
Aspects of Succession in the Coniferous Forests of the...
18
Chapter 14 Aspects of Succession in the Coniferous Forests of the Pacific Northwest Jerry F. Franklin and Miles A. Hemstrom Introduction Many forest phenomena achieve ultimate expression in the coniferous forests of coastal northwestern America. Coniferous species dominance is, in itself, unusual in a moist temperate region (Waring and Franklin 1979). Nearly every arboreal genus represented attains a size and life span at or near the generic maximum. Except for some Eucalyptus forests in Australia, biomass accumulations and leaf areas routinely exceed levels encountered in any other forest type. Accumulations of coarse woody debris are similarly large. Time scales are expanded by species longevity, infrequent disturbances, and relatively slow vegetative regrowth. In this chapter we shall explore aspects of succession in the northwestern forests found from the crest of the Cascade Range to the Pacific Ocean. Emphasis is on newer findin g s since general descriptions of successional sequences are already available (e.g., Franklin and Dyrness 1973). These include the persistent effects of the pioneer species, the variable nature of seres as revealed in age structure analyses, and impor- tance of coarse woody debris. Our objectives are to: (1) provide contrasts with succession in temperate deciduous forests as well as with conifer forests in boreal and arid regions; (2) contrast recent understanding about succession in the Pacific Northwest with previous concepts; (3) examine the role of coarse woody debris in forest succession; and (4) su g gest some problems northwestern forests present in successional modeling. Regional Setting The coastal Pacific Northwest is a mountainous region dominated by the Cascade Range, Olympic Mountains, Coast Ranges, and Siskiyou Moun- tains of Washington and Oregon (Heilman et al. 1979). Topograph y is Franklin, Jerry F.; Hemstrom, Miles A. Aspects of succession in the coniferous forests of the Pacific Northwest. In: West, Darrell C.; Shugart, Herman H.; Botkin, Daniel B., eds. Forest succession: concepts and application. New York: Springer-Verlag; 1981: chapter 14.
Transcript of Aspects of Succession in the Coniferous Forests of the...

Chapter 14
Aspects of Succession in theConiferous Forests of the Pacific
Northwest
Jerry F. Franklin and Miles A. Hemstrom
IntroductionMany forest phenomena achieve ultimate expression in the coniferousforests of coastal northwestern America. Coniferous species dominanceis, in itself, unusual in a moist temperate region (Waring and Franklin1979). Nearly every arboreal genus represented attains a size and life spanat or near the generic maximum. Except for some Eucalyptus forests inAustralia, biomass accumulations and leaf areas routinely exceed levelsencountered in any other forest type. Accumulations of coarse woodydebris are similarly large. Time scales are expanded by species longevity,infrequent disturbances, and relatively slow vegetative regrowth.
In this chapter we shall explore aspects of succession in thenorthwestern forests found from the crest of the Cascade Range to thePacific Ocean. Emphasis is on newer findin gs since general descriptions ofsuccessional sequences are already available (e.g., Franklin and Dyrness1973). These include the persistent effects of the pioneer species, thevariable nature of seres as revealed in age structure analyses, and impor-tance of coarse woody debris. Our objectives are to: (1) provide contrastswith succession in temperate deciduous forests as well as with coniferforests in boreal and arid regions; (2) contrast recent understanding aboutsuccession in the Pacific Northwest with previous concepts; (3) examinethe role of coarse woody debris in forest succession; and (4) su ggest someproblems northwestern forests present in successional modeling.
Regional Setting
The coastal Pacific Northwest is a mountainous region dominated by theCascade Range, Olympic Mountains, Coast Ranges, and Siskiyou Moun-tains of Washington and Oregon (Heilman et al. 1979). Topograph y is
Franklin, Jerry F.; Hemstrom, Miles A. Aspectsof succession in the coniferous forests of thePacific Northwest. In: West, Darrell C.;Shugart, Herman H.; Botkin, Daniel B., eds.Forest succession: concepts and application.New York: Springer-Verlag; 1981: chapter 14.

