
Attenuated Francisella asiatica iglC mutant induces protective immunity to francisellosis in tilapia
6
Vaccine 29 (2011) 593–598 Contents lists available at ScienceDirect Vaccine journal homepage: www.elsevier.com/locate/vaccine Attenuated Francisella asiatica iglC mutant induces protective immunity to francisellosis in tilapia Esteban Soto a , Judy Wiles a , Philip Elzer a,b , Kevin Macaluso a , John P. Hawke a,∗ a Department of Pathobiological Sciences, Louisiana State University (LSU)-School of Veterinary Medicine, Skip Bertman Dr., Baton Rouge, LA 70803, USA b LSU AgCenter, Department of Veterinary Science, Dalrymple Dr., Baton Rouge, LA 70803, USA article info Article history: Received 22 February 2010 Received in revised form 15 May 2010 Accepted 10 June 2010 Available online 25 June 2010 Keywords: Francisella asiatica Tilapia Vaccine Immunity abstract Francisella asiatica is a Gram-negative, facultative intracellular bacteria that causes fish francisellosis. Fish francisellosis is a severe sub-acute to chronic granulomatous disease with high mortalities and high infectivity rates in cultured and wild fish. To date, there is no approved vaccine for this widespread emer- gent disease. The goal of this study was to characterize the efficacy of a defined F. asiatica mutant (iglC) as a live attenuated vaccine against subsequent immersion challenge with the wild-type (WT) organism. In previous work, the iglC was found to be attenuated upon intraperitoneal injection and immersion challenges. In vitro, the iglC exhibited reduced growth in tilapia head-kidney derived macrophages, and was significantly attenuated (p < 0.001) as demonstrated by cytopathogenic and apoptosis assays. In this study, the iglC was tested to determine its ability to protect tilapia against challenge with high doses (lethal dose 80) of WT bacteria. Naïve tilapia vaccinated by immersion with a suspension of the iglC and subsequently challenged with WT F. asiatica were protected (90% mean percent survival) from the lethal challenges. F. asiatica-specific antibodies produced in response to immunization with the iglC were subsequently found to protect naïve tilapia against high-dose F. asiatica challenge in passive immuniza- tion experiments. Significant protection (p < 0.001) was obtained when fish were passively immunized and challenged with 10 4 and 10 5 CFU/fish of WT F. asiatica; but not when challenged with 10 6 CFU/fish. This is the first report of a defined live attenuated strain providing protection against F. asiatica in fish. © 2010 Elsevier Ltd. All rights reserved. 1. Introduction Francisella asiatica is a Gram-negative facultative intracellular bacterium, and the causative agent of francisellosis in fish. The bac- terium is an emergent pathogen of tilapia (Oreochromis niloticus), hybrid striped bass (Morone chrysops × M. saxatilis), Atlantic salmon (Salmo salar L.), three-line grunt (Parapristipoma trilineatum), and many other important cultured fish species [1–16]. Infected fish present with non-specific clinical signs such as erratic swim- ming, anorexia, anemia, exophthalmia and high mortality. Upon gross and microscopic examination, several internal organs (mainly spleen and kidney) are enlarged and contain widespread multifo- cal white nodules. Histological examination reveals the presence of multifocal granulomatous lesions, with the presence of numerous small, pleomorphic, cocco-bacilli [15,17]. In previous work, it was found that as few as 23 F. asiatica colony- forming units (CFUs) injected in the peritoneum were capable of causing mortalities in tilapia, and that even fewer were enough to cause serious pathological lesions in important organs like the ∗ Corresponding author. Tel.: +1 225 578 9705; fax: +1 225 578 9701. E-mail address: [email protected] (J.P. Hawke). head-kidney and spleen. Nevertheless, the pathogenic mechanisms that underlie its remarkable infectivity and its capacity to cause disease in a broad range of fish hosts are not known [17,18]. Some of the most interesting virulence genes identified in Fran- cisella tularensis are the genes of the intracellular growth locus (iglA, iglB, iglC, and iglD) present as part of a 30 kb pathogenicity island previously described [19]. The functions of the conserved proteins corresponding to the genes of the Francisella Pathogenicity Island (FPI) are elusive. Overall, Igl proteins appear to be essential for the ability of F. tularensis to survive inside the macrophages and cause disease [19–21]. The intracellular growth locus C pro- tein (IglC) was one of the first of the FPI-encoded proteins found to be highly induced following F. tularensis infection of macrophages [21]. As with other genes mutants in the FPI, iglC mutants fail to grow in macrophages, are deficient in ability to escape from phago- somes, and fail to down-regulate the proinflammatory response in macrophages [20,22–24]. In previous work, we identified homologues to the F. tularen- sis iglA, iglB, iglC, and iglD genes in F. asiatica strain LADL 07-285A isolated from diseased tilapia [18]. An insertion mutation in the iglC gene of F. asiatica LADL 07-285A was constructed by allelic exchange, and the iglC mutant was found to be attenuated fol- lowing intraperitoneal and immersion challenges in tilapia [18]. 0264-410X/$ – see front matter © 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2010.06.040
-
Upload
esteban-soto -
Category
Documents
-
view
213 -
download
1
Transcript of Attenuated Francisella asiatica iglC mutant induces protective immunity to francisellosis in tilapia

Af
Ea
b
a
ARRAA
KFTVI
1
bth(mpmgscms
fct
0d
Vaccine 29 (2011) 593–598
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
ttenuated Francisella asiatica iglC mutant induces protective immunity torancisellosis in tilapia
steban Sotoa, Judy Wilesa, Philip Elzera,b, Kevin Macalusoa, John P. Hawkea,∗
Department of Pathobiological Sciences, Louisiana State University (LSU)-School of Veterinary Medicine, Skip Bertman Dr., Baton Rouge, LA 70803, USALSU AgCenter, Department of Veterinary Science, Dalrymple Dr., Baton Rouge, LA 70803, USA
r t i c l e i n f o
rticle history:eceived 22 February 2010eceived in revised form 15 May 2010ccepted 10 June 2010vailable online 25 June 2010
eywords:rancisella asiaticailapiaaccine
a b s t r a c t
Francisella asiatica is a Gram-negative, facultative intracellular bacteria that causes fish francisellosis.Fish francisellosis is a severe sub-acute to chronic granulomatous disease with high mortalities and highinfectivity rates in cultured and wild fish. To date, there is no approved vaccine for this widespread emer-gent disease. The goal of this study was to characterize the efficacy of a defined F. asiatica mutant (�iglC)as a live attenuated vaccine against subsequent immersion challenge with the wild-type (WT) organism.In previous work, the �iglC was found to be attenuated upon intraperitoneal injection and immersionchallenges. In vitro, the �iglC exhibited reduced growth in tilapia head-kidney derived macrophages, andwas significantly attenuated (p < 0.001) as demonstrated by cytopathogenic and apoptosis assays. In thisstudy, the �iglC was tested to determine its ability to protect tilapia against challenge with high doses
mmunity (lethal dose 80) of WT bacteria. Naïve tilapia vaccinated by immersion with a suspension of the �iglC andsubsequently challenged with WT F. asiatica were protected (90% mean percent survival) from the lethalchallenges. F. asiatica-specific antibodies produced in response to immunization with the �iglC weresubsequently found to protect naïve tilapia against high-dose F. asiatica challenge in passive immuniza-tion experiments. Significant protection (p < 0.001) was obtained when fish were passively immunizedand challenged with 104 and 105 CFU/fish of WT F. asiatica; but not when challenged with 106 CFU/fish.
