Aquaculture 238 (2004) 239 – 247 Www.elsevier.com/Locate/Aqua-Online
9
The combined effects of temperature and salinity on growth and survival of juvenile mud crabs (Scylla serrata Forska ˚l) Ian M. Ruscoe * , Colin C. Shelley, Graham R. Williams Darwin Aquaculture Centre, Fisheries Group, Department of Business, Industry and Resource Development, GPO Box 3000, Darwin, NT 0801, Australia Received 12 December 2003; received in revised form 29 March 2004; accepted 17 May 2004 Abstract An experiment was conducted to determine the effects of temperature and salinity on the growth and survival of the mud crab Scylla serrata, and the combined optima for these parameters during the nursery phase. Instar 2 crablets, mean weight 18.43 F 0.42 mg were individually reared at combinations of temperature (20, 25, 30, and 35 jC) and salinity (0x,5x, 10x, 20x, 30x, and 40x) for 18 days. Crablets were fed an artificial penaeid diet to satiety each day and at harvest were weighed, and the carapace width was measured. All crablets held at 0x, regardless of temperature, were dead the morning after stocking and were therefore excluded from any data analysis. Survival was significantly different among treatments due to temperature but not due to salinities between 5x and 40x. The mean survival for the various temperatures were 36.0%, 98.0%, 96.0%, and 94.0% for temperatures of 20, 25, 30, and 35 jC, respectively. Growth, as measured by individual wet weight at harvest, carapace width, instar 3 intermoult duration, and weight-specific growth rate (WSGR percentage per day), was influenced more by temperature than by salinity, although both had significant effects on all parameters examined. Analyses indicate that maximum WSGR of around 16% day 1 was achieved at 30 jC and 10–20x salinity. Based on the findings of this experiment, S. serrata juveniles from the Northern Territory of Australia should be reared at approximately 30 jC with salinities in the range of 10 – 25x in order to achieve maximal production. D 2004 Elsevier B.V. All rights reserved. Keywords: Mud crab; Scylla serrata; Temperature; Salinity; Growth; Survival; Crablet 0044-8486/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.aquaculture.2004.05.030 * Corresponding author. Tel.: +61-8-8924-4265; fax: +61-8-8924-4277. E-mail address: [email protected] (I.M. Ruscoe). www.elsevier.com/locate/aqua-online Aquaculture 238 (2004) 239 – 247
-
Upload
chuaz-teekaa -
Category
Documents
-
view
385 -
download
7
Transcript of Aquaculture 238 (2004) 239 – 247 Www.elsevier.com/Locate/Aqua-Online

www.elsevier.com/locate/aqua-online
Aquaculture 238 (2004) 239–247
The combined effects of temperature and salinity on
growth and survival of juvenile mud crabs
(Scylla serrata Forskal)
Ian M. Ruscoe*, Colin C. Shelley, Graham R. Williams
Darwin Aquaculture Centre, Fisheries Group, Department of Business, Industry and Resource Development,
GPO Box 3000, Darwin, NT 0801, Australia
Received 12 December 2003; received in revised form 29 March 2004; accepted 17 May 2004
Abstract
An experiment was conducted to determine the effects of temperature and salinity on the growth
and survival of the mud crab Scylla serrata, and the combined optima for these parameters during the
nursery phase. Instar 2 crablets, mean weight 18.43F 0.42 mg were individually reared at
combinations of temperature (20, 25, 30, and 35 jC) and salinity (0x, 5x, 10x, 20x, 30x, and
40x) for 18 days. Crablets were fed an artificial penaeid diet to satiety each day and at harvest were
weighed, and the carapace width was measured. All crablets held at 0x, regardless of temperature,
were dead the morning after stocking and were therefore excluded from any data analysis. Survival
was significantly different among treatments due to temperature but not due to salinities between 5xand 40x. The mean survival for the various temperatures were 36.0%, 98.0%, 96.0%, and 94.0% for
temperatures of 20, 25, 30, and 35 jC, respectively. Growth, as measured by individual wet weight at
harvest, carapace width, instar 3 intermoult duration, and weight-specific growth rate (WSGR
percentage per day), was influenced more by temperature than by salinity, although both had
significant effects on all parameters examined. Analyses indicate that maximum WSGR of around
16% day� 1 was achieved at 30 jC and 10–20x salinity. Based on the findings of this experiment,
S. serrata juveniles from the Northern Territory of Australia should be reared at approximately 30 jCwith salinities in the range of 10–25x in order to achieve maximal production.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Mud crab; Scylla serrata; Temperature; Salinity; Growth; Survival; Crablet
0044-8486/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.aquaculture.2004.05.030
* Corresponding author. Tel.: +61-8-8924-4265; fax: +61-8-8924-4277.
