
Aortic smooth muscle cells are able to convert angiotensin I to angiotensin II
6
Vol. 173, No. 3, 1990 December 31, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Pages 1137-1142 AORTIC SMOOTH MUSCLE CELLS ARE ABLE TO CONVERT ANGIOTENSIN I TO ANGIOTENSIN II Philippe ANDRE 1, Chrism SCHOTT, Herrade NEHLIG and Jean-Claude STOCLET Laboratoire de Pharmacologic Mol6culaire et Cellulaire CNRS URA600, Universit6 Louis Pasteur de Strasbourg, B.P. 24, 67401 Illkirch, France Received October 5, 1990 SUMMARY: The role of vascular smooth muscle cells (VSMC) in the intraparietal conversion of angiotensin I (AngI) to angiotensin II (AngII) was investigated in rat aortic tissue. The responses of rat aortic vascular smooth muscle cells to AngI and AnglI were assessed by studying contraction of endothelium-denuded aortic rings and by measuring intracellular Ca ++ ion concentration in primary cultures of VSMC free of endothelial cells. In both preparations, AngI and AnglI induced identical responses which were inhibited by saralasin, a blocker of AnglI receptors. In the presence of captopril, an inhibitor of the angiotensin converting enzyme, the increase in calcium caused by AngI was abolished in VSMC cultures and the contractile effect of this peptide in aortic rings was strongly decreased, whereas the responses to AngII remained unaffected. These results demonstrate that VSMC are able to convert AngI to AngH. © 1990 Academic Press, Inc. Angiotensin converting enzyme (ACE) is a dipeptidyl carboxipeptidase (EC 3.4.15.11) which cleaves AngI to AngII, a potent vasoconstrictor agent. It is well established that circulating AngI, produced by renin, has no direct vasoconstrictor effect but it is converted into AngII, by circulating and endothelial ACE (1-2). This mechanism has a major importance in the regulation of vascular tone. However, evidence exists suggesting the presence of a renin-angiotensin system in the vessel wall: the presence of renin in smooth muscle and endothelial cells (3) and of angiotensinogen mRNA (4) in the rat aorta adventitia and periaortic brown adipose tissue have previously been reported. Futhermore, AngI induces contractions of isolated endothelium- denuded vessels (5). It has been also suggested that the local production of AnglI could play an autocrine or paracrine role on VSMC (6). However, the localization of a functional extra- endothelial ACE in the vascular wall is uncertain, due to the presence of various cell types. The aim of the present work was to examine whether vascular smooth muscle cells are able to convert AngI to AngII. An ACE activity-dependent effect of AngI on AngII receptors has been demonstrated on rat aortic smooth muscle cells in these experiments. 1To whom correspondence should be addressed. 1137 0006-291X/90 $1.50 Copyright © 1990 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
philippe-andre -
Category
Documents
-
view
212 -
download
0
Transcript of Aortic smooth muscle cells are able to convert angiotensin I to angiotensin II

Vol. 173, No. 3, 1990
December 31, 1990
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 1137-1142
AORTIC SMOOTH MUSCLE CELLS ARE ABLE TO CONVERT ANGIOTENSIN I TO ANGIOTENSIN II
Philippe ANDRE 1, Chrism SCHOTT, Herrade NEHLIG and Jean-Claude STOCLET
Laboratoire de Pharmacologic Mol6culaire et Cellulaire CNRS URA600,
Universit6 Louis Pasteur de Strasbourg, B.P. 24, 67401 Illkirch, France
Received October 5, 1990
SUMMARY: The role of vascular smooth muscle cells (VSMC) in the intraparietal conversion of angiotensin I (AngI) to angiotensin II (AngII) was investigated in rat aortic tissue. The responses of rat aortic vascular smooth muscle cells to AngI and AnglI were assessed by studying contraction of endothelium-denuded aortic rings and by measuring intracellular Ca ++ ion concentration in primary cultures of VSMC free of endothelial cells. In both preparations, AngI and AnglI induced identical responses which were inhibited by saralasin, a blocker of AnglI receptors. In the presence of captopril, an inhibitor of the angiotensin converting enzyme, the increase in calcium caused by AngI was abolished in VSMC cultures and the contractile effect of this peptide in aortic rings was strongly decreased, whereas the responses to AngII remained unaffected. These results demonstrate that VSMC are able to convert AngI to AngH. © 1990 Academic Press, Inc.