defi
This is the first report of a. Introduction
Francisella asiatica is a Gram-negative facultative intracellularacterium, and the causative agent of francisellosis in fish. The bac-erium is an emergent pathogen of tilapia (Oreochromis niloticus),ybrid striped bass (Morone chrysops × M. saxatilis), Atlantic salmonSalmo salar L.), three-line grunt (Parapristipoma trilineatum), and
any other important cultured fish species [1–16]. Infected fishresent with non-specific clinical signs such as erratic swim-ing, anorexia, anemia, exophthalmia and high mortality. Upon
ross and microscopic examination, several internal organs (mainlypleen and kidney) are enlarged and contain widespread multifo-al white nodules. Histological examination reveals the presence ofultifocal granulomatous lesions, with the presence of numerous
mall, pleomorphic, cocco-bacilli [15,17].
In previous work, it was found that as few as 23 F. asiatica colony-orming units (CFUs) injected in the peritoneum were capable ofausing mortalities in tilapia, and that even fewer were enougho cause serious pathological lesions in important organs like the
∗ Corresponding author. Tel.: +1 225 578 9705; fax: +1 225 578 9701.E-mail address: [email protected] (J.P. Hawke).
264-410X/$ – see front matter © 2010 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2010.06.040
ned live attenuated strain providing protection against F. asiatica in fish.© 2010 Elsevier Ltd. All rights reserved.
head-kidney and spleen. Nevertheless, the pathogenic mechanismsthat underlie its remarkable infectivity and its capacity to causedisease in a broad range of fish hosts are not known [17,18].
Some of the most interesting virulence genes identified in Fran-cisella tularensis are the genes of the intracellular growth locus(iglA, iglB, iglC, and iglD) present as part of a 30 kb pathogenicityisland previously described [19]. The functions of the conservedproteins corresponding to the genes of the Francisella PathogenicityIsland (FPI) are elusive. Overall, Igl proteins appear to be essentialfor the ability of F. tularensis to survive inside the macrophagesand cause disease [19–21]. The intracellular growth locus C pro-tein (IglC) was one of the first of the FPI-encoded proteins found tobe highly induced following F. tularensis infection of macrophages[21]. As with other genes mutants in the FPI, iglC mutants fail togrow in macrophages, are deficient in ability to escape from phago-somes, and fail to down-regulate the proinflammatory response inmacrophages [20,22–24].
In previous work, we identified homologues to the F. tularen-
sis iglA, iglB, iglC, and iglD genes in F. asiatica strain LADL 07-285Aisolated from diseased tilapia [18]. An insertion mutation in theiglC gene of F. asiatica LADL 07-285A was constructed by allelicexchange, and the �iglC mutant was found to be attenuated fol-lowing intraperitoneal and immersion challenges in tilapia [18].
5 cine 29
RtItccmH
miOem
2
2
(�uah(TbUoaEf
2
54Spffi(If5c5dH
2
tcwltatcm3
94 E. Soto et al. / Vac
ecently, F. asiatica wild-type (WT) and �iglC mutant were foundo be resistant to killing by normal and heat-inactivated serum.t was also found that the WT bacterium was able to invadehe tilapia head-kidney derived macrophages (HKDMs) and repli-ate vigorously within them; causing significant apoptosis andytopathogenicity at 24 and 36 h post-infection [17]. The �iglCutant was found to be defective for survival and replication withinKDM, and its cytotoxicity was significantly reduced in vitro [17].
Here we investigated the ability of an attenuated F. asiatica �iglCutant, given by immersion as a vaccine, to stimulate adaptive
mmune protection against fish francisellosis in tilapia fingerlings.ur results show that the attenuated strain may be used as anffective immersion vaccine against F. asiatica and that the vaccine-ediated protection is partially dependent on antibodies.
. Materials and methods
.1. Bacteria
F. asiatica LADL 07-285A WT was isolated from cultured tilapiaOreochromis sp.) and was described in previous work [15]. The
iglC mutant isolate was made by homologous recombinationsing a PCR product, and its attenuation was demonstrated in vivond in vitro [17,18]. F. asiatica isolates were grown in cystineeart agar supplemented with bovine hemoglobin solution (CHAH)Becton-Dickinson (BD) BBL, Sparks, MD, USA) for 48–72 h at 28 ◦C.he liquid medium consisted of Mueller-Hinton II cation adjustedroth supplemented with 2% IsoVitaleX (BD BBL, Sparks, MD,SA) and 0.1% glucose (MMH) [25]. Broth cultures were grownvernight at 25 ◦C in a shaker at 175 rpm, and bacteria were frozent −80 ◦C in the broth media containing 20% glycerol for later use.scherichia coli DH5� was grown using Luria-Bertani broth or agaror 16–24 h at 37 ◦C.
