
Apoplastic and Symplastic Pathways of Atrazine and Glyphosate
8
Plant Physiol. (1986) 82, 1000-1007 0032-0889/86/82/1000/08/$0 1.00/0 Apoplastic and Symplastic Pathways of Atrazine and Glyphosate Transport in Shoots of Seedling Sunflower' Received for publication October 28, 1985 and in revised form July 30, 1986 JOHN J. JACHETTA*2, ARNOLD P. APPLEBY, AND LARRY BOERSMA Department of Crop Science (J.J.J., A.P.A.) and Department of Soil Science (L.B.) Oregon State University, Corvallis, Oregon 97331 ABSTRACI I'4ClAtrazine (2-chloro4-lethylaminol-6-Iisopropylamino-s-triazine) and [4Clglyphosate (N-Iphosphonomethyllglycine) were xylem fed to sunflower shoots at 100 micromolar for 1 hour in the light, then placed in the dark at 100% relative humidity for 1, 4, 7, or 10 hours. The distribution of atrazine and glyphosate between shoot parts, in the leaves, and between the aoplast and symplast of the leaf was determined. The apoplastic concentrations and distribution patterns of atrazine and gly- phosate in the leaves were evaluated using a pressure dehydration tech- nique, our results were compared to the previously reported distribution patterns of the naturally occurring apoplastic leaf solutes, and the apo- plastic dye PTIS (trisodium 3-hydroxy-5,8,10-pyrenetrisulfonate). The pattern of atrazine and glyphosate distribution in the shoot, and between the leaf apoplast and symplast, was found to reflect the potential of these herbicides to enter the shoot symplast. The results of this study are discussed with respect to current theories of xenobiotic transport in plants, and have been found to be consistent with the intermediate permeability hypothesis for xenobiotic transport. The translocation pattern of an herbicide within a plant de- pends on the site of application of the compound and also on its ability to penetrate the plasmalemma. Two major patterns of herbicide transport have been extensively described (2), namely the apoplastic pattern and the symplastic pattern. Movement in the apoplast occurs mainly via the xylem and cell walls while movement in the symplast occurs via the interconnected proto- plasts of the parenchyma and phloem (3). Herbicides which move in the symplastic transport pattern, when applied to the leaves, stems, or roots of plants, must initially move into and through the apoplast. These materials are then in position to penetrate the plasmalemma-bound symplast where they are available for mobilization and transport, via the phloem, to the sites of active metabolism. Herbicides that move only in the apoplastic transport pattern, when applied to the leaves, are not exported from the leaves. When these materials are applied to stems or roots, they apparently move into the apoplast and are carried in the transpiration stream to the leaves. There is virtually no symplastic movement of these compounds out of the leaves (15). Chemicals which move in both patterns are termed ambi- mobile (9). The question of why some chemicals are transported via the 'Contribution of the Oregon Agricultural Experiment Station, Oregon State University, Corvallis, Tech. Paper No. 7701. 2 Currently, Sr. Plant Physiologist, Union Carbide Agr. Prod. Co., P. 0. Box 12014. Research Triangle Park, NC 27709. symplast and others are not is of considerable importance, and the resolution of this question may provide the information necessary to predict the systemic properties of pesticidal and growth regulating organic molecules. At the tissue level, clarifi- cation would appear to depend upon an understanding of the penetration of the compound from the water-filled apoplast into the membrane-bound symplast. Currently, two hypotheses explain the observed transport pat- terns in plants. Both assume that the potential for a compound to penetrate the symplast, and for the symplast to retain that compound, can be correlated with the overall pattern of transport it achieves in the plant as a whole. The first hypothesis is the historical view. Earlier references indicate that compounds which display the apoplastic transport pattern do not move readily into the protoplasts of plant cells, and thus are essentially confined to the apoplast, moving with the transpiration stream (see review in Ashton and Crafts [1]). Herbicides which move in the sym- plastic transport pattern were thought to be preferentially ab- sorbed into the symplast, and subject to long-distance transport in the phloem (1). More recent experimental evidence has dem- onstrated that several compounds which display the apoplastic transport pattern easily penetrate the plasmalemma of plant cells (6, 20, 22, 24, 25, 30, 31). Peterson and Edgington (22) suggested that chemicals which penetrate into the symplast but display an apoplastic transport pattern be called pseudoapoplastic. They suggest that because transport in the transpiration stream is much more rapid than transport in the symplast, and because the plasmalemma is readily permeable to pseudoapoplastic compounds, the symplast cannot retain these compounds, and consequently they are leached into the apoplast and carried away with the transpiration stream. This theory has resulted in the development of the 'intermediate permeability' hypothesis (21, 22, 36), a comple- mentary theory for symplastic and ambimobile compounds which penetrate the plasmalemma by passive diffusion and do not act like weak acids, such as 2,4-D. According to the intermediate permeability mechanism, a substance that can penetrate the symplasm of the phloem will be retained for long-distance transport to the carbohydrate sinks if the permeability of the compound is low enough to allow the material to be retained within the sieve tube long enough to be transported to the sink regions. A herbicide would display am- bimobile translocation if, as symplastic transport occurs, the compound continuously leaks out of the phloem into the apo- plast to be redistributed in the xylem. The degee ofambimobility would depend on the rate of leakage from the symplast, with symplastic transport occurring when there is a high membrane resistance to leakage, and apoplastic transport occurring with a low membrane resistance to leakage. These two theories of transport predict very different intercel- lular concentations of herbicides that follow the apoplastic pat- 1000 Downloaded from https://academic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 November 2021
Transcript of Apoplastic and Symplastic Pathways of Atrazine and Glyphosate

Plant Physiol. (1986) 82, 1000-10070032-0889/86/82/1000/08/$0 1.00/0
Apoplastic and Symplastic Pathways of Atrazine and GlyphosateTransport in Shoots of Seedling Sunflower'
Received for publication October 28, 1985 and in revised form July 30, 1986
JOHN J. JACHETTA*2, ARNOLD P. APPLEBY, AND LARRY BOERSMADepartment ofCrop Science (J.J.J., A.P.A.) and Department ofSoil Science (L.B.) Oregon StateUniversity, Corvallis, Oregon 97331
ABSTRACI
I'4ClAtrazine (2-chloro4-lethylaminol-6-Iisopropylamino-s-triazine)and [4Clglyphosate (N-Iphosphonomethyllglycine) were xylem fed tosunflower shoots at 100 micromolar for 1 hour in the light, then placedin the dark at 100% relative humidity for 1, 4, 7, or 10 hours. Thedistribution of atrazine and glyphosate between shoot parts, in the leaves,and between the aoplast and symplast of the leaf was determined. Theapoplastic concentrations and distribution patterns of atrazine and gly-phosate in the leaves were evaluated using a pressure dehydration tech-nique, our results were compared to the previously reported distributionpatterns of the naturally occurring apoplastic leaf solutes, and the apo-plastic dye PTIS (trisodium 3-hydroxy-5,8,10-pyrenetrisulfonate). Thepattern of atrazine and glyphosate distribution in the shoot, and betweenthe leaf apoplast and symplast, was found to reflect the potential oftheseherbicides to enter the shoot symplast. The results of this study arediscussed with respect to current theories of xenobiotic transport inplants, and have been found to be consistent with the intermediatepermeability hypothesis for xenobiotic transport.
The translocation pattern of an herbicide within a plant de-pends on the site of application ofthe compound and also on itsability to penetrate the plasmalemma. Two major patterns ofherbicide transport have been extensively described (2), namelythe apoplastic pattern and the symplastic pattern. Movement inthe apoplast occurs mainly via the xylem and cell walls whilemovement in the symplast occurs via the interconnected proto-plasts of the parenchyma and phloem (3). Herbicides whichmove in the symplastic transport pattern, when applied to theleaves, stems, or roots of plants, must initially move into andthrough the apoplast. These materials are then in position topenetrate the plasmalemma-bound symplast where they areavailable for mobilization and transport, via the phloem, to thesites of active metabolism. Herbicides that move only in theapoplastic transport pattern, when applied to the leaves, are notexported from the leaves. When these materials are applied tostems or roots, they apparently move into the apoplast and arecarried in the transpiration stream to the leaves. There is virtuallyno symplastic movement of these compounds out of the leaves(15). Chemicals which move in both patterns are termed ambi-mobile (9).The question of why some chemicals are transported via the
'Contribution of the Oregon Agricultural Experiment Station, OregonState University, Corvallis, Tech. Paper No. 7701.
