
Amyloidogenic Propensities and Conformational Properties of ProIAPP and IAPP in the Presence of...
14
Amyloidogenic Propensities and Conformational Properties of ProIAPP and IAPP in the Presence of Lipid Bilayer Membranes Suman Jha 1 , Daniel Sellin 1 , Ralf Seidel 2 and Roland Winter 1 ⁎ 1 Physical Chemistry I—Biophysical Chemistry, Department of Chemistry, TU Dortmund University, Otto-Hahn Str. 6, D-44227 Dortmund, Germany 2 Department Physical Biochemistry, Max-Planck- Institute for Molecular Physiology, Otto-Hahn Str. 11, D-44227 Dortmund, Germany Received 16 December 2008; received in revised form 15 April 2009; accepted 30 April 2009 Available online 7 May 2009 Human islet amyloid polypeptide (hIAPP), which is considered the primary culprit for β-cell loss in type 2 diabetes mellitus patients, is synthesized in β- cells of the pancreas from its precursor pro-islet amyloid polypeptide (proIAPP), which may be important in early intracellular amyloid formation as well. We compare the amyloidogenic propensities and conformational properties of proIAPP and hIAPP in the presence of negatively charged lipid membranes, which have been discussed as loci of initiation of the fibrillation reaction. Circular dichroism studies verify the initial secondary structures of proIAPP and hIAPP to be predominantly unordered with small amounts of ordered secondary structure elements, and exhibit minor differences between these two peptides only. Using attenuated total reflection–Fourier transform infrared spectroscopy and thioflavin T fluorescence spectroscopy, as well as atomic force microscopy, we show that in the presence of negatively charged membranes, proIAPP exhibits a much higher amyloido- genic propensity than in bulk solvent. Compared to hIAPP, it is still much less amyloidogenic, however. Although differences in the secondary structures of the aggregated species of hIAPP and proIAPP at the lipid interface are small, they are reflected in morphological changes. Unlike hIAPP, proIAPP forms essentially oligomeric-like structures at the lipid interface. Besides the interaction with anionic membranes [1,2-dioleoyl-sn- glycero-3-phosphocholine (DOPC)+ x1,2-dioleoyl-sn-glycero-3-[phospho- rac-(1-glycerol)]], interaction with zwitterionic homogeneous (DOPC) and heterogeneous (1,2-dipalmitoyl-sn-glycero-3-phosphocholine:DOPC:cho- lesterol 1:2:1 model raft mixture) membranes has also been studied. Both peptides do not aggregate significantly at DOPC bilayers. In the presence of the model raft membrane, hIAPP aggregates markedly as well. Conversely, proIAPP clusters into less ordered structures and to a minor extent at raft membranes only. The addition of proIAPP to hIAPP retards the hIAPP fibrillation process also in the presence of negatively charged lipid bilayers. In excess proIAPP, increased aggregation levels are finally observed, however, which could be attributed to seed-induced cofibrillation of proIAPP. © 2009 Elsevier Ltd. All rights reserved. Edited by S. Radford Keywords: protein aggregation; amyloid; lipid bilayer; ATR-FTIR; CD *Corresponding author. E-mail address: [email protected]. Abbreviations used: hIAPP, human islet amyloid polypeptide; proIAPP, pro-islet amyloid polypeptide; DOPC, 1,2-dioleoyl-sn-glycero-3-phosphocholine; T2DM, type 2 diabetes mellitus; PC, prohormone convertase; ATR-FTIR, attenuated total reflection–Fourier transform infrared spectroscopy; AFM, atomic force microscopy; HFIP, hexafluoroisopropanol; TFA, trifluoroacetic acid; ThT, thioflavin T; DOPG, 1,2-dioleoyl-sn-glycero-3-[phospho-rac-(1- glycerol)]; DPPC, 1,2-dipalmitoyl-sn-glycero-3-phosphocholine; Chol, cholesterol; IRE, internal reflection element; FSD, Fourier self-deconvolution; LUV, large unilamellar vesicle; D 2 O, deuterium oxide; NaCl, sodium chloride. doi:10.1016/j.jmb.2009.04.077 J. Mol. Biol. (2009) 389, 907–920 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2009 Elsevier Ltd. All rights reserved.
Transcript of Amyloidogenic Propensities and Conformational Properties of ProIAPP and IAPP in the Presence of...

doi:10.1016/j.jmb.2009.04.077 J. Mol. Biol. (2009) 389, 907–920
Available online at www.sciencedirect.com
Amyloidogenic Propensities and ConformationalProperties of ProIAPP and IAPP in the Presence of LipidBilayer Membranes
Suman Jha1, Daniel Sellin1, Ralf Seidel2 and Roland Winter1⁎
1Physical ChemistryI—Biophysical Chemistry,Department of Chemistry, TUDortmund University,Otto-Hahn Str. 6, D-44227Dortmund, Germany2Department PhysicalBiochemistry, Max-Planck-Institute for MolecularPhysiology, Otto-Hahn Str. 11,D-44227 Dortmund, GermanyReceived 16 December 2008;received in revised form15 April 2009;accepted 30 April 2009Available online7 May 2009
*Corresponding author. E-mail addAbbreviations used: hIAPP, huma
1,2-dioleoyl-sn-glycero-3-phosphochattenuated total reflection–Fourier trhexafluoroisopropanol; TFA, trifluorglycerol)]; DPPC, 1,2-dipalmitoyl-snFourier self-deconvolution; LUV, lar
0022-2836/$ - see front matter © 2009 E
Human islet amyloid polypeptide (hIAPP), which is considered the primaryculprit for β-cell loss in type 2 diabetes mellitus patients, is synthesized in β-cells of the pancreas from its precursor pro-islet amyloid polypeptide(proIAPP), which may be important in early intracellular amyloid formationas well. We compare the amyloidogenic propensities and conformationalproperties of proIAPP and hIAPP in the presence of negatively charged lipidmembranes, which have been discussed as loci of initiation of the fibrillationreaction. Circular dichroism studies verify the initial secondary structures ofproIAPP and hIAPP to be predominantly unordered with small amounts ofordered secondary structure elements, and exhibit minor differencesbetween these two peptides only. Using attenuated total reflection–Fouriertransform infrared spectroscopy and thioflavin T fluorescence spectroscopy,as well as atomic force microscopy, we show that in the presence ofnegatively charged membranes, proIAPP exhibits a much higher amyloido-genic propensity than in bulk solvent. Compared to hIAPP, it is still muchless amyloidogenic, however. Although differences in the secondarystructures of the aggregated species of hIAPP and proIAPP at the lipidinterface are small, they are reflected in morphological changes. UnlikehIAPP, proIAPP forms essentially oligomeric-like structures at the lipidinterface. Besides the interaction with anionic membranes [1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC)+x1,2-dioleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)]], interaction with zwitterionic homogeneous (DOPC) andheterogeneous (1,2-dipalmitoyl-sn-glycero-3-phosphocholine:DOPC:cho-lesterol 1:2:1 model raft mixture) membranes has also been studied. Bothpeptides do not aggregate significantly at DOPC bilayers. In the presence ofthe model raft membrane, hIAPP aggregates markedly as well. Conversely,proIAPP clusters into less ordered structures and to a minor extent at raftmembranes only. The addition of proIAPP to hIAPP retards the hIAPPfibrillation process also in the presence of negatively charged lipid bilayers.In excess proIAPP, increased aggregation levels are finally observed,however, which could be attributed to seed-induced cofibrillation ofproIAPP.
© 2009 Elsevier Ltd. All rights reserved.
Edited by S. Radford
Keywords: protein aggregation; amyloid; lipid bilayer; ATR-FTIR; CDress: [email protected] islet amyloid polypeptide; proIAPP, pro-islet amyloid polypeptide; DOPC,oline; T2DM, type 2 diabetes mellitus; PC, prohormone convertase; ATR-FTIR,ansform infrared spectroscopy; AFM, atomic force microscopy; HFIP,oacetic acid; ThT, thioflavin T; DOPG, 1,2-dioleoyl-sn-glycero-3-[phospho-rac-(1--glycero-3-phosphocholine; Chol, cholesterol; IRE, internal reflection element; FSD,ge unilamellar vesicle; D2O, deuterium oxide; NaCl, sodium chloride.
lsevier Ltd. All rights reserved.

