Analysis of gene expression during development in the...
13
/. Embryol exp. Morph. Vol. 34, 1, pp. 19-31, 1975 19 Printed in Great Britain Analysis of gene expression during development in the homeotic mutant Contrabithorax of Drosophila melanogaster By GINES MORATA 1 From the Institute) de Genetica y Antropologia, Centro de Investigaciones Biologicas, Madrid, Spain SUMMARY Contrabithorax, a mutant of the bithorax system in Drosophila melanogaster produces a partial homeotic transformation of mesothorax (wing) into metathorax (haltere). The wing of a fly homozygous or heterozygous for the mutant is a mosaic of wing and haltere structures. A genetic analysis of the mutant suggests that its phenotype is due to some form of derepression in the wing of two other genes of the bithorax system {bithorax and post- bithorax) which are not normally active there. This repression is not complete. The activity of the two genes is below the normal level resulting in only a partial transformation of wing into haltere. Clones of marked cells were generated by X-rays and were found to include both transformed (haltere) and untransformed (wing) territory; this was true even for those generated late in development. Thus the final expression of a cell depends not on its immediate ancestry but perhaps on the level of the products of the wild-type alleles of bithorax and postbithorax. INTRODUCTION Mutants belonging to a pseudoallelic series, the 'bithorax' system, produce several kinds of homeotic transformations involving the mesothoracic, meta- thoracic and first abdominal segments of Drosophila melanogaster (Lewis, 1963, 1964). Amongst them, the mutant Contrabithorax (Cbx) leads to the differentia- tion in the mesothorax (wing) of structures typical of the metathorax (haltere). At the phenotypic level the main characteristic of Cbx is that it presents partial expression, that is, only a fraction of the cells in the wing segment show mutant phenotype, the rest of them being wild type. Since the cells are genetically identical and develop in the same developmental system, we are faced with the problem of what mechanisms are responsible for this differential behaviour. Moreover, assuming that the development of a haltere or a wing requires the function of not one but several specific genes, the expression in the Cbx meso- thoracic cells of the mutant (haltere) or wild-type (wing) phenotype requires the co-ordinate induction and/or repression of specific sets of genes. 1 Author's address: MRC Laboratory of Molecular Biology, University Postgraduate Medical School, Hills Road, Cambridge, U.K.
Transcript of Analysis of gene expression during development in the...

/. Embryol exp. Morph. Vol. 34, 1, pp. 19-31, 1975 19Printed in Great Britain
Analysis of gene expressionduring development in the homeotic mutantContrabithorax of Drosophila melanogaster
By GINES MORATA1
From the Institute) de Genetica y Antropologia, Centro deInvestigaciones Biologicas, Madrid, Spain
SUMMARYContrabithorax, a mutant of the bithorax system in Drosophila melanogaster produces
a partial homeotic transformation of mesothorax (wing) into metathorax (haltere). Thewing of a fly homozygous or heterozygous for the mutant is a mosaic of wing and halterestructures. A genetic analysis of the mutant suggests that its phenotype is due to some formof derepression in the wing of two other genes of the bithorax system {bithorax and post-bithorax) which are not normally active there. This repression is not complete. The activityof the two genes is below the normal level resulting in only a partial transformation of winginto haltere.
Clones of marked cells were generated by X-rays and were found to include both transformed(haltere) and untransformed (wing) territory; this was true even for those generated late indevelopment. Thus the final expression of a cell depends not on its immediate ancestry butperhaps on the level of the products of the wild-type alleles of bithorax and postbithorax.
INTRODUCTION
Mutants belonging to a pseudoallelic series, the 'bithorax' system, produceseveral kinds of homeotic transformations involving the mesothoracic, meta-thoracic and first abdominal segments of Drosophila melanogaster (Lewis, 1963,1964). Amongst them, the mutant Contrabithorax (Cbx) leads to the differentia-tion in the mesothorax (wing) of structures typical of the metathorax (haltere).