E-mail address: [email protected] (I.M. Ruscoe).

I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247240
1. Introduction
Mud crabs of the genus Scylla have recently been the focus of international
collaborative research, and progress has been made in hatchery technology in order
to supply seed stock (Ruscoe et al., 2004). In the Northern Territory of Australia, these
relatively large crabs are usually found in intertidal and subtidal zones of estuaries and
in mangrove systems (O’Grady et al., 2003). This shallow water tropical environment,
characterised by a 5-month warm, wet season and a 7-month cooler, dry season,
exposes the crabs to wide annual, and often daily, fluctuations in temperature and
salinity.
The apparent wide temperature and salinity tolerances of S. serrata, as evidenced
by its natural range and preferred environment, makes this species an attractive can-
didate for aquaculture. However, it is necessary to characterise the effects of tem-
perature and salinity on crab performance to optimise industry development and farm
practise.
The biological effects of variations in temperature and salinity are considered to be
among the most important factors influencing marine organisms (Ponce-Palafox et al.,
1997). Furthermore, temperature is considered to be the most important modifier of energy
flow and hence growth, while salinity imposes the greatest additional load on metabolic
requirements of aquatic animals (Brett, 1979).
There are numerous studies which focus on the effect of a single environmental variable
on growth and survival of crustaceans (Hill, 1980, S. serrata—temperature; Vijayan and
Diwan, 1995, Penaeus indicus—temperature, salinity, pH; Hai et al., 1998, S. serrata—
salinity; Coman et al., 2002, Penaeus japonicus—temperature; Hamasaki, 2003, S.
serrata—temperature), but relatively few which study the interrelationship of two or
more factors (Hill, 1974, S. serrata—temperature and salinity; Chen and Chia, 1996, S.
serrata—temperature and salinity; Ponce-Palafox et al., 1997, Litopenaeus vannamei—
temperature and salinity), and additionally most studies have used larval animals. The
optimal salinity for survival of S. serrata zoea larvae was reported to be 32x, and
additionally, this variable did not affect the zoeal stage duration (Baylon et al., 2001). It
has also more recently been found that the best temperature for survival of the same
species larvae was 29 jC, and that there was a significant shortening in the duration of
larval rearing with increasing temperature between 23 and 32 jC (Hamasaki, 2003). It is,
however, the immature and stable juvenile stages of development, those not subject to
differentiation, that are considered the best candidates in which to study the governing
mechanisms of growth (Brett, 1979).
The interaction of temperature and salinity on the growth and survival of juve-
nile stone crabs Menippe mercinaria and Menippe adina has been assessed (Brown
et al., 1991). These authors found that temperature (5–40 jC), but not salinity
(10–40x), significantly affects moult frequency, and that both parameters affect
survival.
The aim of this study was to investigate the effect of temperature and salinity on growth
and survival of S. serrata juveniles, and to establish any interactive effects of the same.
The information generated will be a useful guide to industry development by elucidating
optimal water requirements for nursery rearing.

I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247 241
2. Materials and methods
2.1. Experimental animals
The instar 2 crablets were produced from a larval-rearing experiment carried out at the
Darwin Aquaculture Centre (DAC). Megalopa were settled in a 3.8-m-diameter flat-
bottomed tank and were fed on-grown Artemia and an artificial crumble diet (500–750 Amsieved, Vital 12 P. japonicus diet, Higashimaru, Kagoshima, Japan) until the second crab
instar. The crablets were harvested by draining onto a submerged screen. Only fully intact
and active crablets were used in the experiment. Fifty-six individuals, which were not then
used in the experiment, were weighed on the day of stocking to obtain a mean stocking
weight. The crablets were gently dried using blotting paper and were individually weighed
on a Denver AA-250 electronic balance (accuracy to 0.0001 g). All subsequent weighing
occurred in the same way.