.2. Preparation of sonicated F. asiatica lysate for ELISA
Approximately 1 × 1012 CFU of F. asiatica were harvested from00 ml of broth culture by centrifugation at 1500 × g for 10 min at◦C in a GSA rotor in an accuspin 3R refrigerated centrifuge (Fishercientific). The pellet was washed three times with Dulbecco’shosphate-buffered saline (PBS; Gibco/Invitrogen, Carlsbad, CA),ollowed by centrifugation at 1500 × g for 10 min. Following thenal wash, the pellet was resuspended in 4 ml of 20 mM Tris–ClpH 8.0) with a protease inhibitor cocktail (Roche Applied Sciences,ndianapolis, IN). The bacteria were sonicated on ice for a 30-s pulse,ollowed by a 30-s rest, 10 times using a Sonic Dismembrator Model00 (Fisher Scientific) at a power of 70%. The samples were thenentrifuged for 1 h at 16,000 × g at 4 ◦C in an Eppendorf centrifuge415 R (Fisher Scientific). Protein concentration of the sonicate wasetermined by the Bradford protein assay (Bio-Rad Laboratories,ercules, CA).
.3. Fish
Adult and fingerling tilapia nilotica (O. niloticus) used duringhe trial were obtained from a source with no history of Fran-isella infection. For verification, a sub-sample of the populationas tested for bacteria by complete clinical, bacteriological, sero-
ogical and molecular analysis as previously described, to ensurehat they were free of F. asiatica [15]. Fingerlings were maintained
t 15 fish per tank in 40-l tanks containing 30 l of water flowinghrough at 25 ◦C, and fed commercial tilapia feed daily (Burris Aqua-ulture Feeds, Franklinton, LA) at 3% fish body weight per day. Theean weight of the fish was 6.4 g. Adults weighing an average of46 g were acclimated for a minimum of 2 months in a flow through
(2011) 593–598
water system at 25 ◦C. Five adult fish were maintained in a 100-ltank containing 80 l of water/tank with constant oxygenation.
2.4. Immunization and challenge
Vaccination trials were conducted using tilapia fingerlings. Fourdifferent �iglC mutant vaccination treatments and a mock immu-nized control treatment were evaluated. Each treatment consistedof eight tanks (15 fish in each tank). The first group of fish was vacci-nated by addition of 107 CFU/ml of the �iglC mutant to 10 l of staticwater and incubation for 180 min. The second group received a doseof 107 CFU/ml of the �iglC mutant but for 30 min. The third groupreceived a dose of 103 CFU/ml of the �iglC mutant for 180 min;and the fourth group received a dose of 103 CFU/ml of the �iglCmutant for 30 min. The control tanks received 100 ml of 1× PBSinto 10 l of static water for a period of 180 min. After either 30 or180 min, the flowing water in each tank was restored to a finalvolume of 30 l of water/tank. Four weeks following a single immer-sion immunization with the �iglC mutant or PBS (control tanks),tilapia fingerlings were challenged by immersion following previ-ously established protocols [18]. Briefly, water volumes in each tankwere adjusted to 10 l of water/tank, and 100 ml of PBS containing F.asiatica suspension was added to each tank for a final concentrationof 108 CFU/ml of WT F. asiatica. The fish colonization and infectionwas allowed to progress for 180 min in static water with oxygena-tion, after which flowing water was re-assumed to a final volume of30 l/tank. Three tanks per treatment were utilized for monitoringmortality every 12 h for 30 days. The remaining tanks were utilizedfor mucus and serum collection and analysis. The protective indexwas calculated according to the formula [26]: RPS = 100% (1 − (%mortality in vaccinated fish/% mortality in control fish)).
2.5. Mucus and serum collection
Mucus and serum collection was performed following previ-ously published protocols [27]. Mucus was sampled from 5 tilapiafingerlings from each immunized or mocked-immunized group at0, 2, 4, 6, 8 and 10 weeks post-vaccination. Fish were euthanizedwith an overdose of MS-222 and mucus collected by swabbing bothsides of the fish 10 times from head to tail with a cotton applica-tor. Swabs were placed in 1.5 ml microcentrifuge tubes containing0.9 ml of PBS supplemented with 0.02% (w/v) sodium azide. Tubeswere stored overnight at 4 ◦C. The next morning, tubes and swabswere vigorously vortexed for 2 min and liquid removed from theswabs by pressing against the side of the tube. The resulting liquidwas centrifuged at 3000 × g for 10 min and the supernatant col-lected and frozen at −20 ◦C in polypropylene tubes until analyzedby ELISA.
Blood samples were obtained from the same fish at each timepoint, by caudal venepuncture and collection with plain Fisher-brand micro-hematocrit capillary tube (Fisher Scientific, Pittsburg,PA). Blood was held at 25 ◦C for 1 h and then separated with cen-trifugation at 400 × g for 5 min and then stored at −20 ◦C for lateranalysis by ELISA.
2.6. ELISA
An ELISA was developed to quantify anti-F. asiatica antibodyproduced by tilapia in serum and mucus. Immulon II 96-well flat-bottom microtiter plates (Thermo Labsystems, Franklin, MA, USA)were coated overnight at 4 ◦C with a 7 �g protein ml−1 solution of
sonicated F. asiatica whole cell antigen in 0.05 M carbonate coat-ing buffer, pH 9.6, at 100 �l per well. Plates were then washedthree times in PBS containing 0.05% Tween-20 (PBST). The wellswere blocked for 1 h at room temperature (RT) with filter-sterilizedPBS containing 0.05% Tween 20 (Sigma) and 1% bovine serum albu-
ine 29
msaw2s(t2adaoawsMTv
2
ttv(5aofaaabfilHpFlaaoLM
2
o1Palwo
2
odlp[
E. Soto et al. / Vacc
in (BSA; Fraction V, Sigma) (PBST-BSA). Tilapia serum and mucusamples were diluted 1:1000 or 1:50 (respectively) in PBST-BSA,nd 100 �l of the resulting solution was added to three replicateells of the microtiter plate. The plate was incubated at 25 ◦C forh and washed 3× with PBST. Mouse anti-tilapia IgM heavy chain
pecific monoclonal antibody kindly provided by Richard ShelbyUSDA/ARS, Auburn, AL) was diluted 1:100 in PBST and 100 �l ofhis solution added to each well [28]. The plate was incubated at5 ◦C for 1 h and washed 3× with PBST. Peroxidase-conjugated goatnti-mouse IgG (Pierce Biotechnology, Rockford, Illinois, USA) wasiluted 1:10,000 in PBST and added to each well. After incubationt 25 ◦C for 1 h, the plate was washed again 3× in PBST and 100 �lf ABTS Peroxidase Substrate System (KPL, Gaithersburg, MD) wasdded to each well. The ELISA reaction was stopped after 30 minith 100 �l 1% sodium dodecyl sulfate (SDS), and the optical den-
ity (OD) of the reactions was read at 405 nm with a SpectraMax2/M2e Microplate Readers (Molecular Devices, Sunnyvale, CA).
he relative amount of specific antibody was measured as the ODalue.
