron." The prokaryotes. Springer New York, 2006. 72-90.
6
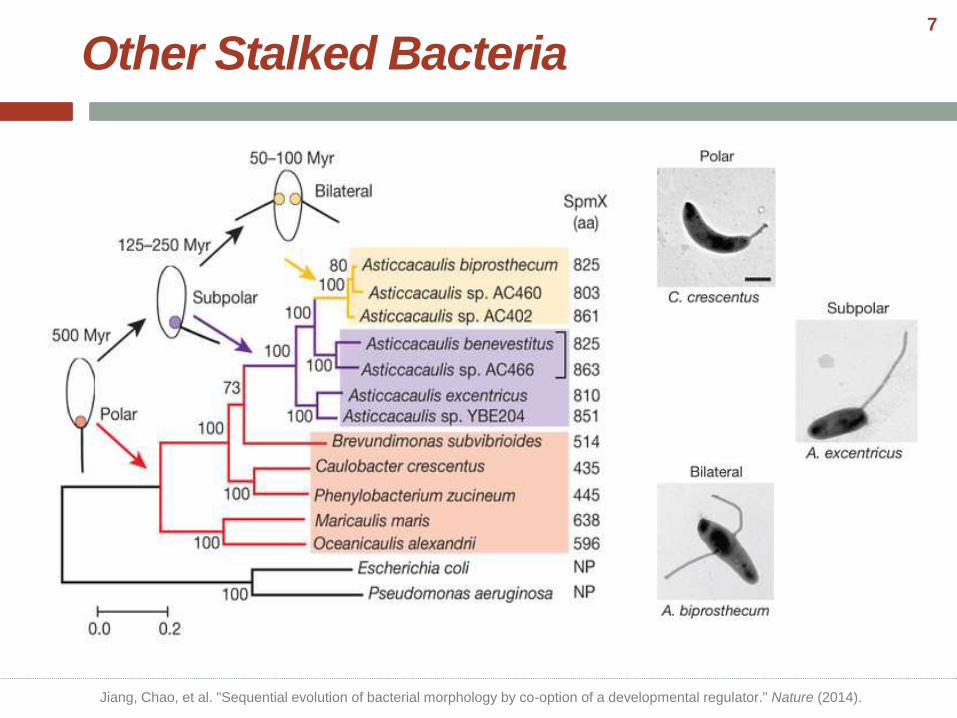
Other Stalked Bacteria
Jiang, Chao, et al. "Sequential evolution of bacterial morphology by co-option of a developmental regulator." Nature (2014).
7
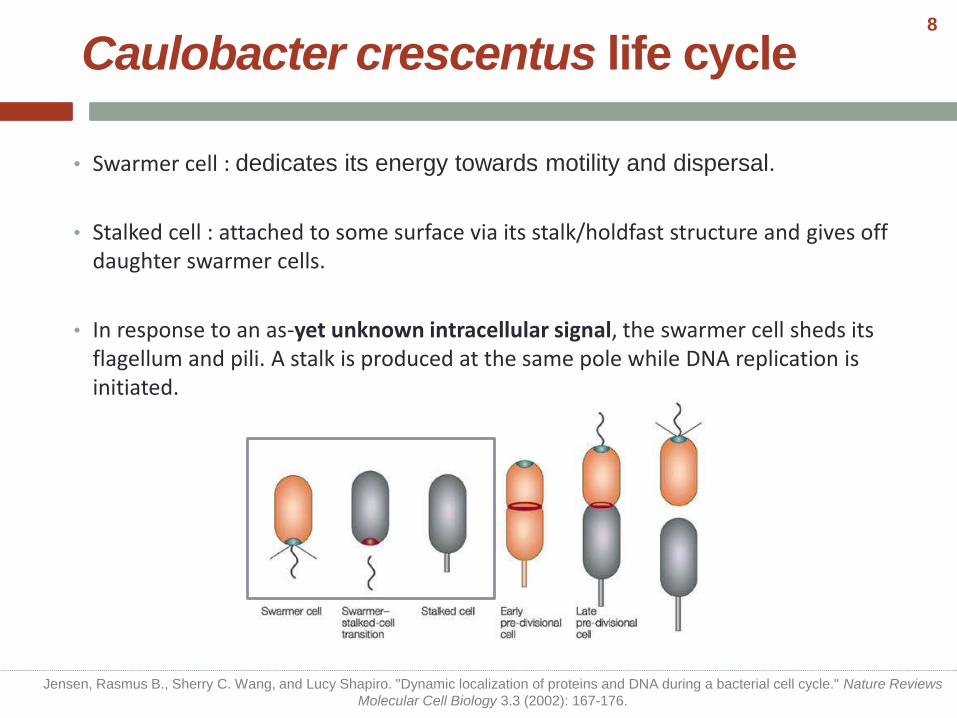
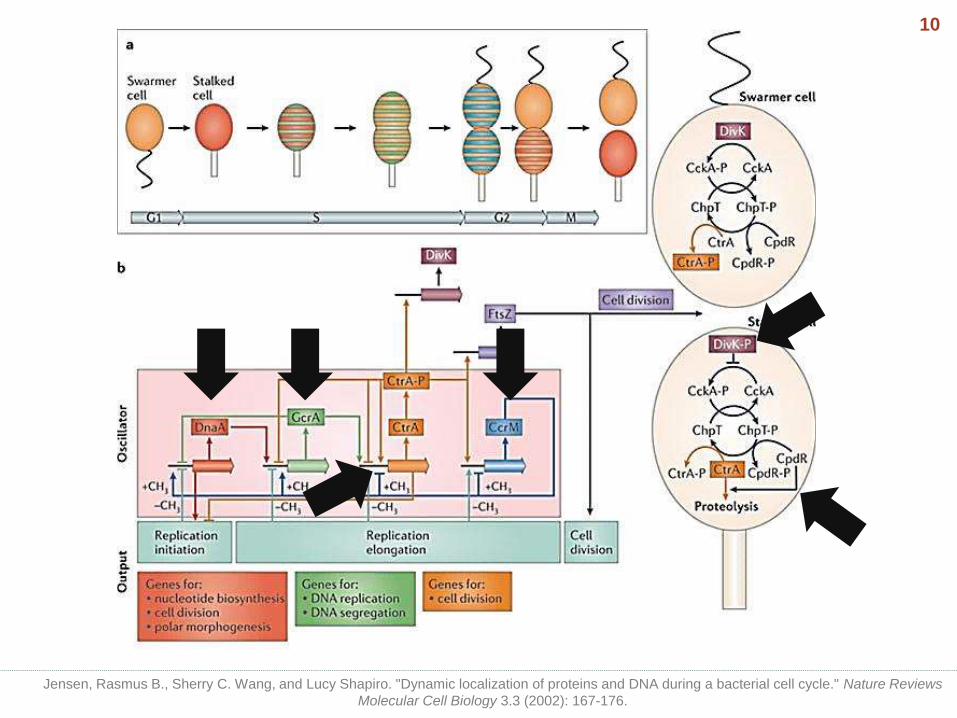
• Swarmer cell : dedicates its energy towards motility and dispersal.
• Stalked cell : attached to some surface via its stalk/holdfast structure and gives off daughter swarmer cells.
• In response to an as-yet unknown intracellular signal, the swarmer cell sheds its flagellum and pili. A stalk is produced at the same pole while DNA replication is initiated.
Caulobacter crescentus life cycle
Jensen, Rasmus B., Sherry C. Wang, and Lucy Shapiro. "Dynamic localization of proteins and DNA during a bacterial cell cycle." Nature Reviews
Molecular Cell Biology 3.3 (2002): 167-176.
8
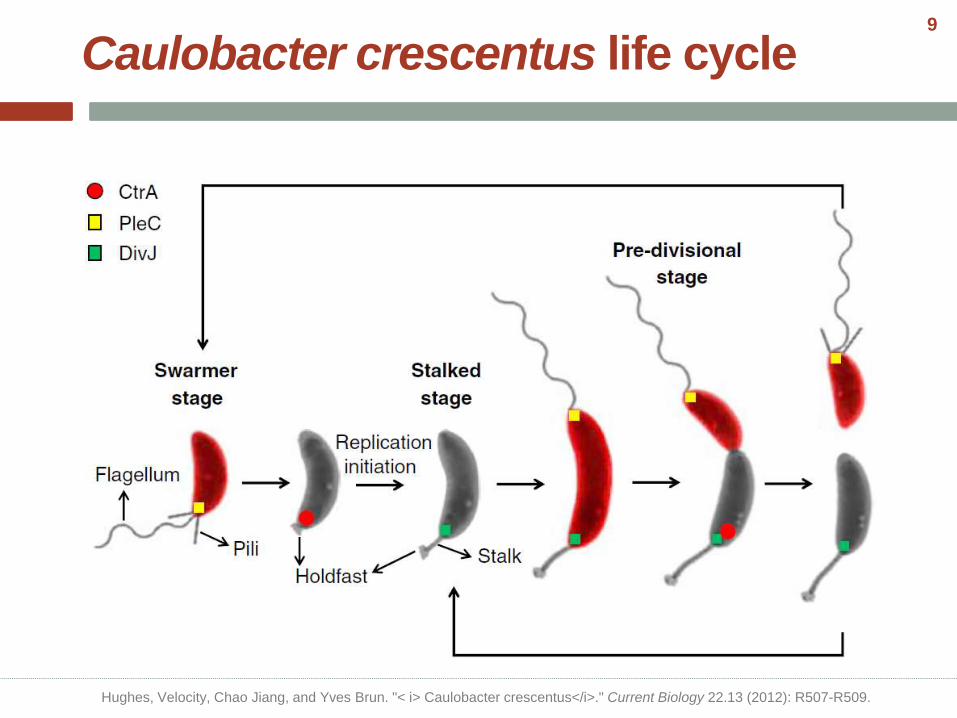
Caulobacter crescentus life cycle
Hughes, Velocity, Chao Jiang, and Yves Brun. "< i> Caulobacter crescentus</i>." Current Biology 22.13 (2012): R507-R509.
9
Jensen, Rasmus B., Sherry C. Wang, and Lucy Shapiro. "Dynamic localization of proteins and DNA during a bacterial cell cycle." Nature Reviews
Molecular Cell Biology 3.3 (2002): 167-176.
10
az http://2014.igem.org/Team:USTC-China/project/cimager
11