2 Currently, Sr. Plant Physiologist, Union Carbide Agr. Prod. Co., P.0. Box 12014. Research Triangle Park, NC 27709.
symplast and others are not is of considerable importance, andthe resolution of this question may provide the informationnecessary to predict the systemic properties of pesticidal andgrowth regulating organic molecules. At the tissue level, clarifi-cation would appear to depend upon an understanding of thepenetration of the compound from the water-filled apoplast intothe membrane-bound symplast.
Currently, two hypotheses explain the observed transport pat-terns in plants. Both assume that the potential for a compoundto penetrate the symplast, and for the symplast to retain thatcompound, can be correlated with the overall pattern oftransportit achieves in the plant as a whole. The first hypothesis is thehistorical view. Earlier references indicate that compounds whichdisplay the apoplastic transport pattern do not move readily intothe protoplasts of plant cells, and thus are essentially confined tothe apoplast, moving with the transpiration stream (see reviewin Ashton and Crafts [1]). Herbicides which move in the sym-plastic transport pattern were thought to be preferentially ab-sorbed into the symplast, and subject to long-distance transportin the phloem (1). More recent experimental evidence has dem-onstrated that several compounds which display the apoplastictransport pattern easily penetrate the plasmalemma ofplant cells(6, 20, 22, 24, 25, 30, 31).
Peterson and Edgington (22) suggested that chemicals whichpenetrate into the symplast but display an apoplastic transportpattern be called pseudoapoplastic. They suggest that becausetransport in the transpiration stream is much more rapid thantransport in the symplast, and because the plasmalemma isreadily permeable to pseudoapoplastic compounds, the symplastcannot retain these compounds, and consequently they areleached into the apoplast and carried away with the transpirationstream. This theory has resulted in the development of the'intermediate permeability' hypothesis (21, 22, 36), a comple-mentary theory for symplastic and ambimobile compoundswhich penetrate the plasmalemma by passive diffusion and donot act like weak acids, such as 2,4-D.According to the intermediate permeability mechanism, a
substance that can penetrate the symplasm of the phloem willbe retained for long-distance transport to the carbohydrate sinksif the permeability of the compound is low enough to allow thematerial to be retained within the sieve tube long enough to betransported to the sink regions. A herbicide would display am-bimobile translocation if, as symplastic transport occurs, thecompound continuously leaks out of the phloem into the apo-plast to be redistributed in the xylem. The degee ofambimobilitywould depend on the rate of leakage from the symplast, withsymplastic transport occurring when there is a high membraneresistance to leakage, and apoplastic transport occurring with alow membrane resistance to leakage.These two theories of transport predict very different intercel-
lular concentations of herbicides that follow the apoplastic pat-1000
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021

APOPLASTIC AND SYMPLASTIC HERBICIDE TRANSPORT
tern of translocation as compared to those that translocate in thesymplastic pattern. The historical view predicts higher concen-trations ofapoplastically transported compounds in the cell wallsand xylem of source leaves, with a lower concentration of thesematerials within the leaf protoplasts. Conversely, there should bea lower concentration of symplastically transported compoundsin the apoplast than in the symplast. The intermediate permea-bility hypothesis predicts the opposite situation, with ambimobileand symplastically transported compounds initially existing inhigher concentrations in the apoplast of source leaves thancompounds which follow the apoplastic transport pattern.The objective of this study was to determine the distribution
of two herbicides with quite different translocation patterns,atrazine and glyphosate, in the apoplast and symplast of intactsunflower leaves. Results of this study were used to distinguishbetween the historical and intermediate permeability transporttheories for herbicide translocation. Atrazine and glyphosatewere chosen for study because both compounds have been shownto move into cells by passive diffusion (10, 13, 22, 24). However,atrazine follows the apoplastic transport patterns (1), while gly-phosate follows the symplastic transport pattern, with a tendencyto be slightly ambimobile (6, 13). The distribution of thesecompounds was compared to the distribution previously reportedfor the naturally occurring leaf solutes, and the fluorescent dyetrisodium 3-hydroxy-5,8,10-pyrenetrisulfonate (18). PTS3 isknown to be confined to the apoplast of the plant, is nontoxic atthe concentrations employed in this study (23), highly watersoluble, and not adsorbed onto cell walls (8, 35).
MATERIALS AND METHODSTranspiration Inhibition by Atrazine and Glyphosate. Sun-
flower plants were grown as previously reported (18). Beforeplants were sampled, they were kept in a darkened, humidifiedgrowth chamber for 12 h at 21°C. At the end of the dark period,plants were removed from the growth chamber and all but thetwo opposing fully expanded first true leaves were removed. Theshoots were then excised under water at the soil surface. Shootswere transferred to flasks containing 100 gM Ca(NO3)2 and placedin the dark growth chamber for 90 min to allow recovery of theshoot water potential to approximately -0.15 MPa. Shoots andflasks were then weighed and placed in a lighted growth chamber(300 umol m-2 s-' at leaf blade level) for 90 min, which wasfound to be sufficient to allow steady state transpiration. Shootswere then transferred in the lighted growth chamber to flaskscontaining 100 Mm Ca(NO3)2 plus 100 Mm atrazine or glyphosate,or herbicide-free 100 Mm Ca(NO3)2 solution for 90 min, afterwhich all shoots were transferred to herbicide-free 100 gMCa(NO3)2 for the remainder of the experiment. The total weightof the shoot and flask was recorded every 1.0 or 1.5 h over a 12h period. Leaf area was measured at the end of the experiment,using a Li-Cor area meter (model Li-3000, Lambda InstrumentCorp., Lincoln, NE). Transpiration was measured by dividingthe change in weight of the flask and shoot by the total leaf areaof the shoot. The experiment was conducted as a completelyrandomized design with three replications per treatment.
Distribution of PTS, Atrazine, and Glyphosate. Sunflowershoots were prepared as in the transpiration inhibition study.After a 90-min period in a dark growth chamber to allow recoveryofthe shoot water potential to approximately 0.15 MPa, followedby 90-min in a lighted growth chamber, excised shoots weretransferred to vials containing 5 ml of 100 gM Ca(N03)2 pluseither 100 Mm [U-ring-'4C]atrazine, or 100 Mm [methyl-'4C]gly-phosate. Shoots and vials were weighed, and plants were main-tained in the lighted growth chamber to allow uptake of thetreatment solution.
Following a 90-min exposure, shoots and vials were reweighedto determine the amount of treatment solution uptake, sealed inprehumidified plastic bags, and placed in a dark growth chamberto retard transpiration and further uptake of the treatment solu-tion. After 1, 4, 7, or 10 h, shoots and vials were removed fromthe plastic bags, one leaf was excised, and xylem sap was ex-pressed in a pressure chamber as previously described (18). Theremaining leafand stem were immediately mounted on a blotterpaper and frozen between sheets of dry ice for subsequent lyoph-ilization, autoradiography, and quantification of radioactivity.Xylem sap from the excised leaf was expressed for approxi-
mately 5 min at each balance pressure; each successive balancepressure was 0.02 to 0.04 MPa greater than the previous balancepressure. Xylem sap was collected on a filter paper disk under ahumidified chamber as described previously (18), after which thefilter paper disk was sealed in a scintillation vial. To determinethe weight of expressed xylem sap, the filter paper disk and vialwere weighed together before and immediately after collectionof the sap. After sap was expressed in pressure increments to atotal of 0.5 MPa, the pressure in the chamber was released, thepetiole was quickly excised, and weights of the petiole and leafblade were recorded. Shoots also were treated with PTS (200mg/l in 100 Mm Ca[NO3]2) in the same manner, and photographsof the leaves under UV light were made for comparison withautoradiographs. Results from a compartmental analysis of ex-pressed sap after 1, 4, 7, or 10-h dark periods were pooled tocalculate means and standard errors unless significantly differentat the 5% level.
