APRIL 1978 Vol. 17/4 Investigative Ophthalmology & Visual...
12
APRIL 1978 Vol. 17/4 Investigative Ophthalmology & Visual Science A Journal of Clinical and Basic Research Articles Training of voluntary torsion Richard Balliet and Ken Nakayama By means of a visual feedback technique, human subjects were trained to make large conjugate cyclorotary eye movements at will. The range of movement increased with training at a rate of approximately 0.8° per hour of practice, reaching 30° at the end of training. Photographs recorded the ability to make voluntary cyclofixations at any amplitude within the subject's range. Cyclotorsional pursuit was also trained, with ability increasing with greater amounts of visual feedback. In addition, torsional saccadic tracking was trained, showing a magnitude vs. peak velocity relationship similar to that seen for normal saccades. Control experiments indi- cate that all of these movements were voluntary, with no significant visual induction. With extended practice, large torsional movements could be made without any visual stimulation. The emergence of voluntary torsion through training demonstrates that the oculomotor system has more plasticity than has generally been assumed, reopening the issue as to whether other movements could also be trained to alleviate the symptoms of strabismus. Key words: eye movements, torsion, saccades, slow pursuit, fixation, orthoptics, oculomotor plasticity c ' yclorotations are defined as rotations about the visual axis of the eye. These rota- tions are considered to be reflexive, with no indication of voluntary control. For example, From the Smith-Kettlewell Institute of Visual Science, Department of Visual Sciences, University of the Pacific, San Francisco, Calif. This paper is based on a Ph.D. dissertation submitted by the first author to the Department of Visual Sciences, University of the Pacific, San Francisco, Calif. 94115. The work was supported in part by NEI Grant EY01582 and the Smith-Kettlewell Eye Research Foundation. Submitted for publication July 6, 1977. Reprint requests: Dr. Richard Balliet, Smith-Kettlewell Institute, 2232 Webster St., San Francisco, Calif. 94115. the eye can undergo a cyclorotation as it moves from one tertiary position of gaze to another, but the amount of this cyclorotation is fixed, being dictated by Listings law. 11 In addition, involuntary cyclovergence has been reported to occur during convergence, 1 and reflexive cycloversions have been demon- strated to occur during lateral head tilt. 14> 18 Cycloversional movements can also be in- duced visually by large field stimuli (25° to 50°). These movements include optokinetic cycloversion, 7 * 8 * 13 fusional cyclovergence, 9 and tonic cycloversions. 8 All of these previ- ously reported cyclorotary eye movements occur either as a necessary consequence of other movements or as part of a sensorimotor 0146-0404/78/0417-0303$01.20/0 © Assoc. for Res. in Vis. and Ophthal., Inc. 303
Transcript of APRIL 1978 Vol. 17/4 Investigative Ophthalmology & Visual...

APRIL 1978 Vol. 17/4
Investigative Ophthalmology& Visual Science
A Journal of Clinical and Basic Research
Articles
Training of voluntary torsionRichard Balliet and Ken Nakayama
By means of a visual feedback technique, human subjects were trained to make large conjugatecyclorotary eye movements at will. The range of movement increased with training at a rate ofapproximately 0.8° per hour of practice, reaching 30° at the end of training. Photographsrecorded the ability to make voluntary cyclofixations at any amplitude within the subject'srange. Cyclotorsional pursuit was also trained, with ability increasing with greater amounts ofvisual feedback. In addition, torsional saccadic tracking was trained, showing a magnitude vs.peak velocity relationship similar to that seen for normal saccades. Control experiments indi-cate that all of these movements were voluntary, with no significant visual induction. Withextended practice, large torsional movements could be made without any visual stimulation.The emergence of voluntary torsion through training demonstrates that the oculomotor systemhas more plasticity than has generally been assumed, reopening the issue as to whether othermovements could also be trained to alleviate the symptoms of strabismus.
Key words: eye movements, torsion, saccades, slow pursuit, fixation,orthoptics, oculomotor plasticity
c' yclorotations are defined as rotationsabout the visual axis of the eye. These rota-tions are considered to be reflexive, with noindication of voluntary control. For example,
From the Smith-Kettlewell Institute of Visual Science,Department of Visual Sciences, University of thePacific, San Francisco, Calif.
This paper is based on a Ph.D. dissertation submitted bythe first author to the Department of Visual Sciences,University of the Pacific, San Francisco, Calif. 94115.The work was supported in part by NEI GrantEY01582 and the Smith-Kettlewell Eye ResearchFoundation.
Submitted for publication July 6, 1977.Reprint requests: Dr. Richard Balliet, Smith-Kettlewell
Institute, 2232 Webster St., San Francisco, Calif.94115.
the eye can undergo a cyclorotation as itmoves from one tertiary position of gaze toanother, but the amount of this cyclorotationis fixed, being dictated by Listings law.11 Inaddition, involuntary cyclovergence has beenreported to occur during convergence,1 andreflexive cycloversions have been demon-strated to occur during lateral head tilt.14> 18
Cycloversional movements can also be in-duced visually by large field stimuli (25° to50°). These movements include optokineticcycloversion,7* 8* 13 fusional cyclovergence,9
and tonic cycloversions.8 All of these previ-ously reported cyclorotary eye movementsoccur either as a necessary consequence ofother movements or as part of a sensorimotor
0146-0404/78/0417-0303$01.20/0 © Assoc. for Res. in Vis. and Ophthal., Inc. 303

304 Balliet and NakayamaInvest. Ophthalmol. Visual Sci.
April 1978
Fig. 1. Photographs showing a tonic voluntary cy-clotorsion of 20°. Bars drawn in across the irisesare similar to those used for photographic analysis.Vertical reference marker (present in all photo-graphs) is seen at right.
reflex. There are no instances where one seespurposeful cyclofixations, or smooth and sac-cadic torsional tracking, phenomena whichare common to voluntary horizontal or verti-cal eye rotations.