Endospore formation in
Bacillus subtilis

Endospore formation
• It is an extreme survival strategy employed by certain low G+C Gram-positive bacteria.
• Spores are resistant to heat, cold, radiation, and other adverse environmental conditions.
• The primary function of endospore formation appears to be the survival and dissemination of the species.
• When the environment becomes more favorable, the endospore can reactivate itself to the vegetative state.
• Time and effort has been expended in the study of this process in the genus Bacillus, particularly Bacillus subtilis.
Higgins, Douglas, and Jonathan Dworkin. "Recent progress in Bacillus subtilis sporulation." FEMS microbiology reviews 36.1
(2012): 131-148.
13
Endospore Formation
Desulfoto-maculum
Thermo-actinomycetes
Sporosarcina
Clostridium
Sporolacto-bacillus
Bacillus
Endospore formation14
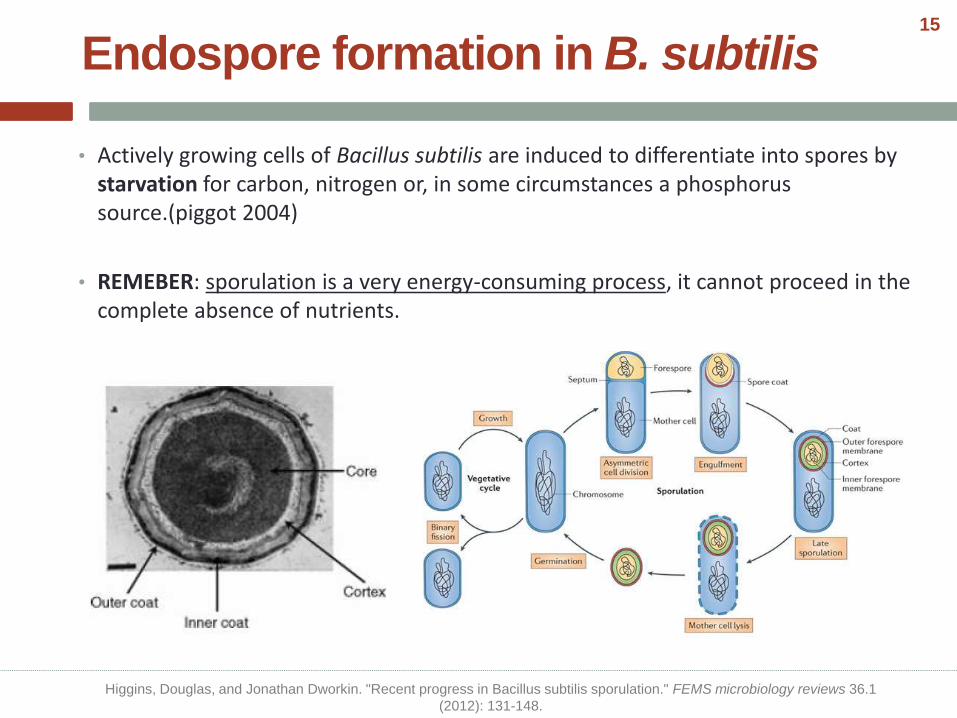
• Actively growing cells of Bacillus subtilis are induced to differentiate into spores by starvation for carbon, nitrogen or, in some circumstances a phosphorus source.(piggot 2004)
• REMEBER: sporulation is a very energy-consuming process, it cannot proceed in the complete absence of nutrients.
Endospore formation in B. subtilis
Higgins, Douglas, and Jonathan Dworkin. "Recent progress in Bacillus subtilis sporulation." FEMS microbiology reviews 36.1
(2012): 131-148.
15
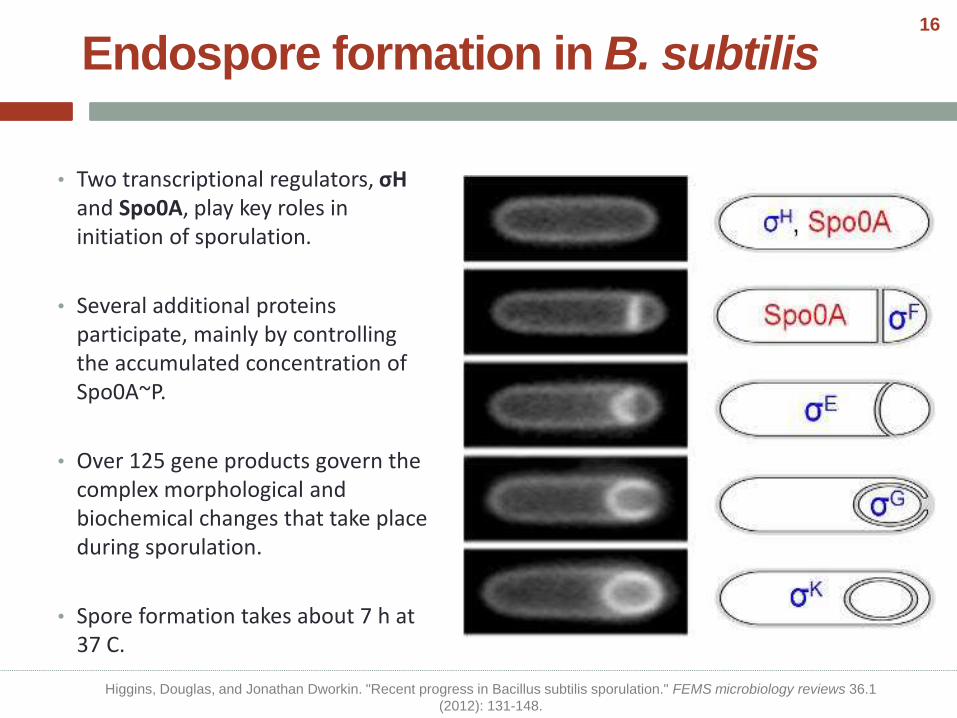
• Two transcriptional regulators, σHand Spo0A, play key roles in initiation of sporulation.
• Several additional proteins participate, mainly by controlling the accumulated concentration of Spo0A~P.
• Over 125 gene products govern the complex morphological and biochemical changes that take place during sporulation.
• Spore formation takes about 7 h at 37 C.
Endospore formation in B. subtilis
Higgins, Douglas, and Jonathan Dworkin. "Recent progress in Bacillus subtilis sporulation." FEMS microbiology reviews 36.1
(2012): 131-148.
16
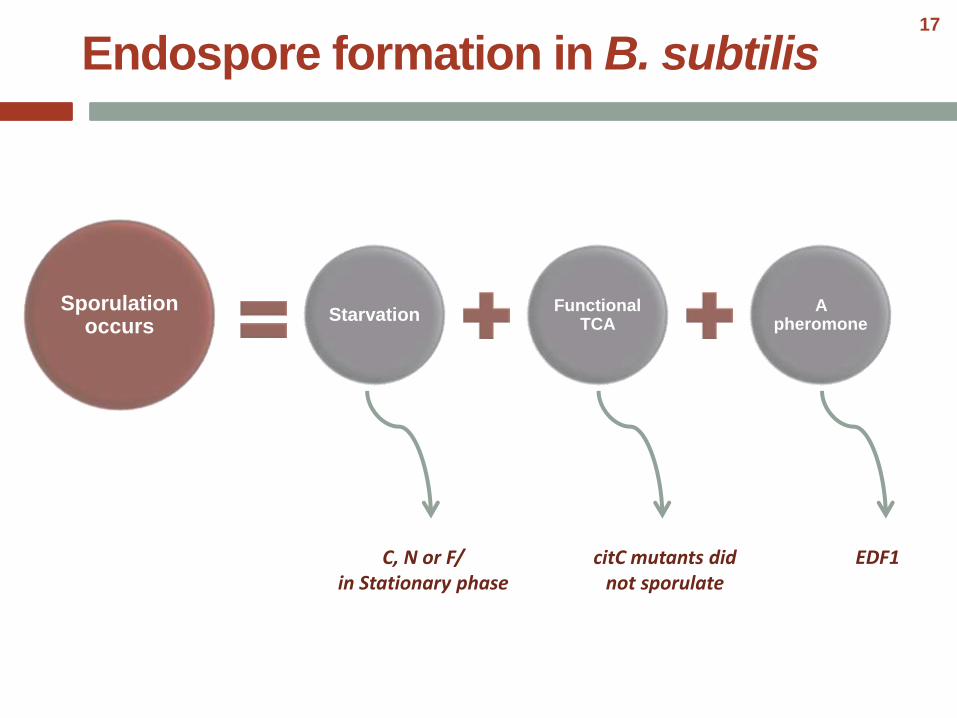
Endospore formation in B. subtilis
StarvationFunctional
TCAA
pheromoneSporulation
occurs
C, N or F/in Stationary phase
citC mutants did not sporulate
EDF1
17
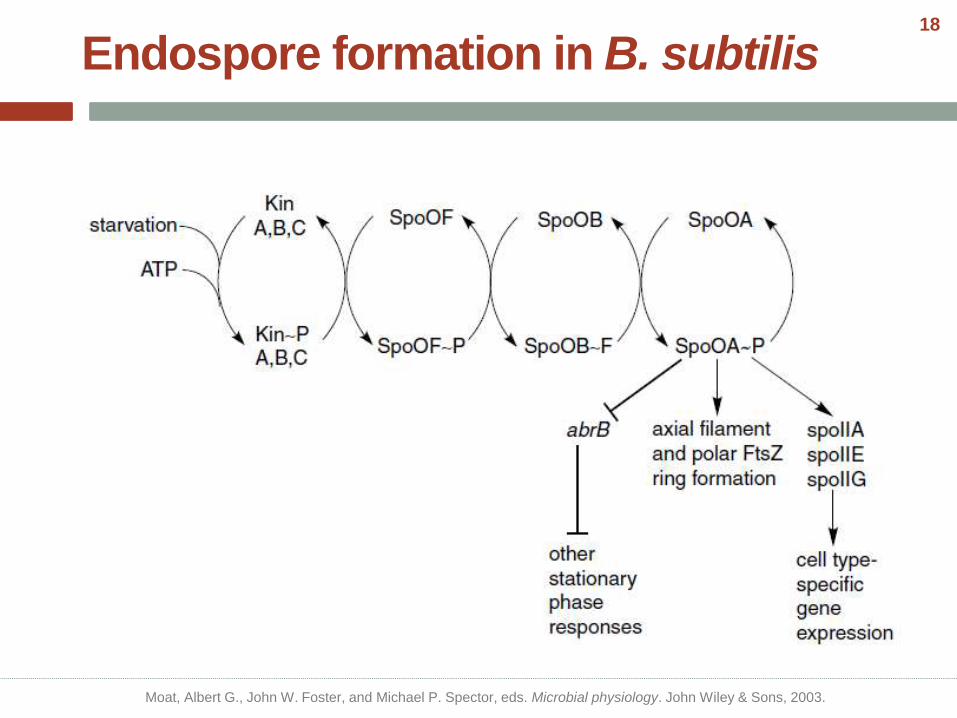
Endospore formation in B. subtilis
Moat, Albert G., John W. Foster, and Michael P. Spector, eds. Microbial physiology. John Wiley & Sons, 2003.
18
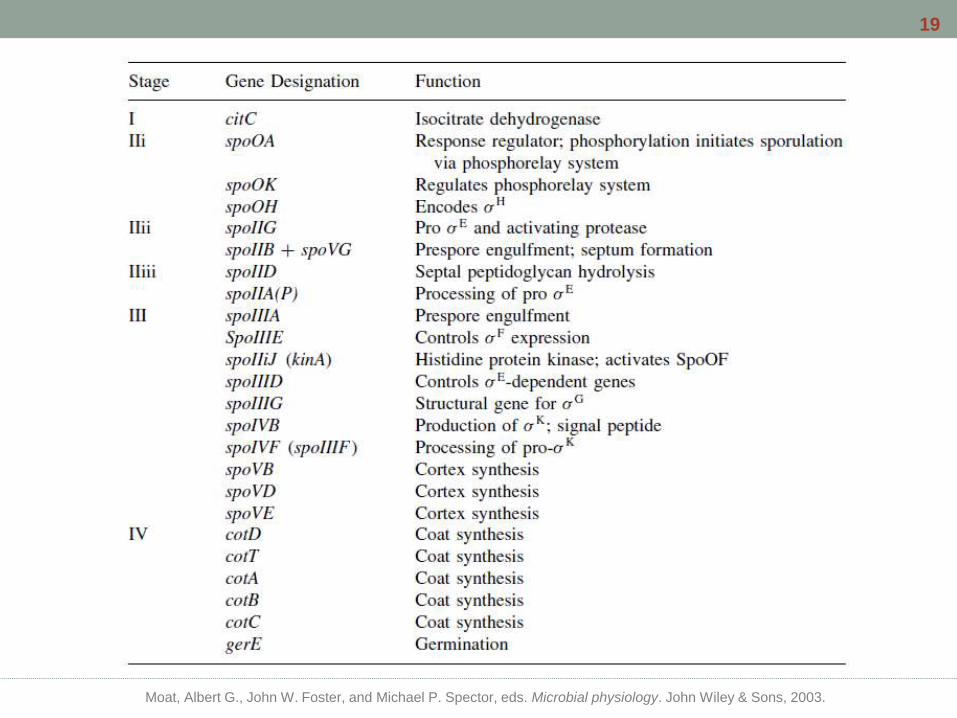
Moat, Albert G., John W. Foster, and Michael P. Spector, eds. Microbial physiology. John Wiley & Sons, 2003.
19
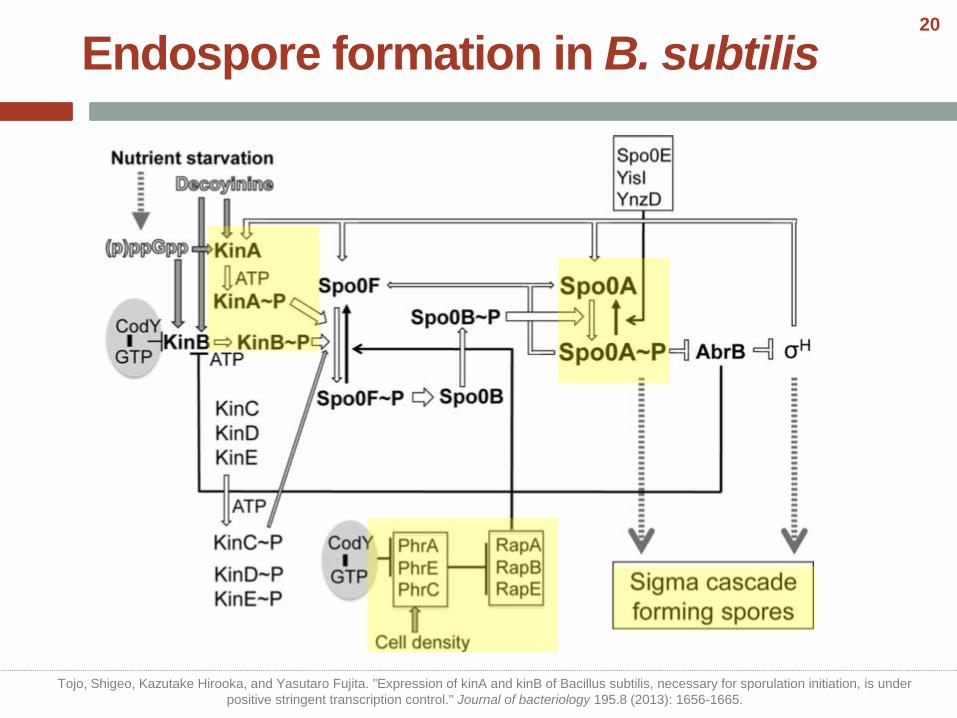
Endospore formation in B. subtilis
Tojo, Shigeo, Kazutake Hirooka, and Yasutaro Fujita. "Expression of kinA and kinB of Bacillus subtilis, necessary for sporulation initiation, is under
positive stringent transcription control." Journal of bacteriology 195.8 (2013): 1656-1665.
20

