
Asymmetric reduction of acetophenone by immobilized Hansenula capsulata cells
6
JOURNAL OFFERMENTATION AND BIOENGINEERING Vol. 85, No. 3, 322-327. 1998 Asymmetric Reduction of Acetophenone by Immobilized Hansenula capsulata Cells YURI HASEGAWA, SHUJI ADACHI,* AND RYUICHI MATSUNO Division of Applied Life Sciences, Graduate School of Agriculture, Kyoto University, Sakyo-ku, Kyoto 606-01, Japan Received 30 October 1997/Accepted 15 December 1997 Immobilized cells of the yeast Hansenula capsulata IF0 0974 could catalyze the asymmetric reduction of acetophenone to Q-1-phenylethanol with almost complete enantioselectivity using glucose as an energy source. Depending on the operational conditions, 1 to 3 mol glucose were required for the reduction of 1 mol of acetophenone. A packed-bed reactor gave a higher conversion than a batch reactor because of its anaerobic conditions and the fact that reverse oxidation was prevented in the former reactor. However, the formation of CO2 bubbles made continuous operation difficult. A shallow-bed reactor, from which CO, bubbles could be easily removed, improved the conversion to over 90%. Continuous production of Q-1-phenylethanol was realized at a significantly high enantiomerlc excess for 220 h by incorporating cell re-cultivation. [Key words: asymmetric reduction, I-phenylethanol, Hansenula capsulata, shallow-bed reactor] Optically active compounds are abundant in our sur- roundings -as flavors, sugars, amino acids, and pro- teins in our daily foods, medicines, pesticides, liquid crystals, and so on. During the synthesis of optically active secondary alcohols as synthons for the synthesis of compounds with chiral center(s), lipase and esterase have been widely used (l-4) due to their high stability and the fact that no cofactor is needed (5). Oxidoreductases, which directly catalyze the formation of the chiral hy- droxyl group from the achiral ketone group instead of it being indirectly formed through stereoselective hydrol- ysis of the alcohol-fatty acid ester, as in the case of lipase, have not yet been adopted for industrial use because of difficulties in cofactor NAD(P)H/NAD(P)+ regeneration and poor cofactor stability in solution (6). For the practical use of oxidoreductases, a cofactor regeneration system that will allow effective use of the expensive cofactor, and hence the economical production of optically active compounds on a larger scale, is required (7, 8). One way to achieve this is to use whole cells as the catalyst. An important advantage of using whole cells is that all of the reaction system for cofactor regeneration is present within the catalytic cells them- selves (9, 10). Other advantages are that purification of the enzyme is not required, simple and cheap com- pounds such as glucose or ethanol can be adopted as an energy source for cofactor regeneration, and the self- replicating feature of viable microorganisms is utilized when whole cells are used. We have previously reported the production of optically active 1-phenylethanol (l-PA) through enantioselective oxidation of its racemate with yeast cells (11, 12). Two yeast strains, Pachysolen tannophilus IF0 1007 and Han- senula capsulata IF0 0974, were proved to have com- plete enantioselectivity for conversion of the @)-isomer of l-PA to the corresponding ketone, acetophenone. Using cells of these yeasts, the continuous production of (R)-l-PA was achieved in a fluidized-bed bioreactor. One difficulty in obtaining the residual isomer through stereoselective oxidation of the other isomer arises from the fact that the enantiomeric excess of the residual * Corresponding author. isomer is largely dependent on the conversion of the other isomer (13). According to the definition of enantio- merit excess, a conversion of 49.8%, based on the race- mate, must be accomplished in order to obtain a product with an enantiomeric excess of 99% using a catalyst with complete enantioselectivity. On the other hand, the enantiomeric excess of the product through reduction of the prochiral ketone is independent of the conversion as long as the biocatalyst is completely enan- tioselective. Asymmetric reduction is thus considered to be an attractive method. The reduction of acetophenone to the @)-isomer of l- PA with a high enantiomeric excess by immobilized H. capsulata cells was investigated in this study. Factors affecting the performance in the yeast-mediated reduc- tion of acetophenone-the concentration of the sub- strate, consumption of the energy source, and operating mode for the reaction-were examined. A shallow-bed reactor of the packed-bed type was adopted for the con- tinuous reduction of acetophenone. MATERIALS AND METHODS Materials The yeast H. capsulata IF0 0974 was obtained from the Institute for Fermentation, Osaka. The cells were stored at 4°C on a 2%-agarose slant of YPD medium (1% (w/v) yeast extract, 2% Polypepton, 2% D-glucose, pH unadjusted). All the chemicals used were of analytical grade and were purchased from either Wako Pure Chemical Industries (Osaka) or Nacalai Tesque (Kyoto) . Preparation of immobilized cells The methods for harvesting cells and immobilizing them in calcium- alginate gel beads were the same as previously reported (12). A seed culture (3.5 ml YPD medium) of the yeast precultivated at 30°C with shaking at 120rpm for 24 h was inoculated into a SOO-ml Sakaguchi culture flask con- taining 150ml YPD medium. The flask was then incubat- ed under the same conditions as the preculture. Cells harvested by centrifugation were rinsed with 0.85% saline, immobilized in 1% Ca-alginate gel beads, and then stored at 4°C until use. In all the experiments described below, CaC12 was added at a concentration of 0.01 mol 322