908 Propensities and Properties of ProIAPP and IAPP
Introduction
“Amyloid” is the term given to a particular typeof protein aggregate in which proteins aggregateessentially in one dimension and form fibrillarstructures also known as “one-dimensionalcrystallization.”1 These fibrillar structures haveparticular secondary structures independent of theprotein, called cross-β-sheets, as they have β-sheetsaligned perpendicular to the fibrillar axis. Thesefibrillar structures and/or their prefibrillar formshave cytotoxic properties2–4 and are involved invarious diseases such as Alzheimer's disease,Parkinson's disease, and type 2 diabetes mellitus(T2DM). Human islet amyloid polypeptide(hIAPP), also known as amylin, is secreted bypancreatic β-cells along with insulin and residualunprocessed proprotein [pro-islet amyloid poly-peptide (proIAPP), proinsulin]. hIAPP has beenconsidered as the primary culprit for β-cell loss inT2DM patients because of its much higher amyloi-dogenic propensity in comparison to insulin,proinsulin, and proIAPP.5 Under normal condi-tions, hIAPP is a soluble 37-residue peptide that issecreted in response to various stimuli to maintainblood glucose level.6–8 Although its biologicalfunctions are not well-known, hIAPP is believedto play a role as a counterregulator to insulin bysuppressing food intake and increasing the bloodglucose level.9hIAPP is synthesized in β-cells of the pancreas
as the prohormone precursor proIAPP along withproinsulin.6 Prior to cosecretion, they undergodisulfide bond formation in the rough endoplasmicreticulum and then are processed with the samesubtilisin protein family, known as prohormoneconvertase (PC) 2 and PC1/3 or furin, within β-cell secretory granules, followed by the activity ofcarboxypeptidase E to remove the basic PC1/3recognition motif (i.e., Lys-Arg) and then the
Scheme 1. Proposed pathway for the normal pro
activity of peptidyl amidating monooxygenasecomplex to remove Gly38 and amidate theexposed carboxy-terminus of Tyr37 in hIAPP (seeScheme 1).10–13 Thus, normal processing of the67-residue proIAPP to yield the 37-residue hIAPPrequires formation of a disulfide bridge betweenresidues Cys13 and Cys18 of proIAPP, followedby cleavage of 11 residues at the N-terminus, 19residues at the C-terminus, 2 basic residues fromthe resultant C-terminus, and then amidation ofthe carboxy end of tyrosine at the C-terminus ofhIAPP.13 Upon secretagogue stimulation, maturehIAPP is secreted into the extracellular matrixalong with mature insulin. In a recent computersimulation and theoretical study on an hIAPPfragment, we have put forward an explanationfor suppressed aggregation in crowded cellularenvironments based on the statistical mechanicsof aggregation phenomena in finite-size systems.The effect observed may play a decisive role inhampering intracellular aggregation of highlyinsoluble amyloidogenic proteins such as hIAPP,whereas aggregation is unavoidable in the extra-cellular space at the same peptide concentration.14
The prediabetic stage of T2DM is accompanied bya higher insulin need that causes release ofpremature secretory granules, through secretagoguestimulation, with a higher amount of residualproprotein (proIAPP and proinsulin). Furthermore,increased levels of free fatty acids have beenobserved in this prediabetic stage of T2DM.15
Work from various groups16–18 suggested that β-cells exposed to hyperlipidemia remodulate theirmembranes by increasing the negatively chargedlipid content. In the prediabetic stage, the anioniclipid content of the membrane seems to increasebeyond the physiological range (i.e., N10–30%).In fact, it has been shown that hIAPP bindsstrongly to membranes that have more than about30% anionic lipids, and this is supposed to lead
cessing of human proIAPP in pancreatic β-cells.
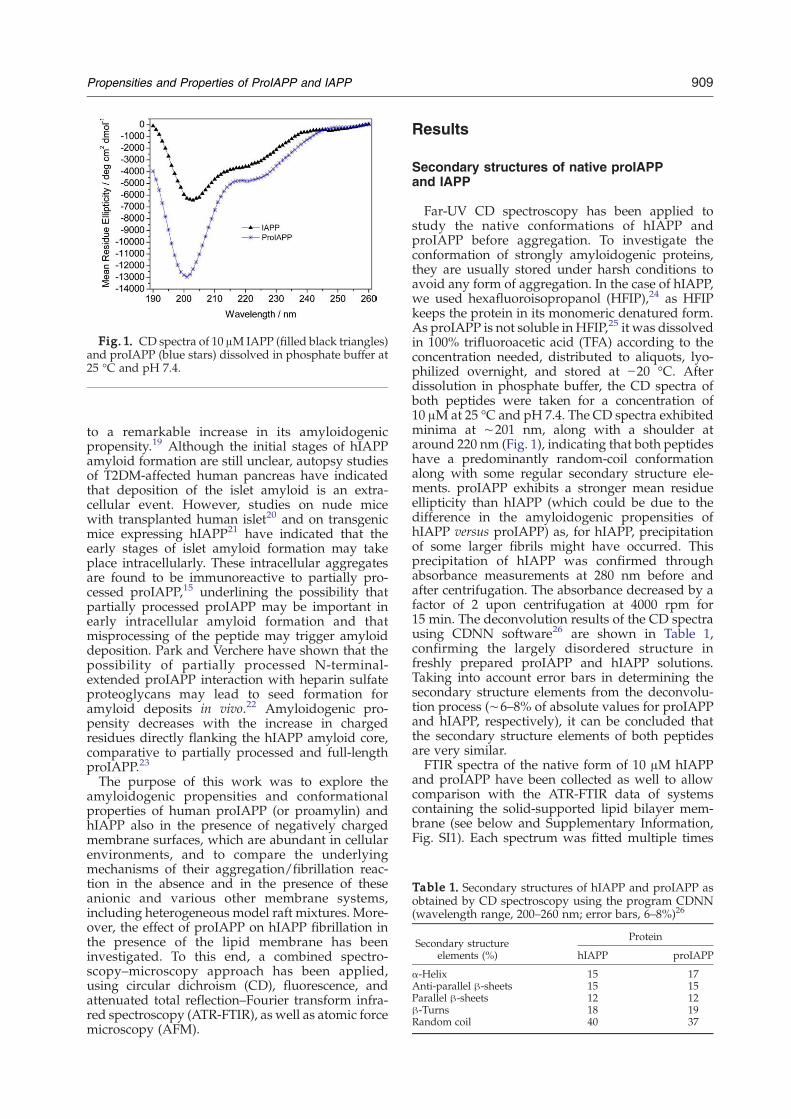
Fig. 1. CD spectra of 10 μM IAPP (filled black triangles)and proIAPP (blue stars) dissolved in phosphate buffer at25 °C and pH 7.4.
Table 1. Secondary structures of hIAPP and proIAPP asobtained by CD spectroscopy using the program CDNN(wavelength range, 200–260 nm; error bars, 6–8%)26
Secondary structureelements (%)
Protein
hIAPP proIAPP
α-Helix 15 17Anti-parallel β-sheets 15 15Parallel β-sheets 12 12β-Turns 18 19Random coil 40 37
909Propensities and Properties of ProIAPP and IAPP
to a remarkable increase in its amyloidogenicpropensity.19 Although the initial stages of hIAPPamyloid formation are still unclear, autopsy studiesof T2DM-affected human pancreas have indicatedthat deposition of the islet amyloid is an extra-cellular event. However, studies on nude micewith transplanted human islet20 and on transgenicmice expressing hIAPP21 have indicated that theearly stages of islet amyloid formation may takeplace intracellularly. These intracellular aggregatesare found to be immunoreactive to partially pro-cessed proIAPP,15 underlining the possibility thatpartially processed proIAPP may be important inearly intracellular amyloid formation and thatmisprocessing of the peptide may trigger amyloiddeposition. Park and Verchere have shown that thepossibility of partially processed N-terminal-extended proIAPP interaction with heparin sulfateproteoglycans may lead to seed formation foramyloid deposits in vivo.22 Amyloidogenic pro-pensity decreases with the increase in chargedresidues directly flanking the hIAPP amyloid core,comparative to partially processed and full-lengthproIAPP.23
The purpose of this work was to explore theamyloidogenic propensities and conformationalproperties of human proIAPP (or proamylin) andhIAPP also in the presence of negatively chargedmembrane surfaces, which are abundant in cellularenvironments, and to compare the underlyingmechanisms of their aggregation/fibrillation reac-tion in the absence and in the presence of theseanionic and various other membrane systems,including heterogeneous model raft mixtures. More-over, the effect of proIAPP on hIAPP fibrillation inthe presence of the lipid membrane has beeninvestigated. To this end, a combined spectro-scopy–microscopy approach has been applied,using circular dichroism (CD), fluorescence, andattenuated total reflection–Fourier transform infra-red spectroscopy (ATR-FTIR), as well as atomic forcemicroscopy (AFM).
