
Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein
12
1 23 Cellular and Molecular Bioengineering ISSN 1865-5025 Volume 6 Number 1 Cel. Mol. Bioeng. (2013) 6:38-47 DOI 10.1007/s12195-012-0260-9 Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein Transport by Varying Structural and Chemical Properties Karthik Uppulury, Artem K. Efremov, Jonathan W. Driver, D. Kenneth Jamison, Michael R. Diehl & Anatoly B. Kolomeisky
Transcript of Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein

1 23
Cellular and MolecularBioengineering ISSN 1865-5025Volume 6Number 1 Cel. Mol. Bioeng. (2013) 6:38-47DOI 10.1007/s12195-012-0260-9
Analysis of Cooperative Behavior inMultiple Kinesins Motor Protein Transportby Varying Structural and ChemicalProperties
Karthik Uppulury, Artem K. Efremov,Jonathan W. Driver, D. KennethJamison, Michael R. Diehl & AnatolyB. Kolomeisky

1 23
Your article is protected by copyright and
all rights are held exclusively by Biomedical
Engineering Society. This e-offprint is for
personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein
Transport by Varying Structural and Chemical Properties
KARTHIK UPPULURY,2 ARTEM K. EFREMOV,1 JONATHAN W. DRIVER,1 D. KENNETH JAMISON,1
MICHAEL R. DIEHL,1,2 and ANATOLY B. KOLOMEISKY2
1Department of Bioengineering, Rice University, Houston, TX 77005, USA; and 2Department of Chemistry, Rice University,Houston, TX 77005, USA
(Received 31 August 2012; accepted 6 November 2012; published online 21 November 2012)
Associate Editor Jung-Chi Liao & Henry Hess oversaw the review of this article.
Abstract—Intracellular transport is a fundamental biologicalprocess during which cellular materials are driven by enzy-matic molecules called motor proteins. Recent optical trap-ping experiments and theoretical analysis have uncoveredmany features of cargo transport by multiple kinesin motorprotein molecules under applied loads. These studies suggestthat kinesins cooperate negatively under typical transportconditions, although some productive cooperation could beachieved under higher applied loads. However, the micro-scopic origins of this complex behavior are still not wellunderstood. Using a discrete-state stochastic approach weanalyze factors that affect the cooperativity among kinesinmotors during cargo transport. Kinesin cooperation is shownto be largely unaffected by the structural and mechanicalparameters of a multiple motor complex connected to a cargo,but much more sensitive to biochemical parameters affectingmotor–filament affinities. While such behavior suggests thenet negative cooperative responses of kinesins will persistacross a relatively wide range of cargo types, it is also shownthat the rates with which cargo velocities relax in time uponforce perturbations are influenced by structural factors thataffect the free energies of and load distributions within amultiple kinesin complex. The implications of these laterresults on transport phenomena where loads change tempo-rally, as in the case of bidirectional transport, are discussed.
Keywords—Intracellular transport, Kinesin, Cooperativity.
INTRODUCTION
Motor proteins are a class of active enzymatic mole-cules that convert chemical energy of ATP hydrolysisinto a mechanical work while transporting cellularmaterials through highly crowded and viscous envi-ronments within cells.1,9,27 Although the properties of
many microtubule and actin-dependent motors shouldallow them to perform these functions as single mole-cules, there are many examples where organelles, vesi-cles and other sub-cellular commodities are transportedsimultaneously by teams of motors that function col-lectively.4,8,13 In addition, cargos are often outfittedwithdifferent types of motors that either move in oppositedirections or along different types of cytoskeletal fila-ments.15,16,22,24,25 The collective properties of motorproteins are therefore important for understandingmechanisms of intracellular transport because thenumber of motor proteins, their types and relativenumbers of different species might be key factors influ-encing regulation and control of intracellular pro-cesses.2,3,10,11,18,19,23 Although significant advances inuncovering many features of cellular transport phe-nomena have been achieved, precise mechanisms ofcollective action of motor proteins in cells are still notwell explained.
Dynamic properties of multiple motor proteinassemblies have been investigated in a variety ofexperimental2,3,11,18,19,23 and theoretical stud-ies,5,10,12,14,21 many of which seek to characterize howcargo motion changes with variation in motor numberand type. Application of engineered complexes ofmotor protein has also been very productive in deci-phering such responses.2,3,6,7,10,11,18,19,23 For instance,our group has employed precision particle tracking andoptical trapping methods to investigate the dynamicproperties of interacting kinesin molecules that areorganized on DNA scaffolds.11,23 As has been found inrecent and independent studies that employ antibodiesto connect kinesin molecules,28 these experiments con-clude that the functions of complexes containing twokinesins are best characterized as net negative cooper-ative since the probability of cargo motion by only oneload-bearing motor within a complex is generally much
Address correspondence to Anatoly B. Kolomeisky, Department
of Chemistry, Rice University, Houston, TX 77005, USA. Electronic
mail: [email protected], [email protected]
Cellular and Molecular Bioengineering, Vol. 6, No. 1, March 2013 (� 2012) pp. 38–47
DOI: 10.1007/s12195-012-0260-9
1865-5025/13/0300-0038/0 � 2012 Biomedical Engineering Society
38
Author's personal copy

higher than the probability that both motors will beengaged in transport and cooperate productively. It hasalso been argued that the weak enhancement of cellulartransport in this case is due to high efficiency of singlekinesin molecules.11,14,23
The above studies of structurally-organized motorcomplexes have also stimulated significant develop-ments in theoretical analysis of collective behavior ofmotor proteins.2,3,10–12,18,19 One of the most successfultheoretical approaches is based on a discrete-state sto-chastic model that captures key elements of single-molecule kinesin dynamics as well as dynamic behaviorof two-kinesin assemblies.5,10,12 This method is uniquesince it (i) explicitly takes into account individualmicroscopic transitions between single-motor-boundconfigurations and a spectrum of two-motor boundconfigurations with different distances between mole-cules along filaments; (ii) parameterizes transition ratesfrom fits obtained for single-kinesin optical trap exper-iments; and (iii) calculates numerically exactly all spa-tial–temporal dynamic properties of multiple-motorsystem. This approach reproduces all features observedin experiments on two-kinesin assemblies, and it alsoprovides microscopic explanations for observeddynamic behavior. For example, this model suggeststhat collections of kinesins face mechanical and kineticchallenges that restrict their ability to adopt filament-bound configurations that support load-sharingbehaviors. Furthermore, discrete state modeling hasalso demonstrated that this behavior is only exacerbatedwhen applied loads vary spatially and temporally.10 Yet,such behaviors naturally raise fundamental questions asto whether such dynamics is general for all motor pro-teins or it is unique for structurally organized kine-sin complexes utilized in our experiments. Herein,we apply the discrete-state stochastic modelingapproach to examine the extent to which multiple
kinesin behaviors change with varying structural andchemical properties of a multiple motor system.