Spores of Actinomycetes
(S. coelicolor)

Actinomycete Spores
• Under nutrient poor conditions filaments differentiate into spores.
• In contrast to endospores, these structures are part of the reproductive process.
• The developmental process to create an actinomycete spore is less complex than that of the endospore. It involves the simple formation of cross walls that divide the filament into sections, each containing a chromosome. These then differentiate into mature spores.
• During this process a tougher cell wall is laid down and there is conversion of the cytoplasm to a dormant state so that the spore becomes more resistant to heat and chemicals, though not as hardy as an endospore.
• Actinomycete spores are capable of surviving for long periods of time (for years) and can germinate into vegetative cells when appropriate growth conditions are present.
Angert, Esther R. "Alternatives to binary fission in bacteria." Nature Reviews Microbiology 3.3 (2005): 214-224.
22
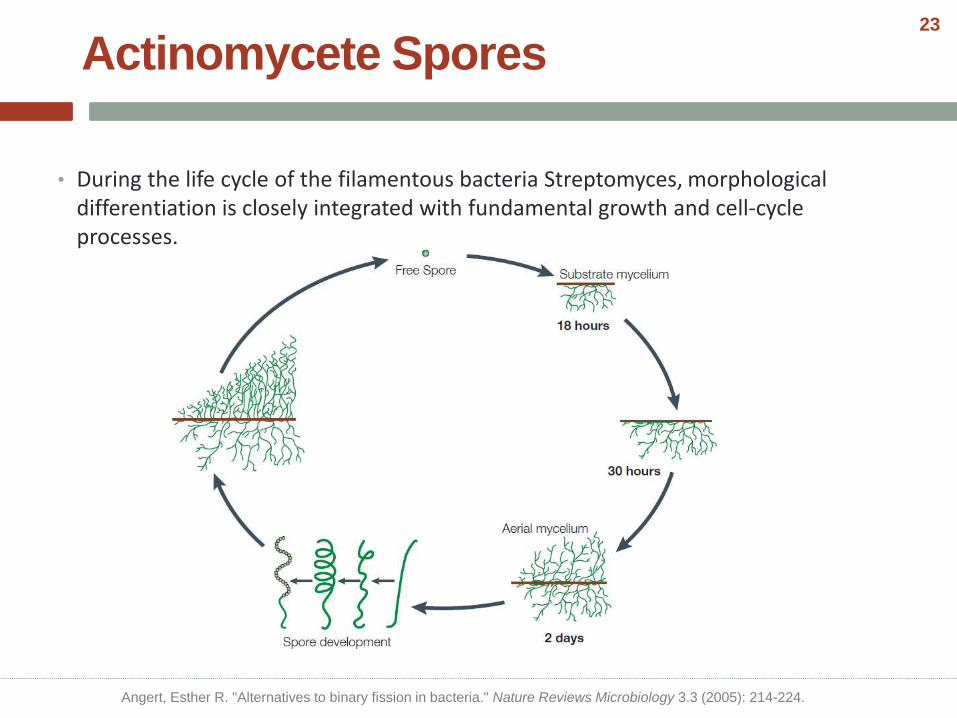
Actinomycete Spores
• During the life cycle of the filamentous bacteria Streptomyces, morphological differentiation is closely integrated with fundamental growth and cell-cycle processes.
Angert, Esther R. "Alternatives to binary fission in bacteria." Nature Reviews Microbiology 3.3 (2005): 214-224.
23
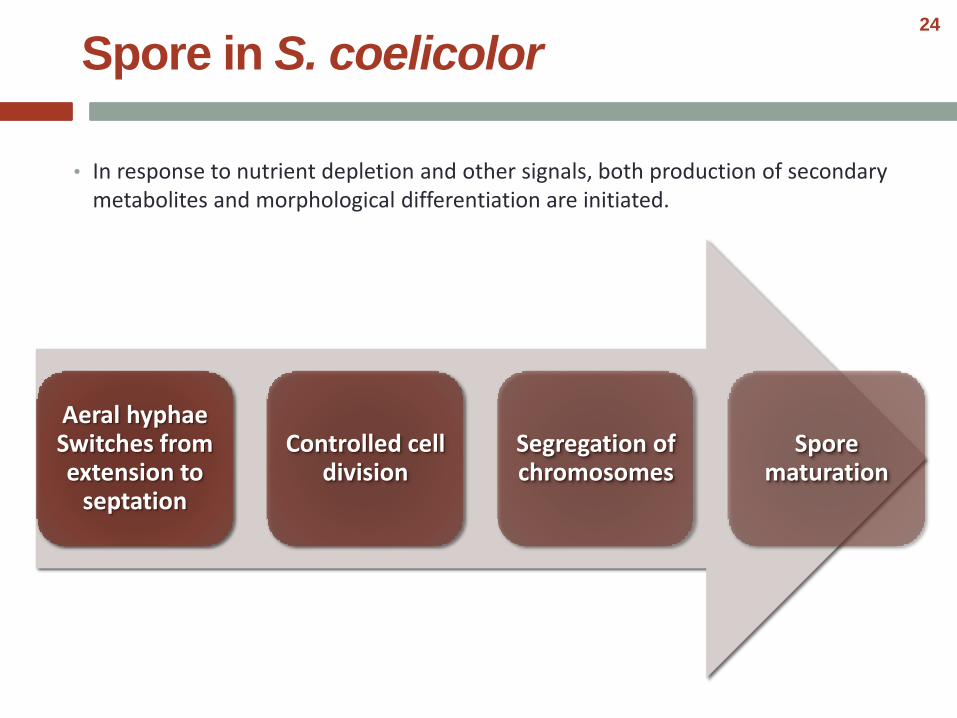
• In response to nutrient depletion and other signals, both production of secondary metabolites and morphological differentiation are initiated.
Spore in S. coelicolor
Aeral hyphae Switches from extension to
septation
Controlled cell division
Segregation of chromosomes
Spore maturation
24

Spore in S. coelicolor
Switch from extension to
septation
Flärdh, Klas, and Mark J. Buttner. "Streptomyces morphogenetics: dissecting differentiation in a filamentous bacterium." Nature Reviews Microbiology 7.1 (2009): 36-49.
25