-
Upload
yuri-hasegawa -
Category
Documents
-
view
213 -
download
0
Transcript of Asymmetric reduction of acetophenone by immobilized Hansenula capsulata cells

JOURNAL OF FERMENTATION AND BIOENGINEERING Vol. 85, No. 3, 322-327. 1998
Asymmetric Reduction of Acetophenone by Immobilized Hansenula capsulata Cells
YURI HASEGAWA, SHUJI ADACHI,* AND RYUICHI MATSUNO
Division of Applied Life Sciences, Graduate School of Agriculture, Kyoto University, Sakyo-ku, Kyoto 606-01, Japan
Received 30 October 1997/Accepted 15 December 1997
Immobilized cells of the yeast Hansenula capsulata IF0 0974 could catalyze the asymmetric reduction of acetophenone to Q-1-phenylethanol with almost complete enantioselectivity using glucose as an energy source. Depending on the operational conditions, 1 to 3 mol glucose were required for the reduction of 1 mol of acetophenone. A packed-bed reactor gave a higher conversion than a batch reactor because of its anaerobic conditions and the fact that reverse oxidation was prevented in the former reactor. However, the formation of CO2 bubbles made continuous operation difficult. A shallow-bed reactor, from which CO, bubbles could be easily removed, improved the conversion to over 90%. Continuous production of Q-1-phenylethanol was realized at a significantly high enantiomerlc excess for 220 h by incorporating cell re-cultivation.
[Key words: asymmetric reduction, I-phenylethanol, Hansenula capsulata, shallow-bed reactor]
Optically active compounds are abundant in our sur- roundings -as flavors, sugars, amino acids, and pro- teins in our daily foods, medicines, pesticides, liquid crystals, and so on. During the synthesis of optically active secondary alcohols as synthons for the synthesis of compounds with chiral center(s), lipase and esterase have been widely used (l-4) due to their high stability and the fact that no cofactor is needed (5). Oxidoreductases, which directly catalyze the formation of the chiral hy- droxyl group from the achiral ketone group instead of it being indirectly formed through stereoselective hydrol- ysis of the alcohol-fatty acid ester, as in the case of lipase, have not yet been adopted for industrial use because of difficulties in cofactor NAD(P)H/NAD(P)+ regeneration and poor cofactor stability in solution (6). For the practical use of oxidoreductases, a cofactor regeneration system that will allow effective use of the expensive cofactor, and hence the economical production of optically active compounds on a larger scale, is required (7, 8). One way to achieve this is to use whole cells as the catalyst. An important advantage of using whole cells is that all of the reaction system for cofactor regeneration is present within the catalytic cells them- selves (9, 10). Other advantages are that purification of the enzyme is not required, simple and cheap com- pounds such as glucose or ethanol can be adopted as an energy source for cofactor regeneration, and the self- replicating feature of viable microorganisms is utilized when whole cells are used.
We have previously reported the production of optically active 1-phenylethanol (l-PA) through enantioselective oxidation of its racemate with yeast cells (11, 12). Two yeast strains, Pachysolen tannophilus IF0 1007 and Han- senula capsulata IF0 0974, were proved to have com- plete enantioselectivity for conversion of the @)-isomer of l-PA to the corresponding ketone, acetophenone. Using cells of these yeasts, the continuous production of (R)-l-PA was achieved in a fluidized-bed bioreactor. One difficulty in obtaining the residual isomer through stereoselective oxidation of the other isomer arises from the fact that the enantiomeric excess of the residual
* Corresponding author.
isomer is largely dependent on the conversion of the other isomer (13). According to the definition of enantio- merit excess, a conversion of 49.8%, based on the race- mate, must be accomplished in order to obtain a product with an enantiomeric excess of 99% using a catalyst with complete enantioselectivity. On the other hand, the enantiomeric excess of the product through reduction of the prochiral ketone is independent of the conversion as long as the biocatalyst is completely enan- tioselective. Asymmetric reduction is thus considered to be an attractive method.