METHODS
Our analysis of motor protein cooperativity is basedon a discrete-state stochastic approach that has beendeveloped to explain optical trapping measurementsobtained for two-kinesin assemblies acting on cargosagainst applied external loads.6,7,26 A generic view ofthe system of two kinesins that move cargo is shown inFig. 1(Left). It is assumed that two kinesins are boundat a distance Sbead apart from each other on the surfaceof a spherical cargo with a diameter D. The motors canattach and detach to and from different microtubulelattice sites that are separated by 8.2 nm, and they areassumed to move only along a single protofilament forsimplicity. The model enumerates for a broad range oftransitions where only one motor protein molecule isconnected to microtubules as well as a wide spectrum oftwo-motor-bound configurations where the separationdistance between binding sites on microtubules,Smicrotubule, varies.
Transition Rate Modeling
The main idea of the discrete state stochasticapproach is to calculate the rates a multiple motorcomplex transitions between different filament-boundconfigurations via explicit calculations of the freeenergy of the system in each state. It is done byassuming that between transitions the system quicklyrelaxes to mechanical equilibrium, although chemicalequilibrium is not assumed.6,7,26 The forces acting onthe cargo and the motors are estimated from the force-balance assumption and parameterizing the stiffness
FIGURE 1. (Left) A schematic representation of the model of cargo transport by two coupled kinesin motor proteins along themicrotubule. The cargo diameter is D, the separation distance between the motors on the cargo surface is Sbead and that on themicrotubules is Smicrotubule. (Right) Plots of motor stiffness (jM) that show the strain-induced stiffening of a single kinesin-bead linkageunder load. The legend indicates the force at which motor stiffness reaches half of its maximal value (f1/2). The red curve(f1/2 = 3.2 pN) approximates the stiffness function of the kinesin motors examined experimentally in Rogers et al.,23 Jamison et al.11,10
Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein Transport 39
Author's personal copy

profile of the kinesin motor from experiments. The freeenergy of the complex, which is called a configura-tional energy (Econfig), can then be calculated explicitlyusing the following expression:
Econfig ¼1
2kTðxT � xbÞ2 þ
X
M
Zlax
l0
jj~Faxjjdl ð1Þ
where,jT,xT,xb andFax are the trap stiffness, positionofthe trap, position of the cargo bead center and the axialload experiencedby thebead respectively.Theparameterl0 denotes the length of the kinesin motor under zeroforce,while the parameter lax is the extended lengthof themotor protein. This equation reflects two contributionsinto the free energy of the state, the potential energy ofthebead in the trap and themechanical energy associatedwith stretching the motor protein—see Driver et al.6 formore detailed explanations.
The way the system of coupled motor proteinsevolves with a time is given by a set of master equa-tions that are numerically solved as explained in detailin our previous works.6,7,26 The calculated free energiesare utilized for estimating transition rates via thedetailed balance conditions. Solving master equationsprovides probability distributions of different states ofthe system at various times, which can be used toestimate average observable quantities such as averagedetachment rates, detachment forces and cargo veloc-ities that, in turn, can be used to assess whether themotors within a complex can cooperate productively.
Model Parameterization and Calculating Observables
Transition rates are input parameters for solvingmaster equations, and they are obtained in the fol-lowing way. Detachment and binding rates in theabsence of external forces are estimated from experi-mental analyses of single kinesin behaviors exclu-sively,7,26 and the rates under loads are determinedusing the detailed balance condition as explained inDriver et al.6 Stepping rates of the motor under appliedloads are determined from fits to single-kinesin opticaltrapping data: see Driver et al.6 for more details. Toproperly describe mechanical properties of the system,the elasticity of motor–cargo links is estimated byusing fits to measurements of single-kinesin stiffness-es.7,11 These results show the non-linear elasticity ofthe system which allows us to take into account theeffects of strain-induced stiffening of connectionsbetween motor proteins and cargo.
