
arnm - plantphysiol.org · plant materials has been reviewed by Simon (21) ... Indiani Canicer...
6
VICKERY INFLUENCE OF LIGHT ON CITRIC ACID 14. VICKERY, H. B. The metabolism of the organic acids of tobacco leaves. VIII. Effect of culture of ex- cised leaves in solutions of L-malate at pH 4 to pH 7. Jour. Biol. Chem. 214: 323-333. 1955. 15. VICKERY, H. B., PUCHER, G. 'AT., WAKE-MAN, A. J. and LEAVEN\\WORTH, C. S. Chemical inivestigations of the tobacco plant. VI. Chemical changes that occur in leaves during culture in light and in dark- ness. Conn. Agr. Expt. Station. Bull. 399. 755- 832. 1937. 16. V7ICKERY, H. B. The formation of starch in leaves of Bryophyllumi calycim ii cultured in darkness. Plant Physiol. 27: 231-239. 1952. 17. VrICKERY, H. B. The formatioin of starch anid the be- havior of isocitric acid in excised leaves of Bryo- phyllimi calvcinzo.m cultured in darkness. Plant Phv siol. 32: 220-224. 1957. 18. VICKERY, H. B. The effect of temperature on the behavior of malic acid and starch in leaves of Bryo- phyllain calycitnuui cultured in darkness. Plant Physiol. 29: 385-392. 1954. 19. VICKERY, H. B. and WILSON, D. G. Preparation of potassium dihydrogen LS ( + ) -isocitrate from Bryo- phylluiii calyciniuni leaves. Jour. Biol. Chem. 233: 14-17. 1958. 20. \WOLF, J. Beitrage zur Kenniitniis des Sauerstoff- wechsels sukkulenter Crassulacean. IV. Beobach- tungen iiber Gehaltsschwankungen von Gesamt-, Apfel- und Zitronensiurein. Planta. 29: 314-324. 1939. 21. ZBINOVSKY, V. anid BURRIs, R. H. Mfetabolism of infiltrated organic acids by tobacco leaves. Plant Physiol. 27: 240-250. 1952. RESPIRATORY AND ASSOCIATED RESPONSES OF CARROT DISCS TO SUBSTITUTED PHENOLS ' B. K. GAUR' AND HARRY BEEVERS DEPARTMENT OF, BIOLOGICAL SCIENCES, PURDUE UNIvERsITy, LAFAyETTE, INDIANA The striking physiological and biochemical effects of the classical uncoupling agent 2,4-dinitrophenol (DNP) are now well known, (4, 5, 15) though still imperfectly understood. The pertinent work with plant materials has been reviewed by Simon (21) and Laties (16). Stenlid, who carried out some of the earliest Nwork on plants, reported that 5 other nitro- phenols brought about similar respiratory responses to those indluced by DNP, and, like DNP, inhibited glucose uptake by wheat roots (24). The behavior of the nitrophenols was comiiparedl to that of azide (23) and later to that of methylene blue (25). In this paper (quantitative comparisons have been mladle of the effects of a series of NO.,-, Cl-, and Br- substituted phenols on respiration andl concommitant glucose uptake. In addition, the effects of these conm- pounds on mitochondrial oxidations have been de- termiiled. The conclusion is dlrawn that, with dif- ferent degrees of efficiency, all of the substituted phenols acted as uncoupling agents. Observations on the effects of the phenols on C'402-output frolm glucose-Cl4 are also presented ancd discussed in relation to the (uestion raised re- cently by Porter aind Runeckles (19) of a possible shift in respiratory substrate resulting from DNP treatmiient. MATERIALS AND AIMETHODS Carrots of 25 to 30 mm average diameter were purchased locally and storedI at 3 to 50 C for at least a week before use. Discs of about 0.3 mm thickness and 7 mnm diameter were prepared, wTashed quickly I Received revised manuscript November 3, 1958. 2 Present address: Biology Division,, A. E. E. T., Indiani Canicer Research Centre, Parel, Bombay, India. in 3 changes of distilled water, drie(l lightly, and separated into weighed samples. The various phenols and other reagents were of the best grade commer- cially available, and were used without further puri- fication. Radioactive glucose, unifornmly labeled with C14 was prepared by hydrolysis and chromatographic separation from a sample of sucrose-C14 prodluced (luring photosynitlhesis in C'40., and generously sup- pliedI by Dr. H. R. Garner. All determinations of radioactivity were carried out in a gas flow window- less counting system, and the results are corrected for background, and in the case of carbonate, for self absorption. Two types of experiments were carried out. In the 1st, the aimii was to determine the relative effects on 0.,-uptake of a group of phenols andI other com- poun(Is, by testing responses over a wide range of concentrations. For such experiments 650 mg samples of (liscs wrere placed in Warburg flasks (with the test compound wvhere necessary) in 2 ml 0.03 M potassium phosphate pH 5.0. O., uptake was meas- ure(l over a 2-hour period in the usual manner at 250 C. The information from these experiments was used to choose, for each compound, concentrations which were: a) slightly stinmulatory, b) strongly stinmulatory, and c) inhibitory to 0,-uptake. In the 2nd series of experiments the test comnpoundIs were supplied at each of these 3 levels and stimultaneous measurements of 0.,-uptake and glucose uptake were made over a longer experimental period (6 to 8 hr). In these experiments 2 series of flasks wvere assembled. The 1st was composed of large vessels (100 ml capacity) and these contained 1.30 g discs, the test compound, and an aliquot of radioactive glucose (0.036 mg) (tip- ped from the side arnm after equilibration) in 4 ml 427 www.plantphysiol.org on June 16, 2018 - Published by Downloaded from Copyright © 1959 American Society of Plant Biologists. All rights reserved.