14: Aspects of Succession in Coniferous Forests of Pacific Northwest
213
pically mountainous and rugged. Gentle intermountain regions, such asPueet Trough and Willamette Valley, are not extensive. Forests ex-
,,:nd from essentially sea level to 1600 m or more. Soils are typicallywuthful but moderately fertile and commonly surprisingly deep due to: 1 ;rositional processes.
The climate of the region favors forest development (Waring andFrankl in 1979). Winters have mild temperatures and heavy precipitation.summe rs are warm and relatively dry. This regime is believed to be a ma-jor factor in the dominance of conifers over deciduous hardwoods. Cli-.mate does vary substantially with geography and elevation. The mildestclima tes are in the coastal Picea sitchensis zone. The coolest environmentsare in the subalpine zones above 800-1200 m elevation, where winterc nowpacks of up to 3 or 4 m may be developed.
S y necological studies in the region have shown that moisture and tem-perature are the primary environmental controls on plant communitycomposition (Zobel et al. 1976). Both vary sharply over short distances inthe broken mountain topography. Consequently, similar contrasts occurin the composition and structure of forest communities.
General Features of the Forests
Forests in the Pacific Northwest differ markedly in composition, struc-ture, and function from the deciduous forests that dominate most mesicsegments of the world's temperate zones. There are relatively few majortree species, and most are conifers (Table 14.1). Productivity can be veryhigh as a record of 36.2 metric tons ha - I yr - I in a 26-year-old Tsugahetcropliylla stand indicates (Fujimori 1972). Nevertheless, forestrecovery following disturbance (as measured by leaf area, for example)t y pically requires several decades, and is substantially slower than decidu-ous forests (Bormann and Likens 1979a). Live above-ground biomassaccumulations in old-growth forests (> 250-year-old) forests average700-900 metric tons/ha, and stands with over 1000 metric tons/ha are notuncommon (Franklin and Waring 1980). Leaf areas (one side only)range from 10 to 13 m 2 /m- in one sample of stands (Franklin and Waring1980). Gholz (1979) reports a maximum value of 20.2 m 2 /m 2 in a Piceasitchensis-Tsuga heterophylla stand.
Several general successional sequences have been described in coastalPacific Northwest forests (Franklin and Dyrness 1973). A typical sere inthe Tsuga heterophylla zone would be
heterophylla Tsuga heterophyllaPseudotsugaThuja plicata

214 J. F. Franklin and M. A. Ilenistrom
In the Picea siwhensis zone a normal sere would involve
Picea sitcheti.5is Tsuga hetcrophyllaTsuga heterophylla
.ft I Abuts rubra stands could conceivably occur as the first stage in eitherof these seres although its successional replacement by conifer stands isstill unproven. A sere for the lower subalpine in the Cascade Range is
Pseudotsuga Inenziesiiand/orAbies procera
Tsuga heteroph.vIlaAbies amabilis
A bies amabilis
Throughout most of the region under consideration Tsuga heteroplollaand Abies amabilis are viewed as the major potential climax species basedupon a variety of evidence, including size class analyses. Special habitatsprovide exceptions, such as the dry sites where Pseudotsuga menziesii orLihocedrus decurrens appear to be self-perpetuating. Some species ofmoderate shade tolerance, such as Thuja plicata and Chamaecyparis floor-katensis, typically reproduce poorly in old-growth stands. This may indi-cate subclimax status, or they may behave similarly to Sequoia semper-virens, which manages a stable age class distribution despite an apparentsparsity of reproduction at any one time (Stephen Viers 1979, pers.comm.).
Most tree species in the Pacific Northwest play multiple successionalroles. In this discussion it is important to distinguish ecological roles fromspecies' shade tolerances. Species colonizing a site early in a sere are play-ing a pioneer role. Species apparently capable of perpetuating themselveson a site in the absence of disturbance are playing a climax role. Althoughshade tolerance is often equated with a climax role and intolerance with apioneer role, this need not be the case. Several intolerant species canform a stable type of climax if environmental conditions
istheir
more tolerant associates. A prime northwestern example is the develop-ment of self-perpetuating stands of Pseudotsuga menziesi on habitats thatare too dry for Tsuga heteroph►lla or Abies grandis. In other cases, en-vironmental conditions need only favor the less tolerant associate, suchas snowpack favoring reproduction of Abies amabilis over that of Tsugahewrophylla (Thornburgh 1969).
Similarly, essentially any of the shade-tolerant species can and do playpioneer roles on most sites where they are also climax. Tsuga heterophyllais conspicuous early in seres on cut-over forest lands in the Picea sitchensiszone. So is Abies amabilis on many high-elevation burns. This can be dueto an absence of seed source for faster-growing intolerant species. Inother cases, the growth rate of the tolerant species may he adequate tostay with or even exceed that of the intolerant species.

14: Aspects of Succession in Coniferous Forests of Pacific Northwest 215
7.; E\O r- C CD vn C, r-O —̂ •.0 e..1
r- r-O C's •.0 M -rrn r,) 0 in CD —
cC
E CV CV CV fV ■ (..) fV t"") C`,1 CN kr, rA
co< C' I ,4.7%kr1 A
CV C)v„11 I ,r) r-
rn A AI -- 'Nr-
c)C
I I rN CCr,A A A
,r1 V-) le.)tntr' rsn CDC' +r- Cr ,r V") 0 V1CD s.0
C.
rJ
-CL)
I I
-r
CD ,n C:'CO— N__.'-' '-' •
,_..,`•••••"'11
r-
0 C. 0kr, oo C,...._, e--,r-... — — ' •• r...1■ I '-' I
I I
c
,e1CV kr,— 0 cs,
. •-•
I
kr,r-
0 0 0,...., ,..,4 ,:.,-- --..s-, CV CN cn r
, . . I I
^
0(-4.-.
'n
(7,..CD.."
c5 v; CDC' r- ,....,
,r1 CD CD i CD,r) eN) C 00CNCD --C
v!,rs-
id, CD CD CD CDr■ V") s. ,S) lrl lrl
-- — — C.+,vl,r-
-4- 4 4- -4- 4- 4- + +++ + +0000000 C occ Coco,n C ,n C 0 0
rn — ,r) kr, s.7.- e-s4 C •zr •cr^ ^ ^
rJ