The present analyses examine how collective kinesinbehaviors change with structural and mechanicalproperties of a multiple motor complex as well asbiochemical properties of its constituent motors. Our
prior experiments utilized a DNA scaffold that sepa-rated the motors by 50 nm on surface of 500 nm sizedbeads.10,11 For the present study, the influence of amotor complex’s structure was examined by alteringthe distance between motor protein connections on thebead Sbead and cargo size D. The effect of mechanicalproperties was also explored by varying elastic prop-erties of motor protein complexes. Again, experimentalmeasurements of force-dependent elasticity of singlekinesins6,11 indicate that the motor protein stiffness hasa non-linear dependence due to strain-induced stiff-ening of the motors (Fig. 1). We have shown that theseresponses can be approximated well by a sigmoidfunction.6,11 To simplify the present analyses, ouroriginal function was modified by altering the force atwhich motor stiffness increase with increasing load. Tolabel different load-dependent stiffness curves weintroduce a quantity f1/2, which corresponds to theforce at which motor stiffness (jM) reaches half of itsmaximal value. Accordingly, high f1/2 corresponds tomore compliant motors, while smaller values describerigid motor proteins. Experimental measurements ofelasticity for kinesins give f1/2 = 3.2 pN nm�1.11 Thenfor each elasticity behavior specified by different curves(Fig. 1Right) dynamic properties of the system areevaluated directly and compared with each other toquantify the effect of motor protein stiffness oncooperativity.
The effect of chemical interactions between kinesinsand microtubules can be taken into account by modi-fying detachment forces and detachment rates. Sucheffects can be explored by modulating the unloadedrate for single motor detachment (e). We also defineand modulate kinesin’s critical detachment force (Fd),which is the characteristic force that exponentiallydecreases the rate of motor protein dissociation fromthe microtubule. The critical detachment force is alsoassociated with the length scale Ld, viaFd = kT/Ld,that describes the distance needed to move the motorprotein molecule away from the microtubule to beconsidered detached. The structurally-defined kinesincomplexes described in Driver et al.6 can be modeledusing the experimentally determined values ofe = 0.312 s�1 and Fd = 3.1 pN. For simplicity, weassume that motor detachment follows Kramer’s-typedependence, although experimental data are betterdescribed by a two-state model.6 We utilize thisapproach to simplify all computations, since experi-mentally observed trends are not affected by thisapproximation and only some small quantitative fea-tures are changed. It is important to note that, incontrast to some treatments, we have shown that thisapproximation can still lead to average detachmentrates for multi-motor complexes that are not expo-nential functions of forces. Thus, we predict multiple
UPPULURY et al.40
Author's personal copy

motor behaviors that exhibit apparent deviations fromBell-model predictions without the need to assume thatmotors exhibit complex, force-dependent detachmentcharacteristics such as slip–catch behaviors.6
To quantify the influence of all these structural,mechanical and chemical properties, we concentrate onseveral observable quantities that are measured inexperiments and that can be easily calculated in thediscrete-state approach. Specifically, in this work, wecalculate the free energy difference, DEconfig, betweensingle-bound and a range of two-motor states sincemotor protein molecules can only cooperate produc-tively when they are bound close together to themicrotubule. Similarly, one could see the effect ofcooperativity by monitoring the ratio of averagedetachment forces for two-motor bound and single-motor bound cargo complexes, F2h i= F1h i. A dynamicview of cooperativity can also be obtained from anal-ysis of the ratio of average velocities for two-motorand single-motor complexes, V2h i= V1h i.
Recent force clamp assays have shown that multiplekinesin complexes may experience loading conditionsthat prevent them from reaching a steady state; forexample, when loads vary spatially or temporally.6
Consequently, understanding mechanisms of motorcooperation also requires analyses of the relaxationbehavior of motor protein complexes under variableloading conditions. Generally, if the applied load ischanged instantaneously from a low to high load, mul-tiple kinesin velocities will also change in time as thesystem progresses towards a new steady state since theprobability that a complexwill achieve productive, load-sharing states increases with increasing load. The char-acteristic relaxation time describing this change can beapproximated using the following exponential function:
Vav tð Þ ¼ A� B exp � t
srelax
� �ð2Þ
In this expression, the parameter A corresponds to astationary-state velocity, while the parameter B givesthe deviation from the stationary-state velocity at
initial times. The values of A, B and srelax are extractedfrom fits to calculated curves.
Relaxation times were calculated for a circumstancewhere the applied load increased instantaneously from4 to 5 pN, and then remained static at 5 pN until thesystem reached its steady state. This condition mimicsthose provided in our prior force clamping assays.6 Acomplete set of parameters used in our calculations ispresented in Table 1.
RESULTS AND DISCUSSIONS
As discussed in Driver et al.11, the distribution of anapplied load between two elastically-coupled kinesinswhen both motors are attached to a microtubule isstrongly dependent on how far apart the motors arebound at the filament. Due to geometric constraints,equitable load sharing only occurs when the motors arebound closely on the filament, and hence, they willonly cooperate productively under an applied load ifthey can transition into these states. To gauge howstructural and mechanical properties of a complexinfluence this ability, we first examined dependence ofconfigurational energy changes of a two-kinesin com-plex on cargo diameter (D), the distance between thesites that the motors are anchored to the cargo surface(Sbead), and the mechanical compliances of motor–cargo linkages (via the parameter f1/2). In each case,configurational energies were calculated for transitionsfrom single-motor bound states to a range of two-motor bound configurations. Generally, the lower thisenergy difference the higher the degree of cooperativitysince the system will transition more frequently intotwo-motor bound states where the motors will sharetheir load (see Figs. 2, 3), and since transition proba-bilities will be controlled by the corresponding Boltz-mann’s factor: exp(�DEconfig/kBT).
The dependence of DEconfig for different sized cargosas a function of the separation distance between themotors on the microtubule are presented for applied
TABLE 1. Model parameters and their estimated values.