Shoots were dried and analyzed for radioactivity in parts: stembelow the leaves, growing point above the leaves, petioles, leaves,and a strip of tissue 2 mm wide adjacent to the main veins ofthe leaf. Radioactivity in the leafand tissue strips were combinedfor analysis of the distribution of label among plant parts. Eachplant part was weighed, ground in a Wiley Mill fitted with a 20-mesh screen, and combusted in a Packard 305 sample oxidizerwith subsequent counting of the liberated 14C02 by scintillationspectrometry. Samples combusted comprise greater than 30% byweight of the leaves and greater than 50% of the other plantparts; all of the excised leaf tissue strips were combusted.
Herbicides in the expressed xylem sap were extracted fromfilter paper disks in the scintillation vial with 1 ml of methanolor water (for atrazine and glyphosate, respectively) for 24 h. Theradioactivity for each sample was determined by liquid scintil-lation spectrometry and all necessary corrections for volume,quenching, and background were made. The concentration ofchemical in the apoplast was determined graphically from theexpressed sap efflux curves between 0.3 and 0.4 MPa appliedpressure as described previously (18). The concentration of her-bicide in the symplast was calculated from the excised tissue stripadjacent to the main veins of the opposite leaf which had nothad xylem sap expressed, using the following equation:
moles of mateial in strip tissue[Sym] - - moles in strip apoplast
total water in leaf strip - water in strip apoplastThe total weight of water in the leaf strip sample was calculatedas:
sample dry weight x
weight of water in sample = (leaf fresh weight- Wd)Wd
where Wd is the dry weight of the leaf after oven drying at 60°Cfor 24 h. The volume of water in the apoplast was estimated, ona percent basis, from pressure-volume analysis of similar leavesfrom sunflowers grown under the same conditions at the sametime as described previously (18); this value was used to calculate
1001
'Abbreviation: PTS, trisodium 3-hydroxy-5,8, I 0-pyrenetrisulfonate.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021
Plant Physiol. Vol. 82, 1986
the moles of herbicide, and volume of water, in the leaf stripapoplast. A symplast-apoplast partitioning quotient for atrazineand glyphosate was calculated by dividing the concentration ofmaterial in the symplast by the concentration in the apoplast.Experiments consisted of one shoot per treatment and wererepeated four times for atrazine and three times for glyphosate.
Identity of Transported Molecules. TLC was used to verifythat the ['4C]atrazine remained intact atrazine. Leaves of shootsthat were allowed to take up ['4C]atrazine for 90 min, sealed inprehumidified plastic bags, and incubated in a dark growthchamber for 10 h were used. Leaves were extracted three timesby homogenization in 10 volumes (about 20 ml) of 80% (v/v)methanol:water (10:1 [v/w] ratio of 80% methanol to freshweight leaves). The homogenate was filtered through WhatmanNo. 1 filter paper, and the three methanol extracts were com-bined. Methanol was removed, and the aqueous extract wasreduced in volume to 5 ml in vacuo by rotary evaporation at40C. The extract was frozen and stored at -70°C until used.
Aliquots (1 ml) of the extract were lyophilized, and the residuewas resuspended in 150 Ml of water for spotting onto 20 by 20cm silica gel thin-layer plates. Two different solvent systems wereused: (a) benzene:acetic acid:water (60:40:3, v/v/v), followed byair-drying for 30 min at room temperature, and (b) n-bu-tanol:acetic acid:water (120:30:50, v/v/v). Zones containing ra-dioactivity were located by autoradiography, scraped into scin-tillation vials, and the radioactivity associated with each zonewas determined. This procedure follows that of Frear and Swan-son (1 1), and resulting Rf values of ['4C]atrazine and radioactivemetabolites were comparable to their values. Thin-layer chro-matograms with only [14C]atrazine also were prepared for com-parison. This analysis was performed for three sets of two leaveseach.A glyphosate metabolism study was not conducted as numer-
ous studies have shown that glyphosate metabolism does notoccur during short-term experiments (12, 19, 26, 27, 41, 42).
Partition Coefficients. Partition coefficients were determinedfor atrazine and glyphosate and compared with the previouslydescribed symplast-apoplast partitioning quotients. Labeled atra-zine or glyphosate was added to 22-ml scintillation vials contain-ing 5 ml of 2-octanol and buffer. Buffer (Hydrion, Micro Essen-tial Laboratory, Brooklyn, NY) solutions ranged from pH 3 topH 8 and pH 10 to pH 12 in pH steps of 1.0. The vials wereplaced in a reciprocating shaker for 8 h and the aqueous andorganic phases were sampled 24 h later. After correcting forcounting efficiency, the partition coefficient was calculated bydividing the number of dpm's in the organic phase by those inthe aqueous phase. The experiment was conducted as a com-pletely randomized design with five replications per treatment.
RESULTS AND DISCUSSION
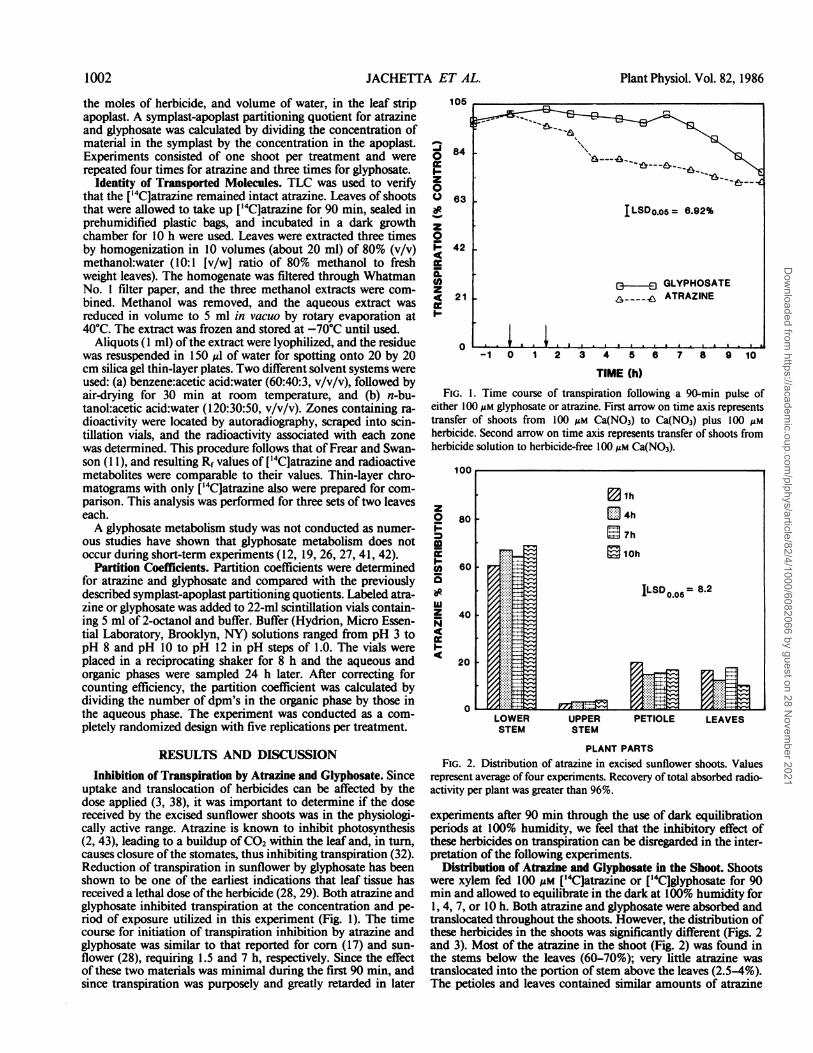
Inhibition of Transpiration by Atrazine and Glyphosate. Sinceuptake and translocation of herbicides can be affected by thedose applied (3, 38), it was important to determine if the dosereceived by the excised sunflower shoots was in the physiologi-cally active range. Atrazine is known to inhibit photosynthesis(2, 43), leading to a buildup ofCO2 within the leaf and, in turn,causes closure of the stomates, thus inhibiting transpiration (32).Reduction of transpiration in sunflower by glyphosate has beenshown to be one of the earliest indications that leaf tissue hasreceived a lethal dose ofthe herbicide (28, 29). Both atrazine andglyphosate inhibited transpiration at the concentration and pe-riod of exposure utilized in this experiment (Fig. 1). The timecourse for initiation of transpiration inhibition by atrazine andglyphosate was similar to that reported for corn (17) and sun-flower (28), requiring 1.5 and 7 h, respectively. Since the effectof these two materials was minimal during the first 90 min, andsince transpiration was purposely and greatly retarded in later
105
0-
.i0c
zl%.w
z
4c
IC
84
63
42
21
0-1 0 1 2 3 4 5 6 7 8 9 10
TIME (h)
FIG. 1. Time course of transpiration following a 90-min pulse ofeither 100 AM glyphosate or atrazine. First arrow on time axis representstransfer of shoots from 100 AM Ca(NO3) to Ca(NO3) plus 100 AMherbicide. Second arrow on time axis represents transfer of shoots fromherbicide solution to herbicide-free 100 Mm Ca(NO3).