216 J. F. Franklin and M. A. ticnistrom
Scale in the Pacific Northwest
Many of the unique aspects of coastal Pacific Northwest forests resultfrom an increase in the size and longevity of the tree species in co m pari-son with other temperate and boreal forest regions. Pacific Northwesterntree species are well known for achieving large sizes and long life spans(Table 14.1). Consequently, compositional effects of a disturbance canpersist for centuries or even millenia. The long-term persistence of seralspecies and infrequent catastrophic disturbances often combine to makeachievement of climax stands hypothetical. Classical old-growth stands inthe Pacific Northwest are actually not climax but, rather, draw much oftheir character from large seral Pseudotsuga (Franklin et al. 1981). This isapparent in stands of Pseudotsuga undisturbed for over 1000 years inMount Rainier National Park (Table 14.2). Climax forests, on the otherhand, are almost exclusively dominated by species of smaller stature andshorter life span, such as Tsuga heterophylla and Abies amabilis, and arenot as impressive as those containing the larger, seral species. In forestswith such extended seres the potential for direct observation of succes-sional processes is obviously very limited.
The impact of the large, long-lived pioneer species extends to struc-tural as well as compositional aspects of succession. These trees becomesources of coarse woody debris—snags and down logs—after dying. Thesestructures are important in many aspects of ecosystem function and com-position (Franklin et al. 1980). Pseudotsuga produces relatively decay-resistant logs of large size. Between 480 and 580 years are believed neces-sary to eliminate 90% of an 80-cm diameter Pseudotsuga log (Joseph E.Means 1980, pers. comm.). In forests containing these species the accu-mulations of woody debris are greater and individual pieces are lar ger andmore persistent than would be the case in a climax forest of Tsuga hetero-phylla or Abies amabilis. In this way the influence of a large sera! tree is ex-tended several centuries beyond the death of the last specimen. Whileclimax composition might be achieved soon after the death of the lastPseudotsuga (say, in 1000 years), a stable forest structure based on theclimax species could be delayed another 500 years. If one views Thuja ph-cata as a seral species (albeit one that extends the duration of its per-sistence on a site by some modest reproduction within the stand), manymillenia would be necessary to achieve the idealized climax forest. Livetrees would persist for several generations, and the large Duna logs areeven more resistant to decay than those of Pseudotsuga.
In spite of these problems, the climax concept is useful, and climaxforests do exist. Forests that compositionally and structurally approximatehypothetical climax conditions are present in both temperate and subal-pine regions of the Pacific Northwest. We suspect that these were typi
-cally (but not exclusively) developed where the climax species (e.g.,Tsu,isqz he/crop/0/u or Ables amabilis) was the major or sole colonizing

14: Aspects of Succession in Coniferous Forests of Pacific Northwest 217
Lode 14.2. Pseudotsuga menziesii presence (percentage of plots where itoccurs) and basal area and total basal area in three old-growth ageclasses at Mount Rainier National Park, Washington, on a series of500- and 1000-m 2 plots located below 1350 m.
PseudotsugaBasal area
.-\e class Number presence(yr) of plots (%) Pseudotsuga All species
350-600 114 67 26.9 98.4'50-1000 55 69 20.1 91.81000 9 33 11.9 73.9
species following the last disturbance. Forests dominated by climaxspecies, with only a minor component of typical seral species, are evenmore common.
Disturbance frequency is a final area of contrast in scale between thePacific Northwest and most other temperate and boreal forests. We hesi-tate to suggest that the geographic scale of the disturbances is necessarilyany greater in the Pacific Northwest, despite the evidence for fire ep-isodes that covered hundreds of thousands of hectares. Hurricanes andtyphoons destroy vast reaches of temperate forests and wildfires destroysimilar areas of boreal forests at a time in other parts of the world. Thedisturbance pattern in the Pacific Northwest is one of infrequent, holo-caustic forest fires, however. A study of fire history over the last 1000years indicates a natural fire rotation averaging 434 years within MountRainier National Park (Flemstrom 1979). Limited data from elsewhere inthe region suggest that natural fire rotations of several centuries are typi-cal. There are, of course, variations associated with habitat, topographicsettings, or geoeraphical locale (e.g., rain shadow of a mountain range).Natural tire rotation may also decline with latitude. The pattern of infre-quent catastrophic, as opposed to frequent, light, burns is quite con-sistent, however.
Several other aspects of catastrophic fire in the Pacific Northwest arerelevant to successional studies. Major fire episodes appear to correlatewith climatic conditions. This is suggested by correspondance of fire ep-isodes at Mount Rainier with reconstructed dry climatic periods (Hem-strom 1979). It is also suggested by the occurrence of similar forest ageclasses over lar ge, but geographically isolated, segments of the region. Inaddition, fuel loadings are almost always heavy in these forests. This isnot an environment in \khich fire suppression leads to unnaturally heavyfuel accumulations. Related to this is the fact that wildfires in theseforests rarel y consume much of the wood. Trees die and become snagsand down logs, but se veral subsequent fires are necessary to consume a