Parameter Estimated/measured values Reference
p0 unloaded binding rate 4.7 s�1 Estimated from Leduc et al.17
e0 unload detachment 0.312 s�1 Estimated from Driver et al.6
u0þ, unloaded stepping rate 1.59 9 1014 s�1 Estimated from Driver et al.6
u0þþ unloaded stepping rate 61.7 s�1 Estimated from Driver et al.6
w0�, unloaded stepping rate 0.654 s�1 Estimated from Driver et al.6
w0�� unloaded stepping rate 1.69 9 109 s�1 Estimated from Driver et al.6
Fd, detachment force 3.1 pN
D, cargo diameter 300–700 nm See text for details
Sbead separation distance 30–70 nm See text for details
f1/2 stiffness at half-maximum 1.2–5.2 pN See text for details
Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein Transport 41
Author's personal copy

FIGURE 2. Difference in configurational energies between two-motor and single-motor states as a function of structural andchemical parameters. (a) DEconfig for varying cargo diameter (D) at 5 pN applied load. (b) DEconfig for varying separation distances(Sbead) at 5 pN. (c) DEconfig for varying motor stiffness at 5 pN load. (d) DEconfig for varying cargo diameter (D) at a 12 pN load.(e) DEconfig for varying separation distances (Sbead) at a 12 pN load. (f) DEconfig for varying motor stiffness at a 12 pN load.
FIGURE 3. Analyses of increases of cargo detachment forces due to motor cooperation. The percentage increaseF2h i= F1h i � 1ð Þ � 100% is computed in the panels a, b and c correspond to the cases of varying cargo diameter (D), separation
distance (Sbead) and motor stiffness (jM). F2h i is the average detachment force compiled from the detachment force distribution of atwo-motor cargo complex while F1h i corresponds to that of a single-motor molecule.
UPPULURY et al.42
Author's personal copy

loads of 5 pN in Fig. 2a, and for 12 pN in Fig. 2d. Theresults of each calculation shows there is a relativelylarge range of intermediate Smicrotubule values whereDEconfig is negligible since transitions into these con-figurations do not change the force balance betweenthe motors in a complex. Outside of this region,DEconfig changes more rapidly with Smicrotubule com-pared to the cargo size D. Although the effects are verysmall, the energy needed to transition into configura-tions where the motors are closely spaced on themicrotubule is found to increase with the increasingcargo size (Figs. 2a, 2d). Moreover, DEconfig is found tobe sensitive to cargo size for transitions into stateswhere the motors are positioned far apart on thefilament (Smicrotubule > 140 nm) and will experienceadditional ‘counter forces’ due to the resultant lead-ing–lagging-motor organization of the complex inthese configurations.11 Nevertheless, given the magni-tude of these differences, we generally expect thatcargo size will not affect the probabilities the motorswill share their loads appreciably.
While also small compared to the dependence ofDEconfig on Smicrotubule, somewhat stronger dependencesof DEconfig are predicted when the separation distancebetween the motors on the bead (Sbead) is increased(Fig. 2b—for 5 pN; and Fig. 2e—for 12 pN). Thesetrends can be understood from the system geometry(Fig. 1Left). When Smicrotubule< 100 nm, increasing thecargo diameterD or Sbead makes it harder for the secondmotor to bind to the microtubule because the motorsmust now stretch a larger distance to reach their corre-sponding lattice sites, which is more energetically costly.It is also important to note that the free energy of thesystem could be further affected by bead rotations anddisplacements against the applied load, as discussed inDriver et al.6 Note also that for Smicrotubule> 100 nm,DEconfig decreases with increasing Sbead, in contrast to thedependence on cargo diameter D. Although neitherstructural parameter is shown to influence DEconfig sig-nificantly compared to the on-filament motor spacing,this distinction occurs since the vectorial properties of theloads experienced by each motor (i.e., the split betweentangential and upward loads on a motor) will changedifferently when cargo size and motor spacing arevaried—due to bead curvature.
The elasticity of motor proteins in a motor-beadlinkage is predicted to have the strongest effect on thefree energy difference between two-motor and single-motor bound states (Figs. 2c, 2f). This response can beexplained as follows. Under the same external load (F),the cargo displacement is given by Dx = F/jM, whichleads to the free-energy contribution of ½ jMDx2 = 1/2 F2/jM. This result states that increasing thestiffness lowers the contribution to free energy. Thissimple argument therefore predicts that more rigid
motors should cooperate better since they do notincrease Econfig as much as do the more compliantmotors. These arguments work well for Smicrotubule <
100 nm. However, the trend reverses for larger sepa-ration distances due to the introduction of counterforces which increase load per each motor molecule(see Fig. S3 in Driver et al.6). Yet, we expect thedependence of Econfig on jM to mimic the trend foundwith Sbead, as opposed to the trend for cargo diameterD, since compliant motors can stretch more easily. Amore flexible complex will therefore behave effectivelyas motors that are spaced further apart on the beadwhen they adopt configurations where Smicrotubule islarge.
A different way of analyzing cooperativity of mul-tiple motor proteins is to compare average detachmentforces of two-motor complexes F2h ið Þ and single-motormolecules F1h ið Þ. These forces can be determined fromdetachment distributions obtained in the static trapmode of experiments.11 If two-motor assembliescooperate positively via load sharing, it will take muchmore energy to remove them from the microtubules.But if the cooperation is weak the detachment forceF2h i for two motor protein molecules should not besignificantly larger than the detachment force F1h i forthe single-motor case. It is important to note that weare considering here the detachment force rather thanthe stalling force which only describes the condition ofno motion for the cargo without providing microscopicdetails on mechanisms of cooperativity.