1lU I
z0
P
0
w
z
4c
I--
4c
80 F
60
40
20
0
LOWERSTEM
UPPERSTEM
PETIOLE LEAVES
PLANT PARTSFIG. 2. Distribution of atrazine in excised sunflower shoots. Values
represent average of four experiments. Recovery of total absorbed radio-activity per plant was greater than 96%.
experiments after 90 min through the use of dark equilibrationperiods at 100% humidity, we feel that the inhibitory effect ofthese herbicides on transpiration can be disregarded in the inter-pretation of the following experiments.
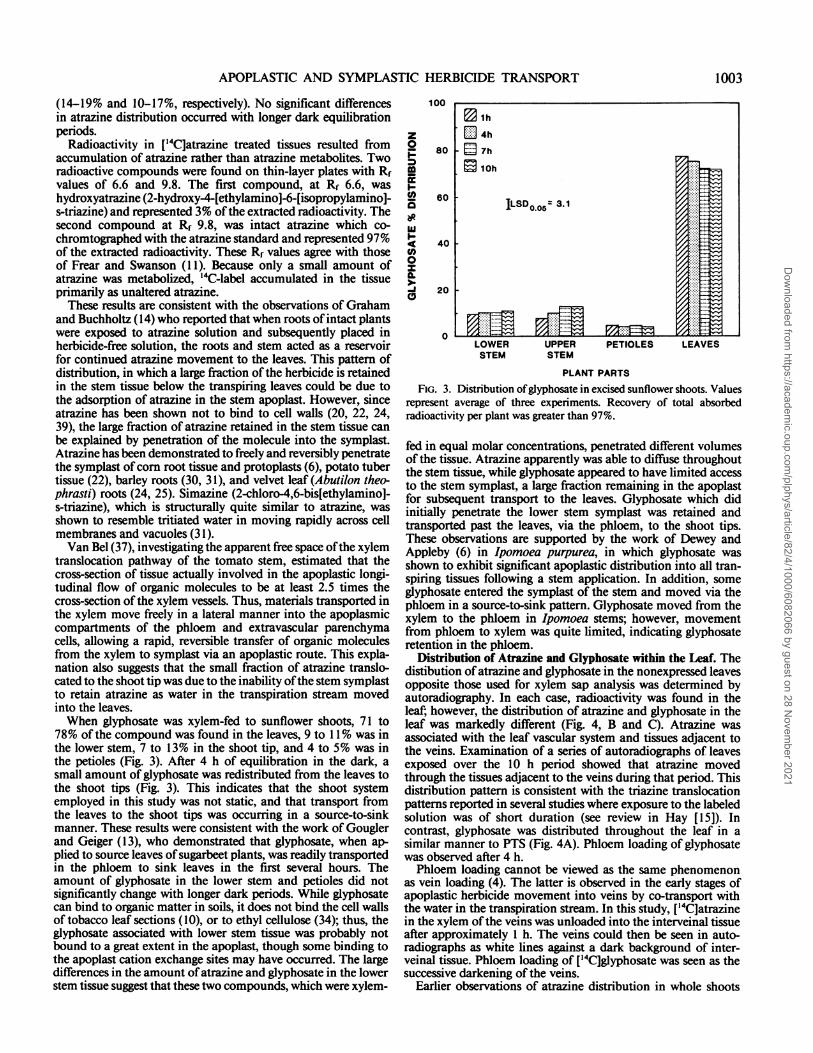
Distribution of Atrazine and Glyphosate in the Shoot. Shootswere xylem fed 100 Mm [4C]atrazine or ['4C]glyphosate for 90min and allowed to equilibrate in the dark at 100% humidity for1, 4, 7, or 10 h. Both atrazine and glyphosate were absorbed andtranslocated throughout the shoots. However, the distribution ofthese herbicides in the shoots was significantly different (Figs. 2and 3). Most of the atrazine in the shoot (Fig. 2) was found inthe stems below the leaves (60-70%); very little atrazine wastranslocated into the portion of stem above the leaves (2.5-4%).The petioles and leaves contained similar amounts of atrazine
S -t---b---6&- L
ILSDo.05 = 6.92%
C B GLYPHOSATEg__ zATRAZINE
1h
0 4h
E 7h
ILSD 8.2
--.. . . .. . I I. . . .
1002 JACHETTA ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021
APOPLASTIC AND SYMPLASTIC HERBICIDE TRANSPORT
(14-19% and 10-17%, respectively). No significant differencesin atrazine distribution occurred with longer dark equilibrationperiods.
Radioactivity in ['4C]atrazine treated tissues resulted fromaccumulation of atrazine rather than atrazine metabolites. Tworadioactive compounds were found on thin-layer plates with Rfvalues of 6.6 and 9.8. The first compound, at Rf 6.6, washydroxyatrazine (2-hydroxy-4-[ethylamino]-6-[isopropylamino]-s-triazine) and represented 3% ofthe extracted radioactivity. Thesecond compound at Rf 9.8, was intact atrazine which co-chromtographed with the atrazine standard and represented 97%of the extracted radioactivity. These Rf values agree with thoseof Frear and Swanson (11). Because only a small amount ofatrazine was metabolized, 14(C-label accumulated in the tissueprimarily as unaltered atrazine.These results are consistent with the observations of Graham
and Buchholtz (14) who reported that when roots ofintact plantswere exposed to atrazine solution and subsequently placed inherbicide-free solution, the roots and stem acted as a reservoirfor continued atrazine movement to the leaves. This pattern ofdistribution, in which a large fraction of the herbicide is retainedin the stem tissue below the transpiring leaves could be due tothe adsorption of atrazine in the stem apoplast. However, sinceatrazine has been shown not to bind to cell walls (20, 22, 24,39), the large fraction of atrazine retained in the stem tissue canbe explained by penetration of the molecule into the symplast.Atrazine has been demonstrated to freely and reversibly penetratethe symplast of corn root tissue and protoplasts (6), potato tubertissue (22), barley roots (30, 31), and velvet leaf (Abutilon theo-phrasti) roots (24, 25). Simazine (2-chloro-4,6-bis[ethylamino]-s-triazine), which is structurally quite similar to atrazine, wasshown to resemble tritiated water in moving rapidly across cellmembranes and vacuoles (31).Van Bel (37), investigating the apparent free space ofthe xylem
translocation pathway of the tomato stem, estimated that thecross-section of tissue actually involved in the apoplastic longi-tudinal flow of organic molecules to be at least 2.5 times thecross-section of the xylem vessels. Thus, materials transported inthe xylem move freely in a lateral manner into the apoplasmiccompartments of the phloem and extravascular parenchymacells, allowing a rapid, reversible transfer of organic moleculesfrom the xylem to symplast via an apoplastic route. This expla-nation also suggests that the small fraction of atrazine translo-cated to the shoot tip was due to the inability ofthe stem symplastto retain atrazine as water in the transpiration stream movedinto the leaves.When glyphosate was xylem-fed to sunflower shoots, 71 to
78% of the compound was found in the leaves, 9 to 11% was inthe lower stem, 7 to 13% in the shoot tip, and 4 to 5% was inthe petioles (Fig. 3). After 4 h of equilibration in the dark, asmall amount of glyphosate was redistributed from the leaves tothe shoot tips (Fig. 3). This indicates that the shoot systememployed in this study was not static, and that transport fromthe leaves to the shoot tips was occurring in a source-to-sinkmanner. These results were consistent with the work of Gouglerand Geiger (13), who demonstrated that glyphosate, when ap-plied to source leaves of sugarbeet plants, was readily transportedin the phloem to sink leaves in the first several hours. Theamount of glyphosate in the lower stem and petioles did notsignificantly change with longer dark periods. While glyphosatecan bind to organic matter in soils, it does not bind the cell wallsof tobacco leaf sections (10), or to ethyl cellulose (34); thus, theglyphosate associated with lower stem tissue was probably notbound to a great extent in the apoplast, though some binding tothe apoplast cation exchange sites may have occurred. The largedifferences in the amount of atrazine and glyphosate in the lowerstem tissue suggest that these two compounds, which were xylem-
100
z0P
0
m
icn
co1-4c
80
60
40
20
0 L I
LOWER UPPER PETIOLES LEAVESSTEM STEM
PLANT PARTS
FIG. 3. Distribution ofglyphosate in excised sunflower shoots. Valuesrepresent average of three experiments. Recovery of total absorbedradioactivity per plant was greater than 97%.