Spore in S. coelicolor
Switch from extension to
septation
Flärdh, Klas, and Mark J. Buttner. "Streptomyces morphogenetics: dissecting differentiation in a filamentous bacterium." Nature Reviews Microbiology 7.1 (2009): 36-49.
25
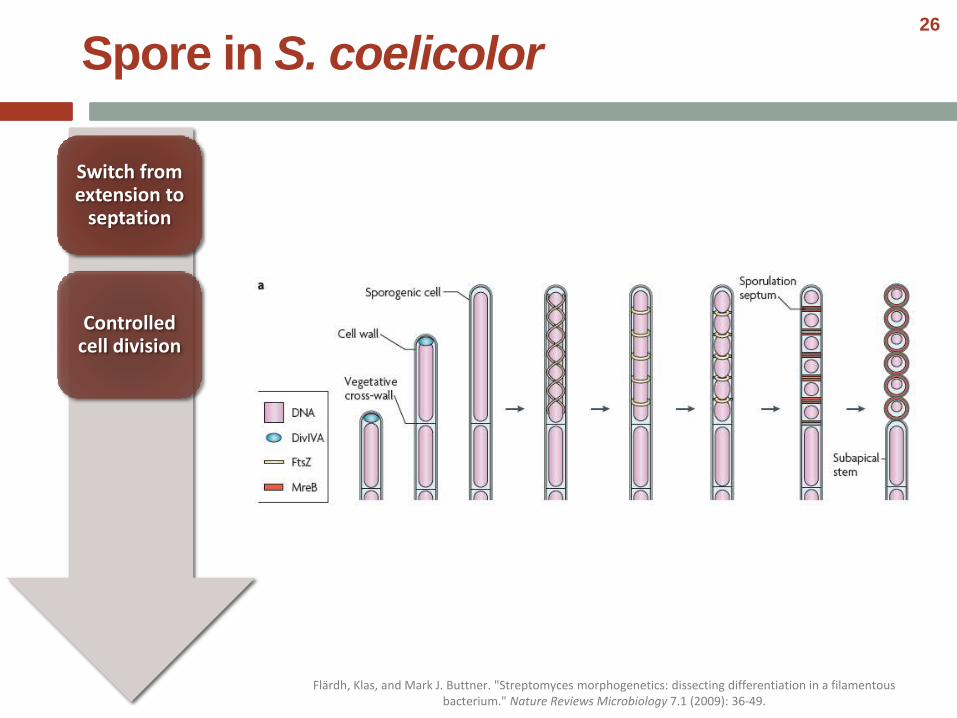
Controlled cell division
Switch from extension to
septation
Spore in S. coelicolor
Flärdh, Klas, and Mark J. Buttner. "Streptomyces morphogenetics: dissecting differentiation in a filamentous bacterium." Nature Reviews Microbiology 7.1 (2009): 36-49.
26
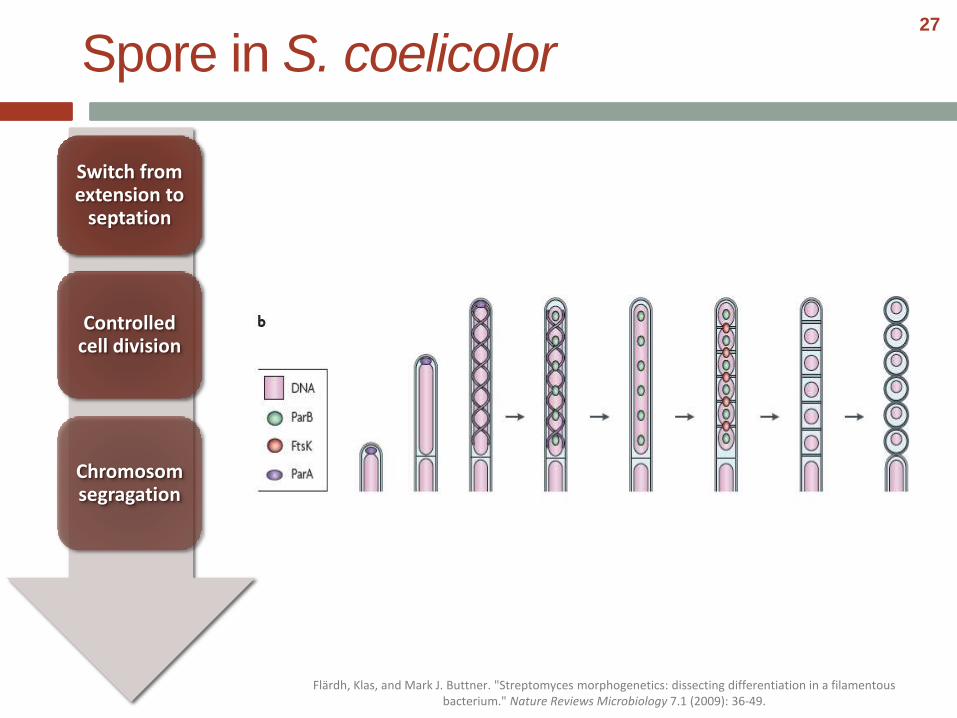
Controlled cell division
Switch from extension to
septation
Chromosomsegragation
Spore in S. coelicolor
Flärdh, Klas, and Mark J. Buttner. "Streptomyces morphogenetics: dissecting differentiation in a filamentous bacterium." Nature Reviews Microbiology 7.1 (2009): 36-49.
27

• Spore maturation involves the production of a thick, lysozyme-resistant spore wall.
• This wall is laid down after sporulation septation is complete.
• The correct assembly of the spore wall depends on mreB, which may explain why Streptomyces and a few other sporulatingactinomycete genera contain genes for this ancestral actin, whereas most other actinobacteria do not.
• The subcellular localization of MreB–EGFP is consistent with a role for MreB in assembly of the spore wall.
• Correct is: first as bands that coincide with sporulation septa at both poles after septal constriction, and then spreading out to surround the spore completely, lining the inner surface of the membrane before disappearing in the mature spores.
Controlled cell division
Switch from extension to
septation
Chromosomsegragation
Spore maturation
Spore in S. coelicolor
Flärdh, Klas, and Mark J. Buttner. "Streptomyces morphogenetics: dissecting differentiation in a filamentous bacterium." Nature Reviews Microbiology 7.1 (2009): 36-49.
28
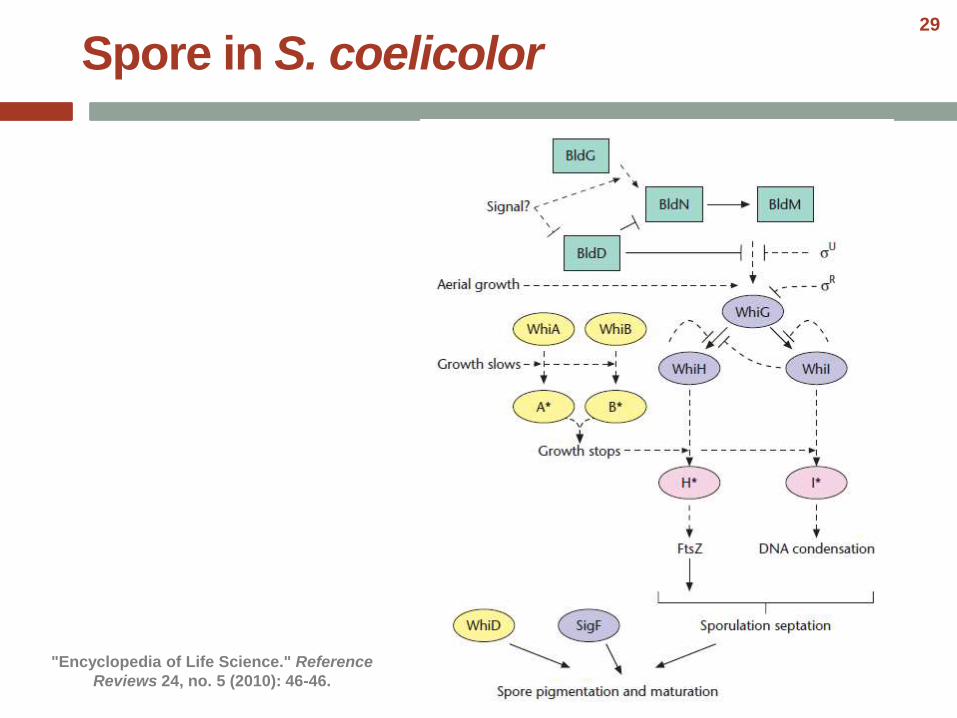
Spore in S. coelicolor
"Encyclopedia of Life Science." Reference
Reviews 24, no. 5 (2010): 46-46.
29

Cysts in Azotobacter
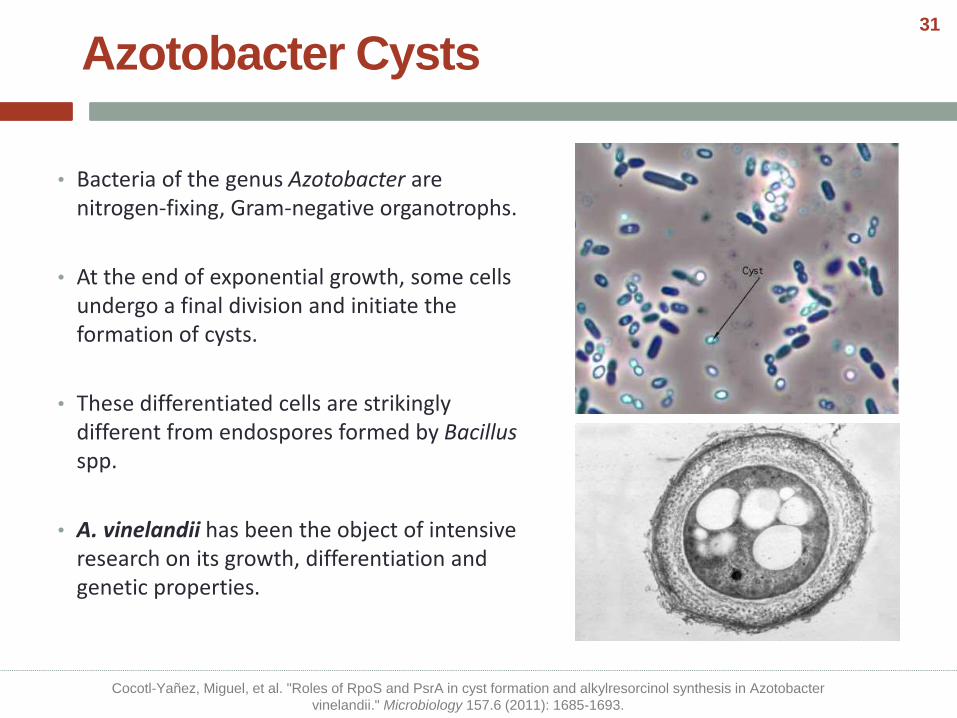
Azotobacter Cysts
• Bacteria of the genus Azotobacter are nitrogen-fixing, Gram-negative organotrophs.
• At the end of exponential growth, some cells undergo a final division and initiate the formation of cysts.
• These differentiated cells are strikingly different from endospores formed by Bacillusspp.
• A. vinelandii has been the object of intensive research on its growth, differentiation and genetic properties.
Cocotl-Yañez, Miguel, et al. "Roles of RpoS and PsrA in cyst formation and alkylresorcinol synthesis in Azotobacter
vinelandii." Microbiology 157.6 (2011): 1685-1693.
31

