
Are old-growth forests special? Or just old? How would you know?
50
Are old-growth forests special? Or just old? How would you know?
-
Upload
domenic-hubbard -
Category
Documents
-
view
217 -
download
1
Transcript of Are old-growth forests special? Or just old? How would you know?

Are old-growth forests special? Or just old?
How would you know?

Hemlock-northern hardwood forests
Shade-tolerant canopy species:Acer saccharum (sugar maple)Fagus grandifolia (American beech)Tsuga canadensis (eastern hemlock)
Less tolerant canopy species:Betula alleghaniensis (yellow birch)Acer rubrum (red maple)
Shade-tolerant sub-canopy species:Abies balsamea (balsam fir)Ostrya virginiana (hop-hornbeam)
Shade-tolerant canopy, swamp forestThuja occidentalisPicea glauca
Less tolerant canopy, swamp forestFraxinus nigra

Textbook: Stands at equilibrium composition - frequent, gap-phase -> statistical self-replacement of shade-tolerant dominants - some ‘gap specialists’ persist (Betula alleghaniensis)
Dscn9985.jpg
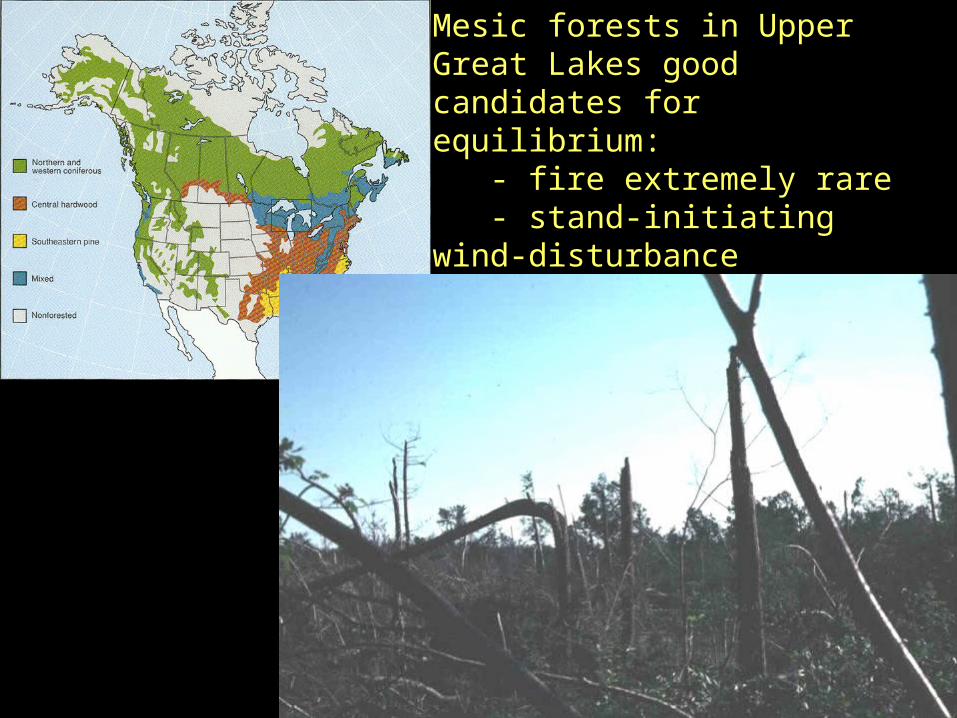
Mesic forests in Upper Great Lakes good candidates for equilibrium: - fire extremely rare - stand-initiating wind-disturbance return time est >2000 yr - large areas with no anthropogenic disturbance

HOWEVER, model rarely tested by direct evidence.
Indirect approaches -- palynology, simulation models, chronosequences -- all have severe limitations in regard to assumptions or scale.

But direct, observational data are difficult to get because SLOW systems.
Dominants can live over 400 years, remain suppressed in understory many decades.