Results
Secondary structures of native proIAPPand IAPP
Far-UV CD spectroscopy has been applied tostudy the native conformations of hIAPP andproIAPP before aggregation. To investigate theconformation of strongly amyloidogenic proteins,they are usually stored under harsh conditions toavoid any form of aggregation. In the case of hIAPP,we used hexafluoroisopropanol (HFIP),24 as HFIPkeeps the protein in its monomeric denatured form.As proIAPP is not soluble in HFIP,25 it was dissolvedin 100% trifluoroacetic acid (TFA) according to theconcentration needed, distributed to aliquots, lyo-philized overnight, and stored at −20 °C. Afterdissolution in phosphate buffer, the CD spectra ofboth peptides were taken for a concentration of10 μM at 25 °C and pH 7.4. The CD spectra exhibitedminima at ∼201 nm, along with a shoulder ataround 220 nm (Fig. 1), indicating that both peptideshave a predominantly random-coil conformationalong with some regular secondary structure ele-ments. proIAPP exhibits a stronger mean residueellipticity than hIAPP (which could be due to thedifference in the amyloidogenic propensities ofhIAPP versus proIAPP) as, for hIAPP, precipitationof some larger fibrils might have occurred. Thisprecipitation of hIAPP was confirmed throughabsorbance measurements at 280 nm before andafter centrifugation. The absorbance decreased by afactor of 2 upon centrifugation at 4000 rpm for15 min. The deconvolution results of the CD spectrausing CDNN software26 are shown in Table 1,confirming the largely disordered structure infreshly prepared proIAPP and hIAPP solutions.Taking into account error bars in determining thesecondary structure elements from the deconvolu-tion process (∼6–8% of absolute values for proIAPPand hIAPP, respectively), it can be concluded thatthe secondary structure elements of both peptidesare very similar.FTIR spectra of the native form of 10 μM hIAPP
and proIAPP have been collected as well to allowcomparison with the ATR-FTIR data of systemscontaining the solid-supported lipid bilayer mem-brane (see below and Supplementary Information,Fig. SI1). Each spectrum was fitted multiple times

Fig. 2. Secondary structure elements of the nativestructures of hIAPP and proIAPP in phosphate buffer at25 °C and pH 7.4, as obtained by FTIR spectroscopy.
Fig. 3. ThT-monitored amyloid fibril formation kineticsof 10 μM hIAPP (open black rectangles), 10 μM proIAPP(filled magenta stars), 10 μM proIAPP+10 μM hIAPP(open red triangles), 30 μM proIAPP+10 μM hIAPP (openblue stars), and 40 μM proIAPP (olive-green open crosses)in the presence of a DOPC:DOPG (7:3, wt/wt) lipidbilayer. Experiments were carried out in phosphate bufferat 25 °C (pD=7.4).
910 Propensities and Properties of ProIAPP and IAPP
with a similar set of subbands and slight variationsin the starting conditions for each fit. Figure 2 showsthe results of the fits, including the correspondingerror bars. We have to keep in mind that, owing tounknown transition dipole moments of differentconformers in the FTIR data, no absolute numberscan be given here, hence relative changes areessentially discussed only. A comparison of second-ary structure contents shows again that bothpeptides have a similar native conformation.proIAPP is slightly richer in α-helical conformations,whereas hIAPP possesses a slightly higher contentof unordered and turn structures.
Fibrillation kinetics in the presence of1,2-dioleoyl-sn-glycero-3-phosphocholine:1,2-dioleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)](7:3, wt/wt) lipid bilayers using thioflavin T assayand comparison with other lipid bilayer systems
Thioflavin T (ThT), a benzothiazole dye, exhibitsenhanced fluorescence upon binding to amyloidfibrils and is commonly used to diagnose amyloidfibrils.27 ThT changes its fluorescent characteristicsremarkably upon interaction with amyloids.Unbound ThT has its excitation maximum at385 nm, and the emission maximum is at 445 nm.Upon interaction with fibrillar or prefibrillar struc-tures, the excitation maximum shifts to 440 nm, andthe emission maximum shifts to 482 nm. Wemonitored the increase in the ThT emission maximaat 25 °C with time for 10 μM hIAPP and 40 μMproIAPP in the presence of 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC):1,2-dioleoyl-sn-glycero-3-[phospho-rac-(1-glycerol)] (DOPG) (7:3, wt/wt),as depicted in Fig. 3 (for 10 μM proIAPP, noaggregation was observed over the time spanshown). The fibrillation kinetics follows a nuclea-tion-dependent aggregation mechanism, including alag phase accompanied by nuclei formation, fol-lowed by an exponential growth phase and then astationary phase, finally displaying a sigmoid-likecurve. The lag phase is generally used to define theamyloidogenic propensity of a protein. The shorter
is the lag phase, the more amyloidogenic is theprotein. As shown in Fig. 3, 10 μM hIAPP has a lagphase of 40 min under these experimental condi-tions, in comparison to 38 h for 40 μm proIAPP.Thus, hIAPP has a markedly higher amyloidogenicpropensity (by a factor of ∼57 for the fourfoldsmaller concentration) than proIAPP. Still, thepresence of the anionic DOPC:DOPG membrane isable to trigger the aggregation/fibrillation processof proIAPP. In the absence of the lipid bilayer, noaggregation has been observed for 40 μM proIAPPat 25 °C up to 7 days. Within this time period,aggregation of proIAPP has also not been observedat membranes with significantly less anionic lipidcontents (experiments were performed at DOPC:DOPG wt/wt ratios of 9:1 and 8:2): at the purezwitterionic DOPC lipid bilayer and at the 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC):DOPC:cholesterol (Chol) (1:2:1) model raft mixture(data not shown).
Conformational changes in hIAPP and proIAPPat solid interfaces and in the presence of lipidmembranes as revealed by ATR-FTIR
First, the conformational changes in hIAPP werestudied in the absence and in the presence of thelipid interface at 25 °C. To evaluate the changes inthe secondary structure of hIAPP at the internalreflection element (IRE) crystal (Ge) and at themembrane interface, we collected ATR-FTIR spectraover a time period of 20 h in 5-min intervals afterinjection of a solution of 10 μM hIAPP into the ATRcell. Due to interfacial adsorption, the local concen-tration close to the IRE crystal surface is increasedwith time t, and aggregation is induced. Duringthe aggregation process, the peak maximum of the

911Propensities and Properties of ProIAPP and IAPP
amide-I′ band of hIAPP shifts from 1644 to1627 cm−1, indicating a decrease in unorderedconformations and a concomitant increase in β-structures. In Fig. 4a, spectra for selected time pointsare presented. On the right-hand side of Fig. 4a, thecorresponding normalized spectra are shown forbetter visualization of the changes that occurred.Examination of Fourier self-deconvolution (FSD)and second derivative of the normalized FTIRspectra (Supplementary Information, Fig. SI2)revealed infrared subbands for the native hIAPP(at t=1 min) at about 1680, 1674, 1665, 1658, 1649,1642, and 1632 cm−1 and a shoulder at 1623 cm−1,reflecting a complex composition of secondarystructure elements. During the aggregation process,some of the bands shift towards larger wavenum-bers, and a broad band appears at 1616–1619 cm−1,reflecting the formation of intermolecular β-sheetswith strong hydrogen bonding. Peak fitting of thenormalized spectra was applied to allow a quanti-tative analysis of secondary structural changes.In Fig. 5a, the time dependence of the secondarystructural changes in hIAPP is shown for thefirst 10 h. The (probably parallel) intermolecularβ-sheet contribution representing the subbandat ∼1635 cm−1 is not shown, since its intensity(∼12.5%) does not change. As can also be seen in
Fig. 4. Time evolution of the amide-I′ bands of hIAPP andATR cell. (a) hIAPP (10 μM) in the absence of membrane. (b)consisting of DOPC:DOPG (7:3, wt/wt). (c) proIAPP (40 μMprimary ATR-FTIR spectra after buffer, noise subtraction, aninfrared spectra are shown.
Fig. 5, intermolecular β-sheet formation (bands at1626 and 1616 cm−1) of hIAPP occurs at the expenseof a decrease in turns/loops, α-helical structures,and unordered structures.In a next step, we followed the fibrillation reaction
of hIAPP in the presence of the anionic DOPC:DOPG membrane. To evaluate secondary structuralchanges, we collected FTIR spectra up to 20 h inintervals of 5 min after the injection of a solution of10 μMhIAPP into the ATR cell containing the spreadlipid bilayer film. The time evolution of the amide-I′band is shown for selected time points in Fig. 4b(left). The gross features of these data are in goodagreement with earlier data.28 Here, we put forwarda more detailed and quantitative analysis of theaggregation reaction by applying resolutionenhancement methods and curve fitting to thespectra. During the aggregation process, the peakmaximum of the amide-I′ band of hIAPP shifts from1646 to 1624 cm−1. On the right-hand side of Fig. 4b,the corresponding normalized spectra are shown aswell. Examination of the FSD and second derivativeof the normalized spectra (Supplementary Informa-tion, Fig. SI2) revealed six distinct bands for thehIAPP conformation after t=6 min at about 1674,1666, 1662, 1652, 1645, and 1638 cm−1, with themost prominent band at 1645 cm−1. During the
proIAPP upon aggregation at 25 °C after injection into thehIAPP (10 μM) in the presence of phospholipid bilayers) in the presence of DOPC:DOPG (7:3, wt/wt). Left: Thed baseline correction are shown. Right: Normalized ATR

Fig. 5. Time evolution of secondary structural changesupon fibrillation of 10 μM hIAPP in the absence (a) and inthe presence (b) of the DOPC:DOPG (7:3, wt/wt)membrane derived from peak fitting of the normalizedATR-FTIR spectra. (c) Time evolution of secondarystructural changes upon aggregation of 40 μM proIAPPin the presence of the DOPC:DOPG (7:3, wt/wt) mem-brane derived from peak fitting of the normalized ATR-FTIR spectra.