At the phenotypic level the main characteristic of Cbx is that it presentspartial expression, that is, only a fraction of the cells in the wing segment showmutant phenotype, the rest of them being wild type. Since the cells are geneticallyidentical and develop in the same developmental system, we are faced withthe problem of what mechanisms are responsible for this differential behaviour.Moreover, assuming that the development of a haltere or a wing requires thefunction of not one but several specific genes, the expression in the Cbx meso-thoracic cells of the mutant (haltere) or wild-type (wing) phenotype requires theco-ordinate induction and/or repression of specific sets of genes.
1 Author's address: MRC Laboratory of Molecular Biology, University PostgraduateMedical School, Hills Road, Cambridge, U.K.

20 G. MORATA
One possible explanation is that phenotypic mosaicism might stem from adeterminative event somewhere in the development of the mutant disc whichproduces separate lineages of cells which either differentiate the mutant or thewild-type phenotype (see Postlethwait & Schneiderman, 1969, 1971).
Taking advantage of the previous genetic information about the bithoraxsystem (Lewis, 1963,1964,1967) and how it controls the development of varioussegments (Morata & Garcia-Bellido, in preparation), I have made a geneticand clonal analysis of the mutant Cbx in an attempt to understand the processesthat lead to the expression of this phenotypic diversity.
The main results suggest that: (1) the partial expression of the Cbx genotypeis due to partial expression of the bithorax system in the mutant wing; (2) thosecells which express the mutant phenotype are not related to each other: smallclones generated by X-irradiation late in development often include both trans-formed and untransformed cells.
MATERIALS AND METHODS
bithorax mutants
The following mutants were used in this work: bx3,pbx, Cbx, Ubx1 and Ubx130.All map at 3-58-8. Several alleles are known of bx and Ubx but only one forpbx and Cbx. The mutant Ubx1Z0 is associated with In (3LR) Ubx130 (see Lindsley& Grell, 1968).
The translocation T(13)P115 was also used. It is an insertional translocation,discovered by E. B. Lewis, carrying a fragment of the third chromosome(including all bithorax genes) to the first chromosome proximal to forked. Theaneuploid segregant Df{3)P115f + is viable but sterile and shows the same pheno-type as Ubxj +.
Cell marker mutants
The mutant multiple wing hairs (mwh) was used. It is located at 3-0-0 andaffects trichome differentiation producing several processes per cell instead ofone. It can be identified even in very small clones in both wing and haltere(Garcia-Bellido & Merriam, 1971a; Morata & Garcia-Bellido, in preparation).
Irradiation
In mitotic recombination experiments, larvae were irradiated at 1000 R(R = 2-58 x 10~4 C/kg) using an X-ray machine Phillips 151 Be at standardworking conditions of 150 kV at 15 mA with a 2 mm aluminium filter.
Larvae of all ages were irradiated and their ages at irradiation calculatedaccording to the time between irradiation and pupariation.
Hatched adults were treated with hot KOH 10 % for lOmin to digest softparts, washed in alcohol and mounted in Euparal for microscopical examination.

Gene expression in Cbx 21
RESULTS
(1) Some characteristics of the bithorax system
Almost all the information known today about the bithorax system comesfrom the studies of Lewis (1963, 1964, 1967) and the reader is strongly advisedto consult his articles for a full description of the different mutants.
The bithorax system contains at least five loci, three of which produce recessivemutations {bithorax, bithoraxoid and postbithorax) and two others (Ultrabi-thorax and Contrabithorax) produce dominant ones. (Another two mutants,Contrabithoraxoid and Ultraabdominal, have been described by Lewis (1968)but as yet little information about them is available.) Although the homeotictransformations produced by these mutants affect both dorsal and ventralstructures, only dorsal transformations will be considered in this report.
The mutants in the bithorax (bx) locus produce a transformation of the an-terior haltere into anterior wing, the mutant postbithorax (pbx) transforms theposterior haltere into posterior wing. Thus flies homozygous for bithorax andpostbithorax have four wings and no halteres. The mutant bithoraxoid (bxd)transforms the first abdominal segment into metathorax and it presents alsopostbithorax transformation because of its effect on the wild allele of post-bithorax located to its right (Lewis, 1963, 1964).