Transcript of arnm - plantphysiol.org · plant materials has been reviewed by Simon (21) ... Indiani Canicer...

VICKERY INFLUENCE OF LIGHT ON CITRIC ACID
14. VICKERY, H. B. The metabolism of the organic acidsof tobacco leaves. VIII. Effect of culture of ex-cised leaves in solutions of L-malate at pH 4 to pH7. Jour. Biol. Chem. 214: 323-333. 1955.
15. VICKERY, H. B., PUCHER, G. 'AT., WAKE-MAN, A. J.and LEAVEN\\WORTH, C. S. Chemical inivestigationsof the tobacco plant. VI. Chemical changes thatoccur in leaves during culture in light and in dark-ness. Conn. Agr. Expt. Station. Bull. 399. 755-832. 1937.
16. V7ICKERY, H. B. The formation of starch in leavesof Bryophyllumi calycimii cultured in darkness.Plant Physiol. 27: 231-239. 1952.
17. VrICKERY, H. B. The formatioin of starch anid the be-havior of isocitric acid in excised leaves of Bryo-phyllimi calvcinzo.m cultured in darkness. PlantPhv siol. 32: 220-224. 1957.
18. VICKERY, H. B. The effect of temperature on thebehavior of malic acid and starch in leaves of Bryo-phyllain calycitnuui cultured in darkness. PlantPhysiol. 29: 385-392. 1954.
19. VICKERY, H. B. and WILSON, D. G. Preparation ofpotassium dihydrogen LS ( + ) -isocitrate from Bryo-phylluiii calyciniuni leaves. Jour. Biol. Chem. 233:14-17. 1958.
20. \WOLF, J. Beitrage zur Kenniitniis des Sauerstoff-wechsels sukkulenter Crassulacean. IV. Beobach-tungen iiber Gehaltsschwankungen von Gesamt-,Apfel- und Zitronensiurein. Planta. 29: 314-324.1939.
21. ZBINOVSKY, V. anid BURRIs, R. H. Mfetabolism ofinfiltrated organic acids by tobacco leaves. PlantPhysiol. 27: 240-250. 1952.
RESPIRATORY AND ASSOCIATED RESPONSES OFCARROT DISCS TO SUBSTITUTED PHENOLS '
B. K. GAUR' AND HARRY BEEVERSDEPARTMENT OF, BIOLOGICAL SCIENCES, PURDUE UNIvERsITy, LAFAyETTE, INDIANA
The striking physiological and biochemical effectsof the classical uncoupling agent 2,4-dinitrophenol(DNP) are now well known, (4, 5, 15) though stillimperfectly understood. The pertinent work withplant materials has been reviewed by Simon (21)and Laties (16). Stenlid, who carried out some ofthe earliest Nwork on plants, reported that 5 other nitro-phenols brought about similar respiratory responsesto those indluced by DNP, and, like DNP, inhibitedglucose uptake by wheat roots (24). The behaviorof the nitrophenols was comiiparedl to that of azide(23) and later to that of methylene blue (25).
In this paper (quantitative comparisons have beenmladle of the effects of a series of NO.,-, Cl-, and Br-substituted phenols on respiration andl concommitantglucose uptake. In addition, the effects of these conm-pounds on mitochondrial oxidations have been de-termiiled. The conclusion is dlrawn that, with dif-ferent degrees of efficiency, all of the substitutedphenols acted as uncoupling agents.
Observations on the effects of the phenols onC'402-output frolm glucose-Cl4 are also presentedancd discussed in relation to the (uestion raised re-cently by Porter aind Runeckles (19) of a possibleshift in respiratory substrate resulting from DNPtreatmiient.
MATERIALS AND AIMETHODS
Carrots of 25 to 30 mm average diameter werepurchased locally and storedI at 3 to 50 C for at leasta week before use. Discs of about 0.3 mm thicknessand 7 mnm diameter were prepared, wTashed quickly
I Received revised manuscript November 3, 1958.2 Present address: Biology Division,, A. E. E. T.,
Indiani Canicer Research Centre, Parel, Bombay, India.
in 3 changes of distilled water, drie(l lightly, andseparated into weighed samples. The various phenolsand other reagents were of the best grade commer-cially available, and were used without further puri-fication. Radioactive glucose, unifornmly labeled withC14 was prepared by hydrolysis and chromatographicseparation from a sample of sucrose-C14 prodluced(luring photosynitlhesis in C'40., and generously sup-pliedI by Dr. H. R. Garner. All determinations ofradioactivity were carried out in a gas flow window-less counting system, and the results are corrected forbackground, and in the case of carbonate, for selfabsorption.
Two types of experiments were carried out. Inthe 1st, the aimii was to determine the relative effectson 0.,-uptake of a group of phenols andI other com-poun(Is, by testing responses over a wide range ofconcentrations. For such experiments 650 mgsamples of (liscs wrere placed in Warburg flasks (withthe test compound wvhere necessary) in 2 ml 0.03 Mpotassium phosphate pH 5.0. O., uptake was meas-ure(l over a 2-hour period in the usual manner at250 C.