2.2. Facilities and protocol
This experiment was carried out in 20 100-l plastic tanks inside an environmentally
controlled room at the DAC. These tanks were arranged in 2 parallel rows of 10.
The 100-l tanks acted as water baths, which were maintained at four experimental
temperatures. These were 20, 25, 30, and 35 jC. The room was cooled so that the water in
the tanks was maintained at 20 jC, unless otherwise heated. All temperatures above 20 jCwere maintained with a 300-W thermostat-controlled immersion heater, and gentle
aeration ensured mixing of the water bath. There were five 100-l tanks maintained at
each temperature in a randomised block design. Within each 100-l tank were spaces for 12
partially submerged 600-ml plastic containers (105 mm diameter� 105 mm deep), which
were used to hold a single crab each. Crabs were stocked at 30 jC and were increased or
decreased at a rate of not more than 3 jC h� 1.
Six salinities were tested. These were 0x, 5x, 10x, 20x, 30x, and 40x. In each
100-l water bath, there were two 600-ml containers at each salinity. Each container held a
single individual and had a lid to prevent evaporation and subsequent changes in
temperature and salinity. In total, 240 individual crabs were used in this experiment.
The various salinity treatments were randomised within each 100-l water bath. Salinity
was either increased or decreased at a rate of not more than 5x h� 1, concurrently with the
temperature adjustment.
Water used to create the various salinities was made from filtered seawater and a
commercial scientific grade marine salt ‘‘Coralife’’ (Energy Savers Unlimited, Carson,
CA, USA). The filtered seawater for all treatments was made up to slightly higher than 40
ppt by the addition of the Coralife salt and was then adjusted with carbon-filtered
freshwater to the various salinities required, except for the 0-ppt treatment, where only
carbon-filtered freshwater was used. This was done so that each salinity received some
commercial salt mixture, some filtered seawater, and some freshwater.
Three times a week, each crab was removed from its container and was placed
immediately into a prepared (temperature- and salinity-adjusted) batch of new water. The
rearing container was cleaned in freshwater, refilled with the appropriate temperature- and

I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247242
salinity-adjusted water, and the crab was then restocked. In this way, a 100% water
exchange was facilitated.
Each day at 1000 h, the crablets were fed 1400–1700 Am sieved Vital 12 artificial diet
(as above) to excess. The following morning, the containers were vacuumed to remove
uneaten food and faeces, and the occurrence of moulting and mortality recorded. Crabs
were grown for 18 days. At harvest, crabs from all tanks were individually weighed (0.001
g), and carapace width measurements were taken (0.1 mm) using vernier calipers.
2.3. Water quality
Temperature and pH of randomly selected containers in each water bath was recorded at
0800 daily with a WP-90 digital water quality meter (TPS, Brisbane, Australia). Salinity of
randomly selected containers within each water bath was measured with a refractometer.
Oxygen was measured with an OxyGuard H01G-Handy Gamma, digital meter (OxyGuard
International, Birkerød, Denmark). Ammonia was measured every 3 days in several
containers that were observed to have the greatest feed consumption using the salicylate
method (Bower and Holm-Hansen, 1980) incorporating a colorimeter.
2.4. Data analysis
Residuals of all data sets were examined to determine a requirement for data
transformation. Block replicates were used to calculate survival, hence these data were
analysed using a two-way ANOVA with a main effects design for the factors temperature
(20, 25, 30, and 35 jC) and salinity (5x, 10x, 20x, 30x, and 40x). Therefore,
comparison of means was not possible for these data set. The proportional survival data
were transformed (arcsine-square root) prior to analysis, although percentage data are
shown in the table for ease of interpretation.
Growth data, recorded as mean weight and carapace width at harvest, as well as weight-
specific growth rate (WSGR, percentage per day; Hopkins, 1992; Evans and Jussila,
1997), was analysed using ANOVAwith a split-plot design where the main plot-factor was
temperature (20, 25, 30, and 35 jC), and the subplot factor was salinity (5x, 10x, 20x,
30x, and 40x). The instar 3 intermoult duration was also recorded and analysed in the
same manner as the growth data, although the data from the 20 jC treatments were not
included in the analysis as so few of the animals in this treatment moulted to instar 4
during the experiment. The third instar was chosen because the majority of crabs reached
the fourth instar. Significant differences between treatments were separated by Tukey’s test
(Sokol and Rohlf, 1981).