The mutants in the Ultrabithorax (Ubx) locus are homozygous lethal, showingin heterozygous condition a slight enlargement of the halteres plus a variableand small (1-6) number of wing-type bristles. All of them show mutant pheno-type when in trans with any of the recessive mutants. It has been shown (Lewis,1963; Morata & Garcia-Bellido, in preparation) by mosaic analysis that thephenotype of Ubx homozygous clones is that of the triple combination for thethree recessive mutants, suggesting that mutants in Ubx locus produce an inacti-vation of the whole bithorax system. This interpretation is supported by the factthat the heterozygous deficiency for the whole bithorax system, DfP115/ +shows Ubx phenotype.
Some of the Ubx alleles, like Ubx130, produce very extreme mutant pheno-types in trans with bx, pbx and bxd which are not distinguishable from thoseproduced by the same mutants in trans with their deficiency. Thus bx3/Ubx1S0
has the same extreme phenotype as bx3/DfPU5, pbx/Ubx1™ has the same aspbx\DfP115, etc.,. . . (Morata, 1973). This result suggests that the mutantUbx1™ produces a complete lack of function of the bithorax system equivalentto the physical deficiency.
All of the mutants described so far affect only the haltere or the first abdominalsegment. Moreover the apparent lack of function of the bithorax system ingenetic combinations like Ubx1S0IDfPU5 (which is lethal in individuals butviable in somatic spots) only affects the differentiation of the cells in the haltereand in the first abdominal segment and does not affect the differentiation of cellsof any other segment including the wing (Morata, 1973). This fact suggests that
22 G. MORATA
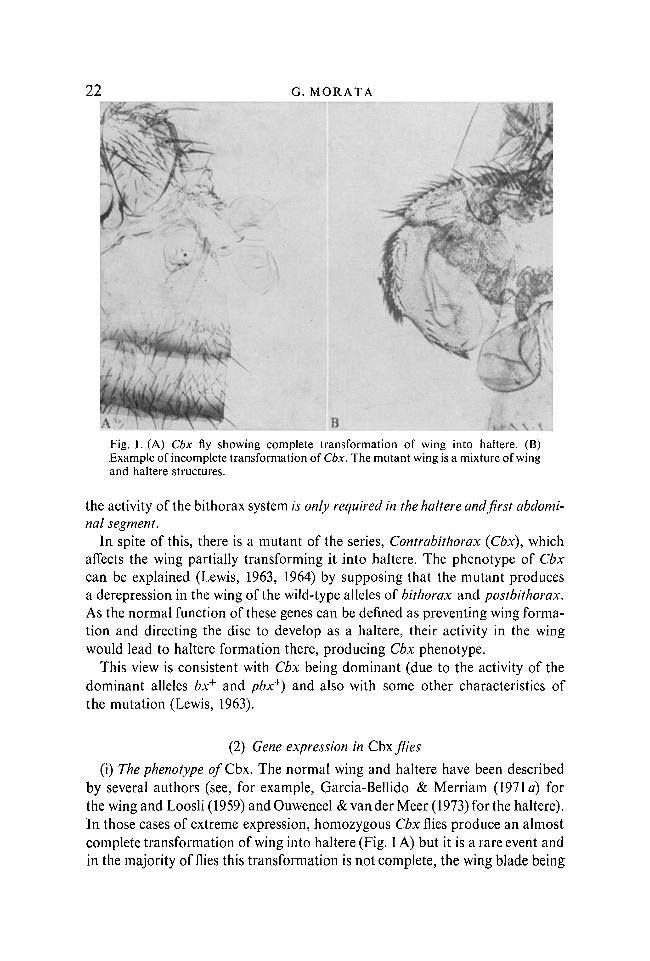
B \Fig. 1. (A) Cbx fly showing complete transformation of wing into haltere. (B)Example of incomplete transformation of Cbx. The mutant wing is a mixture of wingand haltere structures.
the activity of the bithorax system is only required in the haltere and first abdomi-nal segment.
