assay of tissue glycogen pre
6
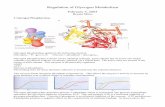
Name: K***** K*** ID#: ********* Lab Partner: V******* S****** Date: Wednesday 22 nd September, 2010 Course Code: BIOL 2363 – Metabolism Title of Lab: Assay of Tissue Glycogen Aim: To compare the levels of glucose present in the liver of a fasted animal with that of a fed animal as well as to compare the glucose levels of either the heart, muscle or kidney of either a fed or fasted animal with the glucose level present in the liver. Theory: Glycogen is the main storage polysaccharide of animal cells. It is a polymer of (α1 → 4)-linked glucose subunits with (α1 → 6)- linked branches. Glycogen is extensively branched and highly compact (Nelson and Cox 2008) (King 2010) (King 2010) After a meal, glucose levels in the blood rise and liver cells link the excess glucose molecules into long, branching chains of glycogen via condensation reactions. When blood glucose levels fall, the liver cells hydrolyse the glycogen to produce single glucose molecules which can be released into the bloodstream. This glucose becomes available to supply energy to the body (Whitney and Rolfes 2008). Fig.2: Structure showing the many branches of glycogen. Fig. 1: Structure showing the types of linkages present in glycogen
-
Upload
karina-khan -
Category
Documents
-
view
326 -
download
2
Transcript of assay of tissue glycogen pre

Name: K***** K***
ID#: *********
Lab Partner: V******* S******
Date: Wednesday 22nd September, 2010
Course Code: BIOL 2363 – Metabolism
Title of Lab: Assay of Tissue Glycogen
Aim: To compare the levels of glucose present in the liver of a fasted animal with that of a fed animal as well as to compare the glucose levels of either the heart, muscle or kidney of either a fed or fasted animal with the glucose level present in the liver.
Theory: Glycogen is the main storage polysaccharide of animal cells. It is a polymer of ( 1 α → 4)-linked glucose subunits with ( 1α → 6)-linked branches. Glycogen is extensively branched and highly compact (Nelson and Cox 2008)
(King 2010) (King 2010)
After a meal, glucose levels in the blood rise and liver cells link the excess glucose molecules into long, branching chains of glycogen via condensation reactions. When blood glucose levels fall, the liver cells hydrolyse the glycogen to produce single glucose molecules which can be released into the bloodstream. This glucose becomes available to supply energy to the body (Whitney and Rolfes 2008).
Glycogen synthesis involves the hormone insulin. When blood glucose levels are too high, insulin is released into the blood stream which stimulates the synthesis of glycogen. The enzymes UDP-glucose pyrophosphorylase, Glycogen synthase and Branching enzyme (amylo-(1-4→1-6) transglycosylase) are involved in glycogen synthesis (Nelson and Cox 2008).
UDP- glucose pyrophosphorylase catalyses the formation of UDP-glucose from UTP and glucose-1-phosphate:
UTP + glucose-1-phosphate → UDP-glucose + PPi
Glycogen synthase transfers the glucosyl residue from UDP-glucose to the C4 OH group at the non-reducing end of a glycogen molecule forming an ( 1 α → 4) glycosidic bond.
Fig.2: Structure showing the many branches of glycogen.
Fig. 1: Structure showing the types of linkages present in glycogen

After a number of glucose units have been joined as a straight chain with ( 1 α → 4) linkages, the branching enzyme breaks one of the 1 α → 4 bonds and transfers a block of residues to another site in the glycogen molecule, reattaching them by an 1α → 6 bond. (Hames and Hooper 2005).
Glycogen degradation involves the hormones epinephrine and glucagon. When blood glucose levels fall glucagon is secreted by cells of the pancreas which act on the liver to stimulate glycogen breakdown to elevate the blood glucose levels. Muscular contraction causes the release of epinephrine which acts on the muscle to increase glycogen breakdown for a ready supply of energy to the cells (Hames and Hooper 2005).
The enzymes glycogen phosphorylase and glycogen-debranching are concerned with glycogen degradation (Nelson and Cox 2008).
Phosphorylase degrades glycogen by catalysing the phosphorolysis reaction where 1α → 4 glycosidic bonds are broken to release glucose units one at a time from the non-reducing end of the glycogen molecule.
Glycogen + Pi ⇆ glycogen + glucose-1-phosphate (n residues) (n-1 residues)
Glycogen- debranching enzyme removes 1α → 6 branches and allows phosphorylase to continue degrading the glycogen molecule (Hames and Hooper 2005).
Blood glucose level can be determined using the glucose oxidase-peroxidase assay. Glucose oxidase catalyses the oxidation of -D-glucose to D-glucono- -lactone with the β δsimultaneous release of hydrogen peroxide. In thepresence of peroxidase this hydrogen peroxide enters into a second reaction involving p-hydroxybenzoic acid and 4-aminoantipyrine with the quantitative formation of a quinoneimine dye complex which is measured at 510 nm (Megazyme 2004). The amount of dye formed is a measure of glucose oxidized.The reactions involved are:
(Glucose oxidase)(1) β-D-Glucose + O2 + H2O D-glucono-lactone + H2O2
(peroxidase)(2) 2 H2O2 + p-hydroxybenzoic acid + 4-aminoantipyrine quinoneimine dye + 4 H2O(Megazyme 2004)