.7. Adoptive transfer studies
Twenty adult fish serum donors were immunized by intraperi-oneal (IP) vaccination with the �iglC mutant. Prior to challengehe fish were anesthetized with MS-222 (100 mg/l). Intraperitonealaccinated fish received a 0.1 ml injection of bacterial suspension107 CFU/fish). Serum collected from the 20 immunized fish at 4,, and 6 weeks post-immunizations were pooled together and wasnalyzed by ELISA. Endpoint titers were reported as the reciprocalf the last dilution yielding an OD more than twice that of the serumrom naïve control fish. Normal fish serum was obtained from 20dult naïve fish injected with 1× PBS and processed the same ways the immunized fish. Heat-inactivated immunized serum (HIIS)nd heat-inactivated normal serum (HINS), were obtained by incu-ating serum obtained from immunized and mocked-immunizedsh at 56 ◦C for 30 min. Two hundred and forty naïve tilapia finger-
ings (20/tank) were injected IP with either 200 �l of pooled HIIS,INS or PBS 24 h before IP challenge with F. asiatica WT. Three tankser treatment were challenged with either 103, 104, 105, or 106 CFU. asiatica/fish by IP injection. During the subsequent 21-day chal-enge period, fish were monitored daily for clinical signs of diseasend mortality. Moribund and dead fish were removed twice daily,nd bacterial samples were aseptically obtained from the spleenf morbid and dead fish to confirm the presence of F. asiatica. TheD50 for each treatment was calculated by the method of Reed anduench [29], at day 21 post-injection.
.8. Direct complement lysis
F. asiatica WT and E. coli DH5�, were cultured as previ-usly described. Bacteria were adjusted to a concentration of× 107 CFU/ml in PBS. A 1:1 ratio of the bacterial isolates to eitherBS, normal (NS), immunized (IS) tilapia sp. serum was combinednd the samples were incubated for 2 h at room temperature. Fol-owing incubation sub-samples of the bacteria/serum mixtures
ere collected at 0, 1, and 2 h, serially diluted in PBS and spottednto either CHAH (F. asiatica) or LB (E. coli) plates.
.9. Opsonophagocytosis assays
To examine the opsonic potential of the immune sera, an
psonophagocytosis assay was established. Tilapia head-kidneyerived macrophages (HKDMs) were collected and purified fol-owing previously established protocols [30,31]. Modifications toreviously established protocols were used to infect tilapia HKDM32,33]. Briefly, 5-day cultures of tilapia HKDM in 96-well plates
(2011) 593–598 595
containing 1-5 × 105 cells/well were used. F. asiatica was grownfor a period of 8 h in MMH at 25 ◦C. Optical density (OD600) of theculture was determined and the bacteria were adjusted to a finalconcentration of 5 × 108 CFU/ml. One ml aliquots of the bacterialsuspension were pelleted at 10,000 × g for 5 min in an Eppendorf5415 D centrifuge (Eppendorf-Brinkman, Westbury, NY), and thepellet was resuspended in either 1 ml of HINS or HIIS. Ten-fold serialdilutions were plated on CHAH after incubation to determine totalbacterial cell viability. After 1 h incubation, the 96-well plate wasinoculated with 10 �l of opsonized bacteria per well to achieve amultiplicity of infection (MOI) of 50 bacteria: 1 macrophage. Theplates were centrifuged for 5 min at 400 × g to synchronize bacte-rial contact with macrophages. Following 2 h incubation at 25 ◦Cwith 5% CO2, the cells were washed three times with warm media(25 ◦C), further incubated with fresh media and lysed for 15 min attime 0, 24, and 48 h by the addition of 100 �l of 1% Saponin in PBS.The lysates were serially diluted and spread onto CHAH plates todetermine viable counts. Experiments were performed in triplicateon a minimum of three separate occasions with similar results.
2.10. Statistical analysis
The Statistical Analysis System (SAS Institute, Inc., 2003) wasused with the general linear models procedure (PROC GLM) toconduct analysis of variance (ANOVA) of a factorial arrangementof treatments. When the overall test indicated significance, pair-wise comparisons of main effects were calculated with Tukey’s test.Interaction effects were examined with pairwise t-test comparisonof least-square means. For the mortality studies the percent mortal-ities were transformed with an arcsine transformation to normalizethe data. To ensure overall protection level of Type I error, onlyprobabilities associated with pre-planned comparisons were used.All comparisons were considered significant at (p < 0.05).