912 Propensities and Properties of ProIAPP and IAPP
aggregation process, new bands at ∼1684, 1626, and1619 cm−1 appear. The strong bands at 1619 and1626 cm−1 indicate the presence of intermolecular β-sheets. Peak fitting of the normalized spectra wascarried out for selected spectra within the first 5 h ofthe measurements to allow quantitative analysis oftime-dependent changes in secondary structure, asshown in Fig. 5b. Comparison of Fig. 5a and b showsthat, besides the different timescale, the aggregationprocess of hIAPP at the solid support (ATR crystal)and in the presence of the anionic membraneproceeds essentially with similar conformationalchanges. In the presence of the anionic lipid inter-face, more intermolecular β-sheets (+5%) and lessturns (−5%) are formed, however.The aggregation mechanism of proIAPP in contact
with the anionic lipid membrane was studied in asimilar manner. To evaluate the changes in thesecondary structure of proIAPP in contact with thephospholipid bilayer, we collected FTIR spectra overa time period of 7 days with intervals of 30 min afterthe injection of a solution of 40 μM proIAPP into themembrane containing an ATR cell (a 10 μMproIAPPconcentration was measured as well but is notshown here as it did not aggregate up to 7 days at25 °C). During the aggregation process, the peak
Fig. 6. Second derivative spectra (a) and FSD (b) of theamide-I′ infrared bands of 40 μM proIAPP duringadsorption to the DOPC:DOPG (7:3, wt/wt) membrane.Left arrows indicate the strong blue shift of the subband at∼1647 cm−1, which is usually assigned to α-helical/disordered conformations. For better visualization, thespectra have been offset.
913Propensities and Properties of ProIAPP and IAPP
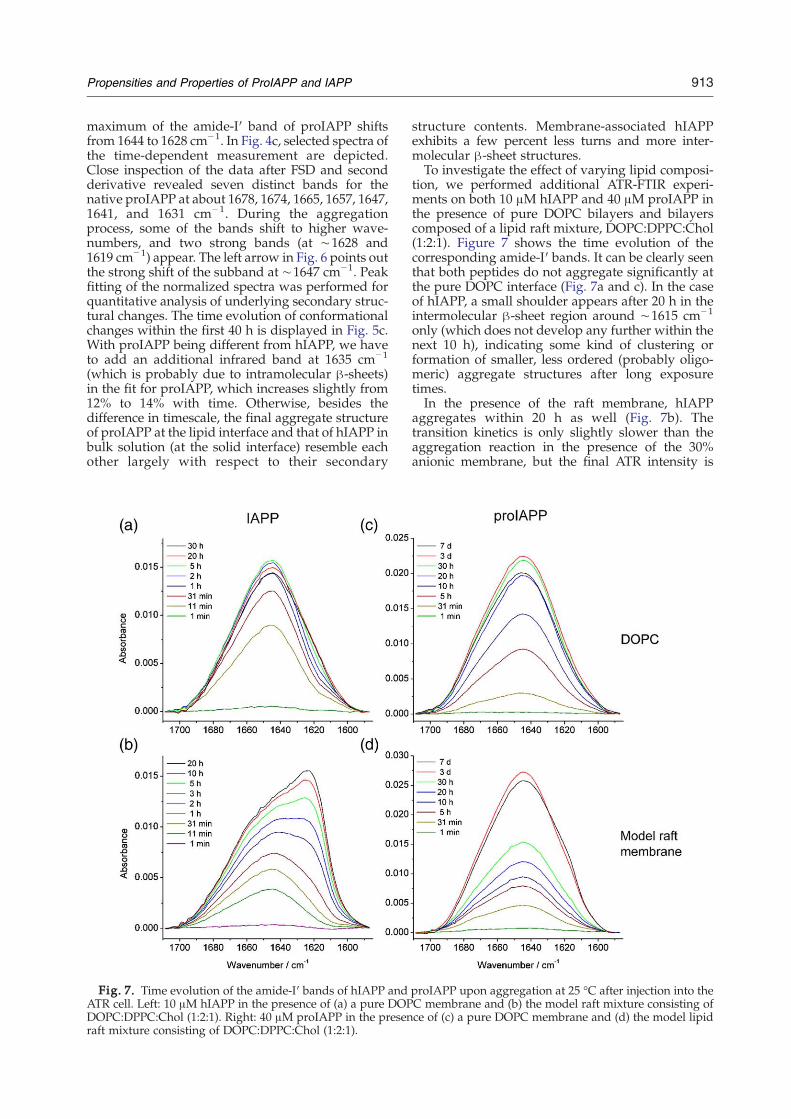
maximum of the amide-I′ band of proIAPP shiftsfrom 1644 to 1628 cm−1. In Fig. 4c, selected spectra ofthe time-dependent measurement are depicted.Close inspection of the data after FSD and secondderivative revealed seven distinct bands for thenative proIAPP at about 1678, 1674, 1665, 1657, 1647,1641, and 1631 cm− 1. During the aggregationprocess, some of the bands shift to higher wave-numbers, and two strong bands (at ∼1628 and1619 cm−1) appear. The left arrow in Fig. 6 points outthe strong shift of the subband at ∼1647 cm−1. Peakfitting of the normalized spectra was performed forquantitative analysis of underlying secondary struc-tural changes. The time evolution of conformationalchanges within the first 40 h is displayed in Fig. 5c.With proIAPP being different from hIAPP, we haveto add an additional infrared band at 1635 cm−1
(which is probably due to intramolecular β-sheets)in the fit for proIAPP, which increases slightly from12% to 14% with time. Otherwise, besides thedifference in timescale, the final aggregate structureof proIAPP at the lipid interface and that of hIAPP inbulk solution (at the solid interface) resemble eachother largely with respect to their secondary
Fig. 7. Time evolution of the amide-I′ bands of hIAPP andATR cell. Left: 10 μM hIAPP in the presence of (a) a pure DOPDOPC:DPPC:Chol (1:2:1). Right: 40 μM proIAPP in the presenraft mixture consisting of DOPC:DPPC:Chol (1:2:1).
structure contents. Membrane-associated hIAPPexhibits a few percent less turns and more inter-molecular β-sheet structures.To investigate the effect of varying lipid composi-
tion, we performed additional ATR-FTIR experi-ments on both 10 μM hIAPP and 40 μM proIAPP inthe presence of pure DOPC bilayers and bilayerscomposed of a lipid raft mixture, DOPC:DPPC:Chol(1:2:1). Figure 7 shows the time evolution of thecorresponding amide-I′ bands. It can be clearly seenthat both peptides do not aggregate significantly atthe pure DOPC interface (Fig. 7a and c). In the caseof hIAPP, a small shoulder appears after 20 h in theintermolecular β-sheet region around ∼1615 cm−1
only (which does not develop any further within thenext 10 h), indicating some kind of clustering orformation of smaller, less ordered (probably oligo-meric) aggregate structures after long exposuretimes.In the presence of the raft membrane, hIAPP
aggregates within 20 h as well (Fig. 7b). Thetransition kinetics is only slightly slower than theaggregation reaction in the presence of the 30%anionic membrane, but the final ATR intensity is
proIAPP upon aggregation at 25 °C after injection into theC membrane and (b) the model raft mixture consisting ofce of (c) a pure DOPC membrane and (d) the model lipid

914 Propensities and Properties of ProIAPP and IAPP
lower. For better visualization of the affinity ofhIAPP for the different membrane systems, adsorp-tion kinetics is shown in Fig. 10a by plotting theamide-I′ band intensity as a function of time. Thecorresponding time evolution of the intermolecularβ-sheet content, which is characteristic of theaggregation propensity of hIAPP in the presence ofvarious membranes, is depicted in Fig. 10b. In thecase of the raft mixture, only 30% of the amount ofaggregate structures are found when compared tohIAPP at the anionic membrane.When exposed to the raft mixture, the amide-I′
band of proIAPP displays a small shoulder in theintermolecular β-sheet region after 3 days only,which further evolved after 7 days (Fig. 7d). Hence,proIAPP seems to aggregate in the presence of theheterogeneous raft membrane to a minor extentonly, but the kinetics is very slow and the aggregatestructure does not seem to be well ordered.