In this paper we will demonstrate that if anadequate sensory feedback loop is created,providing subjects with a very sensitive indi-cator of their own eye position, voluntary cy-clotorsional eye movements can be trained.Methodology and results will be presentedwhich will substantiate that once this re-lationship is established, large voluntary cy-clorotations can be learned and broughtunder relatively fine control.
Tonic cycloversion
MethodsSubjects. We report the successful training of
three subjects. Our first subject, M. N. (age 31)had unilateral intermittent exotropia, his left eyedeviating 30°. Normally he allowed this exotropiato be manifest. Otherwise, he had normal stere-opsis, eye movements, and corrected acuity. Ini-tially both eyes were tested for possible deviationsfrom Listing's law. They showed no systematicdeviation, with the mean torsion in each eye beingpredictable by the direction of gaze16 We did see,
however, a larger than average torsional responsevariability in all positions of gaze (S.D. = 0.75°).We reasoned that although small, M. N.'s tor-sional variability was clearly perceptible andperhaps sufficient to extend further by exploratorytraining procedures. As our first and pilot subjectwe worked closely with him, up to 5 hr/week overa period of 13 months, testing many methods todevelop an apparently optimal approach, Then,having found successful procedures for M. N. (seelater), we trained two normal individuals whoshowed no oculomotor abnormalities.
Subjects R. B, (one of the authors, age 30) andC. H. (age 26) were chosen because of their highmotivation. These subjects had normal stereopsisand eye movements and normal or corrected nor-mal acuity. Contact lenses were worn by C. H.Both subjects were trained and tested over aperiod of 2 months.
Training and testing procedure. Many differenttraining procedures were used with M. N. Wereport only upon the final method used most suc-cessfully with him and exclusively with C. H. andR, B. In general, the following procedures wereconducted during both training and testing.
During visual feedback training, the subjectwas seated in a dark room with the left eyealways occluded. Torsional head movement wasfixed to within 6' of arc by a full-mouth impres-sion bite plate (see Ditchbum10). Fixating a tar-get with the right eye, the subject activated abrief vertical electronic flash (11 x 14°), leadingto the formation of a clear vertical afterimage.By horizontally sliding the base of the biteplate mechanism, the subject then imaged theafterimage parallel to a vertical luminous real-line of equal visual angle which was viewed at100 om distance in primary position. The real-line was a partially occluded "white" 110 voltluminescent panel. It was bisected by a 15'circular black fixation point. The subject con-trolled the luminance of the real-line over arange of 0.2 to 1.0 log foot-lamberts to preventvisual suppression of the afterimage line by thetarget line (or vice versa). He also controlled aThis rotation angle could be observed by the sub-ject directly and/or by a digital voltmeter readout.The meter had an accuracy of ±4 ' .
The subject was instructed to keep his afterim-age matched parallel only by cyclorotating his eyeto the real-line which he progressively rotatedmore and more (left or right) from the vertical astraining progressed. Usually 20' of arc incrementswere used. No instructions were given as to whattype of eye movement was to be used to arrive at

Volume 17Number 4 Training of voluntary torsion 305
this position, e.g., slow pursuit, saccades, nystag-mus. Such matches which could be subjectivelyheld for a minimum duration of 5 sec were definedas "cyclofixations." Subjects were encouraged totrain equally in both incyclotorsional and excyclo-torsional directions, training usually at the rate of1 hr/day.
Photographic measurement of eye torsion. Ob-jective measures of cyclofixations were obtainedfrom over 1,500 photographs taken with a 35 mmNikon F-2 motor-driven camera with lenses andbellows attachments equivalent to a lens with afocal length of 400 mm. Two Vivitar 292 strobeflash units provided illumination. High-resolutioncolor photographs on Kodachrome II film weretaken at 100 cm distance with a first-surface mirrormounted at 30 cm distance in front of the subjectat a 45° angle, so positioned to be 5.0° off thesubject's primary visual axis. At this angle sys-tematic inaccuracies due to perspective distortionare minimal, not exceeding 3%. (See Appendix ofNakayama and Balliet16.)
The camera was activated by a button whichcould be pressed by the subject while he simulta-neously held a subjectively determined cyclofixa-tional match. Fixed to the bite bar was a verticalphotographic reference marker positioned nearthe outer can thus. Thus each photograph con-tained all the information necessary to determineeye torsion, eliminating errors due to possiblecamera or film misalignment. During photo-graphic analysis slides were magnifided 12 timesin size by a Kodak Carousel projector. The imagewas rear-projected onto a horizontal table screenmade of translucent "white" Mylar. Landmarksused in measurement were two identifiablelimbal-scleral blood vessel junctions which bor-dered a diameter of the iris and approximatelybisected the pupil. The orientation of a lineformed between these two points relative to thestationary vertical marker was used in determiningobjective cyclotorsion. Fig. 1 illustrates an exam-ple of this technique, showing a 20° voluntary cy-clotorsional difference between the two photo-graphs; the bar represents the line joining the twolandmarks. All photographic cyclotorsional mea-surements were defined relative to average eyeposition when the observer was relaxed and notmaking voluntary eye torsions. Torsional mea-surements from the same photograph were re-peatable to within ±5 ' of arc.