In spite of this, there is a mutant of the series, Contrabithorax (Cbx), whichaffects the wing partially transforming it into haltere. The phenotype of Cbxcan be explained (Lewis, 1963, 1964) by supposing that the mutant producesa derepression in the wing of the wild-type alleles of bithorax and postbithorax.As the normal function of these genes can be defined as preventing wing forma-tion and directing the disc to develop as a haltere, their activity in the wingwould lead to haltere formation there, producing Cbx phenotype.
This view is consistent with Cbx being dominant (due to the activity of thedominant alleles bx+ and pbx+) and also with some other characteristics ofthe mutation (Lewis, 1963).
(2) Gene expression in Cbx flies
(i) The phenotype of Cbx. The normal wing and haltere have been describedby several authors (see, for example, Garcia-Bellido & Merriam (1971 a) forthe wing and Loosli (1959) andOuweneel & van der Meer (1973) for the haltere).In those cases of extreme expression, homozygous Cbx flies produce an almostcomplete transformation of wing into haltere (Fig. 1 A) but it is a rare event andin the majority of flies this transformation is not complete, the wing blade being

Gene expression in Cbx 23
Fig. 2. (A) Mosaic wing showing wing (W) and haltere (H) trichomes. Notice thepresence in the haltere tissue of several small bristles typical of the haltere (arrows).(B) Trichomes of intermediate size between wing and haltere in the mutant wing(arrow). Compare with those of typical wing (above right) and of typical haltere(below). (C) Small group of haltere trichomes in the anterior wing (arrow). (D)Typical wing costa and triple row bristles surrounded by haltere trichomes.
a mosaic of wing and haltere differentiations (Fig. 1 B). Mature Cbx wing discsalso show characteristics of both wing and haltere discs.
In the majority of cases, each cell differentiates wing or haltere structures andintermediates do not appear (Fig. 2 A). However, trichomes can be detectedwhich are an intermediate size between those of wing and haltere (Fig. 2B).Those trichomes might be secreted by 'hybrid' wing-haltere cells.
Usually the mutant wing is composed of large patches of wing and halteretissue as those shown in Fig. 2 A but together with these, other small halterepatches can frequently be detected (Fig. 2C). The minimum size of these smallpatches is about 30 cells.
It is interesting to note that in the case of extreme expression, the only wingstructures seen are wing bristles that are completely surrounded by halteretrichomes (Fig. 2D).
The expression of the mutant genotype is variable in the different regions ofthe mutant wing. Fig. 3 shows the percentage of transformation in the major

24 G. MORATA
Fig. 3. Degree of transformation in different regions of the wing. Percentages werecalculated in a sample of 100 Cbx homozygous wings. NO, Notum; SC, scutellum;CO, costa; TR, triple row; II, III, IV, V, veins.
TR
Fig. 4. Correspondence between different regions of wing and haltere. MS, Mesono-tum; CO,costa (P, proximal; M, medial; D,distal); TR, triple row; MT, metanotum;SCA, scabellum; PE, pedicellum; CA, capitellum.

Gene expression in Cbx 25
Table 1. Homeotic transformations of haltere into wing (i/-> W) and wing intohaltere {W->H) in different genotypes involving Cbx and other mutants of thebithorax system
The symbol + indicates slight transformation and + + more complete transformation.
Phenotype
Genotype H -> W W -»H
Cbx/+ - + +Cbxjbx^ - + +Cbx/pbx — + +Cbx I Ubx1 + +Cbx I Ubx1™ + +CbxjDfPUS + +
regions of the wing. In the posterior part (defined by the line between veins IIIand IV) the expression is 100 % (a complete transformation). In the anteriorregions there is variable degree of transformation ranging from 90 % in C to11 % in A'. The degree of transformation is weaker the more anterior are theregions in the wing blade.