The information from these experiments was usedto choose, for each compound, concentrations whichwere: a) slightly stinmulatory, b) strongly stinmulatory,and c) inhibitory to 0,-uptake. In the 2nd seriesof experiments the test comnpoundIs were supplied ateach of these 3 levels and stimultaneous measurementsof 0.,-uptake and glucose uptake were made over alonger experimental period (6 to 8 hr). In theseexperiments 2 series of flasks wvere assembled. The1st was composed of large vessels (100 ml capacity)and these contained 1.30 g discs, the test compound,and an aliquot of radioactive glucose (0.036 mg) (tip-ped from the side arnm after equilibration) in 4 ml
427
www.plantphysiol.orgon June 16, 2018 - Published by Downloaded from Copyright © 1959 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
0.03 M phosphate pH 5.0. In order to measure glu-cose uptake, samples (0.1 ml, containing initiallyabout 400 cpm) were removed at hourly intervalsfrom these large flasks. These were dried under aninfrared lamp on steel planchets and counted directly.The decrease in radlioactivity was used to calculateglucose uptake.
The 2nd, parallel, series was made up of snmallflasks each containing 650 mg discs in 2 ml of solutionwith proportionate anmounts of glucose and the testcompounds. These flasks were used to obtain a con-tinuous record of 0. -uptake of controls and treatedsamples during the experiment; they were not openedwhen samples were removedl from the larger flasks.The respired CO., was trapped in 20 % sodium hy-droxide and the radioactivity determined at the endof the experinment by converting to barium carbonate,plating and counting. For each of the compoundstestedl an experiment of this kind was carried out andthen repeatedl at least 3 times. The results were thenpooled and averaged. In the experiments witlh nito-chondria, castor beans were used; the methods werethose described by Beevers and Walker (3).
RESULTS
A. EXPERIMENTS IN WIHICII RESPIRATION ONLYVVAS MEASURED: When different concentrations of2,4-dinitrophenol were applied, the anticipatedstimulatory and inhibitory effects were establishedfrom the outset, and O.-uptake continued unchangedat these new levels throughout the period of measure-ment. From data obtainedl in this way dosage re-sponse curves were constructed for each compound.
The forms of the various response curves weresimilar; for each compound only a rather narrowrange of concentrations induced stimulated O.,-uptake,
and inhibitory effects ensued when higher levels wereapplied. The response curves differed from eachother in 2 respects; the extent of the stimulation in-duced at the most effective levels of the test compoundand in the placing of the response curve along theconcentration axis.
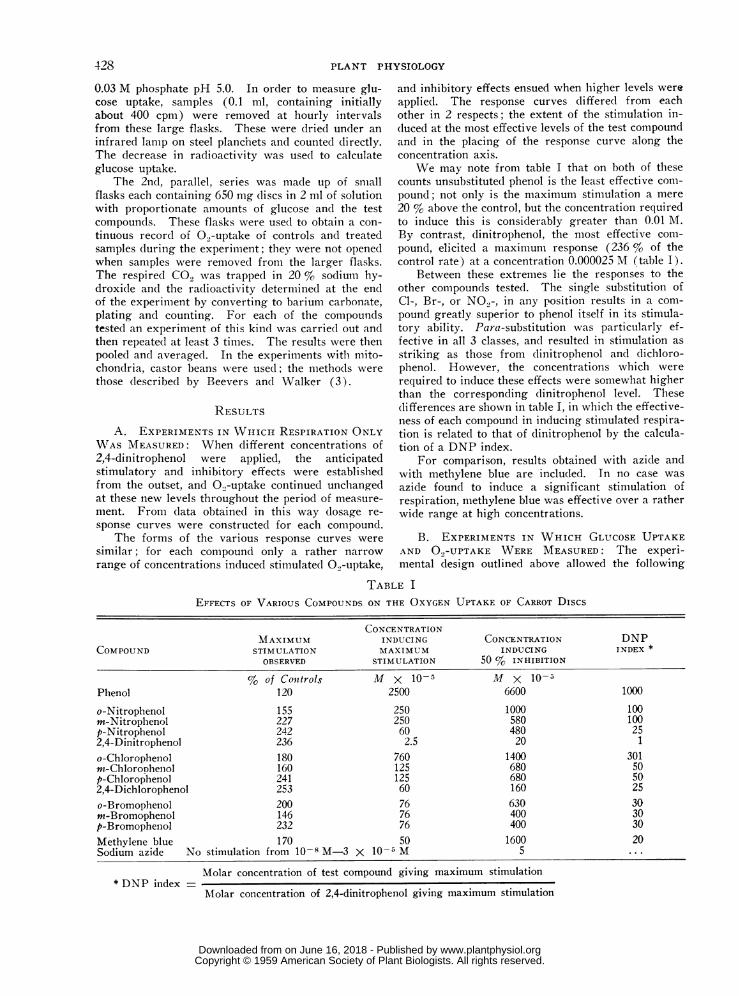
We may note from table I that on both of thesecounts unsubstituted phenol is the least effective com-pound; not only is the maximum stimulation a mere20 % above the control, but the concentration requiredto induce this is considerably greater than 0.01 M.By contrast, dinitrophenol, the most effective com-pound, elicited a maximum response (236 % of thecontrol rate) at a concentration 0.000025 M (table I).