Angiotensin converting enzyme (ACE) is a dipeptidyl carboxipeptidase (EC 3.4.15.11) which
cleaves AngI to AngII, a potent vasoconstrictor agent. It is well established that circulating
AngI, produced by renin, has no direct vasoconstrictor effect but it is converted into AngII, by
circulating and endothelial ACE (1-2). This mechanism has a major importance in the regulation
of vascular tone. However, evidence exists suggesting the presence of a renin-angiotensin
system in the vessel wall: the presence of renin in smooth muscle and endothelial cells (3) and
of angiotensinogen mRNA (4) in the rat aorta adventitia and periaortic brown adipose tissue
have previously been reported. Futhermore, AngI induces contractions of isolated endothelium-
denuded vessels (5). It has been also suggested that the local production of AnglI could play an
autocrine or paracrine role on VSMC (6). However, the localization of a functional extra-
endothelial ACE in the vascular wall is uncertain, due to the presence of various cell types.
The aim of the present work was to examine whether vascular smooth muscle cells are able to
convert AngI to AngII. An ACE activity-dependent effect of AngI on AngII receptors has been
demonstrated on rat aortic smooth muscle cells in these experiments.
1To whom correspondence should be addressed.
1137
0006-291X/90 $1.50 Copyright © 1990 by Academic Press, Inc.
All rights of reproduction in any form reserved.

Vol. 173, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
MATERIALS AND METHODS
Cell cultures Primary cultures of aortic smooth muscle cells from 10 to 12 week-old Wistar rats were obtained as previously described (7). The aorta was treated with CLII collagenase at 70 U/ml in Hank's balanced salt solution for 30 rain at 37°C. The adventitia was stripped mechanically and the endothelium was removed with a fine paint brush after the vessel had been opened longitudinally. The aorta was incubated with an elastase-collagenase digestion medium. After 90 min, the tissue was homogenized and centrifuged. The pellet was resuspended in MEM with 2% Ultroser G and the cells were plated in 25 cm 2 flasks. After a 2 h-preplating, the supernatant was transferred to a new flask and seeded at 3x105 cells/ml on sterile glass coverslips. The medium was changed every two days. Endothelial cells were collected from fresh bovine aorta using collagenase digestion as described by Schini et al (8). The endothelial cells were cultivated on 25 cm 2 plastic flasks in MEM/HAM F12 (1:1) supplemented with 20% foetal calf serum and were used at the third passage.
Fluorescence microscopy Endothelial cells and vascular smooth muscle cells were distinguished by the selective kinetic uptake of acetylated-low density lipoproteins (9). Briefly, confluent endothelial cells or VSMC were incubated for 4 hours at 37°C with 10 ~tg/ml of lipoproteins labelled with the fluorescent probe 1,1'dioctadecyl 3,3,3',3'tetramethyl-indocarbocyanine perchlorate (Di I-Ac-LDL). VSMC and endothelial cells were washed three times with a physiological solution for 10 min and fixed in 4% paraformaldehyde in 0.12M Na/K phosphate buffer saline.The samples were examined with a fluorescence microscope (Zeiss).
Intracellular calcium measurement Intracellular free calcium concentration (Ca+÷i) was measured according to the method used by Cornwell and Lincoln (10). Fifteen day-old VSMC cultures, grown on glass coverslips, were washed 3 times with 2 ml of balanced salt solution (BSS). The glass coverslip was inserted in quartz spectrophotometric cuve. 3 ml of Fura-2/AM,5 p.M, were added to the cells and incubated 30 min at 37°C. The cells were washed 3 times with BSS without bovine serum albumin and incubated for an additional 20 min to ensure complete hydrolysis of the Fura-2/acetoxymethyl ester. Ca~i measurements were made in a SPEX 1681 spectrometer Fluorescence measurements were made with two different excitation wavelengths, 340 and 380 nm. The emission was measured at 510 nm. Calculation of Ca++i was done as described previously by Grynkiewicz et al.(11)
Contractile experiments Two mm-long aortic rings from 10-12 week old male Wistar rats were rubbed with fine forceps to remove the endothelium and were mounted under an initial tension of 2 g. The bath containing 20 mi of physiological solution (composition mM: NaC1 118; NaHCO3 25; glucose 10; KC1 4.7; CaC12 1.25; MgSO4 1.19; KH2PO4 1.14) was maintained at 37°C and bubbled with a mixture of 95% 02-5% CO2. After a 60 min equilibration, the tension was readjusted to 2 g. The absence of functional endothelium was assessed by the absence of a relaxing effect of acetylcholine 10-6M added when the maximal tension had been reached with 10-6M noradrenaline. Thereafter, tissues were washed every 15 min during 60 min. When indicated, saralasin or captopril were added 15 min before AngI or AngII addition. Isometric responses to AngI and AngII were recorded
Statistical analysis Statistical analysis was performed using the Wilcoxon test, taking P<0.05 as the limit of significance. The results were expressed as mean + standard error of the mean. n denotes the number of experiments.