The fact that most of the mutant wings are mosaics of wing and halterestructures allows us to make correlation between the type of haltere cells andtheir location in the wing. This is expressed in Fig. 4 and shows a correspondencebetween homologous regions along the proximo-distal axis so that the proximalhaltere regions (metanotum) appear in the proximal wing regions (notum) theintermediate haltere regions (scabellum and pedicellum) appear in the inter-mediate wing regions (proximal and medial costa) and the distal haltere (capi-tellum) appear in distal wing (distal costa and distal wing blade). This type ofhomologous correspondence has also been described for other homeotic mutants(Gehring, 1966; Gloor & Kobel, 1966; Postlethwait & Schneiderman, 1971) andalso appears in the opposite transformations by bithorax and postbithoraxmutants (Ouweneel, 1973; Morata & Garcia-Bellido, in preparation).
(ii) Possible causes of the partial expression of Cbx. It has been observed byLewis (1963) that Ultrabithorax mutants in trans with Cbx produce a slightbithorax and postbithorax phenotypes which is stronger than that produced bythe same Ubx mutants in trans with a wild-type chromosome. This observationsuggests that there is some hypofunction of bx+ and pbx+ genes of the Cbxchromosome.
In order to confirm this hypothesis flies of different genetic constitutions weresynthesized and their phenotypes were analysed for homeotic transformationboth in wings and halteres. As shown in Table 1, flies of the genotypes Cbx/bx3
and Cbx/pbx have a Cbx phenotype as expected and also show no sign of trans-formation in the halteres. However, flies of the genotypes Cbx/Ubx1, Cbx/Ubx130

26 G. MORATA
Table 2. Number and size o/mwh clones found after X-irradiationat different developmental ages
Age calculated in hours between irradiation and pupariation. W clones embracingonly wing trichomes. H clones embracing only haltere trichomes. W + H clonesembracing both. Average size of clones in parentheses.
Age atirradiation
0-2424^848-7272-9696-120
No. ofwings
5010444
180146
No. ofW clones
48 (40)40(190)7 (68-6)
15(163)5(680)
No. ofH clones
52 (4-3)29 (28-7)2 (400)
21 (127)3 (400)
No. ofW + H clones
06 (52-6)2 (850)4(204)6(630)
and Cbx/DfPlJS presented together with Cbx phenotype a slight but clearbithorax and postbithorax phenotype in the halteres, confirming the earlierobservation of Lewis and inicating that there is some hypofunction of wild-typealleles of bx and pbx when in cis with Cbx.
Thus, the evidence suggests that although Cbx flies have activity of bx+ andpbx+ alleles in the mesothorax, this activity is below normal. Concentrationsof bx+ or pbx+ products that are near to threshold could explain the variedeffect on mesothoracic cells. Thus partial phenotypic expression could reflectpartial genetic activity.
(3) Clonal analysis
The clonal analysis was performed in the mutant wing disc to study the lineagerelationships of the wing and haltere structures, specially to investigate whetherthere are independent cell lineages for each type of structure.
Larvae of the genetic constitution Cbx/mwh were irradiated at different agesand wings of hatched adults were searched for mwh spots. Because mwh ismuch easier to detect in the wing blade, only clones in this region were considered.
The clones found were classified into three groups: (1) forming exclusivelywing structures, (2) forming exclusively haltere structures, (3) forming bothwing and haltere structures.
The number and average cell numbers in clones of the three types which aroseafter irradiation at different developmental ages are shown in Table 2. Clonesembracing wing and haltere structures were found in all ages except in fliesirradiated 0-24 h before pupariation (Fig. 5). This result suggests that there arenot separate cell lineages for wing and haltere during most of development.The lack of this type of clone in the flies irradiated in the latest stage may bedue to the small size (Table 2) of the clones so that the probability of crossinga border between wing and haltere is very low. As shown in Table 2 as theclones are larger the proportion of clones embracing wing and haltereincreases.

Gene expression in Cbx 27
Fig. 5. mwh clones embracing wing and haltere trichomes.