3. Results
3.1. Immersion vaccination with �iglC protected tilapiafingerlings against homologous F. asiatica immersion challenge
To evaluate the efficacy of �iglC in protecting tilapia fingerlingsagainst virulent F. asiatica immersion challenge, tilapia fingerlingswere vaccinated by immersion by four different treatments. Vac-cination with a dose of 107 CFU/ml of water for a period of 30 or180 min conferred 68.75% and 87.5% relative percent survival (RPS),respectively, against otherwise lethal (80% mortality) immersionchallenge with the WT isolate during a period of 30 days. Vacci-nation with a dose of 103 CFU/ml of water for a period of 30 minor 180 min conferred 56.25% and 62.5% RPS, respectively, againstimmersion challenge with the WT isolate. Mock (PBS)-vaccinatedfish succumbed to the infection by day 7, presenting clinical signs ofthe disease, including ascites and widespread granulomas in spleenand kidney. Fish vaccinated with either treatments of �iglC had sig-nificantly higher survival rates than those mocked vaccinated withPBS after challenge with WT F. asiatica (p < 0.05) (Fig. 1). Juveniletilapia vaccinated with either treatment of �iglC generated a weakserum and mucosal antibody response that was not significantlydifferent than that of controls at 2, 4 and 6 weeks post-vaccination(Figs. 2 and 3). However, after the WT immersion challenge, theserum and mucosal samples from iglC vaccinated fish with a doseof 107 CFU/ml of water for a period of 30 and 180 min, resulted
in a significantly greater secondary antibody response at weeks 8and 10 post-initial vaccination (p < 0.05) (Figs. 2 and 3). The non-immunized fish showed an increased primary antibody responseafter WT challenge when compared to antibodies levels at week 0(Figs. 2 and 3).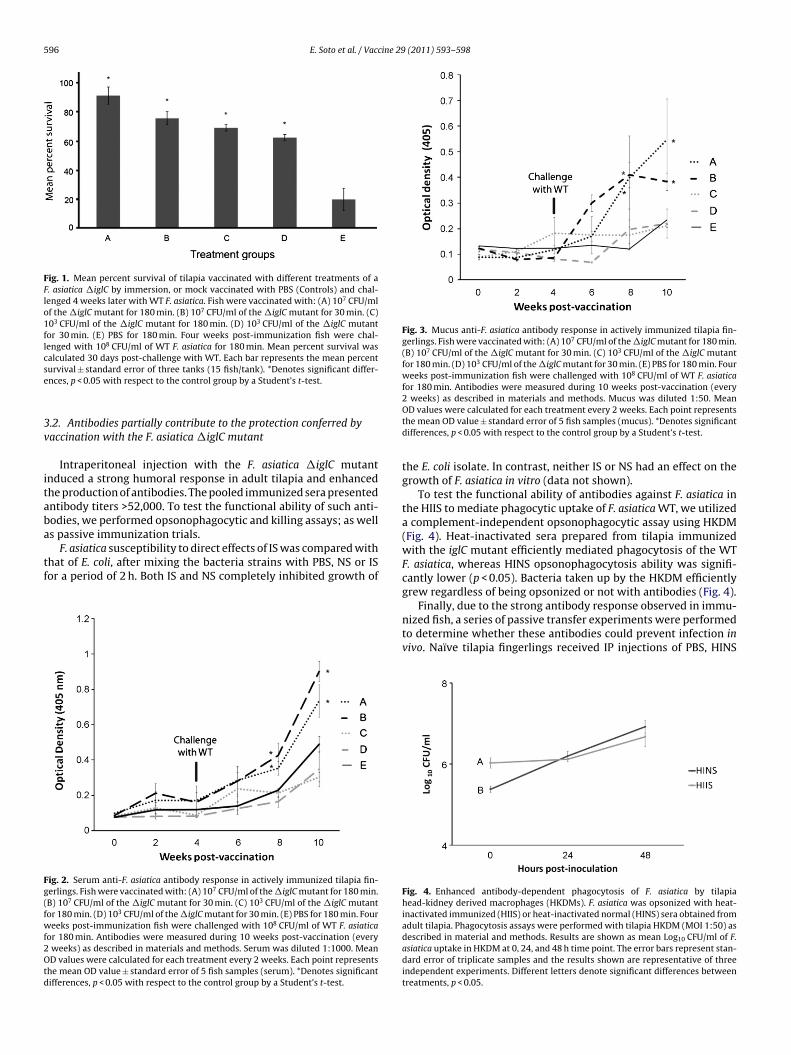
596 E. Soto et al. / Vaccine 29 (2011) 593–598
Fig. 1. Mean percent survival of tilapia vaccinated with different treatments of aF. asiatica �iglC by immersion, or mock vaccinated with PBS (Controls) and chal-lenged 4 weeks later with WT F. asiatica. Fish were vaccinated with: (A) 107 CFU/mlof the �iglC mutant for 180 min. (B) 107 CFU/ml of the �iglC mutant for 30 min. (C)103 CFU/ml of the �iglC mutant for 180 min. (D) 103 CFU/ml of the �iglC mutantfor 30 min. (E) PBS for 180 min. Four weeks post-immunization fish were chal-lenged with 108 CFU/ml of WT F. asiatica for 180 min. Mean percent survival wascse
3v
itaba
tf
Fg(fwf2Otd
Fig. 3. Mucus anti-F. asiatica antibody response in actively immunized tilapia fin-gerlings. Fish were vaccinated with: (A) 107 CFU/ml of the �iglC mutant for 180 min.(B) 107 CFU/ml of the �iglC mutant for 30 min. (C) 103 CFU/ml of the �iglC mutantfor 180 min. (D) 103 CFU/ml of the �iglC mutant for 30 min. (E) PBS for 180 min. Fourweeks post-immunization fish were challenged with 108 CFU/ml of WT F. asiaticafor 180 min. Antibodies were measured during 10 weeks post-vaccination (every
alculated 30 days post-challenge with WT. Each bar represents the mean percenturvival ± standard error of three tanks (15 fish/tank). *Denotes significant differ-nces, p < 0.05 with respect to the control group by a Student’s t-test.
.2. Antibodies partially contribute to the protection conferred byaccination with the F. asiatica �iglC mutant
Intraperitoneal injection with the F. asiatica �iglC mutantnduced a strong humoral response in adult tilapia and enhancedhe production of antibodies. The pooled immunized sera presentedntibody titers >52,000. To test the functional ability of such anti-odies, we performed opsonophagocytic and killing assays; as well
s passive immunization trials.F. asiatica susceptibility to direct effects of IS was compared withhat of E. coli, after mixing the bacteria strains with PBS, NS or ISor a period of 2 h. Both IS and NS completely inhibited growth of
ig. 2. Serum anti-F. asiatica antibody response in actively immunized tilapia fin-erlings. Fish were vaccinated with: (A) 107 CFU/ml of the �iglC mutant for 180 min.B) 107 CFU/ml of the �iglC mutant for 30 min. (C) 103 CFU/ml of the �iglC mutantor 180 min. (D) 103 CFU/ml of the �iglC mutant for 30 min. (E) PBS for 180 min. Foureeks post-immunization fish were challenged with 108 CFU/ml of WT F. asiatica
or 180 min. Antibodies were measured during 10 weeks post-vaccination (everyweeks) as described in materials and methods. Serum was diluted 1:1000. MeanD values were calculated for each treatment every 2 weeks. Each point represents
he mean OD value ± standard error of 5 fish samples (serum). *Denotes significantifferences, p < 0.05 with respect to the control group by a Student’s t-test.
2 weeks) as described in materials and methods. Mucus was diluted 1:50. MeanOD values were calculated for each treatment every 2 weeks. Each point representsthe mean OD value ± standard error of 5 fish samples (mucus). *Denotes significantdifferences, p < 0.05 with respect to the control group by a Student’s t-test.
the E. coli isolate. In contrast, neither IS or NS had an effect on thegrowth of F. asiatica in vitro (data not shown).