The reduction of acetophenone to the @)-isomer of l- PA with a high enantiomeric excess by immobilized H. capsulata cells was investigated in this study. Factors affecting the performance in the yeast-mediated reduc- tion of acetophenone-the concentration of the sub- strate, consumption of the energy source, and operating mode for the reaction-were examined. A shallow-bed reactor of the packed-bed type was adopted for the con- tinuous reduction of acetophenone.
MATERIALS AND METHODS
Materials The yeast H. capsulata IF0 0974 was obtained from the Institute for Fermentation, Osaka. The cells were stored at 4°C on a 2%-agarose slant of YPD medium (1% (w/v) yeast extract, 2% Polypepton, 2% D-glucose, pH unadjusted). All the chemicals used were of analytical grade and were purchased from either Wako Pure Chemical Industries (Osaka) or Nacalai Tesque (Kyoto) .
Preparation of immobilized cells The methods for harvesting cells and immobilizing them in calcium- alginate gel beads were the same as previously reported (12). A seed culture (3.5 ml YPD medium) of the yeast precultivated at 30°C with shaking at 120rpm for 24 h was inoculated into a SOO-ml Sakaguchi culture flask con- taining 150ml YPD medium. The flask was then incubat- ed under the same conditions as the preculture. Cells harvested by centrifugation were rinsed with 0.85% saline, immobilized in 1% Ca-alginate gel beads, and then stored at 4°C until use. In all the experiments described below, CaC12 was added at a concentration of 0.01 mol
322

VOL. 85, 1998 ASYMMETRIC REDUCTION OF ACETOPHENONE 323
1-r to the substrate solution in order to maintain the mechanical strength of the gel beads.
Batch reactions The effects of the concentrations of substrate and energy source on the initial reaction rate were examined using batch reactors. Unless other- wise specified, 0.5 g immobilized yeast and 5 ml substrate solution were put into a IOO-ml glass bottle and incubat- ed at 30°C in a water-bath shaker (EX-11; Taitech, Tokyo) at 150 rpm. A portion of the broth (0.3 ml) was removed from the bottle at intervals and used for the analysis.
The effect of the substrate concentration was observed by varying the initial acetophenone concentration bet- ween 10 and 40mmol I-‘. The molar ratio of glucose to acetophenone was fixed at 1 : 1 in each run. The effect of the molar ratio of the energy source (glucose) to substrate was examined at a fixed acetophenone concentra- tion (10 or 20mmol l-r), the molar ratio being varied between 1 : 1 and 10 : 1. In addition to the above batch reactions, reduction under relatively anaerobic condi- tions was also tested by filling the headspace of the batch reactor with nitrogen gas.
Comparison of packed-bed and batch reactors Con- tinuous reduction was carried out in a packed-bed reactor and the conversion obtained at a variety of feed flow rates was compared with that observed in a batch reactor using the design equations for a plug-flow reac- tor (PFR) and a batch reactor, assuming that the liquid flow in the packed-bed reactor was a plug flow. The design equation for the PFR is known to be identical to the one for a batch reactor.
Under the assumption that the effectiveness factor of immobilized yeast cells is unity, the design equation for a batch reactor is
where k’ is the volume of the reaction mixture, Cm is the initial concentration of the substrate, V, is the volume of the immobilized yeast gel beads, r is the reaction rate based on unit gel volume, and t is the time. The conver- sion xs is defined as follows:
Xs= Go - cs
Go where Cs is the substrate concentration at any given time.
The design equation for a PFR is expressed as fol- lows:
(3)
where Z is the bed height, u. is the superficial velocity, &b is the bed voidage, and Cso is the substrate concentra- tion at the inlet. Since the right-hand term of Eq. 1 is the same as that of Eq. 3, V,t/V in a batch reactor corresponds to (1 - &~,)Z/U~ in a PFR.