Between these extremes lie the responses to theother compounds tested. The single substitution ofCl-, Br-, or NO.,-, in any position results in a com-pound greatly superior to phenol itself in its stimula-tory ability. Para-substitution was particularly ef-fective in all 3 classes, and resulted in stimulation asstriking as those from (linitrophenol and dichloro-phenol. However, the concentrations which wererequired to induce these effects were somewhat higherthan the corresponding dinitrophenol level. Thesedifferences are shown in table I, in which the effective-ness of each compound in inducing stimulated respira-tion is related to that of dinitrophenol by the calcula-tion of a DNP index.
For comparison, results obtained with azide andwith methylene blue are includled. In no case wasazide found to induce a significant stimulation ofrespiration, methylene blue was effective over a ratherwide range at high concentrations.
B. EXPERIMENTS IN WHICH GLUCOSE UPTAKEAND O.,-UPTAKE WERE MEASURED: The experi-mental design outlined above allowed the following
BLE I
EFFECTS OF VARIOUS COMPOUNDS ON THE OXYGEN UPTAKE OF CARROT DISCS
CONCENTRATIONMAXIMUM INDUCING CONCENTRATION DNP
COMPOUND STIMULATION MAXIMUM INDUCING INDEX *OBSERVED STIMULATION 50 % INHIBITION
% of Conttrols Al' x 10-5 M x 10-aPhenol 120 2500 6600 1000
o-Nitrophenol 155 250 1000 100m-Nitrophenol 227 250 580 100p-Nitrophenol 242 60 480 252,4-Dinitrophenol 236 2.5 20 1
o-Chlorophenol 180 760 1400 301rm-ChloroDhenol 160 125 680 50p-Chlorophenol 241 125 680 502,4-Dichlorophenol 253 60 160 25o-Bromophenol 200 76 630 30m-Bromophenol 146 76 400 30p-Bromophenol 232 76 400 30Methvlene blue 170 50 1600 20Sodium azide No stimulation from 10-8 M-3 X 10-5 M 5
* DNP index =Molar concentration of test compound giving maximum stimulation
Molar concentration of 2,4-dinitrophenol giving maximum stimulation
42
www.plantphysiol.orgon June 16, 2018 - Published by Downloaded from Copyright © 1959 American Society of Plant Biologists. All rights reserved.
429GAUR AND) BEEVERS--RESPONSES TO SUBSTITUTED PHENOLS
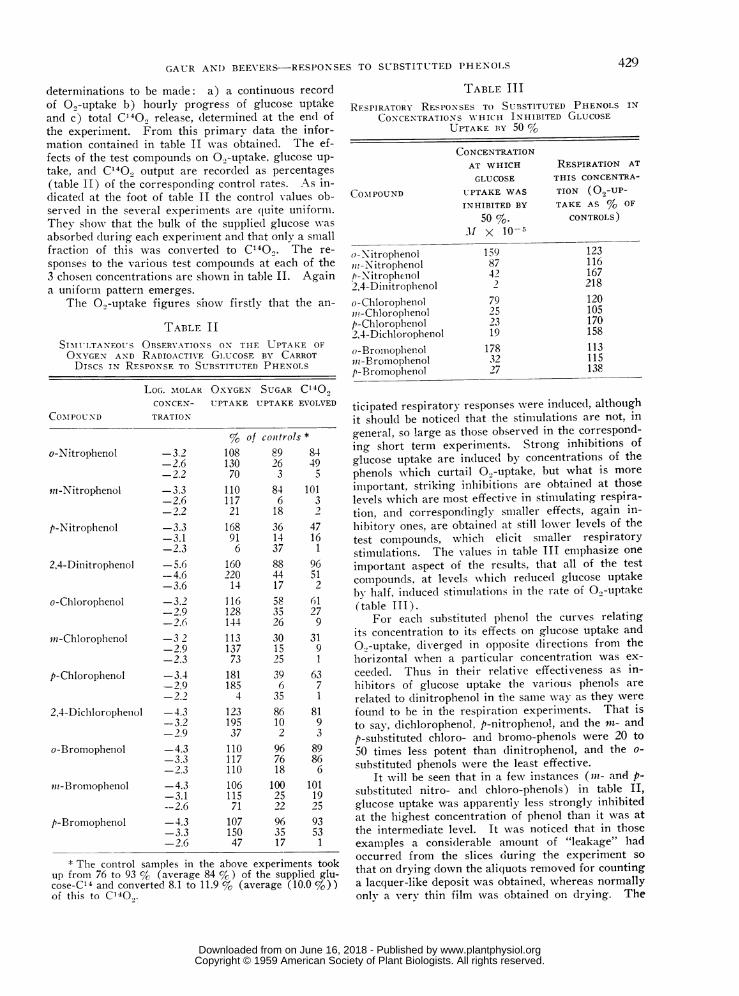
determinations to be made: a) a continuous recordof 09-uptake b) hourly progress of glucose uptakeand c) total C1400 release, determined at the end ofthe experiment. From this primary data the infor-mation containe(l in table II was obtained. The ef-fects of the test compounds on 0,-uptake, glucose up-take, and C140., output are recordedI as percentages(table II) of the corresponding control rates. As in-dicated at the foot of table II the control values ob-servedl in the several experiments are (quite uniformii.They showN, that the bulk of the suppliedI glucose wasabsorbedl during each experiment and that only a smallfraction of this was converted to C140.. The re-sponses to the various test compounds at each of the3 chosen concentrations are shown in table II. Againa uniform pattern emerges.