This can be discouraging -- UNLESS...
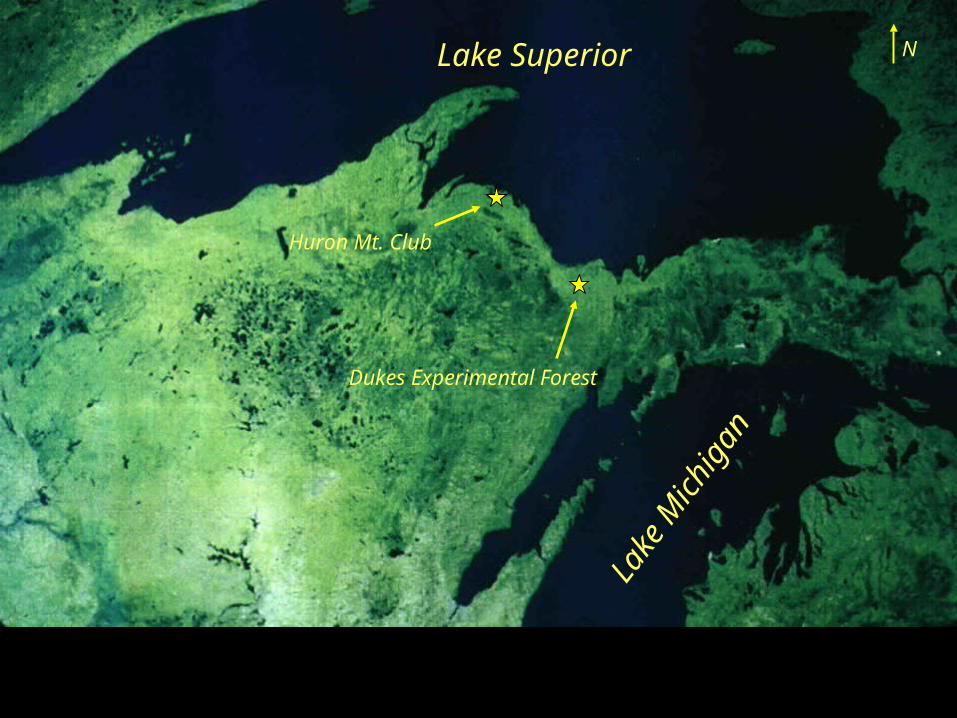
Lake Superior
Lake
Mic
higa
n
Huron Mt. Club
Dukes Experimental Forest
N
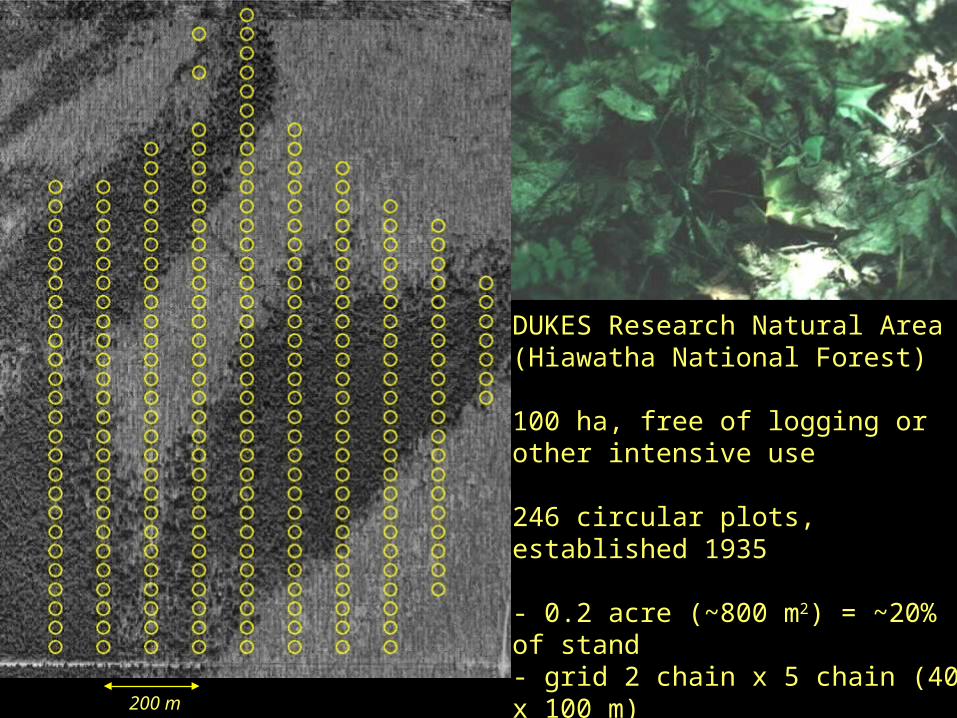
200 m
DUKES Research Natural Area(Hiawatha National Forest)
100 ha, free of logging or other intensive use
246 circular plots, established 1935
- 0.2 acre (~800 m2) = ~20% of stand- grid 2 chain x 5 chain (40 x 100 m)

200 m
High Ca,Acer
High Ca,Acer
Peat,conifers
Peat,conifers Hardpan,
mixed
Hardpan,mixed
podzol,Tsuga
podzol,Tsuga
Habitat is spatially patchy

y vs x
East
No
rth
200 m
2D Graph 2
X Data
0 10 20 30 40 50
Y D
ata
0
5
10
15
20
25
30
'4485 vs '4486
2D Graph 1
X Data
0 10 20 30 40 50
Y D
ata
0
5
10
15
20
25
30
Acer saccharumFagus grandifoliaBetula alleghaniensisTsuga canadensisAcer rubrumOther species
Symbol area scaled to total basal area; maximum basal area = 86.3 m2/ha
20 m
Data set is spatially complicated