218 J. F. Franklin and M. A. Ilemstrom
majority of this woody debris. Even the tree foliage often escapes burn-ing. Numerous examples of reburn, at least during historic times, sug-gests that young stands (e.g., 25-75 years) are more susceptible to burn-in g than later forest stages..
Revised Concepts about Succession inPseudotsuga Forests
General descriptions of succession in northwestern Pseudotsuga forestshave been extant for many decades (e.g., Isaac 1943 ., Munger 1930,1940). Pseudotsuga is described as rapidly occupying burned sites, fol-lowed by reinvasion of the more shade-tolerant and fire-sensitive Tsugaheterophylla and Thuja plicata. Establishment of the Tsuga and Thuja con-tinue indefinitely under the canopy of the Pseudotsuga. Ultimately thesespecies replace Pseudotsuga, which are lost to wind throw, pathogens, orother agents of mortality.
Such descriptions were largely anecdotal accounts rather than quantita-tive analyses, however. They were based on observations of forest restab-lishment following extensive wildfires in the mid-1800s and the Yacholt(1902) and Tillamook (1933) burns. Later stages of succession were in-ferred from species size-class patterns in mature and old- g rowth stands.Modern successional studies have also, by necessity, relied heavily oninferences drawn from size-class analyses. The great sizes and agesachieved by trees limited the usefulness of tree coring and direct observa-tion on sample plots.
Age structure analyses and historical reconstructions of Pseudotsugaforests are now underway, however, utilizing sites scheduled for timbercutting. Quantitative data on succession are accumulating rapidly and pro-vidin g new insights, many of which run counter to previous dogma.
One surprising discovery from age structure analyses is the wide rangeof ages in dominant Pseudotsuga in at least one age class of old-growth(400-500-year-old) forests. This phenomenon was observed on Experi-mental Watershed No. 10 at the 11. J. Andrews Experimental Forest. This10-ha watershed is located at 400-600 m elevation on the western slopesof the central Ore gon Cascade Range. In its pristine state the watershedwas occupied primarily by old-growth forests of Pseudotsuga, Tsugaheterophylla, Thuia plicata, Pi,uuis lamhertiana, and Castanopsis chrrsophylla(Hawk 1979). Prior to logging, all trees g reater than 5 cm diameter atbreast height were mapped, measured, and tagged. Followinging loggingin 1975, approximately 600 of 2800 trees, including a lar ge proportion ofthe dominant Pseudotsuga, were relocated and a ged from ring counts onstumps.
Watershed No. 10 appears to have been burned sometime around

14: Aspects of Succession in Coniferous Forests of Pacific Northwest 219
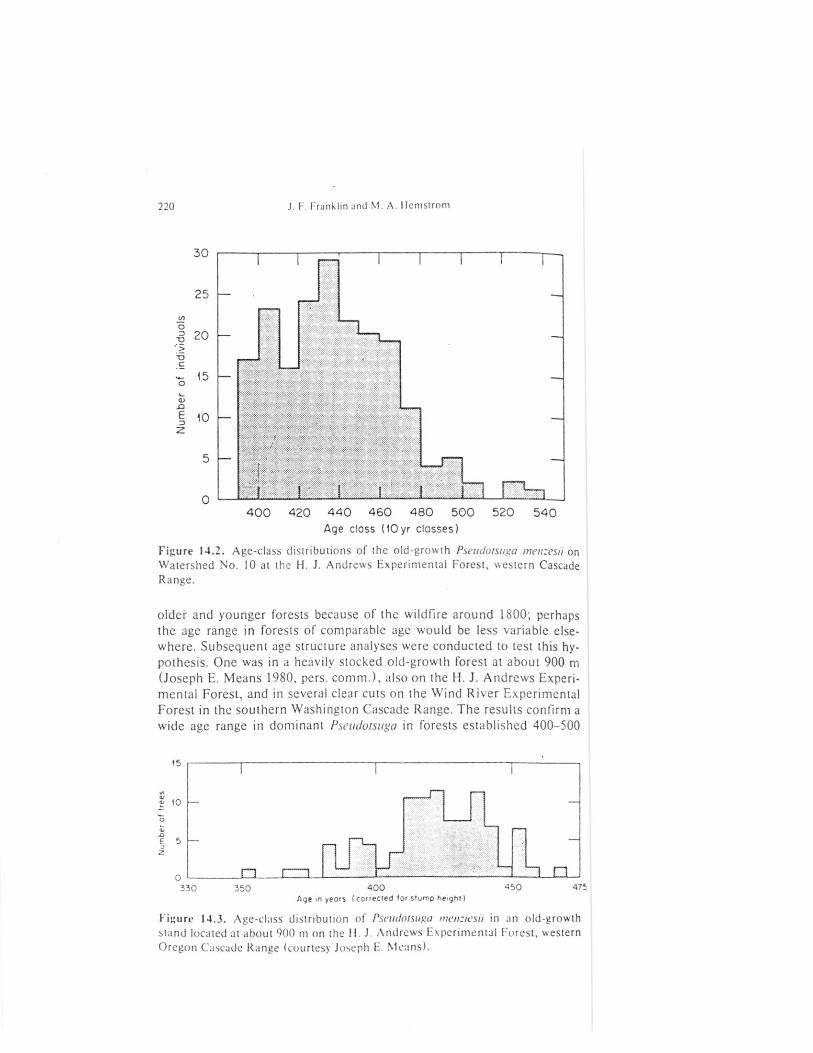
1800, based on a younger generation of Pseudotsuga (Fig. 14.1) and spar-sity of Tsuga over 200 years old. The most notable feature of the ages t ructure is the broad range of ages in the old-growth Pseudotsuga, how-ever (Fig. 14.2). This age structure indicates a very slow reestablishment-f the new forest, following the catastrophic disturbance(s) that destroyedt he forest present on the site over 500 years ago.
These results were so unexpected that confirmation was needed fromJ oe structure analyses in other stands before they were accepted. Much ofWatershed No. 10 was relatively droughty and covered by a mosaic of
70
60
50
0
E 30
20
10
0575 475 375 275 175
Tree ages (25 yr classes )
Figure 14.1. Age-class distribution of all Pseudotsuga men:iesii by 25-year ageclasses on Watershed No 10 at the II. J. Andrews Experiment Forest, westernOregon Cascade Ranee.
75
220 J. F. Franklin and M. A. 11cmstrom
30
25
• >
D
0
150
E 107
20
0400 420 440 460 480 500 520 540
Age class (10 yr classes)
Figure 14.2. Age-class distributions of the old-growth P.seutlotsuga men:esii onWatershed No. 10 at the 11. J. Andrews Experimental Forest, western CascadeRange.
older and younger forests because of the wildfire around 1800; perhapsthe age range in forests of comparable age would be less variable else-where. Subsequent age structure analyses were conducted to test this hy-pothesis. One was in a heavily stocked old-growth forest at about 900 m(Joseph E. Means 1980, pers. comm.), also on the 11. J. Andrews Experi-mental Forest, and in several clear cuts on the Wind River ExperimentalForest in the southern Washin gton Cascade Range. The results confirm awide age range in dominant Pseudotsuga in forests established 400-500
15
6
Ez
0330 350 400 450 475
Age in years (Corrected for stump height)
Figure 14.3. Age-class distribution of Psetulotsuga menziesn in an old-growthstand located at about 900 m on the 11. J. Andrews Experimental Forest, westernOregon Cascade Range (courtesy Joseph E. Means).