To test the functional ability of antibodies against F. asiatica inthe HIIS to mediate phagocytic uptake of F. asiatica WT, we utilizeda complement-independent opsonophagocytic assay using HKDM(Fig. 4). Heat-inactivated sera prepared from tilapia immunizedwith the iglC mutant efficiently mediated phagocytosis of the WTF. asiatica, whereas HINS opsonophagocytosis ability was signifi-cantly lower (p < 0.05). Bacteria taken up by the HKDM efficientlygrew regardless of being opsonized or not with antibodies (Fig. 4).
Finally, due to the strong antibody response observed in immu-nized fish, a series of passive transfer experiments were performedto determine whether these antibodies could prevent infection invivo. Naïve tilapia fingerlings received IP injections of PBS, HINS
Fig. 4. Enhanced antibody-dependent phagocytosis of F. asiatica by tilapiahead-kidney derived macrophages (HKDMs). F. asiatica was opsonized with heat-inactivated immunized (HIIS) or heat-inactivated normal (HINS) sera obtained fromadult tilapia. Phagocytosis assays were performed with tilapia HKDM (MOI 1:50) asdescribed in material and methods. Results are shown as mean Log10 CFU/ml of F.asiatica uptake in HKDM at 0, 24, and 48 h time point. The error bars represent stan-dard error of triplicate samples and the results shown are representative of threeindependent experiments. Different letters denote significant differences betweentreatments, p < 0.05.

E. Soto et al. / Vaccine 29
Fig. 5. Adoptive transfer of heat-inactivated normal serum (HINS), heat-inactivatedimmunized serum (HIIS) or PBS to naïve tilapia fingerlings. Immune sera was col-lected from 20 adult tilapia vaccinated by intraperitoneal injection (IP) with the�iglC mutant 4, 5 and 6 weeks post-vaccination. Sera were pooled and antibod-ies titers were measured before passively immunized the fingerlings. Normal serawere collected and pooled form 20 adult tilapia injected with PBS 4, 5 and 6 weekspost-injection. Naïve fingerlings (60 fish/treatment) were injected IP with 200 �l ofpooled HINS, HIIS or PBS 24 h before IP challenge with 103, 104, 105 or 106 CFU/fish ofF. asiatica WT. Animals were monitored daily for morbidity and mortality. Results arerepresentative of two independent experiments. Mean percent mortality for eachtmnt
o1emcapmtP
4
rohRetlta6twofeanays[Sa
F
reatment was calculated 21 days post-challenge with WT. Each bar represents theean percent mortality ± standard error of three tanks (20 fish/tank). *Denotes sig-
ificant differences, p < 0.05 with respect to the control group (PBS) by a Student’s-test.
r HIIS sera (200 �l) collected from adult tilapia immunized with07 CFU/fish. The tilapia fingerlings were then challenged (IP) withither 103, 104, 105 or 106 CFU/fish of WT F. asiatica and wereonitored daily for health and survival for a total of 21 days post-
hallenge. Although passive immunization of HIIS did not protectgainst high doses of the bacterium (106 CFU/fish) injected in theeritoneum of naïve fingerlings, significant (p < 0.05) reductions inortality were observed in HIIS immunized fish when challenged
o 104 and 105 CFU/fish and compared to those immunized withBS or HINS (Fig. 5).
. Discussion
A live attenuated vaccine given to the fish by the immersionoute, has the advantage of directly targeting the natural routesf attachment and penetration of the bacteria into the fish andence inducing protective immunity at the primary site of infection.esults showed that an immersion vaccination with four differ-nt treatments of a �iglC mutant significantly (p < 0.05) protectsilapia fingerlings against homologous F. asiatica immersion chal-enge (Fig. 1). Results of immunization trials indicated that whenhe �iglC mutant vaccine was delivered for either 30 or 180 mt a dose of 107 CFU/ml, relative percent survival (RPS) values of8.75% and 87.5% were obtained, demonstrating the potential ofhe vaccine to prevent francisellosis in tilapia. During the first 4eeks post-vaccination, a relatively small antibody response was
bserved in immunized fish, and they were not significantly dif-erent to those observed in the control groups. However, uponxposure to WT F. asiatica, a significantly higher (p < 0.05) mucosalnd humoral antibody response was evident in the fish vacci-ated with a dose of 107 CFU/ml (Figs. 2 and 3). The importance ofntibody-mediated protection at the mucosal or systemic level haset to be determined for F. asiatica. However, antibody has beenhown to correlate with protection for Flavobacterium columnare
27]; F. psychrophilum [34], Vibrio spp. [35], Yersinia ruckeri [36],treptococcus iniae [37], A. hydrophila [38], Edwardsiella tarda [39],nd E. ictaluri [40] in cultured fish species.The passive immunity studies described here demonstrate that. asiatica-specific antibodies mediate protection after IP injection
(2011) 593–598 597
of different concentration of F. asiatica WT (Fig. 5). Thus we believethat the F. asiatica-specific antibody response is a useful componentof the protective immune response to lethal F. asiatica infectionin fish. Since F. asiatica is a facultative intracellular organism, thebacteria can exist in an extracellular form in the tilapia, thus it isconceivable that antibodies are able to prevent the systemic spreadof bacteria. Since mucosal responses observed in this study demon-strated that vaccinated fish presented higher amount of anti-F.asiatica antibodies at 6, 8 and 10 weeks post-vaccination, it canalso be speculated that mucosal antibody might serve to inhibit thedamage or possible external bacterial colonization in fish tissues(Figs. 2 and 3).
In many Gram-negative bacteria, the protective antibody mayact in conjunction with complement in vivo in a direct bacteri-cidal response, or aid phagocytes in their ability to engulf andkill the pathogen. Antibodies may directly influence the phago-cytic activity of Fc receptor-bearing cells, such as macrophagesand NK cells, to enhance phagocytosis of intracellular pathogens.Our results demonstrate that neither NS nor IS obtained fromadult tilapia, damaged the bacteria in vitro, however sera from�iglC mutant-vaccinated animals exhibited enhanced antibody-mediated phagocytosis of F. asiatica in HKDM (Fig. 4). In F. tularensis,antibody-mediated bacterial clearance and in vitro analysis ofopsonophagocytosis revealed that IFN-� treatment of macrophagesinduced rapid killing of intracellular bacteria; but opsonophagocy-tosis via Fc�Rs in the absence of IFN-� failed to control bacterialreplication [41,42]. These results suggested that Fc�R-mediatedcellular activation synergizes with IFN-� for intracellular bacterialkilling [41,42]. In this study, non-activated HKDM from naïve fishfailed to control intracellular bacterial replication, even thoughtthey were opsonized with antibodies against F. asiatica (Fig. 4). Therole of cell-mediated immunity has yet to be determined in thecontrol of fish francisellosis; but activation of fish macrophageshave been shown to play a role in the defense against impor-tant fish pathogens like Aeromonas salmonicida and Renibacteriumsalmoninarum [43,44]. In F. tularensis, cell mediated immunity haslong been believed to be critical for protection, furthermore, syn-ergy between antibodies, T cell-derived cytokines, and phagocytesappears to be critical to achieve sterilizing immunity against thepathogen. Defined F. novicida mutant strains like �iglB and �iglC,protected mice against homologous challenges via the participa-tion of IFN-� producing cells such as natural killers (NKs) and Tcells [44,45].