3. Results
The mean weight (F S.E.) of the instar 2 crablets at stocking was 18.43F 0.42 mg.
Mean temperature (jC; F S.E.) of the treatments at 0800 h was 20.0F 0.1, 24.9F 0.1,
29.8F 0.1, and 34.8F 0.1. No salinity change from the set values was measured
with the refractometer within the experimental units throughout the experiment.

I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247 243
Ammonia–nitrogen ranged between < 0.1 and 2.0 mg l� 1. Oxygen remained above
4.0 mg l� 1, and pH ranged between 7.78 and 8.16.
Table 1 shows the summarised data for the various treatments in this experiment. All
crablets held at 0x, regardless of temperature, were dead the morning after stocking.
These animals were excluded from any data analysis.
For the remaining data, there was a highly significant difference in survival among
treatments due to temperature (P < 0.0001), but not due to salinity in the range 5–40x(P= 0.1137). The mean survival for the various temperatures were 36.0%, 98.0%, 96.0%,
and 94.0% for 20, 25, 30, and 35 jC, respectively.There were highly significant differences in the mean weight at harvest attributable to
both temperature (P < 0.0001) and salinity (P= 0.0027), but with no interaction effect
(P= 0.38). The crabs held at 30 jC in salinities ranging from 5x to 20xwere the heaviest at
harvest. Similarly, highly significant differences in carapace width at harvest were
attributable to both temperature (P < 0.0001) and salinity (P= 0.027) but with no interaction
Table 1
Means of crablet survival (%), harvest weight (g), carapace width (mm), instar 3 intermoult duration (days), and
weight-specific growth rate (%/day; F S.E.) for various combinations of temperature and salinity used in this
experiment
Temperature
(jC)Salinity
(x)
Survival
(%)
Harvest
weight
(g)
Harvest
carapace
width (mm)
Instar 3
intermoult
duration
WSGR
(%/day)
(N) (days)
20 0 0
5 30 0.042F 0.024 c 6.9F 3.9 f 1 13 4.37F 1.01 ef
10 40 0.045F 0.023 c 7.0F 3.5 f 0 4.99F 0.26 ef
20 40 0.080F 0.040 c 8.1F 4.1 ef 2 7.50F 1.5 7.75F 1.28 def
30 40 0.042F 0.020 c 6.9F 3.4 f 0 4.49F 0.32 f
40 30 0.052F 0.030 c 7.1F 4.1 f 0 5.76F 0.38 ef
25 0 0
5 100 0.142F 0.045 bc 9.9F 3.1 def 10 7.5F 0.31 cd 10.89F 0.72 bcd
10 100 0.156F 0.049 b 10.2F 3.2 cdef 10 7.2F 0.29 bcd 11.48F 0.66 bcd
20 100 0.200F 0.063 b 10.9F 3.4 bcde 10 7.7F 0.45 cd 12.68F 0.77 abc
30 90 0.135F 0.045 bc 9.8F 3.3 def 9 7.6F 0.41 cd 10.56F 0.77 cd
40 100 0.120F 0.038 bc 9.5F 3.0 def 10 8.2F 0.63 d 10.35F 0.23 cde
30 0 0
5 80 0.291F 0.103 a 13.4F 4.7 ab 10 4.3F 0.21 a 15.24F 0.36 ab
10 100 0.337F 0.105 a 13.7F 4.3 a 10 4.3F 0.21 a 16.06F 0.34 a
20 100 0.333F 0.105 a 13.7F 4.3 a 10 4.9F 0.23 ab 15.98F 0.36 a
30 100 0.291F 0.919 a 12.9F 4.1 abc 10 5.5F 0.22 abc 15.10F 0.54 ab
40 100 0.215F 0.068 ab 11.7F 3.5 abcd 10 6.9F 0.28 bcd 13.16F 0.87 abc
35 0 0
5 80 0.195F 0.068 b 11.5F 4.1 abcd 10 5.3F 0.45 abc 12.84F 0.41 abc
10 100 0.197F 0.062 b 11.6F 3.7 abcd 10 4.8F 0.42 ab 12.90F 0.59 abc
20 90 0.197F 0.066 b 11.5F 3.8 abcd 10 6.1F 0.28 abcd 12.59F 0.60 abc
30 100 0.172F 0.054 b 10.9F 3.5 bcde 10 5.3F 0.40 abc 11.93F 0.79 bcd
40 100 0.135F 0.042 bc 10.0F 3.1 def 9 6.6F 0.44 abcd 10.40F 0.93 cde
Means followed by the same letters are not significantly different from one another ( Pz 0.05).