14: Aspects of Succession in Coniferous Forests of Pacific Northwest 221
15
10a
0
E
250 300 350 400
450Age in yeors
Figure 14.4. Age-class distribution of Pseudotsuga menziesii in old-growth standslocated around the periphery of the T. T. Munger Research Natural Area in theWind River Valley, southern Washin g ton Cascade Range.
years ago. Pseudotsuga occurred only as dominants in this stand andcovered an age span of about 140 years (Fig. 14.3). Stands at Wind Riverhad the widest range of Pseudotsuga ages yet encountered-230-460 years(Fig. 14.4).
A number of hypotheses have been proposed to explain these results.Slow recolonization is an obvious possibility. The disturbance episodethat occurred around 500 years ago was certainly extensive, coveringhundreds of thousands of hectares in the Cascade Range. The episodemay also have been exceptionally severe involving repeated wildfires.Such circumstances could result in a lack of seed source. The age distribu-tions resemble those proposed for such a scenario by Harper (1977a).Multiple disturbances are a second possibility. Young Pseudotsuga forestsare particularly susceptible to wildfire during their first 75-100 years. Par-tial reburns during this time can provide opportunities for establishmentof additional Pseudotsuga cohorts. This has been documented in youngPseudotsuga stands (Mark Klopsch 1980, pers. comm.) but is very hard toverify in old-growth stands, where there are fewer trees and greater errorsin aging. Surface erosion on steep sites or climatic damage early in standhistory could also contribute to the wide age range in dominant, old-growth Pseudotsuga. Another possibility is competition from shrub orhardwood trees, such as Ccanothus and Abuts. They could delay coniferestablishment or suppress early growth of conifer seedlings and saplings.There is very little evidence for the suppression phenomenon in earlygrowth patterns of the dominant conifers, however.
222 J. F. Franklin and M. A. Hemstrom
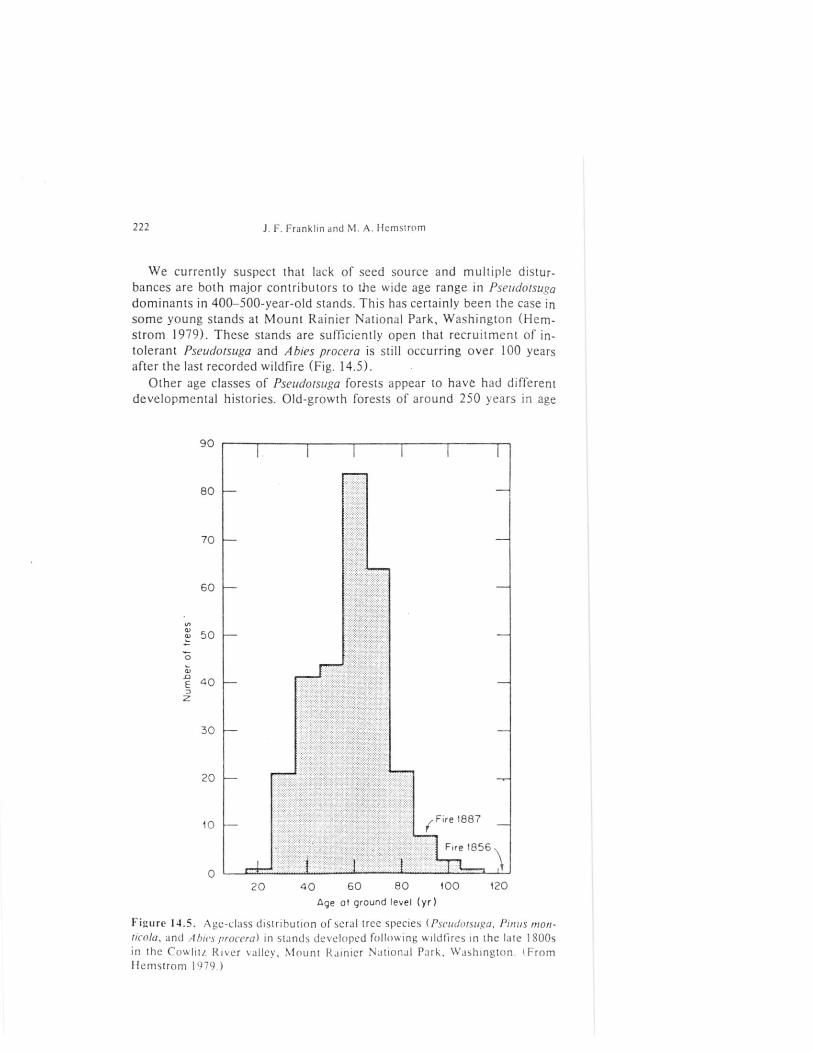
We currently suspect that lack of seed source and multiple distur-bances are both major contributors to the wide age range in Pseudotsugadominants in 400-500-year-old stands. This has certainly been the case insome young stands at Mount Rainier National Park, Washington (Hem-strom 1979). These stands are sufficiently open that recruitment of in-tolerant Pseudotsuga and Abies procera is still occurring over 100 yearsafter the last recorded wildfire (Fig. 14.5). •
Other age classes of Pseudotsuga forests appear to have had differentdevelopmental histories. Old-growth forests of around 250 years in age
r Fire 1887
Fire 1856
20 40 60 80 100 120Age at ground level (yr )
Figure 14.5. Age-class distribution of sera! tree species (Pseudotsuga, Pinus mon-ticola, and Abies procera) in stands developed following wildfires in the late 1800sin the Cowlitz River valle y , Mount Rainier National Park, Washington. ( Fromflemstrom 1979.)
90
80
70
60
a) 500
E 40
30
20
10