Morphology of proIAPP aggregates at the lipidinterface
AFM is a well-known technique that is used toreveal the morphology of protein aggregates, also inthe presence of membranous interfaces.29,30 We usedthe spin-coating method to prepare the samples andfreeze–dry them, so that it takes a minimum time todry and to minimize surface-induced aggregationeffects. Unlike other membrane-induced fibrillatingsystems such as hIAPP,29,31,32 we found proIAPP toform oligomeric or small fibrillar structures only.Notably, there is no major spectral difference in theATR-FTIR data for these two peptides, hence thedifferent aggregates (large oligomeric versus fibril-lar) must essentially differ in the packing propertiesof the aggregating units only. Figure 8 depictscharacteristic AFM images of 40 μM aggregatedproIAPP in the presence of the anionic lipid bilayer.Essentially oligomeric structures were found withheights in the range of 2.5–3.5 nm. Along withthese oligomeric species, a minor population ofsmall fibrillar structures having lengths of 50–55 nmwas found as well. A similar analysis for 40 μM
Fig. 8. AFM image of proIAPP aggregated at a concentratiomembrane at 25 °C for 7 days (left) and corresponding height
proIAPP in bulk solution under the same conditionsup to 7 days did not show either significantsecondary structural changes (measured by ATR-FTIR) or changes in morphology (by AFM) (datanot shown).
Amyloidogenic propensity of hIAPP in thepresence of proIAPP
To investigate how proIAPP influences the amy-loidogenicity of hIAPP at the lipid interface, we firstinjected a mixture of 10 μM hIAPP and 10 μMproIAPP after rigorous mixing for 3 h (using anoverhead shaker) into the membrane containing theATR cell. Spectra were collected every 30 min over atime course of 96 h. During this time period, no shiftin the amide-I′ band maximum was observed. Asseen in Fig. 9a, the amide-I′ band increases withtime, displaying a low overall intensity (hencesmaller signal-to-noise ratio) even after 20 h. Thecorresponding ThT binding assay, which was con-ducted under the same conditions, showed amarked emission peak at 482 nm, however, indicat-ing that aggregation has essentially occurred in thebulk phase during the mixing time. The ThT bindingassay for the peptide mixture hIAPP:proIAPP (1:1and 1:3) with shorter mixing times of 4 min ofvortexing only revealed aggregation, but withprolonged lag phases compared to hIAPP (Fig. 3).The corresponding ATR-FTIR measurements nowshow aggregation at the anionic lipid interface aswell (Fig. 9b), and intermolecular β-sheet formationis slightly retarded compared to that of pure hIAPPin the presence of the anionic membrane.The fibrillation kinetics observed by the ThT
binding assay can be compared with the relativeintensity of intermolecular β-sheet formation inFig. 10. The adsorption kinetics for hIAPP isslightly faster than that for the 1:1 peptide mixtureat the anionic membrane (Fig. 10a), and the sameholds true for fibrillation kinetics. Notably, the finaladsorption and intermolecular β-sheet intensitiesare smaller for hIAPP in comparison to the peptidemixture, which could be attributed to seed-induced
n of 40 μM in the presence of the DOPC:DOPG (7:3, wt/wt)scale (right).

Fig. 9. Time course of the ATR-FTIR spectra of a 1:1 mixture of hIAPP and proIAPP (each 10 μM) after the sample wasshaken (a) for 3 h or (b) 4 min before it was injected into the ATR cell containing the spread DOPC:DOPG (7:3) lipid bilayermembrane.
915Propensities and Properties of ProIAPP and IAPP
cofibrillation of proIAPP. The longer lag andelongation phase of the proIAPP+IAPP mixturecould be attributed to the interaction and complexformation between the highly amyloidogenic coreof hIAPP and distinct regions, such as the pro-region, of proIAPP molecules.
Discussion
hIAPP is the major constituent of pancreaticamyloid in T2DM. It has been shown to exhibit astrong amyloidogenic and cytotoxic potential invitro.19 In vivo, hIAPP is stored as proIAPP insecretory granules and secreted upon processing inresponse to insulin secretagogue. The main focus ofthis work was to explore the amyloidogenicpropensity and conformational properties ofproIAPP in the presence of negatively chargedmembranes, such as DOPC:DOPG (7:3, wt/wt)
Fig. 10. (a) Adsorption kinetics of a mixture of hIAPP and(DOPC:DOPG, 7:3) membrane; that of hIAPP in the presencmodel raft membrane; and that of a pure DOPC lipid bilayerbetween 1710 and 1585 cm−1. (b) The corresponding intermoarea in the presence of the anionic membrane and that of hIAP
phospholipid bilayers, in comparison to hIAPP.Specific aims were to reveal the timescale andunderlying mechanisms of the aggregation/fibrilla-tion process of the two peptides in the presence oflipid membranes. Besides anionic membranes, purezwitterionic and model raft mixtures were also usedas model membrane systems for the interactionstudies. In fact, lipid membranes seem to constitutethe preferred loci of aggregation and probably alsoof cytotoxicity.29,31,32 Moreover, the effect ofproIAPP on hIAPP fibrillation was explored. Tothis end, CD, fluorescence, and ATR-FTIR, as well asAFM methods, were applied.The secondary structures of the two peptides have
first been determined shortly after injection of thesamples to capture their initial native-like conforma-tion, using CD spectroscopy and FTIR. The resultsconfirm earlier results33–35 that hIAPP in the nativestate consists mainly of unordered structures (ca40%) with some amounts of α-helical, β-sheet, turn,
proIAPP (1:1; each 10 μM) in the presence of an anionice of the anionic membrane, the DOPC:DPPC:Chol (1:2:1)membrane, followed by integration of the amide-I′ bandlecular β-sheet content with respect to the whole amide-I′P in the presence of the anionic and model raft membrane.

916 Propensities and Properties of ProIAPP and IAPP
and loop conformations. Interestingly, the second-ary structure of proIAPP is found to be very similarto that of hIAPP in bulk solution. proIAPP is slightlyricher in α-helical conformations, whereas hIAPPpossesses a slightly higher content of unordered andturn structures only.By examining the fibrillation kinetics monitored
with the ThT assay (Fig. 3), it can be seen that hIAPPin the presence of the anionic membrane has a lagphase of∼40min under the experimental conditionschosen (10 μM, 25 °C), whereas proIAPP exhibits alag phase of ∼39 h for a fourfold higher concentra-tion. Conversely, in bulk solution, 40 μM proIAPPdid not aggregate when left over a time period of7 days. Certainly, different lag and growth times areseen in different sample environments, such as in theATR cell, where no extended lag phase is observedfor hIAPP and the lag time for proIAPP is in therange of∼4 h. Differences in timescale are due to thedifferent sample geometry and vesicle preparations.In the presence of supported phospholipid bilayers(ATR experiment), the signal is due to the concen-tration increase in the peptides upon adsorption atthe flat lipid interface, while in the ThT experiment,the peptides interact with large unilamellar vesicles(LUVs).Our data thus clearly show that a strong self-
associating potential is achieved for proIAPP in thepresence of anionic membranes. Nevertheless,proIAPP is found to be markedly less amyloido-genic than hIAPP. This decreased amyloidogenicityof proIAPP may be rationalized by the increased netcharge on proIAPP (see Scheme 1) and the increasein in-register charges in the C-terminal extensionsequence directly flanking the hIAPP amyloidogeniccore.23
Comparison of Fig. 5a and b shows that, besidesthe different timescale, the aggregation process ofhIAPP in the absence and in the presence of theanionic membrane proceeds largely with similarconformational changes. For hIAPP aggregated inthe presence of the lipid membrane, more inter-molecular β-sheets (+5%) and less turns (−5%) areformed, however. Interestingly, the comparison ofthe different aggregating systems investigatedreveals that the final secondary structure of aggre-gated hIAPP in the absence of a lipid interface andthat of aggregated proIAPP in contact with theanionic membrane are rather similar.This difference in the secondary structure content
of the aggregated species of hIAPP and proIAPP atthe membranous interface is also reflected inmorphological changes. Unlike hIAPP, proIAPPformsmainly small oligomeric-like disordered struc-tures at the lipid membrane, having heights of∼3.5 nm. These heights correspond to the heightsof normal fibrils, which vary typically from 2 nm (inthe case of hIAPP fragments36,37 and α-synuclein38)to 6 nm (e.g., for IAPP39 and insulin40,41). Thisdifference in the morphological structure of proIAPPmay be rationalized by the presence of the pro-region(i.e., its C-terminal and N-terminal ends flanking theamyloidogenic core), which largely corresponds
to the sequence of the much more amyloidogenichIAPP. It may be rationalized that, due to theseadditional ends, efficient packing into orderedfibrillar structures is hampered, thus leading to themore disordered oligomeric aggregate structuresobserved by AFM for proIAPP.Both peptides hIAPP and proIAPP do not aggre-
gate significantly in the presence of the purezwitterionic fluid DOPC membrane within a timeperiod of up to 7 days. Conversely, in the presence ofzwitterionic but heterogeneous model raft mem-branes, hIAPP aggregates markedly as well. Thetransition kinetics is only slightly slower, but thecontents of aggregate structures were about 60%lower than that in the presence of the anionicmembrane. One may speculate that the initialadsorption of hIAPP molecules at the rim of thelipid domains may serve as nucleation sites. Thedecrease in the line tension of neighboring domainsby peptide insertion might serve as the driving forceof this process. On the contrary, proIAPP seems tocluster into less ordered structures in the presence ofthe heterogeneous raft membrane and to a minorextent only, and the kinetics is very slow.We have also shown that the addition of proIAPP
to hIAPP delays the rate of IAPP fibril formation,probably by some kind of complex formation. A 1:1molar mixture of proIAPP and hIAPP increased thelag time by a factor of ∼3 (Fig. 3). Thus, it appearsreasonable to speculate that the pro-region of theproIAPP could serve to prolong the fibrillationkinetics of the highly amyloidogenic peptide hIAPP.In fact, it has been speculated that part of the functionof hIAPP intermediates such as proIAPP may be toprevent aggregation and amyloidogenesis in theearly stages of hIAPP biosynthesis and transport.23
Interestingly, in particular in the case of excessproIAPP, increased aggregation levels have finallybeen observed, however, which could be attributedto seed-induced cofibrillation of proIAPP. Reports onthe aggregation of other systems where similarproteins have been compared, such as differentinsulin constructs or homologues of lysozyme,42,43
are still scarce, so that generalizations are difficult tomake. What seems to be clear, however, is thatseeding fibril formation depends strongly on thesimilarity of their sequences, which is largely fulfilledfor hIAPP and its pro-peptide proIAPP.