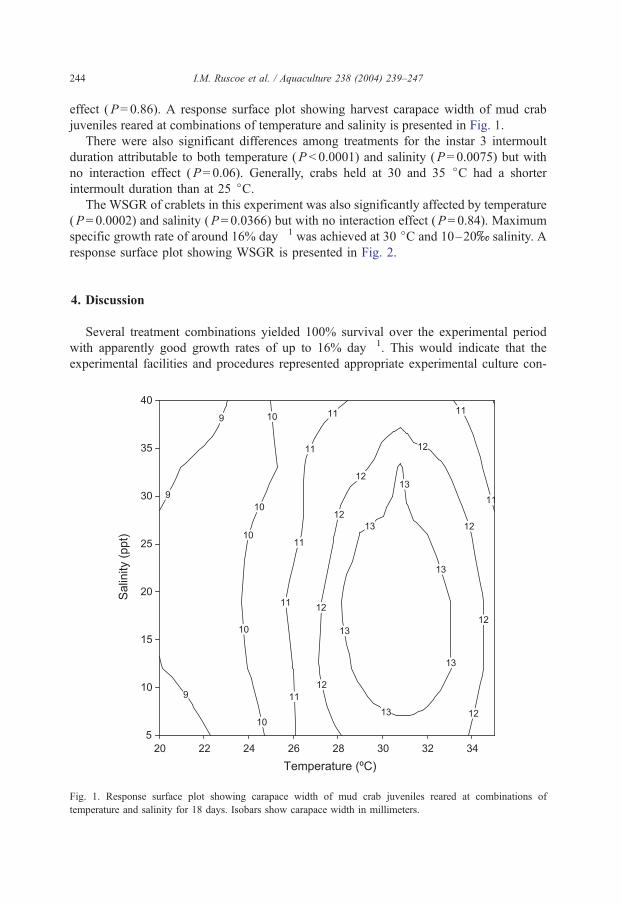
I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247244
effect (P= 0.86). A response surface plot showing harvest carapace width of mud crab
juveniles reared at combinations of temperature and salinity is presented in Fig. 1.
There were also significant differences among treatments for the instar 3 intermoult
duration attributable to both temperature (P < 0.0001) and salinity (P= 0.0075) but with
no interaction effect (P= 0.06). Generally, crabs held at 30 and 35 jC had a shorter
intermoult duration than at 25 jC.The WSGR of crablets in this experiment was also significantly affected by temperature
(P= 0.0002) and salinity (P= 0.0366) but with no interaction effect (P= 0.84). Maximum
specific growth rate of around 16% day� 1 was achieved at 30 jC and 10–20x salinity. A
response surface plot showing WSGR is presented in Fig. 2.
4. Discussion
Several treatment combinations yielded 100% survival over the experimental period
with apparently good growth rates of up to 16% day� 1. This would indicate that the
experimental facilities and procedures represented appropriate experimental culture con-
Fig. 1. Response surface plot showing carapace width of mud crab juveniles reared at combinations of
temperature and salinity for 18 days. Isobars show carapace width in millimeters.

Fig. 2. Response surface plot showing weight-specific growth rate (WSGR) of mud crab juveniles reared at
combinations of temperature and salinity for 18 days. Isobars show WSGR (percentage per day).
I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247 245
ditions for this species. The results have shown that juvenile mud crabs (S. serrata) can
tolerate a wide range of temperature and salinity, although variations from the optimal of
both of these factors will have substantial effects on growth, survival, and production.
Temperature was by far the most influential factor affecting survival and growth in this
experiment (for salinities between 5x and 40x). For every indicator of growth assessed
in this experiment, the temperature effect accounted for greater variance in growth than the
salinity effect. Similar to the results of this experiment, Cadman and Weinstein (1988)
found that both temperature and salinity affected growth of the blue crab Callinectes
sapidus, although temperature had a greater effect.