Ages not shown:
1 age of 5951 oge of 6491 age of 725
Fire obout 264t years ago
1
14: Aspects of Succession in Coniferous Forests of Pacific Northw-est 223
occur at scattered locations in the Cascade Range. These forests havebeen noted for their relatively hi g h densities of rather uniformly sizedpseudotsuga dominants. Tree ages in several stands, were much moreiiohtly clustered than in the older forests (Fig. 14.6). More abundantseed supplies and an absence of reburns would both contribute to themore rapid complete restocking of the site. Age structure analyses ofstands on sites of small, single-burn wildfires may reveal even tightercohorts—comparable to these proposed by Munger (1930, 1940) andIsaac (1943).
Recent data show species composition to be as variable as stocking lev-els in early stages of forest succession in the Pacific Northwest. Manyspecies play varying successional roles, depending on site and seedsource. Shade-tolerant species, such as Tsuga hetcrophylla or Abies ania-hilts, have the potential for direct reestablishment on many disturbedsites. Even more commonly species mixtures, such as Pseudotsuga andTsuga, become established on disturbed sites, followed by structural dif-ferentiation due to contrasting species erowth rates. Dr. Chadwick Oliverand his students at- the University of Washington have studied severaly oung forest stands in the Pacific Northwest that follow this pattern, in-cludin g mixtures of Pseudotsuga and Tsuga heterophylla (Wierman et al1979) and Alma rubra and associated conifers (Stubblefield et al 1978).
Analyses of age structure and developmental history are just beginningin the Pacific Northwest, but current work has been revealing. The seres
20
15
E
0 10al
E
180 200 220 240
260 280 300Age (yr)
Figure 14.6. Age-class distribution of PseudorsiiKa in a mature forest near RagbyResearch 'natural Area, Clackamas River drainage, western Oregon CascadeRange.