The percentage increase F2h i= F1h i � 1ð Þ � 100% as afunction of the cargo size, geometry and motor stiff-ness is shown in Fig. 3 and are plotted as a function ofthe critical detachment forces Fd, which is a force thatcharacterizes the sensitivity of motor detachment tostrain.6,7 Increasing the cargo diameter lowers the ratioF2h i= F1h i as expected (Fig. 3a) since in this case itbecomes easier energetically to detach two-motorcomplexes, leading to lower cooperativity. Similareffect is observed for increasing the distance betweenbound motors (Fig. 3b), although the effect is smaller.In addition, more rigid motors also increase thecooperativity with respect to force production(Fig. 3c); in agreement with our free-energy argu-ments. Nevertheless, one can clearly see that theF2h i= F1h iratio for all ranges of reasonable parameters isonly slightly larger than 1, indicating weak cooperativegains and a generic insensitivity to a multiple motorsystem’s structural and mechanical properties. Thelargest gains and dependencies on the structural andmechanical properties of the complex are observedwhen Fd is assumed to be very large (6–7 pN) whichmost probably are not realistic for typical cellularconditions. This is due to the fact that larger Fd
corresponds to stronger interactions between motor
Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein Transport 43
Author's personal copy
proteins and microtubules, and it leads to longer life-times of two-motor complexes under load, yieldinghigher probability for cooperative interactions withload sharing.
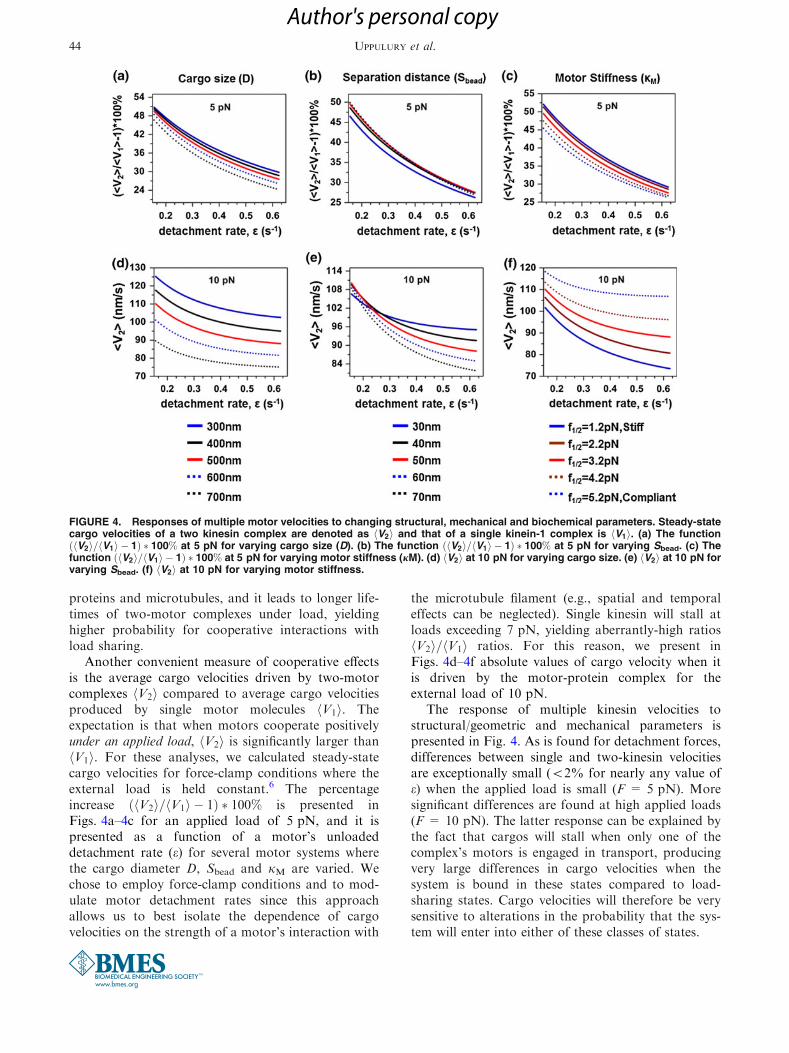
Another convenient measure of cooperative effectsis the average cargo velocities driven by two-motorcomplexes V2h i compared to average cargo velocitiesproduced by single motor molecules V1h i. Theexpectation is that when motors cooperate positivelyunder an applied load, V2h i is significantly larger thanV1h i. For these analyses, we calculated steady-statecargo velocities for force-clamp conditions where theexternal load is held constant.6 The percentageincrease V2h i= V1h i � 1ð Þ � 100% is presented inFigs. 4a–4c for an applied load of 5 pN, and it ispresented as a function of a motor’s unloadeddetachment rate (e) for several motor systems wherethe cargo diameter D, Sbead and jM are varied. Wechose to employ force-clamp conditions and to mod-ulate motor detachment rates since this approachallows us to best isolate the dependence of cargovelocities on the strength of a motor’s interaction with
the microtubule filament (e.g., spatial and temporaleffects can be neglected). Single kinesin will stall atloads exceeding 7 pN, yielding aberrantly-high ratiosV2h i= V1h i ratios. For this reason, we present inFigs. 4d–4f absolute values of cargo velocity when itis driven by the motor-protein complex for theexternal load of 10 pN.