Table 3. Proportion of wing and haltere cells in all mwh clones that werefound to embrace wing and haltere trichomes
Age calculated in hours between irradiation and pupariation. Except for the verysmall clones in which the number of trichomes was counted directly, the size wascalculated according to the area occupied by each type of trichome with correc-tions for different cell densities.
Age atirradiation (h)
24-48
48-72
72-96
96-120
Total no.of cells
402412
10010040
70100
160300100250
300300
100010001000200
No. ofwing cells
3066
602010
525
15015030
100
100280500300200100
No. ofhaltere cells
10186
408030
6575
1015070
150
20020
500700800100
Ratio wing/haltere cells
3/11/31/13/21/41/3
1/131/3
15/11/13/72/3
1/214/11/13/71/41/1

28 G. MORATA
The continuity of all of the clones embracing wing and haltere argues againstthe possibility of two independent events of mitotic recombination since theprobability of conjunction of two independent clones to form a continuouspattern must be very low.
The supposed influence of division rate on homeotic expression has beenemphasized several times (Ouweneel, 1969; see Postlethwait & Schneiderman,1971, for discussion on this point). The results presented here argue againstthis hypothesis since the sizes of clones producing haltere or wing are more orless similar, suggesting that division rate does not influence the final differentia-tion. It also has been shown that the normal haltere and wing had very similar ifnot equal division rate (Morata & Garcia-Bellido, in preparation). Furthermore,the analysis of clones differentiating wing and haltere (Table 3) shows that theproportion of wing and haltere tissue in those clones is very variable, suggestingthat the division rate of the cells within a given clone does not affect thedifferentiation.
DISCUSSION
As shown previously (p. 30) in Cbx flies the homeotic transformation of winginto haltere is likely due to the derepression in the mesothorax of the bithoraxand postbithorax genes which are those responsible for the anterior and posteriortransformation respectively. However, the genetic analysis shows that despitetheir derepression in the mesothorax, the activity of bx+ and pbx+ when in ciswith Cbx is below the level found in wild-type flies. This hypofunction mayexplain the partial expression of Cbx genotype; that is, some cells might haveenough bx+ or pbx+ product to differentiate mutant haltere phenotype whileothers would not and so produce normal wing cells.
In this context it is interesting to note that in many aspects the developmentalpathways leading to wing and haltere formation are similar. Both discs growduring the same larval stages, have similar division rate and have an apparentlyhomologous process of compartmentalization (Morata & Garcia-Bellido, inpreparation; Garcia-Bellido, Ripoll & Morata, 1973). These results suggest thatonly the presence of bx+ and pbx+ gene products is enough to direct develop-ment into haltere while their absence produces wing structures.
Moreover, it has been inferred from mosaic analysis (Morata & Garcia-Bellido,in preparation) that during most of the developmental period of the halteredisc the cells require the function of bithorax and postbithorax genes continuallyso that the removal of the wild-type alleles is expressed directly by a change inthe state of determination of the affected cells. Only when the change is inducedafter 24 h before puparium formation (that is, in the late development of thedisc) does the removal of the wild-type alleles fail to affect the final differentiationof the cells. This inability of the cells to respond to a change in the genetic consti-tution after a given moment has been called 'perdurance' by Garcia-Bellido &Merriam (1971 b). These results are in line with those obtained with other genes

Gene expression in Cbx 29
like hairy and Hairy wing (Hw) (Garcia-Bellido & Merriam, 19716) engrailed(Garcia-Bellido & Santamaria, 1972) and spineless-aristapedia (Postlethwait &Girton, 1974) and suggest that this continuous requirement is a general featureof the homeotic genes. One can speculate that all show perdurance in the latestages because of the persistence of the normal gene products during the remain-ing few divisions, rather than because of the attainment of an irreversible stateof determination. This interpretation is supported by the observation of Garcia-Bellido (unpublished) that the perdurance of bithorax is lost after in vivo culture.