ResultsTraining of cyclofixation. The training of
cyclorotary eye movements requires a com-
C.H.
R.B.
10 15 20 25HOURS OF TRAINING
Fig. 2. Training of voluntary tonic cycloversion forsubjects C. H. and R. B. Hours of training at 1hr/day vs. the total range (degrees) of psy-chophysically measured torsion is plotted by thesmall symbols. These are maximum tonic rangeswhich can be held for a 5 sec duration. The threelarge squares and the one large circle are simulta-neous measures obtained photographically.
bination of optimal stimulus arrangements aswell as physical and mental effort. Very smalldifferences in the brightness of the afterim-age and the real line led to the inhibition ofthe dimmer by the brighter, necessitatingfine control of the intensity of the real-line toensure the visibility of both. Attentive con-centration was necessary to refrain from mov-ing the afterimage with an attempted headmovement. For example, during initial train-ing, two subjects broke bite plates, attempt-ing unsuccessfully to rotate the afterimage bymaking lateral head movements.
Slowly, however, the subjects succeededin the task, learning to increase their range oftorsional eye movements with greater andgreater facility. Fig. 2 shows the remarkablysteady increase of voluntary torsion as afunction of hours of practice. Small symbolsrepresent the maximum range of torsionaldifference measured psychophysically. Thelarge symbols (square and circle) show thetorsional ranges derived from photographicmeasurements; standard errors are small,within the size of the symbols. At the end oftraining, subjects had the ability to make cy-clofixations at any cyclotorsional amplitudewithin the ranges specified.
Subject C. H. acquired a cyclofixation

306 Balliet and NakayamaInvest. Ophthalmol. Visual Sci.
April 1978
12 10 8 6 4 2 0 2 4 6 8 10 12INCYCLO VERTICAL EXCYCLO
OBJECTIVE TORSION-FIXATING R.E.. DEC
Fig. 3. Conjugacy of trained torsional eye move-ments. Comparison of the torsion in each eye(measured photographically) as the subject ismatching the afterimage tilt to designated anglesof the comparison line. Viewing is monocularthrough the right eye. Solid line represents per-fect conjugacy.
range of 26.5° (12.0° incyclo, 14.5° excyclo) inapproximately 30 hr. Subject R. B. acquireda cyclofixation range of about 20° (9.0° in-cyclo, 11.0° excyclo) in approximately 25 hr.Both subjects showed about an 0.8°/hr train-ing rate. These torsional ranges should not beconsidered as the maximum possible; forexample, C. H. is clearly not at an asymptoticlevel at the end of 35 hr of training. SubjectM. N. had a final objectively measured cy-clofixation range of approximately 20° (10.0°incyclo, 10.0° excyclo). His training resultsare not shown because of differing proce-dures used in training of 250 hr over a periodof 13 months (as described in the Subjectssection).
It should be emphasized that the effort re-quired to make these torsional movementsdecreased dramatically over the trainingperiod. Near the end of training, for exam-ple, torsional fixations of up to 6° to 8° were"virtually effortless" and could be made withthe same ease as with ordinary fixations. Italso appears that the acquired capacity tomake these movements is long-lasting. M.
N., for example, could return to previouspositions after having been away for 2months. Furthermore, after approximately 1year without any intervening experience,both C. H. and R. B. still had the capacity tomake torsional movements of about one-halftheir previous torsional range.
Trained cyclofixation is not visually in-duced. As mentioned earlier, very smallamounts of tonic eye torsion can be inducedby the static inclination at a multiple-linedfield.8 As such, these movements appear todiffer from the currently described move-ments in two significant respects. (1) Theyare much smaller (less than 1.0°). (2) They areinvoluntary. In order to determine howmuch of the measured cyclofixation was vi-sually induced by the real stimulus line weconducted a control experiment. After train-ing, subjects were instructed to relax andsimply look for 30 sec at the real-line, whichwas held at different fixed lateral angles in3.0° increments. This was done both with andwithout the usual vertical afterimage line. Atthe end of the 30 sec, photographs of theright (fixating) eye were taken. The time in-terval of 30 sec was comparable to the longesttime required to obtain a afterimage/real-linematch using voluntary torsion.
Nearly identical results were found wheth-er or not the subject had an afterimage. Theyindicate that without voluntary effort there isessentially no effect; eye torsion remainsnearly constant for any angle of the line. OnlyR. B. showed a slight trend, attaining a max-imal torsional response of less than 1°. Theseresults are similar to that found by Crone8
and demonstrate that in our stimulus situa-tion optostatically induced eye torsion is veryweak.
Binocular aspects. The training of torsionwas an exclusively monocular task, with notraining or measurement of the occluded lefteye. It is of interest to know the rotationalstatus of this occluded eye as the trained eyeundergoes large torsional changes. For ex-ample, cyclotorsional eye movements havebeen reported to accompany vergence,1 withextorsion increasing for increasing amounts ofconvergence. This relationship raises the

Volume 17Number 4 Training of voluntary torsion 307
question as to whether our trained torsionsare simply an expression of this relation orwhether they indeed represent a new andseparate type of eye movement.