The response of multiple kinesin velocities tostructural/geometric and mechanical parameters ispresented in Fig. 4. As is found for detachment forces,differences between single and two-kinesin velocitiesare exceptionally small (<2% for nearly any value ofe) when the applied load is small (F = 5 pN). Moresignificant differences are found at high applied loads(F = 10 pN). The latter response can be explained bythe fact that cargos will stall when only one of thecomplex’s motors is engaged in transport, producingvery large differences in cargo velocities when thesystem is bound in these states compared to load-sharing states. Cargo velocities will therefore be verysensitive to alterations in the probability that the sys-tem will enter into either of these classes of states.
FIGURE 4. Responses of multiple motor velocities to changing structural, mechanical and biochemical parameters. Steady-statecargo velocities of a two kinesin complex are denoted as V2h i and that of a single kinein-1 complex is V1h i. (a) The function
V2h i= V1h i � 1ð Þ � 100% at 5 pN for varying cargo size (D). (b) The function V2h i= V1h i � 1ð Þ � 100% at 5 pN for varying Sbead. (c) Thefunction V2h i= V1h i � 1ð Þ � 100% at 5 pN for varying motor stiffness (jM). (d) V2h i at 10 pN for varying cargo size. (e) V2h i at 10 pN forvarying Sbead. (f) V2h i at 10 pN for varying motor stiffness.
UPPULURY et al.44
Author's personal copy

The dependence of multiple kinesin velocities on thestructural and mechanical properties of a complex gen-erally follows the trends found with cargo detachmentforces; again, these changes are negligible when appliedloads are small (Figs. 4a–4c). The only exception isfoundwithmotor stiffness. Increasedmotor compliance(increasing f1/2) produces lower cargo velocities whenapplied loads are below kinesin’s stalling force (Fig. 4c).However, this trend is reversed at large loads (Fig. 4f).Webelieve that this behavior stems from the fact that thecompliant motors will stretch very large distances underthese loads. Such behavior therefore increases the rangeof motor bound configurations where the motors willshare their load equitably; hence, allowing themotors tostep more rapidly compared to those in a rigid complexthat is bound in similar configurations (this effect isillustrated in Fig. S3 in Jamison et al.11). This responsetherefore illustrates the importance of analyzing thevectorial properties of load distributionswithinmultiplemotor complexes when assessing cooperative effects.Nevertheless, it is also important to recognize that thelow average detachment forces produced by compliantmotor complexeswill reduce the probability that a cargowill be transported to such loads. Thus, despite the trendfound in Fig. 4f, the weak cooperative behaviors foundin those analyses will likely dominate multiple kinesinbehaviors.
As was found with the dependence of multiplemotor detachment forces on Fd, our calculations showthat motor cooperativity is generally much more sen-sitive to biochemical factors affecting motor affinitythan the structural and mechanical properties of thecomplexes, especially at small applied loads. Forexample, significant changes to two-motor velocitiesare predicted when the unloaded detachment rate of a
single motor molecule (e) is varied (Fig. 4). This can beunderstood in the following way. Decreasing thedetachment rate increases the time that motors spendbound to microtubules; thus, increasing the probabilitya complex will generate high-velocity and cooperativeload-sharing states. At the same time, cooperativity isreduced for large detachment rates since the motorswill detach before these states are reached. Given thestrong sensitivity of multiple kinesin velocities to e, weconclude that factors influencing how tightly the basalaffinity of motors to their filaments as well as thesensitivity of motor–microtubule interactions to loadwill influence multiple kinesin behaviors more signifi-cantly than the structural and mechanical properties ofthe complexes, which primarily determine how load isdistributed within a complex.
Finally, we also examined how the multiple kinesinvelocities change in time by examining how fast cargovelocities relax to a steady-state velocity after aninstantaneous change in the applied load. Althoughneglected in many multiple motor analyses,understanding such behavior is important since theserelaxation times will influence whether a multiple motorsystem will achieve a steady-state during transportscenarios where applied loads vary temporally. Relax-ation occurs since the distributions of multiple motor-bound configurations depend on the applied load on acargo. Relaxation rates/times will naturally depend onhow complexes transition between different configura-tions in time via motor binding, detachment and step-ping. To analyze this behavior, we calculated relaxationtimes by starting computations of the two-kinesin sys-tem at applied load Fap,0 and changing it to anotherapplied load Fap,1. The data presented in Fig. 5 corre-sponds to case where the load was jumped from 4 to
FIGURE 5. The relaxation time (srelax) for different structural, mechanical and chemical properties. (a) srelax at 5 pN applied load forvarying cargo sizes (d). (b) srelax at 5 pN for varying separation distances (Sbead). (c) srelax at 5 pN for varying motor stiffness jM.
Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein Transport 45
Author's personal copy

5 pN, but as we checked to confirm that qualitativeresults are not strongly dependent on the magnitude ofthese changes in load. Relaxation times were deter-mined using the procedures described in the ‘‘Methods’’section.