In summary, an attenuated strain of F. asiatica (�iglC) wascharacterized as a potential live-vaccine for fish francisellosis.Immunization of tilapia nilotica with this strain by immer-sion delivery provided long lasting protective immune responses(p < 0.05), as demonstrated by antibodies levels, and the antibodiesdirected to F. asiatica were protective as shown in passive immunitytrials.
Acknowledgments
We gratefully thank Dr. Richard Shelby (USDA/ARS, Auburn, AL)for sharing the mouse anti-tilapia IgM antibodies used in this study.We also thank Dr. Ron Thune, Natthida Petchampai, Dr. PiyanateSunyakumthorn, Dr. Walairat Pornwiroon and Dr. Chutima Thep-parit from the Pathobiological Sciences Department, LSU-School ofVeterinary Medicine for their skillful technical assistance.
References
[1] Kamaishi T, Fukuda Y, Nishiyama M, Kawakami H, Matsuyama T, YoshinagaT, et al. Identification and pathogenicity of intracellular Francisella bacteriumin three-line grunt Parapristipoma trilineatum. Fish Pathol 2005;40(2):67–71.

5 cine 29
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[44] Ellis AE. Immunity to bacteria in fish. Fish Shellfish Immunol
98 E. Soto et al. / Vac
[2] Hsieh CY, Tung MC, Tu C, Chang CD, Tsai SS. Enzootics of visceral granulomasassociated with Francisella-like organism infection in tilapia (Oreochromis spp.).Aquaculture 2006;254(1–4):129–38.
[3] Ostland VE, Stannard JA, Creek JJ, Hedrick RP, Ferguson HW, Carlberg JM,et al. Aquatic Francisella-like bacterium associated with mortality of inten-sively cultured hybrid striped bass Morone chrysops × M. saxatilis. Dis AquatOrg 2006;72(2):135–45.
[4] Kay W, Petersen BO, Duus JØ, Perry MB, Vinogradov E. Characterization of thelipopolysaccharide and �-glucan of the fish pathogen Francisella victoria. FebsJ 2006;273(13):3002–13.
[5] Nylund A, Ottem KF, Watanabe K, Karlsbakk E, Krossøy B. Francisella sp. (familyFrancisellaceae) causing mortality in norwegian cod (Gadus morhua) farming.Arch Microbiol 2006;185(5):383–92.
[6] Olsen AB, Mikalsen J, Rode M, Alfjorden A, Hoel E, Straum-Lie K, et al. A novelsystemic granulomatous inflammatory disease in farmed Atlantic cod, Gadusmorhua L., associated with a bacterium belonging to the genus Francisella. J FishDis 2006;29(5):307–11.
[7] Mikalsen J, Olsen AB, Tengs T, Colquhoun DJ. Francisella philomiragia subsp.noatunensis subsp. nov., isolated from farmed atlantic cod (Gadus morhua L.).Int J Syst Evol Microbiol 2007;57(9):1960–5.
[8] Ottem KF, Nylund A, Karlsbakk E, Friis-Møller A, Krossøy B, Knappskog D.New species in the genus Francisella (gammaproteobacteria; Francisellaceae);Francisella piscicida sp. nov. isolated from cod (Gadus morhua). Arch Microbiol2007;188(5):547–50.
[9] Mauel MJ, Soto E, Moralis JA, Hawke J. A piscirickettsiosis-like syndrome incultured Nile tilapia in Latin America with Francisella spp. as the pathogenicagent. J Aquat Anim Health 2007;19(1):27–34.
10] Hsieh C, Wu Z, Tung M, Tsai S. PCR and in situ hybridization for the detection andlocalization of a new pathogen Francisella-like bacterium (FLB) in ornamentalcichlids. Dis Aquat Org 2007;75(1):29–36.
11] Ottem KF, Nylund A, Karlsbakk E, Friis-Møller A, Krossøy B. Characterization ofFrancisella sp., GM2212, the first Francisella isolate from marine fish, Atlanticcod (Gadus morhua). Arch Microbiol 2007;187(5):343–50.
12] Birkbeck TH, Bordevik M, Frøystad MK, Baklien Å. Identification of Francisellasp. from Atlantic salmon, Salmo salar L., in Chile. J Fish Dis 2007;30(8):505–7.
13] Ottem KF, Nylund A, Isaksen TE, Karlsbakk E, Bergh Ø. Occurrence of Francisellapiscicida in farmed and wild Atlantic cod, Gadus morhua L., in Norway. J Fish Dis2008;31(7):525–34.
14] Ottem KF, Nylund A, Isaksen TE, Karlsbakk E, Bergh Ø. Occurrence of Francisellapiscicida in farmed and wild Atlantic cod, Gadus morhua L., in Norway. J Fish Dis2008;31(8):637.
15] Soto E, Hawke JP, Fernandez D, Morales AJ. Francisella sp., an emerging pathogenof tilapia, Oreochromis niloticus (L.), in Costa Rica. J Fish Dis 2009;32(8):713–22.
16] Ottem KF, Nylund A, Karlsbakk E, Friis-Møller A, Kamaishi T. Elevation ofFrancisella philomiragia subsp. noatunensis Mikalsen et al. (2007) to Francisellanoatunensis comb. nov. [syn. Francisella piscicida Ottem et al. (2008) syn. nov.]and characterization of Francisella noatunensis subsp. orientalis subsp. nov., twoimportant fish pathogens. J Appl Microbiol 2009;106(4):1231–43.
17] Soto E, Fernandez D, Thune R, Hawke JP. Interaction of Francsiella asiaticawith Tilapia nilotica (Oreochromis niloticus) innate immunity. Infect Immun2010;78(5):2070–8.