The 00-uptake figures show firstly that the an-
TABLE II
SIMTULTANEO1US OBSERVATIONS ON THF UPTAKE OFOXYGEN AND RADIOACTINVE Gi.uCOSE BY CARROTDISCS IN RESPONSE TO SUBSTITUTED PHE-NOLS
LOG. MOLAR OXYGEN SUGAR C140,CONCEN- UPTAKE UPTAKE EVOLVED
COMIPOUNS D TRATIONN
% of cowtrols *
o-Nitrophenol -3.2 108 89 84-2.6 130 26 49-2.2 70 3 5
ini-Nitrophenol -3.3 110 84 101-2.6 117 6 3-2.2 21 18 2
p-Nitrophenol -3.3 168 36 47-3.1 91 14 16-2.3 6 37 1
2,4-Diniitrophenol -5.6 160 88 96-4.6 220 44 51-3.6 14 17 2
o-Cl1lorophenol -3.2 116 58 61-2.9 128 35 27-2.6 144 26 9
mt-Chlorophenol -3 2 113 30 31-2.9 137 15 9-2.3 73 25 1
p-Chlorophenol -3.4 181 39 63-2.9 185 6 7-2.2 4 35 1
2,4-Dichlorophelnol -4.3 123 86 81-3.2 195 10 9-2.9 37 2 3
o-BromopheInol -4.3 110 96 89-3.3 117 76 86-2.3 110 18 6
ni-Bromophenlol -4.3 106 100 101-3.1 115 25 19--2.6 71 22 25
p-Bromophenol -4.3 107 96 93-3.3 150 35 53-2.6 47 17 1
g The cointrol samples in the above experiments tookup from 76 to 93 % (average 84 %) of the supplied glu-cose-C14 and converted 8.1 to 11.9 % (average (10.0 %))of this to C' 40.,.
TABLE IIIRESPIRATORY RESPONSES TO SUBSTITUTED PHENOLS IN
CONCENTRATIONS WN-HICH INHIBITED GLUCOSEUPTAKE BY 50 %
COMAIPOUND
o-NitrophenolJin-Nitrophenolh-Nitrophenol2,4-Dinitrophenolo- ChlorophenolmP!-Chlorophenolp-Chlorophlciol2,4-Dichlorophenolo-Broiioplhenoliii-Bromophenolp-Bromophenol
CONCENTRATIOIAT WHICHGLUCOSE
UPTAKE WASINHIBITED BY
50 %.Jf x 10-5
1598742
792523191783227
RESPIRATION ATTHIS CONCENTRA-TION (09-UP-TAKE AS % OF
CONTROILS)
123116167218120105170158113115138
ticipated respiratory responses were induced, althouighit should be notice(l that the stimulations are not, ingeneral, so large as those observed in the correspond-ing short term experiments. Strong inhibitions ofglucose uptake are induced by concentrations of thephenols which curtail 0.,-uptake, but what is more
important, striking inhibitions are obtained at thoselevels which are most effective in stimulating respira-tion, and correspondingly smaller effects, again in-hibitory ones, are obtaine(d at still lower levels of thetest compounds, which elicit smaller respiratorystimulations. The values in table III emphasize one
important aspect of the results, that all of the testcompounds, at levels wlhich reduced glucose uptakebv half, induced stimulations in the rate of 0.)-uptake(table III).
For each substituted plhenol the curves relatingits concentration to its effects on glucose uptake and0.,-uptake, diverged in opposite (lirections from thehorizontal when a particular concentration was ex-
ceeded. Thus in their relative effectiveness as in-hibitors of glucose uptake the various phenols are
related to dinitrophenol in the same way as they werefound to be in the respiration experiments. That isto say, dichlorophenol, p-nitrophenol, and the Sn- andp-substituted chloro- and bromo-phenols were 20 to50 times less potent than dinitrophenol, and the o-substituted phenols were the least effective.
It will be seen that in a few instances (inz- andl p-substituted nitro- andl chloro-phenols) in table II,glucose uptake was apparently less strongly inhibitedat the highest concentration of phenol than it was atthe intermediate level. It was noticed that in thoseexamples a considerable amount of "leakage" hadoccurred from the slices (luring the experiment sothat on drying down the aliqluots removed for countinga lacquer-like deposit was obtained, whereas normallyonly a very thin film was obtained on drying. The
www.plantphysiol.orgon June 16, 2018 - Published by Downloaded from Copyright © 1959 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
effect of the materials lost from the slices would be toincrease self absorption and so to give a lower count-ing rate. This would lead to a spuriously high valuefor glucose uptake. WVe do not feel justified there-fore in attaching significance to the apparently lowereffectiveness of the highest concentrations of someof the test compounds.
The figures for C140. output during the experi-ments with glucose-C14 are shown in the final columnof table II. It should be emphasized that, of the totalglucose absorbed by the controls, only about 10 %was converted to C0. Nevertheless it is clear thatthe test compounds drastically reduced the productionof C140, at whatever level they were applied. Thevery close parallel between the responses of C1402production and glucose uptake to individual concentra-tions of the test compound snould be especially noted.