The packed-bed reactor equipment used in this experi- ment was configured and operated as follows. The im- mobilized yeast gel beads were packed into a glass column of 1.4 cm I.D. and 10 cm height. The feed solu- tion containing 20 mmol 1-l acetophenone, 20 mmol I-’ glucose, and 0.01 mol I-’ CaClz (pH unadjusted) was pumped upward into the column at various flow rates. The reactor was kept at 30°C by circulating thermo-
-7 30 mm
FIG. 1. Dimensions of the shallow-bed reactor.
regulated water through the jacket of the column. At the same time, the batch reaction was carried out using 10 ml substrate solution and 0.5 g (wet weight) immobi- lized yeast gel beads.
Continuous operation by shallow-bed reactor The dimensions of the shallow-bed reactor, which was made of acrylic resin, are given in Fig. 1. Using this reactor, the relationship between the conversion and the mean residence time was examined under various conditions. (S)-I-PA was continuously produced with cell re-cultiva- tion under the following operating conditions: 20 mmol I- 1 acetophenone and 40 mmol 1-l glucose dissolved in 0.1 mol I-’ [2-(N-morpholino)ethanesulfonic acid]-Na (MES-Na) buffer, pH 7.0, containing 0.01 mol 1-l CaC12, a 50ml reactor volume, 48 g immobilized yeast gel beads, and a 0.34ml min-’ feed flow rate, correspond- ing to a mean residence time of 147 min. The re-cultiva- tion procedure adopted in this experiment was as follows (12): the continuous reaction was stopped when the con- version was less than half of the initial value, and the beads were removed from the reactor. They were then washed three times with 0.01 mol I-’ sterile CaC&, and transferred into two 500-ml Erlenmeyer flasks, each con- taining 200 ml of the YPD medium with 0.01 mol 1-l CaQ. The flasks were incubated in a water bath shaker at 30°C under shaking at 120 rpm for 27 h, after which the contents were recovered and washed with 0.01 mol I-’ sterile CaC&. The re-cultivated beads were then packed into the reactor again and continuous reaction was carried out under the same conditions as in the previous operation.
Analysis The acetophenone and l-PA concentra- tions were measured by the same procedure as previously described (12) using a high-performance liquid chromato- graph (L-6000; Hitachi, Tokyo) equipped with an ODS column (Cosmosil 5C18, 4.6 mm4 x 150 mm; Nacalai Tesque). Benzoic acid (5 mmol I- *) was mixed with the same volume of a sample as the internal standard. A mixture of 0.01 mol 1-l MES-Na buffer (pH 7.0) and acetonitrile (50 : 50 (v/v)) was used as the eluent at a flow rate of 1 ml min’. The absorbance of the effluent at 220nm was monitored by a UV detector (Yanaco M- 315; Yanagimoto Mfg., Kyoto).
Assuming no formation of by-products, the conver- sion was calculated based on the following equation:

324 HASEGAWA ET AL.
Cl-PA conversion [%I = C, .-PA+ CA, x 100 (4)
where C, _ PA and CAP are the concentrations of l-PA and acetophenone, respectively.
A sample for the measurement of the enantiomeric excess of l-PA was prepared by extracting 0.1 ml of the reaction medium with the same volume of n-hexane. The hexane phase was applied to the chromatograph using an enantioseparating column, Chiralcel OB (4.6 mm$ x 250 mm; Daicel Chemical Industries, Tokyo). The eluent used was a mixture of n-hexane and 2-propanol (90: 10 (v/v)) at a flow rate of 0.5 ml min-I. The absorbance at 220nm was monitored and recorded; the retention times of (s)- and @)-l-PA were about 10 and 12min, respectively.
The concentration of n-glucose was determined by the glucose oxidase/peroxidase method using the Glucose C- II Test Wako (Wako Pure Chemical Industries).
.J. FERMENT. BIOENG.,
RESULTS AND DISCUSSION
Effect of substrate concentration Figure 2 shows the asymmetric reductions in a batch reaction at different initial substrate concentrations with a fixed energy source to substrate (glucose : acetophenone= 1 : 1) molar ratio. Maximum progress was observed at 30mmol I-* acetophenone and glucose; at an initial concentration of 40mmol I-‘, the reaction rate decelerated compared to the reduction at 30mmol 1-i. Figure 3 illustrates the relationship between l-PA produced and glucose con- sumed by the yeast cells during the reaction. When the concentrations of the substrate and energy source were both 30 mmol I-l, more l-PA was produced per mole of glucose consumed (ACp/( - AC,) = ca. 1.4 mol mall l) than in 20mmol I-’ (A&/(-ACo)=ca. l.Omol mol-*). The conversion of energy from glucose into the asym- metric reduction of acetophenone was greater at an initial concentration of 30 mm01 1-l.