Materials and Methods
Materials
Sodium hydrogen phosphate, deuterium oxide (D2O),deuterium chloride, and sodium chloride (NaCl) wereobtained from Sigma-Aldrich (St. Louis, MO, USA).HFIP was obtained from Fluka, whereas chloroformand ThT were obtained from Merck. Amylin (hIAPP)was obtained from Calbiochem (La Jolla, CA, USA). Thelipids DOPC (zwitterionic) and DOPG (anionic) wereobtained from Avanti Polar Lipids (Alabaster, AL, USA).Aqueous solutions were prepared using deionized water

917Propensities and Properties of ProIAPP and IAPP
throughout the study, except for the ATR-FTIR study,which was performed in D2O medium. For purification ofproIAPP, all solvents used were of HPLC grade, and Ni-NTA resin was purchased from Sigma. The enterokinaseenzyme was purchased from Novagen (Wisconsin, USA).Muscovite mica for the AFM studies was obtained fromPlano GmbH (Wetzlar, Germany).
Expression and purification of proIAPP
A plasmid encoding human proIAPP was a generousgift from Prof. Dr. A. Kapurniotu (Technical University ofMunich, Munich, Germany) and was transformed intoEscherichia coli BL21(DE3) strain. Transformed E. coli BL21(DE3) cells were grown at 37 °C in 2× YT mediumcontaining 100 mg/l ampicillin. Overexpression wasinduced by IPTG for 3 h at 37 °C. Cells were harvestedby centrifugation (4000g, 15 min, 4 °C), and cell pelletswere resuspended in 25 mMTris–HCl (pH 8.0) and 50mMNaCl. Cells were disrupted in a Microfluidizer (Micro-fluidics Corporation, Newton, MA, USA) and centrifuged(48,000g, 30 min, 7 °C). GdnHCl (8 M) in 10 mM Tris–HCl(pH 8.0) and 100 mM NaCl was added to the supernatantup to a final concentration of 6 M GndHCl. Thesupernatant containing the expressed protein was passedthrough Ni-NTA resin and eluted with 100 mM imidazole,10 mM Tris–HCl (pH 8.0), and 100 mM NaCl. The eluatewas dialyzed against 50 mM Tris–HCl (pH 8) and 50 mMNaCl, and the resulting precipitant was immediatelydissolved to final concentrations of 4 M GdnHCl, 10 mMTris–HCl (pH 8.0), and 100 mM NaCl. The dissolvedprotein solution was loaded on a C18 column (ProntoSILHPLC, Germany). An elution gradient used was asfollows: 10 min at 10% buffer B (90% acetonitrile and0.05% TFA in H2O) in 90% buffer A (0.06% TFA in H2O);60 min from 10% to 90% buffer B in buffer A. The flow ratewas 2 ml/min throughout the run, and detection of thepeptide was performed at 214 nm. The fusion proteinfraction was confirmed by electrospray ionization–massspectrometry, then lyophilized and dissolved in enteroki-nase cleavage buffer [20 mM Tris–HCl (pH 7.4), 50 mMNaCl, and 2 mM CaCl2]. The cleavage reaction wasperformed with an enterokinase enzyme for 16 h at roomtemperature (20 °C). The cleavage product was lyophi-lized, dissolved in 10% acetic acid, and purified on RP-HPLC with the same elution profile as followed for thefusion protein. The purity and identity of human proIAPPwere determined with SDS-PAGE and electrosprayionization–mass spectrometry (LCQ Advantage MAX;Thermo). The protein concentration was estimated fromabsorbance at 280 nm in water, using an extinctioncoefficient of 3230 M−1 cm−1.44 Following concentrationdetermination, the proIAPP fraction was solved in 100%TFA and then distributed to aliquots and lyophilized. Itwas stored at −20 °C for further use.
CD measurements
Far-UV CD measurements were performed on a JASCOJ715 spectrometer (Jasco Corporation, Japan) by dissol-ving lyophilized proIAPP or hIAPP aliquots to aconcentration of 10 μM. Spectra were recorded using a0.1-cm path-length cuvette over the range of 190–260 nmwith a scanning speed of 50 nm/min and a response timeof 1 s at a spectral resolution of 1 nm. Five spectra wereaveraged and corrected against 10 mM phosphate buffer(pH 7.4). Secondary structure estimation was performedusing the CDNN program.26
ThT binding
The lyophilized proIAPP and hIAPP aliquots weredissolved in a reaction buffer and incubated in a quartzcuvette at 25 °C without stirring. Following incubation,fluorescence intensity was recorded with excitation at440 nm, and an emission spectrum was recorded in therange of 450–540 nm on a K2 fluorometer (ISS, Urbana, IL,USA). Figure 2 shows the relative fluorescence intensities atλ482 nm after addition of 50 μM ThTwith time. The spectrawere background-subtracted against the ThT fluorescencein the reaction buffer without protein. The reaction bufferused was 10 mM phosphate buffer (pH 7.4) with 100 mMNaCl and 0.5 mg of DOPC:DOPG (7:3, wt/wt).
Attenuated total reflection–Fourier transform infraredspectroscopy
ATR-FTIR measurements45 were performed using aNicolet 6700 FTIR Spectrometer equipped with a liquid-nitrogen-cooled MCT-A detector. The spectrometer cham-ber was continuously purgedwith dry air to remove watervapor. The ATR out-of-compartment accessory consists ofa liquid jacketed Piketech ATR flow-through cell made ofsteel. A trapezoidal Ge-crystal (80 mm×10 mm×4 mm,45° angle of incidence; Piketech, Madison, WI, USA) wasused as internal reflection unit (IRE). A backgroundspectrum was collected using the same IRE before thesample measurements. Typically, spectra of 128 scanswere taken with a resolution of 2 cm−1. The measurementswere usually performed at 25 °C. Processing of the ATR-FTIR spectra was performed using GRAMS software(Thermo Electron). After subtraction of a spectrum of thelipid membrane together with the buffer and subtractionof noise from water vapor, the spectra were baseline-corrected between 1710 and 1585 cm−1 and also offset-corrected. To follow the changes in secondary structurewith time, the spectra were normalized by setting the areabetween 1710 and 1585 cm−1 of the amide-I′ band to 1. Tocompare differences in adsorption propensities at thecrystal and membrane surface, respectively, no suchnormalization was used. No smoothing has been appliedto the spectra.For all infrared spectroscopic experiments, a phosphate
buffer (10 mM PO43−; pD=7.4) in D2O containing NaCl
(total ionic strength, 100 mM) was used. Stock solutions ofthe lipids were prepared by dissolving a mixture of 7 mgof DOPC and 3 mg of DOPG in chloroform up to a totallipid concentration of 10 mg/ml. Fifty microliters (con-taining 0.5 mg of lipids) of this solution was dried under astream of nitrogen. When all chloroform had evaporated,the samples were placed in a Speed-Vac for 3 h in order toremove residual chloroform.