fed in equal molar concentrations, penetrated different volumesof the tissue. Atrazine apparently was able to diffuse throughoutthe stem tissue, while glyphosate appeared to have limited accessto the stem symplast, a large fraction remaining in the apoplastfor subsequent transport to the leaves. Glyphosate which didinitially penetrate the lower stem symplast was retained andtransported past the leaves, via the phloem, to the shoot tips.These observations are supported by the work of Dewey andAppleby (6) in Ipomoea purpurea, in which glyphosate wasshown to exhibit significant apoplastic distribution into all tran-spiring tissues following a stem application. In addition, someglyphosate entered the symplast of the stem and moved via thephloem in a source-to-sink pattern. Glyphosate moved from thexylem to the phloem in Ipomoea stems; however, movementfrom phloem to xylem was quite limited, indicating glyphosateretention in the phloem.
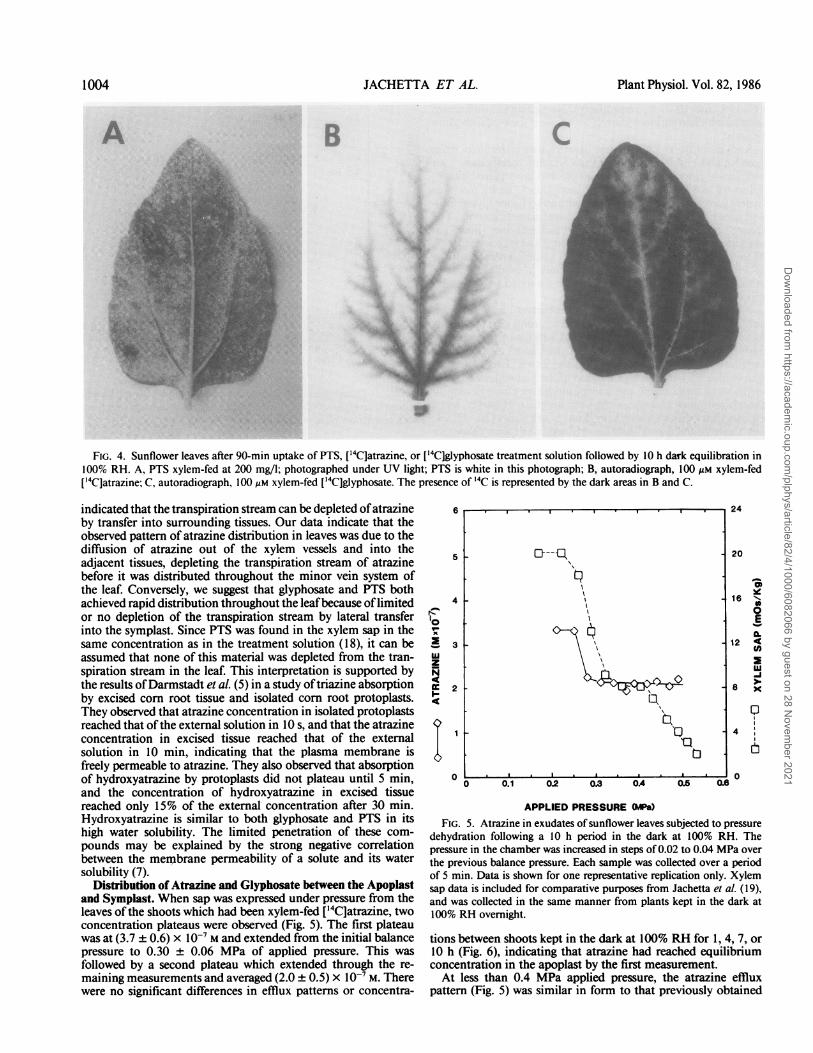
Distribution of Atrazine and Glyphosate within the Leaf. Thedistibution ofatrazine and glyphosate in the nonexpressed leavesopposite those used for xylem sap analysis was determined byautoradiography. In each case, radioactivity was found in theleaf; however, the distribution of atrazine and glyphosate in theleaf was markedly different (Fig. 4, B and C). Atrazine wasassociated with the leaf vascular system and tissues adjacent tothe veins. Examination of a series of autoradiographs of leavesexposed over the 10 h period showed that atrazine movedthrough the tissues adjacent to the veins during that period. Thisdistribution pattern is consistent with the triazine translocationpatterns reported in several studies where exposure to the labeledsolution was of short duration (see review in Hay (15]). Incontrast, glyphosate was distributed throughout the leaf in asimilar manner to PTS (Fig. 4A). Phloem loading of glyphosatewas observed after 4 h.Phloem loading cannot be viewed as the same phenomenon
as vein loading (4). The latter is observed in the early stages ofapoplastic herbicide movement into veins by co-transport withthe water in the transpiration stream. In this study, [14C]atrazinein the xylem ofthe veins was unloaded into the interveinal tissueafter approximately 1 h. The veins could then be seen in auto-radiographs as white lines against a dark background of inter-veinal tissue. Phloem loading of ['4C]glyphosate was seen as thesuccessive darkening of the veins.
Earlier observations of atrazine distribution in whole shoots
1003
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021
Plant Physiol. Vol. 82, 1986
AN B"A C~E
I.
FIG. 4. Sunflower leaves after 90-min uptake of PTS, ['4C]atrazine, or ['4C]glyphosate treatment solution followed by 10 h dark equilibration in100% RH. A, PTS xylem-fed at 200 mg/l; photographed under UV light; PTS is white in this photograph; B, autoradiograph, 100 Mm xylem-fed['4C]atrazine; C, autoradiograph, 100 Mm xylem-fed ['4C]glyphosate. The presence of '4C is represented by the dark areas in B and C.
indicated that the transpiration stream can be depleted ofatrazineby transfer into surrounding tissues. Our data indicate that theobserved pattern of atrazine distribution in leaves was due to thediffusion of atrazine out of the xylem vessels and into theadjacent tissues, depleting the transpiration stream of atrazinebefore it was distributed throughout the minor vein system ofthe leaf. Conversely, we suggest that glyphosate and PTS bothachieved rapid distribution throughout the leafbecause oflimitedor no depletion of the transpiration stream by lateral transferinto the symplast. Since PTS was found in the xylem sap in thesame concentration as in the treatment solution (18), it can beassumed that none of this material was depleted from the tran-spiration stream in the leaf. This interpretation is supported bythe results ofDarmstadt et al. (5) in a study oftriazine absorptionby excised corn root tissue and isolated corn root protoplasts.They observed that atrazine concentration in isolated protoplastsreached that ofthe external solution in lO s, and that the atrazineconcentration in excised tissue reached that of the externalsolution in 10 min, indicating that the plasma membrane isfreely permeable to atrazine. They also observed that absorptionof hydroxyatrazine by protoplasts did not plateau until 5 min,and the concentration of hydroxyatrazine in excised tissuereached only 15% of the external concentration after 30 min.Hydroxyatrazine is similar to both glyphosate and PTS in itshigh water solubility. The limited penetration of these com-pounds may be explained by the strong negative correlationbetween the membrane permeability of a solute and its watersolubility (7).