Effect of molar ratio of energy source to acetopbenone Asymmetric reduction of acetophenone to l-PA requires an energy source for regeneration of the cofactor NAD(P)H. In a preliminary study on the asymmetric reduction of secondary alcohols by H. capsulata, we employed ethanol and glucose as energy sources; because
IO, I I I I
2 4
Time @]
FIG. 2. Effect of substrate concentration on production of l- phenylethanol (l-PA). The molar ratio of glucose to acetophenone was 1 : 1 in each run. Symbols: 0, A, 0, and 0, indicate initial concentrations of 10, 20, 30, and 40 mmol I-r, respectively.
k I I
U 5 10 I5
-ACo[mmol 1-11
FIG. 3. Relationship between l-PA produced (AC,) and glucose consumed (-AC,) in the experiment shown in Fig. 2. Symbols: 0 and A, indicate initial substrate concentrations of 20 and 30mmol I-‘, respectively.
the efficiency of ethanol in cofactor regeneration was found to be not as high as that of glucose, we used glucose as the energy source throughout this study. The optimum ratio of glucose to acetophenone was examined using batch reactors. Figure 4 shows the effect of the molar ratio on the time course of l-PA production. At lower molar ratios, the concentration of l-PA initially increased as the relative concentration of acetophenone was reduced; however, the increases ceased as the reac- tion continued, and then turned into gradual decreases. l-PA eventually completely disappeared from the reac- tion medium at molar ratios of 0.5 : 1 or less. Exhaus- tion of the energy source seemed to reverse the direction of the reduction reaction, resulting in the oxidation of l- PA into acetophenone. This implied that the reaction sys- tem would be catalyzed by an NADH/NAD+-dependent oxidoreductase rather than by an NADPH/NADP+- dependent one. In contrast, higher molar ratios (2 : 1 or more) resulted in very similar initial reaction rates, though at a ratio of 10 : 1 the rate decreased slightly.
0 2 Time @:
6
FIG. 4. Effect of molar ratio of glucose to acetophenone on production of l-PA. This experiment was conducted in batch reactors without filling with Nz gas. The concentration of acetophenone was fixedat lOmmolI-r. Symbols: 0, A, q , 0, 0, A, m, and +, indicate glucose/acetophenone molar ratios of 0.1, 0.2, 0.5, 1, 2, 3, 5, and 10mol mol-r, respectively.
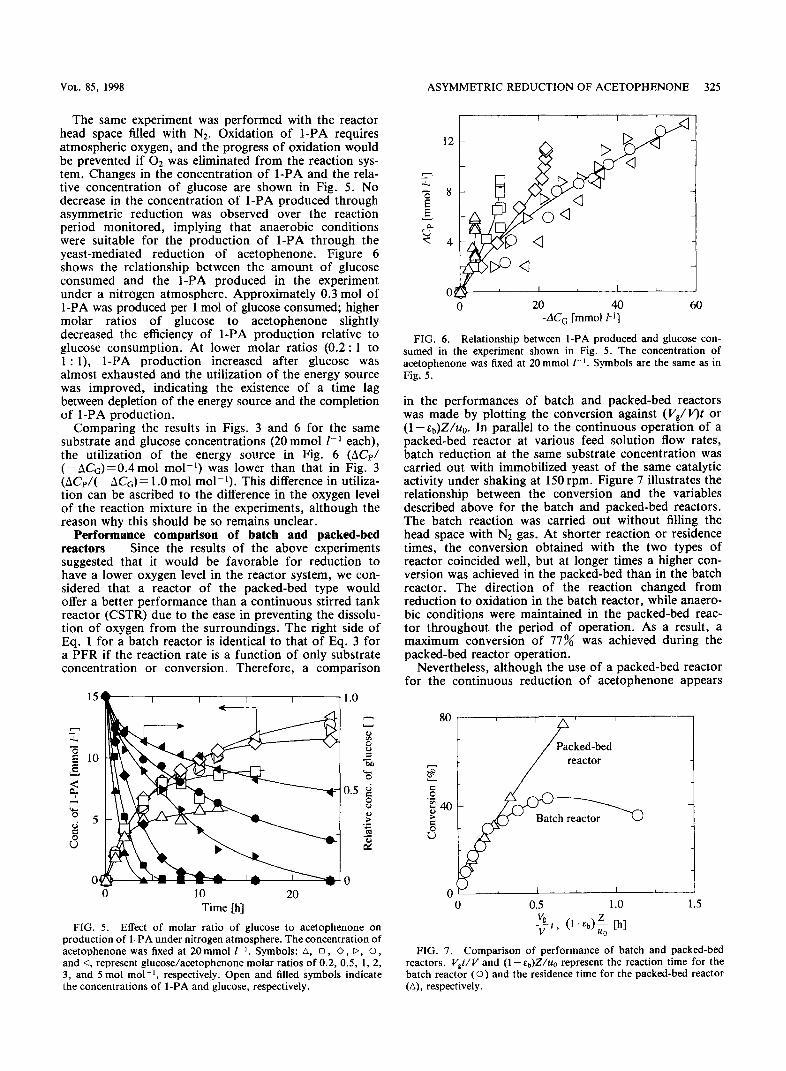
VOL. 85, 1998 ASYMMETRIC REDUCTION OF ACETOPHENONE 325
The same experiment was performed with the reactor head space filled with N2. Oxidation of l-PA requires atmospheric oxygen, and the progress of oxidation would be prevented if O2 was eliminated from the reaction sys- tem. Changes in the concentration of l-PA and the rela- tive concentration of glucose are shown in Fig. 5. No decrease in the concentration of l-PA produced through asymmetric reduction was observed over the reaction period monitored, implying that anaerobic conditions were suitable for the production of l-PA through the yeast-mediated reduction of acetophenone. Figure 