
CHEMISTRY STOICHIOMETRY & COLLOIDS STOICHIOMETRY & COLLOIDS.

Stoichiometry and Population Dynamics
Andersen et al. 2004 Ecology LettersLauren Yamane
1Tuesday, November 13, 12

Food quality is important not just quantity
• Preference for high quality food (high nutrient: C ratio) in terrestrial studies (White 1993, Sterner & Elser 2002)
2Tuesday, November 13, 12

Problem: Consumers require relatively constant nutrient
ratio from low nutrient autotrophs
• Autotroph nutrient: C ratio often highly variable
• Variability due to physiological plasticity under varying conditions
3Tuesday, November 13, 12

Constraints in growth and population dynamics due to
food quality
• Link: Threshold Elemental Ratio (TER)
• TER = critical C: nutrient ratio
• Above TER get nutrient limitation
• Transfer efficiency decreases
• Below TER get C (Energy) limitation
4Tuesday, November 13, 12

Herbivores can have both negative and positive density dependent effects on
plant nutrient composition
• Negative: Herbivores remove the good stuff (traditional; Turchin 2003)
• Positive: Herbivores reduce biomass but nutrient input remains same + recycle nutrients Nutrient: C
5Tuesday, November 13, 12
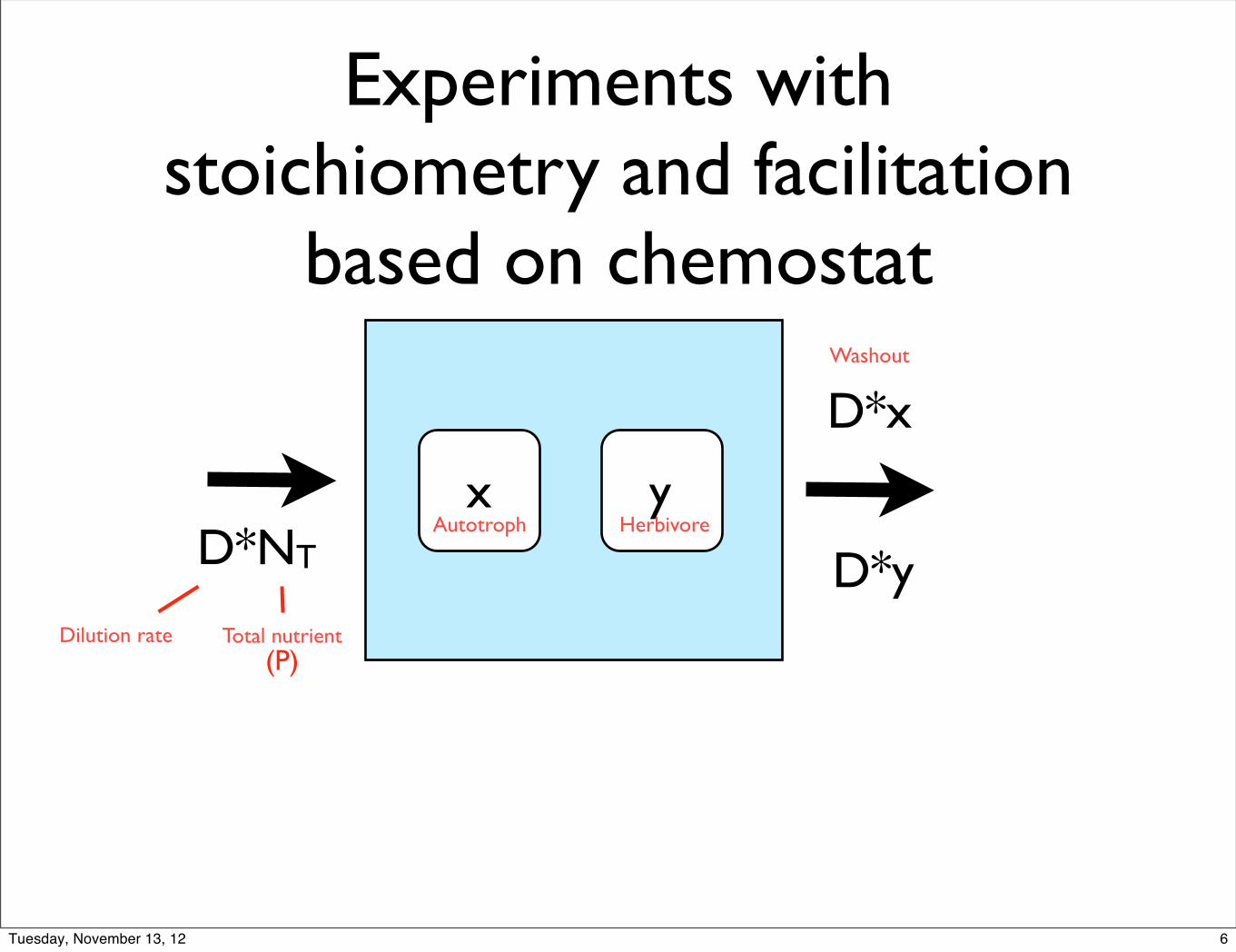
Experiments with stoichiometry and facilitation
based on chemostat
x yD*NT
Total nutrientDilution rate
Autotroph Herbivore
D*x
D*y
Washout
(P)
6Tuesday, November 13, 12
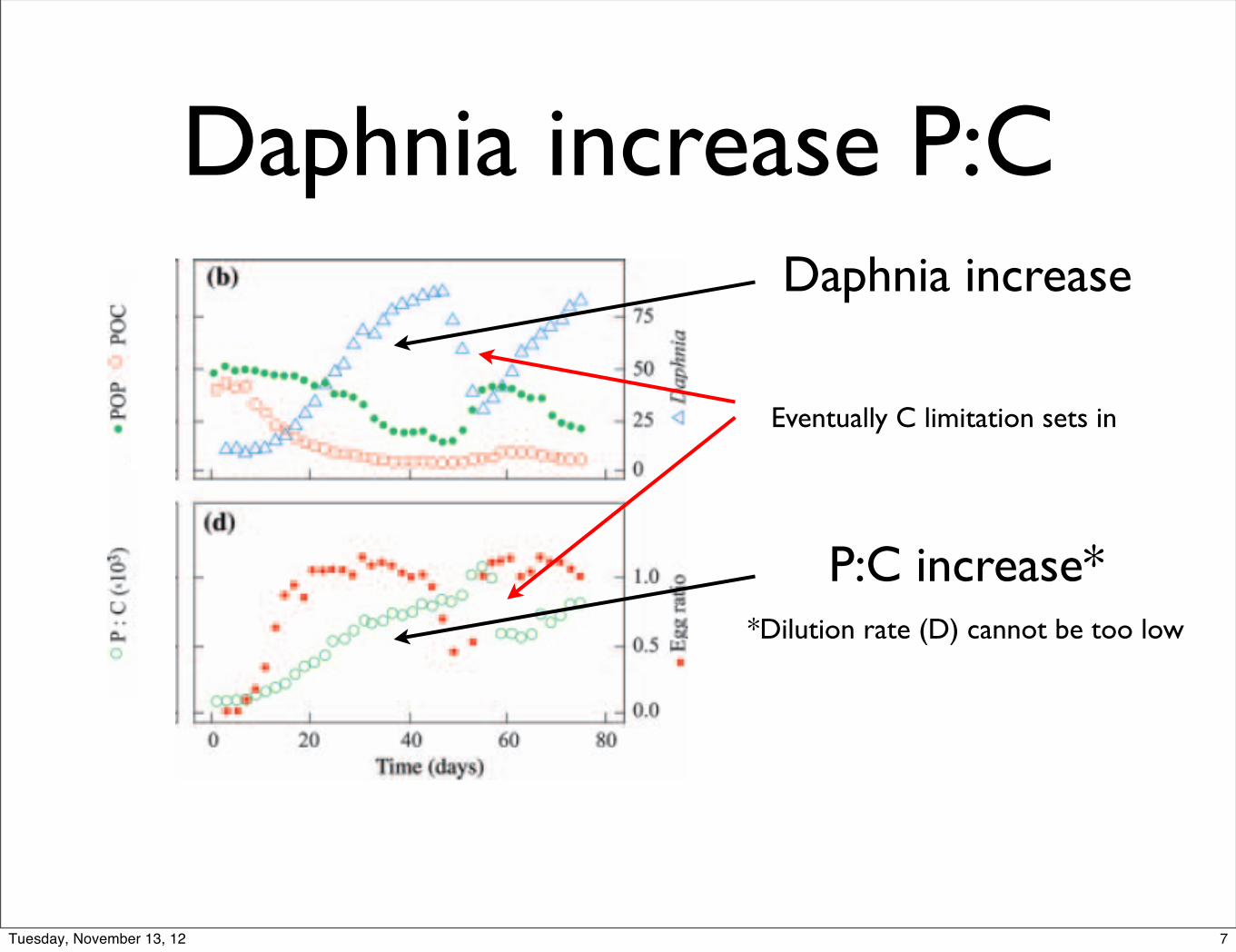
Daphnia increase P:C Daphnia increase
P:C increase**Dilution rate (D) cannot be too low
Eventually C limitation sets in
7Tuesday, November 13, 12