To deal with this possibility, we separatelyphotographed the rotations of each eye fordifferent cyclofixations made with the righteye. We did this photographing of the righteye as described in the Methods sectionabove and photographing the occluded lefteye as it viewed darkness. During experi-mentation, M. N.'s left eye was exodeviatedby 30°, necessitating the camera to be ap-proximately positioned to photograph alongthe visual axis. Thus, for a cyclotorsional fixa-tion made by the right eye as defined by aparticular angle of the afterimage match, therotations of either eye could be separatelymeasured and compared. The degree of tor-sional conjugacy between the right and lefteyes was determined by plotting the torsionof the viewing eye against the torsion of theoccluded eye for different matching angles ofthe real-line. Fig. 3 shows a near-perfect cor-respondence between the cyclorotations ofthe two eyes in all three subjects.
Horizontal movements of the left eye weredetermined by measuring the relationshipbetween the corneal reflex and the pupil cen-ter.10 This position remained constant towithin ±0.5° (experimental error ± 0.2°) overthe whole range of torsional movements ofthe right eye. This exceptional stability ofhorizontal eye position of the occluded eyeduring eye torsion effectively rules out anyexplanation of our results based on changes invergence.* All our voluntary cyclotorsions
*All subjects demonstrated an exceptional degree ofhorizontal and vertical stability of the occluded eye and anear-perfect conjugacy of torsion. This was even true forM. N.'s left eye, which was exodeviated by 30°. Consid-ering the presumed primary actions of the oblique andvertical muscles in 30° abduction,6' l9 it may behypothesized that either the combination of neural in-puts to the six muscles to produce torsion is greatly al-tered for M. N. or that the primary mechanical actions ofthe muscles remain invariant in an eye-based coordinatesystem. In the former hypothesis, torsion would be or-ganized at a high level in the oculomotor system, thusrequiring a "torsion command" system to rotate the eyearound the visual axis, independent of gaze direction. .
are conjugate wheel-like cycloversions whichoccur around the visual axes.
Subjective observations. During the train-ing and testing of all voluntary cycloversions(including voluntary cycloversional slow pur-suit and saccadic tracking—see below), sub-jects experienced sensations of lateral rolling.They also experienced during initial trainingoccasional nausea, headache, and body fa-tigue. Most notable and most consistent wasthe sensation of the body turning in the samedirection as the eye, with concomitant sensa-tions of stationary objects appearing to rotatein the opposite direction when environmen-tal cues were available. A more quantitativedescription of these and related illusions ispresented elsewhere.3
Dynamic cycloversions
Voluntary conjugate eye movements canbe divided into two distinct categories: slowpursuit and saccadic tracking. Because ourtrained cyclofixations were clearly voluntary,it was of interest to see whether theirdynamic characteristics were comparable tovoluntary horizontal and vertical movements,specifically in the degree to which they hadthe appropriate smooth and saccadic charac-teristics. Alternatively, it was also of interestto see whether the dynamics were compara-ble to the nystagmic pattern normally seenduring reflexive cyclorotations.7' 8> l3
Method. In order to characterize the ability tomake voluntary dynamic cycloversions in detail,motion picture records were taken in one sessioneach for two subjects, M. N. and C. H. This oc-curred after only 10 additional training sessions of1 hr each. The subjects' ability to make large cy-clofixations gave them the ability to make dynamiccyclorotary pursuit or tracking of a moving targetwithin this short duration. Considered alone, thetraining of dynamic tracking was not difficult;however, it was made difficult by the bright il-lumination required for high-resolution photogra-phy. The over-all apparatus and procedure werealso similar to the methods described earlier, withdifferences noted below.
Motion picture technique. Measures of dynamictorsion were made from photographs taken with a16 mm Bolex H16 reflex movie camera running ateither 24 frames/sec for smooth or 64 frames/sec
308 Balliet and NakayamaInvest. Ophthalmol. Visual Sci.
April 1978
C.H.
TIME. SECONDSTIME. SECONDS
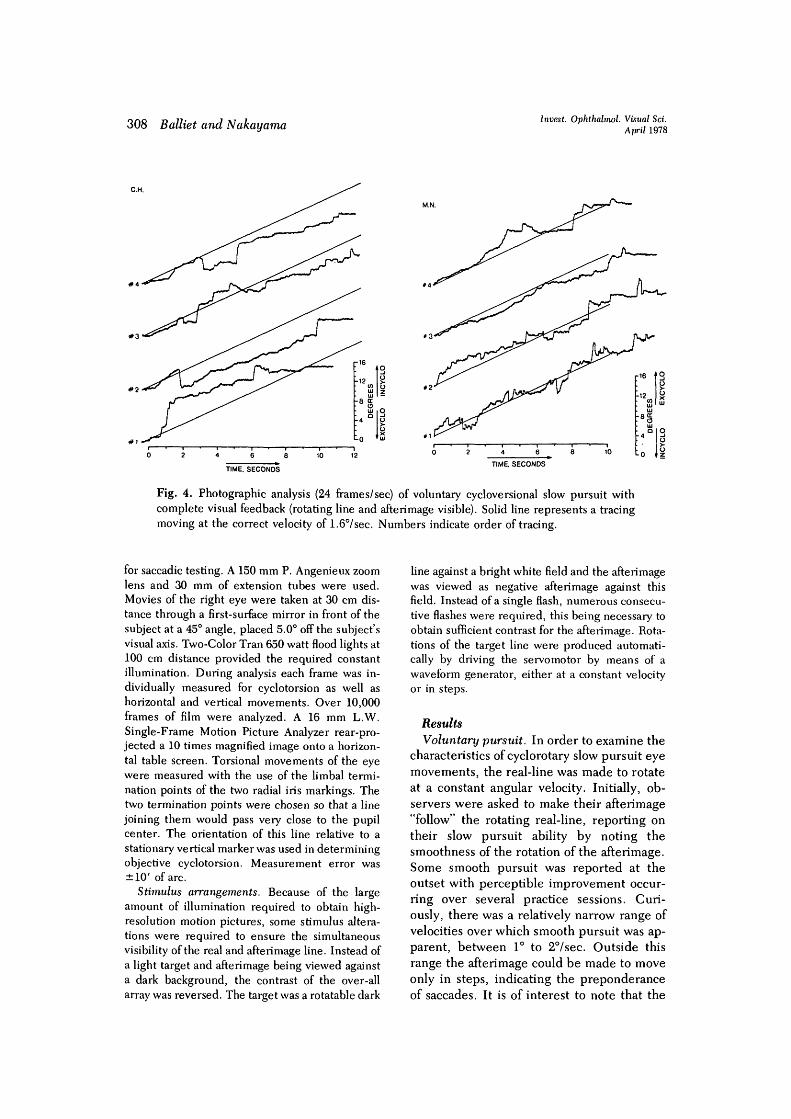
Fig. 4. Photographic analysis (24 frames/sec) of voluntary cycloversional slow pursuit withcomplete visual feedback (rotating line and afterimage visible). Solid line represents a tracingmoving at the correct velocity of 1.67sec. Numbers indicate order of tracing.