C. EXPERIMENTS WITH MITOCHONDRIA: Theresponses of plant mitochondria to DNP have beendescribed by several authors (1, 6, 16, 17, 18) and itis clear that P/0 ratios are drastically reduced.These experiments have usually been carried out inthe presence of the hexokinase-glucose trappingsystem, which, although it allows the inhibitorv effectof DNP on phosphate esterification to be shown, maypreclude the demonstration of what is an equally dis-tinctive effect, namely that of stimulated 0.-uptake.In castor bean mitochondria such a stimulatorv effectof DNP has been described, (1), and we have nowinvestigated the ability of other substituted phenolsto bring about this response. For each compounda series of concentrations was applied and responsecurves were constructed from the results of triplicateexperiments. The values in table IV were read offfrom the curves (table IV). Striking stimulationswere induced by all of the phenols; again the di-substituted phenols were particularly effective, with
TABLF IVEFFECTS OF VARIOUS SUBSTITUTED PHENOLS, SODIUMI
AZIDE AND METHYLENE BLUE ON THE OXYGENCONSUMPTION BY MITOCHONDRIA FROM
CASTOR BEAN ENDOSPERM *
MOLAR CONC MOLAR CONCTEST COMPOUND MAXIMUM GIVING GIVING
STIMULATION MAXIMUM 50 %STIMULATION INHIBITION
% of control M X 10-3 M X 10-3o-Nitrophenol 184 5.0 9.8m-Nitrophenol 205 1.3 5.5p-Nitrophenol 204 0.4 5.3Dinitrophenol 204 0.3 6.3o-Chlorophenol 146 3.7 12.0m-Chlorophenol 141 2.5 4.9p-Chlorophenol 189 2.5 7.6Dichlorophenol 182 0.5 2.8Sodium azide 107 0.5 3.1Methylene blue 145 0.3 47.9
the p-, in-, and o-substituted phenols following in thatorder. Methylene blue induced a considerable stim-ulation while azide had virtually no such effect. In-hibitions of 09-uptake ensued when the concentrationsof all the compounds were raised above the stimulatingone.
DISCUSSION
It is clear from the results that all of the substi-tuted phenols, when applied at appropriate concentra-tions have the following properties in common withthe classical uncoupling agent DNP: a) They stim-ulate the respiration of carrot discs; b) at levelsstimulatory to 0-uptake, they inhibit the associatedendergonic process of glucose uptake; and c) theystimulate the 09-uptake of castor bean mitochondriaoxi(lizing °c-ketoglutarate.
It is concluded that such compounds, like DNP,are uncoupling agents.
From their different abilities to induce the re-sponses described (tables I, II and IV), the followingconclusions were drawn about the relative effective-ness of the various phenols. 2,4-dinitrophenol wasthe most effective compound, i.e., it induced responsesat the lowest levels. This was followed in turn by2,4-dichlorophenol, and the para-substituted nitro-,chloro- and bromo-phenols; the ortho-substituted com-poundls were less effective but considerably moreactive than phenol itself. The responses to the addi-tion of various substituents in the molecule thus con-form in general to a pattern which has emerged frommore extensive comparative work with other ma-terials, in which usually only one response, i.e., arespiratory effect or an inhibitory action, has beenmeasured. Among this work may be mentioned par-ticularly that of Stenlid on wheat roots (24), Clowesand Krahl (7, 14) on sea urchin eggs, of Simon andBlacknman (20, 22) on Trichodermiia viride, Leinnamiiinor and Brassica alba and of Field, Martin andField (9) and more recently Deken (8) on yeast.We suppose, with the previous authors, that the dif-ferences in effectiveness most probably reflect, to alarge degree, the ease of penetration of the phenolsto the points of primary attack, which, one would nowsuppose, are the enzymes in the mitochondria concern-ed with electron transfer.
Azide failed to induce respiratory stimulations inthe experiments reported here, although effects whichcan be reasonably ascribed to an uncoupling actionhave been described in plants by Stenlid (23) and byHarley and his colleagues (12). Presumably re-spiratory stimulations can only be dlemonstrated ifthe level of azide required is lower than that at whichthe oxidation enzymes are thlemselves inhibited. Asin Stenlid's experiments (25) methylene blue inducedeffects on respiration and glucose uptake which weresimilar to those due to the phenols. This parallelismhas now been shown to extend to the mitochondria.It should be noticed however, that the mechanism ofaction of methylene blue may be rather different fromthat of the phenols, since it is itself autooxidizable
v ne mitocnonoria were prepared as descriued uyBeevers and Walker and 0,-uptake was measured over a30-minute period with ,c-ketoglutarate (0.005 M) as sub-strate (3).
40
* Irt- -------A -- A----:I,-A 'k-
www.plantphysiol.orgon June 16, 2018 - Published by Downloaded from Copyright © 1959 American Society of Plant Biologists. All rights reserved.

GAUR AND BEEVERS-RESPONSES TO SUBSTITUTED PHENOLS
and may effectively by-pass normal electron transfersteps (13).
In considering the C140, results it should first beemphasized that the carrot tissue contains ample avail-able carbohydrate reserves, and its respiration, witha respiratory quotient close to unity is not increasedby the small amounts of glucose supplied. At alllevels of DNP (and, by inference, of the otherphenols) which stimulate 0,-uptake, C02-output isstimulated to at least the same extent (2). But, atthese stimulatory levels, as shown in table III C1402output was drastically reduced, so that the specificactivity of the respiredl CO, declines with increasingDNP. Such an observation was one of the argumentsused by Porter and Runeckles (19) in support ofan otherwise well founded suggestion that in tobaccoleaves DNP induced the respiration of non-carbohy-drate material. However, the final 2 columns intable II show that there is a striking parallelism be-tween the inhibitory effects of each phenol on glucoseuptake and on C140 release. In the carrot tissuethen, it is possible to ascribe the lowered specificactivity of the CO, directly to a reduced uptake of theradioactive glucose, and it is not necessary to invokea change in respiratory substrate as an explanation.It should be noted that in contrast to the carrot tissueused in these experiments anid those described previ-ously (2) the tobacco leaf tissue had a low RQ (0.7),its respiration was strikingly stimulated by the addedsugar, and the experiments were carried out in 100 %02. Whatever may be the explanation of the re-sponses to DNP in the tobacco tissue, the present re-sults confirm that no change is niecessary in the con-clusions from our previous experiments, (2, 10),namely that the respiratory responses to DNP in thetissues used can be ascribed to an increased rate ofglycolysis brought about as an indirect result of theuncoupling action at the electron transfer level.