1935 1948 1974-1980 1989-2004
n = 238 (8 ‘transitional’plots unsampled)
Stems > 5 in (12.7 cm)dbh tallied by 2.5 cm category
n = 123 (alternating along N-S lines)
Stems > 5 in (12.7 cm)dbh tallied by 2.5 cm category
n = 242
Stems > 0.5 in (1.3 cm)dbh tallied by 0.1 in category
n = 197
Upland: stems > 5 cmmapped full plot, >1 cm8 m radius subplot, remeasured 5-yr cycle.
Swamp: stems >5 cm full plot, >1 cm 8 m sub-plot tallied only
And temporally even more complicated
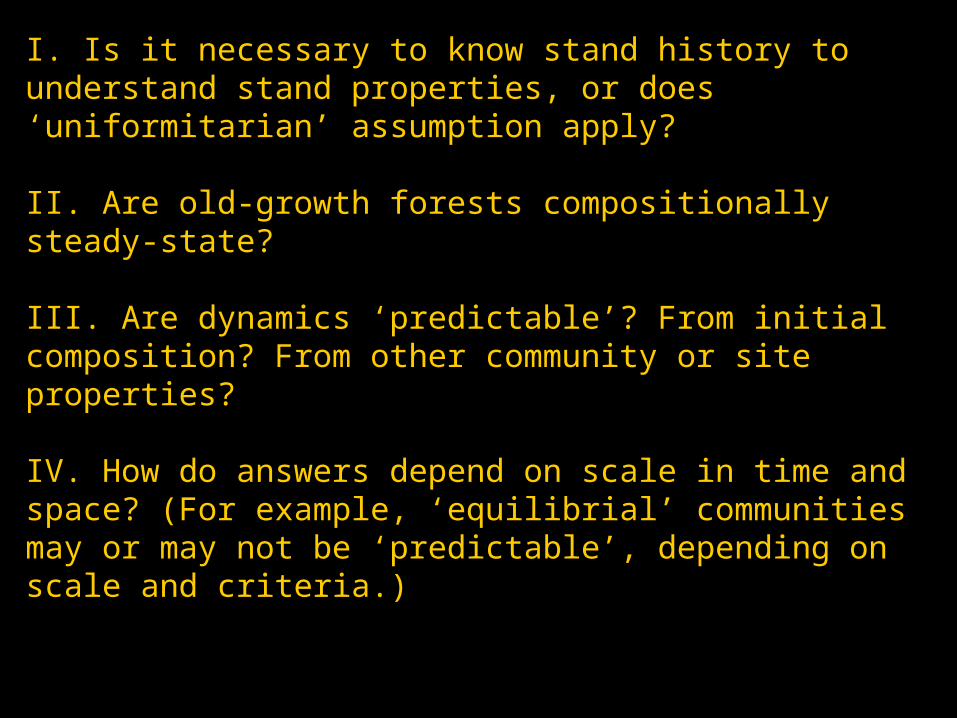
I. Is it necessary to know stand history to understand stand properties, or does ‘uniformitarian’ assumption apply?
II. Are old-growth forests compositionally steady-state?
III. Are dynamics ‘predictable’? From initial composition? From other community or site properties?
IV. How do answers depend on scale in time and space? (For example, ‘equilibrial’ communities may or may not be ‘predictable’, depending on scale and criteria.)
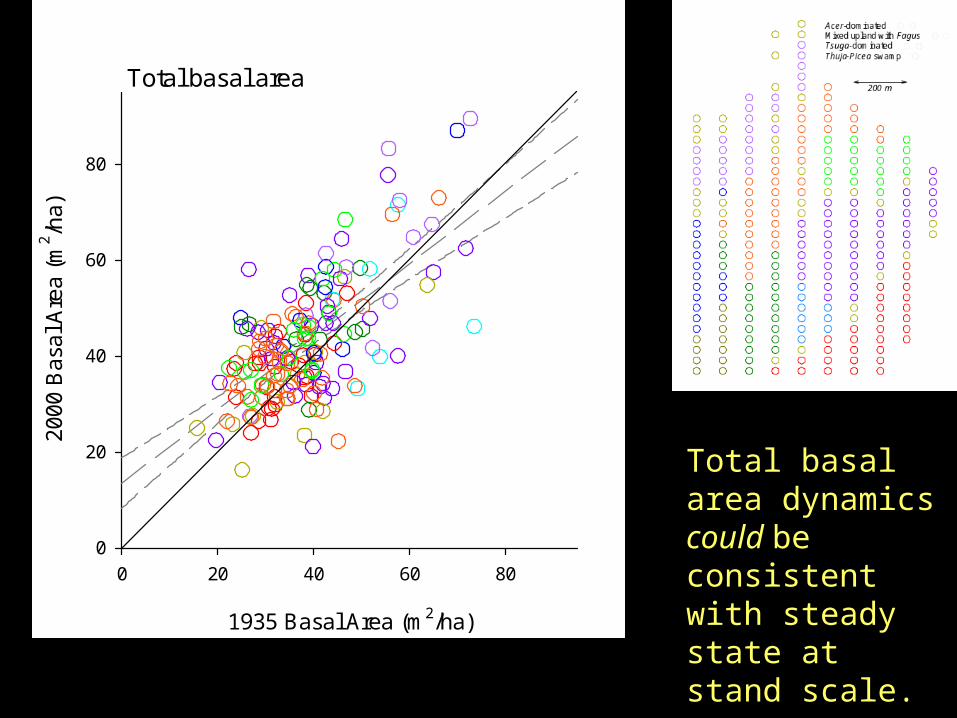
1935 Basal Area (m2/ha)
0 20 40 60 80
20
00
Ba
sal A
rea
(m
2 /ha
)
0
20
40
60
80
Total basal area
Total basal area dynamics could be consistent with steady state at stand scale.

0 10 20 30 40 50
20
00
Ba
sal A
rea
(m
2/h
a)
0
10
20
30
40
50
Acer saccharum
0 10 20 30 40 50 60
0
10
20
30
40
50
60
Tsuga canadensis
1935 Basal Area (m2/ha)
0 2 4 6 8 10 12 14 16 18
20
00
Ba
sal A
rea
(m
2/h
a)
0
2
4
6
8
10
12
14
16
18Fagus grandifolia
1935 Basal Area (m2/ha)
0 20 40 60 80
0
20
40
60
80Thuja occidentalis
Shade-tolerant species are increasing.
But population trends say otherwise