Distribution of Atrazine and Glyphosate between the Apoplastand Symplast. When sap was expressed under pressure from theleaves of the shoots which had been xylem-fed ['4C]atrazine, twoconcentration plateaus were observed (Fig. 5). The first plateauwas at (3.7 + 0.6) x IO-' M and extended from the initial balancepressure to 0.30 ± 0.06 MPa of applied pressure. This wasfollowed by a second plateau which extended through the re-maining measurements and averaged (2.0 ± 0.5) x 10-' M. Therewere no significant differences in efflux patterns or concentra-
6
5
I0x
N4
4
3
2
00 0.1 02 0.3 0.4 0.5 0.6
24
20
16 -00E
0.
w
-I
8 x
4
APPLIED PRESSURE OMPa)FIG. 5. Atrazine in exudates of sunflower leaves subjected to pressure
dehydration following a 10 h period in the dark at 100% RH. Thepressure in the chamber was increased in steps of 0.02 to 0.04 MPa overthe previous balance pressure. Each sample was collected over a periodof 5 min. Data is shown for one representative replication only. Xylemsap data is included for comparative purposes from Jachetta et al. ( 19),and was collected in the same manner from plants kept in the dark at100% RH overnight.
tions between shoots kept in the dark at 100% RH for 1, 4, 7, or10 h (Fig. 6), indicating that atrazine had reached equilibriumconcentration in the apoplast by the first measurement.At less than 0.4 MPa applied pressure, the atrazine effiux
pattern (Fig. 5) was similar in form to that previously obtained
w X w * |wII
I I* afs s
1004 JACHETTA ET AL.
I
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021
APOPLASTIC AND SYMPLASTIC HERBICIDE
0
I--I
0.
0
a.4c
6 7
65
54
70
3 X
I-
241-a.
0
0
0
LUI-400 2
a.
1I01
APOPLAST SYMPLAST
TISSUE SYSTEM
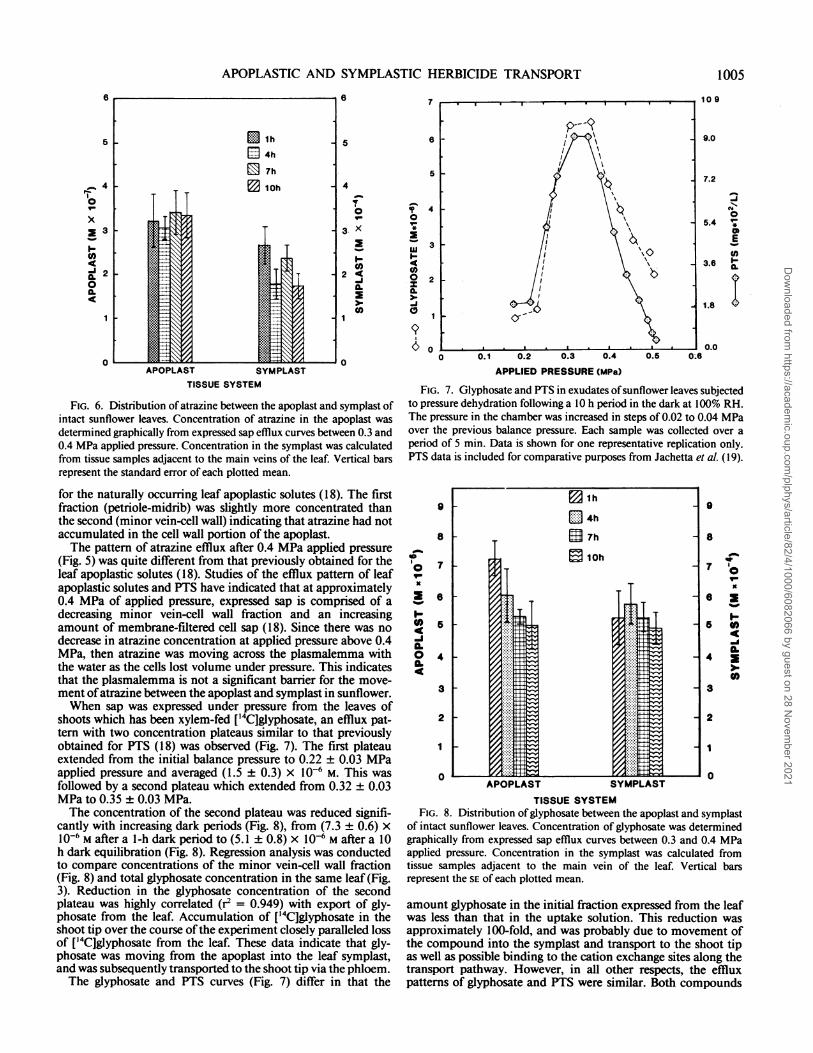
FIG. 6. Distribution of atrazine between the apoplast and symplast ofintact sunflower leaves. Concentration of atrazine in the apoplast wasdetermined graphically from expressed sap efflux curves between 0.3 and0.4 MPa applied pressure. Concentration in the symplast was calculatedfrom tissue samples adjacent to the main veins of the leaf. Vertical barsrepresent the standard error of each plotted mean.
for the naturally occurring leaf apoplastic solutes (18). The firstfraction (petriole-midrib) was slightly more concentrated thanthe second (minor vein-cell wall) indicating that atrazine had notaccumulated in the cell wall portion of the apoplast.The pattern of atrazine efflux after 0.4 MPa applied pressure
(Fig. 5) was quite different from that previously obtained for theleaf apoplastic solutes (18). Studies of the efflux pattern of leafapoplastic solutes and PTS have indicated that at approximately0.4 MPa of applied pressure, expressed sap is comprised of adecreasing minor vein-cell wall fraction and an increasingamount of membrane-filtered cell sap (18). Since there was nodecrease in atrazine concentration at applied pressure above 0.4MPa, then atrazine was moving across the plasmalemma withthe water as the cells lost volume under pressure. This indicatesthat the plasmalemma is not a significant barrier for the move-ment ofatrazine between the apoplast and symplast in sunflower.When sap was expressed under pressure from the leaves of
shoots which has been xylem-fed ['4C]glyphosate, an efflux pat-tern with two concentration plateaus similar to that previouslyobtained for PTS (18) was observed (Fig. 7). The first plateauextended from the initial balance pressure to 0.22 ± 0.03 MPaapplied pressure and averaged (1.5 ± 0.3) x 106 M. This wasfollowed by a second plateau which extended from 0.32 ± 0.03MPa to 0.35 ± 0.03 MPa.The concentration of the second plateau was reduced signifi-
cantly with increasing dark periods (Fig. 8), from (7.3 ± 0.6) x106 M after a 1-h dark period to (5.1 ± 0.8) x 106 M after a 10h dark equilibration (Fig. 8). Regression analysis was conductedto compare concentrations of the minor vein-cell wall fraction(Fig. 8) and total glyphosate concentration in the same leaf (Fig.3). Reduction in the glyphosate concentration of the secondplateau was highly correlated (r2 = 0.949) with export of gly-phosate from the leaf. Accumulation of ['4C]glyphosate in theshoot tip over the course ofthe experiment closely paralleled lossof [14C]glyphosate from the leaf. These data indicate that gly-phosate was moving from the apoplast into the leaf symplast,and was subsequently transported to the shoot tip via the phloem.The glyphosate and PTS curves (Fig. 7) differ in that the
0 0.1 0.2 0.3 0.4APPLIED PRESSURE (MPa)
0.5 0-6
10 9
9.0
7.2
-IN0
5.4 *;
3.6 I
1.8
0.0
FIG. 7. Glyphosate and PTS in exudates ofsunflower leaves subjectedto pressure dehydration following a 10 h period in the dark at 100% RH.The pressure in the chamber was increased in steps of 0.02 to 0.04 MPaover the previous balance pressure. Each sample was collected over aperiod of 5 min. Data is shown for one representative replication only.PTS data is included for comparative purposes from Jachetta et aL (19).