6 shows the relationship between the amount of glucose consumed and the l-PA produced in the experiment under a nitrogen atmosphere. Approximately 0.3 mol of l-PA was produced per 1 mol of glucose consumed; higher molar ratios of glucose to acetophenone slightly decreased the efficiency of l-PA production relative to glucose consumption. At lower molar ratios (0.2 : 1 to 1 : l), l-PA production increased after glucose was almost exhausted and the utilization of the energy source was improved, indicating the existence of a time lag between depletion of the energy source and the completion of l-PA production.
8
4
00 0 20 40 60
-ACG [mmol L-11
FIG. 6. Relationship between l-PA produced and glucose con- sumed in the experiment shown in Fig. 5. The concentration of acetophenone was fixed at 20 mmol I-r. Symbols are the same as in Fig. 5.
Comparing the results in Figs. 3 and 6 for the same substrate and glucose concentrations (20 mmol 1-l each), the utilization of the energy source in Fig. 6 (ACp/ (-A&)=0.4 mol mol-l) was lower than that in Fig. 3 (ACr/( -AC,)= 1.0 mol mol-l). This difference in utiliza- tion can be ascribed to the difference in the oxygen level of the reaction mixture in the experiments, although the reason why this should be so remains unclear.
Performance comparison of batch and packed-bed reactors Since the results of the above experiments suggested that it would be favorable for reduction to have a lower oxygen level in the reactor system, we con- sidered that a reactor of the packed-bed type would offer a better performance than a continuous stirred tank reactor (CSTR) due to the ease in preventing the dissolu- tion of oxygen from the surroundings. The right side of Eq. 1 for a batch reactor is identical to that of Eq. 3 for a PFR if the reaction rate is a function of only substrate concentration or conversion. Therefore, a comparison
in the performances of batch and packed-bed reactors was made by plotting the conversion against (V./V)t or (1 - Q,)Z/U~. In parallel to the continuous operation of a packed-bed reactor at various feed solution flow rates, batch reduction at the same substrate concentration was carried out with immobilized yeast of the same catalytic activity under shaking at 150 rpm. Figure 7 illustrates the relationship between the conversion and the variables described above for the batch and packed-bed reactors. The batch reaction was carried out without filling the head space with N2 gas. At shorter reaction or residence times, the conversion obtained with the two types of reactor coincided well, but at longer times a higher con- version was achieved in the packed-bed than in the batch reactor. The direction of the reaction changed from reduction to oxidation in the batch reactor, while anaero- bic conditions were maintained in the packed-bed reac- tor throughout the period of operation. As a result, a maximum conversion of 77% was achieved during the packed-bed reactor operation.