224 .1 F. Franklin and M. A. I lemstrom
following each disturbance episode appear very individualistic. Initialforests have been widely divergent in composition. stockin g levels, andtimes to canopy closure. Forest chronosequences must be interpretedconservatively. For example, a 450-year-old Psetalotsuga forest has notnecessarily evolved through stages comparable to existing 135-250-year-old stands.
Role of Woody Debris in Forest Succession
Coarse woody debris in the form of snags and do%^ n logs is a major struc-tural feature of northwestern forests (Franklin et al. 1981). An avera ge of194 metric tons/ha was measured in a series of 10 natural stands rangingfrom 100 to 1000 years in age (Franklin and Waring 1980). Down logsaveraged 106.7 metric tons/ha with a range of 60-156 in this data set. Themaximum value recorded for down logs was 418.5 metric tons/ha in anintact stand in the Carbon River, Mount Rainier National Park.
Trends in coarse woody debris were weak across the chronosequenceof stands due partially to surprisingly high levels in three young(100-150-year-old) stands. Large amounts of dead wood are apparentlycarried over from previous stand, since massive boles of the fire-killedtrees are typically not consumed. This material persists until the replace-ment stand begins to provide woody debris of comparable size.
Table 14.3. Densities and total numbers of Picea sitchensis and Tsuga hereto-phylla reproduction (stems S 8 m tall) by substrate type and logdecay class in a mature forest on a high terrace along the SouthFork I loh River, Olympic National Park, Washington."
Density (number/m2) Absolute (number/ha)Picea Tsuga Pic ea Tsuga
Substrate typePicea logs 36.0 15.3 14,500 6170Tsuga logs 29.9 9.6 15,300 4910Unknown logs 30.0 7.5 4920 1230Stumps and root wads 5.I 1.6 550 170Ground humus 0.08 0.01 700 90
Log decay classEarly (II) 24.8 15.5 7030 4340Middle (III) 38.5 10.7 19,000 5250Late (IV) 28.6 9.2 7800 2500Very late (V) 28.2 8.2 820 170
From McKee ct al. (in press).

14: Aspects of Succession in Coniferous Forests of Pacific Northwest 225
Rotten wood seedbeds are very important throu ghout the coastal Piceasitchensis zone (Minore 1972), but nurse logs attain maximum impor-t ance in the forests of Picea sitchensis and Tsuga heteraphylla found in thevalleys of the western Olympic Mountains. Reproduction in these alluvialfores ts is essentially confined to log substrates (Table 14.3) (McKee et al.in press). The absence of seedlin gs .on the forest floor contrasts sharply^vith the dense reproduction crowded onto logs, which occupy only 11%of the area. Logs also vary markedly in their suitability as a seedbed withsta o e of decay (moderate stages of decay have highest seedlin g densities)and log species (Tsuga lo gs are poorer seedbeds than those of Piceasitchens is or Pseudotsuga).
Past patterns in amount and distribution of woody debris may alsohave strongly influenced the structure of existing mature Picea-Tsugaforests in the western Olympic Mountains. Tree densities and stand basalareas and biomass values are less than expected in this climatically favor-able forest environment, resulting in relatively open and well lightedstands. The lower densities could result from the fact that suitableseedbeds (logs) were limited at the time of stand establishment and havestill not been produced insufficient numbers to allow development of fullstocking. In any case, Picea sitchensis reproduces successfully and attainsclimax status in these relatively open alluvial forests in contrast to its seralrole elsewhere in the dense forests typical of the Picea sitchensis zone(Franklin and Dyrness 1973).
Long-Lived Seral Species in SuccessionModeling
We are interested in developing and utilizing successional models for Pa-cific Northwestern conifer forests for use in both basic and appliedresearch. The philosophical construct that underlies the JABOWA-FOR ET family of models appears to us as a concise general statementsummarizing our knowledge of succession: vegetational or ecosystemchan g e is a function of life histories of available species interactin g withenvironmental conditions (includin g disturbances) and with stochasticelements to produce a probabilistic array of alternative outcomes. Thisconstruct encompasses the diversity of ecosystems and processes outlinedin this book and provides a basis for developin g and testin g hypotheses.Some of the distinctive features of Pacific Northwest forests require spe-cial consideration in a FOR FT-type model (Shugart and West 1977).Rates and scales rather than types of processes distinguish succession inthese forests and require model modification. Specific modeling, concernsIn the northwestern forests include the following: (1) mortality must be

226 J. F. Franklin and M. A. Ilemstrom
modeled on an annual time step; (2) plot size is critical; and (3) previousstand history is important.
Long-lived trees complicate the task of developing mortality proba bil-ity functions. Two to four generations of shorter-lived species may ger-minate, mature, and die during the life span of one Pseudotsuga, Thuja pli-cata, or Chamaecyparis nootkatensis. Annual mortality of these short- andlong-lived species is not constant between species nor within species bydiameter. There seem to be five patterns of suppression-related mortality(Fig. 14.7):
Long-lived early seral species, which undergo high mortality ratesin small, suppressed size classes but are able to endure long periods ofslow growth in large size classes (e.g. Pseudotsuga, PinUS ponderosa) (Fig.14.7a).
Short-lived early seral species, which experience constant annualmortality (e.g., Alnus rubra) (Fig. 14.7b).
Long-lived early to late seral species, which are relatively shadetolerant and experience little suppression mortality in small or large sizes(e.g., Chamaeoparis nootkatensis and Thuja plicata) (Fig.14.7c).
Long-lived early to mid-seral species, which experience moderatesuppression mortality in small sizes, reach minimum suppression mortal-ity at about half their maximum diameter, then experience increasingsuppreSsion mortality in lar ge sizes (e.g., Abies procera) (Fig. 14.7d).
5. Relatively shorter-lived, mid- to late seral, shade-tolerant species,which experience the least suppression mortality at small sizes and slowlyincreasing suppression mortality in larger sizes (e.g., Abies amabilis andTsuga heterophvlla) (Fi g . 14.7e).
The current model requires that trees die any year based on their es-timated annual mortality probability. Because all species are relativelylong-lived and at least a few usually survive to old age in a given stand,annual mortality probabilities are very small (Table 14.4). In addition,fires or wind storms, which occur at intervals of 100-1000 years, causeepisodes of mortality. Episodic disturbances must be included to accu-rately simulate patterns of regeneration and growth in understory treesand inputs of dead wood to the forest floor.
Large trees also place some constraints on plot size for simulatedstands. Internal computer storage limits require that number of treesmodeled as individuals be relatively small. A plot size for simulatedstands large enough to prevent a single large Pseudotsuga from completelydominating the stand could contain large numbers of small trees. Raisingthe minimum recruitment size to compensate results in loss of informa-tion about re g eneration patterns. In addition, the simulated death of anold-growth Pseudotsuga in a small plot causes a sharp increase in availablelight; gap-phase reproduction becomes possible. In reality, a small openarea in a matrix of trees 80 m tall does not receive enou g h light for suc-