SIUMMARYThe effects of a series of substituted phenols on
02-uptake and glucose accumulation by carrot cliscshave been determined. The response to each com-pound was a respiratory stimulation, which reached amaximum and then declined sharply to an inhibitionas the concentration was raised over a rather narrowrange. The striking changes in respiration as theconcentrations were increased were accompanied bythe progressive inhibition of glucose absorption; thoseconcentrations which induced maximally stimulated02-uptake drastically curtailed glucose uptake. Thevarious phenols induced stimulations of O,-uptakeby castor bean mitochondria which were similar tothose induced by dinitrophenol. The various sub-stituted phenols were thus supposed to be acting, withvarying degrees of efficiency, as uncoupling agents.
The relative effectiveness was gauged by compar-ing the concentrations of the compounds required toinduce maximally stimulated respiration, which inmost instances amounted to over 200 % of the controlrates. 2,4-Dinitrophenol was the most effective com-
pound, followed, in turn by 2,4-dichlorophenol,p-nitrophenol, the bromophenols, mn- and p-chloro-phenol, mn- and o-nitrophenol, o-chlorophenol andphenol itself, which induced at best only 20 % stimula-tion.
In parallel experiments with methylene blue it wasshown that effects similar to those of the substitutedphenols were induced. Azide induced only respira-tory inhibitions and corresponding curtailment ofglucose accumulation in carrot discs.
It was also demonstrated that the appearance ofradioactive carbon in the respired CO2 was inhibitedonly to the same degree as the uptake of the glucose-C'4 was itself prevented. For this reason the effectsof substituted phenols on C14O production cannot beconsidered as evidence for a changeover in respira-tory substrate in response to their addition.
This work has been supported in part by theAtomic Energy Commission, (Contract no. AT-(11-1)-330). WXe record our appreciation of thissupport.
LITERATURE CITED1. AKAZANVA, T. and BEEVERs, H. Mitochondria in
the endosperm of the germinating castor bean: adevelopmental study. Biochem. Jour. 67: 115-118.1957.
2. BEEVERs, H. 2,4-Dinitrophenol and plant respiration.Amer. Jour. Bot. 40: 91-96. 1953.
3. BEEVERs, H. and WALKER, D. A. The oxidativeactivity of particulate fractions from germinatingcastor beans. Biochem. Jour. 62: 114-120. 1956.
4. BONNER, J. Limiting factors an(d growth inhibitorsin the growth of Avcnan colcoptile. Amer. Jour.Bot. 36: 323-332. 1949.
5. BON NER. J. Relations of respiration and growth inAvenia colcoptile. Amer. Jour. Bot. 36: 429-436.1949.
6. BONNER, J. and MILLERD, A. Oxidative phosphory-lation by plant mitochondria. Arch. Biochem.Biophys. 42: 135-148. 1953.
7. CLOWEs, G. H. A. and KRAHL, M. E. Studies oncell metabolism and cell division. I. On the rela-tion between molecular structures, chemical proper-ties and biological activities of nitrophenols. Jour.Gen. Physiol. 20: 145-171. 1936.
8. DEKEN, DE R. H. Relations entre la structuremoleculaire et le pouvoir inhibiteur des nitro ethalophenols. Biochim. Biophys. Acta 17: 494-502.1955.
9. FIELD, J., MARTIN, A. W. and FIELD, S. M. Theeffect of 2,4-dinitrophenol on the oxygen consump-tion of yeast. Jour. Cellular Comp. Physiol. 4:405-420. 1934.
10. FRENCH, R. C. and BEEVERS, H. Respiratory andgrowth responses induced by growth regulators andallied compounds. Amer. Jour. Bot. 40: 660-666.1953.
11. GAUR, B. K. Some effects of uncoupling agents onplant tissues and mitochondria. Ph.D. thesis, Pur-due University, Lafayette 1957.
12. HARLEY, J. L., MCCREADY, C. C., BRIERLEY, J. K.and JENNINGS, D. H. The salt respiration of ex-cised beech mycorrhizas. II. The relationship be-
4.31
www.plantphysiol.orgon June 16, 2018 - Published by Downloaded from Copyright © 1959 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
tween oxygen consumption and phosphate absorp-tion. New Phytologist 55: 1-28. 1956.
13. JUDAH, J. D. and WILLIAMs-ASHMAN, H. G. Theinhibition of oxidative phosphorylation. Biochem.Jour. 48: 33-42. 1951.
14. KRAHL, M. E. and CLOWES, G. H. A. Studies oncell metabolism and ceil division. II. Stimulationof cellular oxidation and reversible inhibition ofcell division by dihalo and trihalophenols. Jour.Gen. Physiol. 20: 173-184. 1936.