Analysis of secondary structure changes in hIAPP andproIAPP
Second derivative and FSD were applied to the normal-ized spectra to identify the components of the amide-I′band region. These peaks were then fitted to the normal-ized raw spectra according to the least squares condition,using a Levenberg–Marquardt curve-fitting routine withbands of Voigt line shape. The starting width at the halfheight of each peak was 8 cm−1. Each peak position wasrestricted to move not farther than 1–3 cm−1 (as deter-mined by shifts in the second derivative and FSD) fromtheir centers during the fitting routine. Such spectral shiftswere allowed to take into account differences in the

918 Propensities and Properties of ProIAPP and IAPP
dielectric environment and conformational dynamics ofthe secondary structures. All spectra were fitted with asimilar set of peaks and parameters. In ambiguous cases(such as the well-known overlap of α-helical and unor-dered band regions), information from CD and NMRexperiments was taken into account was well. The areaunder each peak represents the fraction of each component(assuming similar transition dipole moments for thedifferent conformers) and was finally used to determinethe percentage of secondary structure components.
Preparation of peptide solutions
hIAPP was supplied as a dry powder in vials of 0.5 mgeach (peptide content, 83%). One hundred microliters ofthe stock was lyophilized and dissolved in 1 ml ofphosphate buffer to yield a final concentration of 10 μMhIAPP.
Preparation of LUVs
Of the lipid mixture (anionic: DOPC:DOPG 7:3, 8:2, or9:1; raft mixture: DOPC:DPPC:Chol 1:2:1; or pure DOPC),0.5 mg was dissolved in 1 ml of buffer and ultrasonicatedfor 10 min. After five cycles of freezing and thawing, ahomogeneous solution of multilamellar vesicles wasobtained. To obtain a solution of LUVs, it was pressed11 times through an extruder with a membrane filter madeof polycarbonate (pore diameter, 0.1 μm; Avanti PolarLipids). In the case of the lipid raft mixture, the extrusionstep was performed above its transition temperature (at60 °C).The freshly prepared solution of LUVs was injected
carefully into the ATR flow cell, which was tempered at25 °C. In the case of the lipid raft mixture, the celltemperature was kept at 60 °C. Spreading of the vesiclestook place spontaneously. After adsorption overnight, themembrane was washed with buffer for a time period of 6 h(in the case of the lipid raft mixture at 30 °C, the cell wasthen tempered at 25 °C before the peptide solution wasadded). For this purpose, 2 ml of buffer was pumped froma reservoir into the sample compartment (1.4 ml/min).Lipid vesicles that did not strongly adhere to the ATRcrystal surface were washed away by the buffer solution.
Adsorption and washing of the lipid membrane
The adsorption of the membrane was followed via theband intensity of lipid bands over time. For this purpose, abuffer spectrum was subtracted, and spectra were base-line-corrected in the region of the acyl chains of the lipidsbetween 3060 and 2820 cm− 1. Then, the area wasdetermined by integration between 3030 and 2810 cm−1.During the washing process, spectra were collected bylikewise performing the same steps. To check whethermembrane preparation was successful and to check thereproducibility of the membrane deposition procedure,the area of the acyl-chain region was determined byintegration between 3030 and 2810 cm−1 (average area,∼2.25 cm−1). As shown in earlier studies, we may assumethat after the washing step, one double layer of lipids isleft on the IRE only.45
Adsorption of hIAPP to the lipid membrane andaggregation
To ensure the integrity of the membrane, the hIAPPsolution was injected into the ATR cell by means of aperistaltic pump running at 1.4 ml/min. Then the change
in the amide-I′ band was followed over time. Spectra werecollected every 5 min for a time period of 20 h.
hIAPP in the absence of any membrane
A solution of 10 μM hIAPP was injected into the ATRflow-through cell. Spectra were collected every 5 min overa time period of 20 h.
proIAPP
A solution of 10 or 40 μM proIAPP was injected into themembrane containing the ATR cell, as in previousexperiments. Spectra were collected every 30 min over atime period of 7 days.
Native structures of hIAPP and proIAPP
For determination of the secondary structures of thenative forms of hIAPP and proIAPP, a 10 μM solution ofeach peptide was injected into the ATR flow cell and, aftera few minutes, an ATR-FTIR spectrum was collected.
Atomic force microscopy
A sample from the ATR-FTIR cell was collected anddissolved in chloroform, and the buffer-soluble part wastaken. After a 10-fold dilution with deionized water, 20 μlof the solution was applied onto freshly cleavedmuscovitemica through spin coating and subjected to lyophilization.AFM images were recorded in tapping-in-air mode at adrive frequency of approximately 300 kHz, using aMultiMode™ SPM microscope equipped with a nano-scope IIIa controller (Veco Instruments, Santa Barbara,CA, USA). PointProbe NCHR aluminum-coated silicontips from nanosensors (Neuchâtel, Switzerland) were usedas probe. Control samples (freshly cleaved mica anddiluted buffer solution) were investigated as well toexclude possible artifacts.
Acknowledgements
Financial support from the Deutsche Forschungs-gemeinschaft and the International Max-PlanckResearch School of Chemical Biology is gratefullyacknowledged.
Supplementary Data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.jmb.2009.04.077
References
1. Jarrett, J. T. & Lansbury, P. T., Jr (1993). Seeding “onedimensional crystallization” of amyloid: a pathogenicmechanism in Alzheimer's disease and scrapie? Cell,73, 1055–1058.
2. Chiman, S., Shaibat, M. A., Jones, C. R., Calero, D. C.,Aizezi, B. & Ishii, Y. (2007). Evidence of fibril-like β-

919Propensities and Properties of ProIAPP and IAPP
sheet structures in a neurotoxic amyloid intermediateof Alzheimer's β-amyloid. Nat. Struct. Biol. 14,1157–1164.
3. Grudzielanek, S., Velkova, A., Shukla, A., Smirnovas,V., Tatarek-Nossol, M., Rehage, H. et al. (2007).Cytotoxicity of insulin within its self-assembly andamyloidogenic pathways. J. Mol. Biol. 370, 372–384.
4. Yoshiike, Y., Akagi, T. & Takashima, A. (2007). Surfacestructure of amyloid-β fibrils contributes to cytotoxi-city. Biochemistry, 46, 9805–9812.
5. Clark, A. & Nilsson, M. R. (2004). Islet amyloid: acomplication of islet dysfunction or an aetiologicalfactor in type 2 diabetes? Diabetologia, 47, 157–169.
6. Kahn, S. E., D'Alessio, D. A., Schwartz, M. W.,Fuijimoto, W. Y., Ensinck, J. W., Taborsky, G. J. &Porte, D. (1990). Evidence of cosecretion of isletamyloid polypeptide and insulin by β-cells. Diabetes,39, 634–638.
7. Cooper, G. J. S., Willis, A. C., Clark, A., Turner, R. C.,Sim, R. B. & Reid, K. B. M. (1987). Purification andcharacterization of a peptide from amyloid-richpancreas of type 2 diabetic patients. Proc. Natl Acad.Sci. USA, 84, 8628–8632.
8. Clark, A., Lewis, C. E., Willis, A. C., Cooper, G. J. S.,Morris, J. F., Reid, K. B. M. & Turner, R. C. (1987). Isletamyloid formed from diabetes-associated peptidemay be pathogenic in type-2 diabetes. Lancet, 330,231–234.
9. Rushing, P. A., Hagen, M. M., Seeley, R. J., Lutz, T. A.,D'Alessio, D. A., Air, E. L. & Woods, S. C. (2001).Inhibition of central amylin signaling increase foodintake and body adiposity in rats. Endocrinology, 142,5035–5038.
10. Marcinkiewicz, M., Ramla, D., Seidah, N. G. &Chretien, M. (1994). Developmental expression ofthe prohormone convertase PC1 and PC2 in mousepancreatic islets. Endocrinology, 135, 1651–1660.
11. Sanke, T., Bell, G. I., Sample, C., Rubenstein, A. H. &Steimer, D. F. (1998). An islet amyloid peptide isderived from an 89-amino acid precursor by proteo-lytic processing. J. Biol. Chem. 263, 17243–17246.
12. Marzaban, L., Trigo-Gonzales, G., Zhu, X., Rhodes,C. J., Halban, P. A., Steiner, D. F. & Verchere, C. B.(2004). Role of β-cell prohormone convertase (PC)1/3 in processing of pro-islet amyloid polypeptide.Diabetes, 53, 141–148.