The fact that there is such a continuous dependence by the cells on the state ofactivity of their homeotic genes may help us to understand the development of theCbx wing. Due to the hypofunction of bx+ and pbx+, the mutant cells may havean unstable state of determination that could change from one cell generation toanother depending on the level of activity in each cell of bx+ andpbx+ products.What actually decides the final fate of the cells is the level of these products inthe last divisions.
This instability of the state of determination is reflected by the clonal analysissince clones embracing wing and haltere structures were found in all except thelast stage examined. The fact that these clones were not found in this last stagemay be due either to the small size of the clones at this period (about fourcells/clone, with little probability of crossing a border between haltere and wing)or to the perdurance of bx+ and pbx+ products in the last two-three divisions.If this latter possibility were true then the cells would be finally committed todifferentiate wing or haltere at a very late stage of development, on averagetwo-three divisions before the disc stops growing. In any case it is clear thatduring most of the development of Cbx wing disc the cells remain undeterminedto produce wing or haltere.
In some respects these results are similar to those of Postlethwait & Schneider-man (1969, 1971) in their experiments with Antpn. They were unable to findclones crossing the border between leg and antenna structures after 72 h ofdevelopment, that is the third larval period. As they point out recently (Postle-thwait & Schneiderman, 1974) the mutation Antpn produces an unstable stateof determination in the mutant antennal disc.
Despite this lack of stable determination in the Cbx wing disc some regionsshow a consistently higher degree of expressivity than others (see Fig. 4). Thereasons that may lead to this region-specific expressivity are not fully understood,they may reflect different synthetic rates of bx+ and pbx+ products or differentregional requirements.
Together with these regional differences some wing structures seem to be moreeasily transformed than others. In the cases of extreme expression it is often found(see Fig. 2D) that the only wing structures that can be observed are the bristleswhereas all of the surrounding trichomes are typical of haltere. This differentialbehaviour of bristles versus trichomes in wing-haltere transformation has also

30 G. MORATA
been observed in experiments (Morata & Garcia-Bellido, in preparation) inwhich bxs/bxs clones (marked with y mwhjv) were induced in normal (bxz/ + )halteres. Clones were detected which presented wing bristles surrounded bymarked (and therefore belonging to the same clone) haltere trichomes.
The causes of this difference between bristles and trichomes are not clear.The bristles appear consistently in the margin and it has been shown (Santamaria& Garcia-Bellido, in preparation) that bristle differentiation in the wing marginis dependent on interactions between cells in the dorsal and ventral surface. Itis conceivable that those specific inductive stimuli demand a longer period ofbithorax gene activity in the affected cells (it is known that presumptive bristlecells perform several extra divisions whereas presumptive epidermal cells do not(Lawrence, 1966), and this may uncover the hypofunction of the controllinggenes in some cells. Other cells outside the margin do not receive the stimuli andremain haltere trichomes.
Another characteristic of the Cbx transformation is the proximo-distalcorrespondence between wing and haltere regions (see Fig. 4). This type ofcorrespondence has been observed previously in other homeotic mutants(Gehring, 1966; Gloor & Kobel, 1966; Postlethwait & Schneiderman, 1969,1971) as well as in bithorax mutants (Ouweneel, 1973; Morata & Garcia-Bellido, in preparation) and has been discussed in detail by Postlethwait &Schneiderman (1971). This correspondence probably indicates identical posi-tional information (Wolpert, 1969) along at least the proximo-distal axis inthe haltere and wing discs.
This identity in the positional information in homologous regions of wingand haltere probably produces the same developmental properties, whether thecells will differentiate haltere or wing depending exclusively on the activity ofinactivity of the bithorax genes.
I thank Drs A. Garcia-Bellido, P. Santamaria and P. Ripoll for fruitful discussions. I ammost grateful to Dr P. A. Lawrence for his comments and help during the preparation ofthe manuscript.
REFERENCES
GARCIA-BELLIDO, A. & MERRIAM, J. B. (1971 a). Parameters of the wing imaginal disc develop-ment of Drosophila melanogaster. Devi Biol. 24, 61-87.