for saccadic testing. A 150 mm P. Angenieux zoomlens and 30 mm of extension tubes were used.Movies of the right eye were taken at 30 cm dis-tance through a first-surface mirror in front of thesubject at a 45° angle, placed 5.0° off the subject'svisual axis. Two-Color Tran 650 watt flood lights at100 cm distance provided the required constantillumination. During analysis each frame was in-dividually measured for cyclotorsion as well ashorizontal and vertical movements. Over 10,000frames of film were analyzed. A 16 mm L.W.Single-Frame Motion Picture Analyzer rear-pro-jected a 10 times magnified image onto a horizon-tal table screen. Torsional movements of the eyewere measured with the use of the limbal termi-nation points of the two radial iris markings. Thetwo termination points were chosen so that a linejoining them would pass very close to the pupilcenter. The orientation of this line relative to astationary vertical marker was used in determiningobjective cyclotorsion. Measurement error was±10' of arc.
Stimulus arrangements. Because of the largeamount of illumination required to obtain high-resolution motion pictures, some stimulus altera-tions were required to ensure the simultaneousvisibility of the real and afterimage line. Instead ofa light target and afterimage being viewed againsta dark background, the contrast of the over-allarray was reversed. The target was a rotatable dark
line against a bright white field and the afterimagewas viewed as negative afterimage against thisfield. Instead of a single flash, numerous consecu-tive flashes were required, this being necessary toobtain sufficient contrast for the afterimage. Rota-tions of the target line were produced automati-cally by driving the servomotor by means of awaveform generator, either at a constant velocityor in steps.
Results
Voluntary pursuit. In order to examine thecharacteristics of cyclorotary slow pursuit eyemovements, the real-line was made to rotateat a constant angular velocity. Initially, ob-servers were asked to make their afterimage"follow" the rotating real-line, reporting ontheir slow pursuit ability by noting thesmoothness of the rotation of the afterimage.Some smooth pursuit was reported at theoutset with perceptible improvement occur-ring over several practice sessions. Curi-ously, there was a relatively narrow range ofvelocities over which smooth pursuit was ap-parent, between 1° to 2°/sec. Outside thisrange the afterimage could be made to moveonly in steps, indicating the preponderanceof saccades. It is of interest to note that the

Volume 17Number 4 Training of voluntary torsion 309
TIME. SECONDS TIME. SECONDS
Fig. 5. Photographic analysis (24 frames/sec) of voluntary cycloversional slow pursuit withpartial visual feedback (afterimage visible, no rotating line). Numbers indicate order of trac-ing.
M.NC.H
#8
TIME. SECONDS TIME, SECONDS
Fig. 6. Photographic analysis (24 frames/sec) ot voluntary cycloversional slow pursuit with norotary visual feedback (neither afterimage or real target line is visible). Numbers indicate or-der of tracing.
optimal velocity for optokinetic following of arotating stimulus is also in this slow range.13
For a photographic analysis of pursuit, aconstant 1.6°/sec angular velocity was usedover a total rotation of 16°, rotating clockwisefrom -8° to +8° for M. N. and the reverse forC. H.
Three different conditions were used: (1)complete visual feedback utilizing both the
afterimage and the target line, (2) partial vi-sual feedback with the afterimage only, and(3) no rotary or tilted visual feedback—thisconsisting of a single black fixation dot. Fig. 4to 6 show time records of the torsional eyemovements, obtained photographically, forthe three conditions for each subject. Thesloping solid line in Fig. 4 corresponds to amovement at the velocity of the target line.

310 Balliet and Nakayama Invest. Ophthalmol. Visual Sci.April 1978
.5—r~1.0 1.0
TIME. SECONDS TIME. SECONDS
Fig. 7. Photographic analysis (64 frames/sec) of voluntary cycloversional saccadic tracking: 4°step stimulus. Arrow represents "correct" destination of the saccadic eye movement. Numbersindicate order of tracing.