The dependence of relaxation times on differentstructural properties of the system is shown in Fig. 5.Interestingly, in contrast to multiple kinesin detach-ment forces and velocities, relaxation times are foundto exhibit strong dependencies on the structural andmechanical properties of a cargo. While the evolutionof motor-complexes bound geometry is a complexprocess involving motor binding detachment andstepping, we believe that there are some general fea-tures of relaxation dynamics that can be understood byanalyzing the configuration-dependent free energies ofthe complexes. For example, relaxation times decreasefor larger cargo sizes (Fig. 5a). The analyses of DEconfig
in Fig. 2a show that the range of states where theenergetic costs associated with transitions from single-to two-motor bound configurations are negligible alsodecreases as cargo diameter (D) increases. Conse-quently, the number of states occupied by a motorcomplex on a large cargo will likely decrease withincreasing the cargo size, yielding smaller relaxationtimes. Multiple kinesin velocities also relax faster whenSbead is small (Fig. 5b) and when their linkages to thebeads are stiff (Fig. 5c). These responses may seem tocontradict the trend found with cargo size. However,closer examination show of the states at large motorseparation distances (Figs. 2b, 2c) as well as load dis-tributions (Fig. S3 in Jamison et al.11) uncovers similareffects. In these cases, the range of ‘energetically neu-tral’ states decreases as Sbead and motor compliancesincrease. Furthermore, analyses of force distributionsin Jamison et al.11 show that the difference between theportion of the load assumed by a primary and sec-ondary load-bearing motor will generally decrease withincreasing motor compliance. Relaxation times for acompliant multiple motor complexes will therefore beslow since their motors will step at similar rates, whichwill reduce the rate that the average motor separationdistance changes in time. Overall, these results showthat multiple kinesin relaxation phenomena will bevery sensitive to structural and mechanical parametersaffecting a complex’s configuration-dependent freeenergies and how loads are distributed between itsmotors.
CONCLUSIONS
The cooperative behaviors of coupled kinesin motorproteins were examined via the discrete-state stochasticapproach. The ability of this method to account for the
majority of the relevant states occupied by a complexas well as the transitions between them, and its successin reproducing key experimental observations foundwith experimental analyses of structurally-definedkinesin complexes, provided a basis to explore howkinesin cooperation is influenced by the structural,mechanical and biochemical properties of a motorcomplex. Overall, the present results suggest thatmultiple kinesin force production and velocities will berelatively insensitive to the organization of motors oncargos, their stiffness and cargo size. Responses tothese factors are found, yet they are generally smalland do not influence the abilities of multiple kinesins tocooperate productively as a team. These predictionssuggest that the net negative cooperative behaviorsfound in previous multiple kinesin studies are likelyrobust, and will apply to various transport scenarioswhere multiple kinesins are organized differently ondifferent types of cargos. Yet, it is important to notethat this may not be the case for other motor proteins.For example, recent experimental and theoreticalstudies on myosins V indicate that dynamic behaviorof multiple myosins V depend stronger on elasticityand cargo sizes. Then collective dynamics of myosins Vcould be tuned by other cellular processes more pre-cisely.10,20 It will be interesting to test these ideasdirectly by studying other motor protein systems viaadvanced theoretical and experimental methods.
In contrast to structural and mechanical parameters,biochemical factors affectingmotor-filament affinities arefound to influence, appreciably, the extent to whichkinesin systems will adopt state where the motors canshare the applied load on a cargo. One main reason forthis distinction stems from the fact that multiple kinesincomplexes generallypossess a broad rangeof stateswhereonlyonemotor in the complexbears the applied load, andhence the differences in the free energy between thesestates and single-motor bound states are negligible.Structural factors that alter load distributions within acomplex tend not to affect the energies of these configu-rations. However, biochemical factors such as the un-loaded free energy of kineins–microtubule associationalter the energiesof allmotor-bound states. Furthermore,these factors affect the timescale that motors remain fil-ament-bound, and hence, influence whether a complexhas the opportunity to evolve its bound geometry from asingle-motor-bound state to productive load-sharingstates.
In contrast to cargo detachment forces and velocities,the relaxation dynamics is found to be very sensitive tostructural, mechanical and chemical parameters. Thus,while dependent on a number of factors, these factors caninfluence whether amultiple kinesin systemwill achieve asteady-state in circumstance where loads changetemporally, as will be the case during bidirectional
UPPULURY et al.46
Author's personal copy

transport. If forces acting on motor proteins changerapidly, some forms of motors complexes might not havetime to adopt their geometries to steady-state conditions,giving rise to hysteresis and other complicated effects thatmight confound interpretations. In other words, devia-tions from stationary-state behavior of motor proteinscould significantly modify cooperativemotors responses,and one should be careful in assessing competitionsbetween motors and cannot necessarily assume thatstrong or weak motors would win a tug of war.
ACKNOWLEDGMENTS
This work was supported by grants from theNational Science Foundation (MCB-0643832), theNational Institute of Health (1R01GM094489-01) andthe Welch Foundation (C-1559 to A.B.K. and C-1625to M.R.D.).
REFERENCES
1Alberts, B., et al. Molecular Biology of the Cell (4th ed.).USA: Taylor & Francis, 2002.2Ali, Y. M., G. G. Kennedy, D. Safer, K. M. Trybus, L. H.Sweeney, and D. M. Warshaw. Myosin Va and myosin VIcoordinate their steps while engaged in an in vitro tug ofwar during cargo transport. Proc. Natl. Acad. Sci. U.S.A.108:E535–E541, 2011.3Ali, Y. M., H. Lu, C. S. Bookwalter, D. M. Warshaw, andK. M. Trybus. Myosin V and Kinesin act as tethers toenhance each others’ processivity. Proc. Natl. Acad. Sci.U.S.A. 105:4691–4696, 2008.4Ally, S., A. G. Larson, K. Barlan, S. E. Rice, and V. I. J.Gelfand. Opposite-polarity motors activate one another totrigger cargo transport in live cells. Cell Biol. 187:1071–1082, 2009.5Campas, O., Y. Kafri, K. B. Zeldovich, J. Casademunt,and J.-F. Joanny. Collective dynamics of interactingmolecular motors. Phys. Rev. Lett. 97:038101, 2006.6Driver, J. W., K. D. Jamison, K. Uppulury, A. R. Rogers,A. B. Kolomeisky, andM. R. Diehl. Productive cooperationamong processive motors depends inversely on their mecha-nochemical efficiency. Biophys. J . 101:386–395, 2011.7Driver, J. W., A. R. Rogers, K. D. Jamison, R. K. Das,A. B. Kolomeisky, and M. R. Diehl. Coupling betweenmotor proteins determines dynamic behaviors of motorprotein assemblies. Phys. Chem. Chem. Phys. 12:10398–10405, 2010.8Holzbaur, E. L. F., and Y. E. Goldman. Coordination ofmolecular motors: from in vitro assays to intracellulardynamics. Curr. Opin. Cell Biol. 22:4–13, 2010.9Howard, J. Mechanics of motor proteins and the cyto-skeleton. Sunderland, MA: Sinauer Associates, 2001.