• When A. vinelandii experiences a lack of nutrients, it will develop into cyst . The cell is then protected against desiccation by a multilayered coat, of which gel-forming alginate is a necessary part.(j. bacteril 2009)
• Depending on the carbon source and the conditions under which growth occurs, poly-b-hydroxybutyrate (PHB) accumulates in vegetative cells, and the extent of encystment in such cultures is related to the intracellular concentration of that polymer.
• The problem studying encystment genetics is the inability to generate useful mutants of the organism.
• Azotobacter cells are refractory to mutagenesis except for those genes involved in nitrogen fixation and resistance to certain antibiotics. The basis for this difficulty may lie in the large amount of deoxyribonucleic acid (DNA) (6.610214 g per cell) that these cells possess.
Azotobacter Cysts
Gimmestad, Martin, et al. "Characterization of three new Azotobacter vinelandii alginate lyases, one of which is involved in cyst
germination." Journal of bacteriology 191.15 (2009): 4845-4853.
32
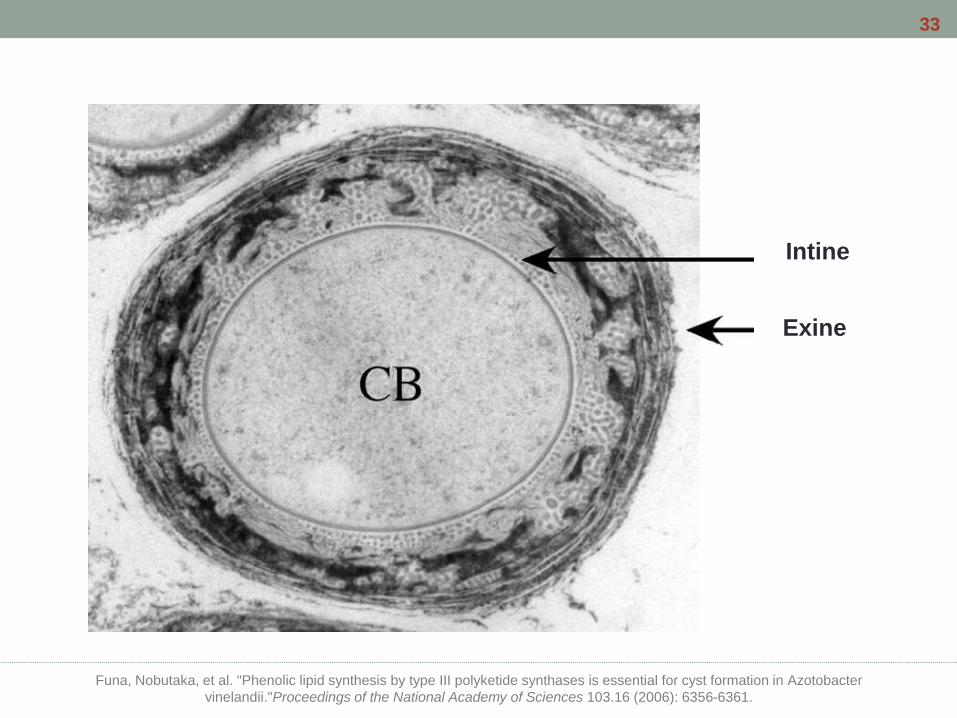
Exine
Intine
Funa, Nobutaka, et al. "Phenolic lipid synthesis by type III polyketide synthases is essential for cyst formation in Azotobacter
vinelandii."Proceedings of the National Academy of Sciences 103.16 (2006): 6356-6361.
33
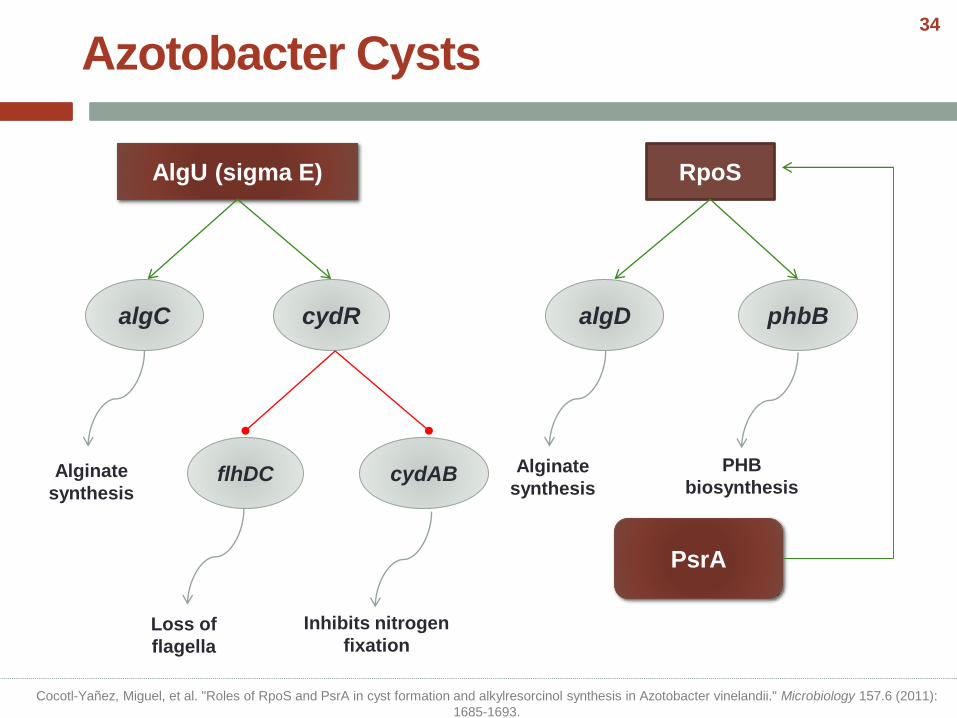
AlgU (sigma E)
algC cydR
RpoS
algD phbB
PsrA
Azotobacter Cysts
Alginate
synthesis
PHB
biosynthesisflhDC cydAB
Loss of
flagella
Inhibits nitrogen
fixation
Alginate
synthesis
Cocotl-Yañez, Miguel, et al. "Roles of RpoS and PsrA in cyst formation and alkylresorcinol synthesis in Azotobacter vinelandii." Microbiology 157.6 (2011):
1685-1693.
34

Heterocyst in cyanobacteria

Heterocyst in cyanobacteria
• Many cyanobacterial species are capable of nitrogen fixation. However, oxygenic photosynthesis and nitrogen fixation are incompatible processes because nitrogenase is inactivated by oxygen.
Mechanisms to separate these
activities
TemporalSpatialy
(differentiation)
Kumar, Krithika, Rodrigo A. Mella-Herrera, and James W. Golden. "Cyanobacterial heterocysts." Cold Spring Harbor perspectives in
biology 2.4 (2010): a000315.
36
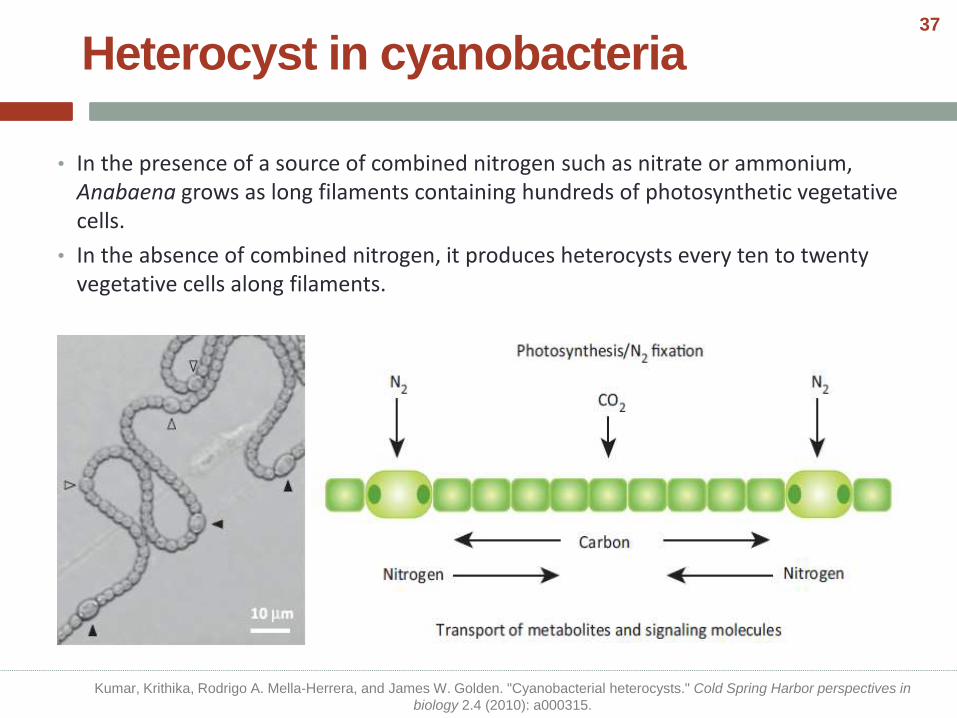
• In the presence of a source of combined nitrogen such as nitrate or ammonium, Anabaena grows as long filaments containing hundreds of photosynthetic vegetative cells.
• In the absence of combined nitrogen, it produces heterocysts every ten to twenty vegetative cells along filaments.
Heterocyst in cyanobacteria
Kumar, Krithika, Rodrigo A. Mella-Herrera, and James W. Golden. "Cyanobacterial heterocysts." Cold Spring Harbor perspectives in
biology 2.4 (2010): a000315.
37

• Oxygen-producing photosystem PSII is dismantled during differentiation and heterocysts show an increased rate of respiration.
• Morphological changes include the deposition of two additional envelope layers around the heterocyst: an inner “laminated” layer composed of two heterocyst specific glycolipids (HGL)and an outer polysaccharide layer (HEP).
• Heterocysts and vegetative cells are mutually interdependent. Because they lack photosystem II and carbon fixation, heterocysts are dependent on vegetative cells for a source of reductant and carbon, which is probably partially supplied as sucrose.
• In Anabaena PCC 7120, vegetative cells must also supply glutamate to heterocysts, which convert it to glutamine and other amino acids.
• In return, newly fixed nitrogen is rapidly exported from heterocysts and distributed to nearby vegetative cells.
Heterocyst in Anabaena
Kumar, Krithika, Rodrigo A. Mella-Herrera, and James W. Golden. "Cyanobacterial heterocysts." Cold Spring Harbor perspectives in
biology 2.4 (2010): a000315.
38