Materials AngI, AngII, acetylcholine, CLII collagenase and noradrenaline, saralasin were purchased from Sigma. Captopril was obtained from Squibb. Fura-2/AM was from Molecular Probes. Di I-Ac-LDL was obtained from Biomedical Technology. Elastase was from Biosys. Ultroser G was from IBF.
1138

Vol. 173, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
RESULTS
Absence of endothelial cells in primary cultures of VSMC
To exclude the possible involvement of endothelial cell ACE in the tranformation of AngI to
AnglI in our VSMC cultures, we evaluated the presence of endothelial cells in VSMC cultures
by fluorescence staining. In the presence of Di I-Ac-LDL, endothelial cells showed an intense
fluorescence (fig. 1A). This staining was predominantly associated with both the cytoplasm and
the plasmalemma (8). In VSMC (fig. 1/3), low background fluorescence was observed. Only the
plasma membrane and the nucleus were slighty fluorescent. There was no staining of the
cytoplasm. No brightly fluorescent cells corresponding to endothelial cells were found in VSMC
cultures showing that our cultures are endothelial-free.
Effects of Angl and AnglI on cytosolic free calcium in cultured VSMC
Due to the rapid desensitization of the response to AnglI, we used only a single maximally active
dose of the peptide in each experiment. The effect of the peptide in the absence or in the presence
of antagonist or ACE inhibitor was compared on cells from the same batch.
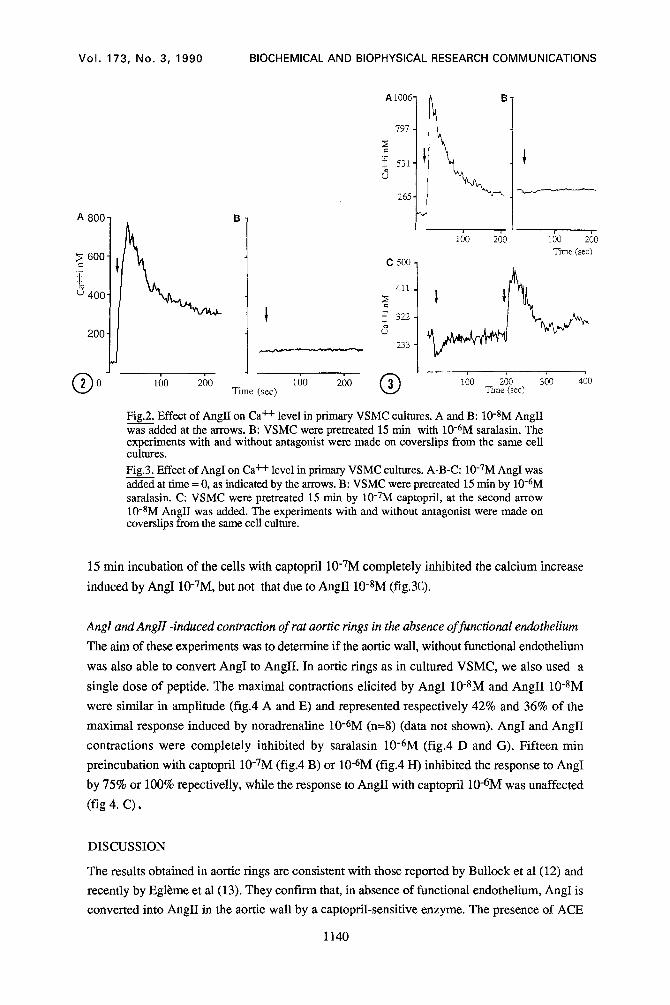
The effect of AngII on a 15 day-old VSMC culture is illustrated in fig 2A, In all experiments, the
addition of AnglI 10-SM induced a rapid rise in Ca+÷i from 2.3+0.5x10-7M (n=6) to
8.8+l.5x10-TM (n=6) (P<0.01) followed by a decrease to a plateau which remained higher than
the basal level for 10 min Incubation of the ceils with EGTA 5x10-3M abolished this plateau
phase and decreased the level of Ca++i by about 50% (data not shown). The addition of the
AnglI receptor antagonist saralasin 10-6M, 15 min before AnglI 10-SM addition, completely
inhibited the effect of AnglI on Ca~i (fig. 2/3)
In the same manner, AngI 10-7M increased the level of Ca÷+i from 2.9 + 1.1xl0-TM (n=7) to
2.3 + 1.6x10-6M (n=7) (P<0.05) (fig.3A) in VSMC, with the same biphasic time course. This
effect was also blocked by a 15 min preincubation with saralasin 10-6M (fig.3/3). Furthermore, a
~D Characterization of (A) bovine aortic endothelial cells and (B) rat aortic VSMC, i I-Ac-LDL fluorescence staining.