0 5 10 15 20 25 30
20
00
Ba
sal A
rea
(m
2/h
a)
0
5
10
15
20
25
30Betula alleghaniensis
0 2 4 6 8 10
0
2
4
6
8
10Abies balsamea
1935 Basal Area (m2/ha)
0 5 10 15 20 25
20
00
Ba
sal A
rea
(m
2/h
a)
0
5
10
15
20
25Acer rubrum
1935 Basal Area (m2/ha)
0 5 10 15 20 25
0
5
10
15
20
25Fagus grandifolia
Less tolerant species and subcanopy species are decreasing.
Fraxinus nigra

Dukes Experimental Forest:pH
Symbol Area Scaled from minimum to maximum
Easting
No
rth
ing
200 m
Dukes Experimental Forest:Calcium
Symbol Area Scaled from Minimum to Maximum
Easting
No
rth
ing
200 m
Complex spatial structure; strong patterning related to substrate for shade-tolerant species

But not for others; implies more likely role of historical factors?

1935 Basal Area (m2/ha)
0 10 20 30 40 50 60
1948
Ba
sal A
rea
(m
2 /ha)
0
10
20
30
40
50
60
Tsuga canadensis
1948 Basal Area (m2/ha)
0 10 20 30 40 50 60
1977
Ba
sal A
rea
(m
2 /ha)
0
10
20
30
40
50
60
1977 Basal Area (m2/ha)
0 10 20 30 40 50 60
1991
Ba
sal A
rea
(m
2 /ha)
0
10
20
30
40
50
60
1991 Basal Area (m2/ha)
0 10 20 30 40 50 60
2000
Ba
sal A
rea
(m
2 /ha)
0
10
20
30
40
50
60
2000 Basal Area (m2/ha)
0 10 20 30 40 50 60
2003
Ba
sal A
rea
(m
2 /ha)
0
10
20
30
40
50
60
1935 Basal Area (m2/ha)
0 10 20 30 40 50 60
Last
Bas
al A
rea
(m
2 /ha)
0
10
20
30
40
50
60
Further differences among species with higher temporal resolution; trends more or less uniform for shade-tolerant species

1935 Basal Area (m2/ha)
0 5 10 15 20 25
19
48
Ba
sal A
rea
(m
2 /ha
)
0
5
10
15
20
25Betula alleghaniensis
1948 Basal Area (m2/ha)
0 5 10 15 20 25
19
77
Ba
sal A
rea
(m
2 /ha
)
0
5
10
15
20
25
1977 Basal Area (m2/ha)
0 5 10 15 20 25
19
91
Ba
sal A
rea
(m
2 /ha
)
0
5
10
15
20
25
1991 Basal Area (m2/ha)
0 5 10 15 20 25
20
00
Ba
sal A
rea
(m
2 /ha
)
0
5
10
15
20
25
2000 Basal Area (m2/ha)
0 5 10 15 20 25
20
03
Ba
sal A
rea
(m
2 /ha
)
0
5
10
15
20
25
1935 Basal Area (m2/ha)
0 5 10 15 20 25
La
st B
asa
l Are
a (
m2 /h
a)
0
5
10
15
20
25
But Betula dynamics pronouncedly non-stable over time

Fagus: all size classes show large increases
Betula: small size classescollapsing
Many other results consistent to produce first stage results: - forest compositionally dynamic - strong population trends (= predictable?) at stand scale - consistent with successional hypothesis, though no direct evidence of initiating event - implies long-lasting importance of historical events - What kind of events? ‘Intermediate’ disturbance?
=> Requires long-term, observational study

21 July 2002: max winds 150 km/hr, return time ca. 250 yr
Dukes RNA


Initial assessment: consistent with inferences from initial results
- Acer and Fagus, low base-line mortality, high storm mortality;
- Reverse pattern for Betula
- base-line mortality hyper-dispersed, storm mortality strongly clustered, producing gaps of ~30 m).


By 2004, abundant Betula (and other) seedlings on mounds.The next cohort?