18] Soto E, Fernandez D, Hawke JP. Attenuation of the fish pathogen Francisella sp.by mutation of the iglC gene. J Aquat Anim Health 2009;(21):140–9.
19] Nano FE, Zhang N, Cowley SC, Klose KE, Cheung KKM, Roberts MJ, et al. A Fran-cisella tularensis pathogenicity island required for intra-macrophage growth. JBacteriol 2004;186(19):6430–6.
20] Nano FE, Schmerk C. The Francisella pathogenicity island. Ann NY Acad Sci2007;1105(June):122–37.
21] Golovliov I, Ericsson M, Sandstrom G, Tarnvik A, Sjostedt A. Identification ofproteins of Francisella tularensis induced during growth in macrophages andcloning of the gene encoding a prominently induced 23-kilo Dalton protein.Infect Immun 1997;65(6):2183–9.
22] Golovliov I, Baranov V, Krocova Z, Kovarova H, Sjostedt A. An attenuated strainof the facultative intracellular bacterium Francisella tularensis can escape thephagosome of monocytic cells. Infect Immun 2003;71(10):5940–50.
23] Lai XH, Golovliov I, Sjostedt A. Expression of IglC is necessary for intracellu-lar growth and induction of apoptosis in murine macrophages by Francisellatularensis. Microb Pathog 2004;37(5):225–30.
[
(2011) 593–598
24] Ludu JS, de Bruin OM, Duplantis BN, Schmerk CL, Chou AY, Elkins KL, et al.The Francisella pathogenicity island protein PdpD is required for full virulenceand associates with homologues of the type VI secretion system. J Bacteriol2008;190(13):4584–95.
25] Baker CN, Hollis DG, Thornsberry C. Antimicrobial susceptibility testing ofFrancisella tularensis with a modified Mueller-Hinton broth. J Clin Microbiol1985;22(2):212–5.
26] Aned DF. Potency testing of fish vaccines. In: Anderson DP, Hennessen H, edi-tors. Fish biologics: serodiagnostics and vaccines. Developments in biologicalstandardization. Basel: Karger; 1981. p. 447–54.
27] Grabowski LD, LaPatra SE, Cain KD. Systemic and mucosal antibody response intilapia, Oreochromis niloticus (L.), following immunization with Flavobacteriumcolumnare. J Fish Dis 2004;27(10):573–81.
28] Shelby RA, Shoemaker CA, Klesius PH. Detection of humoral response to Strep-tococcus iniae infection of Nile tilapia, Oreochromis niloticus, by a monoclonalantibody-based ELISA. J Appl Aquat 2002;(12):23–31.
29] Reed LJ, Muench H. A simple method of estimating fifty percent end points. AmJ Hyg 1938;(27):493–7.
30] Neumann NF, Barreda D, Belosevic M. Production of a macrophage growthfactor(s) by a goldfish macrophage cell line and macrophages derived fromgoldfish kidney leukocytes. Dev Comp Immunol 1998;22(4):417–32.
31] Secombes CJ. Isolation of salmonid macrophages and analysis of their killingactivity. In: Stolen JS, Fletcher TC, Anderson DP, Robertson BS, van MuiswinkelWB, editors. Techniques in fish immunology, vol. 1. New Jersey: SOS Publica-tions; 1992. p. 137–54.
32] Lauriano CM, Barker JR, Yoon S-, Nano FE, Arulanandam BP, Hassett DJ, et al.MgIA regulates transcription of virulence factors necessary for Francisellatularensis intra-amoebae and intra-macrophage survival. Proc Natl Acad SciUSA 2004;101(12):4246–9.
33] Lauriano CM, Barker JR, Nano FE, Arulanandam BP, Klose KE. Allelicexchange in Francisella tularensis using PCR products. FEMS Microbiol Lett2003;229(2):195–202.
34] LaFrentz BR, LaPatra SE, Jones GR, Cain KD. Passive immunization of rainbowtrout, Oncorhynchus mykiss (walbaum), against Flavobacterium psychrophilum,the causative agent of bacterial coldwater disease and rainbow trout fry syn-drome. J Fish Dis 2003;26(7):377–84.
35] Akhlaghi M. Passive immunization of fish against vibriosis, com-parison of intraperitoneal, oral and immersion routes. Aquaculture1999;180(3–4):191–205.
36] Furones MD, Rodgers CJ, Munn CB. Yersinia ruckeri, the causal agent of entericredmouth disease (ERM) in fish. Annu Rev Fish Dis 1993;3C:105–25.
37] Shelby RA, Klesius PH, Shoemaker CA, Evans JJ. Passive immunization of tilapia,Oreochromis niloticus (L.), with anti-Streptococcus iniae whole sera. J Fish Dis2002;25(1):1–6.
38] Ruangpan L, Kitao T, Yoshida T. Protective efficacy of Aeromonas hydrophilavaccines in Nile tilapia. Vet Immunol Immunopathol 1986;12(1–4):345–50.
39] Gutierrez MA, Miyazaki T, Harta H, Kim M. Protective properties of eggyolk IgY containing anti-Edwardsiella tarda antibody against paracolo dis-ease in the Japanese eel, Anguilla japonica Temminck & Schlegel. J Fish Dis1993;(16):113–22.
40] Klesius PH, Sealey WM. Characteristics of serum antibody in enteric septicemiaof catfish. J Aquat Anim Health 1995;(7):205–10.
41] Pammit MA, Raulie EK, Lauriano CM, Klose KE, Arulanandam BP. Intranasal vac-cination with a defined attenuated Francisella novicida strain induces gammainterferon-dependent antibody-mediated protection against tularemia. InfectImmun 2006;74(4):2063–71.
42] Kirimanjeswara GS, Olmos S, Bakshi CS, Metzger DW. Humoral and cell-mediated immunity to the intracellular pathogen Francisella tularensis.Immunol Rev 2008;225(1):244–55.
43] Ellis AE. Innate host defense mechanisms of fish against viruses and bacteria.Dev Comp Immunol 2001;25(8–9):827–39.
1999;9(4):291–308.45] Cong Y, Yu J-, Guentzel MN, Berton MT, Seshu J, Klose KE, et al. Vaccination
with a defined Francisella tularensis subsp. novicida pathogenicity island mutant(�iglB) induces protective immunity against homotypic and heterotypic chal-lenge. Vaccine 2009;27(41):5554–61.


