1139
Vol. 173, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
A 800-
:~ 600
% D 400'
200
0
t
100 2()0 Time (sec) 160 200 @
A 1006,
797,
~ 531'
265.
C 500
411
~ 322
233
16o £o 1~o 260 Time (sec)
1;0 2;0 3;0 400 Time (see)
Fig.2. Effect of AngH on Ca ++ level in primary VSMC cultures. A and B: 10-SM AngII was added at the arrows. B: VSMC were pretreated 15 min with 10-6M saralasin. The experiments with and without antagonist were made on coverslips from the same cell cultures. Fig.3. Effect of AngI on Ca ++ level in primary VSMC cultures. A-B-C: 10"7M AngI was added at time = 0, as indicated by the arrows. B: VSMC were pretreated 15 min by 10-6M saralasin. C: VSMC were pretreated 15 min by 10-TM captopril, at the second arrow 10-SM AnglI was added. The experiments with and without antagonist were made on coverslips from the same cell culture.
15 rain incubation of the cells with captopril 10-7M completely inhibited the calcium increase
induced by AngI 10-7M, but not that due to AnglI 10-SM (fig.3C).
AngI and Angll -induced contraction of rat aortic rings in the absence of functional endothelium
The aim of these experiments was to determine if the aortic wall, without functional endothelium
was also able to convert AngI to AngII. In aortic tings as in cultured VSMC, we also used a
single dose of peptide. The maximal contractions elicited by AngI 10-8M and AngII 10-8M
were similar in amplitude (fig.4 A and E) and represented respectively 42% and 36% of the
maximal response induced by noradrenaline 10-6M (n=8) (data not shown). AngI and AngII
contractions were completely inhibited by saralasin 10-6M (r iga D and G). Fifteen min
preincubation with captopril 10-TM (fig.4 B) or 10-6M (fig.4 H) inhibited the response to AngI
by 75% or 100% repectivelly, while the response to AngII with captopril 10-rM was unaffected
(fig 4. C),
DISCUSSION
The results obtained in aortic rings are consistent with those reported by Bullock et al (12) and
recently by Egl~me et al (13). They confn-m that, in absence of functional endothelium, AngI is
converted into AngII in the aortic wall by a captopril-sensitive enzyme. The presence of ACE
1140

Vol. 173, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
~5
2
2
1
I
0
A B C D E F G H
±
Fig.4. Effects of AngI and AngIl on tension of rat aortic rings in the absence of functional endothelium. AngI 10"8M (A), captopril 10-7M + AngI 10-8M (B), captopril 10-TM + AngI 10-8M + AnglI 10-SM (C), saralasin 10-6M + AngI 10-SM (D), AnglI 10-8M (E), captopril 10-TM + AnglI 10-SM (F), saralasin 10-6M + AnglI 10-8M (G), captopril 10-6M + AngI 10-SM. (n=8) Tension of the rings was normalized to the dry tissue weight.
activity in rat aortic homogenates has been also described previously (14-16). However the rat
aorta contains a large proportion of various types other than smooth muscle cells, due to the
presence of adventitia. In addition we cannot exclude the presence of residual ACE containing
endothelial cell membranes after removal of functional endothelinm.