Traces are numbered in the order taken.With complete visual feedback both subjectsdemonstrated a moderate ability to makevoluntary cyclotorsional slow pursuit move-ments. Particularly accurate are trace 2 for C.H. and trace 3 for M. N., where movementsat the actual target velocity were common(see Fig. 4). Intermixed with these pursuitmovements are small corrective saccades.
With partial visual feedback (Fig. 5), sub-jects had only a single afterimage line and ablack fixation dot, and they were required to"imagine" a line moving at the same velocityas before (1.6°/sec). Under this condition,subjects had more difficulty, showing lesstime in performing smooth movements andshowing more saccades instead. It should benoted, however, that C. H. was able to makesome long, slow pursuit movements in theabsence of a moving line using only the af-terimage. Trace 5 begins with a movementhaving a velocity of 1.6°/sec. Her other traces6 and 7 have episodes with a velocity of about3°/sec. M. N. also shows some slow pursuit ofabout 3°/sec in his trace 7. Such findingsmight be considered analogous to experi-ments showing smooth horizontal trackingwith afterimages alone.12
Ordinarily, horizontal and vertical pursuiteye movements cannot be made in the ab-sence of perceived movement. Except underspecial circumstances,22 a stationary observerviewing a static scene is not capable of pro-
ducing any pursuit, producing only a series ofsaccades. As a further point of comparisonbetween trained torsional pursuit and ordi-nary pursuit, we had our subjects attempt tomake cyclotorsional pursuit movementswithout any stimulus other than a single fixa-tion dot. Under these conditions, movementsconsisted mostly of a series of steplike sac-cades, with M. N. having peculiar overshoots(see below) but with little evidence of smootheye movements (Fig. 6). This inability tomake torsional pursuit in the absence of per-ceived torsional movement provides yet an-other comparison to the known characteris-tics of voluntary horizontal and vertical pur-suit movements.
It should be emphasized that even thoughboth subjects showed little evidence ofsmooth pursuit without a moving target, theycould rotate their eyes over a large range, upto 24° (Fig. 6, M. N.), without any visualstimulus. This underscores the fact that thetorsional eye movements are not visually in-duced. Subjects need no stimulus at all tovoluntarily rotate their eyes.
Voluntary saccades. During the trainingand testing of voluntary cyclorotary saccades,the target line was moved in steps of 4°, 8°,and 16°. M. N. used a clockwise rotation from0° to +4° or +8°, or -8° to +8°, whereas themovements were in the opposite direction forC. H. Subjects were instructed to track thevertical line using their afterimage. Motion

Volume 17Number 4 Training of voluntary torsion 311
M.N,
C.H.
L 0
.5 1.0 1.0 1.5
TIME. SECONDSTIME. SECONDS
Fig. 8. Photographic analysis (64 frames/sec) of voluntary cycloversional saccadic tracking: 16°step stimulus. Arrow represents "correct" destination of the saccadic eye movement. Numbersindicate order of tracing.
picture speed was increased to 64 frames/sec,maximizing the temporal resolution thatcould be obtained photographically. Figs. 7and 8 show sample records of the 4° and 16°saccades.
It is apparent that voluntary saccadic cyclo-torsional eye movements exceeding 12° arepossible (Fig. 8, C. H.). From the records itcan be seen that the voluntary saccades canbe single, sequential, or nearly overlapping.Accuracy of saccades, however, appears lessthan for ordinary saccades, showing errors asgreat as 4°. The records from M. N. weremore irregular, some showing frequent drifts(0.2° to 1.0°/sec) with a magnitude of 1.0° orless.
During the 4° voluntary cycloversional sac-cadic tests (Fig. 7), either the subjects greatlyovershot the correct stimulus position or, inthe case of M. N., the appropriate eyepositions were achieved by making smallfixation drifts and/or small saccades. Bothsubjects reported that an appropriate 4° sac-cade was virtually impossible to make andhad to be coaxed to even attempt such a
movement. The 8° and 16° saccadic taskswere judged to be far easier. These resultssuggest a possible inability to accurately con-trol cyclotorsional saccadic trackings of lessthan 4°.
C .H. demonstrated normal glissadic over-shoots when compared to normal horizontalsaccades,23 and the saccadic tracking of M. N.often showed large dynamic overshoots of upto 2.0°, following the normal velocity-ampli-tude relationship for saccades2 (see Fig. 8).Although M. N. is a unilateral intermittentexotrope, it should be remembered thatthese traces are for his nonstrabismic eye.Also, we found no unusual horizontal eyemovements for either of his eyes when mea-sured by an infrared photoelectric technique,accurate to 20'. We have one possible expla-nation. Bahill and Stark4 have found that theoccurrence and magnitude of dynamic over-shoots increases with fatigue. M.N. had re-ported feeling very tired for several daysprior to and through the day of these tests.In addition, both subjects reported largeamounts of saccadic suppression. Afterimages

312 Balliet and NakayamaInvest. Ophthalmol. Visual Sci.
April 1978
1000:
1.0 10MAGNITUDE, DEC
1.0 10MAGNITUDE. DEG
Fig. 9. Comparison of voluntary cycloversional and horizontal saccades in terms of magnitudevs. peak velocity. Dots represent this analysis for voluntary cyclotorsional saccades. The re-gression line represents a best fit for a comparable analysis of horizontal saccades obtained byBahill et al.3 with bars indicating the range of their data.
disappeared during and somewhat after vol-untary saccades greater than about 4° or 5°,thus diminishing retinal feedback which re-lated eye position to the subject. Unusualoculomotor control may have occurred be-cause of these two factors.