GARCIA-BELLIDO, A. & MERRIAM, J. B. (1971/?). Genetic analysis of cell heredity in imaginaldiscs of Drosophila melanogaster. Proc. natn. Acad. Sci. U.S.A. 68, 2222-2226.
GARCIA-BELLIDO, A., RIPOLL, P. & MORATA, G. (1973). Developmental compartmentaliza-tion of the wing disc of Drosophila. Nature New Biology 245, 251-253.
GARCIA-BELLIDO, A. & SANTAMARIA, P. (1972). Developmental analysis of the wing disc inthe mutant engrailed of Drosophila melanogaster. Genetics, Princeton 72, 87-104.
GEHRING, W. (1966). Bildung eines vollstandigen Mittelbeines mit Sternopleura in derAntennenregion bei der Mutante Nasobemia (Ns) von Drosophila melanogaster. Arch.Julius Klaus-Stift VerebForsch. 41, 44-54.
GLOOR, H. & KOBEL, H. (1966). Antennapedia (ssAnt) eine homeotische Mutante bei Droso-phila hydei. Revue suisse Zool. 73, 229-252.

Gene expression in Cbx 31LAWRENCE, P. (1966). The development of hairs and bristles in the milkweed bug Oncopeltus
fasciatus. J. Cell Sci. 1, 475-498.LEWIS, E. B. (1963). Genes and developmental pathways. Am. Zool. 3, 33-56.LEWIS, E. B. (1964). Genetic control and regulation of developmental pathways. In The Role
of Chromosomes in Development, pp. 231-252. New York: Academic Press.LEWIS, E. B. (1967). Genes and gene complexes. In Heritage from Mendel. Madison, Wisconsin:
University of Wisconsin Press.LEWIS, E. B. (1968). Genetic control of developmental pathways in Drosophila melanogaster.
Proc. \2th Int. Conf. Genet. (Abstract), pp. 96-97.LINDSLEY, D. L. & GRELL, E. H. (1968). Genetic variations of Drosophila melanogaster.
Publs Carnegie Instn no. 627.LOOSLI, R. (1959). Vergleich von Entwicklungspotenzen in normalen, transplantierter und
mutierter Halteren-Imaginalscheiben von Drosophila melanogaster. Devi Biol. 1, 24-64.MORATA, G. (1973). Herencia de la determinacion en Drosophila. Ph.D. thesis, University of
Madrid.OUWENEEL, W. J. (1969). Influence of environmental factors on the homeotic effect of
loboid-ophthalmoptera in Drosophila melanogaster. Wilhelm Roux Arch. EntwMech. Org.164, 15-36.
OUWENEEL, W. J. (1973). Replacement pattern of homeotic wing structures in halteres.Drosoph. Inf. Serv. 50, 102.
OUWENEEL, W. J. & VAN DER MEER, J. M. (1973). Differentiation capacities of the dorsalmetathoracic (Haltere) disc of Drosophila melanogaster. Wilhelm Roux Arch. EntwMech.Org. 172, 149-161.
POSTLETHWAIT, J. H. & GIRTON, J. R. (1974). Development in genetic mosaics of aristapedia,a homeotic mutant of Drosophila melanogaster. Genetics, Princeton 76, 767-774.
POSTLETHWAIT, J. H. & SCHNEIDERMAN, H. A. (1969). A clonal analysis of determination inAntennapedia, a homeotic mutant of Drosophila melanogaster. Proc. natn. Acad. Sci. U.S.A.64, 176-183.
POSTLETHWAIT, J. H. & SCHNEIDERMAN, H. A. (1971). Pattern formation and determinationin the antenna of the homeotic mutant Antennapedia oi Drosophila melanogaster. Devi Biol.25, 606-640.
POSTLETHWAIT, J. H. & SCHNEIDERMAN, H. A. (1974). Developmental genetics of Drosophilaimaginal discs. A. Rev. of Genetics 7, 381-433.
WOLPERT, L. (1969). Positional information and the spatial pattern of cellular differentiation./ . theor. Biol. 25, 1-47.
{Received 15 November 1974, revised 10 March 1975)
EMB 34