Stoichiometry alters parameters in Lotka-Volterra
predator-prey model
• Modeling autotroph (A), herbivore (H) dynamics
• Like chemostat model
• (constant) Dilution rate sets nutrient input and washout of 2 species
Autotroph growth rate Herbivore function response
Herbivore numerical response
8Tuesday, November 13, 12

Functional responses (f.r.) reflect ingestion and handling
time constraintsHolling’s Type I
Holling’s Type 2 (handling time)
9Tuesday, November 13, 12

10Tuesday, November 13, 12

Traditional Lotka-Volterra models
Density independent growth (k1), Type I (I1)
Density independent growth (k1), Type II
(I2)
Density dependent growth (k2), Type I (I1)
Density dependent growth (k2), Type II
(I2)
Rosenzweig-MacArthur model
-Neutral stability -Asymptotic instability
y nullcline moves R
-Asymptotic stability
-Hopf bifurcation from stable focus to limit cycle Autotroph biomass (x)
Herbivore biomass (y)
11Tuesday, November 13, 12
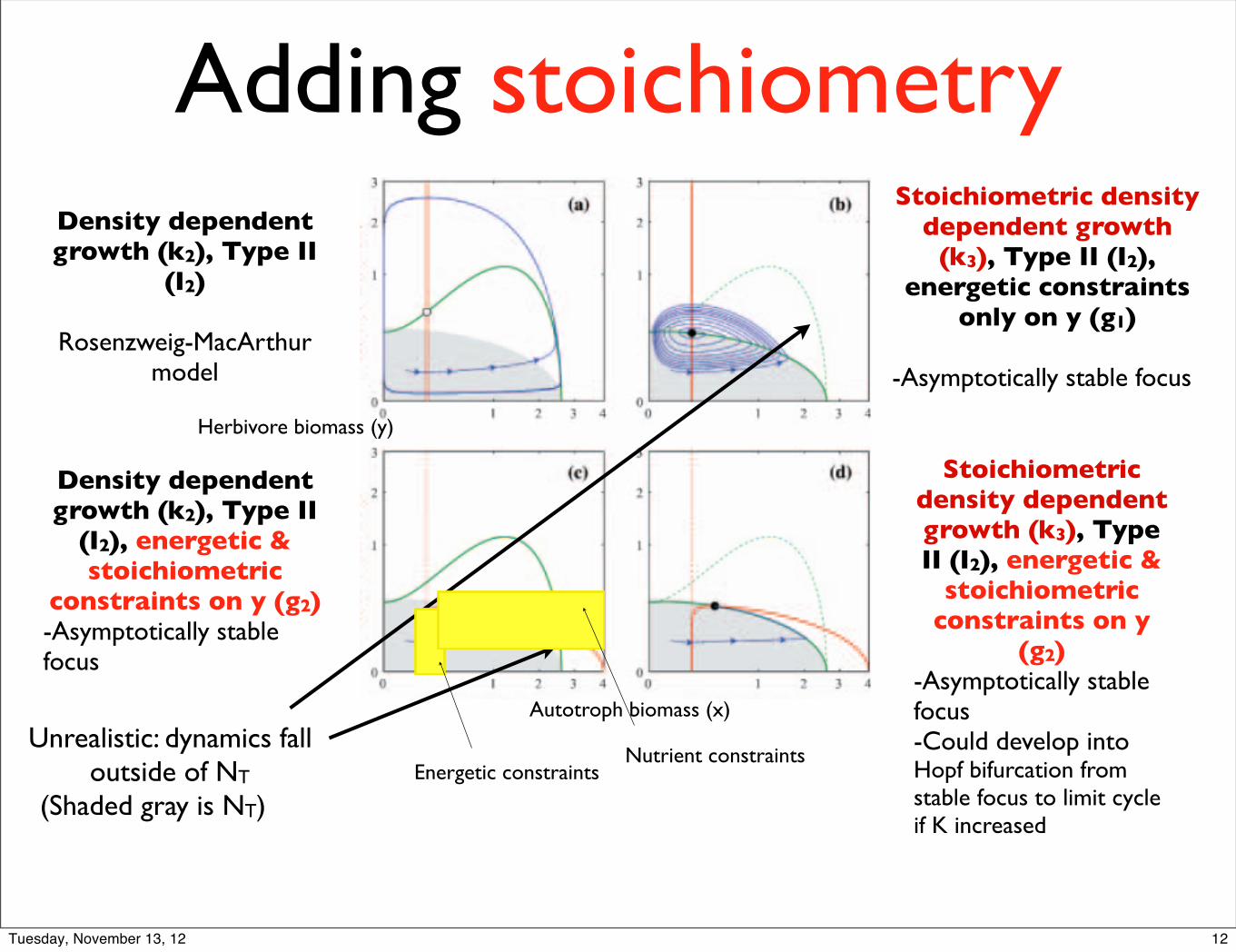
Adding stoichiometry
Unrealistic: dynamics fall outside of NT
Density dependent growth (k2), Type II
(I2)
Rosenzweig-MacArthur model
(Shaded gray is NT)
Stoichiometric density dependent growth
(k3), Type II (I2), energetic constraints
only on y (g1)
-Asymptotically stable focus
Density dependent growth (k2), Type II
(I2), energetic & stoichiometric
constraints on y (g2)-Asymptotically stable focus