13. Roberts, A. N., Leighton, B., Todd, J. A., Cockburn, D.,Schofield, P. N., Sutton, R. et al. (1989). Molecular andfunctional characterization of amylin, a peptideassociated with type 2 diabetes mellitus. Proc. NatlAcad. Sci. USA, 86, 9662–9666.
14. Singh, G., Brovchenko, I., Oleinikova, A. & Winter, R.(2008). Peptide aggregation in finite systems. Biophys.J. 95, 3208–3221.
15. Paulsson, J. F., Andersson, A., Westermark, P. &Westermark, G. T. (2006). Intracellular amyloid-likedeposits contain unprocessed pro-islet amyloid poly-peptide (proIAPP) in β-cells of transgenic mice overexpressing the gene for human IAPP and transplantedhuman islets. Diabetologia, 49, 1237–1246.
16. Clement, L., Kim-Sohn, K. A., Magnan, C., Kassis, N.,Adnot, P., Kergoat, M. et al. (2002). Pancreatic beta-cellalpha2A adrenoceptor and phospholipid changes inhyperlipidemic rats. Lipids, 37, 501–506.
17. Wolf, B. A., Pasquals, S. M. & Turk, J. (1991). Free fattyacid accumulation in secretagogue-stimulated pan-creatic islets and effects of arachidonate on depolar-ization-induced insulin secretion. Biochemistry, 30,6372–6379.
18. Turk, J., Wolf, B. A., Lefkowith, J. B., Stump, W. T.& McDaniel, M. L. (1986). Glucose-induced phos-pholipid hydrolysis in isolated pancreatic islets:quantitative effects on the phospholipid content ofarachidonate and other fatty acids. Biochim. Biophys.Acta, 879, 399–409.
19. Knight, J. D. & Miranker, A. D. (2004). Phospholipidcatalysis of diabetic amyloid assembly. J. Mol. Biol.341, 1175–1187.
20. Westermark, G. T., Westermark, P., Eizirik, D. L.,Hellerstrom, C., Fox, N., Steiner, D. F. & Anderson, A.(1999). Difference in amyloid deposition in islets oftransgenic mice expressing human islet amyloidpolypeptide versus human islets implanted into nudemice. Metab. Clin. Exp. 48, 448–454.
21. Westermark, G. T., Steiner, D. F., Gebre-Medhin, S.,Engstrom, U. & Westermark, P. (2000). Pro isletamyloid polypeptide (proIAPP) immunoreactivity inthe islets of Langerhans. Upsala J. Med. Sci. 105,97–106.
22. Park, K. & Verchere, C. B. (2001). Identification of aheparin binding domain in the N-terminal cleavagesite of pro-islet amyloid polypeptide. Implicationfor islet amyloid formation. J. Biol. Chem. 276,16611–16616.
23. Yonemoto, I. T., Kroon, G. J. A., Dyson, H. J., Balch, W.E. & Kelly, J. W. (2008). Amylin proprotein processinggenerates progressively more amyloidogenic peptidesthat initially sample the helical state. Biochemistry, 47,9900–9910.
24. Wood, S. J., Maleeff, B., Hart, T. & Werzel, R.(1996). Physical, morphological and functionaldifferences between pH 5.8 and 7.4 aggregates ofthe Alzheimer's amyloid peptide Aβ. J. Mol. Biol.256, 870–877.
25. Krampert, M., Bernhagen, J., Schmucker, J., Horn, A.,Schumauder, A., Brunner, H. et al. (2000). Amyloido-genicity of recombinant human pro-islet amyloidpolypeptide (proIAPP). Chem. Biol. 7, 856–871.
26. Böhm, G., Muhr, R. & Jaenicke, R. (1992). Quantitativeanalysis of protein far UV circular dichroism spectraby neural networks. Protein Eng. 5, 191–195.
27. Naiki, H., Higuchi, K., Hosokawa, M. & Takeda, T.(1989). Fluorometric determination of amyloid fibrilsin vitro using the fluorescent dye, thioflavin T. Anal.Biochem. 177, 244–249.
28. Mishra, R., Bulic, B., Sellin, D., Jha, S., Waldmann, H.& Winter, R. (2008). Small-molecule inhibitors of isletamyloid polypeptide fibril formation. Angew. Chem.Int. Ed. 47, 4679–4682.
29. Lopes, D. H., Meister, A., Gohlke, A., Hauser, A.,Blume, A. & Winter, R. (2007). Mechanism of isletamyloid polypeptide fibrillation at lipid interfacesstudied by infrared reflection absorption spectro-scopy. Biophys. J. 93, 3132–3141.
30. Canal, C., Torrassa, S., Rispoli, P., Relini, A., Rolandi,R., Bucciantini, M. et al. (2006). Natively folded HypF-N and its early amyloid aggregates interact withphospholipid monolayers and destabilize supportedphospholipid bilayers. Biophys. J. 91, 4575–4588.
31. Meng, X., Fink, A. L. & Uversky, V. N. (2008). Theeffect of membranes on the in vitro fibrillation ofan amyloidogenic light-chain variable-domain SMA.J. Mol. Biol. 381, 989–999.
32. Maarten, F. M., Engel, L. K., Kleijer, C. C., Meeldijk, H.J. D., Jacobs, J., Verkleij, A. J. et al. (2008). Membranedamage by human islet amyloid polypeptide throughfibril growth at the membrane. Proc. Natl Acad. Sci.USA, 105, 6033–6038.

920 Propensities and Properties of ProIAPP and IAPP
33. Yan, L. -M., Tatarek-Nossol,M., Velkova, A., Kazantzis,A. & Kapurniotu, A. (2006). Design of a mimic ofnonamyloidogenic and bioactive human islet amyloidpolypeptide (IAPP) as nanomolar affinity inhibitor ofIAPP cytotoxic fibrillogenesis.Proc.Natl Acad. Sci. USA,103, 2046–2951.
34. Knight, J. D., Hebda, J. A. & Miranker, A. D. (2006).Conserved and cooperative assembly of membrane-bound α-helical states of islet amyloid polypeptide.Biochemistry, 45, 9496–9508.
35. Higham, C. E., Jaikaran, E. T. A. S., Fraser, P. E., Gross,M. & Clark, A. (2000). Preparation of synthetic humanislet amyloid polypeptide (IAPP) in a stable con-formation to enable study of conversion to amyloid-like fibrils. FEBS Lett. 470, 55–60.
36. Kayed, R., Bernhagen, J., Greenfield, N., Sweimeh, K.,Brunner, H., Voelter, W. & Kapurniotu, A. (1999).Conformational transitions of islet amyloid polypep-tide (IAPP) in amyloid formation in vitro. J. Mol. Biol.287, 781–796.
37. Radovan, D., Smirnovas, V. & Winter, R. (2008). Effectof pressure on islet amyloid polypeptide aggregation:revealing the polymorphic nature of fibrillationprocess. Biochemistry, 47, 6352–6360.
38. Apetri,M.M.,Maiti, N. C., Zagorski,M.G., Carey, P. R.& Anderson, V. E. (2006). Secondary structure of α-synuclein oligomers: characterization by Raman andatomic force microscopy. J. Mol. Biol. 355, 63–71.
39. Marek, P., Abedini, A., Song, B. B., Kanungo, M.,Johnson, M. E., Gupta, R. et al. (2007). Aromaticinteractions are not required for amyloid fibrilformation by islet amyloid polypeptide but doinfluence the rate of fibril formation and fibrilmorphology. Biochemistry, 46, 3255–3261.
40. Jansen, R., Dzwolak, W. & Winter, R. (2005). Amyloi-dogenic self-assembly of insulin aggregates probed byhigh resolution atomic force microscopy. Biophys. J. 88,1344–1353.
41. Ortiz, C., Zhang, D., Ribbe, A. E., Xie, Y. & Ben-Amotz,D. (2007). Analysis of insulin amyloid fibrils byRaman spectroscopy. Biophys. Chem. 128, 150–155.
42. Hong, D.-P., Ahmad, A. & Fink, A. I. (2006). Fibrilla-tion of human insulin A and B chains. Biochemistry, 45,9342–9353.
43. Krebs, M. R. H., Morozova-Roche, L. A., Daniel, K.,Robinson, C. V. & Dobson, C. M. (2004). Observationof sequence specificity in the seeding of proteinamyloid fibrils. Protein Sci. 13, 1933–1938.
44. Pace, C. N., Vajdos, F., Fee, L., Grimsley, G. &Gray, T. (1995). How to measure and predict themolar absorption coefficient of a protein. Protein Sci.4, 2411–2423.
45. Tatulian, S. A. (2003). Attenuated total reflectionFourier transform infrared spectroscopy: a methodof choice for studying membrane proteins and lipids.Biochemistry, 42, 11898–11907.


