10Jamison, K. D., J. W. Driver, and M. R. J. Diehl. Coop-erative responses of multiple kinesins to variable and con-stant loads. Biol. Chem. 287:3357–3365, 2012.
11Jamison, K. D., J. W. Driver, A. R. Rogers, P. E.Constantinou, and M. R. Diehl. Two kinesins transport
cargo primarily via the action of one motor: implicationsfor intracellular transport. Biophys. J . 99:2967–2977, 2010.
12Klumpp, S., and R. Lipowsky. Cooperative cargo trans-port by several molecular motors. Proc. Natl. Acad. Sci.U.S.A. 102:17284–17289, 2005.
13Kulic, I. M., A. E. X. Brown, H. Kim, C. Kural, B. Blehm,P. R. Selvin, P. C. Nelson, and V. I. Gelfand. The role ofmicrotubule movement in bidirectional organelle transport.Proc. Natl. Acad. Sci. U.S.A. 105:10011–10016, 2008.
14Kunwar, A., and A. Mogilner. Robust transport by mul-tiple motors with nonlinear force-velocity relations andstochastic load sharing. Phys. Biol. 7:016012, 2010.
15Kural, C., H. Kim, S. Syed, G. Goshima, V. I. Gelfand,and P. R. Selvin. Kinesin and dynein move a peroxisomein vivo: a tug-of-war or coordinated movement? Science308:1469–1472, 2005.
16Laib, J. A., J. A. Marin, R. A. Bloodgood, and W. H.Guilford. The reciprocal coordination and mechanics ofmolecular motors in living cells. Proc. Natl. Acad. Sci.U.S.A. 106:3190–3195, 2009.
17Leduc,C.,O.Campas,K.B.Zeldovich,A.Roux,P. Jolimaitre,L. Bourel-Bonnet, B. Goud, J.-F. Joanny, P. Bassereau, andJ. Prost. Cooperative extraction of membrane nanotubes bymolecular motors. Proc. Natl. Acad. Sci. U.S.A. 101:17096–17101, 2004.
18Leduc, C., N. Pavin, F. Julicher, and S. Diez. Collectivebehavior of antagonistically acting kinesin-1 motors. Phys.Rev. Lett. 105:128103, 2010.
19Leduc, C., F. Ruhnow, J. Howard, and S. Diez. Detectionof fractional steps in cargo movement by the collectiveoperation of kinesin-1 motors. Proc. Natl. Acad. Sci.U.S.A. 104:10847–10852, 2007.
20Lu, H., A. K. Efremov, C. S. Bookwalter, E. B. Kre-mentsova, J. W. Driver, K. M. Trybus, and M. R. Diehl.Collective dynamics of elastically coupled myosin Vmotors. J. Biol. Chem. 287(33):27753–27761, 2012.
21Muller, M. J. I., S. Klumpp, and R. Lipowsky. Tug-of-waras a cooperative mechanism for bidirectional cargo trans-port by molecular motors. Proc. Natl. Acad. Sci. U.S.A.105:4609–4614, 2008.
22Ou, G. S., O. E. Blacque, J. J. Snow, M. R. Leroux, andJ. M. Scholey. Functional coordination of intraflagellartransport motors. Nature 436:583–587, 2005.
23Rogers, A. R., J. W. Driver, P. E. Constantinou, K. D.Jamison, and M. R. Diehl. Negative interference dominatescollective transport of kinesin motors in the absence ofload. Phys. Chem. Chem. Phys. 11:4882–4889, 2009.
24Rogers, S. L., I. S. Tint, P. C. Fanapour, and V. I. Gelfand.Regulated bidirectional motility of melanophore pigmentgranules along microtubules in vitro. Proc. Natl. Acad. Sci.U.S.A. 94:3720–3725, 1997.
25Ross, J. L., K. Wallace, H. Shuman, Y. E. Goldman, andE. L. F.Holzbaur. Processive bidirectionalmotion of dynein–dynactin complexes in vitro. Nat. Cell Biol. 8:562–570, 2006.
26Uppulury, K., A. K. Efremov, J. W. Driver, D. K. Jamison,M. R. Diehl, and A. B. J. Kolomeisky. How the interplaybetween mechanical and nonmechanical interactions affectsmultiple kinesin dynamics. Phys. Chem. B 116:8846–8855,2012.
27Vale, R. D. The molecular motor toolbox for intracellulartransport. Cell 112:467–480, 2003.
28Xu, J., Z. Shu, S. J. King, and S. P. Gross. Tuning multiplemotor travel via single motor velocity.Traffic 13:1198–1205,2012.
Analysis of Cooperative Behavior in Multiple Kinesins Motor Protein Transport 47
Author's personal copy


