Plot trajectories: 1935-2004
NMS Axis One
-2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
NM
S A
xis
Tw
o
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
SO, some ‘predictability’ at population level. But evidence of coherent pattern at community level?
Acer
Fagus
Tsuga
UPLAND PLOTS ONLY
- In full species-space, Mantel test suggests significant predictability, but - Apparent coherence low in ordination space
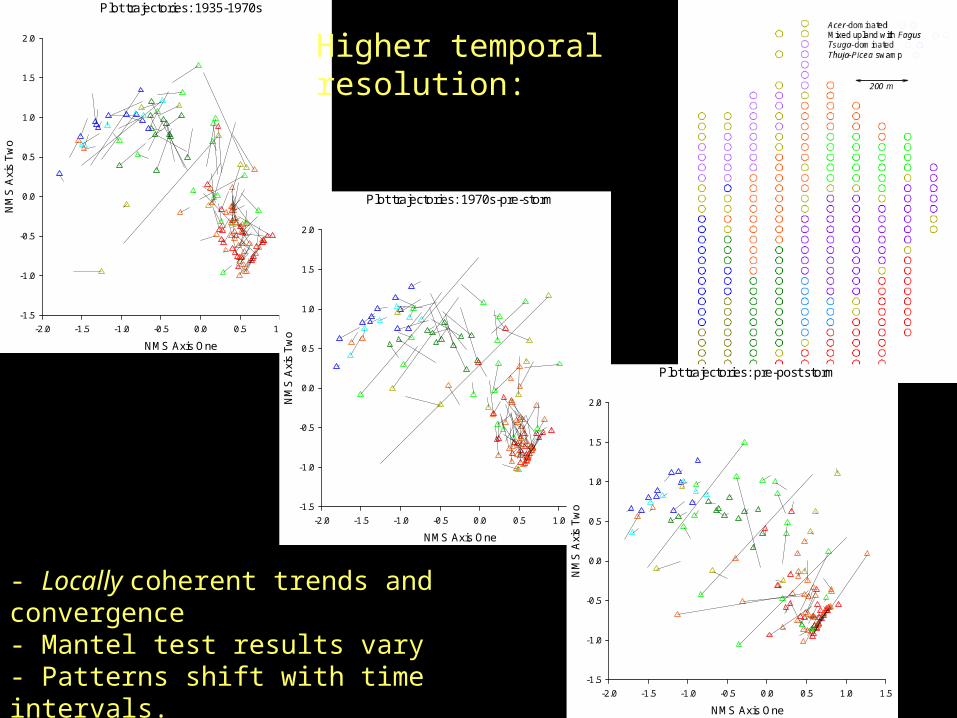
Plot trajectories: 1935-1970s
NMS Axis One
-2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
NM
S A
xis
Tw
o
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
Plot trajectories: 1970s-pre-storm
NMS Axis One
-2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
NM
S A
xis
Tw
o
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
Plot trajectories: pre-post storm
NMS Axis One
-2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
NM
S A
xis
Tw
o
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
Higher temporal resolution:
- Locally coherent trends and convergence - Mantel test results vary- Patterns shift with time intervals.
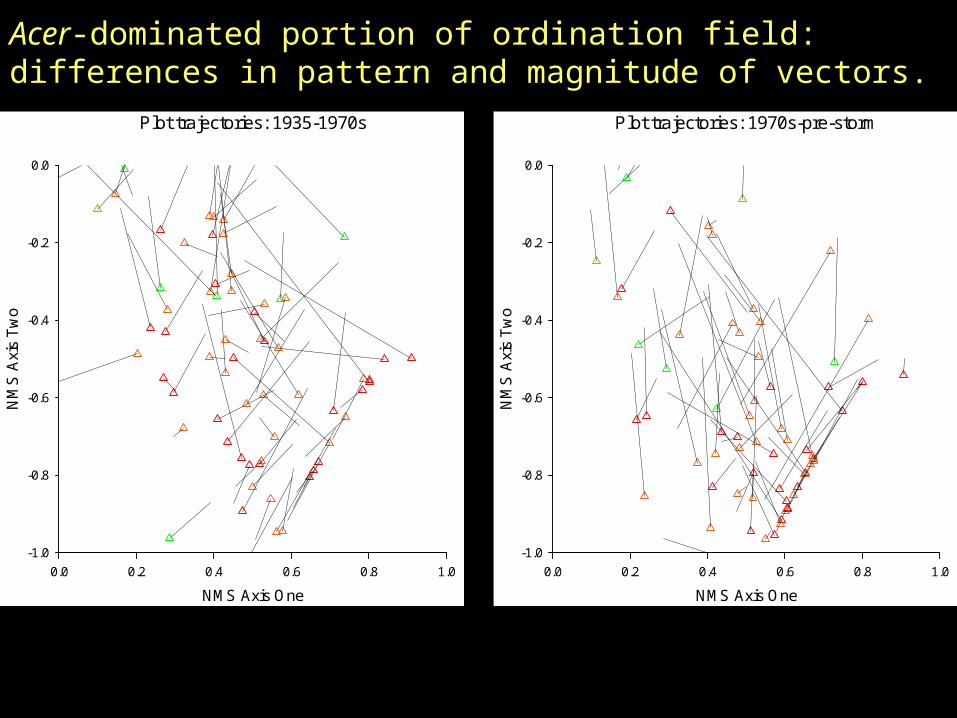
Acer-dominated portion of ordination field: differences in pattern and magnitude of vectors.
Plot trajectories: 1935-1970s
NMS Axis One
0.0 0.2 0.4 0.6 0.8 1.0
NM
S A
xis
Tw
o
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
Plot trajectories: 1970s-pre-storm
NMS Axis One
0.0 0.2 0.4 0.6 0.8 1.0
NM
S A
xis
Tw
o
-1.0
-0.8
-0.6
-0.4
-0.2
0.0

NMS ordination, swamp plots, 1935-1970s
NMS Axis One
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5
NM
S A
xis
Tw
o
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
NMS ordination, swamp plots, 1970s - current
NMS Axis One
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5
NM
S A
xis
Tw
o
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
But little evidence of coherent trajectories in swamp forests.(Mantel test, p=.07, .03)
Thuja

Are different areas or types behaving differently? MRPP comparisons of change vectors
1935-1970s
1970s-2000