Systematic analysis indicates that horizon-tal and vertical saccades are highly stereo-typed. This is best understood by consideringthe well-documented magnitude vs. peak ve-locity function for normal saccades,3' 24 a re-lationship which emphasizes the fact thathorizontal and vertical saccades do not comein all sizes and velocities but that for anygiven size there is a characteristic velocityand duration. The dots in Fig. 9 show peakvelocities of torsional saccades as a function ofamplitude. The velocities have been multi-plied by a factor of 1.43, compensating forthe fact that our frame rate of 64 frames/sec is effectively limited in its high-frequen-cy response to 32 Hz. Such a limitationcan be expected to reduce saccadic peak ve-locity by 30%,3 thereby requiring a multipli-cative factor of 1.43 to obtain the best esti-mate of the true peak velocity. Comparativedata from horizontal saccades are plottedas a regression line with error bars,these data being derived from Bahill et al.,3
utilizing a high-frequency recording sys-tem (1,000 Hz cutofi). What stands outis the remarkable congruence of the twosets of data, suggesting a common or analo-gous origin for both types of saccades.
TRAINED DYNAMIC CYCLOVERSION IS NOT VISU-
ALLY I N D U C E D . R o t a t i n g s t imu l i c an visual ly
induce cyclotorsion.7'8> 13 As a further controlto clarify the voluntary nature of our cyclo-torsional slow pursuit and saccadic tracking,we had subjects view the rotating real-line asin all previous test situations, instructing thesubjects to relax and simply observe therotating stimulus (either 1.6°/sec pursuitramp; 4° or 16° saccadic step). This was donefor both observers, with and without the af-terimage line. Under all conditions no traceof systematic nystagmus or any movementswere found. No movements exceeded 1°.
Discussion
Voluntary nature of the trained cyclover-sions. Reflexive cyclotorsions have beenknown to exist for over 100 years. What isnew* is the existence of a cyclotorsional eyemovement which fails to fall into any class ofreflexive movements, showing instead char-acteristics which are decidedly voluntary.Two sets of observations should further em-phasize this point. First is the finding thatneither static tilt nor rotary motion issufficient to visually induce our reported cy-
* Although not reported as such, we note that Noji,17 in asupposed study of induced cycloversions, used afterim-ages paired with real-lines, reporting increasing "induc-tion" up to 15° only after 5 successive days of "exercise".Rather than induction, we consider this result to be theprobable training of voluntary torsion.

Volume 17Number 4 Training of voluntary torsion 313
clorotary movements. Second and equallyimportant is the fact that large cyclorotaryeye movements, up to 26.5°, can be made inthe absence of any tilted or rotary stimuluswhatever (Fig. 6). Vision alone cannot drivethe large torsional movements; they requirevoluntary effort.
Plasticity and limits of plasticity in the ocu-lomotor system. The emergence of voluntaryeye torsion through training indicates an un-suspected plasticity of the human oculomotorsystem. For example, under ordinary cir-cumstances voluntary fixations are thought tobe governed by Listing's law such that, interms of its resting position, the eye rotateswith only two degrees of freedom, with thethird degree of freedom (torsion) determinedby the horizontal and vertical direction ofgaze. The nonmechanical nature of this onedegree of rotational constraint implies theexistence of a supranuclear neural network,assuring a fixed and remarkably systematicdegree of eye torsion for each gaze di-rection.15' 24 Clearly, the existence of volun-tary torsion demonstrated in this paper indi-cates that such a seemingly "hard-wired"network can be modified or overridden.
It should be noted that despite the largedegree of oculomotor plasticity demonstratedby our training procedures, there are someaspects of the newly trained movementswhich are obviously familiar and clearly notnew. For example, the conditions underwhich one finds slow pursuit are similar, bothfor torsional as well as for normal pursuit,with greater amounts of slow pursuit appar-ent with greater amounts of movement in-formation. For saccades, the magnitude vs.peak velocity relation affords the most sys-tematic comparison. This relation is a con-tinuous one for normal saccades25 and is ap-plicable to the fast phase of nystagmus aswell.20 What is of interest is the fact that ournewly trained movements should also followthis rule very closely. We suggest that thissimilarity is not coincidental but reflects thefact that all saccades, including torsional sac-cades, are mediated by a primitive and rela-tively "hard-wired" neural network, original-ly designed for the quick phase of nystagmus.
Clinical applications. The success of ourtraining procedure raises the issue as towhether other types of oculomotor organi-zations are also possible and, if this is so,whether or not such reorganization wouldhave value in the treatment of eye movementdisorders. The sensory feedback techniquewhich we used to acquire this control is inprinciple similar to other methods whichhave used sensory feedback to gain controlover responses which are thought to be in-voluntary; these include heart rate, visceralfunction, peripheral blood flow, and thepermanent return of muscle control in pa-tients with various manifestations of dis-turbed neuromotor control. (For a review ofrecent developments in sensory feedback,see Schwartz and Beatty.21) One of the keyfeatures for successful training of these in-voluntary responses is the availability of ahigh-gain sensory feedback signal indicatingminute changes in direction of the desiredmotor response. With this information, thesubject gradually gains access to previouslyinaccessible visceral or motor control mech-anisms.