Crabs held at 20 jC in this experiment were observed to have a reduced activity and
food consumption, although these variables were not quantified. Conversely, crabs held at
35 jC were hyperactive especially at the introduction of food, and their consumption
levels, along with the crabs held at 30 jC, were observed to be higher than at 20 and 25
jC. These observations are consistent with those of Ponce-Palafox et al. (1997) for L.
vannamei, and Niu et al. (2003) for Macrobrachium rosenbergii. Caution should be
exercised however in making use of this temperature optima information, as Wyban et al.
(1995) suggested that the temperature optima for L. vannamei is size-specific and

I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247246
decreases as the prawns grow. In addition, for some species, there may be genotypes
within a population with temperature optima significantly different from the population
mean (Coman et al., 2002). This study utilised siblings from a single mating. Given the
large geographic range of S. serrata, the potential for site specificity in regard to
temperature optima needs to be considered. Larvae of South African S. serrata suffered
high mortality at temperatures above 25 jC (Hill, 1974), whereas Japanese S. serrata
larvae had the best survival at 29 jC (Hamasaki, 2003).
Hai et al. (1998) found that a reduction in salinity affected moult frequency of S.
serrata juveniles, whereby crabs held between 18x and 30x moulted more frequently
than crabs held at 6x and 12x, while all crabs held at 0 ppt died. This is similar to the
results of this experiment. A difference between the two studies however is that our
animals were reduced to zero salinity within 6 h, whereas Hai et al. (1998) reduced salinity
at a rate of 2x day� 1. During the 15 days of acclimation, the crablets in that experiment
actually reached instar 3, whereas in this study, mortality occurred overnight, suggesting
that the rapid dilution undertaken in this experiment was too stressful for the animals. A
slower dilution should be considered for future experiments, especially if salinities below
5x are to be tested. Interestingly, salinity in the range 5–40x had no impact on survival,
hence reducing salinity to as low as 5x in 6 h can occur without mortality, especially at
optimum temperatures.
Haemolymph osmolality of subadult S. serrata (198.58F 35.98 g) has been shown to
reach constant levels after 5 days when crabs are transferred to salinities ranging from 14xto 44x (Chen and Chia, 1997). In that study, osmoregulation was largely accounted for by
ion regulation in salinities below 33.4x (968.6 mOsm kg� 1). Above this salinity, crabs
were osmoconformers. In another experiment, adult S. serratawere held at salinities ranging
from 1x to 42x and did not exhibit any ability to discriminate between various salinities
(Davenport and Wong, 1987). Further experimentation could assess the growth capacity of
this species at salinities below 5x, although this experiment has established that the optimal
salinity for growth of juveniles is around 10–20x. Chen and Chia (1996) found that oxygen
consumption and nitrogen excretion of juvenile S. serrata (0.49F 0.11 g), indicators of
metabolic effort, were both lower at 25x compared to 15x, 20x, and 30x.
Based on the findings of this experiment, S. serrata juveniles from theNorthern Territory of
Australia should be reared at approximately 30 jC and at salinities in the range of 15–25x in
order to achieve optimal growth and survival in nursery culture prior to pond stocking.
Acknowledgements
This work was funded by the Fisheries Research and Development (FRDC) under grant
number 2000/210. The authors would like to thank Keith Newman for valuable technical
assistance throughout this project.
References
Baylon, L., Failaman, A., Vengano, E.L., 2001. Effect of salinity on survival and metamorphosis from zoea to
megalopa of the mud crab Scylla serrata Forskal (Crustacea: Portunidae). J. Asian Fish. Sci. 14, 143–151.

I.M. Ruscoe et al. / Aquaculture 238 (2004) 239–247 247
Bower, C.E., Holm-Hansen, T., 1980. A salicylate–hypochlorite method for determining ammonia in seawater.
Can. J. Fish. Aquat. Sci. 37, 794–798.
Brett, J.R., 1979. Environmental factors and growth. In: Hoar, W.S., Randall, D.J., Brett, J.R. (Eds.), Fish
Physiology. Bioenergetics and Growth, vol. 8. Academic Press, London.
Brown, S.D., Bert, T.M., Tweedale, W.A., 1991. A comparison of some temperature and salinity effects on two
species of stone crab (Menippe spp.) post-settlement juveniles. Am. Zool. 31, 91–98.