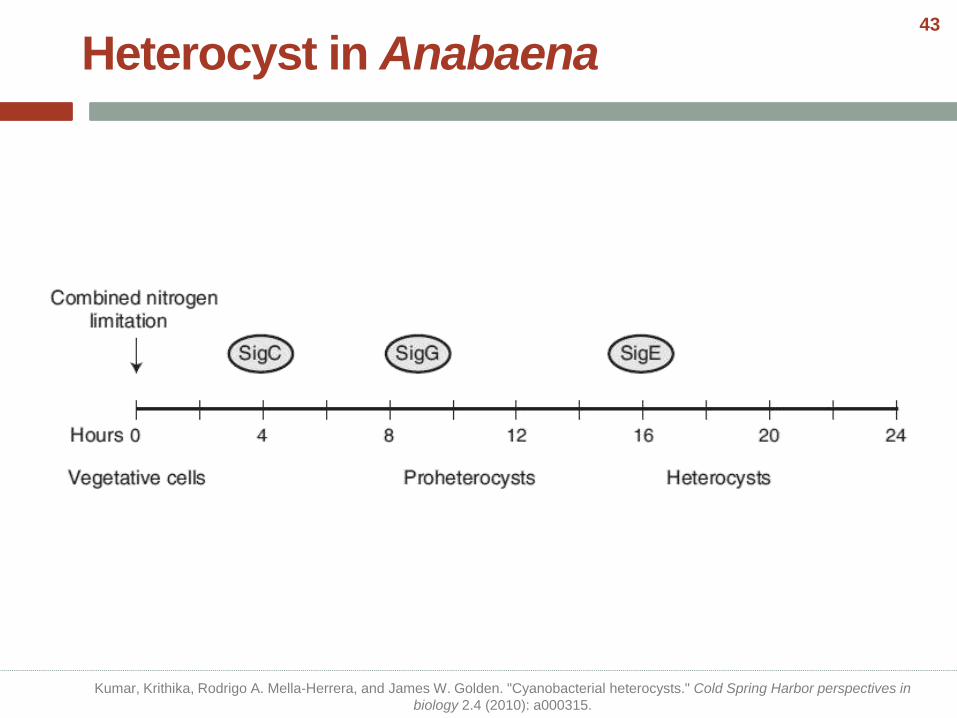
• The timeline of heterocyst development begins with sensing combined-nitrogen limitation and culminates with nitrogen fixation in the mature heterocyst.
• Heterocyst development is complete in about 20 hours at 30 ◦C reversibly when the combined nitrogen source is available.
Heterocyst in Anabaena
Kumar, Krithika, Rodrigo A. Mella-Herrera, and James W. Golden. "Cyanobacterial heterocysts." Cold Spring Harbor perspectives in
biology 2.4 (2010): a000315.
Picture from: Cumino, Andrea C., et al. "Carbon cycling in Anabaena sp. PCC 7120. Sucrose synthesis in the heterocysts and possible
role in nitrogen fixation." Plant physiology 143.3 (2007): 1385-1397.
39
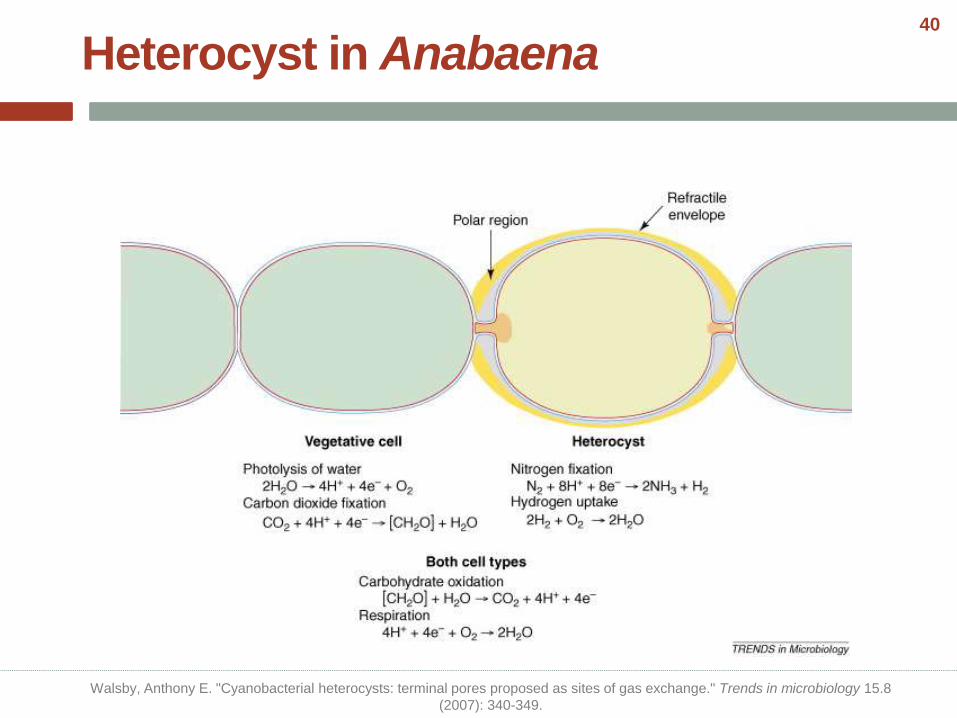
Heterocyst in Anabaena
Walsby, Anthony E. "Cyanobacterial heterocysts: terminal pores proposed as sites of gas exchange." Trends in microbiology 15.8
(2007): 340-349.
40
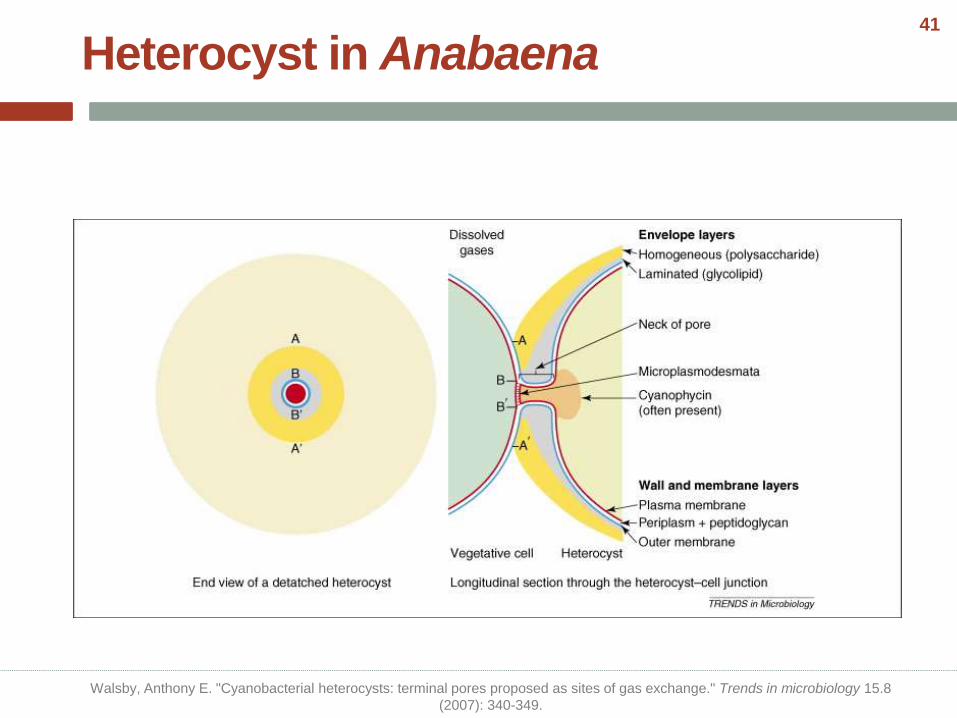
Heterocyst in Anabaena
Walsby, Anthony E. "Cyanobacterial heterocysts: terminal pores proposed as sites of gas exchange." Trends in microbiology 15.8
(2007): 340-349.
41

• In cyanobacteria, 2-oxoglutarate, an intermediate in the Krebs cycle, constitutes the signal for nitrogen deprivation.
• The Krebs cycle in cyanobacteria is incomplete because of the lack of 2-oxoglutarate dehydrogenase 2-oxoglutarate’s main function is to serve as a precursor in a variety of biosynthetic reactions. It is the primary carbon skeleton for incorporation of ammonium and is considered the metabolic junction between carbon and nitrogen balance in cyanobacteria.
• Nitrogen limiting conditions result in an increase in the levels of 2-oxoglutarate.
• NtcA, a transcriptional regulator belonging to the CRP (cyclic AMP receptor protein) family of proteins, senses 2-oxoglutarate levels.
• In Anabaena PCC 7120, NtcA is required for the expression of the genes in pathways for ammonium and nitrate assimilation, as well as heterocyst development.
Heterocyst in Anabaena
Kumar, Krithika, Rodrigo A. Mella-Herrera, and James W. Golden. "Cyanobacterial heterocysts." Cold Spring Harbor perspectives in
biology 2.4 (2010): a000315.
42
Heterocyst in Anabaena
Kumar, Krithika, Rodrigo A. Mella-Herrera, and James W. Golden. "Cyanobacterial heterocysts." Cold Spring Harbor perspectives in
biology 2.4 (2010): a000315.
43

Heterocyst in Anabaena
Kumar, Krithika, Rodrigo A. Mella-Herrera, and James W. Golden. "Cyanobacterial heterocysts." Cold Spring Harbor perspectives in
biology 2.4 (2010): a000315.
44
II. Multicellular Differentiations
1. Fruiting body in Myxobacteria
2. Hyper flagellated swarmer cells
3. Biofilm formation
45

Fruiting body in
Myxobacteria

Fruiting body in Myxobacteria
• Myxobacteria are a group of Gram-negative bacteria that are predominantly found in soil.
• During starvation, growth is arrested and a developmental program is initiated, which culminates in the formation of spore-bearing fruiting bodies that have a well-defined shape .
Claessen, Dennis, et al. "Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies." Nature Reviews
Microbiology 12.2 (2014): 115-124.
47

• The first signs of fruiting body formation are evident 4–6 hours after the exhaustion of resources.
• This is accompanied by changes in cell motility and the formation of aggregationcenters.
• Within 24 hours, the aggregation process is complete, and the nascent fruiting bodies each contain approximately 105 densely packed cells that differentiate into spores.
Fruiting body in Myxobacteria
Claessen, Dennis, et al. "Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies." Nature Reviews
Microbiology 12.2 (2014): 115-124.
48
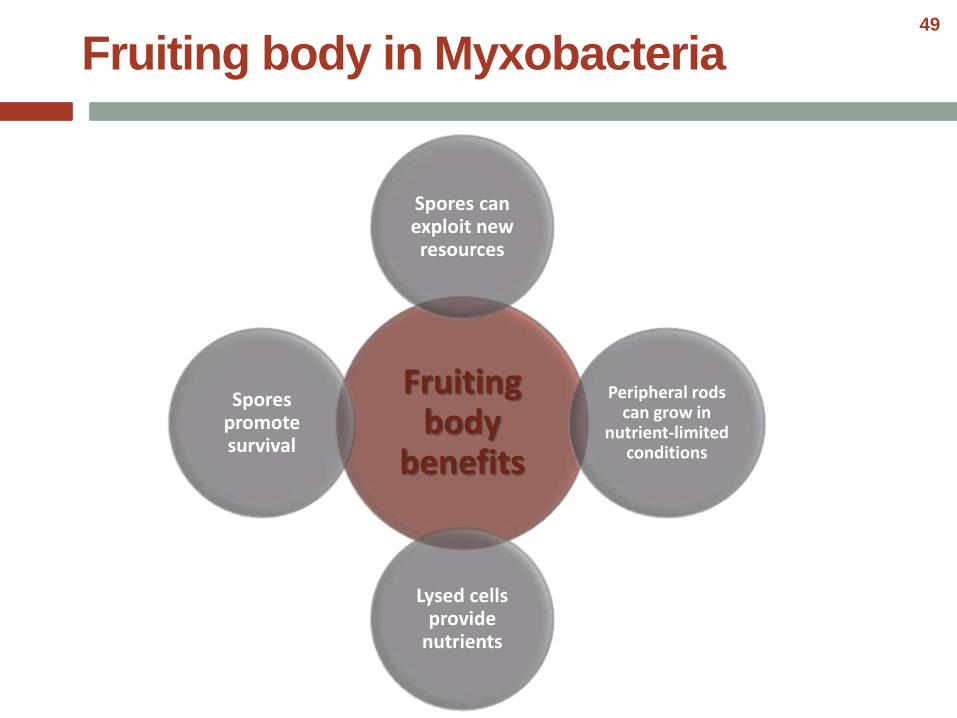
Fruiting body
benefits
Spores can exploit new resources
Peripheral rods can grow in
nutrient-limited conditions
Lysed cells provide
nutrients
Spores promote survival
Fruiting body in Myxobacteria49