15. LARDY, H. A. and WELLMAN, H. Oxidative phos-phorylations: role of inorganic phosphate and ac-ceptor systems in control of metabolic rates. Jour.Biol. Chem. 195: 215-244. 1952.
16. LATIES, G. G. The physical environment and oxi-dative and phosphorylative capacities of higher plantmitochondria. Plant Physiol. 28: 557-575. 1953.
17. LATIES, G. G. Respiration and cellular work andthe regulation of the respiration rate in plants.Survey of Biol. Progress. 3: 215-299. 1957.
18. LIEBERMAN, M. an1d BIALE, J. B. Oxidative plhos-phorylation by sweet potato mitochondria and itsinhibition by polyphenols. Plant Physiol. 31: 420-424. 1956.
19. PORTER, H. K. and RUNECKI.ES, VT. C. Some effects
of 2,4-dinitrophenol upon the metabolism of tobaccoleaf discs. Biochim. Biophys. Acta 20: 100-108.1956.
20. SImON, E. W. The action of nitrophenols on respira-tion and on glucose assimilation in yeast. Jour.Expt. Bot. 4: 377-392. 1953.
21. SIMiON, E. W. Mechanisms of dinitrophenol tox-icity. Biol. Reviews 28: 453-479. 1953.
22. SxIroN, E. W. and BLACKMAN, G. E. Studies onthe principles of phytotoxicity. IV. The effects ofthe degree of nitration oIn the toxicity of phenoland other substituted benzenes. Jour. Expt. Bot.4: 235-250. 1953.
23. STENNLID, G. Some notes on the effect of sodiumazide, dinitrophenol and o-phenanthroline uponoxygen consumption in green leaves. Physiol.Plantarum 2: 61-69. 1949.
24. STENL1IrD, G. The effect of 2,4-dinitrophenol uponoxygen consumption and glucose uptake in youngwheat roots. Physiol. Plantarum 2: 350-371.1949.
25. STFNLID, G. AMethylene blue anid TC-Cj dipyridyl,two different types of inhibitors for aerobic metab-olism in young wheat roots. Physiol. Plantarum3: 197-203. 1950.
STUDIES ON THE MECHANISMI OF HERBICIDAL ACTION OF2-CHLORO-4,6-BIS (ETHYLAMITNO)-S-TRIAZINE ", 2
=-CTh- D. E. MORELAND 3, W. A. GENTNER 4, J. L. HILTON 4 AND K. L. HILL 3
NORTH CAROLINA AGRICULTURAL EXPERITMENT STATION AND U. S. D. A., RALEIGH, NORTH CAROLINA
Recent studies have shown that certain triazinederivatives can be used to control weeds in specificcrops at relatively low rates of application (2, 3).At higher rates of application they can be used assoil sterilants for general weed control on non-croppe(dareas. One of these conmpoundIs, 9-chloro-4.6-lbis-(ethylamino)-s-triazine Fsimazin] 5, applied as a pre-emergent spray at 2 to 4 pounds per acre has givenexcellent control of annual weeds in corn for prac-tically the entire growing season (3). Corn appearsto be especially tolerant to the chemical and very high
Received November 24, 1958.2 Cooperative investigations of the Crops Research
Division, Agricultural Research Service, U. S. Depart-ment of Agriculture and the North Carolina AgriculturalExperiment Station. Published with the approval of theNorth Carolina Agricultural Experiment Station as Paperno. 933.
3 Plant Physiologists, Agricultural Research Service,U. S. Department of Agriculture, N. C. State College,Raleigh, N. C.
4 Plant Physiologists, Crops Research Divisioni, Agri-cultural Research Service, U. S. Department of Agricul-ture, Beltsville, Maryland.
5 The experimental samples used in these studies weresupplied by personnel of the Geigy Chemical Corporation,Ardsley, New York.
rates of application have been used successfully insome studies. Barley and other small grains are notable to tolerate pre-emergent applications of even 2pounds per acre of simazin (2, 3).
The objectives of the studies reported herein werea) to determine the effects of simazin on the phloto-chemical activity of chloroplasts isolated from barleyplants and b) to determine whether the inhibitoryeffects of simazin could be overcome by supplyingtreated barley plants with carbohydrates from anexternal source. A preliminary report (4) of thesefindings has been presented.
MATERIALS AND METHODS
STUDIES WITH ISOLATED CHLOROPLASTS: Thechloroplasts used in this studly were obtained from10-day-old greenhouse-grown barley plants (Hordeuinivutlgare L. variety Dayton, CI 9517) using the pro-cedure given by Spikes et al (6). Following the ini-tial isolation they were washed 3 times with 0.5 Msucrose.
The photochemical activity of the isolated chloro-plasts was measured in small beakers. A 10-ml reac-tion mixture was usedl which had the following com-position: 0.0005 'M potassium ferricyanide, 0.01 Mpotassium chloride, 0.17 M sucrose, 0.05 M potassium
432
www.plantphysiol.orgon June 16, 2018 - Published by Downloaded from Copyright © 1959 American Society of Plant Biologists. All rights reserved.

![Auxin Signaling1[OPEN] - plantphysiol.org wielded by the conductor of an orchestra: “When the auxin baton points your way, it’s your turn to play whatevermusicalinstrumentyouhappentobeholding](https://static.fdocuments.us/doc/165x107/5b275bbc7f8b9a4b7e8b59d7/auxin-signaling1open-wielded-by-the-conductor-of-an-orchestra-when-the-auxin.jpg)
