Stoichiometric density dependent growth (k3), Type II (I2), energetic &
stoichiometric constraints on y
(g2)-Asymptotically stable focus -Could develop into Hopf bifurcation from stable focus to limit cycle if K increased
Energetic constraintsNutrient constraints
Autotroph biomass (x)
Herbivore biomass (y)
12Tuesday, November 13, 12

Stability and persistence with stoichiometry depend on
nutrient level and D
Herbivore biomass (y)
Autotroph biomass (x)
Low NT, Low D
Low NT, High D
High NT, High D
*High NT, Low D
13Tuesday, November 13, 12

Stability and persistence with stoichiometry depend on
nutrient level and D
*
14Tuesday, November 13, 12

Time series of full stoichiometric model
Autotroph
Herbivore
Nutrient limited
Nutrient sufficient Nutrient limited
Energy limited
15Tuesday, November 13, 12

Conclusions
• Stoichiometry can have stabilizing effects on predator-prey dynamics
• Must include stoichiometric effects in both autotroph growth rate and herbivore numerical response for realistic dynamics
• Future directions: additional work on stoichiometry changes due to ontogeny and effects on pop dynamics
16Tuesday, November 13, 12

Questions
• Would functional response ever have stoichiometric constraints? Alter satiation in parameter?
• Stoichiometry important beyond nutrient-limited autotroph herbivore model?
17Tuesday, November 13, 12


