Cadman, L.R., Weinstein, W.P., 1988. Effects of temperature and salinity on the growth of laboratory-reared
juvenile blue crabs Callinectes sapidus Rathburn. J. Exp. Mar. Biol. Ecol. 121, 193–207.
Chen, J., Chia, P., 1996. Oxygen uptake and nitrogen excretion of juvenile Scylla serrata at different temperature
and salinity levels. J. Crustac. Biol. 16, 437–442.
Chen, J., Chia, P., 1997. Osmotic and ionic concentrations of Scylla serrata (Forskal) subjected to different
salinity levels. Comp. Biochem. Physiol. 117A, 239–244.
Coman, G.J., Crocos, P.J., Preston, N.P., Fielder, D., 2002. The effects of temperature on the growth, survival and
biomass of different families of juvenile Penaeus japonicus Bate. Aquaculture 214, 185–199.
Davenport, J., Wong, T.M., 1987. Responses of adult mud crabs (Scylla serrata) (Forskal) to low salinity and low
oxygen tension. Comp. Biochem. Physiol. 86A, 43–47.
Evans, L.H., Jussila, J., 1997. Freshwater crayfish growth under culture conditions: proposition for a standard
reporting approach. J. World Aquac. Soc. 28, 11–19.
Hai, T.N., Hassan, A.B., Law, A.T., Shazili, N.A.M., 1998. Effects of reduced water salinity on juveniles of the
mud crab Scylla serrata. International Forum on the Culture of Portunid Crabs. 1–4 December, 1998,
Boracay, Philippines. Asian Fisheries Society, Quezon City, Philippines, p. 57.
Hamasaki, K., 2003. Effects of temperature on the egg incubation period, survival and development of larvae of
the mud crab Scylla serrata (Forskal) (Brachyura: Portunidae) reared in the laboratory. Aquaculture 219,
561–572.
Hill, B.J., 1974. Salinity and temperature tolerance of zoea of the Portunid crab Scylla serrata. Mar. Biol. 25,
21–24.
Hill, B.J., 1980. Effects of temperature on feeding activity in the crab Scylla serrata. Mar. Biol. 59, 189–192.
Hopkins, K.D., 1992. Reporting fish growth: a review of the basics. J. World Aquacult. Soc. 23, 173–179.
Niu, C., Lee, D., Goshima, S., Nakao, S., 2003. Effects of temperature on food consumption, growth and oxygen
consumption of freshwater prawn Macrobrachium rosenbergii (de Man 1879) postlarvae. Aquac. Res. 34,
501–506.
O’Grady, A., Hay, T., Coleman, A., 2003. Mud crab fishery status report 2002. In: Coleman, A. (Ed.), Fishery
Status Reports 2002. Fishery Report No. 69. Northern Territory Government, Department of Business,
Industry and Resource Development, Darwin, Australia, pp. 53–62.
Ponce-Palafox, J., Martinez-Palacios, C.A., Ross, L.G., 1997. The effects of salinity and temperature on the
growth and survival rates of juvenile white shrimp, Litopenaeus vannamei, Boone, 1931. Aquaculture 157,
107–115.
Ruscoe, I.M., Williams, G.R., Shelley, C.C., 2004. Limiting the use of rotifers to the first zoeal stage in mud crab
(Scylla serrata Forskal) larval rearing. Aquaculture 231, 517–527.
Sokol, R.R., Rohlf, F.J., 1981. Biometry. Freeman, San Francisco, p. 859.
Vijayan, K.K., Diwan, A.D., 1995. Influence of temperature, salinity, pH and light on moulting and growth in the
Indian white prawn Penaeus indicus (Crustacea: Decapoda: Penaeidae) under laboratory conditions. Asian
Fish. Sci. 8, 63–72.
Wyban, J., Walsh, W.A., Godin, D.M., 1995. Temperature effects on growth, feeding rate and feed conversion of
the Pacific white shrimp. Aquaculture 138, 267–279.











![APPENDIX978-1-4757-6134... · 2017. 8. 28. · References [1] S. Abiteboul and A. Bonner, Objects views, ACM SIGMOD Int. Conf on the Management of Data, 1991, pp. 238-247. [2] S.](https://static.fdocuments.us/doc/165x107/610f13c1c957580be10675e9/appendix-978-1-4757-6134-2017-8-28-references-1-s-abiteboul-and-a.jpg)