City-block distance
0 2 4 6 8
Fre
que
ncy
0.00
0.02
0.04
0.06
0.08
1935
1997-2001
Are plot trajectories convergent?Convergence should change distribution of inter-plot distances in species-space.
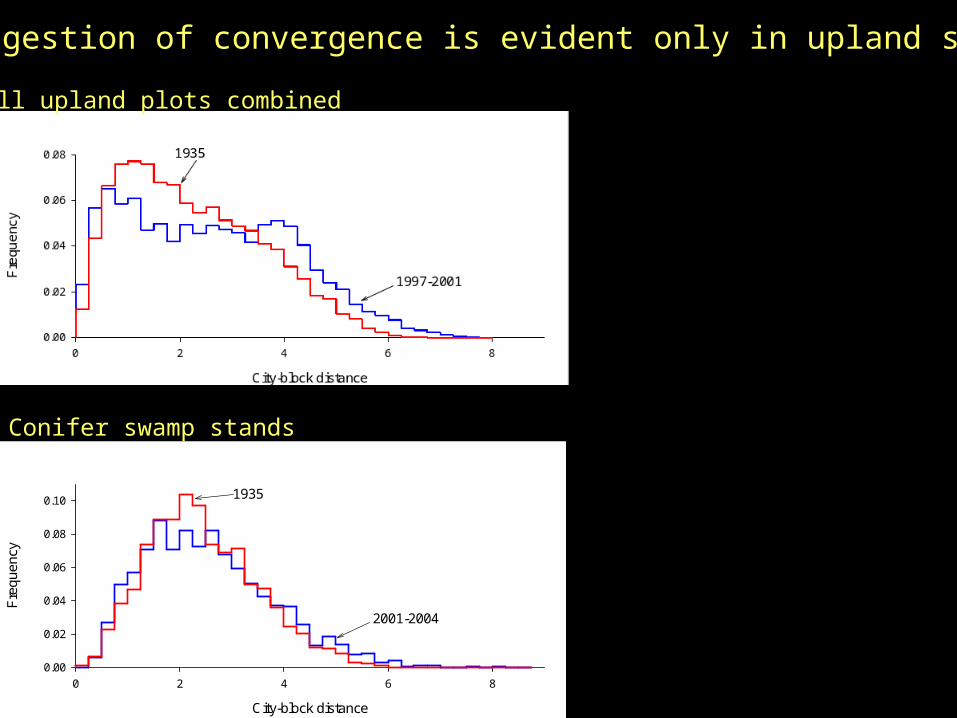
City-block distance
0 2 4 6 8
Fre
quen
cy
0.00
0.02
0.04
0.06
0.08
0.10 1935
2001-2004
But suggestion of convergence is evident only in upland stands
All upland plots combined
Conifer swamp stands
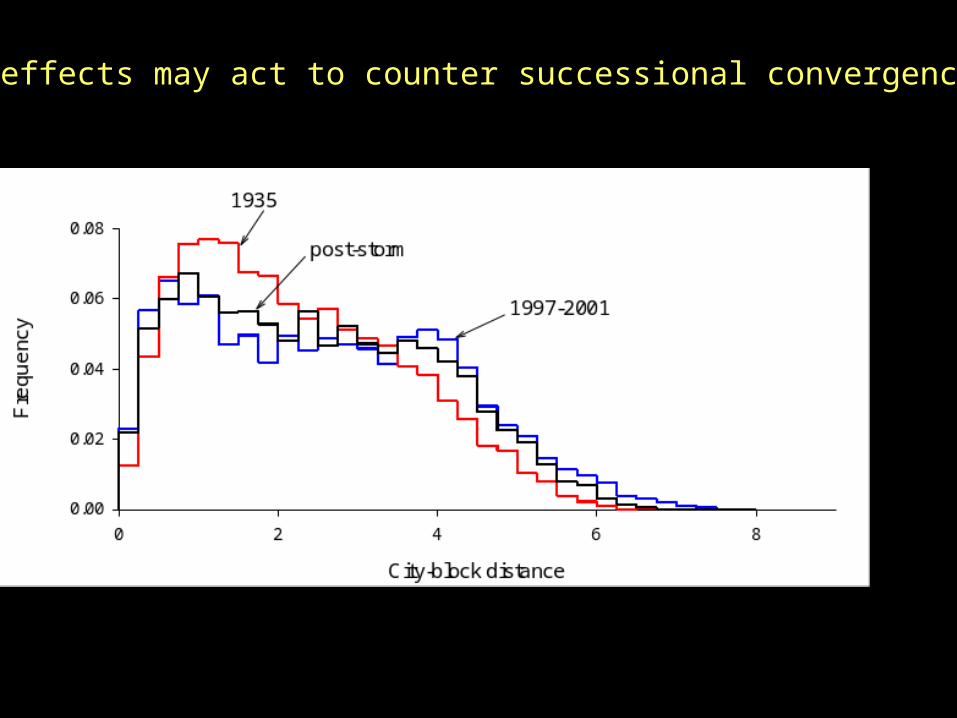
Storm effects may act to counter successional convergence

Summary of ecological results:
- Ancient forests, with no recent disturbance, can be dynamic.
- Dynamics can be influenced by site-conditions, current composition, and infrequent historical events, and all of these factors interact.
- Changes may be predictable at stand scale as results of competitive relationships and environmental change. BUT Current composition relatively weak predictor of trajectories at plot scale.

Methodological and conceptual needs and challenges:
- spatial and temporal scale and resolution are important.
- Historical data sets are of critical value-- but inherited sampling design may constrain how questions can be addressed
SO
- Need data management and analysis tools capable of dealing with variable sampling protocols and intervals and other ‘inherited’ problems with historical data.
- Need focus on development and evaluation of analytical tools for assessing relationships in multi-dimensional data sets over variable scales.

National Science Foundation, Andrew W. Mellon Foundation, US Forest Service, Huron Mt. Wildlife Foundation for support
Thanks to:
Fred Metzger, Eric Bourdo, Jan Schultz for sharing data

AND MANY Bennington College undergraduates for doing the work.