Considering this recent work on the train-ing of involuntary responses and our ownwork on the training of voluntary torsion, it isof interest to determine whether othersterotyped oculomotor routines are also mal-leable. For example, both Hering's law andListing's law are well-known rules of theoculomotor system, reflecting the fact thatthe pattern of innervation to the muscles isvery orderly and seemingly permanent.Normally, this neural permanence is thoughtto be functional, enabling the muscles to di-rect the eyes in a precise and repeatablemanner. In strabismus, and especially withincomitant strabismus, however, these laws,instead of being helpful, are harmful. What isrequired in the case of a muscle weakness is anew innervation pattern; the old patternleads to symptoms. What is needed is amodification of neural patterns in the appro-priate direction, i.e., a functional violation ofHering's law and Listing's law. The questionis whether such fixed patterns of coordinationcan be reorganized.

314 Balliet and NakayamaInvest. Ophthalmol. Visual Sci.
April 1978
Our results suggest that the limits of volun-tary oculomotor control are as yet unknown.Since effortless voluntary torsion can bemaintained over a significant range (6° to 8°minimum), it is possible that other eyemovements can be trained as well. Forexample, voluntary cyclovergence might beapplied to patients with superior obliquepalsies, with the limits of such therapy de-termined by the specific feedback trainingprocedure, patient motivation, and time.
We thank Drs. Lawrence Stark and Brian Brown forhelpful comments. We especially thank subjectsMatthew Noble and Candus Hedberg.
R E F E R E N C E S
1. Allen, M. J.: Dependence of cyclophoria on con-vergence, elevation and the system of axes, Am. J.Optom. & Arch. Am. Acad. Optom. 31:297, 1954.
2. Bahill, T. A., Clark, M. R., and Stark, L.: Dynamicovershoot in saccadic eye movements is caused byneurological control signal reversals, Exp. Neurol.48:107, 1975.
3. Bahill, T. A., Clark, M. R., and Stark, L.: The mainsequence, a tool for studying eye movements, Math.Biosci. 24:191, 1975.
4. Bahill, T. A., and Stark, L.: Overlapping saccadesand glissades are produced by fatigue in the saccadiceye movement system, Exp. Neurol. 48:95, 1975.
5. Balliet, R., and Nakayama, K.: An illusion of selfrotation during voluntary torsion, Science (submit-ted, 1978).
6. Boeder, P.: The cooperative action of extraocularmuscles, Br. J. Ophthalmol. 46:397, 1962.
7. Brecher, G. A.: Die optokinetische Auslosung vonAugenrollung und rotatoischem Nystagmus, Pflue-gers Arch. Ges. Physiol. 234:13, 1934.
8. Crone, R. A.: Optically induced eye torsion. II. Op-tostatic and optokinetic cycloversion, Albrecht vonGraefes Arch. Klin. Exp. Ophthalmol. 196:1, 1975.
9. Crone, R. A., and Everhard-Halm, Y.: Optically in-duced eye torsion. I. Fusional cyclovergence. Al-brecht von Graefes Arch. Klin. Exp. Ophthalmol.195:231, 1975.
10. Ditchburn, R. W.: Eye Movements and Visual Per-ception, Oxford, England, 1973, Clarendon Press.
11. Helmholtz, H.: Treatise on Physiological Optics,translated from Third German edition, Southall, J.P. C , editor, New York, 1910, Dover Publications.
12. Heywood, S., and Churcher, J.: Eye movementsand the afterimage. I. Tracking the afterimage, Vi-sion Res. 11:1163, 1971.
13. Kertesz, A. E., and Jones, R. W.: The effect of angu-lar velocity of stimulus on human torsional eyemovements, Vision Res. 9:995, 1969.
14. Miller, E. F.: Counterrolling of the human eyesproduced by head tilt with respect to gravity, U.S.Naval School of Aviation Medicine Research ReportNo. 75, 1962.
15. Nakayama, K.: Coordination of extraocular muscles.In Lennerstrand, G., and Bach-y-Rita, P., editors:Basic Mechanisms of Ocular Motility and TheirClinical Implications, Oxford, England, 1975, Per-gamon Press, pp. 193-207.
16. Nakayama, K., and Balliet, R.: Listings Law, eyeposition sense, and the perception of the vertical,Vision Res. 17:453, 1977.
17. Noji, R.: Uber optisch Erzwungene paralleleRollungen der Augen, Arch. Ophthalmol. 122:562,1929.
18. Petrov, A. P., and Zenkin, G. M.: Torsional eyemovements and constancy of the visual field, VisionRes. 13:2465, 1973.
19. Robinson, D. A.: A quantitative analysis of extraocu-lar muscle coordination and squint, INVEST. OPH-THALMOL. 14:801, 1975.
20. Ron, S., Robinson, D. A., and Skavenski, A. A.:Saccades and the quick phase of nystagmus, VisionRes. 12:2015, 1972.
21. Schwartz, G. E., and Beatty, J.: Biofeedback—Theory and Research, New York, 1977, AcademicPress, Inc.
22. Steinbach, M. J.: Pursuing the perceptual ratherthan the retinal stimulus, Vision Res. 16:1371, 1976.
23. Weber, R. B., and Daroff, R. B.: Correctivemovements following refixation saccades: type andcontrol system analysis, Vision Res. 12:467, 1972.
24. Westheimer, G.: Saccadic eye movements. In Zik-mund, V., Editor: The Oculomotor System andBrain Function, Bratislava, Czechoslovakia, 1973.Publishing House of the Slovak Academy of Science.
25. Zuber, B. L., and Stark, L.: Micro saccades and thevelocity amplitude relationship for saccadic eyemovement, Science 150:1459, 1965.