The interpretation of the experiments performed on cultured VSMC does not meet the same
problems as isolated aorta experiments. Our results with primary cultured VSMC show that in
the absence of endothelial cells, VSMC themselves are able to convert AngI into active AnglI
and to respond to the latter by a rapid, saralasin-sensitive rise in Ca++i. The presence of specific
binding sites for ramipril, an ACE inhibitor, has been reported by Larrue et al (17) in human
arterial VSMC. Rosenthal et al (18) have found an ACE activity only at pH 8.5 in cultured rat
aortic VSMC at the 5 th and 6 th passage, while in rat aortic extracts, ACE activity was higher
and was detected between pH 7 and pH 9.5. The present results constitute the first evidence that
VSMC possess functional ACE in conditions in which they also possess functional AnglI
receptors (as characterized by the calcium signal and the inhibition by saralasin).
It is interesting to note that several authors have found the presence of other compounds
implicated in the renin-angiotensin system in the vessel wall: Dzan et al (19) have shown by
immunostaining, the presence of renin in vascular media and intima, while Cassis et al (20) have
described the presence of angiotensinogen mRNA in rat adventitia and in periaortic brown
adipose tissue (but not in VSMC). It appears that rat aorta possess the three components of the
renin-angiotensin system: renin, angiotensinogen and ACE. Thus, local conversion of Ang I to
Ang II by VSMC may influence vascular tone.
In conclusion, we demonstrate here for the first time the presence of a functional, captopril-
sensitive ACE activity in rat aortic VSMC in primary culture.
Acknowledgments: We are grateful to Dr. Herrade Stoeckel for her assistance in the fluorescence study. We would also like to express our appreciation to RIOM-Laboratoires CERM for their financial contribution.
1141

Vol. 173, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
REFERENCES
1. Ryan, J.W., Chung, A., Ammons, C., and Carlton, M.L. (1977) Biochem. J. 167. 501- 504.
2. Ondetti, M.A., and Cushman, D.W. (1982) Annu. Rev. Biochem. 51. 238-308. 3. Dzau,V.J., (1989) J. Cardiovasc. Pharmacol. 14 (Suppl.4). S1-$5. 4. Cassis, L.A., Lynch, K.R., and Peach, M.J. (1988) Circ. Res. 62. 1259-1262. 5. Juul, B., Aalkjaer, C., and Mulvany, M.J.(1986) Eur. J. Pharmacol. 135.61-68. 6. Dzau, V.J., and Gibbons, G.H. (1987) Am. J. Cardiol. 60. 991-1006. 7. Travo, P., Barret, G., and Burnstock, G. (1980) Blood Vessels 17. 110-116. 8. Schini, V, Grant, N.J., Miller, R., and Takeda, K. (1988) Eur. J. Cell Biol. 47.53-61. 9. Voyta, J.C., Via, D.P., Butterfield, C.E., and Zetter, B.R. (1984) J. Cell. Biol. 90. 2034-
2040. 10. Comwell, T.L., and Lincoln, T.M. (1989) J. Biol. Chem. 264. 1146-1155. 11. Grynkiewicz, G., Poenie, M., and Tsien, R.Y. (1985) J. Biol. Chem. 260. 3440-3450 12. Bullock, G.R., Taylor, S.G., and Weston, A.H. (1986) Br. J. Pharmacol. 89. 525-532. 13. Egl~me, C., Cressier, F., and Wood, J.M. (1990) Br. J. Pharmacol. 100. 237-240. 14. Welsch, C., Grima, M., Giesen, E.M., Helwig J.J., Barthelmebs, M., Coquard, C., and
Imbs, J.L. (1989) J. Cardiovasc. Pharmacol. 14 (Suppl. 4). $26-$31. 15. Johnson, H., and Drummer, O.H. (1988) Biochem. Pharmac. 37. 1131-1136. 16. Okunishi, H., Miyazaki, M,. Okamura, T., and Toda, N. (1987) Biochem. Biophys. Res.
Commun. 149. 1186-1192. 17. Larrue, J., Demond-Henri, J., and Daret, D. (1989) J. Cardiovasc. Pharmacol. 14 (Suppl.
4). $43-$45. 18. Rosenthal, J.H., Pfeile, B., Michailov, M.L., Pschorr, J., Jacob, I.C.M., and Dahlheim,
H. (1983) Hypertension 6. 383-390. 19. Dzau, V.J. (1987) In: Nijhoff M, ed. Arterial and Venous Systems in Essential
Hypertension. Amsterdam: Martinus Nijhoff,: 153-164. 20. Cassis, L.A., Saye, J., and Peach, M.J. (1988) Hypertension 11. 591-596.
1142


