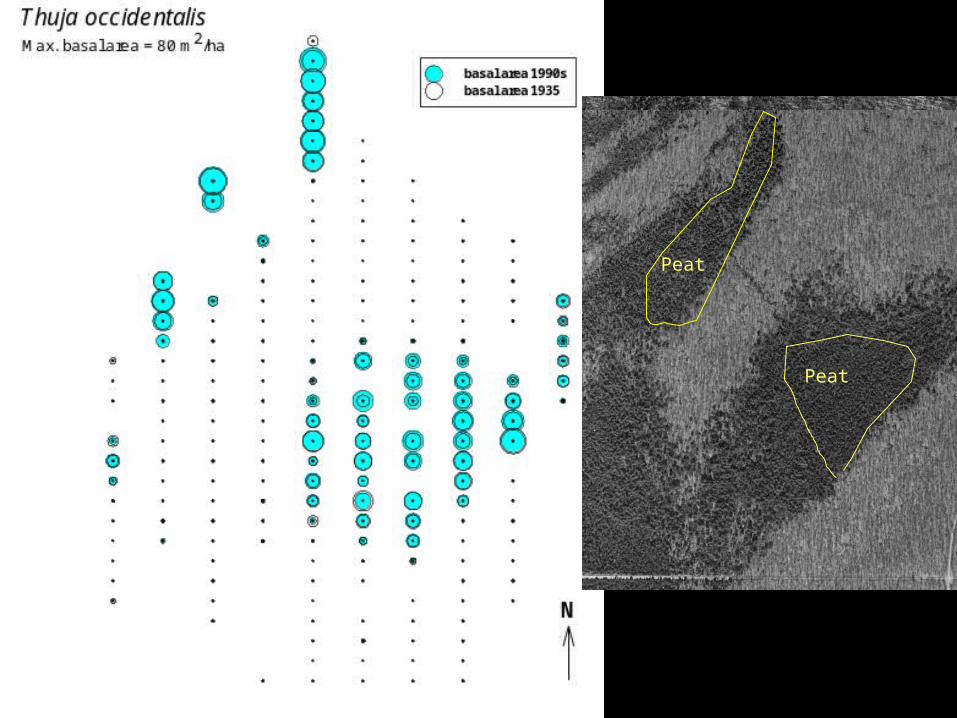
Peat
Peat


Monitoring of individualsallows further demographic insights: growth rates and mortality related.
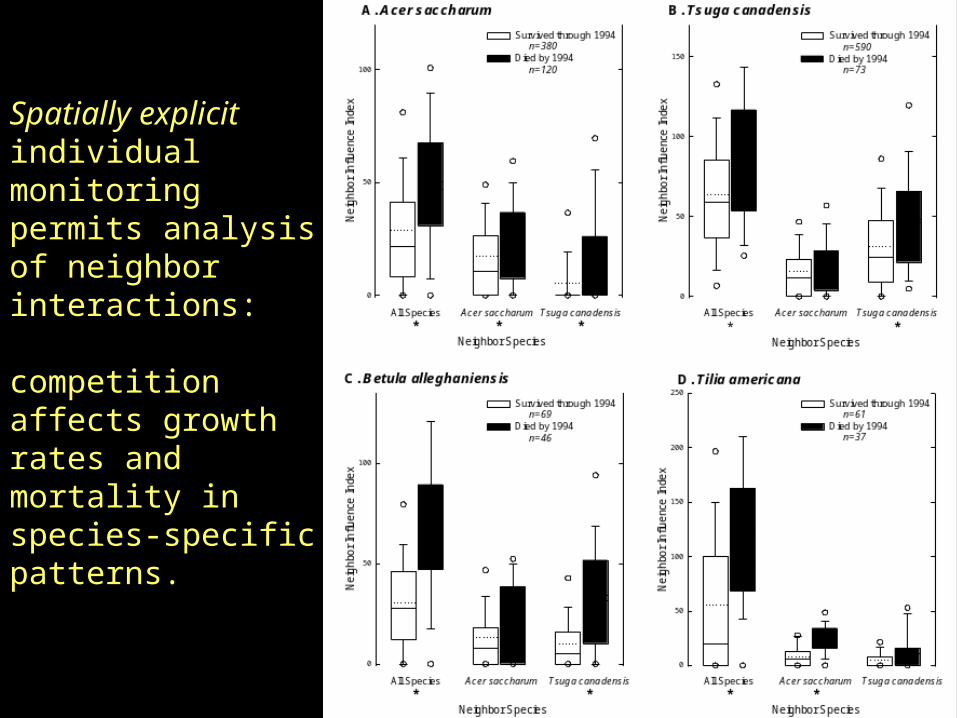
Spatially explicit individual monitoring permits analysis of neighbor interactions:
competition affects growth rates and mortality in species-specific patterns.
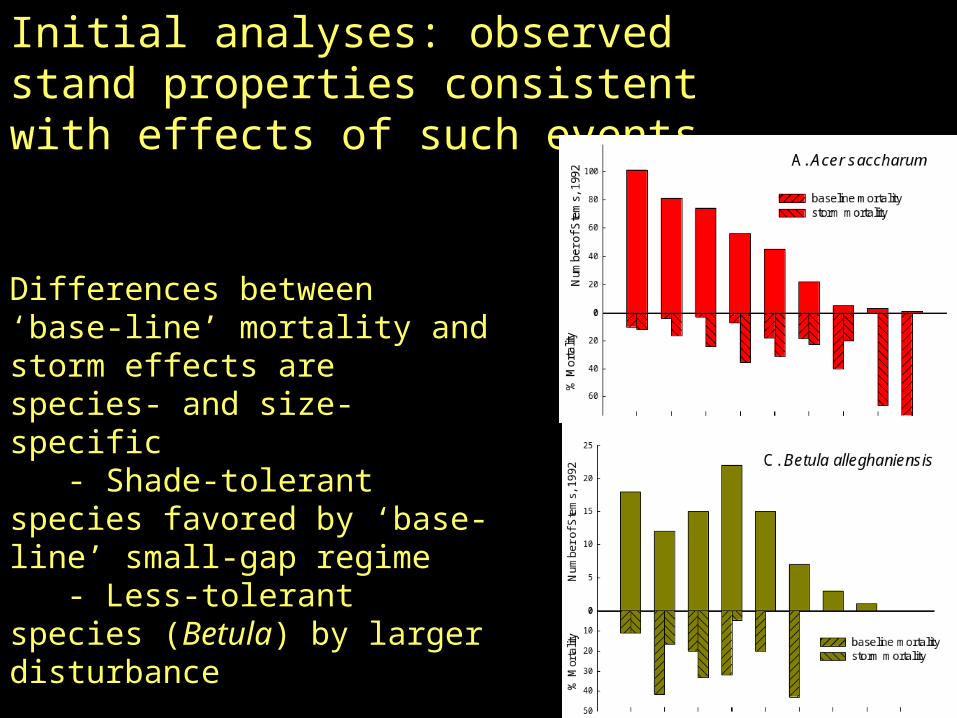
Initial analyses: observed stand properties consistent with effects of such events
Differences between ‘base-line’ mortality and storm effects are species- and size-specific - Shade-tolerant species favored by ‘base-line’ small-gap regime - Less-tolerant species (Betula) by larger disturbance