Myxococcus xanthus life cycle
• Nutrition is depleted
• Cell density is high
• Solid support is available
Kaiser, Dale, Mark Robinson, and Lee Kroos. "Myxobacteria, polarity, and multicellular morphogenesis." Cold Spring Harbor perspectives in
biology 2.8 (2010): a000380.
50
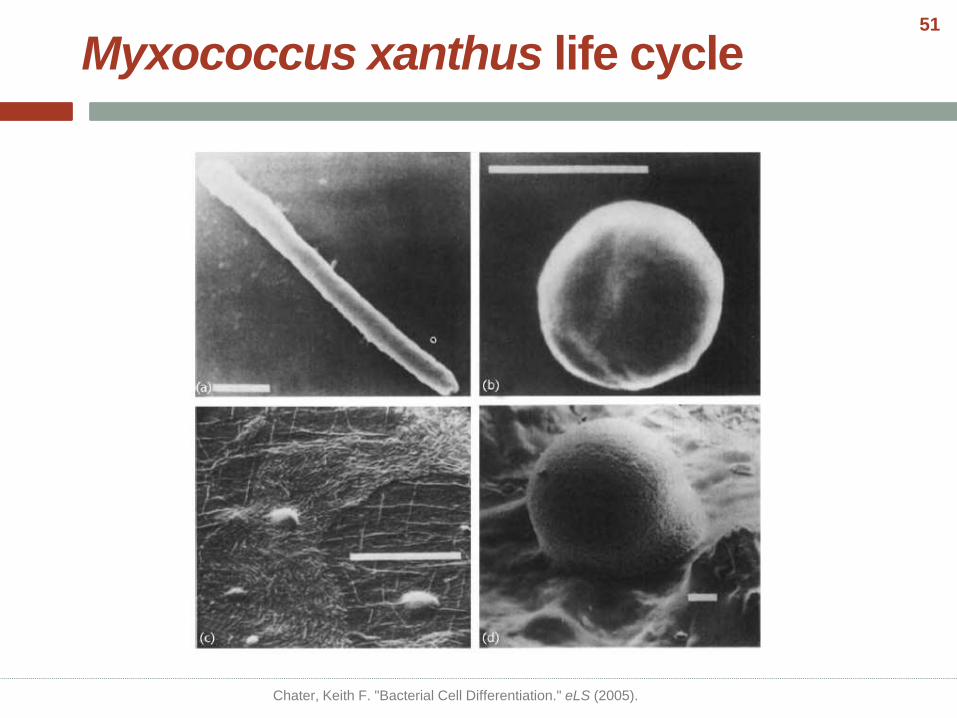
Myxococcus xanthus life cycle
Chater, Keith F. "Bacterial Cell Differentiation." eLS (2005).
51
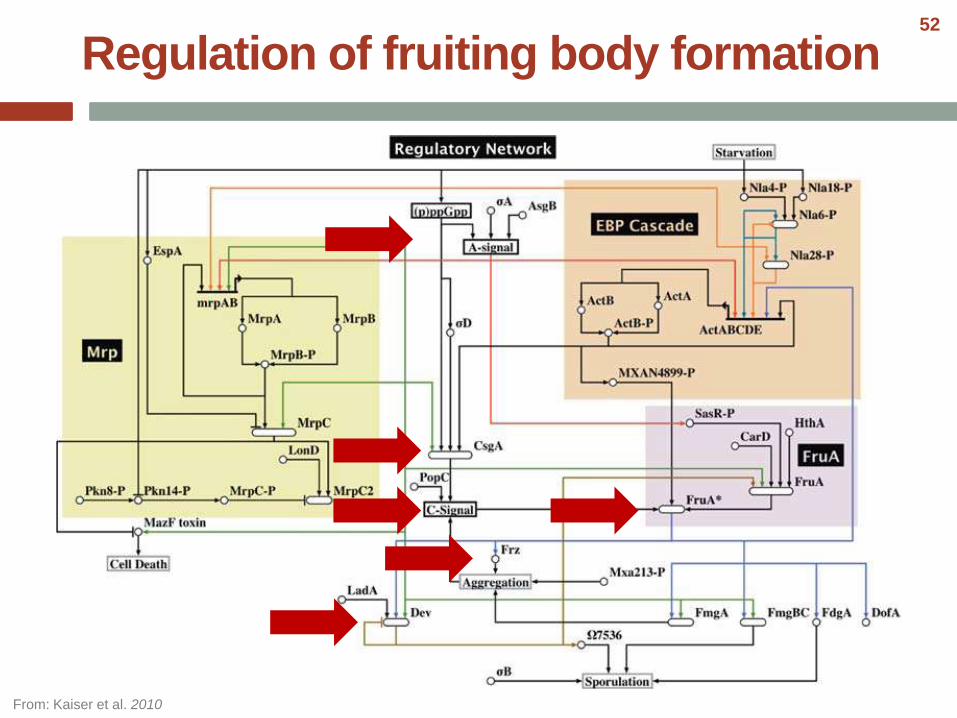
Regulation of fruiting body formation
From: Kaiser et al. 2010
52
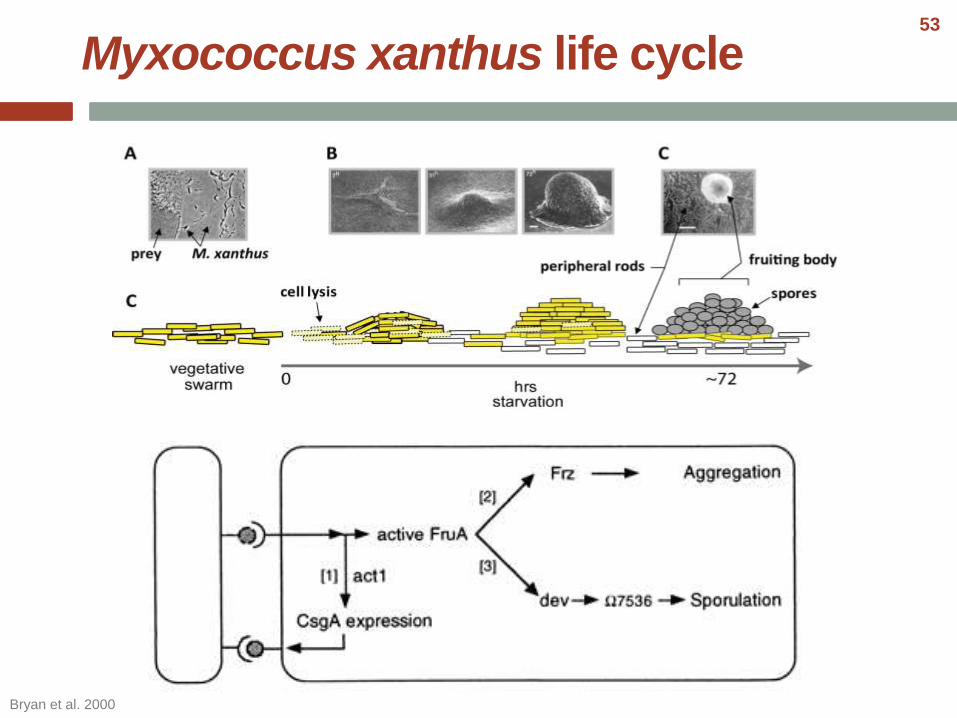
Myxococcus xanthus life cycle
Bryan et al. 2000
53