Procedure:
Materials1. Heart, liver, muscle and kidney from freshly killed rats/guinea pigs (fed and fasted).
2. KOH (300g/L)
3. Saturated Na2SO4
4. ETOH - 95% (v/v)
5. 2.4M HCl
6. 1.0M NaOH
7. Stock glucose solution (0.2 µmol/mL)
8. Dissecting instruments
9. Reagents for the estimation of glucose:Glucose oxidase 12.5 mgPeroxidase 4.0 mgo-dianisidine HCl - 1% (w/v) in EtOH 0.5 mLDissolve all of the above in:0.5M sodium phosphate buffer, pH 7.2 100mL
Two millilitres of fed liver, fasted liver, and were collected in small test tubes and labelled and placed in an ice bucket. 0.2mL of saturated Na2SO4 was added to each tube and vortexed. 5mL of ethanol was then added to all the tubes and left to stand for five minutes. The precipitated glycogen was pelleted by centrifuging the tubes at 3000rpm for 10 minutes. The supernatant liquid was then poured off and discarded. 5mL of distilled water was added to the pellet to dissolve and the tubes were gently warmed in a water bath at 37°C.
Tube containing glycogen from the liver of a fasted animal: The dissolved glycogen was poured into a 10mL measuring cylinder and the test tube was rinsed with 2mL of distilled water. All washings were poured into the measuring cylinder. The volume of solution in the measuring cylinder was brought up to the 10mL mark by distilled water. The contents of the tube were thoroughly mixed.
Tube containing glycogen from the liver of a fed animal: The dissolved glycogen was poured into a 100mL measuring cylinder and the test tube was rinsed with 2mL of distilled water. All washings were poured into the measuring cylinder. The volume of solution in the measuring cylinder was brought up to the 100mL mark with distilled water. The contents of the tube were thoroughly mixed.

Tubes containing glycogen from other tissues of both fed and fasted animals: Each tube of dissolved glycogen was poured into a 25mL measuring cylinder and the test tube was rinsed with 8mL of distilled water. All washings were poured into the measuring cylinders. The volumes of the solutions in the measuring cylinders were brought up to the 25mL mark with distilled water and both were thoroughly mixed.
1mL aliquots of prepared glycogen from the measuring cylinders were pipetted into separate labelled test tubes twice and 1mL of 2.4M HCl was added to each tube which was then thoroughly mixed. The tubes were covered with foil and heated in a boiling water bath for one hour.
After boiling, the solutions were neutralised by adding mL of 1M NaOH. The samples were then diluted to 5mL using distilled water. 0.3 and 0.5mL of liver/muscle from fed/fasted animals were pipetted into separate test tubes and 0.5 and 1.0mL of heart/kidney samples were pipette into other tubes. Distilled water was added to ensure that the volume of each tube was 1.0mL. 2.5mL of enzyme cocktail was then added to each tube and incubated at 37°C for 45minutes. The tubes were cooled to room temperature and the absorbance read at 450nm.
Calibration Curve: Seven test tubes were labelled 1-7 and stock glucose solution was added to the test tubes in the following volumes, 0.0mL, 0.2mL, 0.2mL, 0.4mL, 0.6mL, 0.6mL and 1.0mL respectively. The volume of each tube was made up to 1.0mL using distilled water where necessary. 2.5mL of enzyme cocktail was added to each tube and incubated at 37°C for 45 minutes. The tubes were then cooled to room temperature and the absorbance read at 450nm. A calibration curve was then plotted for glucose.

References
Hames, David, and Nigel Hooper. Instant Notes Biochemistry 3/e. New York: Taylor & Francis Group, 2005.
King, Michael W. The Medical Biochemistry Page. March 24, 2010. http://themedicalbiochemistrypage.org/carbohydrates.html (accessed September 21, 2010).
Megazyme. Glucose Oxidase Assay Procedure. Ireland, 2004.
Nelson, David L, and Michael M Cox. Lehninger Principles of Biochemistry. United States of America: W.H. Freeman and Company, 2008.
Whitney, Elle, and Sharon Rady Rolfes. Understanding Nutrition 11th Edition. California: Wadsworth Publishing, 2008.