9
8
Q940
I-
0
1-I
0
4c
7
5
4
3
2
1
0
9
8
60
OZ%
7 X
V.
3
2
%w
IL
TISSUE SYSTEMFIG. 8. Distribution ofglyphosate between the apoplast and symplast
of intact sunflower leaves. Concentration of glyphosate was determinedgraphically from expressed sap efflux curves between 0.3 and 0.4 MPaapplied pressure. Concentration in the symplast was calculated fromtissue samples adjacent to the main vein of the leaf. Vertical barsrepresent the SE of each plotted mean.
amount glyphosate in the initial fraction expressed from the leafwas less than that in the uptake solution. This reduction wasapproximately 100-fold, and was probably due to movement ofthe compound into the symplast and transport to the shoot tipas well as possible binding to the cation exchange sites along thetransport pathway. However, in all other respects, the effluxpatterns of glyphosate and PTS were similar. Both compounds
I-I0
I-
C-r-
w w | sTRANSPORT 1005
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021
.PlantPhysiol. Vol. 82, 1986
accumulated in the second minor vein-cell wall phase of theefflux curve. This indicates a resistance to movement into thesymplast compared to the movement of atrazine (Fig. 5).Glyphosate and PTS (Fig. 7) both demonstrated a rapid de-
crease in concentration with each increment of applied pressure
following the second plateau, indicating that glyphosate in theleaf symplast is unable to quickly penetrate the plasmalemma,and is retained in the leaf symplast as the cells lose volume underpressure.
These results support the intermediate permeability hypothesis(36), i.e. that compounds exhibiting the symplastic transport
pattern penetrate the symplast slowly, but once absorbed, areretained for long-distance transport. Compounds which displaythe apoplastic transport pattern are freely mobile between theapoplast and symplast.
Apparent Partition Coefficient. The apparent partition coeffi-cient, a measure of the lipid solubility and charge characteristicof a molecule, was determined for atrazine (Fig. 9A) and gly-phosate (Fig. 9B) in a 2-octanol/water system. These data indi-cate that atrazine is partitioned into the lipid phase of the systemto a much greater extent than glyphosate. The apparent partitioncoefficient for both atrazine and glyphosate remained essentially
25
w20
L 0
z2 15
I-
z
N
4 5
I.-
.4
, 60
w-
UL 48w
0
z 360
W-
: 24
I--
4 12co0
a.
0
3 2 4 6 8 10 12
B
_-
I .
IV,~~~~~~~,
2 4t 6 8 10 12
pH
FIG. 9. Apparent octanol/water partition coefficient as a function ofpH of the aqueous phase for (A) atrazine and (B) glyphosate. Verticalbars represent the SE of each plotted mean.
unchanged from pH 4 to 8, the range of physiologically signifi-
cant pH. These partition coefficients strongly reflect the watersolubility of the two herbicides; the water solubility for atrazineand glyphosate is 33 and 12,000 ppm, respectively (40). Sincethe pH of the buffer solution had no effect on the partitioncoefficient for glyphosate, a weak acid mechanism for glyphosateuptake cannot play a significant role in the symplastic transportof this compound (see also Gougler and Geiger [13]).The symplast/apoplast partition quotients of atrazine and
glyphosate (TableI) in tissue adjacent to the main veins of theleaf were determined. Since glyphosate and atrazine were presentin the whole leaf in such different concentrations (Figs. 2 and 3)and were distributed differently (Fig. 4), the symplastic concen-tration was determined only for the tissue near the veins, as thesetissues contained somewhat comparable concentrations of theherbicides. The tissue strips contained (4.1 ± 1.0) xl00 mol/gdry weight ofglyphosate and (2.3 ± 1.3) x 10-6 mol/g dry weightof atrazine.
The partition quotients for atrazine for a 1, 4, 7, or h darkperiod were approximately 7 times greater than those for gly-phosate. These data indicate that atrazine partitions into thesymplast to a greater extent than glyphosate. These data showthat approximately equal amounts of glyphosate and atrazinewere present in the symplast, though the concentration of atra-zine was an order of magnitude lower in the apoplast than was
glyphosate (Figs. 6 and 8).In an ideal situation, a compound which can diffuse rapidly
into or out of cells should have a symplast/apoplast quotient of1. The results reported for atrazine in this study would suggestaccumulation against a concentration gradient (Table I) and doesnot agree with previous reports of free and reversible penetrationin root tissues and protoplasts isolated from roots (6, 22, 24, 25,30, 31). However, the situation inChl containing tissues may bequite different from that observed in root tissues. It is quitepossible that the symplast/apoplast quotient determined for atra-zine in our system reflects the binding of this material to itstarget site in the chloroplast and partitioning into cell membranesin addition to its concentration in the symplastic fluid. Izawaand Good (1 6) reported three simultaneous absorption processesfor atrazine in isolated spinach chloroplasts: an irreversible bind-ing accounting for one molecule of inhibitor for every thousandChl molecules, a partitioning between the biological and aqueoussolvent phases which was independent of atrazine concentration,and a reversibly bound concentration which corresponded closelyto the degree of inhibition. More recently, Stemler and Murphy(33) have demonstrated the existence of both high and lowaffinity binding sites for atrazine in maize chloroplast, one highaffinity binding site per PSII complex was reported. Interactionsof this nature would remove atrazine from the cytoplasm andresult in an overestimate of the symplastic concentration of this
TableI. Symplast/Apoplast Partition Quotientsfor Atrazine andGlyphosate
Apoplast concentrations were determined graphically from expressedsap efflux curves between 0.3 and 0.4 MPa applied pressure. Symplastconcentrations were calculated from excised tissue samples adjacent to
the main veins of the leaf. Data are expressed as the mean ± SE for four
experiments for atrazine and three experiments for glyphosate.
Dark Equilibration Partition QuotientTime Atrazine Glyphosate
h533±268 76±34
4 544±287 96±257 710±62 99±2110 569 ± 278 102 ± 26
A
++S,---4 -4- ~~--- ---- 1- ,
1006 JACHETTAET AL.
i
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021

APOPLASTIC AND SYMPLASTIC HERBICIDE TRANSPORT
material.Our data for glyphosate demonstrates a high resistance to
movement across the plasmalemma (Fig. 7). From these results,we would predict an initial symplast/apoplast quotient of lessthan 1. While our data shows much less glyphosate partitioninginto the symplast than atrazine (Table I), the results presentedhere are considerably greater than predicted, and may indicatesome binding to cation exchange sites in the leaf apoplast.The difference in atrazine and glyphosate concentration in the
apoplast (Figs. 6 and 8) do reflect the differential permeability ofthese compounds observed in this study (Figs. 5 and 7). Theseresults are entirely consistent with the intermediate permeabilityhypothesis (36). Since the rate limiting step in membrane pene-tration is passage through the water-membrane interface (7), andthe activation energy for crossing this interface will be determinedby the same intermolecular forces which govern lipid:water par-tition coefficients (7), it would seem reasonable that the symplast/apoplast partitioning quotients for atrazine and glyphosate reflecttheir respective partition coefficients. This suggests that glyphos-ate's polar nature and resulting high water solubility are primarilyresponsible for its slower rate of penetration into the symplastand resulting accumulation in the apoplast.