Across stand at plot scale, spatial pattern complex, but difficult to interpret.

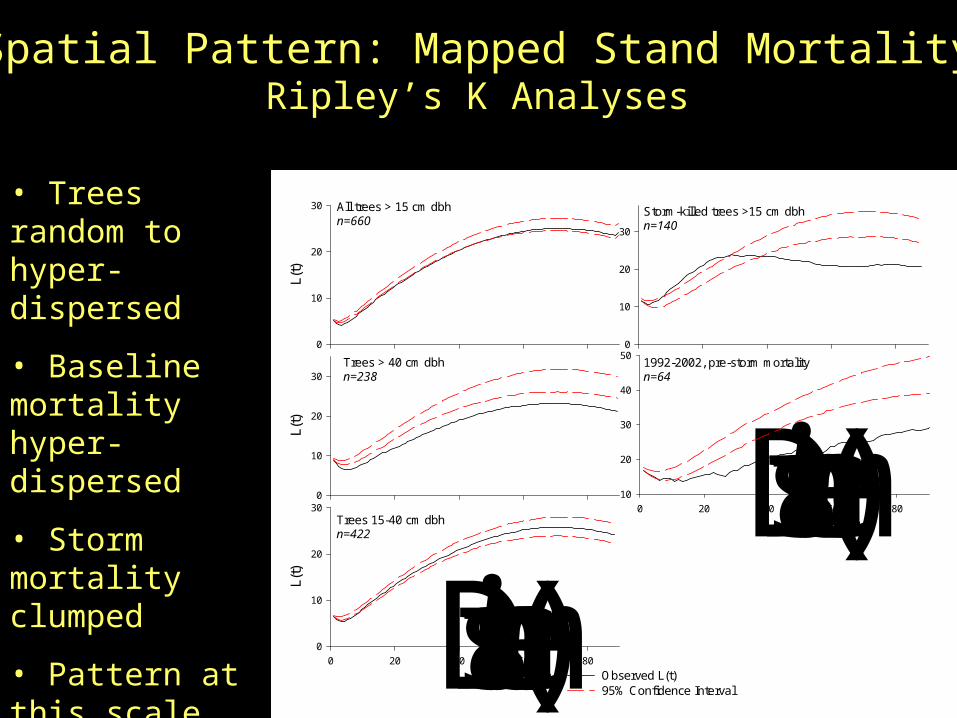
All trees > 15 cm dbhn=660
L(t)
0
10
20
30
Observed L(t)95% Confidence Interval
Storm-killed trees >15 cm dbhn=140
0
10
20
30
Trees > 40 cm dbhn=238
L(t)
0
10
20
30
Trees 15-40 cm dbhn=422
Distance (m)
0 20 40 60 80
L(t)
0
10
20
30
1992-2002, pre-storm mortalityn=64
Distance (m)
0 20 40 60 80
10
20
30
40
50
Spatial Pattern: Mapped Stand MortalityRipley’s K Analyses
• Trees random to hyper-dispersed
• Baseline mortality hyper-dispersed
• Storm mortality clumped
• Pattern at this scale not detectable from plot data

BUT NEW OBSERVATION:
Acer seedlings show extensivestem-layering.

Can layered seedlingsbe released to becomeviable saplings?
YES
In fact, most saplings in‘regeneration patches’appear to have been layered.

SO,- fitness payoff for long- persisting seedlings even greater?- episodic, patchy dynamics even more fundamental?
Even where deer destroy regeneration in smallgaps, large gaps may offer episodic, local escape from browse.


