Hyper flagellated
Swarmer cells

Swarmer cell differentiation
• Swarming is the fastest known bacterial mode of surface translocation and enables the rapid colonization of a nutrient-rich environment and host tissues.
• It requires functional flagella and is coupled to the production of a viscous slime layer.
• It is widespread in many genera of Gram negative and Gram-positive flagellated bacteria and is typically assayed on a solidified medium, containing 0.5–2% agar.
• Species such as Proteus mirabilis and Vibrio parahaemolyticus, which are capable of vigorous swarming even on high-agar medium.
• Swarming has been studied extensively in P. mirabilis, in which elongated, multinucleated and hyper-flagellated swarmer cells can spread as multicellular rafts across surfaces.
Verstraeten, Natalie, et al. "Living on a surface: swarming and biofilm formation." Trends in microbiology 16.10 (2008): 496-506.
55
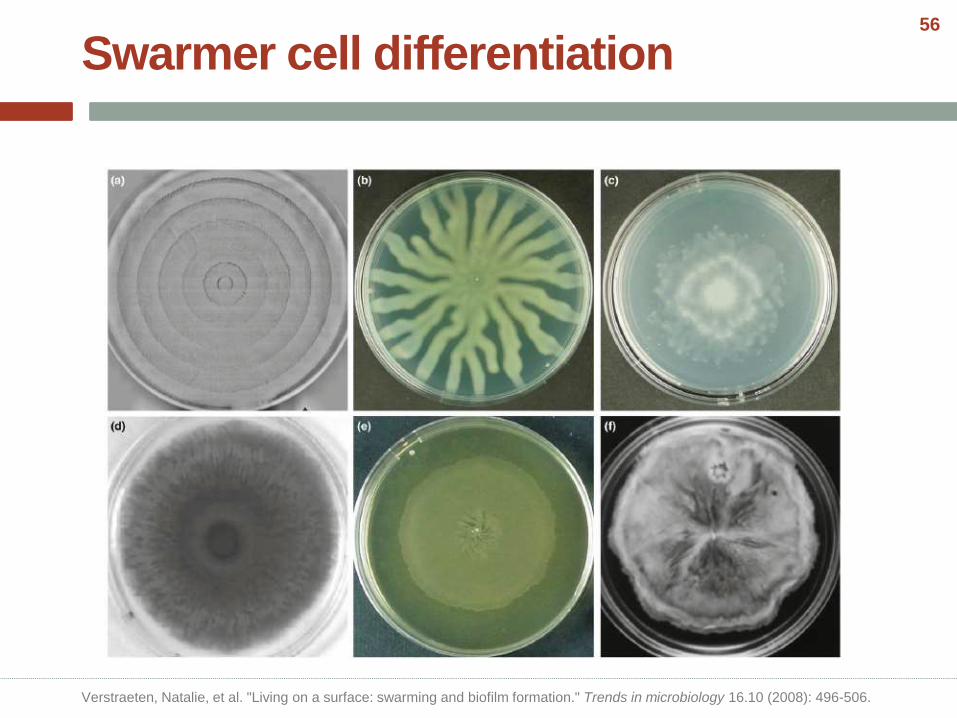
Swarmer cell differentiation
Verstraeten, Natalie, et al. "Living on a surface: swarming and biofilm formation." Trends in microbiology 16.10 (2008): 496-506.
56
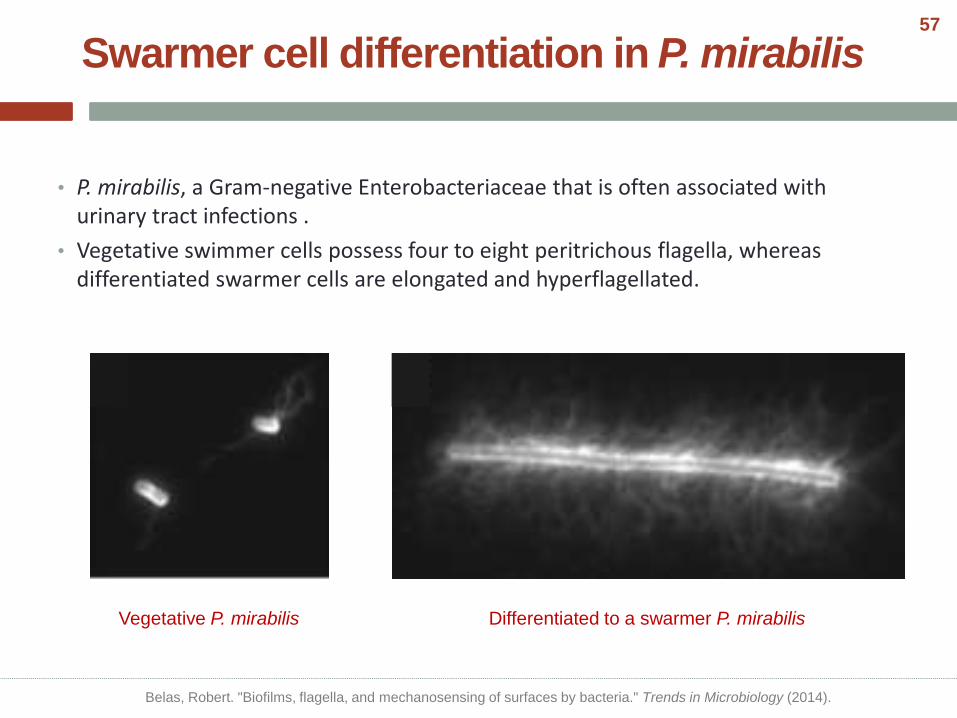
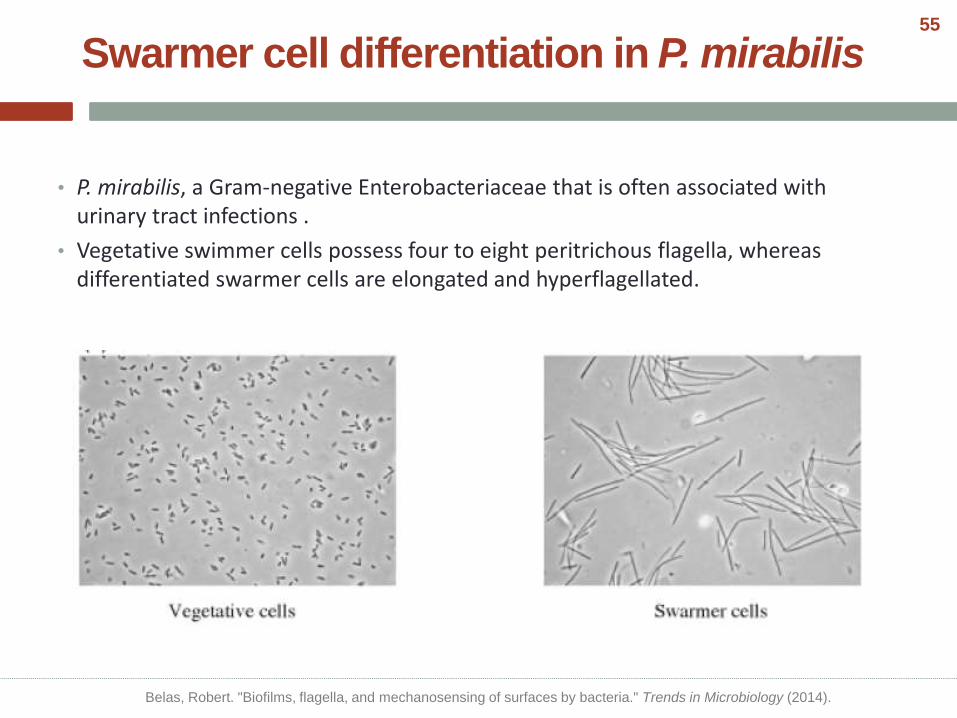
Swarmer cell differentiation in P. mirabilis
• P. mirabilis, a Gram-negative Enterobacteriaceae that is often associated with urinary tract infections .
• Vegetative swimmer cells possess four to eight peritrichous flagella, whereas differentiated swarmer cells are elongated and hyperflagellated.
Vegetative P. mirabilis Differentiated to a swarmer P. mirabilis
Belas, Robert. "Biofilms, flagella, and mechanosensing of surfaces by bacteria." Trends in Microbiology (2014).
57
Swarmer cell differentiation in P. mirabilis
• P. mirabilis, a Gram-negative Enterobacteriaceae that is often associated with urinary tract infections .
• Vegetative swimmer cells possess four to eight peritrichous flagella, whereas differentiated swarmer cells are elongated and hyperflagellated.
Belas, Robert. "Biofilms, flagella, and mechanosensing of surfaces by bacteria." Trends in Microbiology (2014).
55

• Although many species swarm including Aeromonas, Azospirillum, B. subtilis, E. coli, Rhodospirillum, Rhizobium, Salmonella, Serratia, and Yersinia, only a few (two notable examples are V. parahaemolyticus and P. mirabilis) do so after a surface-induced physiological differentiation that results in an elongated, highly flagellated swarmer cell.
• P. mirabilis swarmer cell differentiation is triggered by physical conditions that inhibit the rotation of the peritrichous flagella of the swimmer cell (belas 2014)
• P. mirabilis flagellum functions as a mechanosensor of the surface signal.
• Both lipopolysaccharide (LPS) and O-antigen play a part in P. mirabilis surface sensing.
• Nutrient availability is crucial to sustain the energy demanding process of swarming (verstraeten)
Swarmer cell differentiation in P. mirabilis
Morgenstein, Randy M., Bree Szostek, and Philip N. Rather. "Regulation of gene expression during swarmer cell differentiation in
Proteus mirabilis." FEMS microbiology reviews 34.5 (2010): 753-763.
58
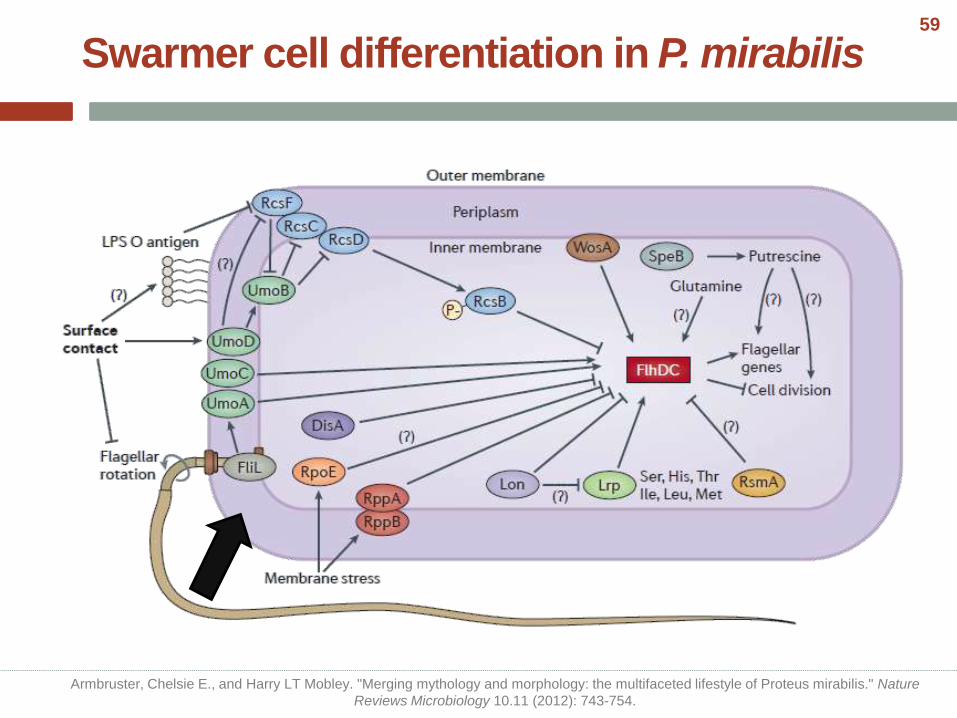
Swarmer cell differentiation in P. mirabilis
Armbruster, Chelsie E., and Harry LT Mobley. "Merging mythology and morphology: the multifaceted lifestyle of Proteus mirabilis." Nature
Reviews Microbiology 10.11 (2012): 743-754.
59
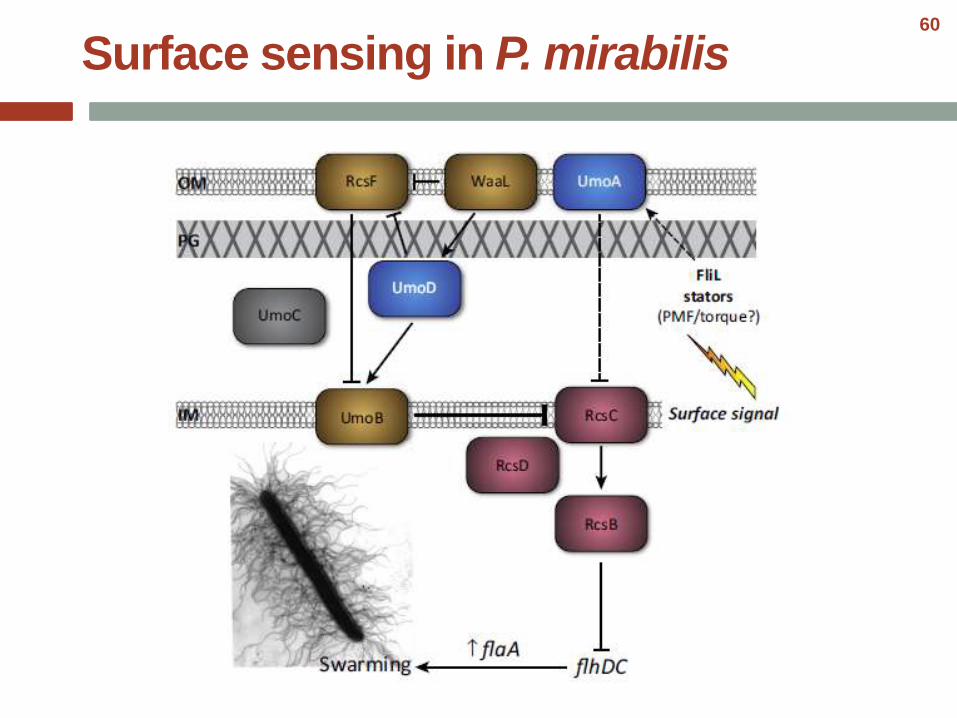
Surface sensing in P. mirabilis60
Summary
Why studying bacterial cell
differentiations?
Basic science is important, isn’t it?
Fighting the bacs!
Making use of ‘em!
61

References to dig deeper!
• Chater, Keith F. "Bacterial Cell Differentiation." eLS (2005).
• Kaiser, Dale. "Cell-Cell Interactions." The Prokaryotes. Springer Berlin
Heidelberg, 2013. 511-528.
• Celiker, Hasan, and Jeff Gore. "Cellular cooperation: insights from
microbes."Trends in cell biology 23.1 (2013): 9-15.
62

Thank You