Nevertheless, although the use of a packed-bed reactor for the continuous reduction of acetophenone appears
15 1.0
80
Batch reactor
0 0 0 10 20
Time [h] 0
0 0.5 1.0 1.5
FIG. 5. Effect of molar ratio of glucose to acetophenone on production of I-PA under nitrogen atmosphere. The concentration of acetophenone was fixed at 20mmol I-i. Symbols: a, 0, 0, D, 0, and a, represent glucose/acetophenone molar ratios of 0.2, 0.5, 1, 2, 3, and 5 mol mol-I, respectively. Open and filled symbols indicate the concentrations of l-PA and glucose, respectively.
FIG. 7. Comparison of performance of batch and packed-bed reactors. V&V and (1 -cb)Z/uO represent the reaction time for the batch reactor (0) and the residence time for the packed-bed reactor (A), respectively.

326 HASEGAWA ET AL. J. FERMENT. BIOENG.,
I I I
0 50 100 150 200
Normalized t [min]
FIG. 8. Relationship between conversion and mean residence time in the shallow-bed reactor. Symbols: 0, A, 0, 0, and v, represent the lst, 2nd, 3rd, 4th, and 5th runs, respectively. The con- centration of acetophenone was 20 mmol 1-l in all runs. The molar ratio of glucose to acetophenone was 1 : 1 in 1st and 2nd runs, and 2 : 1 in other runs. As solvents, 0.01 mol I-’ CaClz was used in the 1st to 3rd runs, and 0.1 mol I-’ MES-Na buffer (pH 7.0) containing 0.01 mol I-’ CaCl* in the 4th and 5th runs.
promising, a problem arose resulting from the genera- tion of carbon dioxide during the metabolism of glucose by the cells: the COZ bubbles formed were retained in the column, accompanied by channelling of the liquid in the reactor, thereby making continuous operation difficult. A means of releasing the bubbles from the column must be devised for the packed-bed reactor to be utilized effectively.
Relationship between conversion and mean residence time in shallow-bed reactor As an alternative, the shallow-bed reactor illustrated in Fig. 1 was used. The short longitudinal distance and slight upward slope of the top facilitated the release of CO2 bubbles from the reactor. The relationship between the conversion and mean residence time of the fluid was obtained at differ- ent feed flow rates using this shallow-bed reactor. Because the specific activity of the immobilized yeast gel beads differed during each immobilization, the mean resi- dence time normalized by dividing the ratio of the initial activity of any immobilized yeast to the immobilized yeast of the lowest activity was adopted as the abscissa in Fig. 8. The initial activities were measured in a batch reactor. Generally, the longer the mean residence time, the higher was the conversion. However, in the first three runs, the conversion leveled off at about 67%, even though the mean residence time was further increased. In the first and second runs, glucose was depleted at the lower feed flow rate, and hence at the higher mean resi- dence times the molar ratio of the energy source to acetophenone in the feed solution was 1 : 1. In the third run, with twice the moles of glucose, residual glucose was eluted from the reactor at a concentration of about 7 mmol 1-l or more throughout the operation (data not shown); exhaustion of glucose thus appeared not to be the main cause of the leveling off of the conversion. Since the solvent used as the feed solution in these runs was 0.01 mol 1-l CaC12 and no adjustment of the pH was made, it was considered that the pH value of the reaction medium might have been significantly lowered by the CO* generation to a level where the cells lost their
Re-cultivatmn
Time [h]
FIG. 9. Time courses during continuous operation of the shal- low-bed reactor. Immobilized yeast was re-cultivated at 120 h. Sym- bols: 0, A, and 0, represent conversion, concentration of glucose, and enantiomeric excess of l-PA, respectively.
catalytic activity. Therefore, fourth and fifth runs were conducted with a buffer of 0.1 mol I-’ MES-Na, pH 7.0, containing 0.01 mol 1-l CaCl* to minimized or eliminate the effect of pH change during operation. In the fifth run, a maximum conversion of 89% was achieved with this MES-Na buffer.