1. 0 (b)
0.5
Mox(0.1) (0.5)DBH max DBH mox
( o)
0 5
0 1
0 (0.11 DBH mox Max
C>-0
C
3(f)
0
1300
00
1 . 0
0.5
0 1
(01) DBH mox1 . 0 "
7- ( c )
Mox
Max
(d)
0.5
0
0.5
1 .0
0(01) DBH max
DBH
DBH mox
(e)
1
14: Aspects of Succession in Coniferous Forests of Pacific Northwest 227
Figure 14.7. Probability of surviving 10 years of suppressed growth, diameter in-crease <0.1 cm/yr, for five groups of important tree species west of the CascadeCrest, Washington and Oregon. (a) long-lived early seral species; (b) short-livedearly sera! species; (c) long-lived early to mid-seral species, low suppression mor-tality; (d) lon g -lived early to mid-seral species, increasing suppression mortality;(e) long-lived late seral species (dbh, diameter at breast height). Curve a is com-piled from NIcArdle (1949) and a chronosequence of stands in western Washing-ton and Ore gon (data on file at Forestry Sciences Laboratory, Corvallis, Oregon).
cessful establishment of intolerant species. Unrealistically high levels ofintolerant species regeneration ma y occur in closed forest situations.Eventually, stand models should he altered to accept larger plot sizes,

228 J. F. Franklin and M. A. Hemstrom
Table 14.4. Example of annual mortality in Pseudotsuga menziesii.a
Age (yr) Plc
r„ 10
20-50 0.940 0.58650-100 0.981 0.825100-150 0.996 0.961150-250 0.999 0.990250-450 0.996 0.961450-750 0.995 0.951750-1000 0.994 0.942
'Compiled from McArdle (1949) and unpublished data on file at the Fores-try Sciences Laboratory, Corvallis, Oregon.b Probability of surviving 1 year of diameter growth less than 0.1 cm.
`Probability of surviving 10 years of diameter growth less than 0.1 cm/yr.
perhaps by modeling cohorts of small trees and then graduating them intothe stand as individuals at a particular diameter.
Finally, stands and disturbance over the previous centuries have animportant bearing on the development of future stands by determiningthe availability of log seedbeds and seed sources. Currently, the model as-sumes time zero to be bare ground with all species available for establish-ment. As emphasized by Franklin and Waring (1980), abundant deadwood carries over from one stand to another even though sequences ofsevere wildfires or other disturbances. Thus, adjustments in the modelmust be made to account for variable initial conditions that follow naturaldisturbances. Disturbance intensity and frequency are important deter-minants of those conditions.
Conclusions
The forests of coastal Pacific Northwest differ from forests in othertemperate and boreal regions in several respects. Trees can grow verylarge and old. Leaf areas can reach 20 m 2 /m 2 , and above-ground, livebiomass values can exceed 1500 metric tons/ha.
Northwestern forests recover slowly after disturbance. Canopy clo-sure may not occur for 40 years or more. Early seral intolerant trees mayhave an age range greater than 100 years in one cohort.
Due to infrequent, catastrophic disturbance and long-lived seralspecies, climax conditions rarely occur when intolerant, early seral treesare important in the initial stand.
Late sera' or "climax — species are often important pioneers.

14: Aspects of Succession in Coniferous Forests of Pacific Northwest 229
Stand history, as it determines the availability of log seedbeds andseed sources, plays an important role in determining the composition andstructure of future stands.
The longevity and size of the early seral species pose interestingproblems in developing FORET-type successional models, particularly inmodelin g mortality, determining plot size, and verifying models.
7. The northwestern forests provide the maximum expression ofmany forest features as well as trends observable in other temperateforest regions. These include size and longevity of species, biomass accu-mulations, and overall importance of coarse woody debris. General suc-cessional theories must take these northwestern phenomena into accountand may find their ultimate tests in this region.
Reproduced by USDA Forest Servicefor official use


