LITERATURE CITED
1. ASHTON FM, AS CRAFTS 1973 Mode of Action of Herbicides. John Wiley andSons, New York
2. ASHTON FM, G ZWEIG, G MASON 1960 The effect ofcertain triazines on `CO2fixation in red kidney beans. Weeds 8: 448-451
3. CRAFTs AS, D CRISP 1971 Phloem Transport in Plants. WH Freeman and Co,San Francisco
4. CRISP CE, M LOOK 1978 Phloem loading and transport of weak acids. In GTBrooks, PC Kearney, eds, Advances in Pesticide Science, Part 3 pp 430-437.Fourth International Congress of Pesticide Chemistry, Zurich Switzerland,Pergamon Press, New York
5. DARMSTADT GL, NE BALKE, TP PRICE 1984 Triazine absorption by excisedcorn root tissue and isolated corn root protoplasts. Pestic Biochem Physiol21: 10-21
6. DEWEY SA, AP APPLEBY 1983 A comparison between glyphosate and assimi-late translocation patterns in tall morning glory (Ipomoea purpurea). WeedSci 31: 308-314
7. DIAMOND JM, EM WRIGHT 1969 Biological membranes: the physical basis ofion and nonelectrolyte selectivity. Annu Rev Physiol 31: 581-646
8. DYBING DC, HB CURRIER 1961 Foliar penetration by chemicals. Plant Physiol36: 169-174
9. EDGINGTON LV, CA PETERSON 1977 Systemic fungicides: theory, uptake andtranslocation. In MP Siegel, HD Sisler, eds, Antifungal Compounds, Vol 2,Interactions in Biological Ecological Systems. Marcel Dekker, New York
10. FERNANDEZ CH 1978 Absorption ofglyphosate [N-phosphonomethyl)-glycine]by leaf sections of tobacco Nicotiana tabacum "Xanthi." PhD Dissertation.University of California, Davis
1 1. FREAR DS, HR SWANSON 1970 The biosynthesis of S-(4-ethylamino-6-isopro-pylamino-2-s-triazine) glutathione: partial purification and properties of aglutathione S-transferase from corn. Phytochemistry 9: 2123-2132
12. GoTTRUP 0, PA O'SULLIVAN, RJ SCHRAA, WH VANDEN BORN 1976 Uptake,translocation, metabolism and selectivity of glyphosate. Weed Res 16: 197-201
13. GOUGLER JA, DR GEIGER 1981 Uptake and distribution of N-phosphonome-thylglycine in sugar beet plants. Plant Physiol 68:668-672
14. GRAHAM JC, K BUCHHOLTZ 1968 Alteration of transpiration and dry matterwith atrazine. Weed Sci 16: 389-392
15. HAY JR 1976 Herbicide transport in plants. In U Audus, ed, HerbicidesPhysiology, Biochemistry, Ecology, Vol 2. Academic Press, New York
16. IZAWA S, NE GOOD 1965 The number of sites sensitive to 3-(3,4-Dichloro-phenyl)-l,l-dimethylurea, 3-(4-Chlorophenyl)-l,I-dimethylurea and 2-chloro-4-(2-propylamino)6-ethylamino-s-triazine in isolated chloroplasts.Biochim Biophys Acta 102: 20-38
17. JACHETTA JJ, SR RADOSEVICH 1981 Enhanced degradation of atrazine by corn(Zea mays L.). Weed Sci 29: 37-44
18. JACHETTA JJ, AP APPLEBY, L BOERSMA 1986 Use of the pressure vessel tomeasure concentrations ofsolutes in apoplastic and membrane-filtered sym-plastic sap in sunflower leaves. Plant Physiol 82: 995-999
19. MARQUIs LY, RD COMES, CP YANG 1979 Selectivity ofglyphosate in creepingred fescue and reed canary grass. Weed Res 19: 335-342
20. ORWICK PL, MM, SCHREIBER, TK HoDGEs 1976 Absorption and efflux ofchloro-s-triazines by Setaria roots. Weed Res 16: 139-143
21. PETERSON CA, PPQ DEWILDT, LV EDGINGTON 1978 A rationale for theambimobile translocation of the nematicide oxamyl in plants. PesticBiochem Physiol 8: 1-9
22. PETERSON CA, LV EDGINGTON 1976 Entry ofpesticides into the plant symplastas measured by their loss from an ambient solution. Pestic Sci 7: 483-491
23. PETERSON CA, M GRIFFITH, NPA HUNER 1985 Permeability of the suberizedmestome sheath in winter rye. Plant Physiol 77: 157-161
24. PRICE TP, NE BALKE 1982 Characterization of rapid atrazine absorption byexcised velvetleaf (Abutilon theophrasti) roots. Weed Sci 30: 633-639
25. PRICE TP, NE BALKE 1983 Characterization of atrazine accumulation byexcised velvetleaf(Abutilon theophrasti) roots. Weed Sci 31: 14-19
26. SANDBERG CL, WF MEGGIrT, D PENNER 1980 Absorption, translocation andmetabolism of 'C-glyphosate in several weed species. Weed Res 20: 195-200
27. SCHULTZ ME, OC BURNSIDE 1980 Absorption, translocation, and metabolismof 2,4-D and glyphosate in hemp dogbane (Apocynum cannabinum). WeedSci 28: 13-20
28. SHANER DL 1978 Effect ofglyphosate on transpiration. Weed Sci 26: 513-51629. SHANER DL, SL LYON 1980 Interaction of glyphosate with aromatic amino
acids in transpiration in Phaseolus vulgaris. Weed Sci 28: 31-3530. SHONE MGT, BO BARTLET, AV WOOD 1974 A comparison of the uptake and
translocation ofsome organic herbicides and a systemic fungicide by barley.II. Relationship between uptake by roots and translocation by shoots. J ExpBot 25: 401-409
31. SHONE MGT, AV WOOD 1974 A comparison of the uptake and translocationof some organic herbicides and a systemic fungicide by barley. I. Absorptionin relation to physiochemical properties. J Exp Bot 25: 390-400
32. SMITH D, KP BUCCHOLTZ 1964 Modification of plant transpiration rate withchemicals. Plant Physiol 39: 572-578
33. STEMLER A, J MURPHY 1984 Inhibition of HCO3-binding to photosystem IIby atrazine at a low-affinity herbicide binding site. Plant Physiol 77: 179-182
34. SPRANKLE P, WF MEGGITT, D PENNER 1975 Absorption, mobility, and micro-bial degradation of glyphosate in the soil. Weed Sci 23: 229-234
35. STRUGGER S 1949 Praktikum der Zell- und Gewebephysiology der Pflanze.Springer-Verlag, Berlin
36. TYREE MT, CA PETERSON, LV EDGINGTON 1979 A simple theory regardingambimobility ofxenobiotics with special reference to the nematicide, oxamyl.Plant Physiol 63: 367-374
37. VAN BEL AJE 1978 The free space of the xylem translocation pathway of thetomato stem. J Exp Bot 29: 295-303
38. VOSTRAL HJ, KP BUCCHOLTZ, CA KUST 1970 Effect of root temperature onabsorption and translocation of atrazine in soybeans. Weed Sci 18: 115-117
39. WARD TM, K HOLLY 1966 The sorption of s-triazines by model nucleophilesas related to their partitioning between water and cyclohexane. J ColloidInterface Sci 22: 221-230
40. Weed Science Society of America 1983 Herbicide Handbook of the WeedScience Society of America, Ed 5. WSSA, Champaign, IL
41. WYRILL JB III, OC BURNSIDE 1976 Absorption, translocation, and metabolismof 2,4-D and glyphosate in common milkweed and hemp dogbane. WeedSci 24: 557-566
42. ZANDSTRA BH, RK NISHIMOTO 1977 Movement and activity of glyphosate inpurple nutsedge. Weed Sci 25: 268-274
43. ZWEIG G, FM ASHTON 1962 The effect of 2-chloro4-)ethylamino)-6-(isopro-pylamino)-s-triazine (atrazine) on distribution of '4C compounds following14CC2 fixation in excised kidney bean leaves. J Exp Bot 13: 5-11
1007
Dow
nloaded from https://academ
ic.oup.com/plphys/article/82/4/1000/6082066 by guest on 28 N
ovember 2021


