Continuous production of (S&l-PA in shallow-bed reactor (&l-PA was continuously produced in the shallow-bed reactor under operating conditions designed to achieve about 90% conversion of acetophenone to @)-l-PA using 20mmol I-’ acetophenone. As shown in Fig. 9, a conversion of about 90% was realized and maintained during the first stage of the operation. However, the conversion gradually decreased by l-2% every half-day, and the rate of decrease became steeper after three days; the conversion had dropped to about a half of the initial value at 120 h, accompanied by the elution of residual glucose into the reactor effluent. This suggested a lowering of the glucose uptake due to deacti- vation of the immobilized cells. The cells were then re- cultivated by simply suspending them in the medium con- taining nutrients, as a result of which they regained their catalytic activity. In the second run following re-cultiva- tion, the conversion recovered to 80% for the first two days, but after 170 h, a relatively rapid decrease oc- curred, and the conversion was only 57.4% at 224 h. Decreased glucose uptake was observed earlier in the second run than in the first. The amounts of glucose con- sumed were different even for the same conversion in the two runs; an excess of glucose was eluted at a concen- tration higher than 10mmol 1-l at 80% conversion of acetophenone in the second run, while no glucose was measured at the exit of the reactor in the first run, even after the conversion decreased to 78%. The reason for this is still unknown. The enantiomeric excess of I-PA produced was complete for the @‘)-isomer over the entire operation period, at least under the conditions of our HPLC analysis. Even after deactivation of the catalytic cells and the conversion were reduced, the optically active product without contamination by another isomer was obtained. These results thus suggested that a shal- low-bed reactor is a good option for reaction systems

VOL. 85, 1998 ASYMMETRIC REDUCTION OF ACETOPHENONE 327
in which the formation of carbon dioxide bubbles con- stitute an inherent problem.
7.
8.
applications of NAD(P)-dependent oxidoreductases in synthe- sis: a survey. Enzyme Microb. Technol., 20, 248-258 (1997). I&e, S. and Hayashida, M.: Two-phase system membrane reac- tor with cofactor recycling. J. Biotechnol., 14, 221-228 (1990). Kragl, U., Kruse, W., Hummel, W., and Wandrey, C.: En- zyme engineering aspects of biocatalysis: cofactor regeneration as example. Biotechnol. Bioeng., 52, 309-319 (1996). Sih, C. J. and Chen, C.-S.: Microbial asymmetric catalysis- enantioselective reduction of ketones. Angew. Chem. Int. Ed. Engl., 23, 570-578 (1984). Kometani, T., Yoshii, H., and Matsuno, R.: Large-scale production of chiral alcohols with baker’s yeast. J. Mol. Catal. B: Enzymatic, 1, 45-52 (1996). Hasegawa, Y., Uosaki, H., Adachi, S., and Matsuno, R.: Production of (R)-1-phenylethanol through enantioselective oxi- dation of @)-isomer from their racemic mixture by Puchysolen funnophilus IF0 1007. Biotechnol. Lett., 18, 367-372 (1996). Hasegawa, Y., Ada&i, S., and Matsuno, R.: Production of homochiral I-phenylethanol through enantioselective oxidation of its racemate with whole cells of the yeast Hunsenula cupsu- latu IF0 0974. _I. Ferment. Bioeng., 83, 346-351 (1997). Tsuji, Y., Fukui, T., Kawamoto, T., and Tanaka, A.: Enan- tioselective dehydrogenation of fi-hydroxysilanes by horse liver alcohol dehydrogenase with a novel in-situ NAD- regeneration system. Appl. Microbial. Biotechnol., 41, 219-224 (1994).
1.
2.
3.
4.
5.
Whitesides, G. M. and Wang, C.-H.: Enzymes as catalysts in synthetic organic chemistry. Angew. Chem. Int. Ed. Engl., 24, 617-638 (1985). Naemura, K., Takahashi, N., Tanaka, S., and Ida, H.: Resolu- tion of the diols of bicyclo[2.2. llheptane, bicyclo[2.2.2]octane and bicyclo[3.2.l]octane by enzymic hydrolysis, and their abso- lute configurations. .I. Chem. Sot. Perkin. Trans., 1, 2337-2343 (1992). Ohrner, N., Orrenius, C., Mattson, A., Norin, T., and Hult, K.: Kinetic resolutions of amine and thiol analogues of sec- ondary alcohols catalyzed by the Cundida antarctica lipase B. Enzyme Microb. Technol., 19, 328-331 (1996). Capewell, A., Wendel, V., Bomscheuer, U., Meyer, H. H., and Scheper, T.: Lipase catalyzed kinetic resolution of 3- hydroxy esters in organic solvents and supercritical carbon dioxide. Enzyme Microb. Technol., 19, 181-186 (1996). Bjorkling, F., Godtfredsen, S. E., and Kirk, 0.: The future impact of industrial lipases. Trends Biotechnol., 9, 360-363 (1991).
6. Devaux-Basseguy, R., Bergel, A., and Comtat, M.: Potential
REFERENCES
9.
10.
11.
12.
13.


















